Наследственность
| Часть серии на |
| Генетика |
|---|
 |
|
|
Наследственность , также называемая наследством или биологическим наследством , является передачей черт от родителей к их потомству; Либо посредством бесполого репродукции или сексуального размножения потомства , клетки или организмы получают генетическую информацию своих родителей. Благодаря наследственности различия между людьми могут накапливаться и привести к тому, виды что развиваются при естественном отборе . Изучение наследственности в биологии - генетика .
Обзор
[ редактировать ]

У людей цвет глаз является примером унаследованной характеристики: человек может наследовать «черту коричневого глаза» от одного из родителей. [ 1 ] Унаследованные признаки контролируются генами организма , а полный набор генов в геноме называется его генотипом . [ 2 ]
Полный набор наблюдаемых черт структуры и поведения организма называется его фенотипом . Эти черты возникают из -за взаимодействия генотипа организма с окружающей средой . [ 3 ] В результате многие аспекты фенотипа организма не унаследованы. Например, замороженная кожа происходит от взаимодействия между генотипом человека и солнечным светом; [ 4 ] Таким образом, Suntans не передаются детям людей. Однако некоторые люди загорают легче, чем другие, из -за различий в их генотипе: [ 5 ] Поразительным примером являются люди с унаследованной чертой альбинизма , которые вообще не загорают и очень чувствительны к солнечным ожогам . [ 6 ]
Известно, что наследственные признаки передаются из одного поколения в другое через ДНК , молекулу , которая кодирует генетическую информацию. [ 2 ] ДНК - это длинный полимер , который включает в себя четыре типа оснований , которые являются взаимозаменяемыми. Последовательность нуклеиновой кислоты (последовательность оснований вдоль определенной молекулы ДНК) указывает генетическую информацию: это сопоставимо с последовательности букв, прописавших проход текста. [ 7 ] Перед тем, как клетка делится через митоз , копируется ДНК, так что каждая из полученных двух клеток будет наследовать последовательность ДНК. Часть молекулы ДНК, которая указывает одну функциональную единицу, называется геном ; Различные гены имеют разные последовательности оснований. Внутри клеток длинные нити ДНК образуют конденсированные структуры, называемые хромосом . Организма наследуют генетический материал от своих родителей в форме гомологичных хромосом , содержащих уникальную комбинацию последовательностей ДНК, которые кодируют для генов. Специфическое местоположение последовательности ДНК в хромосоме известно как локус . Если последовательность ДНК в определенном локусе варьируется между индивидуумами, различные формы этой последовательности называются аллелями . Последовательности ДНК могут меняться с помощью мутаций , производя новые аллели. Если мутация происходит в гене, новый аллель может влиять на признак, которую контролирует ген, изменяя фенотип организма. [ 8 ]
Однако, хотя в некоторых случаях это простое соответствие между аллелем и чертой работает, большинство признаков являются более сложными и контролируются множественными взаимодействующими генами внутри и среди организмов. [ 9 ] [ 10 ] Биологи развития предполагают, что сложные взаимодействия в генетических сетях и связь между клетками могут привести к наследственным изменениям, которые могут лежать в основе некоторых механиков в развития пластичности и канализации . [ 11 ]
Недавние результаты подтвердили важные примеры наследственных изменений, которые не могут быть объяснены прямым агентством молекулы ДНК. Эти явления классифицируются как эпигенетические системы наследования, которые причинно или независимо развиваются по генам. Исследования режимов и механизмов эпигенетического наследования все еще находятся в его научном младенчестве, но эта область исследований привлекла гораздо недавнюю деятельность, поскольку она расширяет масштаб наследуемости и эволюционной биологии в целом. [ 12 ] ДНК-метилирование, отмечающие хроматин , самоокупаемые метаболические петли , молчание генов путем интерференции РНК и трехмерная конформация белков (таких как прионы )-это области, где были обнаружены эпигенетические системы наследования. [ 13 ] [ 14 ] Наследственность также может возникнуть в еще больших масштабах. Например, экологическое наследование в процессе нишевой конструкции определяется регулярной и повторной деятельностью организмов в их окружающей среде. Это генерирует наследие эффекта, которое изменяет и подается обратно в режим отбора последующих поколений. Потомки наследуют гены плюс характеристики окружающей среды, генерируемые экологическими действиями предков. [ 15 ] Другие примеры наследуемости в эволюции, которые не находятся под прямым контролем генов, включают наследство культурных признаков , групповой наследуемости и симбиогенеза . [ 16 ] [ 17 ] [ 18 ] Эти примеры наследуемости, которые работают над геном, широко охватываются под названием многоуровневого или иерархического отбора , который был предметом интенсивных дебатов в истории эволюционной науки. [ 17 ] [ 19 ]
Отношение к теории эволюции
[ редактировать ]Когда в 1859 году Чарльз Дарвин предложил свою теорию эволюции , одной из основных проблем стал отсутствие основного механизма для наследственности. [ 20 ] Дарвин верил в сочетание смешивания наследования и наследства приобретенных признаков ( пангенезис ). Смешивание наследования приведет к однородности среди населения всего за несколько поколений, а затем удалит вариации от популяции, на которой может действовать естественный отбор. [ 21 ] Это привело к тому, что Дарвин принял некоторые ламаркские идеи в более поздних изданиях о происхождении видов и его более поздних биологических работах. [ 22 ] Основной подход Дарвина к наследственности состоял в том, чтобы рассказать о том, как он, по-видимому, работает (замечая, что признаки, которые явно не были выражены у родителей во время воспроизведения, могут быть унаследованы, что определенные признаки могут быть связаны с полом и т. Д.), А не предполагают механизмы Полем [ Цитация необходима ]
Первоначальная модель наследия Дарвина была принята, а затем сильно модифицирована его двоюродным братом Фрэнсисом Гальтоном , который заложил основу для биометрической школы наследственности. [ 23 ] Гальтон не нашел никаких доказательств в поддержку аспектов модели пангенезиса Дарвина, которая опиралась на приобретенные черты. [ 24 ]
Было показано, что наследство приобретенных признаков имело небольшую основу в 1880 -х годах, когда август Вайсманн отрезал хвосты от многих поколений мышей и обнаружил, что их потомство продолжало развивать хвосты. [ 25 ]
История
[ редактировать ]
Ученые в древности имели множество представлений о наследственности: Теофраст предположил, что мужские цветы заставляли созревать женские цветы; [ 26 ] Гиппократ предположил, что «семена» были получены различными частями тела и передавались потомству во время зачатия; [ 27 ] и Аристотель думал, что мужские и женские жидкости смешаны в концепции. [ 28 ] Aeschilus , в 458 году до нашей эры, предложил мужчине в качестве родителя с женщиной в качестве «медсестры для молодой жизни, посеянной внутри нее». [ 29 ]
Древнее понимание наследственности перешло к двум обсуждению доктрины в 18 веке. Доктрина эпигенеза и доктрины преформации были двумя разными взглядами на понимание наследственности. Доктрина эпигенеза, созданная Аристотелем , утверждала, что эмбрион постоянно развивается. Модификации черт родителей передаются эмбриону в течение его жизни. Основа этой доктрины была основана на теории наследования приобретенных признаков . В прямой оппозиции доктрина преформации утверждала, что «как генерирует подобное», где зародыш будет развиваться, чтобы уступить потомство, похожее на родителей. Преформационная точка зрения считает, что размножение было актом раскрытия того, что было создано задолго до этого. Тем не менее, это было оспорено созданием теории клеток в 19 веке, где фундаментальная единица жизни является ячейка, а не некоторых предварительно сформированных частей организма. Различные наследственные механизмы, в том числе смешивание наследования, также были предусмотрены без надлежащего тестирования или количественного определения, а затем были оспорены. Тем не менее, люди смогли развивать домашние породы животных, а также урожай в результате искусственного отбора. Наследование приобретенных признаков также составляло часть ранних ламаркских идей об эволюции. [ Цитация необходима ]
В течение 18 -го века голландский микроскопист Антони Ван Леувенхук (1632–1723) обнаружила «животные» в сперме людей и других животных. [ 30 ] Некоторые ученые предположили, что они видели «маленького человека» ( Homunculus ) внутри каждой спермы . Эти ученые сформировали школу мысли, известную как «спермисты». Они утверждали, что единственным вкладом женщины в следующее поколение была матка, в которой рос Homunculus, и пренатальные влияния матки. [ 31 ] Овисты, противостоящая школа мышления, полагали, что будущий человек был в яйце, и что сперма просто стимулировала рост яйца. Овисты думали, что женщины несли яйца, содержащие детей и детей, и что пол потомства был определен задолго до зачатия. [ 32 ]
Ранняя исследовательская инициатива появилась в 1878 году, когда Альфей Хаятт провел исследование, чтобы изучить законы наследственности путем составления данных о семейных фенотипах (размер носа, формы уха и т. Д.) И выражение патологических условий и ненормальных характеристик, особенно в отношении возраста. внешности. Одним из целей проекта было табличное сведения данных, чтобы лучше понять, почему определенные признаки последовательно выражаются, в то время как другие очень нерегулярны. [ 33 ]
Грегор Мендель: отец генетики
[ редактировать ]
Идея наследования частиц генов может быть связана с моравской [ 34 ] МОНК ГРЕГОР МЕНДЕЛЬ , который опубликовал свою работу над растениями гороха в 1865 году. Однако его работа не была широко известна и была заново открыта в 1901 году. Первоначально предполагалось, что наследство Мендельяна только что приходилось на большие (качественные) различия, такие как те, которые видели Мендель в Его гороховые растения - и идея аддитивного эффекта (количественных) генов не была реализована до тех пор, пока не была статья Рай Фишер (1918) получена . были наследуемыми. Его демонстрация горохового растения стала основой изучения мендельских черт. Эти черты можно проследить на одном локусе. [ 35 ]
Современное развитие генетики и наследственности
[ редактировать ]В 1930 -х годах работа Фишера и других привела к сочетанию менделевских и биометрических школ в современный эволюционный синтез . Современный синтез преодолел разрыв между экспериментальными генетиками и натуралистами; и между палеонтологами, заявляя, что: [ 36 ] [ 37 ]
- Все эволюционные явления могут быть объяснены в соответствии с известными генетическими механизмами и наблюдательными свидетельствами натуралистов.
- Эволюция является постепенной: небольшие генетические изменения, рекомбинация, упорядоченная естественным отбором . Разрывы среди видов (или других таксонов) объясняются постепенно происходящим посредством географического разделения и вымирания (не солнения).
- Отбор в подавляющем большинстве является основным механизмом изменений; Даже небольшие преимущества важны при продолжении. Объектом отбора является фенотип в окружающей среде. Роль генетического дрейфа является двусмысленной; он был убедительно поддержан Несмотря на то, что Добжанский , он был понижен позже, так как были получены результаты экологической генетики.
- Примат популяционного мышления: генетическое разнообразие, переносимое в естественных популяциях, является ключевым фактором в эволюции. Сила естественного отбора в дикой природе была больше, чем ожидалось; Эффект экологических факторов, таких как нишевая оккупация и значение барьеров для потока генов, все важны.
Идея о том, что видообразование возникает после того, как популяции репродуктивно изолированы, много обсуждается. [ 38 ] У растений полиплоидия должна быть включена в любой вид видообразования. Такие составы, как «эволюция, в основном состоит из изменений в частотах аллелей между одним поколением и другим», были предложены довольно позже. Традиционное мнение состоит в том, что биология развития (« evo-devo ») сыграла небольшую роль в синтезе, но рассказ о Гэвина де Пира работе Стивена Джей Гулд предполагает, что он может быть исключением. [ 39 ]
Практически все аспекты синтеза иногда сталкивались с различной степенью успеха. Однако нет никаких сомнений в том, что синтез был большим значением эволюционной биологии. [ 40 ] Это прояснило много путаников и было непосредственно ответственным за стимулирование большого исследования в эпоху после Второй мировой войны .
Трофим Лизенко, однако, вызвал негативную реакцию того, что сейчас называется лизенкоизмом в Советском Союзе , когда он подчеркнул ламарккианские идеи о наследстве приобретенных признаков . Это движение повлияло на сельскохозяйственные исследования и привело к нехватке продовольствия в 1960 -х годах и серьезно повлияло на СССР. [ 41 ]
Появляется все больше доказательств того, что существует трансгенерационное наследование эпигенетических изменений у людей [ 42 ] и другие животные. [ 43 ]
Общие генетические расстройства
[ редактировать ]Типы
[ редактировать ]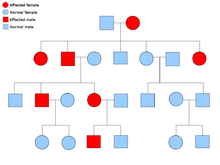
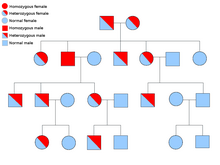
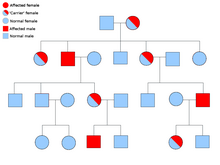
Описание способа биологического наследования состоит из трех основных категорий:
- 1. Количество вовлеченных локусов
- Моногенетический (также называемый «простой») - один локус
- Олигогенный - несколько локусов
- Полигенетический - Много локусов
- 2. вовлечены хромосомы
- Аутосомные - локусы не расположены на сексуальной хромосоме
- Гоносомал - локусы расположены на сексуальной хромосоме
- X-Chromosomal-локусы расположены на X-хромосоме (более распространенный случай)
- Y-хромосомальный-локусы расположены на Y-хромосоме
- Митохондрия - локусы расположены на митохондриальной ДНК
- 3. Корреляционный генотип - фенотип
- Доминирующий
- Промежуточный (также называемый « кодоминант »)
- Рецессивный
- Чрезмерный
- Удержан
Эти три категории являются частью каждого точного описания способа наследования в вышеуказанном порядке. Кроме того, можно добавить больше спецификаций следующим образом:
- 4. Случайно и экологические взаимодействия
- Проницательность
- Полный
- Неполный (процентное число)
- Выразительность
- Неизбежно
- Переменная
- Наследственность (в полигенетических, а иногда и в олигогенетических способах наследования)
- Явления материнского или отцовского отпечатка (также см. Эпигенетику )
- Проницательность
- 5. Секс-связанные взаимодействия
- Связанное с полом наследование ( гоносомальные локусы)
- Ограниченная полом экспрессия фенотипа (EG, крипторхизм )
- Наследование через материнскую линию (в случае митохондриальных локусов ДНК)
- Наследование через отцовскую линию (в случае y-хромосомных локусов)
- 6. Взаимодействие Locus -Locus
- Эпистаз с другими локусами (например, передоминирование )
- Связание генов с другими локусами (также см .
- Гомозиготы смертельных факторов
- Полулетальные факторы
Определение и описание способа наследования также достигаются главным образом посредством статистического анализа данных родословных. В случае известны вовлеченные локусы, методы молекулярной генетики также могут быть использованы.
Доминирующие и рецессивные аллели
[ редактировать ]является Говорят, что аллель доминирующим, если он всегда выражается в появлении организма (фенотип) при условии, что присутствует по крайней мере одна его копия. Например, в городе аллель для зеленых стручков, g , доминирует для того, чтобы для желтых стручков, g . Таким образом, растения гороха с парой аллелей либо GG (гомозигота) , либо GG (гетерозигота) будут иметь зеленые стручки. Аллель для желтых стручков рецессив. Эффекты этого аллеля наблюдаются только тогда, когда он присутствует в обеих хромосомах, GG (гомозигота). Это происходит из Zygosity , степени, в которой обе копии хромосомы или гена имеют одинаковую генетическую последовательность, другими словами, степень сходства аллелей в организме.
-
 Наследственные дефекты в ферментах, как правило, унаследованы аутосомно, потому что существует больше нех-хромосом, чем X-хромосомы, и рецессивную моду, потому что ферменты из незатронутых генов, как правило, достаточны для предотвращения симптомов у носителей.
Наследственные дефекты в ферментах, как правило, унаследованы аутосомно, потому что существует больше нех-хромосом, чем X-хромосомы, и рецессивную моду, потому что ферменты из незатронутых генов, как правило, достаточны для предотвращения симптомов у носителей. -
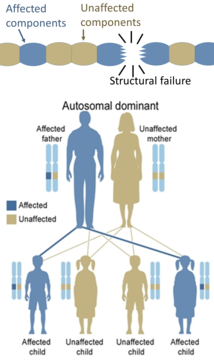 С другой стороны, наследственные дефекты в структурных белках (такие как несовершенство остеогенеза , синдром Марфана и многие синдромы Элеров -Данлос ), как правило, являются аутосомно -доминантными, потому что достаточно, чтобы некоторые компоненты были дефектными, чтобы сделать всю структуру дисфункциональной. Это доминантно-негативный процесс, в котором продукт мутированного гена отрицательно влияет на не мутированный генный продукт в одной и той же клетке.
С другой стороны, наследственные дефекты в структурных белках (такие как несовершенство остеогенеза , синдром Марфана и многие синдромы Элеров -Данлос ), как правило, являются аутосомно -доминантными, потому что достаточно, чтобы некоторые компоненты были дефектными, чтобы сделать всю структуру дисфункциональной. Это доминантно-негативный процесс, в котором продукт мутированного гена отрицательно влияет на не мутированный генный продукт в одной и той же клетке.


Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Штурм Р.А.; Frudakis TN (2004). «Цвет глаз: порталы в гены пигментации и происхождение». Тенденции Genet . 20 (8): 327–332. doi : 10.1016/j.tig.2004.06.010 . PMID 15262401 .
- ^ Jump up to: а беременный Пирсон Х (2006). "Генетика: что такое ген?" Полем Природа . 441 (7092): 398–401. Bibcode : 2006natur.441..398p . doi : 10.1038/441398a . PMID 16724031 . S2CID 4420674 .
- ^ Visscher PM; Холм wg; Рэй Н.Р. (2008). «Наследственность в эпоху геномики - концепции и заблуждения». НАТ Преподобный Генет . 9 (4): 255–266. doi : 10.1038/nrg2322 . PMID 18319743 . S2CID 690431 .
- ^ Shoag J; и др. (Январь 2013). «Коактиваторы PGC-1 регулируют MITF и реакцию загара» . Моллют . 49 (1): 145–157. doi : 10.1016/j.molcel.2012.10.027 . PMC 3753666 . PMID 23201126 .
- ^ Pho ln; Leachman SA (февраль 2010 г.). «Генетика пигментации и меланомы предрасположенность» . Gital Dermatol Venereol . 145 (1): 37–45. PMID 20197744 . Архивировано из оригинала 2019-03-28 . Получено 2013-03-26 .
- ^ Oetting ws; Блестящий MH; Кинг Р.А. (1996). «Клинический спектр альбинизма у людей и действием». Молекулярная медицина сегодня . 2 (8): 330–335. doi : 10.1016/1357-4310 (96) 81798-9 . PMID 8796918 .
- ^ Гриффитс, Энтони, JF; Весслер, Сьюзен Р.; Кэрролл, Шон Б.; Doebley J (2012). Введение в генетический анализ (10 -е изд.). Нью -Йорк: WH Freeman and Company. п. 3. ISBN 978-1-4292-2943-2 .
{{cite book}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Futuyma, Douglas J. (2005). Эволюция Сандерленд, Массачусетс: Sinauer Associates, Inc. ISBN 978-0-87893-187-3 .
- ^ Phillips PC (2008). «Эпистаз - важная роль генных взаимодействий в структуре и эволюции генетических систем» . НАТ Преподобный Генет . 9 (11): 855–867. doi : 10.1038/nrg2452 . PMC 2689140 . PMID 18852697 .
- ^ Wu r; Лин М (2006). «Функциональное картирование - как отображать и изучать генетическую архитектуру динамических сложных признаков». НАТ Преподобный Генет . 7 (3): 229–237. doi : 10.1038/nrg1804 . PMID 16485021 . S2CID 24301815 .
- ^ Jablonka, E.; Лэмб, MJ (2002). «Изменная концепция эпигенетики» (PDF) . Анналы нью -йоркской академии наук . 981 (1): 82–96. Bibcode : 2002nyasa.981 ... 82J . doi : 10.1111/j.1749-6632.2002.tb04913.x . PMID 12547675 . S2CID 12561900 . Архивировано из оригинала (PDF) 2011-05-11.
- ^ Jablonka, E.; Раз, Г. (2009). «Трансгенерационное эпигенетическое наследование: распространенность, механизмы и последствия для изучения наследственности и эволюции» (PDF) . Ежеквартальный обзор биологии . 84 (2): 131–176. Citeseerx 10.1.1.617.6333 . doi : 10.1086/598822 . PMID 19606595 . S2CID 7233550 . Архивировано (PDF) из оригинала 2011-07-15 . Получено 2011-02-18 .
- ^ Bossdorf, O.; Arcuri, D.; Ричардс, CL; Pigliucci, M. (2010). «Экспериментальное изменение метилирования ДНК влияет на фенотипическую пластичность экологически значимых признаков у Arabidopsis thaliana » (PDF) . Эволюционная экология . 24 (3): 541–553. doi : 10.1007/s10682-010-9372-7 . S2CID 15763479 . Архивировано (PDF) из оригинала с 2020-03-01 . Получено 2019-08-15 .
- ^ Jablonka, E.; Лэмб, М. (2005). Эволюция в четырех измерениях: генетические, эпигенетические, поведенческие и символические . MIT Press. ISBN 978-0-262-10107-3 Полем Архивировано из оригинала 2021-12-27 . Получено 2015-06-27 .
- ^ Лаленд, кн; Sterelny, K. (2006). «Перспектива: семь причин (не), чтобы пренебрегать нишей строительство» . Эволюция 60 (8): 1751–1762. doi : 10.1111/j.0014-3820.2006.tb00520.x . PMID 17089961 .
- ^ Чепмен, MJ; Маргулис Л. (1998). «Морфогенез с помощью симбиогенеза» (PDF) . Международная микробиология . 1 (4): 319–326. PMID 10943381 . Архивировано из оригинала (PDF) 2014-08-23.
- ^ Jump up to: а беременный Уилсон, DS; Wilson, EO (2007). «Переосмысление теоретической основы социобиологии» (PDF) . Ежеквартальный обзор биологии . 82 (4): 327–348. doi : 10.1086/522809 . PMID 18217526 . S2CID 37774648 . Архивировано из оригинала (PDF) 2011-05-11.
- ^ Биджма, P.; Уэйд, MJ (2008). «Совместные эффекты родственного, многоуровневого отбора и косвенных генетических эффектов на реакцию на генетический отбор» . Журнал эволюционной биологии . 21 (5): 1175–1188. doi : 10.1111/j.1420-9101.2008.01550.x . PMID 18547354 . S2CID 7204089 .
- ^ VRBA, ES; Гулд, С.Дж. (1986). «Иерархическое расширение сортировки и выбора: сортировка и выбор не могут быть приравниваются» (PDF) . Палеобиология . 12 (2): 217–228. Bibcode : 1986pbio ... 12..217v . doi : 10.1017/s0094837300013671 . S2CID 86593897 . Архивировано из оригинала (PDF) 2016-08-04 . Получено 2011-02-18 .
- ^ Гриффитс, Энтони, JF; Весслер, Сьюзен Р.; Кэрролл, Шон Б.; Доубли, Джон (2012). Введение в генетический анализ (10 -е изд.). Нью -Йорк: WH Freeman and Company. п. 14. ISBN 978-1-4292-2943-2 .
{{cite book}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Чарльзворт, Брайан и Чарльзворт, Дебора (ноябрь 2009 г.). «Дарвин и генетика» . Генетика . 183 (3): 757–766. doi : 10.1534/Genetics.109.109991 . PMC 2778973 . PMID 19933231 . Архивировано из оригинала 2019-04-29 . Получено 2013-03-26 .
- ^ Бард, Джонатан Бл (2011). «Следующий эволюционный синтез: от Ламарка и Дарвина до геномной вариации и системной биологии» . Клеточная связь и передача сигналов . 9 (30): 30. doi : 10.1186/1478-811x-9-30 . PMC 3215633 . PMID 22053760 .
- ^ «Фрэнсис Гальтон (1822-1911)» . Научный музей . Архивировано с оригинала 30 января 2016 года . Получено 26 марта 2013 года .
- ^ Лю Й. (май 2008 г.). «Новый взгляд на пангенезис Дарвина». Biol Rev Camb Philos Soc . 83 (2): 141–149. doi : 10.1111/j.1469-185x.2008.00036.x . PMID 18429766 . S2CID 39953275 .
- ^ Липтон, Брюс Х. (2008). Биология веры: раскрыть силу сознания, материи и чудес . Hay House, Inc. с. 12 . ISBN 978-1-4019-2344-0 .
- ^ Негеби, Моше (лето 1995). «Мужчина и женщина в ботанических работах Теофраста». Журнал истории биологии . 28 (2): 317–332. doi : 10.1007/bf01059192 . S2CID 84754865 .
- ^ Hipócrates (1981). Гиппократические трактаты: о поколении - природа ребенка - болезнь IC . Уолтер де Грютер. п. 6. ISBN 978-3-11-007903-6 .
- ^ «Биология Аристотеля - 5.2. От расследования до понимания; от Хоти до диоти» . Стэнфордский университет. 15 февраля 2006 года. Архивировано из оригинала 7 мая 2019 года . Получено 26 марта 2013 года .
- ^ Eumenides 658-661
- ^ Снег, Курт. «Удивительный маленький маленький« Animalcules »Антони Ван Леувенхук » . Лебен. Архивировано из оригинала 24 апреля 2013 года . Получено 26 марта 2013 года .
- ^ Лоуренс, Сера Р. (2008). Эскиз Харсоекера Homunculus от Essai de Dioptrique . Эмбрион проект энциклопедия. ISSN 1940-5030 . Архивировано из оригинала 2013-04-09 . Получено 26 марта 2013 года .
- ^ Готлиб, Гилберт (2001). Индивидуальное развитие и эволюция: генезис нового поведения . Психология пресса. п. 4. ISBN 978-1-4106-0442-2 .
- ^ Scientific American, «наследственность» . Munn & Company. 1878-11-30. п. 343. Архивировано из оригинала 2022-05-18 . Получено 2021-08-06 .
- ^ Хениг, Робин Маранц (2001). Монах в саду: потерянный и найденный гений Грегора Менделя, отца генетики . Хоутон Миффлин. ISBN 978-0-395-97765-1 Полем
Статья, написанная неясным моравским монахом по имени Грегор Мендель
- ^ Jump up to: а беременный Карлсон, Нил Р. (2010). Психология: наука о поведении , с. 206. Торонто: Пирсон Канада. ISBN 978-0-205-64524-4 . OCLC 1019975419
- ^ Mayr & приходит с 1998 года
- ^ Mayr E. 1982. Рост биологической мысли: разнообразие, эволюция и наследство . Гарвард, Камбс. С. 567 и след.
- ^ Палумби, Стивен Р. (1994). «Генетическая дивергенция, репродуктивная изоляция и морское видообразование». Ежегодный обзор экологии и систематики . 25 : 547–572. doi : 10.1146/annurev.es.25.110194.002555 .
- ^ Gould SJ Ontogeny и Phyologgeny . Гарвард, 1977. С. 221–222
- ^ Handschuh, Стефан; Mitteroecker, Филипп (июнь 2012 г.). «Эволюция - расширенный синтез. Исследовательское предложение, достаточно убедительное для большинства эволюционных биологов?». Бюллетень человеческой обтерологии . 27 (1–2): 18–21. ISSN 2224-4476 .
- ^ Харпер, Питер С. (2017-08-03). «Человеческая генетика в беспокойные времена и места» . Hereditas . 155 : 7. doi : 10.1186/s41065-017-0042-4 . ISSN 1601-5223 . PMC 5541658 . PMID 28794693 .
- ^ Szyf, M (2015). «Негенетическое наследование и трансгенерационная эпигенетика». Тенденции в молекулярной медицине . 21 (2): 134–144. doi : 10.1016/j.molmed.2014.12.004 . PMID 25601643 .
- ^ Кишимото, с; и др. (2017). «Стрессы в окружающей среде вызывают трансгенерационно наследуемые преимущества выживаемости посредством общения за зародышевой линией в Caenorhabditis elegans» . Природная связь . 8 : 14031. BIBCODE : 2017Natco ... 814031K . doi : 10.1038/ncomms14031 . HDL : 2433/217772 . PMC 5227915 . PMID 28067237 .