Врожденная стационарная куриная слепота
| Врожденная стационарная куриная слепота | |
|---|---|
 | |
| нарушение передачи от фоторецепторов внешнего ядерного слоя к биполярным клеткам внутреннего ядерного слоя . В основе CSNB лежит | |
| Специальность | Офтальмология |
Врожденная стационарная куриная слепота ( ВСНБ ) — редкое непрогрессирующее заболевание сетчатки . Люди с CSNB часто испытывают трудности с адаптацией к условиям низкой освещенности из-за нарушения фоторецепторов передачи . У этих пациентов также может наблюдаться снижение остроты зрения, близорукость , нистагм , аномалии глазного дна и косоглазие . [ 1 ] [ 2 ] CSNB имеет две формы — полную, также известную как тип 1 (CSNB1), и неполную, также известную как тип 2 (CSNB2), которые отличаются вовлечением различных путей сетчатки. В CSNB1 нижестоящие нейроны, называемые биполярными клетками , не способны обнаруживать нейротрансмиссию от фоторецепторных клеток. CSNB1 может быть вызван мутациями в различных генах, участвующих в обнаружении нейротрансмиттеров, включая NYX . При CSNB2 сами фоторецепторы имеют нарушенную функцию нейротрансмиссии; это вызвано в первую очередь мутациями в гене CACNA1F , который кодирует потенциалзависимый кальциевый канал, важный для высвобождения нейромедиаторов. CSNB был идентифицирован у лошадей и собак в результате мутаций TRPM1 (Horse, «LP»). [ 3 ] , ГРМ6 (Лошадь, «ЦСНБ2») [ 4 ] ( и LRIT3 Собака , CSNB) [ 5 ] .
Врожденная стационарная куриная слепота (ВСНБ) может наследоваться по Х-сцепленному, аутосомно-доминантному или аутосомно-рецессивному типу, в зависимости от задействованных генов.
Две формы CSNB также могут поражать лошадей: одна связана с леопардовым комплексом окрасов лошадиной шерсти, а другая встречается у некоторых пород лошадей. Оба являются аутосомно-рецессивными. [ 6 ] [ 7 ]
Симптомы и признаки
[ редактировать ]Х-сцепленные разновидности врожденной стационарной куриной слепоты (ВСНБ) можно отличить от аутосомных форм по наличию близорукости , которая обычно отсутствует при аутосомных формах. У пациентов с ЦСНБ часто наблюдаются нарушения ночного зрения, близорукость , снижение остроты зрения , косоглазие и нистагм . Лица с полной формой CSNB (CSNB1) имеют сильно нарушенную чувствительность палочек (снижение примерно в 300 раз), а также дисфункцию колбочек . У пациентов с неполной формой может наблюдаться как близорукость, так и дальнозоркость . [ 8 ]
Причина
[ редактировать ]CSNB вызван сбоями в нейротрансмиссии от фоторецепторов палочек и колбочек к биполярным клеткам сетчатки. [ 9 ] В этом первом синапсе информация от фоторецепторов разделяется на два канала: ВКЛ и ВЫКЛ. Путь ВКЛ обнаруживает появление света, а путь ВЫКЛ обнаруживает смещение света. [ 10 ] Нарушения в CSNB1 специфически влияют на путь ON, препятствуя способности биполярных клеток ON-типа обнаруживать нейромедиаторы, высвобождаемые из фоторецепторов. [ 9 ] Палочки, отвечающие за зрение при слабом освещении, вступают в контакт только с биполярными клетками ON-типа, тогда как колбочки, отвечающие за зрение при ярком свете, вступают в контакт с биполярными клетками обоих подтипов ON и OFF. [ 11 ] Поскольку чувствительные палочки при слабом освещении питаются только по пути ВКЛ, у людей с CSNB1 обычно возникают проблемы с ночным зрением, в то время как зрение в хорошо освещенных условиях сохраняется. [ 9 ] При CSNB2 высвобождение нейромедиатора из фоторецепторов нарушается, что приводит к вовлечению путей включения и выключения.
Электроретинограмма ( ЭРГ ) является важным инструментом диагностики ЦСНБ. a-волна ЭРГ, которая отражает функцию каскада фототрансдукции в ответ на световые вспышки, обычно нормальна у пациентов с CSNB, хотя в некоторых случаях также нарушается фототрансдукция, что приводит к уменьшению a-волны. b-волна ЭРГ, которая в первую очередь отражает функцию ON-биполярных клеток, значительно снижена в случаях CSNB2 и полностью отсутствует в случаях CSNB1. [ 9 ] [ 12 ]
Генетика
[ редактировать ]только три мутации родопсина , связанные с врожденной стационарной куриной слепотой (ВСНБ). Было обнаружено [ 13 ] Две из этих мутаций обнаружены во второй трансмембранной спирали родопсина в Gly-90 и Thr-94. В частности, эти мутации представляют собой Gly90Asp. [ 14 ] и Thr94Ile, о котором сообщалось последним. [ 15 ] Третья мутация — Ala292Glu, она расположена в седьмой трансмембранной спирали , вблизи места прикрепления Lys-296 к сетчатке. [ 16 ] Мутации, связанные с CSNB, затрагивают аминокислотные остатки вблизи связи протонированного основания Шиффа (PSB). Они связаны с изменением конформационной стабильности и протонированного статуса азота ПСБ. [ 17 ]
Патофизиология
[ редактировать ]ЦСНБ1
[ редактировать ]Полная форма Х-сцепленной врожденной стационарной куриной слепоты, также известная как никталопия , вызвана мутациями в гене NYX (никталопин на Х-хромосоме ), который кодирует небольшой белок семейства богатых лейцином повторов (LRR) с неизвестной функцией. [ 18 ] [ 19 ] Этот белок состоит из N-концевого сигнального пептида и 11 LRR (LRR1-11), окруженных богатыми цистеином LRR (LRRNT и LRRCT). На С-конце белка имеется предполагаемый сайт якоря GPI . Хотя функция NYX еще полностью не изучена, считается, что он локализован внеклеточно. Естественная делеция 85 оснований в NYX у некоторых мышей приводит к фенотипу «nob» (отсутствие b-волны), который очень похож на фенотип, наблюдаемый у пациентов с CSNB1. [ 20 ] NYX экспрессируется преимущественно в палочках и колбочках сетчатки. В настоящее время известно почти 40 мутаций NYX, связанных с CSNB1 (табл. 1), расположенных по всему белку. Поскольку функция белка никталопина неизвестна, эти мутации не были дополнительно охарактеризованы. Однако прогнозируется, что многие из них приведут к укороченным белкам, которые, предположительно, нефункциональны.
| Мутация | Позиция | Ссылки | |
|---|---|---|---|
| Нуклеотид | Аминокислота | ||
| с.?-1_?-61дел | 1_20дел | Последовательность сигналов | [ 19 ] |
| Сращивание | Интрон 1 | [ 21 ] | |
| c.?-63_1443-?del | 21_481 часть | [ 19 ] | |
| c.48_64del | L18RfsX108 | Последовательность сигналов | [ 21 ] |
| c.85_108del | R29_A36del | N-концевой LRR | [ 18 ] |
| c.G91C | C31S | ЛРРНТ | [ 19 ] |
| c.C105A | C35X | ЛРРНТ | [ 19 ] |
| ок.C169A | P57T | ЛРРНТ | [ 22 ] |
| ок.C191A | А64Е | ЛРР1 | [ 22 ] |
| c.G281C | Р94П | ЛРР2 | [ 23 ] |
| c.301_303del | на I101s | ЛРР2 | [ 19 ] |
| c.T302C | И101Т | ЛРР2 | [ 23 ] |
| c.340_351del | E114_A118del | ЛРР3 | [ 19 ] [ 21 ] |
| c.G427C | А143П | ЛРР4 | [ 19 ] |
| c.C452T | П151Л | ЛРР4 | [ 18 ] |
| c.464_465insAGCGTGCCCCGAGCGCCTCCTG | S149_V150dup+P151_L155dup | ЛРР4 | [ 18 ] |
| c.C524G | P175R | ЛРР5 | [ 19 ] |
| c.T551C | Л184П | ЛРР6 | [ 18 ] |
| c.556_618делинс | H186?fsX260 | ЛРР6 | [ 18 ] |
| c.559_560delinsAA | А187К | ЛРР6 | [ 19 ] |
| c.613_621dup | 205_207дуп | ЛРР7 | [ 18 ] [ 19 ] |
| ок.628_629 дюймов | R209_S210insCLR | ЛРР7 | [ 18 ] |
| c.T638A | L213Q | ЛРР7 | [ 18 ] |
| ок.A647G | Н216С | ЛРР7 | [ 18 ] [ 21 ] |
| c.T695C | Л232П | ЛРР8 | [ 18 ] |
| ок.727_738del | 243_246del | ЛРР8 | [ 19 ] |
| c.C792G | Н264К | ЛРР9 | [ 18 ] |
| c.T854C | Л285П | ЛРР10 | [ 18 ] |
| c.T893C | F298S | ЛРР10 | [ 18 ] |
| c.C895T | Q299X | ЛРР10 | [ 21 ] |
| c.T920C | L307P | ЛРР11 | [ 19 ] |
| c.A935G | N312S | ЛРР11 | [ 19 ] |
| c.T1040C | L347P | ЛРРКТ | [ 19 ] |
| c.G1049A | W350X | ЛРРКТ | [ 18 ] |
| c.G1109T | Г370В | ЛРРКТ | [ 19 ] |
| ок.1122_1457дел | S374RfsX383 | ЛРРКТ | [ 19 ] [ 21 ] |
| ок.1306del | L437WfsX559 | C-терминал | [ 21 ] |
| LRR: повтор, богатый лейцином , LRRNT и LRRCT: N- и C-концевые LRR, богатые цистеином. | |||
ЦСНБ2
[ редактировать ]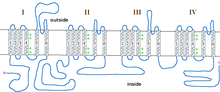
Неполная форма врожденной стационарной куриной слепоты, сцепленной с Х-хромосомой (CSNB2), вызвана мутациями в гене CACNA1F, который кодирует потенциалзависимый кальциевый канал Ca V 1.4, активно экспрессируемый в сетчатке . [ 24 ] [ 25 ] Одним из важных свойств этого канала является то, что он инактивируется с чрезвычайно низкой скоростью. Это позволяет ему производить устойчивый Ca 2+ вход при деполяризации. Поскольку фоторецепторы деполяризуются в отсутствие света, каналы Ca V 1.4 обеспечивают устойчивое высвобождение нейромедиатора при деполяризации. [ 26 ] Это было продемонстрировано на мутантных мышах CACNA1F, у которых заметно снижены сигналы кальция фоторецепторов. [ 27 ] В настоящее время существует 55 мутаций CACNA1F, расположенных по всему каналу, Таблица 2 и Рисунок 1. Хотя большинство этих мутаций приводят к усечению и, вероятно, нефункциональным каналам, ожидается, что они предотвращают способность света гиперполяризовать фоторецепторы. Из мутаций с известными функциональными последствиями четыре производят каналы, которые либо полностью нефункциональны, а две приводят к образованию каналов, открывающихся при гораздо более гиперполяризованных потенциалах, чем у дикого типа. Это приведет к тому, что фоторецепторы продолжат высвобождать нейромедиаторы даже после гиперполяризации, вызванной светом.
| Мутация | Позиция | Эффект | Ссылки | |
|---|---|---|---|---|
| Нуклеотид | Аминокислота | |||
| c.C148T | Р50Х | N-конец | [ 28 ] | |
| c.151_155делАГААА | R51PfsX115 | N-конец | [ 29 ] | |
| c.T220C | C74R | N-конец | [ 29 ] | |
| c.C244T | R82X | N-конец | [ 28 ] [ 29 ] | |
| c.466_469delinsGTAGGGGTGCT CCACCCCGTAGGGGTGCTCCACC |
С156ВделШпилькиГВХОВГВЛХ | Д1С2-3 | [ 28 ] [ 30 ] [ 31 ] | |
| Сращивание | Интрон 4 | [ 28 ] | ||
| c.T685C | С229П | Д1С4-5 | [ 29 ] | |
| c.G781A | G261R | D1-пора | [ 29 ] | |
| c.G832T | E278X | D1-пора | [ 21 ] [ 32 ] | |
| c.904insG | R302AfsX314 | D1-пора | [ 30 ] | |
| c.951_953delCTT | F318del | D1-пора | [ 28 ] | |
| c.G1106A | G369D | Д1С6 | Активирует ~20 мВ более отрицательно, чем дикий тип, увеличивает время достижения пикового тока и уменьшает инактивацию, увеличивает Ca 2+ проницаемость. | [ 24 ] [ 26 ] [ 28 ] [ 29 ] [ 33 ] |
| c.1218delC | W407GfsX443 | Д1-2 | [ 25 ] [ 28 ] [ 32 ] | |
| c.C1315T | Q439X | Д1-2 | [ 29 ] | |
| c.G1556A | Р519К | Д1-2 | Снижение экспрессии | [ 24 ] [ 34 ] |
| ок.C1873T | Р625Х | Д2С4 | [ 28 ] [ 29 ] | |
| c.G2021A | G674D | Д2С5 | [ 26 ] [ 28 ] [ 30 ] | |
| c.C2071T | Р691Х | D2-пора | [ 22 ] | |
| c.T2258G | F753C | Д2С6 | [ 29 ] | |
| c.T2267C | I756T | Д2С6 | Активирует на ~35 мВ более отрицательно, чем дикий тип, инактивирует медленнее. | [ 35 ] |
| Сращивание | Интрон 19 | [ 29 ] | ||
| c.T2579C | L860P | Д2-3 | [ 29 ] | |
| c.C2683T | Р895Х | Д3С1-2 | [ 21 ] [ 22 ] [ 25 ] [ 28 ] | |
| Сращивание | Интрон 22 | [ 29 ] [ 30 ] | ||
| Сращивание | Интрон 22 | [ 29 ] | ||
| c.C2783A | А928Д | Д3С2-3 | [ 26 ] [ 28 ] | |
| c.C2905T | Р969Х | Д3С4 | [ 24 ] [ 29 ] | |
| c.C2914T | Р972Х | Д3С4 | [ 32 ] | |
| Сращивание | Интрон24 | [ 28 ] | ||
| c.C2932T | Р978Х | Д3С4 | [ 30 ] | |
| c.3006_3008delCAT | I1003del | Д3С4-5 | [ 28 ] | |
| c.G3052A | G1018R | Д3С5 | [ 29 ] | |
| c.3125delG | G1042AfsX1076 | D3-пора | [ 28 ] | |
| c.3166insC | L1056PfsX1066 | D3-пора | [ 24 ] [ 25 ] [ 28 ] [ 29 ] | |
| c.C3178T | Р1060Вт | D3-пора | [ 24 ] [ 29 ] | |
| c.T3236C | L1079P | D3-пора | Не открывается без BayK, активирует на ~5 мВ больше отрицательного, чем дикий тип | [ 29 ] [ 33 ] |
| c.3672delC | L1225SfsX1266 | Д4С2 | [ 25 ] [ 28 ] | |
| c.3691_3702del | G1231_T1234del | Д4С2 | [ 24 ] [ 29 ] | |
| c.G3794T | S1265I | Д4С3 | [ 22 ] | |
| c.C3886A | Р1296С | Д4С4 | [ 22 ] | |
| c.C3895T | Р1299Х | Д4С4 | [ 25 ] [ 28 ] [ 29 ] | |
| Сращивание | Интрон 32 | [ 29 ] | ||
| c.C4075T | Q1359X | D4-пора | [ 24 ] [ 29 ] | |
| в. Т4124А | L1375H | D4-пора | Снижение экспрессии | [ 24 ] [ 29 ] [ 34 ] |
| Сращивание | Интрон 35 | [ 29 ] | ||
| c.G4353A | W1451X | C-терминал | Нефункциональный | [ 25 ] [ 26 ] [ 28 ] [ 33 ] |
| c.T4495C | C1499R | C-терминал | [ 29 ] | |
| c.C4499G | П1500Р | C-терминал | [ 29 ] | |
| c.T4523C | L1508P | C-терминал | [ 29 ] | |
| Сращивание | интрон 40 | [ 28 ] | ||
| c.4581delC | F1528LfsX1535 | C-терминал | [ 36 ] | |
| c.A4804T | К1602Х | C-терминал | [ 24 ] [ 29 ] | |
| c.C5479T | Р1827Х | C-терминал | [ 29 ] | |
| c.5663delG | S1888TfsX1931 | C-терминал | [ 28 ] | |
| c.G5789A | Р1930Х | C-терминал | [ 22 ] | |
Диагностика
[ редактировать ]Этот раздел пуст. Вы можете помочь, добавив к нему . ( июнь 2022 г. ) |
Куриная слепота является симптомом у многих пациентов, и диагноз часто осуществляется с помощью различных тестов, включая электроретинограмму, чтобы выявить любые нарушения сетчатки «в целом». [ 37 ] [ 38 ] [ 39 ] Проведенные тесты могут также включать исследование поля зрения, исследование глазного дна и микроскопию с щелевой лампой в дополнение к измерениям, предоставляемым электроретинограммой (ЭРГ). [ 40 ] [ 41 ] [ 42 ]
Сноски
[ редактировать ]- ^ Бай, Дунъэ; Го, Руру; Хуан, Даньдан; Цзи, Цзянь; Лю, Вэй (15 марта 2024 г.). «Сложные гетерозиготные мутации в GRM6, вызывающие полную врожденную стационарную куриную слепоту типа Шуберта-Борншейна» . Гелион . 10 (5): e27039. Бибкод : 2024Heliy..1027039B . дои : 10.1016/j.heliyon.2024.e27039 . ISSN 2405-8440 . ПМЦ 10907788 . ПМИД 38434377 .
- ^ Зейтц, Кристина; Робсон, Энтони Г.; Аудо, Изабель (01 марта 2015 г.). «Врожденная стационарная куриная слепота: анализ и обновление генотип-фенотипических корреляций и патогенетических механизмов» . Прогресс в исследованиях сетчатки и глаз . 45 : 58–110. doi : 10.1016/j.preteyeres.2014.09.001 . ISSN 1350-9462 . ПМИД 25307992 .
- ^ Беллоне Р.Р., Холл Х., Сеталури В., Деви С., Маддоди Н., Арчер С. и др. (22 октября 2013 г.). «Доказательства того, что ретровирусная вставка в TRPM1 является причиной врожденной стационарной куриной слепоты и леопардовой пятнистости у лошадей» . ПЛОС ОДИН . 8 (10): е78280. Бибкод : 2013PLoSO...878280B . дои : 10.1371/journal.pone.0078280 . ПМЦ 3805535 . ПМИД 24167615 .
- ^ Хак Ю.Л., Крэбтри Э.Э., Авила Ф., Саттон Р.Б., Гран Р., О А. и др. (март 2021 г.). «Полногеномное секвенирование идентифицирует миссенс-мутацию в GRM6 как вероятную причину врожденной стационарной куриной слепоты у прогулочной лошади из Теннесси» . Ветеринарный журнал лошадей . 53 (2): 316–323. дои : 10.1111/evj.13318 . ПМИД 32654228 . S2CID 220500585 .
- ^ Дас Р.Г., Беккер Д., Джаганнатан В., Гольдштейн О., Сантана Э., Карлин К. и др. (октябрь 2019 г.). «Полногеномное исследование ассоциаций и полногеномное секвенирование выявили делецию в LRIT3, связанную с врожденной стационарной куриной слепотой у собак» . Научные отчеты . 9 (1): 14166. Бибкод : 2019NatSR...914166D . дои : 10.1038/s41598-019-50573-7 . ПМК 6775105 . ПМИД 31578364 .
- ^ «Панель 2 Аппалузы | Лаборатория ветеринарной генетики» . vgl.ucdavis.edu . Проверено 11 октября 2022 г.
- ^ «Врожденная стационарная куриная слепота (CSNB2) у шагающих лошадей Теннесси | Лаборатория ветеринарной генетики» . vgl.ucdavis.edu . Проверено 11 октября 2022 г.
- ^ Бойкот К.М., Пирс В.Г., Мусарелла М.А., Велебер Р.Г., Мэйбаум Т.А., Берч Д.Г. и др. (апрель 1998 г.). «Доказательства генетической гетерогенности при Х-сцепленной врожденной стационарной куриной слепоте» . Американский журнал генетики человека . 62 (4): 865–875. дои : 10.1086/301781 . ПМК 1377021 . ПМИД 9529339 .
- ^ Перейти обратно: а б с д Зейтц С., Робсон АГ, Аудо I (март 2015 г.). «Врожденная стационарная куриная слепота: анализ и обновление генотип-фенотипических корреляций и патогенетических механизмов». Прогресс в исследованиях сетчатки и глаз . 45 : 58–110. doi : 10.1016/j.preteyeres.2014.09.001 . ПМИД 25307992 . S2CID 45696921 .
- ^ Эйлер Т., Хаверкамп С., Шуберт Т., Баден Т. (август 2014 г.). «Биполярные клетки сетчатки: элементарные строительные блоки зрения». Обзоры природы. Нейронаука . 15 (8): 507–519. дои : 10.1038/nrn3783 . ПМИД 25158357 . S2CID 16309488 .
- ^ Данн Ф.А., Вонг Р.О. (ноябрь 2014 г.). «Модели проводки в сетчатке мыши: сбор данных через коннектом, физиология и световая микроскопия» . Журнал физиологии . 592 (22): 4809–4823. дои : 10.1113/jphysicalol.2014.277228 . ПМЦ 4259528 . ПМИД 25172948 .
- ^ Аудо И, Робсон А.Г., Холдер Г.Е., Мур А.Т. (2008). «Отрицательная ЭРГ: клинические фенотипы и механизмы развития внутренней дисфункции сетчатки». Обзор офтальмологии . 53 (1): 16–40. doi : 10.1016/j.survophthal.2007.10.010 . ПМИД 18191655 .
- ^ Гаррига П., Маньоса Дж. (сентябрь 2002 г.). «Белок фоторецептора глаза родопсин. Структурные последствия заболеваний сетчатки». Письма ФЭБС . 528 (1–3): 17–22. дои : 10.1016/s0014-5793(02)03241-6 . ПМИД 12297272 . S2CID 41860711 .
- ^ Рао В.Р., Коэн ГБ, Оприан Д.Д. (февраль 1994 г.). «Мутация родопсина G90D и молекулярный механизм врожденной куриной слепоты». Природа . 367 (6464): 639–42. Бибкод : 1994Natur.367..639R . дои : 10.1038/367639a0 . ПМИД 8107847 . S2CID 4311079 .
- ^ Н. аль-Джандал, Г. Дж. Фаррар, А. С. Кианг, М. М. Хамфрис, Н. Бэннон, Дж. Б. Финдли, П. Хамфрис и П. Ф. Кенна Хум. Мутат. 13 (1999), стр. 75–81.
- ^ Дрия Т.П., Берсон Э.Л., Рао В.Р., Оприан Д.Д. (июль 1993 г.). «Гетерозиготная миссенс-мутация гена родопсина как причина врожденной стационарной куриной слепоты». Природная генетика . 4 (3): 280–3. дои : 10.1038/ng0793-280 . ПМИД 8358437 . S2CID 7682929 .
- ^ Просеивание П.А., Ричардс Дж.Э., Наарендорп Ф., Бингхэм Э.Л., Скотт К., Альперн М. (январь 1995 г.). «Тьма-свет: модель куриной слепоты из-за мутации человеческого родопсина Gly-90 → Asp» . Труды Национальной академии наук Соединенных Штатов Америки . 92 (3): 880–4. Бибкод : 1995PNAS...92..880S . дои : 10.1073/pnas.92.3.880 . ПМК 42724 . ПМИД 7846071 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот Бек-Хансен Н.Т., Нейлор М.Дж., Мэйбаум Т.А., Спаркс Р.Л., Куп Б., Берч Д.Г. и др. (ноябрь 2000 г.). «Мутации в NYX, кодирующем богатый лейцином протеогликан никталопин, вызывают Х-сцепленную полную врожденную стационарную куриную слепоту». Природная генетика . 26 (3): 319–323. дои : 10.1038/81619 . ПМИД 11062471 . S2CID 10223880 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д Пуш С.М., Цейтц С., Брандау О., Пеш К., Ахац Х., Фейл С. и др. (ноябрь 2000 г.). «Полная форма Х-сцепленной врожденной стационарной куриной слепоты вызвана мутациями в гене, кодирующем богатый лейцином повторяющийся белок». Природная генетика . 26 (3): 324–327. дои : 10.1038/81627 . ПМИД 11062472 . S2CID 42428370 .
- ^ Грегг Р.Г., Мухопадьяй С., Кандилль С.И., Болл С.Л., Пардью М.Т., Макколл М.А., Пичи Н.С. (январь 2003 г.). «Идентификация гена и мутации, ответственной за фенотип мышиного nob» . Исследовательская офтальмология и визуальные науки . 44 (1): 378–384. дои : 10.1167/iovs.02-0501 . ПМИД 12506099 .
- ^ Перейти обратно: а б с д и ж г час я Зито И., Аллен Л.Е., Патель Р.Дж., Мейндл А., Брэдшоу К., Йейтс Дж.Р. и др. (февраль 2003 г.). «Мутации в генах CACNA1F и NYX в британских семьях CSNBX». Человеческая мутация . 21 (2): 169. doi : 10.1002/humu.9106 . ПМИД 12552565 . S2CID 13143864 .
- ^ Перейти обратно: а б с д и ж г Зейтц С., Минотти Р., Фейл С., Матьяс Г., Кремерс Ф.П., Хойнг С.Б., Бергер В. (март 2005 г.). «Новые мутации CACNA1F и NYX в голландских семьях с Х-сцепленной врожденной стационарной куриной слепотой». Молекулярное видение . 11 : 179–183. ПМИД 15761389 .
- ^ Перейти обратно: а б Сяо X, Цзя X, Го X, Ли С, Ян Цз, Чжан Ц (2006). «CSNB1 в китайских семьях связан с новыми мутациями в NYX» . Журнал генетики человека . 51 (7): 634–640. дои : 10.1007/s10038-006-0406-5 . ПМИД 16670814 .
- ^ Перейти обратно: а б с д и ж г час я дж Стром Т.М., Ньякатура Г., Апфельштедт-Силла Е., Хеллебранд Х., Лоренц Б., Вебер Б.Х. и др. (июль 1998 г.). «Ген кальциевого канала L-типа мутировал при неполной Х-сцепленной врожденной стационарной куриной слепоте». Природная генетика . 19 (3): 260–263. дои : 10.1038/940 . ПМИД 9662399 . S2CID 34467174 .
- ^ Перейти обратно: а б с д и ж г Бек-Хансен Н.Т., Нейлор М.Дж., Мэйбаум Т.А., Пирс В.Г., Куп Б., Фишман Г.А. и др. (июль 1998 г.). «Мутации потери функции в гене альфа1-субъединицы кальциевого канала в Xp11.23 вызывают неполную Х-сцепленную врожденную стационарную куриную слепоту». Природная генетика . 19 (3): 264–267. дои : 10.1038/947 . ПМИД 9662400 . S2CID 42480901 .
- ^ Перейти обратно: а б с д и МакРори Дж.Э., Хамид Дж., Деринг С.Дж., Гарсия Э., Паркер Р., Хэмминг К. и др. (февраль 2004 г.). «Ген CACNA1F кодирует кальциевый канал L-типа с уникальными биофизическими свойствами и распределением в тканях» . Журнал неврологии . 24 (7): 1707–1718. doi : 10.1523/JNEUROSCI.4846-03.2004 . ПМК 6730460 . ПМИД 14973233 .
- ^ Мансерг Ф., Ортон Н.К., Весси Дж.П., Лалонд М.Р., Стелл В.К., Трембле Ф. и др. (октябрь 2005 г.). «Мутация гена кальциевого канала Cacna1f нарушает передачу сигналов кальция, синаптическую передачу и клеточную организацию в сетчатке мыши» . Молекулярная генетика человека . 14 (20): 3035–3046. дои : 10.1093/hmg/ddi336 . ПМИД 16155113 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т Бойкот К.М., Мэйбаум Т.А., Нейлор М.Дж., Велебер Р.Г., Робитайл Дж., Мияке Ю. и др. (февраль 2001 г.). «Сводка 20 мутаций CACNA1F, выявленных в 36 семьях с неполной Х-сцепленной врожденной стационарной куриной слепотой, и характеристика вариантов сплайсинга». Генетика человека . 108 (2): 91–97. дои : 10.1007/s004390100461 . ПМИД 11281458 . S2CID 2844173 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и Вутц К., Зауэр С., Цреннер Е., Лоренц Б., Алитало Т., Брогхаммер М. и др. (август 2002 г.). «Тридцать различных мутаций CACNA1F в 33 семьях с неполным типом профиля экспрессии XLCSNB и Cacna1f в сетчатке мыши» . Европейский журнал генетики человека . 10 (8): 449–456. дои : 10.1038/sj.ejhg.5200828 . ПМИД 12111638 .
- ^ Перейти обратно: а б с д и Накамура М., Ито С., Терасаки Х., Мияке Ю. (июнь 2001 г.). «Новые мутации CACNA1F у японских пациентов с неполной врожденной стационарной куриной слепотой». Исследовательская офтальмология и визуальные науки . 42 (7): 1610–1616. ПМИД 11381068 .
- ^ Накамура М., Ито С., Пяо Ч., Терасаки Х., Мияке Ю. (июль 2003 г.). «Атрофия сетчатки и диска зрительного нерва, связанная с мутацией CACNA1F в японской семье». Архив офтальмологии . 121 (7): 1028–1033. дои : 10.1001/archopht.121.7.1028 . ПМИД 12860808 .
- ^ Перейти обратно: а б с Аллен Л.Е., Зито И., Брэдшоу К., Патель Р.Дж., Бёрд А.С., Фитцке Ф. и др. (ноябрь 2003 г.). «Корреляция генотипа-фенотипа в британских семьях с Х-сцепленной врожденной стационарной куриной слепотой» . Британский журнал офтальмологии . 87 (11): 1413–1420. дои : 10.1136/bjo.87.11.1413 . ПМК 1771890 . ПМИД 14609846 .
- ^ Перейти обратно: а б с Хода Дж.К., Загетто Ф., Кошак А., Стрессниг Дж. (январь 2005 г.). «Врожденная стационарная куриная слепота 2-го типа мутации S229P, G369D, L1068P и W1440X изменяют ворота каналов или функциональную экспрессию Ca2+-каналов Ca(v)1.4 L-типа» . Журнал неврологии . 25 (1): 252–259. doi : 10.1523/JNEUROSCI.3054-04.2005 . ПМК 6725195 . ПМИД 15634789 .
- ^ Перейти обратно: а б Хода Дж.К., Загетто Ф., Сингх А., Кошак А., Стрессниг Дж. (март 2006 г.). «Влияние врожденной стационарной куриной слепоты типа 2 мутаций R508Q и L1364H на функцию и экспрессию Ca2+-канала Cav1.4 L-типа». Журнал нейрохимии . 96 (6): 1648–1658. дои : 10.1111/j.1471-4159.2006.03678.x . ПМИД 16476079 . S2CID 25987619 .
- ^ Хемара-Вахануи А., Берджуков С., Хоуп С.И., Дирден П.К., Ву С.Б., Уилсон-Уилер Дж. и др. (май 2005 г.). «Мутация CACNA1F, выявленная при Х-сцепленном заболевании сетчатки, сдвигает зависимость активации канала Cav1.4 от напряжения» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (21): 7553–7558. Бибкод : 2005PNAS..102.7553H . дои : 10.1073/pnas.0501907102 . ПМК 1140436 . ПМИД 15897456 .
- ^ Якоби Ф.К., Хамель К.П., Арно Б., Блин Н., Брогхаммер М., Якоби ПК. и др. (май 2003 г.). «Новая мутация CACNA1F во французской семье с неполным типом Х-сцепленной врожденной стационарной куриной слепоты». Американский журнал офтальмологии . 135 (5): 733–736. дои : 10.1016/S0002-9394(02)02109-8 . ПМИД 12719097 .
- ^ Риггс, Лоррин А. (1 июля 1954 г.). «Электроретинография при куриной слепоте*» . Американский журнал офтальмологии . 38 (1): 70–78. дои : 10.1016/0002-9394(54)90011-2 . ISSN 0002-9394 . ПМИД 13180620 .
- ^ Михаэлидис, Мишель; Холдер, Грэм Э; Мур, Энтони Т. (2017), «Наследственные заболевания сетчатки» , Детская офтальмология и косоглазие Тейлора и Хойта , Elsevier, стр. 462–486.e2, doi : 10.1016/b978-0-7020-6616-0.00046-3 , ISBN 978-0-7020-6616-0 , получено 22 июля 2024 г.
- ^ Хендерсон, Роберт Х. (01 января 2020 г.). «Наследственные дистрофии сетчатки» . Педиатрия и здоровье детей . 30 (1): 19–27. дои : 10.1016/j.paed.2019.10.004 . ISSN 1751-7222 .
- ^ Бай, Дунъэ; Го, Руру; Хуан, Даньдан; Цзи, Цзянь; Лю, Вэй (15 марта 2024 г.). «Сложные гетерозиготные мутации в GRM6, вызывающие полную врожденную стационарную куриную слепоту типа Шуберта-Борншейна» . Гелион . 10 (5): e27039. Бибкод : 2024Heliy..1027039B . дои : 10.1016/j.heliyon.2024.e27039 . ISSN 2405-8440 . ПМЦ 10907788 . ПМИД 38434377 .
- ^ Шуберт, Г.; Борншайн, Х. (18 марта 2010 г.). «Вклад в анализ электроретинограммы человека» . Офтальмологика . 123 (6): 396–413. дои : 10.1159/000301211 . ISSN 0030-3755 . ПМИД 14957416 .
- ^ Риггс, Лоррин А. (1 июля 1954 г.). «Электроретинография при куриной слепоте*» . Американский журнал офтальмологии . 38 (1): 70–78. дои : 10.1016/0002-9394(54)90011-2 . ISSN 0002-9394 . ПМИД 13180620 .