Цикл Кальвина
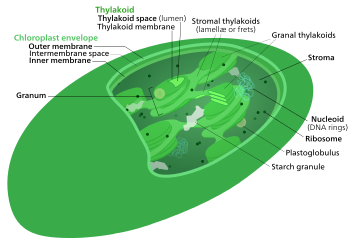
Цикл Кальвина , независимые от света реакции , фаза , темные реакции или фотосинтетическое восстановление углерода ( ПЦР ) био- синтетическая [ 1 ] фотосинтеза - это серия химических реакций, которые преобразуют диоксид углерода и водородные соединения в глюкозу . Цикл Кальвина присутствует во всех фотосинтетических эукариотах, а также во многих фотосинтетических бактериях. У растений эти реакции встречаются в строме , заполненная жидкостью область хлоропласта за пределами тилакоидных мембран . Эти реакции принимают продукты ( АТФ и NADPH ) светозависимых реакций и выполняют дальнейшие химические процессы на них. Цикл Кальвина использует химическую энергию АТФ и снижающую мощность NADPH от реакций, зависящих от света для получения сахара для использования растения. Эти субстраты используются в серии реакций снижения окисления ( окислительно-восстановительного ) для получения сахара в пошаговом процессе; Не существует прямой реакции, которая превращает несколько молекул CO 2 в сахар. Существует три фазы на светозависимые реакции, которые в совокупности называют циклом Кальвина: карбоксилирование , реакции восстановления и регенерация 1,5-бисфосфата (рубла).
Хотя это также называется «темной реакцией», цикл Кальвина на самом деле не происходит в темноте или в ночное время. Это связано с тем, что процесс требует NADPH , который является недолгим и исходит от светозависимых реакций . В темноте растения вместо этого высвобождают сахарозу в флоэму из своих запасов крахмала , чтобы обеспечить энергию для растения. Таким образом, цикл Кальвина происходит, когда свет доступен независимо от вида фотосинтеза ( C3, фиксация углерода , фиксация углерода C4 и метаболизм кислотной кислоты (CAM) ); Камуляционные растения хранят летовому кислоте в вакуолях каждую ночь и днем выпускают ее, чтобы заставить этот процесс работать. [ 2 ]
Связь с другими метаболическими путями
[ редактировать ]Реакции цикла Кальвина тесно связаны с транспортной цепью тилакоида , [ 3 ] В качестве энергии, необходимой для уменьшения углекислого газа, обеспечивается NADPH, полученным во время реакций зависимых от света . Процесс фотопороширования , также известный как цикл C2, также связан с циклом Кальвина, поскольку он является результатом альтернативной реакции фермента Rubisco , а его окончательный побочный продукт-еще одна глицеральдегида-3-p молекула .
Цикл Кальвина
[ редактировать ]В этом разделе нужны дополнительные цитаты для проверки . ( декабрь 2023 г. ) |

Цикл Кальвина , цикл Кальвина -Бенсон -Бассам (CBB) восстановительный пентозофосфатный цикл (цикл RPP) или цикл C3 представляет собой серию биохимических окислительно -восстановительных реакций, которые происходят в строме хлоропластов , в фотосинтетических организмах . Цикл был обнаружен в 1950 году Мелвин Кальвин , Джеймс Бассм и Эндрю Бенсон в Калифорнийском университете, Беркли [ 4 ] Используя радиоактивный изотоп углерод-14 . [ Цитация необходима ]
Фотосинтез происходит на двух этапах в ячейке. молекулы энергии На первом этапе светозависимые реакции захватывают энергию света и используют его, чтобы сделать АТФ носителя умеренной энергии и водородного . Цикл Кальвина использует эти соединения для преобразования углекислого газа и воды в органические соединения [ 5 ] Это может быть использовано организмом (и животными, которые питаются им). Этот набор реакций также называется углеродной фиксацией . Ключевой фермент цикла называется Rubisco . В следующих биохимических уравнениях химические виды (фосфаты и карбоновые кислоты) существуют в равновесиях между их различными ионизированными состояниями, как это управляется pH . [ Цитация необходима ]
Ферменты в цикле Кальвина функционально эквивалентны большинству ферментов, используемых в других метаболических путях, таких как глюконеогенез и пентозофосфатный путь , но ферменты в цикле Кальвина обнаружены в строме хлоропластов вместо клеточного цитозоля , разделяющие реакции. Они активированы в свете (именно поэтому название «темная реакция» вводит в заблуждение), а также продуктами светозависимой реакции. Эти регуляторные функции предотвращают дыхание цикла Кальвина к углекислым газе. Энергия (в форме АТФ) будет потрачена впустую на выполнение этих реакций, когда у них нет чистой производительности . [ Цитация необходима ]
Сумма реакций в цикле Кальвина является следующей: [ Цитация необходима ]
- 3 co
2 + 6 NADPH + 9 ATP + 5 ч
2 O → глицеральдегид-3-фосфат (G3P) + 6 NADP + + 9 ADP + 8 p i (p i = неорганический фосфат )
Гексоза (шестиуглеродные) сахары не являются продуктами цикла Кальвина. Хотя многие тексты перечисляют продукт фотосинтеза как c
6 часов
12 o
6 , это в основном для удобства, чтобы соответствовать уравнению аэробного дыхания , где в митохондриях окисляется шестиуглеродные сахары. Углеводные продукты цикла Кальвина представляют собой трехуглеродные молекулы фосфата сахара или «триозофосфаты», а именно глицеральдегид-3-фосфат (G3P). [ Цитация необходима ]
Шаги
[ редактировать ]На первой стадии цикла Кальвина молекула CO 2 включена в одну из двух трехглеродных молекул ( глицеральдегид 3-фосфат или G3P), где используется две молекулы ATP и две молекулы NADPH , которые были получены На светозависимой стадии. Три шага связаны: [ Цитация необходима ]
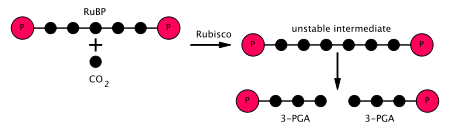

- Фермент Rubisco катализирует карбоксилирование рибулозы-1,5-бисфосфата , RUBP, 5-углеродного соединения, диоксидом углерода (всего 6 углерода) в двухэтапной реакции. [ 6 ] Продуктом первого этапа является энедиол-энзимный комплекс, который может захватить CO
2 или о
2 Таким образом, энедиол-энзимный комплекс является настоящей карбоксилазой/оксигеназой. Ко
2 , который фиксируется Enediol на втором этапе, создает нестабильное шестиуглеродистое соединение, называемое 2-карбокси 3-кето 1,5-бифосфориботол (CKABP [ 7 ] ) (или 3-кето-2-карбоксиарабинитол 1,5-бисфосфат), который немедленно рассыпается на 2 молекулы 3-фосфоглицерата (также написано как 3-фосфоглицельная кислота, PGA, 3PGA или 3-PGA), 3-углеродное соединение Полем [ 8 ] - Фермент фосфоглицерат киназа катализирует фосфорилирование 3-PGA с помощью АТФ (которая была получена на светозависимой стадии). 1,3-бисфосфоглицерат (глицерат-1,3-бисфосфат) и ADP являются продуктами. (Однако обратите внимание, что два 3-PGA производятся для каждого CO
2 , который входит в цикл, поэтому этот шаг использует два ATP на CO
2 фиксировано.) [ Цитация необходима ] - Фермент глицеральдегид 3-фосфатдегидрогеназы катализирует восстановление 1,3BPGA с помощью NADPH (что является еще одним продуктом светозависимой стадии). Глицеральдегид 3-фосфат (также называемый G3P, GP, TP, PGAL, GAP) получен + Полем Опять же, два NADPH используются для CO CO
2 исправлено. [ Цитация необходима ]

Следующий этап в цикле Кальвина - регенерировать Rubp. Пять молекул G3P продуцируют три молекулы RUBP, используя три молекулы АТФ. Поскольку каждая молекула CO 2 производит две молекулы G3P, три молекулы CO 2 продуцируют шесть молекул G3P, из которых пять используются для регенерации RUBP, оставляя чистый прирост одной молекулы G3P на три молекулы CO 2 (как и ожидалось от количества Атомы углерода вовлечены). [ Цитация необходима ]


Стадия регенерации может быть разбита на серию шагов.
- Триозофосфат изомераза превращает один из G3P обратимо в дигидроксиацетонфосфат (DHAP), а также 3-углеродную молекулу. [ Цитация необходима ]
- Альдолаза и фруктоза-1,6-бисфосфатаза преобразуют G3P и DHAP в фруктозу 6-фосфат (6C). Фосфатный ион теряется в растворе. [ Цитация необходима ]
- Затем фиксация другого
2 генерирует еще два G3P. [ Цитация необходима ] - F6P имеет два углерода, удаленные транскетолазой , давая эритрозе-4-фосфат (E4P). Два углерода на транскетлазе добавляются в G3P, что дает кетоз ксилулозо-5-фосфат (xu5p). [ Цитация необходима ]
- E4P и DHAP (образованный из одного из G3P со второго
CO
2 fixation) are converted into sedoheptulose-1,7-bisphosphate (7C) by aldolase enzyme.[citation needed]
- Седогептолоза-1,7-бисфосфатаза (один из трех ферментов цикла кальвина, которые являются уникальными для растений) расщепляет седедогептолозу-1,7-бисфосфат в седогептолозу-7-фосфат , высвобождая неорганический фосфатный ион в раствор. [ Цитация необходима ]
- Фиксация третьего со стороны
2 генерирует еще два G3P. Ketose S7P имеет два углерода, удаленные транскетолазой , давая рибозу-5-фосфат (R5P), а два углерода, оставшиеся на транскетлазе, переносятся в один из G3P, что дает еще один Xu5p. Это оставляет один G3P в качестве продукта фиксации 3 CO
2 , с генерацией трех пентоз, которые могут быть преобразованы в RU5P. [ Цитация необходима ] - R5P превращается в рибулозо-5-фосфат (RU5P, RUP) с помощью фосфопентозой изомеразы . Xu5p превращается в RUP с помощью фосфопентозной эпимеразы . [ Цитация необходима ]
- Наконец, фосфорибулокиназа (еще один энзимент растительного университета пути) фосфорилирует RUP в RUBP, рибулозу-1,5-бисфосфат, завершая цикл Кальвина . Это требует ввода одного АТФ. [ Цитация необходима ]
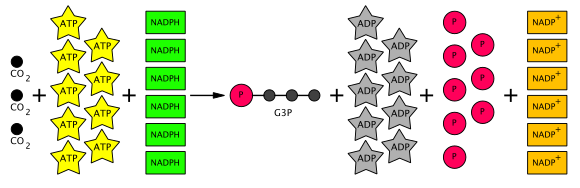
Таким образом, из шести произведенных G3P пять используются для изготовления трех молекул RUBP (5C) (всего 15 углерода), причем только один G3P доступен для последующего преобразования в гексозу. Для этого требуется девять молекул АТФ и шесть молекул NADPH на три CO
2 молекулы. Уравнение общего цикла Кальвина отображается в диаграмме ниже. [ Цитация необходима ]
Рубиско также реагирует на конкуренцию с O
2 вместо co
2 в фотострадаторе . Скорость фотоспечательства выше при высоких температурах. Фотографирование превращает Rubp в 3-PGA и 2-фосфогликолят, 2-углеродную молекулу, которая может быть преобразована через гликолят и глиоксалат в глицин. С помощью системы расщепления глицина и тетрагидрофолата два глицина превращаются в Serine Plus Co
2 Серин может быть преобразован обратно в 3-фосфоглицерат. Таким образом, только 3 из 4 углерода из двух фосфогликолятов могут быть преобразованы обратно в 3-PGA. Можно видеть, что фотоспект имеет очень негативные последствия для растения, потому что, вместо того, чтобы фиксировать
2 , этот процесс приводит к потере CO
2 C4 фиксация углерода эволюционировала, чтобы обойти фотоподранство, но может возникнуть только у определенных растений, родных для очень теплого или тропического климата - например. Кроме того, Rubiscos, катализирующий светозависимые реакции фотосинтеза, как правило, демонстрирует улучшенную специфичность для CO 2 по сравнению с O 2 , чтобы минимизировать реакцию оксигенации. Эта улучшенная специфичность развивалась после того, как Rubisco включила новую субъединицу белка. [ 9 ]
Продукция
[ редактировать ]Непосредственные продукты одного поворота цикла Кальвина-2 молекулы глицеральдегида-3-фосфата (G3P), 3 ADP и 2 NADP + Полем (ADP и NADP + на самом деле не «продукты». Они регенерируются, а затем снова используются в светозависимых реакциях ). Каждая молекула G3P состоит из 3 углерода. Чтобы цикл Кальвина продолжался, RUBP (рибулоза 1,5-бисфосфат) должен быть восстановлен. Таким образом, для этой цели используются 5 из 6 углеродов из 2 молекул G3P. Следовательно, для каждого поворота производится только 1 сетевой углерод. Для создания 1 избыточного G3P требуется 3 углерода, и, следовательно, 3 поворота цикла Кальвина. Для создания одной молекулы глюкозы (которая может быть создана из 2 молекул G3P) потребует 6 видов цикла Кальвина. Избыток G3P также может использоваться для формирования других углеводов, таких как крахмал, сахароза и целлюлоза, в зависимости от того, что нужно растению. [ 10 ]
Светозависимая регуляция
[ редактировать ]Эти реакции не встречаются в темноте или ночью. Существует светозависимая регуляция велосипедных ферментов, поскольку третий шаг требует NADPH. [ 11 ]
Есть две системы регулирования, когда цикл должен быть включен или выключен: система активации тиоредоксина / ферредоксина , которая активирует некоторые из ферментов цикла; и активация фермента Рубиско , активная в цикле Кальвина, которая включает в себя собственную активазу. [ 12 ]
Система тиоредоксина/ферредоксина активирует ферменты глицеральдегид-3-P, дегидрогеназа, глицеральдегид-3-P-фосфатазу, фруктозу-1,6-бисфосфатазу, седогептилозу-1,7-бисфосфатаза и риболезо-5-хеффатаза, которые являются Kieplose-1,7-бисфосфатазой и риболезо-5-фаназой, которые являются Kieptulase-1,7-бисфосфатазой, и риболезо-5-фаназазазазазазазазазазазазазазазазазазазазазазазазой, седогептаза-1,7-бисфосфатазо процесса. Это происходит, когда доступен свет, так как белок ферредоксина снижается в комплексе фотосистемы I цепи тилакоидной электронов, когда электроны циркулируют через нее. [ 13 ] Затем ферредоксин связывается и уменьшает белок тиоредоксина, который активирует ферменты цикла, разрывая цистинную связь, обнаруженную во всех этих ферментах. Это динамический процесс, так как та же связь снова образуется другими белками, которые деактивируют ферменты. Последствия этого процесса заключаются в том, что ферменты остаются в основном активированными днем и деактивированы в темноте, когда не более снижены ферредоксина. [ Цитация необходима ]
Фермент Rubisco имеет свой собственный, более сложный процесс активации. Требуется, чтобы специфическая лизина аминокислота была карбамилирована для активации фермента. Этот лизин связывается с RUBP и приводит к нефункциональному состоянию, если его не оставить некварбамилированным. Специфический фермент активизии, называемый rubisco activase , помогает этому процессу карбамилирования путем удаления одного протона из лизина и делая возможным связывание молекулы диоксида углерода. Даже тогда фермент Rubisco еще не функционален, так как ему нужен ион магния, связанный с лизином для функционирования. Этот ион магния высвобождается из тилакоидного просвета, когда внутренний рН падает из -за активной накачки протонов из электрона. Сама активаза Rubisco активируется повышенными концентрациями АТФ в строме, вызванной ее фосфорилированием . [ 14 ]
Ссылки
[ редактировать ]- ^ Сильверштейн, Элвин (2008). Фотосинтез . Книги двадцать первого века. п. 21. ISBN 9780822567981 .
- ^ Кушман, Джон С. (2001). «Пластическая фотосинтетическая адаптация к засушливым средам» . Физиология растений . 127 (4): 1439–1448. doi : 10.1104/pp.010818 . PMC 1540176 . PMID 11743087 .
- ^ Саинис, Джаяшри Кришна; Дани, Дикша Нархар; Дей, Гаутам Кумар (2003-01-01). «Участие тилакоидных мембран в супрамолекулярной организации ферментов цикла Кальвина inanacystis nidulans» . Журнал физиологии растений . 160 (1): 23–32. doi : 10.1078/0176-1617-00849 . ISSN 0176-1617 .
Результаты показали, что целостность тилакоидных мембран может иметь важное значение для организации последовательных ферментов цикла Кальвина in vivo и облегчить их функционирование.
- ^ Bassham J, Benson A, Calvin M (1950). «Путь углерода в фотосинтезе» (PDF) . J Biol Chem . 185 (2): 781–7. doi : 10.2172/910351 . PMID 14774424 . Архивировано из оригинала (PDF) на 2009-02-19 . Получено 2013-07-03 .
- ^ Кэмпбелл, Нил А.; Брэд Уильямсон; Робин Дж. Хейден (2006). Биология: изучение жизни . Бостон, Массачусетс: Пирсон Прентис Холл. ISBN 0-13-250882-6 .
- ^ Farazdaghi H (2009). «Моделирование кинетики активации и реакции Рубиско из газообмена». Фотосинтез в Силико . Достижения в области фотосинтеза и дыхания. Тол. 29. С. 275–294. doi : 10.1007/978-1-4020-9237-4_12 . ISBN 978-1-4020-9236-7 .
- ^ Лоример, GH; Эндрюс, TJ; Пирс, Дж.; Schloss, JV (1986). «2´-карбокси-3-кето-D-арабинитол 1,5-бисфосфат, шестиуглеродистый промежуток реакции рибулозы бисфосфат карбоксилазы» . Фил. Транс. R. Soc. Лонд Беременный 313 (1162): 397–407. Bibcode : 1986rsptb.313..397L . doi : 10.1098/rstb.1986.0046 .
- ^ Кэмпбелл и Рис Биология: 8 -е издание, стр. 198. Бенджамин Каммингс, 7 декабря 2007 г.
- ^ Schulz, L; Го, Z; Zarzycki, J; Steinchen, W; Шуллер, JM; Хеймерл, т; Принц, S; Мюллер-Каджар, о; Erb, TJ; Хохберг, GKA (14 октября 2022 г.). «Эволюция повышенной сложности и специфичности на рассвете формы I Rubiscos» . Наука . 378 (6616): 155–160. Bibcode : 2022sci ... 378..155s . doi : 10.1126/science.abq1416 . PMID 36227987 . S2CID 252897276 .
- ^ Russell, Wolfe et al. Биология: Изучение разнообразия жизни . Колонто: Колледж Нельсона, 1 -е изд, вып. 1, 2010, стр. 151
- ^ Шрайер, Тина; Хибберд, Джулиан (1 марта 2019 г.). «Вариации в цикле Кальвина - Бенсона: давление и оптимизация отбора?» Полем Журнал экспериментальной ботаники . 70 (6) (опубликовано 27 марта 2019 г.): 1697–1701. doi : 10.1093/jxb/erz078 . PMC 6436154 . PMID 30916343 - Via Oxford Academic.
- ^ Konwarh, Rocktotpal; Абда, Эбрагим М.; Haregu, Simatsidk; Сингх, Ананд Пратап (2022-01-01), Хан, Раджу; Мурали, с.; Gogoi, Satyabrat (Eds.), «Глава 7 - Примерное свидетельство био -нано перекрестных помех между углеродными точками и системами растений» , углеродные точки в сельскохозяйственных системах , Academic Press, стр. 155–173, ISBN 978-0-323-90260-1 Получено 2023-04-22
- ^ Бесс, я; Бьюкенен Б. (1997). «Тиоредоксин-связанные с животными и растительными процессами: новое поколение». Бот. Бык Академический Грех 38 : 1–11.
- ^ Рууска, Сари А.; Эндрюс, Т. Джон; Барсук, Мюррей Р.; Прайс, Г. Дин; Von Caemmerer, Susanne (2000-02-01). «Роль хлоропластного транспорта и метаболитов в модуляции активности рубиско в табаке. Понимания трансгенных растений с уменьшенным количеством цитохрома B/F или глицеральдегид 3-фосфатдегидрогеназы» . Физиология растений . 122 (2): 491–504. doi : 10.1104/pp.122.2.491 . ISSN 1532-2548 . PMC 58886 . PMID 10677442 .
- Bassham JA (2003). «Картирование цикла сокращения углерода: личная ретроспектива». Фотосинты. Резерв 76 (1–3): 35–52. doi : 10.1023/a: 1024929725022 . PMID 16228564 . S2CID 528544452 .
- Диван, Джойс Дж. (2005). «Фотосинтетическая темная реакция» . Биохимия и биофизика, Политехнический институт Ренсселера. Архивировано из оригинала 2005-03-16 . Получено 2012-10-24 .
- Портис, Арчи; Парри, Мартин (2007). «Открытия в Rubisco (рибулоза 1,5-бисфосфат-карбоксилаза/оксигеназа): историческая перспектива» (PDF) . Фотосинтез исследования . 94 (1): 121–143. doi : 10.1007/s11120-007-9225-6 . PMID 17665149 . S2CID 39767233 . Архивировано из оригинала (PDF) 2012-03-12.
Дальнейшее чтение
[ редактировать ]- Rubisco Activase, из онлайн-сайта Plant Physology Archived 2009-02-18 на The Wayback Machine
- Тиоредоксины, от онлайн-сайта Plant Physology Archived 2008-06-16 на The Wayback Machine
Внешние ссылки
[ редактировать ]- Биохимия цикла Кальвина в Политехническом институте Ренсселера
- Цикл Кальвина и пентозофосфатный путь от биохимии , пятое издание Джереми М. Берга, Джона Л. Тимокко и Луберта Страйера. Опубликовано WH Freeman and Company (2002).
Филиалы химии |
|---|