Гребневик
| Гребенчатое желе Временной диапазон: | |
|---|---|
 | |
| «Гребневик» (гребневик) | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Гребневик Эшшольц , 1829 г. |
| Классы | |
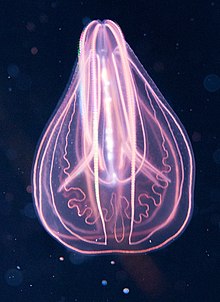
Гребневик ( / t ə ˈ n ɒ f / ə ə tə- NOF -ər -ə ; сг.: гребневик / ˈ t ɛ n ə f ɔːr , ˈ t iː n ə - / TEN -ə-for, TEE -ne- ; от древнегреческого κτείς (kteis) «гребень» и φέρω (pherō) «нести») [6] представляют собой морских тип беспозвоночных , широко известных как гребневики , обитающих в морских водах по всему миру. Они примечательны группами ресничек , которые они используют для плавания (обычно называемыми «гребешками»), и являются крупнейшими животными, плавающими с помощью ресничек.
В зависимости от вида взрослых гребневиков варьируется от нескольких миллиметров размер до 1,5 м (5 футов). В настоящее время известно только 186 современных видов. [7]
Их тела состоят из массы желе со слоем толщиной в две клетки снаружи и еще одним слоем, выстилающим внутреннюю полость. Этот тип имеет широкий диапазон форм тела, включая яйцевидных цидиппидов с парой втягивающихся щупалец, которые захватывают добычу, плоских, как правило, безгребешковых платиктенид и бероидов с большим ртом , которые охотятся на других гребневиков.
Почти все гребневики действуют как хищники , пожирая добычу от микроскопических личинок и коловраток до взрослых мелких ракообразных ; Исключение составляют молодь двух видов, паразитирующая на сальпах , которыми питаются взрослые особи их вида.
Несмотря на их мягкие студенистые тела, окаменелости, которые, как полагают, представляют собой гребневики, появляются в Лагерштеттене еще в раннем кембрии , около 525 миллионов лет назад. Положение гребневиков на «древе жизни» уже давно обсуждается в исследованиях молекулярной филогенетики. Биологи предположили, что гребневики составляют вторую по возрасту ветвящуюся линию животных, а губки являются сестринской группой всех других многоклеточных животных (гипотеза Porifera Sister). [8] Другие биологи утверждают, что гребневики появились раньше, чем губки (гипотеза Ctenophora Sister), которые сами появились до разделения между книдариями и билатериями . [9] [10] Пизани и др. повторно проанализировали данные и предположили, что компьютерные алгоритмы, использованные для анализа, были введены в заблуждение наличием специфических генов гребневиков, которые заметно отличались от таковых у других видов. [11] [12] Последующий анализ, проведенный Whelan et al. (2017) [13] дало дополнительную поддержку гипотезе Ctenophora Sister, и этот вопрос остается предметом таксономических споров. [14] [15] Шульц и др. (2023) обнаружили необратимые изменения синтении у сестры гребневиков — Myriazoa , состоящей из остальных животных. [16]
Отличительные особенности
[ редактировать ]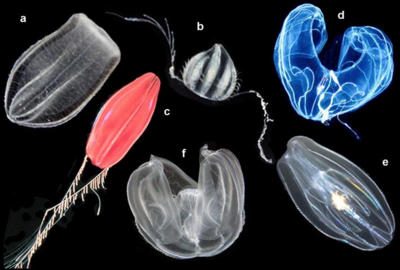
a Beroe ovata , b неопознанный цидиппид, c цидиппид "Tortugas red",
d Bathocyroe Fosteri , e Mnemiopsis leidyi и f Ocyropsis sp. [17]
Среди типов животных гребневики более сложны, чем губки , примерно так же сложны, как книдарии ( медузы , морские анемоны и т. д.), и менее сложны, чем билатерии (к которым относятся почти все другие животные). В отличие от губок, и гребневики, и книдарии имеют:
- клетки, связанные межклеточными связями и
- ковровые базальные мембраны ;
- мышцы ;
- нервная система ; и
- органы чувств (у некоторых, не у всех).
Гребневики отличаются от всех других животных наличием колобластов , которые липкие и прикрепляются к добыче, хотя у некоторых видов гребневиков они отсутствуют. [18] [19]
Как и книдарии, гребневики имеют два основных слоя клеток, между которыми находится средний слой желеобразного материала, который называется мезоглеей у книдарий и гребневиков ; у более сложных животных есть три основных клеточных слоя и нет промежуточного желеобразного слоя. Следовательно, гребневиков и книдарий традиционно называли диплобластическими . [18] [20] И гребневики, и книдарии имеют тип мышц , который у более сложных животных возникает из среднего клеточного слоя , [21] и в результате в некоторых недавних учебниках гребневики классифицируются как триплобластные . [22] в то время как другие все еще считают их диплобластическими. [18] Гребенчатые медузы содержат более 80 различных типов клеток , что превышает число других групп, таких как плакозойные, губки, книдарии и некоторые глубоковетвистые двулатеральные организмы. [23]
Размер варьируется от 1 миллиметра (0,04 дюйма) до 1,5 метра (5 футов), [22] [24] гребневики — крупнейшие неколониальные животные, которые используют реснички («волосы») в качестве основного метода передвижения. [22] У большинства видов есть восемь полосок, называемых гребешковыми рядами, которые проходят по всей длине их тела и несут гребнеобразные полосы ресничек, называемые «ктенами», сложенные вдоль рядов гребешков так, что, когда реснички бьются, реснички каждого гребня касаются гребня. ниже. [22] Название «гребневик» означает «гребень», от греческого κτείς (форма стебля κτεν- ), означающего «гребень», и греческого суффикса -φορος, означающего «несущий». [25]
| Губки [26] [27] | Книдарийцы [18] [20] [28] | Гребневики [18] [22] | билатерия [18] | |
|---|---|---|---|---|
| Книдоциты | Нет | Да | Лишь у некоторых видов (получено из проглоченных книдарий) | |
| микроРНК | Да | Да | Нет | Да |
| Hox-гены | Нет | Да | Нет | Да |
| Коллобласты | Нет | У большинства видов [19] | Нет | |
| пищеварения и кровообращения Органы | Нет | Да | ||
| Анальные поры | Нет | Да | В основном да | |
| Количество слоев основных клеток | Две, между ними желеобразная прослойка. | Спор о том, есть ли два [18] или три [21] [22] | Три | |
| Клетки каждого слоя связаны друг с другом | Нет, за исключением того, что у гомосклероморфов есть базальные мембраны. [29] | Да: межсотовые соединения; базальные мембраны | ||
| чувств Органы | Нет | Да | ||
| Глаза (например, глаза ) | Нет | Да | Нет | Да |
| Апикальный орган | Нет | Да | Да | У видов с первичными реснитчатыми личинками |
| Численность клеток в среднем «желейном» слое | Много | Немного | [непригодный] | |
| Клетки внешнего слоя может перемещаться внутрь и менять функции | Да | Нет | ||
| Нервная система | Нет | Да, просто | От простого к сложному | |
| Мышцы | Никто | Преимущественно эпителиально-мышечная | Преимущественно миоэпителиальный | Преимущественно миоциты |
Описание
[ редактировать ]Для типа, насчитывающего относительно мало видов, гребневики имеют широкий диапазон строения тела. [22] Прибрежные виды должны быть достаточно прочными, чтобы противостоять волнам и кружащимся частицам отложений, в то время как некоторые океанические виды настолько хрупки, что их очень трудно поймать в целости и сохранности для изучения. [19] Кроме того, океанические виды плохо сохраняются, [19] и известны главным образом по фотографиям и запискам наблюдателей. [30] Поэтому наибольшее внимание до недавнего времени концентрировалось на трех прибрежных родах – Pleurobrachia , Beroe и Mnemiopsis . [19] [31] По крайней мере, в двух учебниках описания гребневиков основаны на цидиппидных Pleurobrachia . [18] [22]
Поскольку тело многих видов почти радиально-симметрично , главная ось орально - аборальная (от рта к противоположному концу). Однако, поскольку только два канала вблизи статоциста заканчиваются анальными порами, гребневики не обладают зеркальной симметрией, хотя многие из них имеют вращательную симметрию. Другими словами, если животное вращается полукругом, оно выглядит так же, как и в начале. [32]
Общие особенности
[ редактировать ]гребневиков Тип имеет широкий спектр форм тела, включая уплощенных глубоководных платиктенид , у которых у взрослых особей большинства видов отсутствуют гребни, и прибрежных бероидов , у которых нет щупалец и которые охотятся на других гребневиков, используя огромные рты, вооруженные группами. больших, жестких ресничек , которые действуют как зубы.
Слои тела
[ редактировать ]
Как и у книдарий ( медуз , актиний и др.), тела гребневиков состоят из относительно толстой желеобразной мезоглеи, зажатой между двумя эпителиями , слоями клеток , связанных межклеточными связями и волокнистой базальной мембраной , которую они образуют. секрет . [18] [22] Эпителий гребневиков имеет два слоя клеток, а не один, а некоторые клетки верхнего слоя имеют по несколько ресничек на клетку. [22]
Наружный слой эпидермиса ( наружная кожа) состоит из: чувствительных клеток; клетки, выделяющие слизь , защищающую организм; и интерстициальные клетки, которые могут трансформироваться в другие типы клеток. В специализированных частях тела внешний слой также содержит колобласты , расположенные на поверхности щупалец и используемые при захвате добычи, или клетки, несущие несколько крупных ресничек для передвижения. Внутренний слой эпидермиса содержит нервную сеть и миоэпителиальные клетки, действующие как мышцы . [22]
Внутреннюю полость образуют: рот, обычно закрывающийся мышцами; глотка («горло» ) ; более широкая область в центре, выполняющая роль желудка ; и система внутренних каналов. Они разветвляются через мезоглею к наиболее активным частям животного: рту и глотке; корни щупалец, если они есть; по всей нижней стороне каждого ряда гребенки; и четыре ветви вокруг сенсорного комплекса на дальнем конце от рта - две из этих четырех ветвей заканчиваются анальными порами. Внутренняя поверхность полости выстлана эпителием — гастродермой . Рот и глотка имеют как реснички , так и хорошо развитые мышцы. В других частях системы каналов гастродерма различна на сторонах, ближайших и дальних от органа, который он снабжает. Ближняя сторона состоит из высоких питательных клеток, которые хранят питательные вещества в вакуолях (внутренних отсеках), зародышевых клеток , которые производят яйцеклетки или сперматозоиды, и фотоцитов , которые производят биолюминесценцию . Сторона, наиболее удаленная от органа, покрыта реснитчатыми клетками, которые циркулируют воду через каналы, перемежающиеся ресничными розетками, порами, которые окружены двойными завитками ресничек и соединяются с мезоглеей. [22]
Питание, выделение и дыхание
[ редактировать ]Когда добыча проглатывается, она разжижается в глотке под действием ферментов и мышечных сокращений глотки. Полученная суспензия проходит через систему каналов за счет биения ресничек и переваривается питательными клетками. Ресничные розетки в каналах могут способствовать транспортировке питательных веществ к мышцам мезоглеи. Анальные поры могут выбрасывать нежелательные мелкие частицы, но большая часть нежелательных веществ отрыгивается через рот. [22]
Мало что известно о том, как гребневики избавляются от продуктов жизнедеятельности клеток. Ресничные розетки в гастродерме могут помочь удалять отходы из мезоглеи, а также регулировать плавучесть животного , перекачивая воду в мезоглею или из нее. [22]
Передвижение
[ редактировать ]На внешней поверхности обычно имеется восемь гребенчатых рядов, называемых плавательными пластинками, которые используются для плавания. Ряды ориентированы так, чтобы идти от рта («оральный полюс») к противоположному концу («аборальный полюс») и расположены более или менее равномерно вокруг тела. [18] хотя схема расположения гребней варьируется в зависимости от вида, и у большинства видов ряды гребней простираются только на часть расстояния от аборального полюса ко рту. «Гребни» (также называемые «ктенами» или «гребенчатыми пластинками») проходят через каждый ряд и каждый состоит из тысяч необычайно длинных ресничек, до 2 миллиметров (0,08 дюйма). В отличие от обычных ресничек и жгутиков, которые имеют структуру нитей, расположенных по схеме 9 + 2, эти реснички расположены по схеме 9 + 3, где предполагается, что сверхкомпактная нить выполняет поддерживающую функцию. [33] Обычно они бьются так, чтобы движение происходило в сторону ото рта, хотя они также могут менять направление движения. гребневики обычно плывут в том направлении, в котором рот ест Следовательно, в отличие от медуз, . [22] Пытаясь спастись от хищников, один вид может ускориться в шесть раз по сравнению с обычной скоростью; [34] некоторые другие виды меняют направление в рамках своего побега, изменяя ход ресничек гребенчатой пластинки.
Неясно, как гребневики контролируют свою плавучесть, но эксперименты показали, что некоторые виды полагаются на осмотическое давление , чтобы адаптироваться к воде различной плотности. [35] Жидкости их тела обычно имеют такую же концентрацию , как морская вода. Если они попадают в менее плотную соленую воду, ресничные розетки в полости тела могут перекачивать ее в мезоглею , чтобы увеличить ее объем и уменьшить плотность, чтобы избежать затопления. И наоборот, если они переходят от солоноватой к полноценной морской воде, розетки могут выкачивать воду из мезоглеи, уменьшая ее объем и увеличивая плотность. [22]
Нервная система и органы чувств
[ редактировать ]У гребневиков нет мозга или центральной нервной системы , но вместо этого имеется субэпидермальная нервная сеть (похожая на паутину), которая образует кольцо вокруг рта и наиболее плотна вблизи таких структур, как ряды гребней, глотка, щупальца (если они есть) и сенсорные органы. комплекс дальше всего от рта. [22] Для связи между нервными клетками используются два разных метода; Обнаружено, что некоторые нейроны имеют синаптические связи , но нейроны нервной сети весьма различимы тем, что они слиты в синцитий , а не соединены синапсами. У некоторых животных вне гребневиков также имеются сросшиеся нервные клетки, но никогда до такой степени, чтобы они образовывали целую нервную сеть. [36] [37] [38] Окаменелости показывают, что кембрийские виды имели более сложную нервную систему с длинными нервами, которые соединялись кольцом вокруг рта. Единственные известные сегодня гребневики с длинными нервами — это Euplokamis из отряда Cydippida. [39] Их нервные клетки возникают из тех же клеток-предшественников, что и коллобласты. [40]
Кроме того, имеется менее организованная мезоглеальная нервная сеть, состоящая из одиночных нейритов. Самым крупным сенсорным элементом является аборальный орган (на противоположном от рта конце), который окружен собственной нервной сетью. [41] Основным компонентом этого органа является статоцист , датчик баланса, состоящий из статолита, крошечного зерна карбоната кальция, поддерживаемого четырьмя пучками ресничек , называемых «балансирами», которые определяют его ориентацию. Статоцист защищен прозрачным куполом из длинных неподвижных ресничек. Гребневик не пытается автоматически удерживать статолит на всех балансирах одинаково. Вместо этого его реакция определяется «настроением» животного, другими словами, общим состоянием нервной системы. Например, если гребневик с висячими щупальцами захватывает добычу, он часто поворачивает несколько рядов гребней в обратном направлении, поворачивая рот в сторону добычи. [22]
Исследования подтверждают гипотезу о том, что ресничные личинки книдарий и двулатерий имеют древнее и общее происхождение. [42] Верхушечный орган личинок участвует в формировании нервной системы. [43] Аборальный орган гребневиков не гомологичен апикальному органу других животных, поэтому формирование их нервной системы имеет иное эмбриональное происхождение. [44]
Нервные клетки и нервная система гребневиков имеют другую биохимию по сравнению с другими животными. Например, у них отсутствуют гены и ферменты, необходимые для производства нейротрансмиттеров, таких как серотонин , дофамин , оксид азота , октопамин , норадреналин и других, которые в противном случае наблюдаются у всех других животных с нервной системой, причем гены кодируют рецепторы для каждого из них. нейротрансмиттеры отсутствуют. [45] Монофункциональная каталаза (CAT), одно из трех основных семейств антиоксидантных ферментов, нацеленных на перекись водорода , важную сигнальную молекулу синаптической и нейрональной активности, также отсутствует, скорее всего, из-за потери генов. [46] Было обнаружено, что они используют L-глутамат в качестве нейромедиатора и имеют необычно большое разнообразие ионотропных рецепторов глутамата и генов, отвечающих за синтез и транспорт глутамата, по сравнению с другими многоклеточными животными. [47] Геномное содержание генов нервной системы является наименьшим из известных среди всех животных и может представлять собой минимальные генетические требования для функциональной нервной системы. [48] Тот факт, что части нервной системы имеют непосредственно слитые нейроны, без синапсов, предполагает, что гребневики могут образовывать сестринскую группу другим многоклеточным животным, развившим нервную систему независимо. [38] Если гребневики являются сестринской группой для всех других многоклеточных животных, нервные системы могли быть либо утрачены у губок и плакозойных, либо возникать более одного раза среди многоклеточных животных. [49]
Цидиппиды
[ редактировать ]
Цидиппидные гребневики имеют тела более или менее округлой формы, иногда почти сферические, а иногда более цилиндрические или яйцевидные; обыкновенный прибрежный «морской крыжовник», Pleurobrachia , иногда имеет яйцевидное тело со ртом на узком конце; [22] хотя некоторые особи имеют более однородную округлую форму. С противоположных сторон тела простирается пара длинных тонких щупалец, каждое из которых заключено в оболочку, в которую его можно втянуть. [18] У некоторых видов цидиппид тело в разной степени уплощено, поэтому в плоскости щупалец оно шире. [22]
Щупальца гребневиков цидиппид обычно окаймлены тентиллами («маленькими щупальцами»), хотя у некоторых родов есть простые щупальца без этих боковых ветвей. Щупальца и тентиллы густо покрыты микроскопическими колобластами , которые захватывают добычу, присасываясь к ней. Коллобласты представляют собой специализированные грибовидные клетки наружного слоя эпидермиса и состоят из трех основных компонентов: куполообразной головки с пузырьками (камерами), содержащими клей; стебель, закрепляющий клетку в нижнем слое эпидермиса или в мезоглее; и спиральная нить, которая обвивает стебель и прикрепляется к головке и корню стебля. Функция спиральной нити неясна, но она может поглощать напряжение, когда добыча пытается убежать, и, таким образом, предотвращать разрыв колобласта. [22] Один вид, Minictena luteola, диаметр которого составляет всего 1,5 мм, имеет пять различных типов клеток колобласта. [50] [51]
Помимо коллобластов, представители рода Haeckelia , которые питаются в основном медузами своих жертв , включают жалящие нематоциты в свои собственные щупальца – некоторые книдарийоядные голожаберники аналогичным образом включают нематоциты в свои тела для защиты. [52] Тентиллы Euplokamis значительно отличаются от таковых у других цидиппид: они содержат поперечнополосатую мускулатуру , тип клеток, ранее неизвестный в типе Ctenophora; и в расслабленном состоянии они скручиваются, в то время как щупальцы всех других известных гребневиков удлиняются в расслабленном состоянии. Тентилла Эуплокамиса имеет три типа движений, которые используются при захвате добычи: они могут вылетать очень быстро (за 40–60 миллисекунд ); они могут извиваться, что может приманить добычу, ведя себя как маленькие планктонные черви; и они обвивают добычу. Уникальное постукивание представляет собой раскручивающееся движение, вызванное сокращением поперечно-полосатой мышцы . Извивающиеся движения производятся гладкими мышцами , но узкоспециализированного типа. Свертывание вокруг добычи осуществляется в основном за счет возвращения щупалец в неактивное состояние, но кольца могут натягиваться гладкими мышцами. [53]
Есть восемь рядов гребней, идущих от рта к противоположному концу и равномерно расположенных по всему телу. [18] «Гребни» бьются в метахронном ритме, напоминающем ритм мексиканской волны . [54] От каждого балансира в статоцисте под куполом отходит ресничная борозда, которая затем разделяется и соединяется с двумя соседними гребенчатыми рядами, а у некоторых видов проходит вдоль гребенчатых рядов. Это образует механическую систему передачи ритма сокращений от гребней к балансирам посредством возмущений воды, создаваемых ресничками. [55]
лопастные
[ редактировать ]
есть У Lobata пара лопастей, которые представляют собой мускулистые, чашеобразные продолжения тела, выступающие за пределы рта. Их незаметные щупальца берут начало из углов рта, проходят извилистыми бороздками и распространяются по внутренней поверхности долей (а не тянутся далеко позади, как у Cydippida). Между долями по обе стороны рта у многих видов лопастных есть четыре ушные раковины, студенистые выступы, окаймленные ресничками, которые создают потоки воды, которые помогают направлять микроскопическую добычу ко рту. Такое сочетание структур позволяет лопастям непрерывно питаться подвешенной планктонной добычей. [22]
У лопастей восемь гребенчатых рядов, начинающихся от аборального полюса и обычно не выходящих за пределы тела на доли; у видов с (четырьмя) ушными раковинами реснички, окаймляющие ушные раковины, являются продолжением ресничек в четырех рядах гребешков. Большинство лопастей довольно пассивны при движении по воде, используя для движения реснички на рядах гребней. [22] хотя у Leucotea длинные и активные ушные раковины, движения которых также способствуют движению. Представители лопастных родов Bathocyroe и Ocyropsis могут спастись от опасности, хлопая лопастями, так что струя выбрасываемой воды очень быстро отбрасывает их назад. [56] В отличие от цидиппид, движения гребней лопастных координируются нервами, а не возмущениями воды, создаваемыми ресничками, однако гребни в том же ряду бьются в том же стиле мексиканской волны , что и механически скоординированные гребенчатые ряды цидиппид и бероидов. [55] Возможно, это позволило лопастям вырасти больше, чем цидиппиды, и иметь менее яйцеобразную форму. [54]
Необычный вид, впервые описанный в 2000 году, Lobatolampea Tetragona , был классифицирован как лопастной, хотя лопасти являются «примитивными», а тело напоминает медузу , когда плавает, и дискообразную, когда отдыхает на морском дне. [30]
Бероиды
[ редактировать ]
, У бероидов также известных как нуда , нет кормовых придатков, но их большая глотка , расположенная прямо внутри большого рта и заполняющая большую часть мешковидного тела, несет на оральном конце «макроцилии». Эти сросшиеся пучки из нескольких тысяч крупных ресничек способны «откусывать» куски добычи, которые слишком велики для того, чтобы их можно было проглотить целиком – почти всегда это другие гребневики. [57] Перед полем макроцилий, на ротовых «губах» у некоторых видов Beroe , находится пара узких полосок адгезивных эпителиальных клеток на стенке желудка, которые «застегивают» рот, когда животное не ест, образуя межклеточные связи с противоположной клейкой полоской. Эта плотная застежка упрощает переднюю часть животного, когда оно преследует добычу. [58]
Другие формы тела
[ редактировать ]есть У Ганешиды пара маленьких ротовых долей и пара щупалец. Тело в поперечном сечении скорее круглое, чем овальное, а глотка простирается на внутренние поверхности долей. [22]
Thalassocalycida , , обнаруженная только в 1978 году и известная только у одного вида [59] Медузоподобные, с укороченными в орально-аборальном направлении телами и короткими гребенчатыми рядами на дальней от рта поверхности, берущими начало вблизи аборального полюса. Добычу они захватывают движениями колокольчика и, возможно, двумя короткими щупальцами. [22]
Цестиды («поясные животные») — планктонные животные лентовидной формы, рот и аборальный орган которых расположены посередине противоположных краев ленты. Вдоль каждого аборального края имеется пара гребнеобразных рядов, а из бороздки по всему оральному краю выходят тентиллы, которые расходятся назад по большей части крыловидной поверхности тела. Цестиды могут плавать, покачивая своим телом, а также взмахивая рядами гребней. Есть два известных вида, распространенных по всему миру в теплых и умеренно-теплых водах: Cestum veneris (« пояс Венеры ») - один из самых крупных гребневиков - длиной до 1,5 метров (4,9 футов), который может медленно или довольно быстро волнообразно двигаться. . Параллельный веламен , длина которого обычно составляет менее 20 сантиметров (0,66 фута), может двигаться намного быстрее, что было описано как «стремительное движение». [22] [60]
Большинство Platyctenida имеют овальные тела, уплощенные в орально-аборальном направлении, с парой щупалец, несущих тентиллы, на аборальной поверхности. Они цепляются за поверхности и ползают по ним, выворачивая глотку и используя ее как мускулистую «ступню». У всех известных видов платиктенид, кроме одного, гребенчатые ряды отсутствуют. [22] Платиктениды обычно имеют загадочную окраску, живут на камнях, водорослях или поверхности тела других беспозвоночных и часто обнаруживаются по длинным щупальцам со множеством боковых ветвей, которые, как видно, стекают с задней части гребневика в течение.
Размножение и развитие
[ редактировать ]
Взрослые особи большинства видов способны регенерировать поврежденные или удаленные ткани. [61] хотя только платиктениды размножаются путем клонирования , отщепляя от краев своих плоских тел фрагменты, которые развиваются в новые особи. [22]
Последний общий предок (LCA) гребневиков был гермафродитом . [62] Некоторые из них являются одновременными гермафродитами, которые могут производить и яйцеклетки, и сперматозоиды одновременно, тогда как другие являются последовательными гермафродитами, у которых яйцеклетки и сперма созревают в разное время. нет Метаморфозы . [63] Известно, что по крайней мере три вида развили отдельные полы ( раздельнополость ); Ocyropsis Crystallina и Ocyropsis maculata из рода Ocyropsis и Bathocyroe Fosteri из рода Bathocyroe . [64] Гонады . располагаются в участках внутренней сети каналов под гребенчатыми рядами, а яйцеклетки и сперматозоиды выходят через поры в эпидермисе Оплодотворение обычно наружное , но платиктениды используют внутреннее оплодотворение и держат яйца в выводковых камерах до тех пор, пока они не вылупятся. Самооплодотворение иногда наблюдалось у видов рода Mnemiopsis . [22] и считается, что большинство гермафродитных видов самоплодны. [19]
Развитие оплодотворенной яйцеклетки прямое; отличительной личиночной формы нет. Молодь всех групп, как правило, является планктонной , и большинство видов напоминают миниатюрных взрослых цидиппид, постепенно развивающих свои взрослые формы тела по мере роста. Однако у рода Beroe молодые особи имеют большие рты и, как и взрослые особи, лишены как щупалец, так и оболочек щупалец. В некоторых группах, таких как плоские, обитающие на дне платиктениды, молодь ведет себя больше как настоящие личинки. Они живут среди планктона и, таким образом, занимают иную экологическую нишу , чем их родители, достигая взрослой формы только в результате более радикального онтогенеза . [22] после падения на морское дно. [19]
По крайней мере, у некоторых видов молодые гребневики способны производить небольшое количество яиц и сперматозоидов, хотя они намного меньше размера взрослых особей, а взрослые особи производят яйцеклетки и сперму до тех пор, пока у них достаточно еды. Если им не хватает еды, они сначала перестают производить яйцеклетки и сперму, а затем уменьшаются в размерах. Когда запасы пищи улучшаются, они вырастают до нормального размера, а затем возобновляют размножение. Эти особенности позволяют гребневикам очень быстро увеличивать свою популяцию. [19] Представители Lobata и Cydippida также имеют форму размножения, называемую дисогенией; две половозрелые стадии: сначала личинка, затем молодь и взрослые особи. Находясь в личиночном состоянии, они способны периодически выпускать гаметы. После того, как их первый репродуктивный период закончился, они не будут производить больше гамет до тех пор, пока это не произойдет. Популяция Mertensia ovum в центральной части Балтийского моря стала педогенетической и состоит исключительно из половозрелых личинок размером менее 1,6 мм. [65] [66]
У Mnemiopsis leidyi передача сигналов оксида азота (NO) присутствует как во взрослых тканях, так и дифференциально экспрессируется на более поздних эмбриональных стадиях, что указывает на участие NO в механизмах развития. [67]
Цвета и биолюминесценция
[ редактировать ]
Большинство гребневиков, обитающих у поверхности, большей частью бесцветны и почти прозрачны. Однако некоторые глубокоживущие виды сильно пигментированы, например, виды, известные как «Красный Тортугас». [68] (см. иллюстрацию здесь), который еще официально не описан. [19] Платиктениды обычно живут прикрепленными к другим донным организмам и часто имеют цвет, похожий на цвет этих организмов-хозяев. [19] Кишечник глубоководного рода Bathocyroe имеет красный цвет, что скрывает биолюминесценцию . копепод проглоченных им [56]
Гребенчатые ряды большинства планктонных гребневиков создают эффект радуги, вызванный не биолюминесценцией , а рассеянием света при движении гребневиков. [19] [69] Большинство видов также являются биолюминесцентными, но свет обычно синий или зеленый, и его можно увидеть только в темноте. [19] Однако некоторые значительные группы, в том числе все известные платиктениды и цидиппиды рода Pleurobrachia , не способны к биолюминесценции. [70]
Когда некоторые виды, в том числе Bathyctena chuni , Euplokamis Stationis и Eurhamphaea vexilligera , потревожены, они производят выделения (чернила), которые люминесцируют практически на тех же длинах волн, что и их тела. Молодые особи будут светиться более ярко в зависимости от размера их тела, чем взрослые особи, свечение которых рассеяно по всему телу. Детальное статистическое исследование не выявило функции биолюминесценции гребневиков и не выявило какой-либо корреляции между ее точным цветом и каким-либо аспектом среды обитания животных, например, глубиной или тем, живут ли они в прибрежных или срединно-океанических водах. [71]
У гребневиков биолюминесценция вызвана активацией активированных кальцием белков, называемых фотопротеинами, в клетках, называемых фотоцитами , которые часто ограничены меридиональными каналами, лежащими под восемью рядами гребней. В геноме Mnemiopsis leidyi десять генов кодируют фотобелки. Эти гены экспрессируются совместно с генами опсина в развивающихся фотоцитах Mnemiopsis leidyi , что повышает вероятность того, что производство света и обнаружение света могут работать вместе у этих животных. [72]
Экология
[ редактировать ]
Распределение
[ редактировать ]Гребневики встречаются в большинстве морских сред: от полярных вод при температуре -2 °С до тропиков при температуре 30 °С; вблизи побережий и в середине океана; от поверхностных вод до глубин океана на высоте более 7000 метров. [73] Лучше всего изучены роды Pleurobrachia , Beroe и Mnemiopsis , поскольку именно эти планктонные прибрежные формы чаще всего собираются у берега. [31] [56] В пресной воде гребневики не обнаружены.
В 2013 году мнемиопсис был зарегистрирован в озере Биркет-Карун, а в 2014 году — в озере Эль-Райан II, оба недалеко от Файюма в Египте, куда они были случайно завезены при транспортировке мальков рыбы (кефали). Хотя многие виды предпочитают солоноватые воды, такие как устья рек и прибрежные лагуны, находящиеся в открытом доступе к морю, это была первая запись во внутренней среде. Оба озера соленые, причем Биркет-Карун является гиперсоленым, что показывает, что некоторые гребневики могут обосноваться в соленой лимнической среде без связи с океаном. В долгосрочной перспективе не ожидается, что популяции выживут. Двумя ограничивающими факторами для соленых озер являются доступность пищи и разнообразный рацион, а также высокие температуры жарким летом. Поскольку в то же время была завезена паразитическая изопода Livoneca redmanii , трудно сказать, какая часть экологического воздействия инвазивных видов вызвана одним только гребневиком. [74] [75] [76]
В некоторых прибрежных районах гребневиков может быть много в летние месяцы, но в других местах они редки и их трудно найти.
В заливах, где они встречаются в очень большом количестве, хищничество гребневиков может контролировать популяции мелких зоопланктонных организмов, таких как копеподы , которые в противном случае могли бы уничтожить фитопланктон (планктонные растения), которые являются жизненно важной частью морских пищевых цепей .
Добыча и хищники
[ редактировать ]Почти все гребневики — хищники — вегетарианцев нет и только один род частично паразитирует . [56] Если еды много, они могут съедать в день в 10 раз больше собственного веса. [77] В то время как берое охотятся в основном на другие гребневики, другие виды поверхностных вод охотятся на зоопланктон (планктонные животные) размером от микроскопических, включая моллюсков и личинок рыб, до мелких взрослых ракообразных, таких как копеподы , амфиподы и даже криль . Представители рода Haeckelia охотятся на медуз (стрекательные клетки) своей жертвы и включают нематоцисты в свои собственные щупальца вместо колобластов . [19] Гребневиков сравнивают с пауками по широкому спектру методов ловли добычи: некоторые неподвижно висят в воде, используя свои щупальца в качестве «паутины», некоторые устраивают засады, как прыгающие пауки Salticid , а некоторые свешивают липкую каплю на конце паука. тонкая нить, как это делают пауки болас . Это разнообразие объясняет широкий спектр форм тела в типе с довольно небольшим количеством видов. [56] Двущупальцевая «цидиппида» Lampea питается исключительно сальпами , близкими родственниками асцидий, образующими большие цепочечные плавающие колонии, а молодь Lampea прикрепляется, как паразиты, к сальпам, слишком большим для того, чтобы они могли их проглотить. [56] Представители рода Cydippid Pleurobrachia и лопастные Bolinopsis часто достигают высокой плотности популяции в одном и том же месте и в одно и то же время, поскольку они специализируются на разных типах добычи: Pleurobrachia длинные щупальца в основном захватывают относительно сильных пловцов, таких как взрослые копеподы, в то время как Bolinopsis обычно питается более мелкие и слабые пловцы, такие как коловратки моллюсков и , личинки ракообразных . [78]
Раньше гребневики считались «тупиками» в морских пищевых цепях, поскольку считалось, что низкое соотношение органических веществ к соли и воде делает их плохой пищей для других животных. Также часто бывает трудно идентифицировать остатки гребневиков в кишках возможных хищников, хотя гребни иногда остаются неповрежденными достаточно долго, чтобы дать подсказку. Детальное исследование кеты Oncorhynchus keta показало, что эти рыбы переваривают гребневики в 20 раз быстрее, чем креветки такого же веса , и что гребневики могут обеспечить хорошую диету, если их вокруг достаточно. Бероиды охотятся в основном на других гребневиков. Некоторые медузы и черепахи поедают большое количество гребневиков, а медузы могут временно уничтожить популяции гребневиков. Поскольку популяция гребневиков и медуз часто имеет большие сезонные колебания, большинство рыб, которые на них охотятся, являются универсальными и могут оказывать большее влияние на популяции, чем специализированные желееды. Это подтверждается наблюдением за растительноядными рыбами, намеренно питающимися студенистым зоопланктоном во время цветения в Красном море. [79] Личинки некоторых актиний паразитируют на гребневиках, как и личинки некоторых плоских червей , паразитирующих на рыбе, когда они достигают зрелого возраста. [80]
Экологические воздействия
[ редактировать ]Большинство видов являются гермафродитами , и молодые особи по крайней мере некоторых видов способны к размножению до достижения размеров и формы взрослой особи. Такое сочетание гермафродитизма и раннего размножения позволяет небольшим популяциям расти с взрывной скоростью.

Гребневики могут сбалансировать морские экосистемы, не давая переизбытку копепод поедать весь фитопланктон (планктонные растения), [81] которые являются доминирующими морскими производителями органических веществ из неорганических ингредиентов. [82]
С другой стороны, в конце 1980-х годов западноатлантический гребневик Mnemiopsis leidyi был случайно завезен в Черное и Азовское моря через балластные цистерны судов, и его обвинили в резком падении уловов рыбы из-за поедания как личинок рыб, так и рыбных личинок. мелкие ракообразные, которыми в противном случае кормили бы взрослую рыбу. [81] Мнемиопсис хорошо приспособлен для вторжения на новые территории (хотя это не было предсказано до тех пор, пока он не столь успешно колонизировал Черное море), поскольку он может очень быстро размножаться и переносить широкий диапазон температур и солености воды . [83] Воздействие усилилось из-за хронического чрезмерного вылова рыбы и эвтрофикации , которая дала всей экосистеме краткосрочный импульс, в результате чего популяция мнемиопсиса росла даже быстрее, чем обычно. [84] – и прежде всего отсутствием эффективных хищников на этих интродуцированных гребневиках. [83] Популяции мнемиопсиса в этих районах в конечном итоге были взяты под контроль в результате случайной интродукции питающегося мнемиопсисом североамериканского гребневика Beroe ovata . [85] и похолоданием местного климата с 1991 по 1993 год, [84] что значительно замедлило метаболизм животного. [83] Однако маловероятно, что численность планктона в этом районе восстановится до уровня, существовавшего до Мнемиопсиса . [86]
В конце 1990-х годов мнемиопсис появился в Каспийском море . Вскоре после этого прибыла Beroe ovata , которая, как ожидается, уменьшит, но не устранит воздействие мнемиопсиса . Мнемиопсис также достиг восточного Средиземноморья в конце 1990-х годов и теперь, по-видимому, процветает в Северном и Балтийском морях . [19]
Таксономия
[ редактировать ]Число известных ныне живущих видов гребневиков неизвестно, поскольку многие из названных и официально описанных оказались идентичны видам, известным под другими научными названиями. По оценкам Клаудии Миллс, существует от 100 до 150 действительных видов, которые не являются дубликатами, и что по крайней мере еще 25, в основном глубоководные формы, признаны отдельными, но еще не проанализированы достаточно подробно, чтобы поддержать формальное описание и наименование. [68]
Ранняя классификация
[ редактировать ]Ранние авторы объединили гребневиков с книдариями в один тип, названный Coelenterata, из-за морфологического сходства между двумя группами. Как и книдарии, тела гребневиков состоят из массы желе, причем один слой клеток находится снаружи, а другой выстилает внутреннюю полость. Однако у гребневиков эти слои имеют глубину в две клетки, а у книдарий - только в одну клетку. Гребневики также напоминают книдарий тем, что полагаются на поток воды через полость тела как для пищеварения, так и для дыхания, а также имеют децентрализованную нервную сеть, а не мозг.Геномные исследования показали, что нейроны гребневиков, которые во многом отличаются от нейронов других животных, развивались независимо от нейронов других животных. [87] а растущее понимание различий между гребневиками и другими кишечнополостными убедило более поздних авторов классифицировать их как отдельные типы . Положение гребневиков в эволюционном генеалогическом древе животных уже давно обсуждается, и в настоящее время большинство взглядов, основанных на молекулярной филогенетике , заключается в том, что книдарии и билатерии более тесно связаны друг с другом, чем любой из них с гребневиками.
Современная таксономия
[ редактировать ]
Традиционная классификация делит гребневики на два класса : со щупальцами ( Tentaculata ) и без щупалец ( Nuda ). К нуде относятся только один отряд ( Beroida ) и семейство ( Beroidae ), а также два рода : Beroe (несколько видов) и Neis (один вид). [68]
Тентакулаты отрядов делятся на следующие восемь : [68]
- Cydippida , животные яйцевидной формы с длинными щупальцами. [22]
- Лобата с парными толстыми лопастями. [22]
- Platyctenida , уплощенные животные, обитающие на морском дне или вблизи него; у большинства взрослых особей нет гребней, и они используют свои глотки как присоски, чтобы прикрепляться к поверхностям. [22]
- Ганешида , с парой маленьких долей вокруг рта, но расширенной глоткой, как у платиктенид. [22]
- Камбоджа
- Криптолобифериды
- Thalassocalycida с короткими щупальцами и медузоподобным «зонтиком». [22]
- Цестида , лентовидные и самые крупные гребневики. [22]
Эволюционная история
[ редактировать ]Несмотря на их хрупкие студенистые тела, окаменелости , которые , как полагают, представляют собой гребневики – по-видимому, без щупалец, но с гораздо большим количеством гребешков, чем у современных форм – были найдены в Лагерштеттене еще в раннем кембрии , около 515 миллионов лет назад . Тем не менее, недавний молекулярно-филогенетический анализ пришел к выводу, что общий предок возник примерно 350 миллионов лет назад ± 88 миллионов лет назад, что противоречит предыдущим оценкам, которые предполагают, что он произошел 66 миллионов лет назад после мел-палеогенового вымирания . [88]
Ископаемая запись
[ редактировать ]Из-за своих мягких студенистых тел гребневики чрезвычайно редки в качестве окаменелостей, а окаменелости, которые были интерпретированы как гребневики, были обнаружены только в лагерштеттенах , местах, где окружающая среда исключительно подходила для сохранения мягких тканей. До середины 1990-х годов были известны только два достаточно хороших для анализа экземпляра, оба из кроновой группы, из раннего девона (эмского) периода . Три дополнительных предполагаемых вида были затем обнаружены в сланцах Бёрджесс и других канадских породах того же возраста, около 505 миллионов лет назад , в середине кембрийского периода. У всех троих отсутствовали щупальца, но было от 24 до 80 рядов гребней, что намного больше, чем 8, типичных для современных видов. У них также, по-видимому, были внутренние органоподобные структуры, в отличие от всего, что можно найти у живых гребневиков. У одного из ископаемых видов, о котором впервые сообщалось в 1996 году, был большой рот, по-видимому, окруженный складчатым краем, который мог быть мускулистым. [4] Данные из Китая, полученные годом позже, позволяют предположить, что такие гребневики были широко распространены в кембрии, но, возможно, сильно отличались от современных видов - например, гребневые ряды одного окаменелости были прикреплены к выступающим лопастям. [89] Самым молодым ископаемым видом, не входящим в группу кроны, является вид Daihuoides из позднего девона, принадлежащий к базовой группе, которая, как предполагалось, вымерла более 140 миллионов лет назад. [90]
Эдиакарская эоандромеда предположительно могла представлять собой гребешок. [2] Он имеет восьмеричную симметрию с восемью спиральными рукавами, напоминающими гребешковые ряды гребневика. Если это действительно гребневик, это ставит группу близко к происхождению Bilateria. [91] Раннекембрийское окаменелость, сидячую ветвь похожая на , Stromatoveris из китайского лагерштетте Чэнцзян и датированная примерно 515 миллионами лет назад , очень похожа на Vendobionta предшествующего эдиакарского периода. Де-Ган Шу, Саймон Конвей Моррис и др. обнаружили на его ветвях то, что они считали рядами ресничек, используемых для фильтрационного питания . Они предположили, что Stromatoveris была эволюционной «тетушкой» гребневиков и что гребневики произошли от сидячих животных, чьи потомки стали пловцами и превратили реснички из механизма питания в двигательную систему. [92] Другими кембрийскими окаменелостями, которые подтверждают идею о том, что гребневики произошли от сидячих форм, являются Dinomischus , Daihua , Xianguangia и Siphusauctum , которые также жили на морском дне, имели органические скелеты и покрытые ресничками щупальца, окружающие их рот, которые были обнаружены кладистическим анализом как члены гребневика стволовой группы [93] [94]
Кембрийские окаменелости возрастом 520 миллионов лет, также найденные в Чэнцзяне в Китае, показывают ныне полностью вымерший класс гребневиков, названный « Scleroctenophora », который имел сложный внутренний скелет с длинными шипами. [95] Скелет также поддерживал восемь мягких створок, которые можно было использовать для плавания и, возможно, для кормления. Одна форма, Таумактена , имела обтекаемое тело, напоминающее тело червей-стрелок , и могла быть ловким пловцом. [5]
Отношения с другими группами животных
[ редактировать ]Филогенетическое очень важно для родство гребневиков с остальными Metazoa нашего понимания ранней эволюции животных и происхождения многоклеточности. Это было предметом дискуссий на протяжении многих лет. Гребневики предположительно являются сестринской линией Bilateria . [96] [97] сестра Книдарии , [98] [99] [100] [101] сестра Cnidaria , Placozoa и Bilateria , [102] [103] [104] и сестра всех других животных. [9] [105]
Вальтер Гарштанг в книге «Личиночные формы и другие зоологические стихи» ( Mülleria and the Ctenophore ) высказал даже теорию, согласно которой гребневики произошли от неотенической Mülleria личинки поликлада .
Серия исследований, в которых рассматривалось наличие и отсутствие членов семейств генов и сигнальных путей (например, гомеобоксов , ядерных рецепторов , сигнального пути Wnt и натриевых каналов ), продемонстрировала доказательства, соответствующие двум последним сценариям, что гребневики являются либо сестринскими, либо сестринскими. к Cnidaria , Placozoa и Bilateria или родственным всем другим типам животных. [106] [107] [108] [109] Несколько более поздних исследований, сравнивающих полные секвенированные геномы гребневиков с секвенированными геномами других животных, также подтвердили, что гребневики являются сестринской линией для всех других животных. [110] [28] [111] [112] Эта позиция предполагает, что типы нервных и мышечных клеток либо были потеряны в основных линиях животных (например, Porifera и Placozoa ), либо эволюционировали независимо в линии гребневиков. [110]
Другие исследователи утверждали, что отнесение Ctenophora к сестре всех других животных является статистической аномалией, вызванной высокой скоростью эволюции геномов гребневиков, и что вместо этого Porifera (губки) является самым ранним дивергентным таксоном животных. [104] [113] [114] [115] [116] У них также чрезвычайно высокие темпы митохондриальной эволюции. [117] и наименьшее известное содержание РНК/белка в геноме мтДНК у животных. [118] Таким образом, Ctenophora, по-видимому, представляет собой базальную кладу диплобластов . В соответствии с последним пунктом, анализ очень большого выравнивания последовательностей в таксономическом масштабе многоклеточных животных (1719 белков, имеющих в общей сложности около 400 000 аминокислотных позиций) показал, что гребневики появляются как вторая по возрасту ветвящаяся линия животных, а губки относятся к сестринской группе. всем остальным многоклеточным животным. [8] Кроме того, исследование генов муцина , которые позволяют животным производить слизь, показывает, что у губок их никогда не было, в то время как все другие животные, включая гребневиков, по-видимому, имеют общие гены общего происхождения. [119] И было обнаружено, что, несмотря на все различия, нейроны гребневиков имеют ту же основу, что и нейроны книдарий, после того, как результаты показали, что нейроны, экспрессирующие пептиды, вероятно, являются предками химических нейротрансмиттеров. [120]
Еще одно исследование категорически отвергает гипотезу о том, что губки являются сестринской группой для всех других современных животных, и устанавливает положение Ctenophora как сестринской группы для всех других животных, а несогласие с последней упомянутой статьей объясняется методологическими проблемами анализа в том, что работа. [13] Ни гребневики, ни губки не имеют путей HIF . [121] их геном экспрессирует только один тип потенциалзависимых кальциевых каналов в отличие от других животных, у которых их три типа, [122] и это единственный известный тип животных, у которого отсутствуют настоящие Hox-гены . [28] Несколько видов из других типов; личинка немертинского пилидия, личинка вида форонид Phoronopsis Harmeri и желудевого червя личинка Schizocardium Californicum также не зависят от hox-генов в своем личиночном развитии, но нуждаются в них во время метаморфоза для достижения взрослой формы. [123] [124] [125] Гены иннексина , которые кодируют белки, используемые для межклеточной коммуникации у животных, также, по-видимому, независимо эволюционировали у гребневиков. [126]
Отношения внутри гребневиков
[ редактировать ]
| ||||||||||||||||||||||||||||
Поскольку все современные гребневики, за исключением бероидов, имеют личинки, похожие на цидиппиды, широко распространено мнение, что их последний общий предок также напоминал цидиппид, имея яйцевидное тело и пару втягивающихся щупалец. Чисто морфологический анализ Ричарда Харбисона в 1985 году пришел к выводу, что цидиппиды не монофилетичны, другими словами, не содержат всех, а только потомков одного общего предка, который сам был цидиппидом. Вместо этого он обнаружил, что различные семейства цидиппид больше похожи на представителей других отрядов гребневиков , чем на других цидиппид. Он также предположил, что последний общий предок современных гребневиков был либо цидипидообразным, либо бероидоподобным. [128] Анализ молекулярной филогении в 2001 году с использованием 26 видов, в том числе 4 недавно открытых, подтвердил, что цидиппиды не монофилетичны, и пришел к выводу, что последний общий предок современных гребневиков был цидиппидоподобным. Было также обнаружено, что генетические различия между этими видами были очень малы – настолько малы, что родство между Lobata, Cestida и Thalassocalycida оставалось неопределенным. Это говорит о том, что последний общий предок современных гребневиков появился относительно недавно и, возможно, пережил мел-палеогеновое вымирание 65,5 миллионов лет назад , в то время как другие линии вымерли. Когда анализ был расширен и включил представителей других типов , он пришел к выводу, что книдарии, вероятно, более тесно связаны с билатериями, чем любая из групп с гребневиками, но этот диагноз неясен. [127] Клада, включающая Mertensia , Charistefane и Euplokamis, может быть сестринской линией для всех других гребневиков. [129] [13]
Время дивергенции, оцененное на основе молекулярных данных, указывает примерно на то, сколько миллионов лет назад (млн лет назад) произошли диверсификации основных клад: 350 млн лет назад для Cydippida по сравнению с другими Ctenophora и 260 млн лет назад для Platyctenida по сравнению с Beroida и Lobata. [13]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Чен, Цзюнь-Юань; Шопф, Дж. Уильям; Боттьер, Дэвид Дж.; Чжан, Чэнь-Ю; Кудрявцев Анатолий Б.; Трипати, Абхишек Б.; Ван, Сю-Цян; Ян, Юн-Хуа; Гао, Сян; Ян, Ин (апрель 2007 г.). «Рамановские спектры зародыша гребневика нижнего кембрия из юго-западной провинции Шэньси, Китай» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (15): 6289–6292. Бибкод : 2007PNAS..104.6289C . дои : 10.1073/pnas.0701246104 . ПМЦ 1847456 . ПМИД 17404242 .
- ^ Перейти обратно: а б Тан, Ф.; Бенгтсон, С.; Ван, Ю.; Ван, XL; Инь, CY (20 сентября 2011 г.). «Эоандромеда и происхождение гребневиков». Эволюция и развитие . 13 (5): 408–414. дои : 10.1111/j.1525-142X.2011.00499.x . ПМИД 23016902 . S2CID 28369431 .
- ^ Стэнли, Джорджия; Штюрмер, В. (9 июня 1983 г.). «Первый ископаемый гребневик из нижнего девона Западной Германии». Природа . 303 (5917): 518–520. Бибкод : 1983Natur.303..518S . дои : 10.1038/303518a0 . S2CID 4259485 .
- ^ Перейти обратно: а б Конвей Моррис, С.; Коллинз, Д.Х. (29 марта 1996 г.). «Среднекембрийские гребневики из формации Стивен, Британская Колумбия, Канада». Философские труды Королевского общества B: Биологические науки . 351 (1337): 279–308. Бибкод : 1996RSPTB.351..279C . дои : 10.1098/rstb.1996.0024 .
- ^ Перейти обратно: а б Шу, Деган; Чжан, Чжифэй; Чжан, Фанг; Солнце, Ге; Хан, Цзянь; Сяо, Шухай; Оу, Цян (июль 2015 г.). «Исчезнувшая история скелетирования кембрийских гребневиков» . Достижения науки . 1 (6): e1500092. Бибкод : 2015SciA....1E0092O . дои : 10.1126/sciadv.1500092 . ПМЦ 4646772 . ПМИД 26601209 .
- ^ Фаулер, Джордж Герберт (1911). . В Чисхолме, Хью (ред.). Британская энциклопедия . Том. 7 (11-е изд.). Издательство Кембриджского университета. п. 593.
- ^ Редакционный совет WoRMS (2024 г.), Всемирный реестр морских видов. Доступ 18 сентября 2019 г. , ВЛИЗ , получено 19 февраля 2024 г.
- ^ Перейти обратно: а б Симион, Пол; Филипп, Эрве; Баурен, Денис; Джагер, Мюриэль; Рихтер, Дэниел Дж.; ДиФранко, Арно; Рур, Беатрис; Сато, Нори; Кеннек, Эрик; Эресковский, Александр; Лапеби, Паскаль; Корре, Эрван; Дельсук, Фредерик; Король, Николь; Верхайде, Герт; Мануэль, Микаэль (2017). «Большой и последовательный набор филогеномных данных подтверждает, что губки являются сестринской группой для всех других животных» . Современная биология . 27 (7): 958–967. Бибкод : 2017CBio...27..958S . дои : 10.1016/j.cub.2017.02.031 . ПМИД 28318975 .
- ^ Перейти обратно: а б Данн, Кейси В.; Хейнол, Андреас; Матус, Дэвид К.; Панг, Кевин; Браун, Уильям Э.; Смит, Стивен А.; Сивер, Элейн; Роуз, Грег В.; Обст, Матиас; Эджкомб, Грегори Д.; Соренсен, Мартин В.; Хэддок, Стивен HD; Шмидт-Раэса, Андреас; Окусу, Акико; Кристенсен, Райнхардт Мёбьерг; Уиллер, Уорд К.; Мартиндейл, Марк К.; Гирибет, Гонсало (2008). «Широкая филогеномная выборка улучшает разрешение древа жизни животных». Природа . 452 (7188): 745–749. Бибкод : 2008Natur.452..745D . дои : 10.1038/nature06614 . ПМИД 18322464 . S2CID 4397099 .
- ^ Баксеванис, Андреас Д.; Мартиндейл, Марк К.; Малликин, Джеймс С.; Вольфсберг, Тайра Г.; Данн, Кейси В.; Хэддок, Стивен HD; Патнэм, Николас Х.; Смит, Стивен А.; Хавлак, Пол; Фрэнсис, Уоррен Р.; Кох, Бернард Дж.; Симмонс, Дэвид К.; Морленд, Р. Трэвис; Нгуен, Ань-Дао; Шницлер, Кристин Э.; Панг, Кевин; Райан, Джозеф Ф. (13 декабря 2013 г.). «Геном гребневика Mnemiopsis leidyi и его значение для эволюции типов клеток» . Наука . 342 (6164): 1242592. doi : 10.1126/science.1242592 . ПМЦ 3920664 . ПМИД 24337300 .
- ^ Пизани, Давиде; Петт, Уокер; Дорманн, Мартин; Феуда, Роберто; Рота-Стабелли, Омар; Филипп, Эрве; Лартильо, Николя; Верхайде, Герт (2015). «Геномные данные не подтверждают, что гребневики являются сестринской группой всех других животных» . Труды Национальной академии наук . 112 (50): 15402–15407. Бибкод : 2015PNAS..11215402P . дои : 10.1073/pnas.1518127112 . ПМЦ 4687580 . ПМИД 26621703 .
- ^ Бервальд, Джули (2017). Бесхребетный: наука о медузах и искусство выращивания позвоночника . Книги Риверхеда. [ нужна страница ]
- ^ Перейти обратно: а б с д Уилан, Натан В.; Кокот, Кевин М.; Мороз Татьяна П.; Мукерджи, Кришану; Уильямс, Питер; Паулай, Густав; Мороз Леонид Л.; Халанич, Кеннет М. (ноябрь 2017 г.). «Отношения с гребневиками и их положение в качестве сестринской группы для всех других животных» . Экология и эволюция природы . 1 (11): 1737–1746. Бибкод : 2017NatEE...1.1737W . дои : 10.1038/s41559-017-0331-3 . ПМЦ 5664179 . ПМИД 28993654 .
- ^ Халанич, Кеннет М.; Уилан, Натан В.; Кокот, Кевин М.; Кон, Андреа Б.; Мороз, Леонид Л. (9 февраля 2016 г.). «Ошибки с губками» . Труды Национальной академии наук . 113 (8): Е946-7. Бибкод : 2016PNAS..113E.946H . дои : 10.1073/pnas.1525332113 . ISSN 0027-8424 . ПМЦ 4776479 . ПМИД 26862177 .
- ^ Телфорд, Максимилиан Дж.; Мороз Леонид Л.; Халанич, Кеннет М. (январь 2016 г.). «Сестринский спор» . Природа . 529 (7586): 286–287. дои : 10.1038/529286a . ISSN 1476-4687 . ПМИД 26791714 . S2CID 4447056 .
- ^ Шульц, Дэррин Т.; Хэддок, Стивен HD; Бредесон, Джессен В.; Грин, Ричард Э.; Симаков Олег; Рохсар, Дэниел С. (17 мая 2023 г.). «Древние генные связи подтверждают, что гребневики являются сестрами других животных» . Природа . 618 (7963): 110–117. Бибкод : 2023Natur.618..110S . дои : 10.1038/s41586-023-05936-6 . ISSN 1476-4687 . ПМЦ 10232365 . ПМИД 37198475 .
- ^ Райан, Джозеф Ф.; Шницлер, Кристин Э.; Тамм, Сидни Л. (декабрь 2016 г.). «Отчет о встрече Ctenopalooza: Первая международная встреча ктенофорологов» . ЭвоДево . 7 (1): 19. дои : 10.1186/s13227-016-0057-3 . hdl : 1912/8430 . S2CID 931968 .
- ^ Перейти обратно: а б с д и ж г час я дж к л Хинде, RT (1998). «Книдарии и гребневики». В Андерсоне, Д.Т. (ред.). Зоология беспозвоночных . Издательство Оксфордского университета. стр. 28–57. ISBN 978-0-19-551368-4 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н Миллс, CE «Гребневики – некоторые заметки эксперта» . Университет Вашингтона . Проверено 5 февраля 2009 г.
- ^ Перейти обратно: а б Руперт, Э.Э.; Фокс, Р.С. и Барнс, Р.Д. (2004). Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр. 111–124 . ISBN 978-0-03-025982-1 .
- ^ Перейти обратно: а б Зайпель, К.; Шмид, В. (июнь 2005 г.). «Эволюция поперечно-полосатых мышц: медузы и происхождение триплобластии» . Биология развития . 282 (1): 14–26. дои : 10.1016/j.ydbio.2005.03.032 . ПМИД 15936326 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также Руперт, Э.Э.; Фокс, Р.С. и Барнс, Р.Д. (2004). Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр. 182–195 . ISBN 978-0-03-025982-1 .
- ^ Мороз Леонид Л.; Норекян Тигран П. (16 августа 2018 г.). «Атлас нервно-мышечной организации гребневика Pleurobrachia bachei (А. Агассис, 1860) ». bioRxiv 10.1101/385435 .
- ^ Виитасало, С.; Лехтиниеми М. и Катаджисто Т. (2008). «Инвазивный гребневик Mnemiopsis leidyi в большом количестве зимует в субарктическом Балтийском море». Журнал исследований планктона . 30 (12): 1431–1436. дои : 10.1093/plankt/fbn088 .
- ^ Трамбл, В.; Браун, Л. (2002). «Гребневик». Краткий Оксфордский словарь английского языка . Издательство Оксфордского университета.
- ^ Руперт, Э.Э.; Фокс, Р.С. и Барнс, Р.Д. (2004). Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр. 76–97 . ISBN 978-0-03-025982-1 .
- ^ Бергквист, PR (1998). «Порифера». В Андерсоне, Д.Т. (ред.). Зоология беспозвоночных . Издательство Оксфордского университета. стр. 10–27. ISBN 978-0-19-551368-4 .
- ^ Перейти обратно: а б с Мороз Леонид Л.; Кокот, Кевин М.; Цитарелла, Мэтью Р.; Досунг, Сон; Норекян Тигран П.; Поволоцкая Инна С.; и др. (июнь 2014 г.). «Геном гребневика и эволюционное происхождение нервных систем» . Природа . 510 (7503): 109–114. Бибкод : 2014Natur.510..109M . дои : 10.1038/nature13400 . ПМЦ 4337882 . ПМИД 24847885 .
- ^ Экспозито, JY.; Клюзель, К.; Гарроне Р. и Летиас К. (2002). «Эволюция коллагенов» . Анатомические записи, часть A: открытия в молекулярной, клеточной и эволюционной биологии . 268 (3): 302–316. дои : 10.1002/ar.10162 . ПМИД 12382326 . S2CID 12376172 .
- ^ Перейти обратно: а б Хорита, Т. (март 2000 г.). «Неописанный лопастной гребневик Lobatolampea Tetragona gen. nov. и spec. nov., представляющий новое семейство из Японии» . Зоологический Медеделинген . 73 (30): 457–464 . Проверено 3 января 2009 г.
- ^ Перейти обратно: а б Хэддок, Стивен HD (ноябрь 2004 г.). «Золотой век джелаты: прошлые и будущие исследования планктонных гребневиков и книдарий». Гидробиология . 530–531 (1–3): 549–556. дои : 10.1007/s10750-004-2653-9 . S2CID 17105070 .
- ^ Мартиндейл, MQ; Генри, JQ (октябрь 1999 г.). «Картирование внутриклеточной судьбы у базальных многоклеточных животных, гребневика Mnemiopsis leidyi , раскрывает происхождение мезодермы и существование неопределенных клеточных линий» . Биология развития . 214 (2): 243–257. дои : 10.1006/dbio.1999.9427 . ПМИД 10525332 .
- ^ Афзелиус, бакалавр (1961). «Тонкое строение ресничек плавательных пластинок гребневиков» . Журнал биофизической и биохимической цитологии . 9 (2): 383–94. дои : 10.1083/jcb.9.2.383 . ПМК 2224992 . ПМИД 13681575 .
- ^ Крепс, Т.А.; Перселл, Дж. Э. и Гейдельберг, КБ (июнь 1997 г.). «Побег гребневика Mnemiopsis leidyi хищника-сцифомедузы Chrysaora quinquecirha от » Морская биология . 128 (3): 441–446. Бибкод : 1997ВторникЧур.128..441K . дои : 10.1007/s002270050110 . S2CID 32975367 .
- ^ Миллс, Клаудия Э. (февраль 1984 г.). «Плотность изменяется у Hydromedusae и гребневиков в ответ на изменения солености» . Биологический вестник . 166 (1): 206–215. дои : 10.2307/1541442 . JSTOR 1541442 .
- ↑ У инопланетных гребневиков нервная система невиданная ранее.
- ^ Медуза с нервной системой, вызывающая дрожь в научном сообществе.
- ^ Перейти обратно: а б Буркхардт, Павел; Колгрен, Джеффри; Медхус, Астрид; Дигель, Леонид; Науманн, Бенджамин; Сото-Анхель, Джоан; Нордманн, Ева-Лена; Сачкова, Мария; Киттельманн, Майке (20 апреля 2023 г.). «Синцитиальная нервная сеть гребневика добавляет понимание эволюции нервных систем» . Наука . 380 (6642): 293–297. Бибкод : 2023Sci...380..293B . дои : 10.1126/science.ade5645 . ПМИД 37079688 . S2CID 258239574 . Проверено 24 апреля 2023 г.
- ^ Парри, Люк А.; Лерози-Обрил, Руди; Уивер, Джеймс К.; Ортега-Эрнандес, Хавьер (2021). «Кембрийские гребневики из Юты освещают раннюю эволюцию нервной и сенсорной систем гребневиков» . iScience . 24 (9): 102943. Бибкод : 2021iSci...24j2943P . дои : 10.1016/j.isci.2021.102943 . ПМЦ 8426560 . ПМИД 34522849 .
- ^ Пенниси, Элизабет (10 января 2019 г.). «Клейкие щупальца гребневиков, возможно, появились, когда впервые появились нервные клетки». Наука . дои : 10.1126/science.aaw6288 . S2CID 92852830 .
- ^ Развилась ли нервная система гребневиков независимо?
- ^ Марлоу, Хизер; Тошес, Мария Антониетта; Томер, Раджу; Стейнмец, Патрик Р.; Лаури, Антонелла; Ларссон, Томас; Арендт, Детлев (29 января 2014 г.). «Оформление тела личинок и апикальные органы сохраняются в эволюции животных» . БМК Биология . 12 (1): 7. дои : 10.1186/1741-7007-12-7 . ПМЦ 3939940 . ПМИД 24476105 .
- ^ Нильсен, Клаус (15 февраля 2015 г.). «Личиночная нервная система: настоящая личинка и не по годам развитая взрослая особь» . Журнал экспериментальной биологии . 218 (4): 629–636. дои : 10.1242/jeb.109603 . ПМИД 25696826 . S2CID 3151957 .
- ^ Нильсен, Клаус (июль 2019 г.). «Ранняя эволюция животных: взгляд морфолога» . Королевское общество открытой науки . 6 (7): 190638. Бибкод : 2019RSOS....690638N . дои : 10.1098/rsos.190638 . ПМК 6689584 . ПМИД 31417759 .
- ^ Фокс, Дуглас (1 августа 2017 г.). «Чужие среди нас» . Эон . Проверено 1 августа 2017 г.
- ^ Хьюитт, Оливия Х.; Дегнан, Сэнди М. (13 февраля 2023 г.). «Антиоксидантные ферменты, действующие на перекись водорода, сохранились во всем животном мире, от губок до млекопитающих» . Научные отчеты . 13 (1): 2510. Бибкод : 2023NatSR..13.2510H . дои : 10.1038/s41598-023-29304-6 . ПМЦ 9925728 . ПМИД 36781921 . S2CID 256811787 .
- ^ Норекян Тигран П.; Мороз, Леонид Л. (15 августа 2019 г.). «Разнообразие нервной системы и рецепторов гребневика Beroe abyssicola» . Журнал сравнительной неврологии . 527 (12): 1986–2008. дои : 10.1002/cne.24633 . ПМИД 30632608 .
- ^ Симмонс, Дэвид К.; Мартиндейл, Марк К. (2015). «Ктенофора». Строение и эволюция нервных систем беспозвоночных . Издательство Оксфордского университета. стр. 48–55. doi : 10.1093/acprof:oso/9780199682201.003.0006 . ISBN 9780199682201 .
- ^ Жакели, Гаспар; Папс, Хорди; Нильсен, Клаус (2015). «Филогенетическое положение гребневиков и происхождение нервных систем» . ЭвоДево . 6 (1): 1. дои : 10.1186/2041-9139-6-1 . ПМК 4406211 . ПМИД 25905000 .
- ^ Пять типов колобласта в гребневике цидиппида, Minictena luteola Carré и Carré: ультраструктурное исследование и цитологическая интерпретация.
- ^ Характеристика функционального биоразнообразия всего типа Ctenophora с использованием физиологических измерений.
- ^ Миллс, CE; Миллер, Р.Л. (февраль 1984 г.). «Проглатывание медузы ( Aegina citrea ) содержащим нематоцисты гребневиком ( Haeckelia rubra , ранее Euchroma rubra ): филогенетические последствия». Морская биология . 78 (2): 215–221. Бибкод : 1984МарБи..78..215М . дои : 10.1007/BF00394704 . S2CID 17714037 .
- ^ Маки, ГО; Миллс, CE; Сингла, CL (март 1988 г.). «Структура и функции цепких щупалец Euplokamis (Ctenophora, Cydippida)». Зооморфология . 107 (6): 319–337. дои : 10.1007/BF00312216 . S2CID 317017 .
- ^ Перейти обратно: а б Крейг, CL; Окубо, А. (апрель 1990 г.). «Физические ограничения на эволюцию размера и формы гребневика». Эволюционная экология . 4 (2): 115–129. Бибкод : 1990EvEco...4..115C . дои : 10.1007/BF02270909 . S2CID 24584197 .
- ^ Перейти обратно: а б Тамм, Сидни Л. (1973). «Механизмы координации ресничек у гребневиков». Журнал экспериментальной биологии . 59 : 231–245. дои : 10.1242/jeb.59.1.231 .
- ^ Перейти обратно: а б с д и ж Хэддок, SHD (декабрь 2007 г.). «Сравнительное пищевое поведение планктонных гребневиков». Интегративная и сравнительная биология . 47 (6): 847–853. дои : 10.1093/icb/icm088 . ПМИД 21669763 .
- ^ Тамм, СЛ; Тамм, С. (1985). «Визуализация изменений конфигурации кончиков ресничек, вызванных скользящим смещением микротрубочек в макроцилиях гребневика Beroe ». Журнал клеточной науки . 79 : 161–179. дои : 10.1242/jcs.79.1.161 . ПМИД 3914479 .
- ^ Тамм, Сидни Л.; Тамм, Сигнхильд (1991). «Обратимая эпителиальная адгезия закрывает рот Бероэ , плотоядного морского желе» . Биологический бюллетень . 181 (3): 463–473. дои : 10.2307/1542367 . JSTOR 1542367 . ПМИД 29304670 .
- ^ Гиббонс, Марк Дж.; Ричардсон, Энтони Дж.; В. Анхель, Мартин; Бючер, Эммануэль; Эсналь, Грасиела; Фернандес Аламо, Мария А.; Гибсон, Рэй; Ито, Хироши; Пью, Фил; Беттгер-Шнак, Рут; Туесен, Эрик (июнь 2005 г.). «Что определяет вероятность обнаружения видов в морском голозоопланктоне: важен ли размер, ареал или глубина?». Ойкос . 109 (3): 567–576. Бибкод : 2005Ойкос.109..567Г . дои : 10.1111/j.0030-1299.2005.13754.x .
- ^ Врубель, Дэвид; Миллс, Клаудия (2003) [1998]. Пелагические беспозвоночные Тихоокеанского побережья: Путеводитель по обычным студенистым животным . Sea Challengers и аквариум Монтерей-Бэй. стр. 108 . ISBN 978-0-930118-23-5 .
- ^ Мартиндейл, MQ (декабрь 1986 г.). «Онтогенез и поддержание свойств симметрии взрослых особей у гребневика Mnemiopsis mccradyi ». Биология развития . 118 (2): 556–576. дои : 10.1016/0012-1606(86)90026-6 . ПМИД 2878844 .
- ^ Сассон, Дэниел А.; Райан, Джозеф Ф. (декабрь 2017 г.). «Реконструкция половых способов эволюции животных» . Эволюционная биология BMC . 17 (1): 242. Бибкод : 2017BMCEE..17..242S . дои : 10.1186/s12862-017-1071-3 . ПМК 5717846 . ПМИД 29207942 .
- ^ Эдгар, Эллисон; Пончано, Хосе Мигель; Мартиндейл, Марк К. (2022). «Гребневики являются непосредственными разработчиками, которые непрерывно размножаются, начиная очень рано после вылупления» . Труды Национальной академии наук . 119 (18): e2122052119. Бибкод : 2022PNAS..11922052E . дои : 10.1073/pnas.2122052119 . ПМК 9170174 . ПМИД 35476523 .
- ^ Харбисон, Греция; Миллер, Р.Л. (февраль 1986 г.). «Не все гребневики являются гермафродитами. Исследования систематики, распространения, сексуальности и развития двух видов Ocyropsis». Морская биология . 90 (3): 413–424. Бибкод : 1986MarBi..90..413H . дои : 10.1007/bf00428565 . S2CID 83954780 .
- ^ Райтцель, AM; Панг, К.; Мартиндейл, MQ (2016). «Экспрессия развития генов, связанных с «зародышевой линией» и «определением пола», у гребневика Mnemiopsis leidyi » . Еводево . 7:17 . дои : 10.1186/s13227-016-0051-9 . ПМЦ 4971632 . ПМИД 27489613 .
- ^ Ясперс, К.; Харальдссон, М.; Болте, С.; Ройш, ТБ; Тайгесен, UH; Кьёрбо, Т. (2012). «Популяция гребневиков пополняется полностью за счет личиночного размножения в центральной части Балтийского моря» . Письма по биологии . 8 (5): 809–12. дои : 10.1098/rsbl.2012.0163 . ПМК 3440961 . ПМИД 22535640 .
- ^ Мороз, Леонид; Мукерджи, Кришану; Романова, Дарья (2023). «Передача сигналов оксида азота в гребневиках» . Передний. Нейроски . 17 : 1125433. дои : 10.3389/fnins.2023.1125433 . ПМЦ 10073611 . ПМИД 37034176 .
- ^ Перейти обратно: а б с д Миллс, CE (май 2007 г.). «Тип Ctenophora: список всех действительных научных названий» . Проверено 10 февраля 2009 г.
- ^ Уэлч, Виктория; Виньерон, Жан Поль; Лусс, Вирджиния; Паркер, Эндрю (14 апреля 2006 г.). «Оптические свойства радужного органа гребневика Beroë cucumis (Ctenophora)». Физический обзор E . 73 (4): 041916. Бибкод : 2006PhRvE..73d1916W . дои : 10.1103/PhysRevE.73.041916 . ПМИД 16711845 .
- ^ Хэддок, SHD; Кейс, Дж. Ф. (декабрь 1995 г.). «Не все гребневики биолюминесцентны: плевробрачия» . Биологический вестник . 189 (3): 356–362. дои : 10.2307/1542153 . JSTOR 1542153 . ПМИД 29244577 .
- ^ Хэддок, SHD; Кейс, Дж. Ф. (8 апреля 1999 г.). «Спектры биолюминесценции мелководного и глубоководного студенистого зоопланктона: гребневиков, медуз и сифонофоров». Морская биология . 133 (3): 571–582. Бибкод : 1999MarBi.133..571H . дои : 10.1007/s002270050497 . S2CID 14523078 .
- ^ Шницлер, Кристин Э.; Панг, Кевин; Пауэрс, Меган Л.; Райтцель, Адам М.; Райан, Джозеф Ф.; Симмонс, Дэвид; Тада, Такаши; Парк, Морган; Гупта, Джьоти; Брукс, Шелис Ю.; Блейксли, Роберт В.; Ёкояма, Сёдзо; Хэддок, Стивен HD; Мартиндейл, Марк К.; Баксеванис, Андреас Д. (2012). «Геномная организация, эволюция и экспрессия генов фотобелков и опсинов у Mnemiopsis leidyi: новый взгляд на фотоциты гребневиков» . БМК Биология . 10 (1): 107. дои : 10.1186/1741-7007-10-107 . ПМК 3570280 . ПМИД 23259493 .
- ^ Винников, Джейкоб Р.; Хэддок, Стивен HD; Будин, Италия (01 ноября 2021 г.). «Адаптация жирных кислот в зависимости от глубины и температуры у гребневиков из экстремальных местообитаний» . Журнал экспериментальной биологии . 224 (21): jeb242800. дои : 10.1242/jeb.242800 . ПМЦ 8627573 . ПМИД 34676421 .
- ^ Эль-Шабрави, Гамаль; Дюмон, Анри (2016). «Первое упоминание гребневика в озерах: гребневик Mnemiopsis leidyi A. Agassiz, 1865 вторгается в Фаюм, Египет» . Отчеты о биоинвазиях . 5 (1): 21–24. дои : 10.3391/бир.2016.5.1.04 .
- ^ Эль-Шабрави, Гамаль; Дюмон, Анри (2016). «Первая запись гребневика в озерах: гребневик Mnemiopsis leidyi A. Agassiz, 1865 вторгается в Фаюм, Египет». Отчеты о биоинвазиях . 5 (1): 21–24. дои : 10.3391/бир.2016.5.1.04 . S2CID 90377075 .
- ^ Мохаммед-Геба, Халед; к. Шеир, Шерин; Игл, Роберт; б. Огберн, Мэтью; час Хайнс, Энсон; Дж. Халафалла, Хусейн; Эль-Каттан, Ахмед; е. Хассаб Эль-Наби, Собхи; Галал-Халлаф, Астма (2019). «Молекулярное и морфологическое подтверждение инвазивного американского изопода; Livoneca redmanii Leach, 1818, от Средиземноморья до озера Карун, Египет» (PDF) . Египетский журнал водной биологии и рыболовства . 23 (4): 251–273. дои : 10.21608/ejabf.2019.54062 .
- ^ Рив, MR; Уолтер, Массачусетс; Икеда, Т. (июль 1978 г.). «Лабораторные исследования приема пищи и использования пищи лопастными и щупальчатыми гребневиками 1: Утилизация пищи гребневиками» . Лимнология и океанография . 23 (4): 740–751. дои : 10.4319/lo.1978.23.4.0740 .
- ^ Костелло, Дж. Х.; Ковердейл, Р. (октябрь 1998 г.). «Планктонное питание и эволюционное значение лопастного строения тела гребневиков». Биологический вестник . 195 (2): 247–248. дои : 10.2307/1542863 . JSTOR 1542863 . ПМИД 28570175 .
- ^ Бос АР; Круз-Ривера Э.; Документ АМ (2016). «Травоядные рыбы Siganus rivulatus (Siganidae) и Zebrasoma desjardinii (Acanthuridae) питаются гребневиками и сцифозоами в Красном море». Морское биоразнообразие . 47 : 243–246. дои : 10.1007/ s12526-016-0454-9 S2CID 24694789 .
- ^ Арай, Мэри Нидлер (июнь 2005 г.). «Хищничество пелагических кишечнополостных: обзор». Журнал Морской биологической ассоциации Соединенного Королевства . 85 (3): 523–536. Бибкод : 2005JMBUK..85..523A . дои : 10.1017/S0025315405011458 . S2CID 86663092 .
- ^ Перейти обратно: а б Чанди, Шонали Т.; Грин, Чарльз Х. (июль 1995 г.). «Оценка хищнического воздействия студенистого зоопланктона» . Лимнология и океанография . 40 (5): 947–955. Бибкод : 1995LimOc..40..947C . дои : 10.4319/lo.1995.40.5.0947 .
- ^ Филд, Кристофер Б.; Беренфельд, Майкл Дж.; Рандерсон, Джеймс Т.; Фальковски, Пол (10 июля 1998 г.). «Первичная продукция биосферы: интеграция наземных и океанических компонентов» . Наука . 281 (5374): 237–240. Бибкод : 1998Sci...281..237F . дои : 10.1126/science.281.5374.237 . ПМИД 9657713 .
- ^ Перейти обратно: а б с Перселл, Дженнифер Э.; Шиганова Тамара А.; Декер, Мэри Бет; Худ, Эдвард Д. (2001). «Гребневик Мнемиопсис в аборигенных и экзотических средах обитания: эстуарии США и бассейн Черного моря». Гидробиология . 451 (1/3): 145–176. дои : 10.1023/А:1011826618539 . S2CID 23336715 .
- ^ Перейти обратно: а б Огуз, Т.; Фач Б. и Салихоглу Б. (декабрь 2008 г.). «Динамика инвазии чужеродного гребневика Mnemiopsis leidyi и ее влияние на гибель хамсы в Черном море» . Журнал исследований планктона . 30 (12): 1385–1397. дои : 10.1093/plankt/fbn094 .
- ^ Байха, КМ; Харбисон, Р.; Макдональд, Дж. Х.; Гаффни, премьер-министр (2004). Предварительное исследование молекулярной систематики инвазивного гребневика Beroe ovata . Серия наук НАТО: IV: Науки о Земле и окружающей среде. Том. 35. С. 167–175. дои : 10.1007/1-4020-2152-6_7 . ISBN 978-1-4020-1866-4 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Камбурская, Л. (2006). «Воздействие Beroe cf ovata на студенистый и другой зоопланктон болгарского побережья Черного моря». В Дюмоне, Х.; Шиганова Т.А.; Нирманн, У. (ред.). Водные инвазии в Черное, Каспийское и Средиземное моря . Серия наук НАТО: IV: Науки о Земле и окружающей среде. Том. 35. Спрингер Нидерланды. стр. 137–154. дои : 10.1007/1-4020-2152-6_5 . ISBN 978-1-4020-1866-4 .
- ^ «Гребенчатые желейные нейроны вызывают дебаты об эволюции» . Журнал Кванта . 25 марта 2015 г. Проверено 12 июня 2015 г.
- ^ Уилан, Невада; Кокот, КМ; Мороз, ТП; Мукерджи, К; Уильямс, П; Поли, Дж; Мороз, LL; Галаныч, К.М. (2017). «Отношения с гребневиками и их положение в качестве сестринской группы для всех других животных» . Нат Эколь Эвол . 1 (11): 1737–1746. Бибкод : 2017NatEE...1.1737W . дои : 10.1038/s41559-017-0331-3 . ПМЦ 5664179 . ПМИД 28993654 .
- ^ Конвей Моррис, С. (2003). «Кембрийский «взрыв» многоклеточных животных и молекулярная биология: будет ли Дарвин удовлетворен?» (PDF) . Международный журнал биологии развития . 47 (7–8): 505–515. ПМИД 14756326 . Архивировано из оригинала (PDF) 24 декабря 2009 г. Проверено 14 февраля 2009 г.
- ^ Клюг, Кристиан; Керр, Джоанна; Ли, Майкл С.Ю.; Клотье, Ришар (2021). «Поздно сохранившийся стебель-гребневик из позднего девона Мигуаши (Канада)» . Научные отчеты . 11 (1): 19039. Бибкод : 2021NatSR..1119039K . дои : 10.1038/s41598-021-98362-5 . ПМЦ 8463547 . ПМИД 34561497 .
- ^ Максмен, Эми (7 сентября 2011 г.). «Древняя морская медуза потрясает эволюционное древо животных» . Научный американец . Проверено 21 июня 2018 г.
- ^ Шу, Д.-Г.; Моррис, Южная Каролина; Хан, Дж; Ли, Ю; Чжан, XL; Хуа, Х; Чжан, ZF; Лю, JN; Го, JF; Яо, Ю; Ясуи, К. (5 мая 2006 г.). «Вендобионты нижнего кембрия из Китая и эволюция раннего диплобласта». Наука . 312 (5774): 731–734. Бибкод : 2006Sci...312..731S . дои : 10.1126/science.1124565 . ПМИД 16675697 . S2CID 1235914 .
- ^ Чжао, Ян; Винтер, Якоб; Парри, Люк А.; Вэй, Фань; Грин, Эмили; Пизани, Давиде; Хоу, Сянгуан; Эджкомб, Грегори Д.; Конг, Пейюнь (апрель 2019 г.). «Кембрийские сидячие, гребневики стеблевой группы, питающиеся суспензией, и эволюция плана гребневидного желеобразного тела» . Современная биология . 29 (7): 1112–1125.e2. дои : 10.1016/j.cub.2019.02.036 . ПМИД 30905603 .
- ^ Чжао, Ян; Хоу, Сяньгуан; Конг, Пэй-юнь (1 января 2023 г.). «Щупальцевая природа «колонны» кембрийского диплобласта Xianguangia sinica» . Журнал систематической палеонтологии . 21 (1). Бибкод : 2023JSPal..2115787Z . дои : 10.1080/14772019.2023.2215787 . ISSN 1477-2019 .
- ^ Минди, Вайсбергер (10 июля 2015 г.). «У древних медуз были колючие скелеты, но не было щупалец» . www.livscience.com .
- ^ Саймон Конвей Моррис, Альберто М. Симонетта, изд. (1991). Ранняя эволюция Metazoa и значение проблемных таксонов . Издательство Кембриджского университета. п. 308. ИСБН 978-0-521-11158-4 .
- ^ Нильсен, Клаус; Шарфф, Николай; Эйби-Якобсен, Дэнни (апрель 1996 г.). «Кладистический анализ животного мира» . Биологический журнал Линнеевского общества . 57 (4): 385–410. Бибкод : 1996BJLS...57..385N . дои : 10.1006/bijl.1996.0023 .
- ^ Лейкарт, Рудольф (1923). О морфологии и взаимоотношениях беспозвоночных животных. Вклад в характеристику и классификацию форм животных . ISBN 978-1-245-56027-6 .
- ^ Геккель, Эрнст Генрих Филипп Август (1896). Систематическая филогения беспозвоночных, Invertebrata, Часть 2: Проект систематической филогении . ISBN 978-1-120-86850-3 .
- ^ Хайман, Либби Генриетта (1940). Беспозвоночные: Том I, Простейшие и гребневики . МакГроу Хилл. ISBN 978-0-07-031660-7 .
- ^ Филипп, Х.; Дерелл, Р.; Лопес, П.; Пик, К.; Борчеллини, К.; Бури-Эсно, Н.; Вейселет, Дж.; Ренар, Э.; Хоулистон, Э.; Кеннек, Э.; Да Силва, К.; Винкер, П.; Ле Гаядер, Х.; Лейс, С.; Джексон, диджей; Шрайбер, Ф.; Эрпенбек, Д.; Моргенштерн, Б.; Верхайде, Г.; Мануэль, ML (28 апреля 2009 г.). «Филогеномика возрождает традиционные взгляды на глубокие взаимоотношения животных» . Современная биология . 19 (8): 706–712. Бибкод : 2009CBio...19..706P . дои : 10.1016/j.cub.2009.02.052 . ПМИД 19345102 . S2CID 15282843 .
- ^ Валлберг, А.; Толлессон, М.; Фаррис, Дж. С.; Джонделиус, У. (декабрь 2004 г.). «Филогенетическое положение гребневиков (Ctenophora) и значение таксономической выборки» . Кладистика . 20 (6): 558–578. дои : 10.1111/j.1096-0031.2004.00041.x . ПМИД 34892961 . S2CID 86185156 .
- ^ Коллинз, АГ (2002). «Филогения Medusozoa и эволюция жизненных циклов книдарий» . Журнал эволюционной биологии . 15 (3): 418–432. дои : 10.1046/j.1420-9101.2002.00403.x . S2CID 11108911 .
- ^ Перейти обратно: а б Пик, Канзас; Филипп, Х.; Шрайбер, Ф.; Эрпенбек, Д.; Джексон, диджей; Вреде, П.; Винс, М.; Алие, А.; Моргенштерн, Б.; Мануэль, М.; Верхайде, Г. (сентябрь 2010 г.). «Улучшенный отбор филогеномных таксонов заметно влияет на недвусторонние отношения» . Молекулярная биология и эволюция . 27 (9): 1983–1987. дои : 10.1093/molbev/msq089 . ПМЦ 2922619 . ПМИД 20378579 .
- ^ Хейнол, А.; Обст, М.; Стаматакис, А.; Отт, М.; Роуз, GW; Эджкомб, Джорджия; Мартинес, П.; Багуна, Дж.; Байи, X.; Йонделиус, У.; Винс, М.; Мюллер, WEG; Сивер, Э.; Уилер, туалет; Мартиндейл, MQ; Гирибет, Г.; Данн, CW (22 декабря 2009 г.). «Оценка корня двулатеральных животных с помощью масштабируемых филогеномных методов» . Труды Королевского общества B: Биологические науки . 276 (1677): 4261–4270. дои : 10.1098/rspb.2009.0896 . ПМК 2817096 . ПМИД 19759036 .
- ^ Райан, Дж. Ф.; Панг, К.; Программа сравнительного секвенирования; Малликин, Дж. К.; Мартиндейл, MQ; Баксеванис, А.Д.; Программа сравнительного секвенирования NISC (2010 г.). «Гомеодоменный состав гребневика Mnemiopsis leidyi предполагает, что Ctenophora и Porifera разошлись до ParaHoxozoa» . ЭвоДево . 1 (1): 9. дои : 10.1186/2041-9139-1-9 . ПМК 2959044 . ПМИД 20920347 .
- ^ Райтцель, AM; Панг, К.; Райан, Дж. Ф.; Малликин, Дж. К.; Мартиндейл, MQ; Баксеванис, А.Д.; Таррант, AM (2011). «Ядерные рецепторы гребневика Mnemiopsis leidyi лишены ДНК-связывающего домена с цинковыми пальцами: специфичная для линии потеря или наследственное состояние при возникновении суперсемейства ядерных рецепторов?» . ЭвоДево . 2 (1): 3. дои : 10.1186/2041-9139-2-3 . ПМК 3038971 . ПМИД 21291545 .
- ^ Панг, К.; Райан, Дж. Ф.; Программа сравнительного секвенирования NISC; Малликин, Дж. К.; Баксеванис, А.Д.; Мартиндейл, MQ (2010). «Геномное понимание передачи сигналов Wnt у раннего дивергентного многоклеточного животного, гребневика Mnemiopsis leidyi » . ЭвоДево . 1 (1): 10. дои : 10.1186/2041-9139-1-10 . ПМК 2959043 . ПМИД 20920349 .
- ^ Либескинд, Б.Дж.; Хиллис, DM; Закон, Х.Х. (2011). «Эволюция натриевых каналов предшествует возникновению нервной системы у животных» . Труды Национальной академии наук . 108 (22): 9154–9159. Бибкод : 2011PNAS..108.9154L . дои : 10.1073/pnas.1106363108 . ПМК 3107268 . ПМИД 21576472 .
- ^ Перейти обратно: а б Райан, Дж. Ф.; Панг, К.; Шницлер, CE; Нгуен, А.-Д.; Морленд, RT; Симмонс, ДК; Кох, Б.Дж.; Фрэнсис, WR; Хавлак, П.; Смит, ЮАР; Патнэм, Нью-Хэмпшир; Хэддок, SHD; Данн, CW; Вольфсберг, ТГ; Малликин, Дж. К.; Мартиндейл, MQ; Баксеванис, А.Д. (13 декабря 2013 г.). «Геном гребневика Mnemiopsis leidyi и его значение для эволюции типов клеток» . Наука . 342 (6164): 1242592. doi : 10.1126/science.1242592 . ПМЦ 3920664 . ПМИД 24337300 .
- ^ Уилан, Натан В.; Кокот, Кевин М.; Мороз Леонид Л.; Галанич, Кеннет М. (5 мая 2015 г.). «Ошибка, сигнал и расположение сестры гребневиков ко всем остальным животным» . Труды Национальной академии наук . 112 (18): 5773–5778. Бибкод : 2015PNAS..112.5773W . дои : 10.1073/pnas.1503453112 . ПМЦ 4426464 . ПМИД 25902535 .
- ^ Боровец, Марек Л.; Ли, Эрнест К.; Чиу, Джоанна С.; Плашецки, Дэвид К. (декабрь 2015 г.). «Извлечение филогенетического сигнала и учет систематической ошибки в наборах полногеномных данных подтверждают, что Ctenophora является сестрой остальных Metazoa» . БМК Геномика . 16 (1): 987. doi : 10.1186/s12864-015-2146-4 . ПМЦ 4657218 . ПМИД 26596625 .
- ^ Филипп, Эрве; Дерелль, Ромен; Лопес, Филипп; Пик, Керстин; Борчеллини, Кэрол; Бури-Эно, Николь; Васелет, Жан; Ренар, Эммануэль; Хоулистон, Эвелин; Кеннек, Эрик; Да Силва, Коринн; Винкер, Патрик; Ле Гюядер, Эрве; Лейс, Салли; Джексон, Дэниел Дж.; Шрайбер, Фабиан; Эрпенбек, Дирк; Моргенштерн, Буркхард; Верхайде, Герт; Мануэль, Микаэль (апрель 2009 г.). «Филогеномика возрождает традиционные взгляды на глубокие взаимоотношения животных» . Современная биология . 19 (8): 706–712. Бибкод : 2009CBio...19..706P . дои : 10.1016/j.cub.2009.02.052 . ПМИД 19345102 . S2CID 15282843 .
- ^ Носенко Татьяна; Шрайбер, Фабиан; Адамска, Майя; Адамски, Марцин; Эйтель, Майкл; Хаммель, Йорг; Мальдонадо, Мануэль; Мюллер, Вернер Э.Г.; Никель, Майкл (1 апреля 2013 г.). «Глубокая филогения многоклеточных животных: когда разные гены рассказывают разные истории». Молекулярная филогенетика и эволюция . 67 (1): 223–233. Бибкод : 2013МОЛПЭ..67..223Н . дои : 10.1016/j.ympev.2013.01.010 . ПМИД 23353073 .
- ^ Пизани, Давиде; Петт, Уокер; Дорманн, Мартин; Феуда, Роберто; Рота-Стабелли, Омар; Филипп, Эрве; Лартильо, Николя; Вёрхайде, Герт (15 декабря 2015 г.). «Геномные данные не подтверждают, что гребневики являются сестринской группой всех других животных» . Труды Национальной академии наук . 112 (50): 15402–15407. Бибкод : 2015PNAS..11215402P . дои : 10.1073/pnas.1518127112 . ПМЦ 4687580 . ПМИД 26621703 .
- ^ Капли, Пасхалия; Телфорд, Максимилиан Дж. (11 декабря 2020 г.). «Зависимая от топологии асимметрия систематических ошибок влияет на филогенетическое расположение Ctenophora и Xenacoelomorpha» . Достижения науки . 6 (10): eabc5162. Бибкод : 2020SciA....6.5162K . дои : 10.1126/sciadv.abc5162 . ПМЦ 7732190 . ПМИД 33310849 .
- ^ Кристиансон, LM; Джонсон, С.Б.; Шульц, Д.Т.; Хэддок, Ш. (2021). «Скрытое разнообразие гребневиков, выявленное с помощью новых митохондриальных праймеров и последовательностей COI» . Ресурсы молекулярной экологии . 22 (1): 283–294. дои : 10.1111/1755-0998.13459 . ПМК 9290464 . ПМИД 34224654 .
- ^ Кон, AB; Цитарелла, MR; Кокот, КМ; Бобкова Ю.В.; Галаныч, К.М.; Мороз, LL (2011). «Быстрая эволюция компактного и необычного митохондриального генома гребневика Pleurobrachia bachei» . Молекулярная филогенетика и эволюция . 63 (1): 203–207. дои : 10.1016/j.ympev.2011.12.009 . ПМК 4024468 . ПМИД 22201557 .
- ^ Бакшани, Кэсси Р.; Моралес-Гарсия, Ана Л.; Альтхаус, Майк; Уилкокс, Мэтью Д.; Пирсон, Джеффри П.; Байтелл, Джон К.; Берджесс, Дж. Грант (4 июля 2018 г.). «Эволюционное сохранение антимикробной функции слизи: первая защита от инфекции» . npj Биопленки и микробиомы . 4 (1): 14. дои : 10.1038/s41522-018-0057-2 . ПМК 6031612 . ПМИД 30002868 .
- ^ В мозг гребневиков: ученые исследуют эволюцию нейронов
- ^ Миллс, Д.Б.; Фрэнсис, WR; Варгас, С; Ларсен, М; Элеманс, КП; Кэнфилд, Делавэр; Верхайде, Г (2018). «У последнего общего предка животных не было пути HIF, и он дышал в среде с низким содержанием кислорода» . электронная жизнь . 7 . doi : 10.7554/eLife.31176 . ПМК 5800844 . ПМИД 29402379 .
- ^ Гауберг, Джулия; Абдаллах, Сальсабиль; Эльхатиб, Васим; Харраксингх, Алисия Н.; Пикут, Томас; Стэнли, Элиза Ф.; Сенатор, Адриано (25 декабря 2020 г.). «Консервативные биофизические особенности гомолога пресинаптического Ca2+-канала CaV2 у рано расходящихся животных Trichoplax adhaerens» . Журнал биологической химии . 295 (52): 18553–18578. дои : 10.1074/jbc.RA120.015725 . ПМЦ 7939481 . ПМИД 33097592 .
- ^ Эволюция и развитие - страница 38. Архивировано 2 марта 2014 г. в Wayback Machine.
- ^ Хиберт, Лорел С.; Маслакова, Светлана А. (2015). «Гены Hox моделируют передне-заднюю ось молоди, но не личинки у беспозвоночных Micrura alaskensis (Nemertea) с максимально непрямым развитием» . БМК Биология . 13:23 . дои : 10.1186/s12915-015-0133-5 . ПМЦ 4426647 . ПМИД 25888821 .
- ^ Гонсиоровски, Людвик; Хейнол, Андреас (2019). «Экспрессия гена Hox во время развития форонид Phoronopsis Harmeri» . Еводево . 11 :2. bioRxiv 10.1101/799056 . дои : 10.1186/s13227-020-0148-z . ПМК 7011278 . ПМИД 32064072 . S2CID 208578827 .
- ^ Независимая радиационная передача сигналов иннексина в гребневиках
- ^ Перейти обратно: а б Подар, Мирча; Хэддок, Стивен HD; Согин, Митчелл Л.; Харбисон, Г. Ричард (ноябрь 2001 г.). «Молекулярная филогенетическая основа типа Ctenophora с использованием генов 18S рРНК». Молекулярная филогенетика и эволюция . 21 (2): 218–230. Бибкод : 2001МОЛПЭ..21..218П . CiteSeerX 10.1.1.384.6705 . дои : 10.1006/mpev.2001.1036 . ПМИД 11697917 .
- ^ Харбисон, Греция (1985). «О классификации и эволюции гребневиков». В Конвее Моррисе, С.; Джордж, доктор юридических наук; Гибсон, Р.; Платт, Х.М. (ред.). Происхождение и взаимоотношения низших беспозвоночных . Кларендон Пресс. стр. 78 –100. ISBN 978-0-19-857181-0 .
- ^ Симион, Пол; Беккуш, Николя; Джагер, Мюриэль; Кеннек, Эрик; Мануэль, Микаэль (2015). «Изучение потенциала малых субъединиц РНК и последовательностей ITS для разрешения филогенетических взаимоотношений внутри типа Ctenophora». Зоология . 118 (2): 102–114. Бибкод : 2015Zool..118..102S . дои : 10.1016/j.zool.2014.06.004 . ПМИД 25440713 .
Дальнейшее чтение
[ редактировать ]- РСК Барнс, П. Калоу, П.Дж.В. Олив, Д.В. Голдинг, Дж.И. Спайсер, Беспозвоночные – синтез , 3-е изд., Блэквелл, 2001, гл. 3.4.3, с. 63, ISBN 0-632-04761-5
- Р. К. Бруска, Г. Дж. Бруска, Беспозвоночные , 2-е изд., Sinauer Associates, 2003, гл. 9, с. 269, ISBN 0-87893-097-3
- Дж. Мур, Введение в беспозвоночных , Кембриджский университет. Пресс, 2001, гл. 5.4, с. 65, ISBN 0-521-77914-6
- В. Шефер, Гребневики, гребневики , в книге В. Вестхайде и Р. Ригер: Специальная зоология, том 1 , Густав Фишер Верлаг, Штутгарт, 1996 г.
- Бруно Венцель, Стеклянные животные моря. Гребневик (Acnidaria) , 1958 г., ISBN 3-7403-0189-9
- Марк Шаша, «Ночь лунных желе» , 1992, Simon & Schuster, ISBN 0-671-77565-0
- Дуглас Фокс, «Инопланетяне среди нас: Что гребневик говорит об эволюции интеллекта» , 2017, Aeon.co.
Внешние ссылки
[ редактировать ] Определения из Викисловаря
Определения из Викисловаря  СМИ из палаты общин
СМИ из палаты общин  Таксоны из Wikispecies
Таксоны из Wikispecies  Данные из Викиданных
Данные из Викиданных
- Plankton Chronicles Короткометражные документальные фильмы и фотографии
- Обзор медуз и гребешков на Смитсоновском океаническом портале
- Гребневики из пролива Сан-Себастьян, Бразилия.
- Видео гребневиков в Национальном зоопарке в Вашингтоне, округ Колумбия
- Древо животной жизни переставило ветви, по мнению биологов-эволюционистов
- Информационный бюллетень об австралийских гребневиках
- The Jelly Connection – поразительные изображения, в том числе Beroe на другого гребневика. нападение экземпляра
- В поисках первых животных