Euscociality

Euscocialility ( греческий εὖ Eu «добрый» и социальный ) является самым высоким уровнем организации социальности . Это определяется следующими характеристиками: уход за кооперативным выводом (включая уход за потомством других людей), перекрывающиеся поколения в колонии взрослых и разделение труда на репродуктивные и нерепродуктивные группы. Разделение труда создает специализированные поведенческие группы в обществе животных, которые иногда называют «кастами». Эусоциальность отличается от всех других социальных систем, потому что люди, по крайней мере, одной касты, обычно теряют способность выполнять поведение, характерное для людей в другой касте. Euscocial Colories можно рассматривать как супер -супер -ведущие .
Эусоциальность развивалась среди насекомых , ракообразных , трематоды и млекопитающих . Это наиболее распространено в гименоптере ( муравьях , пчелах и осах ) и в блюттодии ( термиты ). Колония имеет кастовые различия: королевы и репродуктивные мужчины играют роль единственных воспроизводителей, в то время как солдаты и работники работают вместе, чтобы создать и поддерживать жизненную ситуацию, благоприятную для вывода. Королевы производят несколько феромонов королевы для создания и поддержания эусоциального состояния в своих колониях; Они также могут есть яйца, отложенные другими женщинами или оказывать доминирование, боя. Есть два эуссоциальных грызуна : голый моль-рат и моль Дамараланда . Некоторые креветки , такие как Synalpheus Regalis , являются эусоциальными. Эо Уилсон и другие утверждают, что люди развили слабую форму эусоциальности. Было высказано предположение, что колониальный и эпифитный папоротник Стагорн также может использовать примитивное эусоциальное разделение труда.
История
[ редактировать ]
Термин «euscocial» был введен в 1966 году Сюзанной Батра , которая использовала его для описания поведения гнездования у пчел Галектидов по шкале субсоциального/одиночного, колониального/общинного, полусоциального и эусоциального, где колония начинается отдельным человеком. Полем [ 1 ] [ 2 ] Бэтра наблюдала кооперативное поведение пчел, мужчин и женщин, поскольку они взяли на себя ответственность по крайней мере за одну обязанность (то есть, норму, клеточная конструкция, яйцекласс ) в колонии. Кооперативность была важна, поскольку деятельность одного подразделения труда сильно повлияла на деятельность другого. Эусоциальные колонии можно рассматривать как супер -супер -ведущие , причем отдельные касты аналогичны различным ткани или типам клеток в многоклеточном организме ; Касты выполняют определенную роль, которая способствует функционированию и выживанию всей колонии, в то же время не способствуя независимому выживанию за пределами колонии. [ 3 ]
В 1969 году Чарльз Д. Михенер [ 4 ] Дальнейшая расширение классификации Батры с его сравнительным изучением социального поведения у пчел. Он наблюдал несколько видов пчел ( апоидеа ), чтобы исследовать различные уровни социальной активности животных, все из которых представляют собой различные этапы, через которые может пройти колония. Euscociality, который является самым высоким уровнем социальной социальности животных, который может достичь вида, в частности, имела три характеристики, которые отличали его от других уровней: [ 1 ]
- Яичные лайиры и люди, похожие на работница среди взрослых женщин (разделение труда)
- Перекрытие поколений (потомство матери и взрослых)
- Кооперативная работа на клетках соты пчел

EO Wilson расширил терминологию, чтобы включить других социальных насекомых, таких как муравьи, осы и термиты. Первоначально он был определен как включать организмы (только беспозвоночные), которые имели следующие три функции: [ 1 ] [ 5 ] [ 6 ] [ 7 ]
- Репродуктивное разделение труда (со стерильными кастами или без него)
- Перекрывающиеся поколения
- Совместная забота о молодых
Эусоциальность была затем обнаружена в группе хордовых , моль-карат. Дальнейшие исследования отличали еще один, возможно, важный критерий для эусоциальности, «смысл отсутствия возврата». Это характеризуется тем, что люди фиксируются в одну поведенческую группу, обычно до репродуктивной зрелости. Это мешает им переходить между поведенческими группами и создает общество с людьми, которые действительно зависят друг от друга для выживания и репродуктивного успеха. Для многих насекомых эта необратимость изменила анатомию рабочей касты, которая является стерильной и обеспечивает поддержку репродуктивной касты. [ 1 ] [ 7 ]
Разнообразие
[ редактировать ]Большинство эусоциальных обществ существуют в членистоногих , в то время как некоторые из них встречаются у млекопитающих . Некоторые папоротники могут демонстрировать примитивную форму эусоциального поведения. [ 8 ] [ 9 ]
У насекомых
[ редактировать ]Euscociality развивалась несколько раз в разных порядках насекомых, включая Hymenopterans, [ 10 ] термиты, [ 11 ] трипсы, [ 12 ] тля, [ 12 ] и жуки. [ 13 ]
В Hymenoptera
[ редактировать ]
Орден Hymenoptera содержит самую большую группу эусоциальных насекомых, включая муравьев , пчел и ошей, разделившихся на касты: репродуктивные королевы , беспилотники , более или менее стерильные работники, а иногда и солдаты, которые выполняют специализированные задачи. [ 14 ] В хорошо изученной социальной осе Полистик Versicolor , [ 15 ] Доминирующие женщины выполняют такие задачи, как строительство новых клеток и яйцеклеток, в то время как подчиненные женщины, как правило, выполняют такие задачи, как кормление личинок и кормление. Дифференциация задач между кастами можно увидеть в том факте, что подчиненные завершают 81,4% от общего объема пищи, в то время как доминанты завершают только 18,6% от общего числа корм. [ 16 ] Эусоциальные виды со стерильной кастой иногда называют гиперсоциальным. [ 17 ]
В то время как только умеренный процент видов у пчел (семейства Apidae и Halictidae ) и ос ( Crabronidae и Vespidae ) являются эуссоциальными, почти все виды муравьев ( Formicidae ) являются эуссоциальными. [ 18 ] Некоторые основные линии осы в основном или полностью эусоциальные, включая подсемейства Polistinae и Vespinae . Корбикулятные пчелы (подсемейство апинаи семейства апиде ) содержат четыре племена различной степени социальной жизни: высокоусоциальные апини (медоносные пчелы) и мелипонини (пчелы без укусов), примитивно эуссоциальные бомбар (шкмель) и в основном одинокие или слабо социальные эйгилоссини и эуссоциальные бомба (орхидеи пчел). [ 19 ] Эусоциальность в этих семьях иногда управляется набором феромонов , которые изменяют поведение специфических каст в колонии. Эти феромоны могут действовать по разным видам, как это наблюдалось в APIS Andriformis (Black Dwarf Honey Bee), где рабочие пчелы отвечали на королеву -феромон от соответствующих API Florea (Red Dwarf Honey Bee). [ 20 ] Феромоны иногда используются в этих кастах, чтобы помочь в поиске пищи. Например, работники австралийской тетрагуновой пчелиной Carbonaria отмечают источники пищи с феромоном, помогая своим товарищам по гнездам найти пищу. [ 21 ]

Репродуктивная специализация, как правило, включает в себя производство стерильных членов вида, которые выполняют специализированные задачи для ухода за репродуктивными членами. Люди могут иметь поведение и морфология, модифицированная для защиты групп, включая самоотверженное поведение . Например, члены стерильной касты муравьев Honeypot, таких как Myrmecocystus, наполняют свои животные жидкими пищами, пока они не станут неподвижными и не висят на потолках подземных гнезд, выступая в качестве хранилища пищи для остальной части колонии. [ 22 ] Не все социальные насекомые имеют различные морфологические различия между кастами. Например, в неотропической социальной осе Synoeca Surinama ряды определяются социальными дисплеями в развивающемся выводке. [ 23 ] Эти касты иногда дополнительно специализируются на своем поведении в зависимости от возраста, как у Scaptotrigona Postica работников . В возрасте от 0 до 40 дней рабочие выполняют задачи в гнезде, такие как обеспечивающие вывода клеток, чистка колоний, прием нектара и обезвоживание. Получив старше 40 дней, работники S. postica выходят за пределы гнезда для защиты колонии и питания. [ 24 ]
В Lasioglossum aeneiventre , пчел Галектичной из Центральной Америки, гнезда могут возглавлять более чем одну женщину; Такие гнезда имеют больше клеток, а количество активных клеток на самку коррелирует с количеством женщин в гнезде, что подразумевает, что наличие большего количества женщин приводит к более эффективному строительству и обеспечению клеток. [ 25 ] У аналогичных видов только одна королева, такая как ласиоглоссам малачурум в Европе, степень эусоциальности зависит от климата, в котором обнаружен вид. [ 26 ]
В термитах
[ редактировать ]
Термиты (порядок Blattodea , Infraorder Isoptera ) составляют еще одну большую часть высокопроизводительных эусоциальных животных. Колония дифференцирована в различные касты: королева и король - единственные воспроизводимые люди; работники добывают и поддерживают продукты питания и ресурсы; [ 27 ] и солдаты защищают колонию от атак муравья. Последние две касты, которые являются стерильными и выполняют высокоспециализированные, сложные социальные поведения, получены из разных стадий плюрипотентных личинок, продуцируемых репродуктивной кастой. [ 11 ] У некоторых солдат есть челюсти настолько расширенные (специализированные на обороне и атаку), что они не могут прокормить себя и должны кормить работники. [ 28 ]
В жуках
[ редактировать ]Austroplatypus oncompertus является видом амброзийского жука, родом из Австралии, и является первым жуком (порядок Coleoptera ), который будет признан как Eusocial. [ 29 ] [ 13 ] Этот вид образует колонии, в которых одна самка оплодотворяется и защищена многими неплощенными женщинами, которые служат работниками, расковивающими туннели на деревьях. У этого вида есть кооперативный уход за выводами, в котором люди заботятся о несовершеннолетних, которые не являются их собственными. [ 13 ]
У индивидуальных насекомых
[ редактировать ]
Некоторые индуцирующие желчные насекомые , в том числе галлевая тля тисаноптера , пемфигус Spyrothecae (порядок гемиптера ), и трипсы , такие как кладотрипсы (порядок ) , описаны как эусоциальные. [ 12 ] [ 30 ] Эти виды имеют очень высокое родство среди людей из-за их бесполого репродукции (стерильные солдатские касты являются клонами, полученными в результате партеногенеза ), но поведение, восушенное желчими, дает этим видам защитный ресурс. Они производят солдатские касты для защиты крепости и защиты колонии от хищников, клептопаразитов и конкурентов. В этих группах эусоциальность производится как с высокой родственностью, так и живой в ограниченной общей области. [ 31 ] [ 32 ]
В ракообразных
[ редактировать ]Euscociality развивалась в трех разных линиях в роде ракообразных колониальных Synalphaeus . S. Regalis , S. Microneptunus , S. filidigitus , S. Elizabethae , S. Chacei , S. Riosi , S. Duffyi и S. Cayoneptunus - восемь зарегистрированных видов паразитических креветок, которые полагаются на защиту крепости и живут группами Связанные люди в тропических рифах и губках. [ 33 ] Они живут с эско- Для членов колонии есть одно общее жилое пространство, а не размножающиеся члены действуют, чтобы защитить его. [ 34 ]
Гипотеза защиты крепости дополнительно указывает, что, поскольку губки обеспечивают как еду, так и укрытие, существует агрегация родственников (потому что креветки не должны рассеиваться, чтобы найти еду) и много конкуренции за эти гнездовывающие места. Быть цели атаки способствует хорошей системе защиты (солдатская каста); Солдаты продвигают пригодность всего гнезда, обеспечивая безопасность и размножение королевы. [ 35 ]
Euscociality предлагает конкурентное преимущество в популяциях креветок. Euscocial виды более распространены, занимают больше среды обитания и используют больше доступных ресурсов, чем неосощественные виды. [ 36 ] [ 37 ] [ 38 ]
В трематоде
[ редактировать ]Трематоды представляют собой класс паразитического плоского червя, также известный как Flukes. Один вид, Haplorchis Pumilio , развил Eusociality с участием колонии, создающей класс стерильных солдат. [ 39 ] Один случайность вторгается в хозяина и устанавливает колонию десятков тысяч клонов, которые работают вместе, чтобы захватить ее. Поскольку конкурирующие виды трематод могут вторгаться и заменить колонию, она защищена специализированной касой стерильных солдат трематод. [ 40 ] Солдаты меньше, более мобильны и развиваются по другому пути, чем сексуально зрелые репродуктивности. Одно отличие состоит в том, что солдатские чашки (глотка) в пять раз больше, чем у воспроизведений. Они составляют почти четверть объема солдата. Эти солдаты не имеют зародышевой массы, никогда не метаморфоза, чтобы быть репродуктивными и, следовательно, являются обязательно стерильными. [ 40 ] Солдаты легко отличаются от незрелых и зрелых репродуктивных червей. Солдаты более агрессивны, чем репродуктивные, атакуя гетероспецифические трематоды, которые заражают их хозяина in vitro . Интересно, что солдаты Х. Пумилио не нападают на конспецифических из других колоний. Солдаты не распределены по всему телу хозяина. Они встречаются в самых высоких количествах в базальной висцеральной массе, где конкурирующие трематоды имеют тенденцию к умножению во время ранней фазы инфекции. Это стратегическое позиционирование позволяет им эффективно защищаться от захватчиков, аналогично тому, как модели распределения солдат видны у других животных с защитными кастами. Они «кажутся обязательно стерильной физической кастой, сродни калике самых продвинутых социальных насекомых». [ 40 ]
У нечеловеческих млекопитающих
[ редактировать ]
Среди млекопитающих два вида в группе грызунов Phiomorpha являются эуссоциальными, голый моль-рат ( гетероцефалия глабер ) и моль-краю Дамараланда ( Fukomys Damarensis ), оба из которых высоко внедрены . [ 41 ] Обычно живущие в суровой или ограничивающей среде, эти моль-скалы помогают воспитывать братьев и сестер и родственников, рожденных от одной репродуктивной королевы. Тем не менее, эта классификация является спорной из -за оспариваемых определений «eusociality». Чтобы избежать инбридинга , крысы молей иногда вытекают и устанавливают новые колонии, когда ресурсов достаточно. [ 42 ] Большинство людей совместно заботятся о выводке одной репродуктивной женщины (королевы), с которой они, скорее всего, связаны. Таким образом, неясно, действительно ли крот -крысы действительно эусоциальными, поскольку их социальное поведение в значительной степени зависит от их ресурсов и окружающей среды. [ 43 ]
Некоторые млекопитающие в плотоядных и приматах имеют эуссоциальные тенденции, особенно сурикаты ( Suricata Suricatta ) и гномов в монгулах ( Helogale parvula ). Они демонстрируют кооперативное разведение и заметные репродуктивные перекосы. В карликовом мангузе размножающаяся пара получает приоритет и защиту пищи от подчиненных и редко должна защищать от хищников. [ 44 ]
У людей
[ редактировать ]Ученые обсуждают, являются ли люди просоциальными или эуссоциальными. [ 45 ] Эдвард О. Уилсон назвал людей евросоциальными обезьянами, выступая за сходство с муравьями, и наблюдая, что ранние гоминины сотрудничают, чтобы воспитывать своих детей, в то время как другие члены той же группы охотились и охотились. [ 46 ] Уилсон и другие утверждали, что благодаря сотрудничеству и командной работе, муравьи и люди формируют супер -суперганизма. [ 47 ] [ 48 ] [ 49 ] Претензии Уилсона были энергично отвергнуты критиками теории отбора групп , которая основала аргумент Уилсона, [ 46 ] [ 50 ] [ 51 ] и потому что человеческий репродуктивный труд не разделен между кастами. [ 50 ]
Хотя спорно, [ 52 ] Было высказано предположение, что мужская гомосексуальность [ 53 ] и женская менопауза [ 54 ] мог бы развиваться с помощью отбора родственников . [ 55 ] [ 56 ] Это будет означать, что у людей иногда проявляется тип аллопарированного поведения, известного как « помощники в гнезде », с несовершеннолетними и сексуально зрелыми подростками, помогающими своим родителям поднять последующие выводки, как у некоторых птиц, [ 57 ] Некоторые неэусоциальные пчелы и сурикаты . [ 58 ] Эти виды не являются эуссоциальными: у них нет каст, и помощники воспроизводят самостоятельно, если это будет предоставлено. [ 59 ] [ 48 ] [ 60 ]
В растениях
[ редактировать ]
Одно растение, эпифитный папоротник Staghorn, Platycerium bifurcatum ( Polypodiaceae ), может демонстрировать примитивную форму эусоциального поведения среди клонов. Доказательства этого заключаются в том, что люди живут в колониях, где они структурированы по -разному, с листьями разного размера и формы, чтобы собирать и хранить воду и питательные вещества для использования колонии. На вершине колонии есть как плиссированные вентиляционные «гнездо», которые собирают и удерживают воду, и ветвы в форме желоба «ремешок», которые направляют воду: ни один уединенный виды платцерия не имеет обоих типов. В нижней части колонии есть «гнездо» листья, которые зажигают ствол дерева, поддерживающего папоротник, и опускают фотосинтетические листья. Утверждается, что они адаптированы для структурной поддержки колонии, то есть, что люди в колонии в некоторой степени специализируются на задачах, разделение труда . [ 8 ] [ 9 ] [ 61 ]
Эволюция
[ редактировать ]Филогенетическое распределение
[ редактировать ]Euscociality является редким, но широко распространенным явлением у видов, по крайней мере, у семи порядков в животном царстве , как показано в филогенетическом дереве (не показанные группы не показаны). Все виды термитов являются эуссоциальными, и считается, что они были первыми эуссоциальными животными, которые развивались, когда -то в верхний период юрского периода (~ 150 миллионов лет назад). [ 62 ] Другие показанные порядки содержат как Eusocial, так и неэусоциальные виды, в том числе многие линии, где предполагается, что Eusociality является исконным состоянием. Таким образом, число независимых эволюций эусоциальности ( клад ) неизвестно. Основные эусоциальные группы показаны в жирном сайте в филогенетическом дереве.
| Эукариоты |
| ||||||










Парадокс
[ редактировать ]До гена, ориентированного на эволюцию , эусоциальность рассматривалась как парадоксальная: если адаптивная эволюция разворачивается в результате дифференциального воспроизводства отдельных организмов, эволюция людей, неспособных передать их гены, представляет собой проблему. В отношении происхождения видов Дарвин назвал существование стерильных каст как «единственную особую трудность, которая сначала казалась мне непреодолимым и фактически смертельным для моей теории». [ 63 ] Дарвин ожидал, что возможное разрешение на парадокс может лежать в близких семейных отношениях, которые, как Хэмильтон, оценил столетие спустя столетие своей теорией инклюзивной физической подготовки 1964 года . После того, как в середине 1970-х годов был разработан ген-ориентированный взгляд на эволюцию, нерепродуктивные индивидуумы рассматривались как расширенный фенотип генов, которые являются основными бенефициарами естественного отбора. [ 64 ]
Инклюзивная фитнес и гаплодиплоидия
[ редактировать ]Аргумент, что галлодиплоидия способствует эусоциальности
[ редактировать ]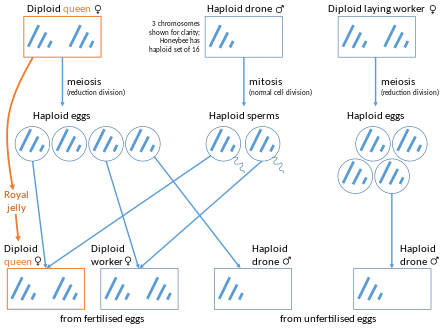
Согласно инклюзивной теории физической подготовки, организмы могут получить пригодность, увеличивая репродуктивную продукцию других людей, которые разделяют свои гены, особенно их близких родственников. Естественный отбор благоприятствует людям, чтобы помочь своим родственникам, когда стоимость помощи меньше, чем выгода, полученная из -за их относительной, умноженной на долю генов, которые они разделяют, то есть, когда затрат <родственность * . В.Д. Гамильтон предположил в 1964 году, что эусоциальность может легче развиваться среди гаплодиплоидных видов, таких как гименоптера, из -за их необычной структуры родства. [ 65 ] [ 66 ] [ 67 ]
У гаплодиплоидных видов самки развиваются из оплодотворенных яиц, а мужчины развиваются из неплощенных яиц. Поскольку мужчина гаплоид, его дочери имеют 100% его генов и 50% их матери. Поэтому они имеют 75% своих генов друг с другом. Этот механизм определения пола порождает то, что В.Д. Гамильтон впервые назвал «суперсхранениями», более тесно связанным со своими сестрами, чем они были бы с их собственным потомством. [ 67 ] Несмотря на то, что работники часто не воспроизводят, они могут передавать больше своих генов, помогая воспитывать своих сестер, чем иметь собственного потомства (каждый из которых будет иметь только 50% своих генов). Эта необычная ситуация, когда женщины могут иметь большую пригодность, когда они помогают задним сестрам, а не производить потомство, часто вызывается, чтобы объяснить множественные независимые эволюции эусоциальности (по меньшей мере девять раз) в гименоптере. [ 68 ]
Аргумент, что галлодиплоидия не способствует эусоциальности
[ редактировать ]Против предполагаемых преимуществ гаплодиплоидии для эусоциальности Роберт Триверс отмечает, что, хотя женщины имеют 75% генов со своими сестрами в галлодиплоидных популяциях, они имеют только 25% своих генов со своими братьями. [ 69 ] Соответственно, средняя родство человека с их братом составляет 50%. Следовательно, поведение помощи - это выгодно, если оно предвзято помогать сестрам, что привело бы население до полового соотношения 1: 3 к женщинам. В этом соотношении мужчины, как более редкий пол, увеличивают репродуктивную ценность, снижая выгоду от инвестиций, представленных женщинами. [ 70 ]
Кроме того, не все эуссоциальные виды являются гаплодиплоидными: термиты, некоторые щекоты, креветки и моль -крысы нет. И наоборот, многие неэусоциальные пчелы являются гаплодиплоидными, а среди эусоциальных видов многие королевы спариваются с несколькими мужчинами, что приводит к тому, что улей сводных сестер, которые имеют только 25% своих генов. Связь между гаплодиплоидией и эусоциальностью ниже статистической значимости. [ 71 ] Таким образом, галлодиплоидия не является ни необходимой, ни достаточной для появления эусоциальности. [ 72 ] Связанность все еще играет роль, так как моногамия (Queens Maring Oningly) является наследственным состоянием для всех исследованных Eusocial Views. [ 73 ] Если отбор родственников является важной силой, стимулирующей эволюцию эусоциальности, моногамия должна быть исконным состоянием, поскольку она максимизирует связанность членов колонии. [ 73 ]
Эволюционная экология
[ редактировать ]Повышенные показатели паразитизма и хищничества являются основными экологическими факторами социальной организации. Group Living обеспечивает защиту членов колонии от врагов, в частности хищников, паразитов и конкурентов, и позволяет им получить преимущество от превосходных методов корма. [ 7 ] Важность экологии в эволюции эусоциальности подтверждается такими доказательствами, как экспериментально индуцированное репродуктивное разделение труда, например, когда обычно одиночные королевы объединяются. [ 74 ] И наоборот, женщины Дамараланда проходят гормональные изменения, которые способствуют рассеиванию после периодов высокого количества осадков. [ 75 ]
Климат также, по -видимому, является селективным агентом, способствующим социальной сложности; В целом по линии пчел и гименоптерам более высокие формы социальности чаще встречаются в тропических, чем умеренная среда. [ 76 ] Аналогичным образом, социальные переходы в пчелах Галиктид , где эусоциальность была получена и потеряна несколько раз, коррелируют с периодами климатического потепления. Социальное поведение в факультативных социальных пчелах часто надежно предсказывается экологическими условиями, а переключения в поведенческом типе экспериментально вызваны путем перемещения потомства одиночного или социального населения в теплый и прохладный климат. В H. Rubicundus женщины производят один выводка в более прохладных регионах и два или более выводках в более теплых регионах, поэтому первое население является одинокой, а последняя - социальная. [ 77 ] У другого вида пчел пота L. Calceatum , социальный фенотип был предсказан с помощью высоты и состава микрохабитата, с социальными гнездами, обнаруженными в более теплых, солнечных местах и одиночных гнездах, найденных в соседних, прохладных, затененных местах. Однако на факультитивно социальные виды пчел, которые включают в себя большинство социальных пчелиных разнообразий, имеют самое низкое разнообразие в тропиках, в значительной степени ограничиваясь умеренными регионами. [ 78 ]
Многоуровневый выбор
[ редактировать ]После того, как присутствуют предварительные адаптации, такие как формирование групп, строительство гнезд, высокая стоимость рассеивания и морфологические различия, конкуренция между группами была предложена в качестве движущей силы перехода к расширенному эусоциальности. Ма Новак, CE Tarnita и EO Wilson предложили в 2010 году, что, поскольку Eusociality производит чрезвычайно альтруистическое общество, эусоциальные группы должны превзойти своих менее кооперативных конкурентов, в конечном итоге устраняя все неэусоциальные группы из вида. [ 79 ] Многоуровневый отбор подвергся жесткой критике за его конфликт с теорией отбора родственников . [ 80 ]
Обращение к солизности
[ редактировать ]Обращение к солизности - это эволюционное явление, при котором потомки эусоциальной группы снова развивают одиночное поведение. Пчелы были модельными организмами для изучения обращения к солизности из -за разнообразия их социальных систем. Каждому из четырех источников эуссоциальности у пчел последовало по крайней мере одно изменение в солирительность, что дало в общей сложности не менее девяти изменений. [ 4 ] [ 5 ] У некоторых видов одиночные и эуссоциальные колонии появляются одновременно в одной популяции, а различные популяции одного и того же вида могут быть полностью одинокими или эусоциальными. [ 77 ] Это говорит о том, что эусоциальность стоит дорого поддерживать и может сохраняться только тогда, когда экологические переменные предпочитают. Недостатки Eusocialility включают стоимость инвестирования в нерепродуктивное потомство и повышенный риск заболевания. [ 81 ]
Все переводы на солирительность произошли среди примитивно эусоциальных групп; Никто не последовал за появлением продвинутой эусоциальности. Гипотеза «точка отсутствия возврата» утверждает, что морфологическая дифференциация репродуктивных и нерепродуктивных каст предотвращает очень эусоциальные виды, такие как педали, от возврата в одиночное состояние. [ 20 ]
Физиология и развитие
[ редактировать ]Феромоны
[ редактировать ]Феромоны играют важную роль в физиологических механизмах эусоциальности. Ферменты, участвующие в производстве и восприятии феромонов, были важны для появления эусоциальности как в термитах, так и в гименоптерах. [ 82 ] Лучшая изучаемая система феромонов королевы у социальных насекомых-это система APIS Mellifera Honey Bee Mellifera . Королева нижнечелюстной железы производит смесь из пяти соединений, трех алифатических и двух ароматических , которые контролируют работников. [ 83 ] Экстракты нижней челюсти железы ингибируют работников строить клетки королевы, что может задержать гормонально поведенческое развитие работников и подавлять их развитие яичников. [ 84 ] [ 83 ] Как поведенческие эффекты, опосредованные нервной системой, часто приводят к распознаванию королев ( релизер ), так и физиологическое воздействие на репродуктивную и эндокринную систему ( праймер ) связаны с одними и теми же феромонами. Эти феромоны улегают или деактивируются в течение тридцати минут, что позволяет работникам быстро реагировать на потерю своей королевы. [ 84 ]
Уровни двух из алифатических соединений быстро увеличиваются у Virgin Queens в течение первой недели после появления куколки , что согласуется с их ролями в качестве половых аттрактантов во время спаривания. [ 83 ] Как только королева спаривается и начинает откладывать яйца, она начинает производить полную смесь соединений. [ 83 ] У некоторых видов муравья репродуктивная активность связана с выработкой феромонов королевами. [ 83 ] Королевы с сожаленными яйцами привлекательны для работников, в то время как молодые крылатые девственные королевы получают небольшой или нет ответа. [ 83 ]

Среди муравьев система королевы феромонов из Fire Ant Solenopsis Invicta включает в себя как релизеры, так и феромоны праймера. Феромон признания королевы (релизера) хранится в ядовитым мешках вместе с тремя другими соединениями. Эти соединения вызывают поведенческий ответ от работников. Несколько эффектов праймеров также были продемонстрированы. Феромоны инициируют репродуктивное развитие у новых крылатых женщин, называемых женскими сексуальными. [ 83 ] Эти химические вещества препятствуют выращиванию мужчин и женщин, подавляют производство яиц в других королевах многочисленных колоний королевы и заставляют работников выполнять избыточные королевы. [ 83 ] [ 84 ] Эти феромоны поддерживают эусоциальный фенотип, причем одна королева поддерживается стерильными работниками и сексуально активными мужчинами ( беспилотники ). В колониях без королевы отсутствие феромонов королевы заставляет крылатых женщин быстро сбросить свои крылья, развивать яичники и откладывать яйца. Эти девственные замены королевы играют роль королевы и начинают производить феромоны королевы. [ 83 ] Точно так же муравьи Queen Weaver oecophylla Longinoda имеют экзокринные железы, которые производят феромоны, которые мешают работникам откладывать репродуктивные яйца. [ 84 ]
Подобные механизмы существуют в эусоциальной осе vespula vulgaris . Чтобы королева доминировала на всех рабочих, обычно насчитывая более 3000 в колонии, она сигнализирует о своем доминировании с феромонами. Рабочие регулярно облизывают королеву, кормя ее, и воздушный феромон из тела королевы предупреждает тех рабочих о ее доминировании. [ 85 ]
Способ действия ингибирующих феромонов, которые предотвращают развитие яиц у работников, был продемонстрирован в Bumble Bee Bombus Tarrestris . [ 84 ] Феромоны подавляют активность эндокринной железы, корпус -аллатума , помешая ей секретировать юношеский гормон . [ 86 ] С низким юношеским гормоном яйца не созревают. Подобные ингибирующие эффекты снижения юношеского гормона были замечены у галектиктинских пчел и полислиновых ос, но не у медоносных пчел. [ 84 ]
Другие механизмы
[ редактировать ]Разнообразные другие механизмы дают королевам различных видов социальных насекомых меру репродуктивного контроля над своими товарищами по гнездам. Во многих осах Полиститов моногамия устанавливается вскоре после формирования колоний путем физического доминирования взаимодействия между основаниями колонии, включая укусы, погоню за проживанием и запрашиванием пищи. Такие взаимодействия создают иерархию доминирования, возглавляемая более крупными, пожилыми людьми с наибольшим развитием яичников. Ранг подчиненных коррелирует со степенью развития яичников. [ 84 ] Рабочие не яйцеклеты , когда присутствуют королевы, по разным причинам: колонии, как правило, достаточно малы, чтобы королевы могли эффективно доминировать в работниках; Квинс практикует селективную офагию ; Поток питательных веществ способствует королеве работникам; и королевы быстро откладывают яйца в новых или освобожденных клетках. [ 84 ]
У примитивно-эусоциальных пчелах (где касты морфологически схожи, а колонии маленькие и недолговечны), королевы часто подталкивают своих скотоводства, а затем возвращаются в гнездо. Это привлекает работников в нижнюю часть гнезда, где они могут реагировать на стимулы для строительства и технического обслуживания клеток. [ 84 ] Быть подталкиваемой королевой может помочь препятствовать развитию яичников; Кроме того, королева ест яйца, отложенные работниками. [ 84 ] Кроме того, временное дискретное производство работников и гинев (фактических или потенциальных королев) может вызвать диморфизмы размера между различными кастами, поскольку размер сильно зависит от сезона, в течение которого на индивидуальность выращивается. Во многих осах каста работников определяется временной схемой, в которой работники предшествуют неработникам того же поколения. [ 87 ] В некоторых случаях, например, в шмелях контроль королевы ослабляется в конце сезона, и развиваются яичники работников. [ 84 ] Королева пытается сохранить свое доминирование путем агрессивного поведения и употребления яйца, приложенных работником; Ее агрессия часто направлена на работника с величайшим развитием яичников. [ 84 ]
В очень эусоциальных ос (где касты морфологически разнородны), как количество, так и качество пищи важны для дифференцировки касты. [ 84 ] Недавние исследования в Осах предполагают, что дифференциальное питание личинок может быть экологическим спуском для личиночной дивергенции для работников или Гиновых. [ 87 ] Все личинки для медоносных пчел изначально питаются королевским желе , которое секретируется работниками, но обычно они переключаются на диету пыльцы и меда по мере их появления; Если их диета является исключительно королевским желе, они растут больше, чем обычно, и дифференцируются в королевы. Это желе содержит специфический белок, Royalactin, который увеличивает размер тела, способствует развитию яичников и сокращает период времени развития. [ 88 ] Дифференциальная экспрессия в полислаях личиночных генов и белков (также дифференциально экспрессируется во время развития королевы и касты у медоносных пчел) указывает на то, что регуляторные механизмы могут работать очень рано в разработке. [ 87 ]
В популярной культуре
[ редактировать ]Научно -фантастическая фантастика Стивена Бакстера 2003 года Коалцентняя представляет собой человеческую эусоциальную организацию, основанную в древнем Риме , в которой большинство людей подвергаются репродуктивной репрессии. [ 89 ] Гарольд Фромм, рассмотренный нащупание для групп Эо Уилсона и других других в обзоре Гудзона , спрашивает, будет ли Уилсон «желание» для людей, чтобы заставить «постоянный рай для людей» означать «быть выбранным групповым на фабриках в стиле из Хаксли [роман 1932 года] «Храбрый новый мир» . [ 90 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый и Креспи, Бернард Дж .; Янега, Дуглас (1995). «Определение эусоциальности». Поведенческая экология . 6 : 109–115. doi : 10.1093/beheco/6.1.109 .
- ^ Jump up to: а беременный Батра, Сюзанна, WT (1 сентября 1966 г.). «Гнезда и социальное поведение галектиновых пчел Индии (Hymenoptera: Halictidae)» . Индийский журнал энтомологии . 28 (3): 375–393.
- ^ Opachaloemphan, Comzit; Ян, Хуа; Лейбхольц, Александра; Десплан, Клод; Рейнберг, Дэнни (2018-11-23). «Последние достижения в области поведенческой (EPI) генетики у эусоциальных насекомых» . Ежегодный обзор генетики . 52 (1): 489–510. doi : 10.1146/annurev-genet-120116-024456 . ISSN 0066-4197 . PMC 6445553 . PMID 30208294 .
- ^ Jump up to: а беременный Миченера, Чарльз Д. (1969). «Сравнительное социальное поведение пчел». Ежегодный обзор энтомологии . 14 : 299–342. doi : 10.1146/annurev.en.14.010169.001503 .
- ^ Jump up to: а беременный Гадагкар, Рагхавендра (1993). "А теперь ... Eusocial Trips!". Текущая наука . 64 (4): 215–216.
- ^ Уилсон, Эдвард О. (1971). «3 Социальные осы; 4 муравья; 6 термитов». Общества насекомых . Кембридж, штат Массачусетс: издательство Belknap of Harvard University Press . ISBN 9780674454903 .
- ^ Jump up to: а беременный в Уилсон, Эдвард О . ; Hölldobler, Bert (20 сентября 2005 г.). «Euscociality: происхождение и последствия» . ПНА . 102 (38): 13367–13371. Bibcode : 2005pnas..10213367W . doi : 10.1073/pnas.0505858102 . PMC 1224642 . PMID 16157878 .
- ^ Jump up to: а беременный Престон, Элизабет (2 июля 2021 года). «Эти растения действуют как пчелы в ульях» . Нью -Йорк Таймс . Получено 7 июля 2021 года .
- ^ Jump up to: а беременный Бернс, KC; Хаттон, Ян; Шепард, Лара (14 мая 2021 г.). "Примитивная эуссоциальность на земельном заводе?" Полем Экология . 102 (9): E03373. Bibcode : 2021ecol..102e3373b . doi : 10.1002/ecy.3373 . ISSN 0012-9658 . PMID 33988245 . S2CID 234496454 . Получено 7 июля 2021 года .
- ^ Данфорт, Брайан Н. (26 декабря 2001 г.). «Эволюция социальности в примитивной эусоциальной линии пчел» . ПНА . 99 (1): 286–290. doi : 10.1073/pnas.012387999 . PMC 117553 . PMID 11782550 .
- ^ Jump up to: а беременный Thorne, BL (1997). «Эволюция эусоциальности в термитах» . Ежегодный обзор экологии, эволюции и систематики . 28 (11): 27–54. doi : 10.1146/annurev.ecolsys.28.1.27 . PMC 349550 .
- ^ Jump up to: а беременный в Стерн, Д.Л. (1994). «Филогенетический анализ эволюции солдат в семействе Aphid Hormaphididae». Труды Королевского общества . 256 (1346): 203–209. BIBCODE : 1994RSPSB.256..203S . doi : 10.1098/rspb.1994.0071 . PMID 8029243 . S2CID 14607482 .
- ^ Jump up to: а беременный в Кент, DS; Симпсон, да (1992). «Eusciality в жуке Austroplatypus incompertus (Coleoptera: Curculionidae)». Естественные науки . 79 (2): 86–87. Bibcode : 1992nw ..... 79 ... 86K . Doi : 10.1007/bf01131810 . S2CID 35534268 .
- ^ Hölldobler, B. (1990). «8 каста и разделение труда». Муравьи . Кембридж, Массачусетс: Belknap Press . С. 298–318.
- ^ Cervo, Rita (2006). «Полишты осы и их социальные паразиты: обзор». Annales Zoologici Fennici . 43 (5/6): 531–549. JSTOR 23736760 .
- ^ Зара, Фернандо; Balestieri, Хосе (2000). «Поведенческий каталог POLISTES Versicolor Olivier (Vespidae: Polistinae) пост-экипировки колонов». Натуралия . 25 : 301–319.
- ^ Ричардс, Мириам Х. (2019). «Определения социальных признаков влияют на эволюционные выводы: филогенетический подход к улучшению социальной терминологии для пчел». Текущее мнение в науке насекомых . 34 : 97–104. doi : 10.1016/j.cois.2019.04.006 . PMID 31247426 . S2CID 151303496 .
- ^ Петерс, Ральф с.; Крогманн, Ларс; Майер, Кристоф; Донат, Александр; Гинкель, Саймон; и др. (Апрель 2017). «Эволюционная история гименоптера» . Текущая биология . 27 (7): 1013–1018. doi : 10.1016/j.cub.2017.01.027 . HDL : 2434/801122 . PMID 28343967 .
- ^ Кардинал, Софи; Данфорт, Брайан Н. (2011). «Древность и эволюционная история социального поведения у пчел» . Plos один . 6 (6): E21086. Bibcode : 2011ploso ... 621086c . doi : 10.1371/journal.pone.0021086 . PMC 3113908 . PMID 21695157 .
- ^ Jump up to: а беременный Wongvilas, S.; Deowanish, S.; Лим, Дж.; Xie, VRD; Гриффит, Оу; Oldroyd, BP (2010). «Межвидовые и конспецифические слияния колоний в API APIS Andriformis и A. Florea ». Насекомые социо . 57 (3): 251–255. doi : 10.1007/s00040-010-0080-7 . S2CID 8657703 .
- ^ Bartareau, T. (1996). «Формажирование поведения тригоны Carbonaria (Hymenoptera: Apidae) на станциях кормления с множественным выбором». Австралийский журнал зоологии . 44 (2): 143. DOI : 10.1071/ZO9960143 .
- ^ Конвей, Джон Р. (сентябрь 1986). «Биология медовых муравьев». Американский учитель биологии . 48 (6): 335–343. doi : 10.2307/4448321 . JSTOR 4448321 .
- ^ West-Eberhard, MJ (1982). «Природа и эволюция роя в тропических социальных ос (Vespidae, Polistinae, Polybini)». Смитсоновский институт тропических исследований .
- ^ из Веена, JW; Самми, MJ; Монкий, Ф. (ноябрь 1997 г.). «Поведение или дроны в Мелипине (Apidae, Melipioninae)». Насекомые сочи . 44 (4): 435–447. doi : 10 1007/S00000000063 . S2CID 36563930 .
- ^ Wciislo, Wt; Will, A.; Orozco, E.E. (1993). Диаграммы страниц aenoptera (helictidae " Ocean Sociaux 40 : 21–40. doi : 10.1007/bf0 . S2CID 686760 .
- ^ Ричардс, Мириам Х. (2000). «Данные географических различий в социальной организации колоний в обязательно социальной пот пчел, Lasioglossum malachurum Kirby (Hymenoptera; Halictidae)». Канадский журнал зоологии . 78 (7): 1259–1266. doi : 10.1139/Z00-064 .
- ^ Коста-Леонардо А.М., Хайфиг И. (2014). Термитная связь во время различных поведенческих действий . В кн.: Биокоммуникация животных. Дортрихт, Спрингер, 161–190.
- ^ Адамс, ES (1987). «Размер территории и ограничения населения в термитах мангровых деревьев». Журнал экологии животных . 56 (3): 1069–1081. Bibcode : 1987 , Janec..56.1069a . doi : 10.2307/4967 . JSTOR 4967 .
- ^ «Наука: австралийский жук, который ведет себя как пчела» . Новый ученый . 9 мая 1992 года . Получено 2010-10-31 .
- ^ Aoki, S.; Имай, М. (2005). «Факторы, влияющие на долю стерильных солдат в растущих колониях тли». Экология населения . 47 (2): 127–136. Bibcode : 2005Popec..47..127a . doi : 10.1007/s10144-005-0218-z . S2CID 2224506 .
- ^ Креспи Б.Дж. (1992). «Eusciality в австралийских желчных трипах». Природа . 359 (6397): 724–726. Bibcode : 1992natur.359..724c . doi : 10.1038/359724A0 . S2CID 4242926 .
- ^ Стерн, Д.; Foster, W. (1996). «Эволюция солдат в тлях». Биологические обзоры . 71 (1): 27–79. doi : 10.1111/j.1469-185x.1996.tb00741.x . PMID 8603120 . S2CID 8991755 .
- ^ Даффи, Дж. Эмметт; Моррисон, Шерил Л.; Риос, Рубен (2000). «Многочисленное происхождение эусоциальности среди губчатых креветков ( Synalpheus )». Эволюция 54 (2): 503–516. doi : 10.1111/j.0014-3820.2000.tb00053.x . PMID 10937227 . S2CID 1088840 .
- ^ Даффи, JE (1998). «О частоте eusociality в смахивающих креветках (Decapoda: Alpheidae), с описанием второго эусоциального вида» . Бюллетень морской науки . 63 (2): 387–400.
- ^ Даффи, JE (2003). «Экология и эволюция эусоциальности в губчатых креветках». Гены, поведение и эволюция социальных насекомых : 217–254.
- ^ Даффи, JE; Macdonald, KS (2010). «Структура родственников, экология и эволюция социальной организации в креветках: сравнительный анализ» . Труды Королевского общества B: Биологические науки . 277 (1681): 575–584. doi : 10.1098/rspb.2009.1483 . PMC 2842683 . PMID 19889706 .
- ^ Hultgren, KM; Даффи, JE (2012). «Экология филогенетического сообщества и роль социального доминирования в креветках, живущих губками». Экологические письма . 15 (7): 704–713. Bibcode : 2012ecoll..15..704h . doi : 10.1111/j.1461-0248.2012.01788.x . PMID 22548770 .
- ^ Макдональд, KS; Rios, R.; Даффи, JE (2006). «Биоразнообразие, специфичность хозяина и доминирование эуссоциальными видами среди креветок с губчатой альфейдом на рифе барьера Белиза» . Разнообразие и распределения . 12 (2): 165–178. Bibcode : 2006divdi..12..165m . doi : 10.1111/j.1366-9516.2005.00213.x . S2CID 44096968 .
- ^ Ричардс, Мириам Х. (10 сентября 2024 г.). «Социальная эволюция и репродуктивные касты у паразитов трематод». Труды Национальной академии наук . 121 (37). doi : 10.1073/pnas.2414228121 .
- ^ Jump up to: а беременный в Метц, Даниэль К.Г.; Хехингер, Райан Ф. (30 июля 2024 г.). «Каста физического солдата инвазивного, инфицирующего человека плоского червя морфологически экстремально и обязательно стерильной» . Труды Национальной академии наук . 121 (31). doi : 10.1073/pnas.2400953121 . PMC 11295071 . PMID 39042696 .
- ^ Бурда, Х. Ханикатт; Begall, S.; Locker-Grutjen, O.; Шарфф А. (2000). "Является ли голые и общие моль-раты эусоциальными и если да, то почему?" Полем Поведенческая экология и социобиология . 47 (5): 293–303. doi : 10.1007/s002650050669 . S2CID 35627708 . Архивировано с оригинала 2016-03-04 . Получено 2007-11-30 .
- ^ O'Riain, MJ; Faulkes, CG (2008). «Африканские моль-раты: эусоциальность, родство и экологические ограничения». Экология социальной эволюции . Спрингер. С. 207–223. doi : 10.1007/978-3-540-75957-7_10 . ISBN 978-3-540-75956-0 .
- ^ O 'Riain, M.; и др. (1996). «Дисперсивное морф в обнаженной моле». Природа . 380 (6575): 619–621. Bibcode : 1996natur.380..619o . doi : 10.1038/380619A0 . PMID 8602260 . S2CID 4251872 .
- ^ Уильямс, SA; Shattuck, MR (2015). «Экология, долголетие и голые моль-скалы: смешанные последствия социальности?» Полем Труды Королевского общества Лондона B: Биологические науки . 282 (1802): 20141664. DOI : 10.1098/rspb.2014.1664 . PMC 4344137 . PMID 25631992 .
- ^ Фостер, Кевин Р.; Ратниккс, Фрэнсис Л.В. (2005). "Новый эуссоциальный позвоночный?" (PDF) . Тенденции в экологии и эволюции . 20 (7): 363–364. doi : 10.1016/j.tree.2005.05.005 . PMID 16701397 . Архивировано из оригинала (PDF) 2012-03-11 . Получено 2011-04-04 .
- ^ Jump up to: а беременный Гинтис, Герберт (2012). «Столкновение Титанов. Обзор книги« Социальное завоевание Земли »Эдварда О. Уилсона» . Биоссака . 62 (11): 987–991. doi : 10.1525/bio.2012.62.11.8 .
- ^ Kesebir, Селин (2012). «Суперорганизм рассказ о человеческой социальности: как и когда человеческие группы похожи на улей». Обзор личности и социальной психологии . 16 (3): 233–261. doi : 10.1177/1088868311430834 . ISSN 1088-8683 .
- ^ Jump up to: а беременный Фостер, Кевин Р.; Ратниккс, Фрэнсис Л.В. (2005). "Новый эуссоциальный позвоночный?" (PDF) . Тенденции в экологии и эволюции . 20 (7): 363–364. doi : 10.1016/j.tree.2005.05.005 . PMID 16701397 .
- ^ Чу, Кэрол; Buchman-Schmitt, Jennifer M.; Стэнли, Ян Х.; Хом, Мелани А.; Такер, Рэймонд П.; Хаган, Кристофер Р.; Роджерс, Меган Л.; Подлогар, Мэтью С.; Chiurliza, Bruno (2017). «Межличностная теория самоубийства: систематический обзор и метаанализ десятилетия межнациональных исследований» . Психологический бюллетень . 143 (12): 1313–1345. doi : 10.1037/bul0000123 . PMC 5730496 . PMID 29072480 .
- ^ Jump up to: а беременный Докинс, Ричард (24 мая 2012 г.). «Спуск Эдварда Уилсона. Книжный обзор« Социальное завоевание Земли »Эдварда О. Уилсона». Перспектива .
- ^ Пинкер, Стивен . «Неверно очарование группы» . Край . Получено 31 июля 2016 года .
- ^ Крамер, Джос; Монье, Жоэль (2016-04-28). «Кин и многоуровневый отбор в социальной эволюции: бесконечный спор?» Полем F1000Research . 5 : F1000 Faculty Rev - 776. doi : 10.12688/f1000research.8018.1 . ISSN 2046-1402 . PMC 4850877 . PMID 27158472 .
- ^ Вандерлаан, Дуг П.; Рен, Чжиюань; Васи, Пол Л. (2013). «Мужская андрофилия в наследственной среде. Этнологический анализ». Человеческая природа . 24 (4): 375–401. doi : 10.1007/s12110-013-9182-z . PMID 24091924 . S2CID 44341304 .
- ^ Хоукс, Кристен; Коксворт, Джеймс Э. (2013). «Бабушки и эволюция долголетия человека: обзор выводов и будущих направлений». Эволюционная антропология . 22 (6): 294–302. doi : 10.1002/evan.21382 . PMID 24347503 . S2CID 37985774 .
- ^ Хупер, Пол Л.; Гервен, Майкл; Винкинг, Джеффри; Каплан, Хиллард С. (2015-03-22). «Инклюзивная физическая форма и дифференциальная продуктивность во всем жизненном курсе определяют переводы между поколениями в небольшом человеческом обществе» . Труды Королевского общества B: Биологические науки . 282 (1803): 20142808. DOI : 10.1098/rspb.2014.2808 . PMC 4345452 . PMID 25673684 .
- ^ Любинский, Марк (2018). «Эволюционные оправдания для репродуктивных ограничений человека» . Журнал вспомогательного размножения и генетики . 35 (12): 2133–2139. doi : 10.1007/s10815-018-1285-3 . PMC 6289914 . PMID 30116921 .
- ^ Джетц, Уолтер; Рубенштейн, Дастин Р. (2011). «Экологическая неопределенность и глобальная биогеография кооперативной размножения у птиц» . Текущая биология . 21 (1): 72–78. doi : 10.1016/j.cub.2010.11.075 . PMID 21185192 .
- ^ Розенбаум, Стейси; Gettler, Lee T. (2018). «С небольшой помощью ее друзей (и семьи), часть I: экология и эволюция не материнской помощи у млекопитающих» . Физиология и поведение . 193 (Pt A): 1–11. doi : 10.1016/j.physbeh.2017.12.025 . PMID 29933836 . S2CID 49380840 .
- ^ Clutton-Brock, Th ; Ходж, SJ; Flower, TP (2008-09-01). «Размер группы и подавление подчиненного воспроизведения в калахари -сурикатах» . Поведение животных . 76 (3): 689–700. doi : 10.1016/j.anbehav.2008.03.015 . ISSN 0003-3472 . S2CID 53203398 .
- ^ «Форум: континуум Eusciality» . Поведенческая экология . 6 (1): 102–108. 1995. doi : 10.1093/beheco/6.1.102 .
- ^ Бернс, Кевин С. (2021-11-02). «О селективном преимуществе колониальности у папоротников Staghorn (Platycerium bifurcatum, polypodiaceae)» . Сигнализация и поведение растения . 16 (11). doi : 10.1080/15592324.2021.1961063 . ISSN 1559-2324 . PMC 8525959 . PMID 34338155 .
- ^ Торн, Бл; Гримальди, да; Кришна, К. (2001) [2000]. «Ранняя ископаемая история термитов». В Abe, T.; Bignell, de; Хигаши, М. (ред.). Термиты: эволюция, социальность, симбиозы, экология . Kluwer Academic Publishers . С. 77–93.
- ^ Дарвин, Чарльз. О происхождении видов , 1859. Глава 8
- ^ Докинс, Ричард (2016) [1982]. «6. Организмы, группы и мемы: репликаторы или транспортные средства?». Расширенный фенотип . Издательство Оксфордского университета . С. 147–178. ISBN 978-0198788911 .
- ^ v7, pp. 1–16 и 17-52.
- ^ Гамильтон, WD (20 марта 1964 г.). «Генетическая эволюция социального поведения я». Журнал теоретической биологии . 7 (1): 1–16.
- ^ Jump up to: а беременный Гамильтон, WD (20 марта 1964 г.). «Генетическая эволюция социального поведения II». Журнал теоретической биологии . 7 (1): 17–52. Bibcode : 1964jthbi ... 7 ... 17h . doi : 10.1016/0022-5193 (64) 90039-6 . PMID 5875340 .
- ^ Quiñones, Andrés E.; Пен, IDO (23 июня 2017 г.). «Единая модель гименоптеранов, которые вызывают эволюционный переход к эйусоциальности» . Природная связь . 8 : 15920. Bibcode : 2017natco ... 815920Q . doi : 10.1038/ncomms15920 . PMC 5490048 . PMID 28643786 .
- ^ Триверс, Роберт Л .; Заяц, Хоуп (1976). «Гаплодиплоидия и эволюция социальных насекомых». Наука . 191 (4224): 249–263. Bibcode : 1976sci ... 191..249t . doi : 10.1126/science.1108197 . PMID 1108197 .
- ^ Alpedrinha, João; Запад, Стюарт А.; Гарднер, Энди (2013). «Гаплодиплоидия и эволюция эусоциальности: воспроизводство работников» . Американский натуралист . 182 (4): 421–438. doi : 10.1086/671994 . HDL : 10023/5520 . PMID 24021396 . S2CID 6548485 .
- ^ Новак, Мартин; Тарнита, Корина; Уилсон, Эдвард О. (26 августа 2010 г.). «Эволюция эусоциальности» . Природа . 466 (7310): 1057–1062. BIBCODE : 2010NATR.466.1057N . doi : 10.1038/nature09205 . PMC 3279739 . PMID 20740005 .
- ^ Уилсон, Эдвард О. (2008-01-01). «Один гигантский прыжок: как насекомые достигли альтруизма и колониальной жизни» . Биоссака . 58 (1): 17–25. doi : 10.1641/b580106 .
- ^ Jump up to: а беременный Хьюз, Уильям О; Бенджамин П. Олдройд; Мадлен Бикман; Фрэнсис Л.В. Ратниккс (2008-05-30). «Принципальная моногамия показывает, что выбор родственников является ключом к эволюции эусоциальности». Наука . 320 (5880): 1213–1216. Bibcode : 2008Sci ... 320.1213H . doi : 10.1126/science.1156108 . PMID 18511689 . S2CID 20388889 .
- ^ Кахан, Ш; Гарднер-Морс, Э. (2013). «Появление репродуктивного разделения труда в принудительных группах королевы муравьев Pogonomyrmex Barbatus » . Журнал зоологии . 291 (1): 12–22. doi : 10.1111/jzo.12071 .
- ^ Molteno, AJ; Беннетт, Северная Каролина (2002). «Освещение, рассеяние и репродуктивное ингибирование у эусоциальных моле Дамараланда ( Cryptomys Damarensis )». Журнал зоологии . 256 (4): 445–448. doi : 10.1017/s0952836902000481 .
- ^ Тот, Ал; Робинсон, GE (2009-01-01). «Evo-Devo и эволюция социального поведения: анализ экспрессии генов мозга у социальных насекомых» . Симпозии Cold Spring Harbor по количественной биологии . 74 : 419–426. doi : 10.1101/sqb.2009.74.026 . PMID 19850850 .
- ^ Jump up to: а беременный Янега Д. (1993). «Экологическое влияние на мужскую производство и социальную структуру в Halictus Rubicund (Hymenoptera: Halictidae)». Насекомые сочи . 40 (2): 169-180. Doi : 10.1007 / bf01240705 . S2CID 44934383 .
- ^ Shell, Wyatt A.; Рехан, Сандра М. (2017-07-24). «Поведенческие и генетические механизмы социальной эволюции: понимание от зарождающихся и факультативно социальных пчел» . Апидология . 49 : 13–30. doi : 10.1007/s13592-017-0527-1 . ISSN 0044-8435 .
- ^ Новак, Массачусетс; Тарнита, CE; Wilson, EO (2010). «Эволюция эусоциальности» . Природа . 466 (7310): 1057–1062. BIBCODE : 2010NATR.466.1057N . doi : 10.1038/nature09205 . PMC 3279739 . PMID 20740005 .
- ^ Аббат, Патрик; и др. (2011). «Инклюзивная теория фитнеса и эусоциальность» . Природа . 471 (7339): E1 - E4. Bibcode : 2011natur.471e ... 1a . doi : 10.1038/nature09831 . PMC 3836173 . PMID 21430721 .
- ^ Зара, Фернандо; Balestieri, Хосе (2000). «Поведенческий каталог POLISTES Versicolor Olivier (Vespidae: Polistinae) пост-экипировки колонов». Натуралия . 25 : 301–319.
- ^ Харрисон, Марк С.; Jongepier, Evelien; Робертсон, Хью М.; Арни, Николас; Bitard-Feildel, Тристан; и др. (2018). «Гемиметаболистые геномы выявляют молекулярную основу термитной эусоциальности» . Природа экология и эволюция . 2 (3): 557–566. Bibcode : 2018natee ... 2..557H . doi : 10.1038/s41559-017-0459-1 . PMC 6482461 . PMID 29403074 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж Варго Э. (1999). «Репродуктивное развитие и онтогенство или производство королевы феромонов в огненном муравьевом Solenopsis Invicta ». Физиологическая энтомология . 24 (4): 370–376. doi : 10.1046/j.1365-3032.1999.00153.x . S2CID 84103230 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не Флетчер, Д.; Росс, К. (1985). «Регуляция размножения в эусоциальных гименоптерах». Ежегодный обзор энтомологии . 30 : 319–343. doi : 10.1146/annurev.ento.30.1.319 .
- ^ Carpenter, JM (1987). «Филогенетические отношения и классификация Vespinae (Hymenoptera: Vespidae)». Систематическая энтомология . 12 (4): 413–431. Bibcode : 1987sysen..12..413c . doi : 10.1111/j.1365-3113.1987.tb00213.x . S2CID 9388017 .
- ^ Feyereisen, R.; Tobe, S. (1981). «Быстрый анализ разделения для обычного анализа юношеского гормона, выпущенного насекомыми Corpora Allata». Аналитическая биохимия . 111 (2): 372–375. doi : 10.1016/0003-2697 (81) 90575-3 . PMID 7247032 .
- ^ Jump up to: а беременный в Охота, Дж.; Wolschin, F.; Хеншоу, М.; Newman, T.; Тот, а.; Амдам, Г. (17 мая 2010 г.). «Дифференциальная экспрессия генов и численность белка, как онтогенетическое смещение в сторону каст в примитивной эусоциальной осе» . Plos один . 5 (5): E10674. Bibcode : 2010ploso ... 510674H . doi : 10.1371/journal.pone.0010674 . PMC 2871793 . PMID 20498859 .
- ^ Камакура, Масаки (май 2011 г.). «RoyalActin вызывает дифференциацию королевы у пчел». Природа . 473 (7348): 478–483. Bibcode : 2011natur.473..478k . doi : 10.1038/nature10093 . HDL : 2123/10940 . PMID 21516106 . S2CID 2060453 .
- ^ Мерфи, Грэм (июль 2008 г.). « Учитывая ее пути»: в (территории) матриархальных утопиях » . Научная фантастика . 35 (2): 266–280.
- ^ Fromm, Harold (2013). «Обзор нащупывания для групп Эдварда О. Уилсона, Джонатана Хайдта, Стивена Митена, Стивена Пинкера и Ричарда Докинса». Обзор Гудзона . 65 (4): 652–658. JSTOR 43489291 .

