Мозжечок
| Мозжечок | |
|---|---|
 Положение человеческого мозжечка (сагиттальный вид) | |
 Анимация мозжечка | |
| Подробности | |
| Произношение | / ˌ s ɛ r ə ɛ ɛləml |
| Часть | Задних изделий |
| Артерия | SCA , AICA , PICA |
| Вена | Верхний , ниже |
| Идентификаторы | |
| латинский | мозжечок |
| Сетка | D002531 |
| Нейроналы | 643 |
| Neurolex Id | Birnlex_1489 |
| TA98 | A14.1.07.001 |
| TA2 | 5788 |
| FMA | 67944 |
| Анатомические термины нейроанатомии | |
Мозжечок латынь ( пл.: Мозга или мозжечка ; для «маленького мозга») является основной особенностью заднего мозга всех позвоночных . он обычно меньше, Хотя обычно у некоторых животных, таких как мормиридные рыбы, он может быть такой же большой, как и даже больше. [ 1 ] У людей мозжечок играет важную роль в моторном контроле и когнитивных функциях, таких как внимание и язык , а также эмоциональный контроль, такие как регулирование реакций страха и удовольствия, [ 2 ] [ 3 ] [ 4 ] Но его функции, связанные с движением, являются наиболее твердыми. Человеческий мозжечок не инициирует движение, но способствует координации , точности и точному времени: он получает вход от сенсорных систем спинного мозга и от других частей мозга и интегрирует эти входные данные с точной настройкой двигательной активности. [ 5 ] Повреждение мозжечка приводит к нарушениям в тонком движении , равновесии , осанке и моторном обучении у людей. [ 5 ]
Анатомически, человеческий мозжечок имеет вид отдельной структуры, прикрепленной к нижней части мозга, спрятанной под полушариями головного мозга . Его корковая поверхность покрыта тонко расположенными параллельными канавками, в результате чего -то контраста с широкими нерегулярными сознаниями коры головного мозга . Эти параллельные канавки скрывают тот факт, что кора мозжечка на самом деле является непрерывным тонким слоем ткани, плотно сложенным в стиле аккордеона . Внутри этого тонкого слоя находятся несколько типов нейронов с очень регулярным расположением, наиболее важными являются клетки Purkinje и гранулярные клетки . Эта сложная нейронная организация дает огромную способность обработки сигналов, но почти весь выход из коры мозжечка проходит через набор небольших глубоких ядер, лежащих во внутренней части белого вещества мозжечка. [ 6 ]
В дополнение к своей прямой роли в моторном управлении, мозжечок необходим для нескольких типов моторного обучения , в частности, обучение для адаптации к изменениям в сенсомоторных отношениях . Было разработано несколько теоретических моделей для объяснения сенсомоторной калибровки с точки зрения синаптической пластичности в мозжечке. Эти модели вытекают из моделей, сформулированных Дэвидом Марром и Джеймсом Альбусом , на основе наблюдения, что каждая ячейка Purkinje Certing получает два резко различных типа ввода: один содержит тысячи слабых входов из параллельных волокнов гранулярных ячеек; Другой - чрезвычайно сильный вклад из одного восходящего волокна . [ 7 ] Основная концепция теории Марра-Альбуса заключается в том, что восходящее волокно служит «сигналом обучения», что вызывает длительное изменение в силе параллельных входов волокон. Наблюдения за долгосрочной депрессией в параллельных входах волокна обеспечили некоторую поддержку теориям этого типа, но их достоверность остается спорной. [ 8 ]
Структура
[ редактировать ]На уровне грубой анатомии мозжечок состоит из плотно сложенного слоя коры , с белым веществом внизу и заполненным жидкостью желудочка у основания. Четыре глубоких ядра мозжечка встроены в белое вещество. [ 9 ] Каждая часть коры состоит из одного и того же небольшого набора нейрональных элементов, изложенных в высоко стереотипной геометрии. На промежуточном уровне мозжечок и его вспомогательные структуры могут быть разделены на несколько сотен или тысячи независимо функционирующих модулей, называемых «микрозонами» или «микрокомплекментами».
Грубая анатомия
[ редактировать ]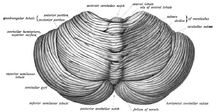
Мозжечок расположен в ямке задних черепной кости . Четвертый желудочек , поны и мозговый мозг находятся перед мозжечкой. [ 10 ] Он отделен от надлежащего мозга слоем кожистой Dura Mater , мозжечка Tentorium ; Все его связи с другими частями мозга проходят через Понс. Анатомисты классифицируют мозжечок как часть Metencephalon , который также включает в себя Pons; Метентфалон является верхней частью ромпенсфалона или «заднего мозга». Как и кора головного мозга, мозжечок делится на два полушария мозжечка ; Он также содержит узкую среднюю зону ( вермии ). Набор больших складок, согласно соглашению, используется для разделения общей структуры на 10 меньших «долей». Из -за большого количества крошечных гранулярных клеток , мозжечок содержит больше нейронов , чем общее количество от остальной части мозга, но занимает только 10% от общего объема мозга. [ 11 ] Количество нейронов в мозжечке связано с количеством нейронов в неокортексе . В мозжечке примерно в 3,6 раза больше нейронов, чем в неокортексе, соотношение, которое сохраняется у многих различных видов млекопитающих. [ 12 ]
Необычный вид поверхности мозжечка скрывает тот факт, что большая часть его объема состоит из очень плотно сложенного слоя серого вещества : коры мозжечка . Каждый хребет или извилина в этом слое называется фолимом . МРТ с высоким разрешением обнаруживает, что взрослой кора мозжечка человека имеет площадь 730 кв. [ 13 ] упакован в объем размеров 6 см × 5 см × 10 см. [ 11 ] Под серого вещество коры лежит белое вещество , состоящее в основном из миелинизированных нервных волокон, бегущих в и обратно. Встроенный в белое вещество, которое иногда называют видами беседки (дерево жизни) из-за его разветвленного, похожая на деревья внешнего вида в поперечном сечении, являются четырьмя глубокими ядрами мозжечка , состоящие из серого вещества. [ 14 ]
Соединение мозжечка с разными частями нервной системы - три парных цветоносителей мозжечка . Это превосходный цветоножка мозжечка , средняя цветоножка мозжечка и низший цветонотчик мозжечка , названные их позицией относительно вермис. Превосходная цветоножка мозжечка является в основном выходом в кору головного мозга, несущая эфферентные волокна через ядра таламуса в верхние моторные нейроны в коре головного мозга. Волокна возникают из глубоких ядер мозжечка. Средний мозжечок соединен с пососом и получает весь свой вход от понгов в основном от ядер понтинов . Вход в поны взят из коры головного мозга и передается из ядер понтинов через поперечные понтиновые волокна к мозжечке. Средняя плодоножка является самой большой из трех, а его афферентные волокна сгруппированы в три отдельных пучков, которые принимают свои входные данные в разные части мозжечка. Нижний цветоносец мозжечка получает вход от афферентных волокон от вестибулярных ядер, спинного мозга и сегментам. Выход из нижнего цветоноса проходит через эфферентные волокна в вестибулярные ядра и ретикулярное образование. Весь мозжечок получает модулирующий вход от нижнего ядра оливари через нижнюю плодоножку мозжечка. [ 6 ]
Подразделения
[ редактировать ]
Основываясь на внешнем виде, можно различить три доли в мозжечке: передняя доля (над первичной трещиной ), задняя доля (ниже первичной трещины) и флоккулонодулярная доля (ниже задней трещины). Эти доли делят мозжечок от рострального на каудальный (у людей, сверху вниз). Однако с точки зрения функции существует более важное различие вдоль среднего и латерального измерения. Оставив флоккулонодулярную долю, которая имеет различные связи и функции, мозжечок может быть функционально анализировать в медиальный сектор, называемый спиноцеребеллумом и большим боковым сектором, называемым цереброцеребеллум . [ 14 ] Узкая полоса выступающей ткани вдоль средней линии называется Vermis мозжечка . ( Вермис является латыни для «червя».) [ 14 ]
Наименьшая область, флоккулонодулярная доля, часто называют вестибулоцеребеллум . Это самая старая часть в эволюционных терминах (archicerebellum) и участвует в основном в балансе и пространственной ориентации; Его первичные соединения связаны с вестибулярными ядрами , хотя он также получает визуальный и другой сенсорный ввод. Ущерб в этом регионе вызывает нарушения баланса и походки . [ 14 ]
Медиальная зона передних и задних лепесток составляет спиноцеребеллум, также известный как палеоцереоток. Этот сектор мозжечка функционирует в основном для тонкой настройки тела и движения конечностей. Он получает проприоцептивный вход от дорсальных столбцов спинного мозга (включая спиноцеребелентный тракт ) и из черепного тройничного нерва , а также из визуальных и слуховых систем. [ 15 ] Он посылает волокна в глубокие ядра мозжечка, которые, в свою очередь, проецируются как в кору головного мозга, так и стержни мозга, что обеспечивает модуляцию нисходящих двигательных систем. [ 14 ]
Боковая зона, которая у людей, безусловно, является самой большой частью, составляет цереберебеллум, также известный как Neocerebellum. Он получает вход исключительно из коры головного мозга (особенно париетальной доли ) через ядра понтинов (образуя кортикопонто-звелковые пути) и посылает выход в основном в вентролатеральный таламус (в свою очередь подключенные к моторным областям премьер-коры и первичного двигателя. Площадь коры головного мозга) и к красному ядру . [ 14 ] Существует разногласия по поводу лучшего способа описать функции бокового мозжечка: считается, что он участвует в движении планирования, которое собирается произойти, [ 16 ] При оценке сенсорной информации для действий, [ 14 ] и в ряде чисто когнитивных функций, таких как определение глагола, который наилучшим образом соответствует определенному существительному (как в «SIT» для «стула»). [ 17 ] [ 18 ] [ 19 ] [ 20 ]
Микроанатомия
[ редактировать ]Два типа нейрона играют доминирующую роль в цепи мозжечка: клетки Пуркинье и гранулярные клетки . Три типа аксонов также играют доминирующие роли: мшистые волокна и восхождение волокон (которые попадают в мозжечок снаружи) и параллельные волокна (которые являются аксонами гранулярных ячеек). Есть два основных пути через мозжечку, происходящие из мшистых волокон и лазантных волокон, оба в конечном итоге заканчиваются в глубоких ядрах мозжечка. [ 11 ]
Мослистые волокна проецируются непосредственно к глубоким ядрам, но также дают следующий путь: мшистые волокна → гранулярные клетки → параллельные волокна → Purkinje -клетки → глубокие ядра. Волокон поднимается на клетки Пуркинье, а также посылает залога непосредственно в глубокие ядра. [ 11 ] Мессизистое волокно и восхождение волокно-волокна каждый несет волокно-специфичную информацию; Cerebellum также получает дофаминергические , серотонинергические , норадренергические и холинергические входы, которые предположительно выполняют глобальную модуляцию. [ 21 ]
Кора мозжечка разделена на три слоя. Внизу лежит толстый гранулированный слой, плотно упакованный гранулярными клетками, наряду с интернейронами , в основном клетки Гольджи , но также включая клетки Лугаро и однополярные клетки кисти . В середине лежит слой Пуркинье, узкую зону, которая содержит клеточные тела клеток Пуркинье и глиальные клетки Бергмана . В верхней части лежит молекулярный слой, который содержит сплюснутые дендритные деревья клеток Пуркинье, а также огромное количество параллельных волокон, проникающих в дендритные деревья клеток Пуркинье. Этот самый внешний слой коры мозжечка также содержит два типа ингибирующего интернейрона: звездчатые клетки и клетки корзины . Как звездчатые, так и корзинские клетки образуют ГАМКергические синапсы на дендриты клеток Пуркинье. [ 11 ]

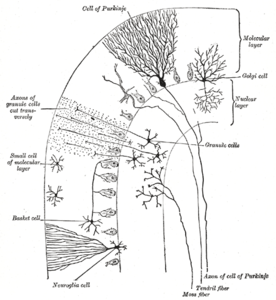
Слои коры мозжечка
[ редактировать ]Молекулярный слой
[ редактировать ]Верхний, самый внешний слой коры мозжечка - это молекулярный слой. Этот слой содержит сплющенные дендритные деревья клеток Пуркинье и огромный набор параллельных волокон, из гранулированного слоя, которые проникают в дендритные деревья клеток пуркинье под прямыми углами. Молекулярный слой также содержит два типа ингибирующего интернейрона: звездчатые клетки и корзину . Как звездчатые, так и корзинские клетки образуют ГАМКергические синапсы на дендриты клеток Пуркинье. [ 11 ]
Пуркинье слой
[ редактировать ]
Клетки Purkinje являются одними из самых отличительных нейронов в мозге, и один из самых ранних типов, которые должны быть признаны - они были впервые описаны чешским анатомистом Яном Евангелиста Пуркин в 1837 году. Они отличаются формой своего дендритного дерева: ветвь дендритов. Очень обильно, но сильно сплющиваются в плоскости, перпендикулярно складкам мозжечка. Таким образом, дендриты ячейки Пуркинье образуют плотную плоскую сеть, через которую параллельные волокна проходят под прямым углом. [ 11 ] Дендриты покрыты дендритными шипами , каждый из которых получает синаптический вход от параллельного волокна. Клетки Purkinje получают больше синаптических входов, чем любой другой тип клеток в мозге - ограничивает количество шипов на одном человеческом Purkinje -клетках до 200 000. [ 11 ] Большие сферические клеточные тела клеток Purkinje упаковываются в узкий слой (толщина одной клетки) коры мозжечка, называемый слоем Purkinje . После излучения коллатералей, которые влияют на близлежащие части коры, их аксоны перемещаются в глубокие ядра мозжечка , где они делают по порядку 1000 контактов, каждый с несколькими типами ядерных клеток, все в небольшом домене. Клетки Purkinje используют GABA в качестве своего нейротрансмиттера и, следовательно, оказывают ингибирующее влияние на их мишени. [ 11 ]
Клетки Purkinje образуют сердце мозжечкового цепи, и их большой размер и характерные модели активности сделали относительно простым изучать их паттерны ответа у животных, используя методы внеклеточной записи. Клетки Purkinje обычно испускают потенциалы действия с высокой скоростью даже в отсутствие синаптического ввода. В бодрствовании, поведении животных средние показатели, в среднем около 40 Гц являются типичными. Поезда Spike показывают смесь так называемых простых и сложных шипов. Простой всплеск - это единственный потенциал действия, за которым следуют рефрактерный период около 10 мс; Сложный всплеск представляет собой стереотипную последовательность потенциалов действия с очень короткими интервалами между шипами и снижением амплитуд. [ 23 ] Физиологические исследования показали, что сложные скачки (которые встречаются с исходными показателями около 1 Гц и никогда не ставками, намного выше, чем 10 Гц), достоверно связаны с активацией восхождения волокна, в то время как простые шипы производятся комбинацией базовой активности и параллельного ввода волокна. За сложными шипами часто сопровождается пауза в несколько сотен миллисекунд, в течение которых простая активность шипа подавляется. [ 24 ]
Специфическая, узнаваемая особенность нейронов Пуркинье является экспрессией кальбиндина . [ 25 ] Окрашивание кальбиндином мозга крысы после одностороннего хронического повреждения седалищного нерва позволяет предположить, что нейроны Пуркинье могут быть вновь генерируются во взрослой мозге, инициируя организацию новых дольков мозжечка. [ 26 ]

Гранулированный слой
[ редактировать ]
Клетки мозжечка , в отличие от клеток Пуркинье, являются одними из самых маленьких нейронов в мозге. Они также являются самыми многочисленными нейронами в мозге: у людей оценки их общего числа среднего числа около 50 миллиардов, что означает, что около 3/4 нейронов мозга являются гранулярными клетками мозжечка. [ 11 ] Их клеточные тела упакованы в толстый слой в нижней части коры мозжечка. Гранулярная ячейка излучает только от четырех до пяти дендритов, каждый из которых заканчивается увеличением, называемым дендритным коготь . [ 11 ] Эти увеличения являются сайтами возбуждающего ввода из мшистых волокон и ингибирующего ввода клеток Гольджи . [ 11 ]
Тонкие, неиелинизированные аксоны гранулярных ячеек поднимаются вертикально до верхнего (молекулярного) слоя коры, где они разделяются по двум, причем каждая ветвь движется горизонтально, образуя параллельное волокно ; Расщепление вертикальной ветви на две горизонтальные ветви приводит к отличительной форме «T». Параллельное волокно человека проходит в среднем 3 мм в каждом направлении от разделения, для общей длины около 6 мм (около 1/10 от общей ширины кортикального слоя). [ 11 ] Когда они бегают, параллельные волокна проходят через дендритные деревья клеток Пуркинье, контактируя с одним из каждых 3–5, которые они проходят, что составляет в общей сложности 80–100 синаптических связей с дендритными шипицами Purkinje. [ 11 ] Гранулярные клетки используют глутамат в качестве своего нейротрансмиттера и, следовательно, оказывают возбуждающее воздействие на их мишени. [ 11 ]

Гранульные ячейки получают все свои вклад от мошных волокон, но превосходят их на 200-1 (у людей). Таким образом, информация в состоянии активности популяции гранулярных клеток совпадает с информацией в мошных волокнах, но перекодирована гораздо более широким образом. Поскольку гранулярные клетки настолько малы и настолько плотно упакованы, трудно записать их активность всплеска в поведении животных, поэтому для теоретизирования мало данных мало для теоретизации. Самая популярная концепция их функции была предложена в 1969 году Дэвидом Марром , который предположил, что они могут кодировать комбинации входов из мшистых волокон. Идея состоит в том, что с каждой гранулярной ячейкой, получающей вход только из 4–5 мошных волокон, гранулярная ячейка не отвечала бы, если бы один из его входов был активным, но отвечал бы, если бы более одного было активным. Эта комбинаторная схема кодирования потенциально позволит мозжечке провести гораздо более мелкие различия между входными шаблонами, чем разрешат только мшистые волокна. [ 27 ]
Мшистые волокна
[ редактировать ]Мосси -волокна попадают в гранулированный слой из их точек происхождения, многие из которых возникают из ядер понтинов , другие из спинного мозга, вестибулярных ядер и т. Д. В мозжечке человека общее количество мшистых волокон было оценено в 200 миллионов. [ 11 ] Эти волокна образуют возбуждающие синапсы с гранулярными клетками и клетками глубоких ядер мозжечка. Внутри гранулированного слоя мшистое волокно генерирует серию увеличения, называемых розетками . Контакты между мшистыми волокнами и дендритами гранулярных клеток происходят в структурах, называемых гломерули . У каждого клубочка есть мошная волоконная розетка в центре, и до 20 гранулярных клеток дендритных когтей, связывающихся с ним. Терминалы из клеток Гольджи проникают в структуру и делают ингибирующие синапсы на дендритах гранулярных клеток. Вся сборка окружена оболочкой глиальных клеток. [ 11 ] Каждое мшистое волокно посылает обеспеченные ветви в несколько мозжечковых фолие, генерируя в общей сложности 20–30 розетков; Таким образом, одно мрачное волокно вступает в контакт с приблизительными 400–600 гранулярными ячейками. [ 11 ]
Волокон поднимается
[ редактировать ]Клетки Purkinje также получают вход от нижнего ядра оливари на контралатеральной стороне ствола мозга через восхождение волокон . Хотя нижняя оливка лежит в мозговом мозге и получает вход от спинного мозга, ствола мозга и коры головного мозга, его вывод полностью идет до мозжечка. Волокна для скалолазания испускает коллатерали в глубокие ядра мозжечка, прежде чем войти в кору мозжечка, где оно разбивает примерно 10 ветвей терминалов, каждая из которых дает вклад в одну ячейку Пуркинье. [ 11 ] В отличие от 100 000 с лишним входов из параллельных волокон, каждая ячейка Purkinje получает вклад из точно одного восходящего волокна; Но это единственное волокно «поднимается» дендриты ячейки Пуркинье, обрушивая их и делая в общей сложности до 300 синапсов по мере ее поступления. [ 11 ] Чистый вход настолько силен, что потенциал одного действия из скалолочного волокна способен создавать расширенный сложный всплеск в ячейке Пуркинье: разрыв нескольких шипов подряд, с уменьшением амплитуды, за которой следует пауза, во время которой подается активность Полем Синапсы скалолочного волокна покрывают тело клетки и проксимальные дендриты; Эта зона лишена параллельных входов волокон. [ 11 ]
Волокон поднимается с низкими показателями, но один потенциал действий для восхождения волокна вызывает взрыв нескольких потенциалов действия в целевой ячейке Пуркинье (сложный всплеск). Контраст между параллельным волокном и поднимающим волокном входов к клеткам Пуркинье (более 100 000 из одного типа по сравнению с именно одним из другого типа) является, пожалуй, самой провокационной особенностью анатомии мозжечка и мотивировал большую часть теоретизирования. Фактически, функция восхождения на волокна является наиболее спорной темой, касающейся мозжечка. Есть две школы мысли: одна после Марра и Альбуса, которые держат вклад в восхождение волокна, служит главным образом как обучающий сигнал, а другая - то, что его функция заключается в том, чтобы напрямую формировать выход мозжечка. Оба взгляда были в значительной степени защищены во многих публикациях. По словам одного обзора, «пытаясь синтезировать различные гипотезы о функции волокон скалолазания, можно взглянуть на рисунок Эшера. Каждая точка зрения, кажется, объясняет определенную коллекцию результатов, но Когда кто -то пытается собрать разные взгляды, последовательная картина того, что делают воспитание волокна, не появляется. Единственное объявление о «неожиданном событии». [ 24 ]
Глубокие ядра
[ редактировать ]
Глубокие ядра мозжечка - это кластеры серого вещества, лежащие в белом веществе в ядре мозжечка. Они, за незначительным исключением из соседних вестибулярных ядер, единственные источники выхода из мозжечка. Эти ядра получают обеспеченные прогнозы из мошных волокон и восхождения волокон, а также ингибирующий ввод от клеток Пуркинье коры мозжечка. Четыре ядра ( зубчатые , шаровидные , эмболиформные и быстрого ) общаются с разными частями мозга и коры мозжечка. (Глобусные и эмболиформные ядра также называются в сочетании в вставленном ядре ). Фастгиальные и вставленные ядра принадлежат к спиноцеребеллуму. Целеобразное ядро, которое у млекопитающих намного больше, чем у других, образуется как тонкий, запутанный слой серого вещества и общается исключительно с боковыми частями коры мозжечка. Флокулус флоккулонодулярной доли является единственной частью коры мозжечка, которая не проецируется на глубокие ядра, - его вывод идет к вестибулярным ядрам. [ 11 ]
Большинство нейронов в глубоких ядрах имеют большие клеточные тела и сферические дендритные деревья с радиусом около 400 мкм и используют глутамат в качестве их нейротрансмиттера. Эти клетки проецируются на различные мишени за пределами мозжечка. С ними смешаны меньшее количество мелких клеток, которые используют ГАМК в качестве нейротрансмиттера и проецируют исключительно на нижнее ядро оливари , источник восхождения волокон . Таким образом, ядро-провальная проекция обеспечивает ингибирующую обратную связь , чтобы соответствовать возбуждающей проекции восхождения волокон с ядрами. Существуют доказательства того, что каждый небольшой кластер ядерных ячеек проецирует в одном и том же кластере оливари, которые посылают в него восходящие волокна; В обоих направлениях есть сильная и соответствующая топография. [ 11 ]
Когда аксон Purkinje Cell входит в одно из глубоких ядер, он ветвет, чтобы установить контакт как с большими, так и с небольшими ядерными клетками, но общее количество обращенных клеток составляет всего около 35 (у кошек). И наоборот, одна глубокая ядерная ячейка получает вход примерно из 860 клеток Пуркинье (опять же у кошек). [ 11 ]
Отсеки
[ редактировать ]
С точки зрения грубой анатомии кора мозжечка, по -видимому, представляет собой гомогенную лист ткани, и с точки зрения микроанатомии все части этого листа, по -видимому, имеют одинаковую внутреннюю структуру. Однако существует ряд признаков, в которых структура мозжечка разделена. Существуют большие отсеки, которые обычно известны как зоны ; Их можно разделить на меньшие отсеки, известные как микрозоны . [ 28 ]
Первые признаки компартментальной структуры были получены из исследований рецептивных полей клеток в различных частях мозжечкового коры. [ 28 ] Каждая часть тела отображается в определенных точках в мозжечке, но существует множество повторений базовой карты, образуя расположение, которое называлось «сломанной соматотопией». [ 29 ] Более четкий показатель компартментализации получается путем иммуноокрашивания мозжечка для определенных типов белка. Самые известные из этих маркеров называются «зебранами», потому что окрашивание для них приводит к сложному рисунку, напоминающему полосы на зебре. Полосы, генерируемые зелеными и другими маркерами компартментализации, ориентированы перпендикулярно складкам мозжечка, то есть они узкие в медиолатеральном направлении, но гораздо более расширены в продольном направлении. Различные маркеры генерируют разные наборы полос, ширина и длина варьируются в зависимости от местоположения, но все они имеют одинаковую общую форму. [ 28 ]
Оскарссон в конце 1970 -х годов предложил, чтобы эти корковые зоны были разделены на более мелкие единицы, называемые микрозонами. [ 30 ] Микрозон определяется как группа клеток Пуркинье, имеющих одинаковое соматотопическое восприимчивое поле. Было обнаружено, что микрозоны содержат по порядку 1000 клеток Пуркинье, расположенные в длинной узкой полосе, ориентированной перпендикулярной кортикальным складкам. [ 28 ] Таким образом, как показывает соседняя диаграмма, дендриты ячейки Purkinje сглаживаются в том же направлении, что и микрозоны, в то время как параллельные волокна пересекают их под прямым углом. [ 11 ]
Это не только восприимчивые поля, которые определяют структуру микрозона: вход для лазания волокна из нижнего ядра оливари в равной степени важен. Ветви скалолочного волокна (обычно нумерация около 10) обычно активируют клетки Пуркинье, принадлежащие к одному и тому же микрозону. Более того, оливские нейроны, которые посылают восходящие волокна в одном и том же микрозоне, как правило, связаны с разрывыми соединениями , которые синхронизируют их активность, вызывая клетки Пуркинье в микрозоне, чтобы показать коррелированную сложную активность всплеска в миллисекундной шкале времени. [ 28 ] Кроме того, клетки Purkinje, принадлежащие к микрозону, все посылают свои аксоны в один и тот же небольшой кластер выходных ядер в глубоких ядрах мозжечка . [ 28 ] Наконец, аксоны клеток корзины гораздо дольше в продольном направлении, чем в медиолатеральном направлении, заставляя их быть в значительной степени ограничиваться одним микрозоном. [ 28 ] Следствием всей этой структуры является то, что клеточные взаимодействия в микрозоне намного сильнее, чем взаимодействия между различными микрозонами. [ 28 ]
В 2005 году Ричард Apps и Мартин Гарвич подвели доказательства того, что сами микрозоны являются частью более крупной сущности, которую они называют многопонанным микрокомплексом. Такой микрокомплекс включает в себя несколько пространственно отделенных кортикальных микрозонов, все из которых проецируются на одну и ту же группу глубоких нейронов мозжечка, а также группу связанных оливских нейронов, которые проецируются на все включенные микрозоны, а также к глубокому ядерному области. [ 28 ]
Кровоснабжение
[ редактировать ]Цереберочный мозжечок обеспечивается кровью из трех парных главных артерий: верхняя мозжечка (SCA), передняя нижняя мозжечковая артерия (AICA) и задняя нижняя мозжечковая артерия (PICA). SCA поставляет верхнюю область мозжечка. Он разделяет на верхней поверхности и ветвет в Pia Mater, где ветвя анастомоза с передней и задней нижней мозжечковой артерией. AICA поставляет переднюю часть нижней поверхности мозжечка. PICA прибывает в нижнюю поверхность, где он делится на медиальную ветвь и боковую ветвь. Медиальная ветвь продолжается назад к выемке мозжечка между двумя полушариями мозжечка; в то время как боковая ветвь снабжает под поверхностью мозжечка, до боковой границы, где она анастомозирует с AICA и SCA.
Функция
[ редактировать ]Самые сильные подсказки к функции мозжечка были получены из -за изучения последствий ущерба для него. Животные и люди с дисфункцией мозжечка показывают, прежде всего, проблемы с моторным контролем, на той же стороне тела, что и поврежденная часть мозжечка. Они по -прежнему могут генерировать двигательную активность, но теряют точность, создавая беспорядочные, несогласованные или неправильно рассмотренные движения. Стандартное испытание функции мозжечка состоит в том, чтобы достичь кончика пальца для цели в длине руки: здоровый человек перемещает кончик пальца в быстрой прямой траектории, тогда как человек с повреждением мозжечка будет медленно и беспорядочно, со многими серединами -Коценена исправления. Дефицит в немоторных функциях сложнее обнаружить. Таким образом, общий вывод, достигнутый десятилетиями назад, заключается в том, что основная функция мозжечка состоит в том, чтобы калибровать подробную форму движения, а не инициировать движения или решать, какие движения для выполнения. [ 14 ]
До 1990-х годов функция мозжечка почти повсеместно считалась чисто моторной, но новые выводы поставили под сомнение эту точку зрения. Исследования функциональной визуализации показали активацию мозжечка в отношении языка, внимания и умственных изображений; Корреляционные исследования показали взаимодействие между мозжечкой и немоторными областями коры головного мозга; И различные немоторные симптомы были признаны у людей с повреждением, которые, по-видимому, ограничены мозжечкой. [ 31 ] [ 32 ] В частности, когнитивный аффективный синдром мозжечка или синдром Шмахмана [ 33 ] был описан у взрослых [ 34 ] и дети. [ 35 ] Оценки, основанные на функциональном отображении мозжечка с использованием функциональной МРТ, позволяют предположить, что более половины мозжечкового коры взаимосвязана с зонами ассоциации коры головного мозга. [ 36 ]
Кенджи Доя утверждает, что функция мозжечка лучше всего понимается не с точки зрения поведения, на которую она влияет, а на нейронные вычисления, которые она выполняет; Cerebellum состоит из большого количества более или менее независимых модулей, все с одной и той же геометрически регулярной внутренней структурой, и, следовательно, все, как предполагается, выполняя одно и то же вычисление. Если входные и выходные соединения модуля связаны с областями двигателя (столь же многие), то модуль будет участвовать в моторном поведении; Но, если соединения связаны с областями, участвующими в немоторном познании, модуль покажут другие типы поведенческих коррелятов. Таким образом, мозжечок участвовал в регуляции многих различных функциональных признаков, таких как привязанность, эмоции, включая восприятие эмоционального языка тела [ 37 ] и поведение. [ 38 ] [ 39 ] Доя, предлагает мозжечок, лучше всего понимается как выбор прогнозирующих действий, основанный на «внутренних моделях» окружающей среды или устройстве для контролируемого обучения , в отличие от базальных ганглиев , которые выполняют обучение подкреплению и кору головного мозга , которая выполняет неконперводное обучение Полем [ 32 ] [ 40 ] Три десятилетия исследования мозга привели к предположению, что мозжечок генерирует оптимизированные ментальные модели и тесно взаимодействует с корой головного мозга, где обновленные внутренние модели воспринимаются как творческая интуиция («А -ха») в рабочей памяти. [ 41 ]
Принципы
[ редактировать ]Сравнительная простота и регулярность анатомии мозжечка привели к ранней надежде, что она может подразумевать аналогичную простоту вычислительной функции, как это выражено в одной из первых книг по электрофизиологии мозжечка, мозга как нейрональная машина от Джона С. Экклса , Масао. Ито и Янос Сцентаготай . [ 42 ] Хотя полное понимание функции мозжечка оставалось неуловимым, по меньшей мере четыре принципа были идентифицированы как важные: (1) обработка при питании, (2) дивергенция и сходимость, (3) модульность и (4) пластичность.
- Обработка прямой связи : мозжечок отличается от большинства других частей головного мозга (особенно коры головного мозга) тем, что обработка сигнала является почти полностью питательной , то есть сигналы перемещаются однонаправленно через систему от ввода к выходу, с очень небольшим повторяющимся внутренней передачей. Небольшое количество рецидивов, которое существует, состоит из взаимного ингибирования; Там нет взаимно возбуждающих цепей. Этот режим работы при питании означает, что мозжечок, в отличие от коры головного мозга, не может генерировать самоподдерживающиеся паттерны нейронной активности. Сигналы входят в схему, обрабатываются каждым этапом в последовательном порядке, а затем оставляют. Как писали Экклс, Ито и Сцентаготаи: «Это устранение при разработке всей возможности реверберационных цепей нейронального возбуждения, несомненно Cerebellum, вероятно, является не каким -то выходным выводом, выражающим операцию сложных реверберационных цепей в мозжечке, а скорее быстрый и четкий ответ на ввод какого -либо конкретного набора информации ». [ 43 ]
- Дивергенция и конвергенция : в мозжечке человека информация от 200 миллионов входов из мшистого волокна расширяется до 40 миллиардов гранулярных ячеек , чьи параллельные выходы волокна затем сходятся в 15 миллионов клеток Пуркинье . [ 11 ] Из -за того, как они выстроены в продольные выстроения, ячейки Purkinje, принадлежащие к микрозону, могут получить вход от 100 миллионов параллельных волокон и сосредоточить свой собственный выпуск на группе менее 50 глубоких ядерных клеток. [ 28 ] Таким образом, сеть мозжечка получает скромное количество входов, очень широко обрабатывает их через его строго структурированную внутреннюю сеть и отправляет результаты через очень ограниченное количество выходных ячеек.
- Модульность : Система мозжечка функционально разделена на более или менее независимые модули, которые, вероятно, насчитывают в сотнях до тысяч. Все модули имеют одинаковую внутреннюю структуру, но разные входы и выходы. Модуль (мультизональная микрокомпментация в терминологии Apps и Garwicz) состоит из небольшого скопления нейронов в нижнем ядре оливари, набора длинных узких полос пункинже в коре мозжечка (микрозоны) и маленького кластера нейрона в одном из глубоких ядер мозжечка. Различные модули разделяют входные данные из мошных волокон и параллельных волокон, но в других отношениях они, по -видимому, функционируют независимо - выход одного модуля, по -видимому, не влияет на активность других модулей. [ 28 ]
- Пластичность : синапсы между параллельными волокнами и клетками Пуркинье, а синапсы между мошенными волокнами и глубокими ядерными клетками подвержены модификации их силы. В одном модуле мозжечка вход из целых миллиардов параллельных волокон сходится к группе из менее 50 глубоких ядерных клеток, и влияние каждого параллельного волокна на эти ядерные клетки регулируется. Это расположение дает огромную гибкость для точной настройки взаимосвязи между входами и выходов мозжечка. [ 44 ]
Обучение
[ редактировать ]Существуют значительные доказательства того, что мозжечок играет важную роль в некоторых типах моторного обучения. Задачи, в которых наиболее четко вступает в игру мозжечок, - это то, что необходимо для того, чтобы выполнять действие. Однако существует много спора о том, происходит ли обучение в самом мозжечке, или он просто служит для предоставления сигналов, которые способствуют обучению в других структурах мозга. [ 44 ] Большинство теорий, которые назначают обучение в схему мозжечка, получены из идей Дэвида Марра [ 27 ] и Джеймс Альбус , [ 7 ] которые постулировали, что восхождение волокон дает учебный сигнал, который вызывает синаптические модификации в параллельных Purkinje . клетчатых синапсах [ 45 ] Марр предположил, что подъем входного волокна приведет к усилению синхронно активированных параллельных волоконных входов. Однако большинство последующих моделей обучения мозжечке следовало за Альбусом, предполагая, что поднимая активность волокна будет сигналом ошибки и приведет к ослаблению синхронно активированных параллельных волоконных входов. Некоторые из этих более поздних моделей, такие как модель адаптивного фильтра Fujita [ 46 ] Сделанные попытки понять функцию мозжечка с точки зрения оптимальной теории контроля .
Идея о том, что восхождение на активность волокна функционирует как сигнал ошибки, была исследована во многих экспериментальных исследованиях, причем некоторые поддерживают ее, но другие ставят под сомнение. [ 24 ] В новаторском исследовании Гилберта и Таха из 1977 года клетки Пуркинье из обезьян, изучающих задачу достижения, показали повышенную сложную активность всплеска, которая, как известно, достоверно указывает на активность ввода лазанного волокна в клетке, когда производительность была плохой. [ 47 ] Несколько исследований моторного обучения у кошек наблюдали сложную активность всплеска, когда произошло несоответствие между предполагаемым движением и движением, которое было фактически выполнено. Исследования вестибуло -кокулярного рефлекса (который стабилизирует визуальное изображение на сетчатке, когда голова поворачивается) обнаружили, что альтинизационная активность волокна указывает на «скольжение сетчатки», хотя и не очень простым способом. [ 24 ]
Одной из наиболее тщательно изученных задач по обучению мозжечка является парадигма кондиционирования глазных изделий , в которой нейтральный кондиционированный стимул (CS), такой как тон или свет, неоднократно в сочетании с безусловным стимулом (США), таким как воздуховодо мигающий ответ. После таких повторяющихся презентаций CS и US CS в конечном итоге вызовет мигание перед США, условно -реакцию или CR. Эксперименты показали, что поражения локализованы либо в определенной части вставленного ядра (одно из глубоких ядер мозжечка), либо в нескольких специфических точках в коре мозжечка, отменит изучение условного отклика. Если результаты мозжечка фармакологически инактивированы, оставляя входные данные и внутриклеточные цепи нетронутыми, обучение происходит, даже когда животное не может показать никакого ответа, тогда как, если внутрицелевые схемы нарушены, нет обучения - эти факты, собранные вместе, убедительно обоснованы, что это убедительно обоснован Обучение, действительно, происходит внутри мозжечка. [ 48 ]
Теории и вычислительные модели
[ редактировать ]
Большая база знаний об анатомической структуре и поведенческих функциях мозжечка сделала его плодородной основой для теоретизирования - возможно, есть больше теорий функции мозжечка, чем любой другой части мозга. Самое основное различие между ними - между «теориями обучения» и «теориями производительности», то есть теориями, которые используют синаптическую пластичность в мозжечке, чтобы объяснить его роль в обучении, по сравнению с теориями, которые учитывают аспекты постоянного поведения Основа обработки сигнала мозжечка. Несколько теорий обоих типов были сформулированы в виде математических моделей и смоделированы с использованием компьютеров. [ 45 ]
Возможно, самой ранней теорией «исполнения» была гипотеза «линии задержки» Валентино Брайтенберга . Первоначальная теория, изложенная Брайтенбергом и Роджером Этвудом в 1958 году, предположила, что медленное распространение сигналов вдоль параллельных волокон накладывает предсказуемые задержки, которые позволяют мозжечке обнаруживать отношения времени в определенном окне. [ 49 ] Экспериментальные данные не подтвердили исходную форму теории, но Брайтенберг продолжал спорить о модифицированных версиях. [ 50 ] Гипотеза о том, что мозжечок функционирует, по существу, как система времени, также защищалась Ричардом Аври . [ 51 ] Другая влиятельная теория «производительности» - теория тензоров Pellionisz и Llinás , которая обеспечила расширенную математическую формулировку идеи о том, что фундаментальные вычисления, выполняемые Cerebellum, предназначена для преобразования сенсорных в моторные координаты. [ 52 ]
Теории в категории «обучение» почти все происходят из публикаций Марра и Альбуса. В статье Марра 1969 года предложила, чтобы мозжечок был устройством для обучения для ассоциации элементарных движений, кодируемых при восхождении волокон с помощью входов из мшистого волокна, которые кодируют сенсорный контекст. [ 27 ] Альбус предложил в 1971 году, что клетка Purkinje мозжечка функционирует как персептрон , неврологическое абстрактное обучение. [ 7 ] Самое основное различие между теориями Марра и Альбуса заключается в том, что Марр предполагал, что поднимая активность волокна приведет к укреплению параллельных синапсов волокна, тогда как Альбус предположил, что они будут ослаблены. Альбус также сформулировал свою версию в качестве программного алгоритма, который он назвал CMAC (контроллер артикуляции модели мозжечка), который был протестирован в ряде приложений. [ 53 ]
Клиническое значение
[ редактировать ]

Повреждение мозжечка часто вызывает симптомы, связанные с двигателем, детали которых зависят от части участия мозжечка и от того, как он поврежден. Повреждение флоккулонодулярной доли может показаться потерей равновесия и, в частности, измененной, нерегулярной ходьбы, с широкой позицией, вызванной трудностями балансировки. [ 14 ] Ущерб боковой зоне обычно вызывает проблемы в квалифицированных добровольных и запланированных движениях, которые могут вызвать ошибки в силе, направлении, скорости и амплитуде движений. Другие проявления включают гипотонию (снижение мышечного тонуса), дискартрию (проблемы с артикуляцией речи), дисметрию (проблемы, оценивающие расстояния или диапазоны движения), дисдиадохокинезия (неспособность выполнять быстрые переменные движения, такие как ходьба), нарушение проверки или феномен -отскок, и и феномен, и и феномен, и и феномен, и и феномен, и и феномен, и и феномен, и и феномен, и феномено. намерение тремор (непроизвольное движение, вызванное чередующимися сокращениями противоположных групп мышц). [ 54 ] [ 55 ] Ущерб средней линии может нарушить движения всего тела, тогда как повреждение, локализованное более в боковом направлении, с большей вероятностью нарушает мелкие движения рук или конечностей. Ущерб верхней части мозжечка имеет тенденцию вызывать нарушения походки и другие проблемы с координацией ног; Ущерб нижней части с большей вероятностью вызывает несогласованные или плохо нацеленные движения рук и рук, а также трудности в скорости. [ 14 ] Этот комплекс моторных симптомов называется атаксией .
Чтобы выявить проблемы мозжечка, неврологическое обследование включает в себя оценку походки (широкая походка, указывающая на атаксию), тесты с указанием пальцев и оценку осанки. [ 5 ] Если указана дисфункция мозжечка, сканирование магнитно -резонансной томографии может использоваться для получения подробной картины любых структурных изменений, которые могут существовать. [ 56 ]
Список медицинских проблем, которые могут привести к повреждению мозжечка, является длинным, включая инсульт , кровоизлияние , отек головного мозга ( отек головного мозга ), опухоли , алкоголизм , физические травмы, такие как огнестрельные ранения или взрывчатые вещества, и хронические дегенеративные состояния, такие как атрофия оливопонтоцерея . [ 57 ] [ 58 ] Некоторые формы мигрени могут также вызывать временную дисфункцию мозжечка, с высокой степенью серьезности. [ 59 ] Инфекция может привести к повреждению мозжечка в таких условиях, как прионные заболевания [ 60 ] и синдром Миллера Фишера, вариант синдрома Гийяна -Барре .
Старение
[ редактировать ]Человеческий мозжечок меняется с возрастом. Эти изменения могут отличаться от других частей мозга. Церебелок является самой молодой областью мозга (и частью тела) у столетников в соответствии с эпигенетическим биомаркером эпохи ткани, известном как эпигенетические часы : он примерно на 15 лет моложе, чем ожидалось у столетия. [ 61 ] Кроме того, паттерны экспрессии генов в мозжечке человека демонстрируют меньшее возрастное изменение, чем в коре головного мозга . [ 62 ] В некоторых исследованиях сообщалось о снижении количества клеток или объема ткани, но объем данных, относящихся к этому вопросу, не очень велик. [ 63 ] [ 64 ]
Развитие и дегенеративные расстройства
[ редактировать ]
Врожденное развитие, наследственные расстройства и приобретенные условия могут повлиять на структуру мозжечка и, следовательно, на функцию мозжечка. Если причинное состояние не является обратимым, единственное возможное лечение - помочь людям жить со своими проблемами. [ 65 ] Визуализация мозжечка плода с помощью ультразвукового сканирования через 18-20 недель беременности может использоваться для скрининга плода на дефекты нервной трубки с уровнем чувствительности до 99%. [ 66 ]
В нормальном развитии передача сигналов эндогенного звукового хеджира стимулирует быструю пролиферацию предшественников нейрона мозжечка (CGNP) во внешнем гранулярном слое (EGL). Развитие мозжечка происходит во время позднего эмбриогенеза и раннего постнатального периода, с пролиферацией CGNP в пика EGL во время раннего развития (послеродовой день 7 у мыши). [ 67 ] По мере того, как CGNP терминально дифференцируются в гранулярные клетки мозжечка (также называемые нейронами мозжечка, CGNS), они мигрируют на внутренний гранул слой (IGL), образуя зрелый мозжечок (к пост-натальному дню 20 у мышей). [ 67 ] Мутации, которые аномально активируют передачу сигналов Sonic Hedgehog, предрасполагают к раку мозжечка ( медуллобластома ) у людей с синдромом Горлина и в генетически инженерных мышиных моделях . [ 68 ] [ 69 ]
Врожденное развитие или недостаточно развитие ( гипоплазия ) мозжечковых словестов является характеристикой как синдрома Денди -Уокера, так и синдрома Жуберта . [ 70 ] [ 71 ] В очень редких случаях весь мозжечок может отсутствовать . [ 72 ] Унаследованные неврологические расстройства Мачадо -Джозеф Болезнь , атаксия Телангиэктазия и атаксия Фридрейха вызывают прогрессирующую нейродегенерацию, связанную с потерей мозжечка. [ 57 ] [ 65 ] Врожденные пороки развития головного мозга вне мозжечка могут, в свою очередь, вызывать грыжу ткани мозжечка , как видно в некоторых формах порока развития Арнольд -Чиари . [ 73 ]
Другие условия, тесно связанные с дегенерацией мозжечка, включают идиопатические прогрессивные неврологические расстройства множественные системы атрофии системы и синдрома Рамсей Хант типа I , [ 74 ] [ 75 ] и аутоиммунного расстройства паранеопластическая дегенерация мозжечка , при которой опухоли в других местах тела вызывают аутоиммунный ответ, который вызывает потерю нейронов в мозжечке. [ 76 ] Атрофия мозжечка может быть результатом острого дефицита витамина В1 ( тиамин ), как это наблюдается при берибери и при синдроме Вернике -Коракофф , [ 77 ] или витамина Е. дефицит [ 65 ]
Атрофия мозжечка наблюдалась во многих других неврологических расстройствах, включая болезнь Хантингтона , рассеянный склероз , [ 60 ] Основной тремор , прогрессирующая эпилепсия миоклонуса и болезнь Ниманна - Фик . Атрофия мозжечка также может возникнуть в результате воздействия токсинов, включая тяжелые металлы или фармацевтические или рекреационные препараты . [ 65 ]
Боль
[ редактировать ]Существует общий консенсус в отношении того, что мозжечок участвует в обработке боли. [ 78 ] [ 79 ] Цереберел получает ввод боли как по нисходящим путям кортико-звелкера, так и по восходящим спино-зеленовому пути через ядра понтинов и нижние оливки. Некоторая часть этой информации передается в моторную систему, вызывая сознательное избегание моторного уклонения от боли, оцениваемой в соответствии с интенсивностью боли.
Считается, что эти прямые входы боли, а также косвенные входы вызывают долгосрочное поведение избегания боли, что приводит к хроническим изменениям осанки и, следовательно, в функциональном и анатомическом ремоделировании вестибулярных и проприоцептивных ядер. В результате хроническая невропатическая боль может вызвать макроскопическое анатомическое ремоделирование заднего мозга, включая мозжечок. [ 26 ] Величина этого ремоделирования и индукция маркеров -предшественников нейронов предполагают вклад взрослого нейрогенеза в эти изменения.
Сравнительная анатомия и эволюция
[ редактировать ]
Схемы в мозжечке похожи на всех классах позвоночных . , включая рыбу, рептилии, птицы и млекопитающие [ 80 ] Существует также аналогичная структура мозга у головоногих с хорошо развитым мозгом, такими как осьминоги . [ 81 ] Это было принято в качестве доказательства того, что мозжечок выполняет функции, важные для всех видов животных с мозгом.
Существует значительное различие в размере и форме мозжечка у разных видов позвоночных. У амфибий это мало развито, и у миноги , а Hagfish мозжечок едва отличается от ствола мозга. Хотя в этих группах присутствует спиноцеребеллум, первичные структуры небольшие, парные нуклеи, соответствующие Vestibulocerebellum. [ 82 ] Церебелок немного больше в рептилиях, значительно больше у птиц и больше у млекопитающих. Большие парные и запутанные доли, обнаруженные у людей, типичны для млекопитающих, но мозжечок, как правило, является единственной медианной долей в других группах и является либо гладким, либо лишь слегка рифленным. У млекопитающих неоцелбеллум является основной частью мозжечка по массе, но у других позвоночных это, как правило, спиноцеребеллум. [ 82 ]
Церебелок хрящевых и костных рыб чрезвычайно большой и сложный. По крайней мере, в одном важном отношении он отличается во внутренней структуре от мозжечка млекопитающих: мозжечок рыбы не содержит дискретных глубоких ядер мозжечка . Вместо этого основными мишенями клеток Пуркинье являются отчетливым типом клеток, распределенных по коре мозжечка, тип, не наблюдаемый у млекопитающих. У мормиридных рыб (семейство слабо электросимментивой пресной рыбы) мозжечок значительно больше, чем у остальной части мозга. Самая большая его часть - это специальная структура, называемая Valvula , которая имеет необычайно регулярную архитектуру и получает большую часть своей входной системы от электросенсорной системы. [ 83 ]
Отличительной чертой мозжечка млекопитающих является расширение боковых дол, чьи основные взаимодействия с неокортексом. По мере того, как обезьяны превратились в большие обезьяны, расширение боковых долей продолжалось, в тандеме с расширением лобных дол. В наследственных гоминидах и в Homo Sapiens до среднего плейстоценового периода мозжечок продолжал расширяться, но лобные доли расширялись быстрее. Однако самый последний период эволюции человека может быть фактически связан с увеличением относительного размера мозжечка, поскольку неокортекс несколько уменьшил его размер, пока мозжечок расширился. [ 84 ] Размер мозжечка человека, по сравнению с остальной частью мозга, увеличивается в размере, в то время как мозговой [ 85 ] С учетом развития и реализации моторных задач, визуальных пространственных навыков и обучения, происходящих в мозжечке, считается, что рост мозжечка имеет некоторую форму корреляции с большими когнитивными способностями человека. [ 86 ] Боковые полушария мозжечка теперь в 2,7 раза больше как у людей, так и у людей, чем у обезьян. [ 85 ] Эти изменения в размере мозжечка не могут быть объяснены большей мышечной массой. Они показывают, что либо развитие мозжечка тесно связано с развитием остальной части мозга, либо что нейронные действия, происходящие в мозжечке, были важны во время эволюции Hominidae . Из -за роли мозжечка в когнитивных функциях увеличение его размера могло сыграть роль в когнитивной экспансии. [ 85 ]
Cerebellum, похожие на структуры
[ редактировать ]Большинство видов позвоночных имеют мозжечок и одну или несколько, подобных мозжечке, областям мозга, которые напоминают мозжечок с точки зрения цитоархитектуры и нейрохимии . [ 80 ] Единственной мозжечкой, подобной структуре, обнаруженной у млекопитающих, является дорсальное кохлеарное ядро (DCN), одно из двух первичных сенсорных ядер, которые получают вход непосредственно из слухового нерва . DCN представляет собой многослойную структуру, с нижним слоем, содержащим гранулярные ячейки, аналогичные клетчатке мозжечка, что приводит к параллельным волокнам , которые поднимаются к поверхностному слою и проходят по нему горизонтально. Поверхностный слой содержит набор ГАМКергических нейронов, называемых клетками , которые напоминают клетки Пуркинье анатомически и химически - они получают параллельный ввод волокна, но не имеют никаких входов, которые напоминают восхождение волокон . Выходными нейронами DCN являются пирамидальными клетками . Они глутаматергические, но также напоминают клетки Пуркинье в некоторых отношениях - у них есть колючие, сплющенные поверхностные дендритные деревья, которые получают параллельный ввод волокна, но они также имеют базальные дендриты, которые получают вход от слуховых нервных волокон, которые проходят через DCN в направлении в Прямые углы на параллельные волокна. DCN наиболее высоко развит у грызунов и других мелких животных и значительно снижен у приматов. Его функция не совсем понятна; Самые популярные предположения связаны с пространственным слухом так или иначе. [ 87 ]
Большинство видов рыб и амфибий обладают системой боковой линии , которая ощущает волны давления в воде. Одна из областей мозга, которая получает первичный вход от органа латеральной линии, медиального октаволатерального ядра, имеет структуру, подобную мозжечке, с гранулярными клетками и параллельными волокнами. В электросчувствительной рыбе вход из электросенсорной системы идет в дорсальный октаволатеральный ядро, которое также имеет структуру, подобную мозжечкам. У рыб с лучами (безусловно, самая большая группа) у зрительного тектума есть слой-предельный слой-который похож на мозжечок. [ 80 ]
Все эти мозжечковые структуры, по-видимому, в основном связаны с сенсорным, а не связанным с двигателем. У всех них есть гранулярные клетки, которые приводят к параллельным волокнам, которые соединяются с пуркинже, подобными нейронам с модифицируемыми синапсами , но ни у кого не имеют лазания волокон, сопоставимых с клетками мозжечка-они получают прямой вклад от периферических сенсорных органов. Ни одна из них не имеет продемонстрированной функции, но наиболее влиятельным предположением является то, что они служат для преобразования сенсорных входов каким -либо сложным способом, возможно, для компенсации изменений в осанке тела. [ 80 ] Фактически, Джеймс М. Бауэр и другие утверждали, отчасти на основе этих структур и частично на основе исследований мозжечка, что сам мозжечок в основном является сенсорной структурой, и что он способствует моторному контролю, перемещая тело в способ, который контролирует полученные сенсорные сигналы. [ 88 ] Несмотря на точку зрения Бауэра, существует также убедительные доказательства того, что мозжечок напрямую влияет на моторную мощность у млекопитающих. [ 89 ] [ 90 ]
История
[ редактировать ]
Описания
[ редактировать ]Даже самые ранние анатомисты смогли распознать мозжечок по его отличительному внешнему виду. Аристотель и Герофил (цитируемый в Галене ) назвали его παρεγκεφαλίς ( parenkephalis ), в отличие от ἐγκέφαλος ( энкефалос ) или собственного мозга. Обширное описание Галена - самое раннее, которое выживает. Он предположил, что мозжечок был источником моторных нервов. [ 91 ]
Дальнейшие значительные события не произошли до эпохи Возрождения . Весалий кратко обсуждал мозжечок, и Анатомия была более тщательно описана Томасом Уиллисом в 1664 году. В течение 18 -го века была проделана больше анатомических работ, но только в начале 19 -го века первым пониманием функции мозжечка были полученный. Луиджи Роландо в 1809 году установил ключевой вывод, что повреждение мозжечка приводит к моторным нарушениям. Жан Пьер муку в первой половине 19 -го века проделала подробную экспериментальную работу, которая показала, что животные с повреждением мозжечка все еще могут двигаться, но с потерей координации (странные движения, неловкая походка и мышечная слабость), и это выздоровление после выздоровления Повреждение может быть почти завершено, если поражение не очень обширно. [ 92 ] К началу 20 -го века было широко признано, что основная функция мозжечка относится к управлению двигателем; Первая половина 20 -го века дала несколько подробных описаний клинических симптомов, связанных с болезнью мозжечка у людей. [ 5 ]
Этимология
[ редактировать ]Название мозжечка является уменьшением головного мозга (мозг); [ 93 ] Это может быть переведено буквально как маленький мозг . Латинское имя является прямым переводом древнегреческого παρεγκεφαλίς ( parenkephalis ), который использовался в работах Аристотеля, первого известного писателя, который описывал структуру. [ 94 ] Никакое другое имя не используется в англоязычной литературе, но исторически использовались различные греческие или латинские имена, в том числе церерам, парну , [ 95 ] энцефалион , [ 96 ] Экранист , [ 95 ] мозг сзади [ 97 ] и Parencephalis . [ 95 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]![]() Эта статья была представлена в Wikijournal of Medicine для внешнего академического рецензирования в 2016 году ( отчеты рецензентов ). Обновленный контент был реинтегрирован на странице Википедии под лицензией CC-By-S-S-3.0 ( 2016 ). Версия записи как рецензировано:
Марион Райт; Уильям Скоггс ; Финн Оруп Нильсен; и др. (30 июня 2016 г.). "Cerebellum" (PDF ) Викиджурнал медицины 3 (1). Doi : 10.15347/ wjm/ 2016.0 ISSN 2002-4 Wikidata Q44001486
Эта статья была представлена в Wikijournal of Medicine для внешнего академического рецензирования в 2016 году ( отчеты рецензентов ). Обновленный контент был реинтегрирован на странице Википедии под лицензией CC-By-S-S-3.0 ( 2016 ). Версия записи как рецензировано:
Марион Райт; Уильям Скоггс ; Финн Оруп Нильсен; и др. (30 июня 2016 г.). "Cerebellum" (PDF ) Викиджурнал медицины 3 (1). Doi : 10.15347/ wjm/ 2016.0 ISSN 2002-4 Wikidata Q44001486
- ^ Hodos W (2009). «Эволюция мозжечка». Энциклопедия нейробиологии . Берлин, Гейдельберг: Спрингер. С. 1240–1243. doi : 10.1007/978-3-540-29678-2_3124 . ISBN 978-3-540-23735-8 .
- ^ Вольф У, Рапопорт М.Дж., Швейзер Т.А. (2009). «Оценка аффективного компонента когнитивного аффективного синдрома мозжечка». Журнал нейропсихиатрии и клинических нейронаук . 21 (3): 245–53. doi : 10.1176/jnp.2009.21.3.245 . PMID 19776302 .
- ^ Schmahmann JD, Caplan D (февраль 2006 г.). «Познание, эмоции и мозжечок» . Мозг . 129 (Pt 2): 290–2. doi : 10.1093/brain/awh729 . PMID 16434422 .
- ^ Шмахманн, Джереми Д. (2019). «Мозжечок и познание» . Нейробиологические буквы . 688 (688): 62–75. doi : 10.1016/j.neulet.2018.07.005 - через Elsevier Science Direct.
- ^ Подпрыгнуть до: а беременный в дюймовый Fine EJ, Ionita CC, Lohr L (декабрь 2002 г.). «История развития мозжечка». Семинары в неврологии . 22 (4): 375–84. doi : 10.1055/s-2002-36759 . PMID 12539058 . S2CID 260317107 .
- ^ Подпрыгнуть до: а беременный Purves D, Augustine GJ, Fitzpatrick D, Hall WC, Lamantia AS, White LE (2011). Нейробиология (5 -е изд.). Сандерленд, Массачусетс: Синауэр. С. 417–423. ISBN 978-0-87893-695-3 .
- ^ Подпрыгнуть до: а беременный в Albus JS (1971). «Теория функции мозжечка». Математика Биоссии . 10 (1–2): 25–61. Citeseerx 10.1.1.14.7524 . doi : 10.1016/0025-5564 (71) 90051-4 .
- ^ Purves D, Augustine GJ, Fitzpatrick D, Hall WC, Lamantia AS, White LE (2007). Нейробиология (4 -е изд.). Нью -Йорк: WH Freeman. С. 197–200. ISBN 978-0-87893-697-7 .
- ^ Дидидзе, морской пехотинец; Levi, Allan (2017-12-15), «Обзор нейроанатомии» , Справочник по нейрохирургии, неврологии и позвоночнике для медсестер и профессионалов в области здравоохранения , Абингдон, Оксон; Нью -Йорк, Нью -Йорк: Routledge, 2018.: Routledge, стр. 53–62, ISBN 978-1-315-38276-0 Получено 2024-08-23
{{citation}}: CS1 Maint: местоположение ( ссылка ) - ^ Standring S, Borley NR, et al., Eds. (2008). "Глава 20". Анатомия Грея: анатомическая основа клинической практики (40 -е изд.). Лондон: Черчилль Ливингстон. п. 297. ISBN 978-0-8089-2371-8 .
- ^ Подпрыгнуть до: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. ведущий с Т в v В х и С Llinas RR, Walton KD, Lang EJ (2004). "Ch. 7 Cerebellum ". В пастух GM (ред.). Синаптическая организация мозга . Нью -Йорк: издательство Оксфордского университета. ISBN 978-0-19-515955-4 .
- ^ Herculano-Houzel S (2010). «Скоординированное масштабирование коры и мозжечкового числа нейронов» . Границы в нейроанатомии . 4 : 12. DOI : 10.3389/fnana.2010.00012 . PMC 2839851 . PMID 20300467 .
- ^ Lyu, Wenjiao; Wu, ye; Хуинх, Хой Минх; Ахмад, Сахар; Яп, Пью-Тайан (2024). «Мультимодальный субмиллиметр МРТ Атлас человеческого мозжечка» . Научные отчеты . 14 (1): 5622. Bibcode : 2024natsr..14.5622L . doi : 10.1038/s41598-024-55412-y . ISSN 2045-2322 . PMC 10920891 . PMID 38453991 .
- ^ Подпрыгнуть до: а беременный в дюймовый и фон глин час я Дж Ghez C, Fahn S (1985). "Церебелл" Inddel er, Schwartz JH (ред.). Принципы нейронной науки, 2 -е издание Нью -Йорк: Elsevier. Стр. 502–5
- ^ Snider RS, Stowell A (1 ноября 1944 г.). «Получение областей тактильных, слуховых и визуальных систем в мозжечке» . Журнал нейрофизиологии . 7 (6): 331–357. doi : 10.1152/jn.1944.7.6.331 . S2CID 146700933 .
- ^ Кингсли Р.Е. (2000). Краткий текст нейробиологии (2 -е изд.). Липпинкотт Уильямс и Уилкинс. ISBN 978-0-683-30460-2 .
- ^ Petersen SE, Fox PT, Posner MI, Mintun M, Raichle ME (1989). «Позитронно -эмиссионные томографические исследования обработки отдельных слов». Журнал когнитивной нейробиологии . 1 (2): 153–70. doi : 10.1162/jocn.1989.1.2.153 . PMID 23968463 . S2CID 35159122 .
- ^ Тимманн Д., Даум I (2007). «Вклад мозжечка в когнитивные функции: отчет о прогрессе после двух десятилетий исследований» . Мозжечок . 6 (3): 159–62. doi : 10.1080/14734220701496448 . PMID 17786810 . S2CID 25671398 .
- ^ Strick PL, Dum RP, Fiez JA (2009). «Церебелл и не моторная функция». Ежегодный обзор нейробиологии . 32 : 413–34. doi : 10.1146/annurev.neuro.31.060407.125606 . PMID 19555291 . S2CID 1066141 .
- ^ Бакнер Р.Л. (октябрь 2013 г.). «Церебелл и когнитивная функция: 25 лет понимания анатомии и нейровизуализации» . Нейрон . 80 (3): 807–15. doi : 10.1016/j.neuron.2013.10.044 . PMID 24183029 .
- ^ Schweighofer N, Doya K, Kuroda S (март 2004 г.). «Аминергическая нейромодуляция мозжечка: к функциональному пониманию». Исследование мозга. Обзоры исследований мозга . 44 (2–3): 103–16. doi : 10.1016/j.brainresrev.2003.10.004 . PMID 15003388 . S2CID 7352039 .
- ^ Felizola SJ, Nakura Y, Ono Y, Kikuchi K, Onodera Y, Ise K, Takase K, Wath A, Jattadady N, Rainey WE, Satoh F, Sasano H (апрель 2014 г.). «PCP4: регулятор синтеза альдостерона в человеческих тканях » Журнал молекулярной экономики 52 (2): 159–6 Doi : 10.1530/jme- 13-0 4103644PMC PMID 24403568
- ^ Eccles JC, Llinás R, Sasaki K (январь 1966 г.). «Возбуждающее синаптическое действие восхождения на волокна на клетках Purkinje в мозжечке» . Журнал физиологии . 182 (2): 268–96. doi : 10.1113/jphysiol.1966.sp007824 . PMC 1357472 . PMID 5944665 .
- ^ Подпрыгнуть до: а беременный в дюймовый Симпсон Джи, Уайли Д.Р., Де Зиув К.И. (1996). «При восхождении сигналов волокна и их последствий». Поведение Brain Sci . 19 (3): 384–398. doi : 10.1017/s0140525x00081486 .
- ^ Whitney ER, Kemper TL, Rosene DL, Bauman ML, Blatt GJ (февраль 2008 г.). «Calbindin-D28K является более надежным маркером клеток Purkinje человека, чем стандартные пятна NISSL: стереологический эксперимент». Журнал методов нейробиологии . 168 (1): 42–7. doi : 10.1016/j.jneumeth.2007.09.009 . PMID 17961663 . S2CID 10505177 .
- ^ Подпрыгнуть до: а беременный Rusanescu G, Mao J (февраль 2017 г.). «Повреждение периферического нерва вызывает нейрогенез мозга взрослых и ремоделирование» . Журнал клеточной и молекулярной медицины . 21 (2): 299–314. doi : 10.1111/jcmm.12965 . PMC 5264155 . PMID 27665307 .
- ^ Подпрыгнуть до: а беременный в Марр Д (июнь 1969 г.). «Теория мозжечкового коры» . Журнал физиологии . 202 (2): 437–70. doi : 10.1113/jphysiol.1969.sp008820 . PMC 1351491 . PMID 5784296 .
- ^ Подпрыгнуть до: а беременный в дюймовый и фон глин час я Дж k Apps R, Garwicz M (апрель 2005 г.). «Анатомические и физиологические основы обработки информации мозжечка». Природные обзоры. Нейробиология . 6 (4): 297–311. doi : 10.1038/nrn1646 . PMID 15803161 . S2CID 10769826 .
- ^ Манни Е., Петросини Л (март 2004 г.). «Столетие соматотопии мозжечка: обсуждаемое представление». Природные обзоры. Нейробиология . 5 (3): 241–9. doi : 10.1038/nrn1347 . PMID 14976523 . S2CID 30232749 .
- ^ Оскарсон О (1979). «Функциональные единицы мозжечка сагиттальных зон и микрозонов». Тенденции Neurosci . 2 : 143–145. doi : 10.1016/0166-2236 (79) 90057-2 . S2CID 53272245 .
- ^ Рапп Б. (2001). Справочник по когнитивной нейропсихологии: какие дефициты показывают о человеческом разуме . Психология пресса. п. 481. ISBN 978-1-84169-044-5 .
- ^ Подпрыгнуть до: а беременный Доя К (декабрь 2000 г.). «Дополнительные роли базальных ганглиев и мозжечка в обучении и моторном контроле». Современное мнение о нейробиологии . 10 (6): 732–9. doi : 10.1016/s0959-4388 (00) 00153-7 . PMID 11240282 . S2CID 10962570 .
- ^ Manto M, Mariën P (2015). «Синдром Шмахмана - идентификация третьего краеугольного камня клинической атаксиологии» . Cerebellum & Ataxias . 2 : 2. DOI : 10.1186/S40673-015-0023-1 . PMC 4552302 . PMID 26331045 .
- ^ Schmahmann JD, Sherman JC (апрель 1998 г.). «Когнитивный аффективный синдром мозжечка» . Мозг . 121 (4): 561–79. doi : 10.1093/Brain/121.4.561 . PMID 9577385 .
- ^ Levisohn L, Cronin-Golomb A, Schmahmann JD (май 2000). «Нейропсихологические последствия резекции опухоли мозжечка у детей: когнитивный аффективный синдром мозжечка в педиатрической популяции» . Мозг . 123 (5): 1041–50. doi : 10.1093/мозг/123.5.1041 . PMID 10775548 .
- ^ Бакнер Р.Л., Криенен Ф.М., Кастелланос А., Диас Дж.С., Йео Б.Т. (ноябрь 2011 г.). «Организация мозжечка человека, оцененная по внутренней функциональной связности» . Журнал нейрофизиологии . 106 (5): 2322–45. doi : 10.1152/jn.00339.2011 . PMC 3214121 . PMID 21795627 .
- ^ Эмоциональный мозжечок . Майкл Адамашке, Марио Манто, Деннис Дж. Л. Шуттер. Чам, Швейцария: Спрингер. 2022. ISBN 978-3-030-99550-8 Полем OCLC 1338132789 .
{{cite book}}: Cs1 maint: другие ( ссылка ) - ^ Hernáez-Goñi P, Tirapu-Ocárroz J, Iglesias-Fernández L, Luna-Lario P (ноябрь 2010). «Участие мозжечка в регуляции привязанности, эмоций и поведения» [роль мозжечка в регуляции привязанности, эмоций и поведения]. Журнал неврологии (на испанском). 51 (10): 597–609. Doi : 10.33588/rn.5110.2010394 . PMID 21069639 .
- ^ Тернер Б.М., Парадисо С., Марвел К.Л., Пирсон Р., Болес Понто Л.Л., Хичва Р.Д., Робинсон Р.Г. (март 2007 г.). «Церебелл и эмоциональный опыт» . Нейропсихология . 45 (6): 1331–41. doi : 10.1016/j.neuropsychologia.2006.09.023 . PMC 1868674 . PMID 17123557 .
- ^ Доя К (октябрь 1999). «Каковы вычисления мозжечка, базальные ганглии и кору головного мозга?». Нейронные сети . 12 (7–8): 961–974. doi : 10.1016/s0893-6080 (99) 00046-5 . PMID 12662639 .
- ^ Manto M, Marvel C, Vandervert L (2022). Новая революция в психологии и нейронаук . Швейцария: Springer Nature. ISBN 9783031060922 .
- ^ Eccles JC, Ito M, Szentágothai J (1967). Мозжечок как нейрональная машина . Springer-Verlag.
- ^ Мозжечок как нейрональная машина , с. 311
- ^ Подпрыгнуть до: а беременный Бойден Е.С., Като А., Рэймонд Дж. Л. (2004). «Обучение Cerebellum-зависимое: роль механизмов множественных пластичности». Ежегодный обзор нейробиологии . 27 : 581–609. doi : 10.1146/annurev.neuro.27.070203.144238 . PMID 15217344 . S2CID 1310007 .
- ^ Подпрыгнуть до: а беременный Houk JC, Buckingham JT, Barto AG (1996). «Модели мозжечка и моторного обучения» (PDF) . Поведение Brain Sci . 19 (3): 368–383. Citeseerx 10.1.1.118.2997 . doi : 10.1017/s0140525x00081474 . Архивировано из оригинала (PDF) 2017-08-09.
- ^ Fujita M (1982). «Модель адаптивного фильтра мозжечка». Биологическая кибернетика . 45 (3): 195–206. doi : 10.1007/bf00336192 . PMID 7171642 . S2CID 3695770 .
- ^ Гилберт П.Ф., Тах, В.Т. (июнь 1977 г.). «Активность клеток Purkinje во время моторного обучения». Исследование мозга . 128 (2): 309–28. doi : 10.1016/0006-8993 (77) 90997-0 . PMID 194656 . S2CID 40799652 .
- ^ Кристиан К.М., Томпсон Р.Ф. (2003). «Нейронные субстраты кондиционирования EyeBlink: приобретение и удержание» . Обучение и память . 10 (6): 427–55. doi : 10.1101/lm.59603 . PMID 14657256 .
- ^ Braitenberg V, Atwood RP (февраль 1958 г.). «Морфологические наблюдения на коре мозжечка». Журнал сравнительной неврологии . 109 (1): 1–33. doi : 10.1002/cne.901090102 . PMID 13563670 . S2CID 8989536 .
- ^ Braitenberg V, Heck D, Sultan F (июнь 1997 г.). «Обнаружение и генерация последовательностей как ключ к функции мозжечка: эксперименты и теория». Поведенческие и мозговые науки . 20 (2): 229–45, обсуждение 245–77. doi : 10.1017/s0140525x9700143x . PMID 10096998 . S2CID 36802745 .
- ^ Ivry RB, Spencer RM, Zelaznik HN, Deadrichsen J (декабрь 2002 г.). «Сроки мозжечка и события» . Анналы нью -йоркской академии наук . 978 (1): 302–17. Bibcode : 2002nyasa.978..302i . doi : 10.1111/j.1749-6632.2002.tb07576.x . PMID 12582062 . S2CID 27237058 .
- ^ Pellionisz A, Llinás R (1982). «Пространственное время представления в мозге. Мозжечок как прогнозирующий метрический тензор пространства-времени». Нейробиология . 7 (12): 2949–70. doi : 10.1016/0306-4522 (82) 90224-х . PMID 7162624 . S2CID 20520737 .
- ^ Horváth G (2003). «CMAC: пересмотр старой нейронной сети» (PDF) . Интеллектуальные системы управления и обработка сигналов . Архивировано из оригинала (PDF) 2020-05-20 . Получено 2009-12-24 .
- ^ Schmitz TJ (2007). «Экспертиза координации». В O'Sullivan SB, Schmitz TJ (Eds.). Физическая реабилитация . Филадельфия: Фа Дэвис. С. 193 –225. ISBN 9780803612471 .
- ^ Mariën P, Manto M (2016). Лингвистический мозжечок . Лондон, Великобритания: академическая пресса. С. 337–351. ISBN 978-0-12-801608-4 .
- ^ Гилман С (март 1998 г.). «Визуализация мозга. Вторая из двух частей». Новая Англия Журнал медицины . 338 (13): 889–96. doi : 10.1056/nejm199803263381307 . PMID 9516225 .
- ^ Подпрыгнуть до: а беременный «Ninds Ataxias и Cerebellar или Spinocerebellar Degeneration страница» . Национальные институты здравоохранения. 16 апреля 2014 года. Архивировано с оригинала 9 февраля 2015 года . Получено 2 февраля 2015 года .
- ^ Yuhas D (15 января 2016 г.). «Ветераны Ирака, Афганистан показывает изменения мозга, связанные с воздействием взрыва» . Scientific American . Архивировано с оригинала 20 января 2016 года . Получено 21 января 2016 года .
- ^ Винсент М., Хаджихани Н. (июнь 2007 г.). «Церебелл и мигрень» . Головная боль . 47 (6): 820–33. doi : 10.1111/j.1526-4610.2006.00715.x . PMC 3761082 . PMID 17578530 .
- ^ Подпрыгнуть до: а беременный «Ниндс страница информации о дегенерации мозжечка» . Национальные институты здравоохранения. 28 февраля 2014 года. Архивировано с оригинала 18 февраля 2015 года . Получено 2 февраля 2015 года .
- ^ Horvath S, Mah V, Lu AT, Woo JS, Choi OW, Jasinska AJ, Riancho JA, Tung S, Coles NS, Braun J, Vinters HV, Coles LS (май 2015 г.). «Старея мозжечка медленно в зависимости от эпигенетических часов» . Старение . 7 (5): 294–306. doi : 10.18632/старение.100742 . PMC 4468311 . PMID 26000617 .
- ^ Фрейзер Х.Б., Хайтович П., Плоткин Дж. Б., Пябо С. , Эйзен М.Б. (сентябрь 2005 г.). «Старение и экспрессия генов в мозгу приматов» . PLOS Биология . 3 (9): E274. doi : 10.1371/journal.pbio.0030274 . PMC 1181540 . PMID 16048372 .
- ^ Андерсен Б.Б., Гандерсен Х.Дж., Паккенберг Б. (ноябрь 2003 г.). «Старение человеческого мозжечка: стереологическое исследование». Журнал сравнительной неврологии . 466 (3): 356–65. doi : 10.1002/cne.10884 . PMID 14556293 . S2CID 7091227 .
- ^ Raz N, Gunning-Dixon F, Head D, Williamson A, Acker JD (2001). «Возрастные и половые различия в мозжечке и вентральных понеде: проспективное исследование MR здоровых взрослых» (PDF) . Американский журнал нейрорадиологии . 22 (6): 1161–7. PMC 7974784 . PMID 11415913 . Архивировано (PDF) из оригинала 2008-12-17.
- ^ Подпрыгнуть до: а беременный в дюймовый Альберт Р.К., Портер Р.С., ред. (2006). Руководство по диагностике и терапии Merck (18 -е изд.). Станция Уайтхаус, Нью -Джерси: библиотеки Merck Research. С. 1886–1887.
- ^ Кэмпбелл Дж., Гилберт В.М., Николаид К.Х., Кэмпбелл С. (август 1987 г.). «Скрининг ультразвука на расщелину позвоночника: черепные и мозжечковые знаки в популяции высокого риска». Акушерство и гинекология . 70 (2): 247–50. PMID 3299184 .
- ^ Подпрыгнуть до: а беременный Хэттен я, Хайнц Н. (1995). «Механизмы нейронного паттерна и спецификации в развивающемся мозжечке». Ежегодный обзор нейробиологии . 18 : 385–408. doi : 10.1146/annurev.ne.18.030195.002125 . PMID 7605067 .
- ^ Polkinghorn WR, Tarbell NJ (май 2007 г.). «Медуллобластома: онкогенез, современная клиническая парадигма и усилия по улучшению стратификации риска». Природа Клиническая практика. Онкология . 4 (5): 295–304. doi : 10.1038/ncponc0794 . PMID 17464337 . S2CID 24461280 .
- ^ Roussel MF, Hatten Me (2011). «Развитие мозжечка и медуллобластома» . Современные темы в биологии развития . 94 : 235–82. doi : 10.1016/b978-0-12-380916-2.00008-5 . ISBN 9780123809162 Полем PMC 3213765 . PMID 21295689 .
- ^ «Страница информации о синдроме Ninds Joubert» . Национальные институты здравоохранения. 23 декабря 2013 года. Архивировано с оригинала 4 января 2015 года . Получено 9 января 2015 года .
- ^ «Информационная страница Ninds Dandy-Walker» . Национальные институты здравоохранения. 14 февраля 2014 года. Архивировано с оригинала 4 января 2015 года . Получено 9 января 2015 года .
- ^ «Ниндс страница информации о гипоплазии мозжечка» . Национальные институты здравоохранения. 29 сентября 2011 года. Архивировано с оригинала 4 января 2015 года . Получено 9 января 2015 года .
- ^ «Информационный бюллетень по развитию развития Chiari» . Национальные институты здравоохранения. 10 декабря 2014 года. Архивировано с оригинала 27 октября 2011 года . Получено 9 января 2015 года .
- ^ «Ninds Dyssynergia cerebellaris myoclonica страница информации» . Национальные институты здравоохранения. 14 февраля 2011 года. Архивировано с оригинала 16 февраля 2015 года . Получено 1 февраля 2015 года .
- ^ «Ninds Olivopontocerebellar Information Atrophy страница» . Национальные институты здравоохранения. 16 апреля 2014 года. Архивировано с оригинала 27 января 2012 года . Получено 9 января 2015 года .
- ^ "НИНДСА ПАРАНЕЛИЧЕСКИХ СИНДРОМ СТРАНИЦА ИНФОРМАЦИЯ" . Национальные институты здравоохранения. 12 марта 2009 г. Архивировано с оригинала 4 января 2015 года . Получено 9 января 2015 года .
- ^ «Ниндс Вернике-Корсакофф Информационная страница» . Национальные институты здравоохранения. 14 февраля 2007 года. Архивировано с оригинала 4 января 2015 года . Получено 9 января 2015 года .
- ^ Moulton EA, Schmahmann JD, Becerra L, Borsook D (октябрь 2010 г.). "Церебелл и боль: пассивный интегратор или активный участник?" Полем Обзоры исследований мозга . 65 (1): 14–27. doi : 10.1016/j.brainresrev.2010.05.005 . PMC 2943015 . PMID 20553761 .
- ^ Бауманн О., Борра Р.Дж., Бауэр Дж. М., Каллен К.Е., Хабас С., Иври Р.Б., Легио М., Маттингли Дж.Б., Молинари М., Моултон Э.А., Полин М.Г., Павлова М.А., Шмахман Д.Д., Соколов А.А. (апрель 2015). «Консенсусная статья: роль мозжечка в процессах восприятия» . Мозжечок . 14 (2): 197–220. doi : 10.1007/s12311-014-0627-7 . PMC 4346664 . PMID 25479821 .
- ^ Подпрыгнуть до: а беременный в дюймовый Bell CC, Han V, Sawtell NB (2008). «Строительные структуры и их последствия для функции мозжечка». Ежегодный обзор нейробиологии . 31 : 1–24. doi : 10.1146/annurev.neuro.30.051606.094225 . PMID 18275284 . S2CID 14536411 .
- ^ Woodhams PL (июль 1977 г.). «Ультраструктура аналога мозжечка в осьминоге». Журнал сравнительной неврологии . 174 (2): 329–45. doi : 10.1002/cne.901740209 . PMID 864041 . S2CID 43112389 .
- ^ Подпрыгнуть до: а беременный Ромер А.С., Парсонс Т.С. (1977). Тело позвоночных . Филадельфия: Holt-Saunders International. п. 531. ISBN 978-0-03-910284-5 .
- ^ Shi Z, Zhang Y, Meek J, Qiao J, Han VZ (август 2008 г.). «Нейрональная организация уникальной специализации мозжечка: Valvula Cerebelli из мормиридной рыбы» . Журнал сравнительной неврологии . 509 (5): 449–73. doi : 10.1002/cne.21735 . PMC 5884697 . PMID 18537139 .
- ^ Уивер А.Х. (март 2005 г.). «Взаимная эволюция мозжечка и неокортекс у окаменелостей» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (10): 3576–80. Bibcode : 2005pnas..102.3576w . doi : 10.1073/pnas.0500692102 . PMC 553338 . PMID 15731345 .
- ^ Подпрыгнуть до: а беременный в Schoenemann Pt (1 декабря 2009 г.). «Эволюция мозга и языка». Изучение языка . 59 : 162–186. doi : 10.1111/j.1467-9922.2009.00539.x . PMID 22230641 .
- ^ Macleod CE, Zilles K, Schleicher A, Rilling JK, Gibson KR (апрель 2003 г.). «Расширение Neocerebellum в Hominoidea». Журнал человеческой эволюции . 44 (4): 401–29. Bibcode : 2003jhume..44..401m . doi : 10.1016/s0047-2484 (03) 00028-9 . PMID 12727461 .
- ^ Робертс П.Д., Портфоры CV (июнь 2008 г.). «Принципы конструкции сенсорной обработки в мозжечковых структурах. Ранняя стадия обработка электросенсорных и слуховых объектов». Биологическая кибернетика . 98 (6): 491–507. doi : 10.1007/s00422-008-0217-1 . PMID 18491162 . S2CID 14393814 .
- ^ Bower JM (1997). «Глава 27 Является ли мозжечок сенсора ради моторики, или двигатель ради Sensory: вид с усов крысы?». Мозжечок: от структуры к контролю . Прогресс в исследовании мозга. Тол. 114. С. 463–96. doi : 10.1016/s0079-6123 (08) 63381-6 . ISBN 978-0-444-82313-7 Полем PMID 9193161 .
- ^ Heiney SA, Kim J, Augustine GJ, Medina JF (февраль 2014 г.). «Точный контроль кинематики движения путем оптогенетического ингибирования активности клеток Пуркинье» . Журнал нейробиологии . 34 (6): 2321–30. doi : 10.1523/jneurosci.4547-13.2014 . PMC 3913874 . PMID 24501371 .
- ^ Witter L, Canto CB, Hoogland TM, de Gruijl Jr, de Zeeuw CI (2013). «Сила и время моторных ответов, опосредованных отскоком стрельбы в ядрах мозжечка после активации клеток Пуркинье» . Границы в нейронных цепях . 7 : 133. doi : 10.3389/fncir.2013.00133 . PMC 3748751 . PMID 23970855 .
- ^ Кларк Э., О'Мэлли CD (1996). «Гл. 11: мозжечок». Человеческий мозг и спинной мозг (2 -е изд.). Норман издательство. п. 629. ISBN 978-0-930405-25-0 .
- ^ Ито М (декабрь 2002 г.). «Исторический обзор значения мозжечка и роль клеток Пуркинье в моторном обучении». Анналы нью -йоркской академии наук . 978 (1): 273–88. Bibcode : 2002nyasa.978..273i . doi : 10.1111/j.1749-6632.2002.tb07574.x . PMID 12582060 . S2CID 22860609 .
- ^ Льюис К.Т., короткий С (1879). Латинский словарь, основанный на издании Эндрюса латинского словаря Фрейнда . Оксфорд: Clarendon Press.
- ^ Маршалл Л.Х., Магун Х.В. (1998). Открытия в человеческом мозге. Нейронаука предыстория, структура мозга и функция . Тотова: Humana Press.
- ^ Подпрыгнуть до: а беременный в Foster FD (1891). Иллюстрированный медицинский словарь . Нью -Йорк: Д. Эпплтон и Компания.
- ^ Краус Ла (1844). Критическая этимологическая медицинская лексика (третье издание) . Göttingen: Verlag из книжного магазина Deutian и Dieterich.
- ^ Schreger CH (1805). Синонимия Анатомика. Синонимика анатомической номенклатуры . Фюрт.
Внешние ссылки
[ редактировать ]- Llinas R, Negello MN (2015). "Cerebellum" . Scholaredia . 10 (1): 4606. Bibcode : 2015schpj..10.4606l . doi : 10.4249/Scholaredia.4606 .
- База данных с центром мозжечка
- Справочник по заболеваниям мозжечка и мозжечка - Manto, M., Gruol, DL, Schmahmann, J., Koibuchi, N., Rossi, F. (Eds.) - Springer - Нью -Йорк
- Окрашенные изображения срезов мозга, которые включают «мозжечок» в проекте Brainmaps
- Неполный мозг человека раскрывает роль мозжечка в мысли и эмоциях
- Женщина, живущая без мозжечка
- Основы заболеваний мозжечка и мозжечка . Gruol, DL, Koibuchi, N., Manto, M., Molinari, M., Schmahmann, JD, Shen, Y. (Eds.). Springer, New York, 2016
- Изображения гистологии мозжечка
- Cerebellum - Journal (Springer Nature)
- Мозжечок и атаксиас - журнал (Biomed Central)