Регуляторная Т -клетка
Регуляторные Т-клетки ( Tregs / ˈ t iː r ɡ ɡ ɡ / или t- рег ), ранее известные как супрессорные Т-клетки , являются субпопуляцией Т-клеток , которые модулируют иммунную систему , поддерживают толерантность к самоантигенам и предотвращают аутоиммунное заболевание Полем T reg Клетки являются иммуносупрессивными и, как правило, подавляют или подавляют индукцию и пролиферацию эффекторных Т -клеток . [ 1 ] Клетки T REG экспрессируют биомаркеры CD4 , FOXP3 и CD25 и, как полагают, получены из той же линии , что и наивный CD4 + ячейки [ 2 ] Поскольку эффекторные Т -клетки также экспрессируют CD4 и CD25, клетки T REG очень трудно эффективно различать по эффекторному CD4 + , затрудняя их учиться. Исследования показали, что бета-фактор роста трансформирующего цитокина (TGF-β) необходим для T- Reg от наивного CD4 дифференцированных клеток + клетки и важны для поддержания t -рег клеток гомеостаза . [ 3 ]
мыши Модели предположили, что модуляция клеток T -Reg может лечить аутоиммунное заболевание и рак и может облегчить трансплантацию органов [ 4 ] и заживление ран . [ 5 ] Их последствия для рака сложны. Клетки T REG , как правило, активируются у людей с раком, и они, по -видимому, рекрутируются на место многих опухолей . Исследования как на людях, так и на моделях на животных причастны к тому, что большое количество клеток Т -рег в микроокружении опухоли указывает на плохой прогноз , и, как полагают, клетки Т -Рег подавляют иммунитет опухоли, что мешает врожденной способности организма к росту раковых раковых ячейки [ 6 ] Исследование иммунотерапии изучает, как регуляция Т -клеток может быть использована при лечении рака. [ 7 ]
Популяции
[ редактировать ]Т -регуляторные клетки являются компонентом иммунной системы, которая подавляет иммунные ответы других клеток. Это важная «самоуверенная», встроенная в иммунную систему для предотвращения чрезмерных реакций. Регуляторные Т-клетки бывают разных форм с наиболее хорошо понятыми, которые экспрессируют CD4, CD25 и FOXP3 (CD4 + CD25 + регуляторные Т -клетки). Эти клетки Т -рег отличаются от вспомогательных Т -клеток . [ 8 ] Другим регуляторным подмножествами Т -клеток является t -reg 17 клетки. [ 9 ] Регуляторные Т -клетки участвуют в закрытии иммунных ответов после того, как они успешно устранены вторжения в организмы, а также в предотвращении аутоиммунитета. [ 10 ]
CD4 + Foxp3 + CD25 (высокие) регуляторные Т -клетки были названы «природными» регуляторными Т -клетками [ 11 ] отличать их от популяций Т -клеток «супрессора», которые генерируются in vitro . Дополнительные регуляторные популяции Т -клеток включают TR1 , T H 3, CD8 + CD28 − и QA-1 ограниченные Т-клетки. Вклад этих групп населения в самоотверждение и иммунный гомеостаз менее четко определен. Foxp3 можно использовать в качестве хорошего маркера для CD4 мыши + CD25 + Т -клетки, хотя недавние исследования также показали доказательства экспрессии FOXP3 в CD4 + CD25 − Т -клетки. У людей FOXP3 также экспрессируется недавно активированными обычными Т -клетками и, следовательно, не идентифицирует T -рег . [ 12 ]
Разработка
[ редактировать ]Все Т -клетки происходят из клеток -предшественников в костном мозге , которые становятся приверженными их линии в тимусе . Все Т -клетки начинаются как CD4 - CD8 - TCR - клетки на стадии DN (двойной негативной), где отдельная ячейка будет переставлять свои гены рецепторов Т -клеток с образованием уникальной, функциональной молекулы, которую они, в свою очередь, испытывают на клетки в тимусной коре для минимального уровня взаимодействия с самого MHC . Если они получают эти сигналы, они пролиферируют и экспрессируют как CD4, так и CD8, становясь двойными клетками. Выбор T REG встречается на радиоприеменных гематопоэтических, полученных MHC класса II, экспрессирующих клеток в мозговом мозге или корпускулах Хассалла в тимусе. На стадии DP (с двойным положительным) они выбираются путем их взаимодействия с клетками внутри тимуса, начинают транскрипцию Foxp3 и становятся T -рег -клетками, хотя они не могут начинать экспрессировать Foxp3 до одноположительной стадии, в этот момент они функциональны . T reg не имеют ограниченной экспрессии TCR NKT или γΔ T -клеток; Т Рег имеет большее разнообразие TCR, чем эффекторные Т-клетки, смещенные в сторону самопептидов.
Процесс отбора T REG определяется сродством взаимодействия с комплексом MHC самопептида. Выбор, чтобы стать T REG - это процесс « Златовласка », то есть не слишком высокий, не слишком низкий, но просто правильно; [ 13 ] Т -клетка, которая получает очень сильные сигналы, будет подвергаться апоптотической смерти; Ячейка, которая получает слабый сигнал, выживет и будет выбран, чтобы стать эффекторной ячейкой. Если Т -клетка получает промежуточный сигнал, то она станет регуляторной ячейкой. Из-за стохастической природы процесса активации Т-клеток все популяции Т-клеток с данным TCR будут в конечном итоге с смесью TFE и T EFF -относительные пропорции, определяемые сродством Т-клет -Mhc. Даже на мышиных моделях с TCR-трансгенными клетками, выбранными на строме специфического антигена, делеция или конверсия не завершены.
После взаимодействия с комплексом MHC с самопептидом Т-клетка должна активировать IL-2R , CD25 и TNFR Superbamily Memporte , OX40 и TNFR2 , чтобы стать CD25 + Foxp3 - T -рег -клетки -предшественник. Чтобы стать зрелым T REG , должен быть активирован фактор транскрипции FOXP3, который обусловлен γ-цепью (CD132), зависимыми цитокинами, в частности IL-2 и/или IL-15. [ 14 ] [ 15 ] Только только IL-2 недостаточно для стимуляции экспрессии Foxp3, необходимы другие цитокины. Принимая во внимание, что IL-2 продуцируется самореактивными тимоцитами, IL-15 продуцируется стромальными клетками тимуса, в основном MTEC и CTEC . [ 14 ]
другое подмножество предшественников T. Недавно было идентифицировано В этом подмножестве отсутствует CD25 и имеет низкую экспрессию Foxp3. Его развитие в основном зависит от IL-15. Это подмножество имеет более низкую аффинность к самогенам, чем CD25 + Foxp3 высокий подмножество. Обе подмножества генерируют зрелые клетки Т- рег после стимуляции с помощью IL-2 с сопоставимой эффективностью как in vitro , так и in vivo . CD25 + Foxp3 высокий Предшественники демонстрируют повышенный апоптоз и развиваются в зрелые клетки T -рег с более высокой кинетикой, чем Foxp3 низкий предшественники. [ 16 ] T regs, полученные из CD25 + Foxp3 высокий Предшественники защищают от экспериментального аутоиммунного энцефаломиелита, тогда как те, которые получены из CD25 + Foxp3 низкий Предшественники защищают от Т-клеточного колита . [ 14 ]
Зрелые CD25+ Foxp3+ Tregs также могут быть разделены на два разных подмножества на основе уровня экспрессии CD25, GITR и PD-1 . Трег, экспрессирующие низкие количества CD25, GITR и PD-1, ограничивают развитие колита путем стимулирования преобразования обычного CD4 + Т -клетки в PTREG. Трегс, высоко экспрессирующие CD25, GITR и PD-1, являются более самореактивными и контрольными лимфопролиферациями в периферических лимфатических узлах-они могут иметь способность защищать от аутоиммунных расстройств. [ 14 ]
Foxp3 + Генерация T reg в тимусе задерживается на несколько дней по сравнению с клетками T EFF и не достигает уровня взрослых ни в тимусе, ни в периферии до примерно трех недель после рода. Клетки T REG требуют CD28 костимуляции , а экспрессия B7.2 в значительной степени ограничена мозговым веществом, развитие которого, по-видимому, параллельна развитию FoxP3 + ячейки Было высказано предположение, что эти два связаны, но никакой окончательной связи между процессами еще не было показано. TGF-β не требуется для функциональности T- reg , в тимусе, поскольку тимусные t- рег , из нечувствительных мышей TGF-β TGFβRII-DN функциональны.
Рециркуляция тимуса
[ редактировать ]Было замечено, что некоторые Foxp3 + Клетки T reg рециркулируют обратно в тимус, где они развивались. Этот t -рег был в основном присутствовал в медедах тимуса, которая является основным сайтом дифференцировки клеток T -рег . [ 17 ] Присутствие этих клеток в тимусе или добавление в культуру ткани тимуса плода подавляет развитие новых клеток T -рег на 34–60%, [ 17 ] Но клетки TCONV не поражены. Это означает, что рециркуляция T reg в тимус ингибировал только De novo Development T -рег -клеток. Молекулярный механизм этого процесса работает из-за способности T REG к адсорбированию IL-2 из микроокружения, тем самым способный индуцировать апоптоз других Т-клеток, которые нуждаются в IL-2 в качестве основного фактора роста. [ 18 ] Рециркулирующие t-рег клетки в тимусе экспрессируют высокое количество высокоаффинной рецепторной α-цепи IL-2 ( CD25 ), кодируемой геном IL2RA , которые собирают IL-2 из медового мозга тимуса и снижают его концентрацию. Новый сгенерированный Foxp3 + Клетки T REG в тимусе не имеют столь большого количества экспрессии IL2RA . [ 17 ] IL-2 является цитокином, необходимым для развития T- рег -клеток в тимусе. Это важно для пролиферации и выживания Т-клеток, но в случае его дефицита IL-15 может быть заменен. Тем не менее, развитие клеток T -рег зависит от IL-2. [ 19 ] У людей была обнаружена популяция CD31 отрицательных T -рег -клеток в тимусе. [ 17 ] CD31 может использоваться в качестве маркера новых генерируемых T -рег -клеток, как и другие Т -лимфоциты. Зрелые и периферические клетки Т -рег снизили его экспрессию. [ 20 ] Таким образом, возможно, что этот регуляторный механизм развития Tymic T -рег также является функциональным у людей.
Вероятно, существует также положительная регуляция развития клеток Tymic T REG, вызванного рециркуляционными клетками TG в тимус. Была обнаружена население CD24 Low Foxp3 + в тимусе с повышенной экспрессией IL-1R2 ( IL1R2 ) по сравнению с периферическими клетками T REG . [ 21 ] [ 22 ] Высокая концентрация IL-1β , вызванная воспалением, уменьшает DE-novo развитие клеток TG в тимусе. [ 22 ] Присутствие рециркуляционных клеток Т- рег в тимусе с высокой экспрессией IL1R2 во время воспалительных условий помогает поглотить IL-1β и снизить его концентрацию в микроокружении мозга, таким образом, они помогают в развитии клеток De novo T REG . [ 22 ] Высокая концентрация IL-1β , вызванная воспалением, уменьшает DE-novo развитие клеток TG в тимусе. [ 22 ] Связывание IL-1β с IL1R2 на поверхности T -рег- клеток не вызывает никакой передачи сигнала, поскольку не существует внутриклеточного ( TIR )-рецепторного домена рецептора-1, который обычно присутствует в врожденных иммунных клетках. [ 23 ]
Функция
[ редактировать ]Иммунная система должна быть в состоянии различать себя и неспособность. Когда самооценка самоотрации/не-самого себя не удается, иммунная система разрушает клетки и ткани тела и в результате вызывает аутоиммунные заболевания . Регуляторные Т-клетки активно подавляют активацию иммунной системы и предотвращают патологическую самореактивность, то есть аутоиммунное заболевание. Критические регуляторные Т -клетки, которые играют в иммунной системе, подтверждается тяжелым аутоиммунным синдромом, который является результатом генетического дефицита в регуляторных Т -клетках ( синдром IPEX - см. Также ниже).
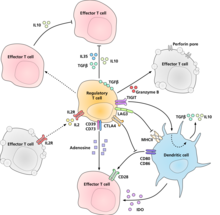
Молекулярный механизм, с помощью которого регуляторные Т -клетки проявляют свою супрессору/регуляторную активность, не был окончательно охарактеризован и является субъектом интенсивных исследований. Эксперименты in vitro дали смешанные результаты относительно потребности контакта клеток к клеткам с подавляемой клеткой. Следующее представляет некоторые из предложенных механизмов иммунного подавления:
- Регуляторные Т -клетки продуцируют ряд ингибирующих цитокинов. К ним относятся TGF-β, [ 24 ] Интерлейкин 35 , [ 25 ] и интерлейкин 10 . [ 26 ] Также представляется, что регуляторные Т-клетки могут индуцировать другие типы клеток для экспрессии интерлейкина-10. [ 27 ]
- Регуляторные Т -клетки могут продуцировать гранзим B , который, в свою очередь, может вызывать апоптоз эффекторных клеток. Сообщается, что регуляторные Т -клетки от мышей с дефицитом гранзима B являются менее эффективными супрессорами активации эффекторных Т -клеток. [ 28 ]
- Обратная передача сигналов посредством прямого взаимодействия с дендритными клетками и индукции иммуносупрессивной индолий 2,3-диоксигеназы . [ 29 ]
- Сигнализация через эктоэферменты CD39 и CD73 с производством иммуносупрессивного аденозина . [ 30 ] [ 31 ]
- Через прямое взаимодействие с дендритными клетками с помощью LAG3 и Tigit . [ 32 ] [ 33 ] Этот обзор взаимодействия T -рег с дендритными клетками обеспечивает различие между механизмами, описанными для клеток человека по сравнению с клетками мыши. [ 34 ]
- Другой механизм управления-через петлю обратной связи IL-2. Активируемые антиген Т-клетки продуцируют IL-2, который затем действует на рецепторы IL-2 на регуляторные Т-клетки, предупреждающие их о том, что высокая активность Т-клеток возникает в области, и они устанавливают супрессорный ответ на них. Это отрицательный цикл обратной связи, чтобы гарантировать, что чрезмерная реакция не происходит. Если фактическая инфекция присутствует, другие воспалительные факторы подавляют подавление. Нарушение петли приводит к гиперреактивности, регуляция может изменить силу иммунного ответа. [ 35 ] Связанное предположение в отношении интерлейкина 2 заключается в том, что активированные регуляторные Т -клетки занимают интерлейкин 2 настолько, что они лишают эффекторных Т -клеток достаточных, чтобы избежать апоптоза. [ 18 ]
- Основным механизмом подавления регуляторными Т-клетками является профилактика костимуляции через CD28 на эффекторных Т-клетках путем действия молекулы CTLA-4 . [ 36 ]
Естественные и индуцированные регуляторные Т -клетки
[ редактировать ]Т -регуляторные лимфоциты развиваются во время онтогеники либо в тимусе , либо на периферии. Соответственно, они разделены на естественные и индуцированные Т -регуляторные клетки. [ 37 ]
Природные Т -регуляторные лимфоциты (TTREGS, NTREGS) характеризуются непрерывной экспрессией рецептора FOXP3 и Т -клеток (TCR) с относительно высокой аутоаффинностью. Эти клетки преимущественно обнаруживаются в организме в кровотоке или лимфатических узлах и служат в основном для обеспечения толерантности к аутоантигенам. [ 37 ]
Индуцированные (периферические) T-регуляторные клетки (ITREGS, PTREGS) возникают в определенных ситуациях в присутствии IL-2 и TGF-B на периферии и начинают экспрессировать FOXP3, становясь функциональным эквивалентом клеток TTREG. Итреги, однако, обнаруживаются в основном в тканях периферических барьеров, где они в основном участвуют в предотвращении воспаления в присутствии внешних антигенов. [ 37 ]
Основные особенности, которые дифференцируют клетки TTREG и ITREG, включают гелиос и нейропилин-1 , присутствие которых предполагает происхождение в тимусе. Другая особенность, отличающая эти две популяции клеток Treg, - это стабильность экспрессии Foxp3 в разных настройках. [ 37 ]
Индуцированные t -регуляторные клетки
[ редактировать ]T (IT Reg ) (CD4 Индуцированные регуляторные клетки + CD25 + Foxp3 + ) являются подавляющими клетками, участвующими в толерантности. подавляют регдации Было показано, что клетки пролиферацию Т -клеток и экспериментальные аутоиммунные заболевания. Эти клетки включают клетки T REG 17 . IT -рег -клетки развиваются из зрелого CD4 + Обычные Т -клетки за пределами тимуса: определяющее различие между природными регуляторными клетками T (nt Reg ) и IT -клетками . Хотя клетки REG и NT REG имеют аналогичную функцию, в последнее время было показано, что клетки Reg Reg являются «важным нередуклевым регуляторным подмножества, которое дополняет клетки NT Reg , частично расширяя разнообразие TCR в пределах регуляторных ответов». [ 38 ] Острое истощение бассейна IT Reg в моделях мышей привело к воспалению и потере веса. Вклад клеток NT REG в сравнении с IT REG -клетками в поддержание толерантности неизвестен, но оба важны. Эпигенетические различия наблюдались между NT REG и IT -клетками , причем первые имеют более стабильную экспрессию FOXP3 и более широкое деметилирование .
Среда на небольшом кишечнике с высоким содержанием витамина А и является местом, где образуется ретиноевая кислота. [ 39 ] Ретиноевая кислота и TGF-бета, продуцируемые дендритными клетками в рамках этой области сигнала для производства регуляторных Т-клеток. [ 39 ] Витамин А и TGF-бета способствуют дифференцировке Т-клеток в регуляторные Т-клетки, противоположные клеткам T H 17 , даже в присутствии IL-6 . [ 40 ] [ 41 ] Кишечная среда может привести к индуцированным регуляторными Т-клетками с TGF-бета и ретиноевой кислотой, [ 42 ] Некоторые из которых экспрессируют лектин-подобный рецептор CD161 и специализируются на поддержании целостности барьера путем ускорения заживления ран. [ 43 ] T -re S в кишечнике дифференцируются от наивных Т -клеток после введения антигена. [ 44 ] Недавно было показано, что регуляторные Т-клетки человека могут быть индуцированы как из наивных, так и из предварительно связанных клеток Th1, так и из клеток Th17 [ 45 ] , полученную из паразитов Используя мимику TGF-β , секретируемое Heligmosomoides Polygyrus и называемым HP -Tgm ( H. polygyrus tgf-β-мимик). [ 46 ] [ 47 ] HP -TGM может индуцировать мышиные FOXP3, экспрессирующие регуляторные Т -клетки, которые были стабильными в присутствии воспаления in vivo . [ 48 ] HP -TGM-индуцированные регуляторные Т-клетки FoxP3+ были стабильны в присутствии воспаления и имели повышенные уровни CD25 , CTLA4 и снижение метилирования в Foxp3 T Reg DEG-Specificated по сравнению с TGF-β-индуцированным TE . [ 45 ]
RORγt+ регуляторные Т -лимфоциты
[ редактировать ]Приблизительно 30–40% клеток Colonic Foxp3+ Treg экспрессируют транскрипционный фактор RORγt. [ 49 ] Итреги способны дифференцироваться в RORγT -экспрессирующие клетки и, таким образом, получать фенотип клеток Th17 . Эти клетки связаны с функциями слизистых лимфоидных тканей, таких как кишечный барьер. У кишечной пластинки пропита обнаружены 20-30% регуляторных клеток Foxp3+ T, экспрессирующих Roryt, и эта высокая доля сильно зависит от присутствия сложного кишечного микробиома. У мышей без зародышей (GF) популяция регуляторных клеток RORγT+ T сильно снижается, тогда как реколонизация специфической микробиотой без патогенов (SPF) восстанавливает нормальное количество этих лимфоцитов в кишечнике. Механизм, с помощью которого микробиота кишечника индуцирует образование клеток RORγT+ Treg, включает в себя продукцию жирных кислот с короткой цепью (SCFA), от которых зависит эта индукция. SCFA-это побочный продукт ферментации и расщепления пищевых волокон, поэтому мыши без микробов имеют очень низкие концентрации как SCFA, так и RORγT Treg-клеток. Индукция клеток RORγT Treg также зависит от присутствия Дендритные клетки у взрослых, клетки TheTis в неонатальном и антигенном представлении MHC II . [ 50 ] [ 51 ]
Клетки RORγT+ Treg не присутствуют в тимусе и не экспрессируют гелиос или нейропилин-1 , но имеют высокую экспрессию CD44 , IL-10 , ICOS, CTLA-4 и нуклеотидаз CD39 и CD73, предполагая сильную регуляторную функцию. [ 50 ]
Функция регуляторных Т -лимфоцитов RORγT+
[ редактировать ]Индукция клеток RORγT+ Treg в лимфатических узлах тонкой кишки имеет решающее значение для установления толерантности к элерансию в кишечнике . Эти клетки особенно важны в профилактике пищевой аллергии. Одним из механизмов является производство супрессивных молекул, таких как цитокин IL-10 . Эти клетки также подавляют популяцию клеток Th17 и ингибируют выработку IL-17 , тем самым подавляя провоспалительный ответ. [ 50 ]
У мышей Colonic RORγT+ Tregs отсутствуют в течение первых двух недель после рождения. Генерация RORγT+ Treg в начале рождения имеет важное значение для предотвращения развития различных иммунопатологий кишечника в более позднем возрасте. Особенно важным является период постепенного перехода от полагаться исключительно на материнское молоко до включения твердой пищи, в возрасте от 15 до 20 дней, когда вводится большое количество микробных антигенов и комменсальная микробиота оседает в кишечнике. В течение этого времени защитные клетки RORγT+ Treg индуцируются микробными антигенами, а нормальный кишечный гомеостаз поддерживается путем индукции толерантности к комменсальной микробиоте. Отсутствие индукции клеток RORγT+ Treg привела у мышей к развитию тяжелого колита . [ 52 ] Количество индуцированных ранней жизнью RORγT+ Tregs влияет материнское молоко, особенно количество антител IgA , присутствующих в материнском молоке. У взрослых мышей RORγt+ Tregs и IgA демонстрируют взаимное ингибирование. Аналогичным образом, мыши, ухаживающие на приемных матерях с более высокими титрами IgA в их молоке, будут развиваться меньше RORγT+ Tregs по сравнению с теми, которые питались молоком, содержащим более низкие титры IgA. [ 53 ]
RORγt+ Tregs также были показаны за их значение в пероральной терпимости и профилактике пищевой аллергии. Младенцы с развитой пищевой аллергией имеют различный состав фекальной микробиоты по сравнению со здоровыми младенцами и имеют повышенную IgE, связанную с фекальной микробиотой и снижением секреторной IGA. У мышей защита от пищевой аллергии была вызвана внедрением видов Clostridiales и Bacteroidales . После их введения происходит расширение клеток кишечника RORγT+ Treg в пользу GATA3+ Treg, опосредуя защиту от аллергии. [ 54 ]
Дефицит триптофана , незаменимая аминокислота, изменяет комменсальный метаболизм микробиоты, что приводит к расширению клеток RORγT+ Treg и восстановлению клеток GATA3+ Treg. Эта индукция, возможно, регулируется стимуляцией арилггковерного рецептора метаболитами, продуцируемыми комменсальными бактериями с использованием триптофана в качестве источника энергии. [ 55 ]
Более низкое количество клеток RORγT+ Treg присутствует у мышей, свободных от зародышевых, колонизированных микробиотой, связанной с воспалительным заболеванием кишечника, по сравнению с мышами без зародышей, колонизированной здоровой микробиотой. Дисрегуляция клеток RORγT+ Treg способствует расширению клеток TH2 , а более низкое количество клеток RORγT+ Treg компенсируется повышенными клетками Helios+ Treg. Как именно может защитить rorγt+ tregs от колита. [ 56 ]
RORγt+ регуляторные Т -лимфоциты при раке
[ редактировать ]Патологический может быть вовлечение регуляторных Т -клеток RORγT+ при колоректальном раке. Было обнаружено, что RORγT+ Tregs, способные экспрессировать IL-17, расширяются при колоректальном раке, и по мере развития рака они теряют способность экспрессировать противовоспалительный IL-10. Точно так же такие RORγT+ Tregs, экспрессирующие IL-17, расширяются в слизистой оболочке пациентов с болезнью Крона. [ 57 ] [ 58 ] Истощение RORγT+ Tregs у мышей с колоректальным раком вызывало повышение реактивности опухолевых специфических Т-клеток и улучшение иммунного наблюдения рака. Это улучшение не вызвано потерей IL-17, поскольку это было доказано, что способствует прогрессированию рака. [ 58 ] У опухолей мышей с условным нокаутом RORγt+ Tregs подтверждается подавление IL-6 , снижение IL-6, экспрессирующих CD11C+ дендритные клетки и сверхэкспрессию CTLA-4 . IL-6 опосредует активацию транскрипционного фактора STAT3 , который является критическим для пролиферации раковых клеток. [ 59 ]
Гата3+ регуляторные Т -лимфоциты
[ редактировать ]Другим важным подмножением клеток Treg являются клетки GATA3+ Treg, которые реагируют на IL-33 в кишечнике и влияют на регуляцию эффекторных Т-клеток во время воспаления. В отличие от клеток RORγT+ TREG, эти клетки экспрессируют Helios и не зависят от микробиома. [ 51 ] [ 60 ]
GATA3+ T REGS являются основными иммуносупрессорами во время воспаления кишечника, и T REG используют GATA3 для ограничения воспаления тканей. Эта клеточная популяция также ограничивает иммунитет Th17 T-клеток в кишечнике, потому что Gata3-дефицитные T REG экспрессируют более высокий RORC и IL-17A . транскрипт [ 61 ]
Болезнь
[ редактировать ]Важным вопросом является то, как иммуносупрессивная активность регуляторных Т -клеток модулируется в течение постоянного иммунного ответа. В то время как иммуносупрессивная функция регуляторных Т -клеток предотвращает развитие аутоиммунного заболевания, это нежелательно во время иммунных реакций на инфекционные микроорганизмы.
Инфекции
[ редактировать ]После встречи с инфекционными микроорганизмами активность регуляторных Т -клеток может быть подавлена, прямо или косвенно, другими клетками, чтобы облегчить устранение инфекции. Экспериментальные данные моделей мыши предполагают, что некоторые патогены могли развиваться для манипулирования регуляторными Т -клетками для иммуносупрессии хозяина и таким образом усилить их собственное выживание. Например, сообщалось, что регуляторная активность Т-клеток увеличивается в нескольких инфекционных контекстах, таких как ретровирусные инфекции (наиболее известными из которых является ВИЧ), микобактериальные инфекции (например, туберкулез [ 62 ] ) и различные паразитические инфекции, включая лейшманию и малярию .
Клетки T REG играют основные роли во время ВИЧ -инфекции. Они подавляют иммунную систему, таким образом ограничивая клетки-мишени и снижая воспаление, но это одновременно нарушает клиренс вируса с помощью клеточного иммунного ответа и усиливает резервуар, выдвигая CD4 + Т -клетки в состояние покоя, включая инфицированные клетки. Кроме того, клетки T REG могут быть инфицированы ВИЧ, увеличивая размер резервуара ВИЧ напрямую. Таким образом, клетки T Reg исследуются как мишени для исследования лечения ВИЧ. [ 63 ] Некоторые стратегии истощения клеток T -рег были протестированы у SIV инфицированных нечеловеческих приматов и показали, что они вызывают вирусную реактивацию и усиление специфичного CD8 SIV CD8 + Т -клеточные ответы. [ 64 ]
Регуляторные Т -клетки играют большую роль в патологии висцерального лейшманиоза и в предотвращении избыточного воспаления у пациентов, вылеченных висцеральным лейшманиозом.
Когда
[ редактировать ]Есть некоторые доказательства того, что клетки T -рег могут быть дисфункциональными и движущими нейровоспалением при боковом амиотрофическом склерозе из -за более низкой экспрессии FOXP3. [ 65 ] В настоящее время в клиническом испытании I экспансия T -рег для последующей аутологичной трансплантации после того, как перспективные результаты были получены в клиническом исследовании I. в настоящее время исследуется [ 66 ]
Беременность
[ редактировать ]В то время как регуляторные Т -клетки увеличиваются благодаря поликлональной экспансии как системной, так и на местном уровне во время здоровых беременностей, чтобы защитить плод от материнского иммунного ответа (процесс, называемый материнской иммунной толерантностью), данные свидетельствуют о том, что это поликлональное расширение нарушено у преэкламптических матерей и их потомков. [ 67 ] Исследования предполагают снижение производства и развития регуляторных Т -клеток во время преэклампсии, может ухудшить материнскую иммунную толерантность, что приводит к гиперактивному иммунному ответу, характерному для преэклампсии. [ 68 ]
Рак
[ редактировать ]
CD4 + Регуляторные Т -клетки часто ассоциируются с солидными опухолями как у людей, так и на мышиных моделях. Повышенное количество регуляторных Т -клеток при раке молочной железы, колоректального и яичников связано с более низким прогнозом. [ 69 ]
CD70 + В-клетки неходжкин-лимфомы индуцируют экспрессию FOXP3 и регуляторную функцию во внутриопухолевом CD4 + CD25 − Т -клетки. [ 70 ]
Большинство опухолей вызывают иммунный ответ у хозяина, который опосредуется опухолевыми антигенами, что отличает опухоль от других нездоровых клеток. Это заставляет большое количество инфильтрирующих опухоль лимфоцитов (TIL) появляться в TME. [ 71 ] Эти лимфоциты могут нацелиться на раковые клетки и, следовательно, медленное или прекращать развитие опухоли. Тем не менее, этот процесс сложен, поскольку клетки T REG , по -видимому, предпочтительнее переносится в TME. В то время как клетки T REG обычно составляют только около 4% CD4 + Т -клетки, они могут составить до 20–30% CD4 + Население вокруг TME. [ 72 ]
Соотношение t reg к эффекторным Т -клеткам в TME является определяющим фактором успеха иммунного ответа рака. Высокие уровни T -рег -клеток в TME связаны с плохим прогнозом при многих видах рака, [ 73 ] такие как яичники, грудь, почечная и рак поджелудочной железы. [ 72 ] Это указывает на то, что клетки T REG подавляют эффекторные Т -клетки и препятствуют иммунному ответу организма на рак. Тем не менее, в некоторых типах рака противоположность верна, а высокие уровни T -рег -клеток связаны с положительным прогнозом. Эта тенденция наблюдается при раке, таких как колоректальная карцинома и фолликулярная лимфома . Это может быть связано со способностью к клеткам T -рег подавлять общее воспаление, которое, как известно, запускает пролиферацию клеток и метастазирование. [ 72 ] Эти противоположные эффекты указывают на то, что роль T R -клеток в развитии рака сильно зависит как от типа, так и от местоположения опухоли.
Хотя до сих пор не совсем понимается, как T -reg -клетки преимущественно перемещаются в TME, химиотаксис, вероятно, обусловлена продукцией хемокинов опухолью. Инфильтрация t в TMEI , облегчаемая связыванием CCR4 CCR4 хемокинового рецептора, который экспрессируется на клетках T -рег , с его лигандом CCL22, который секретируется многими типами опухолевых клеток. [ 74 ] TG Расширение клеток в месте опухоли может также объяснить повышенные уровни T -рег -клеток. Известно, что цитокин, TGF-β, который обычно продуцируется опухолевыми клетками, индуцирует дифференцировку и расширение клеток T REG . [ 74 ]
Белок 3 -го ящика 3 ( FOXP3 ) в качестве фактора транскрипции является важным молекулярным маркером клеток T REG . Полиморфизм FOXP3 (RS3761548) может быть вовлечен в прогрессирование рака желудка посредством влияния функции TG и секреции иммуномодулирующих цитокинов, таких как IL-10 , IL-35 и TGF-β . [ 75 ]
Клетки T REG, присутствующие в TME, могут быть либо индуцированными , либо природными (тимическими) TER , которые развиваются из наивных предшественников. Тем не менее, связанные с опухолью T REG также может происходить из IL-17A + Foxp3 + T res , которые развиваются из клеток Th17. [ 76 ] [ 77 ]
В целом, иммунодепрессия TMEHA в значительной степени способствовало неудачным результатам многих лечения иммунотерапии рака. Истощение клеток T REG в моделях животных показало повышенную эффективность иммунотерапевтических методов лечения, и, следовательно, многие иммунотерапевтические методы лечения в настоящее время включают истощение TER . [ 2 ]
Терапия рака, нацеленная на регуляторные Т -лимфоциты
[ редактировать ]В TME , обильно эффекторные режимы . которые чрезмерно экспрессируют иммуносупрессивные молекулы, такие как CTLA-4 Антитела против CTLA-4 вызывают истощение TER и , таким образом, увеличивают CD8 + Т -клетки эффективны против опухоли. Антитело против CTLA-4 ипилимумаб был одобрен для пациентов с продвинутой меланомой. Молекула иммунной проверки PD-1 ингибирует активацию как обычных Т-клеток, так и T- регулировки , и использование антител против PD-1 может привести к активации и иммуносупрессивной функции TER . Устойчивость к лечению против PD-1-MAB, вероятно, вызвана повышенной T- рег активностью клеток . Быстрое прогрессирование рака на блокаде PD-1 называется гиперпрогрессивным заболеванием. Терапия, нацеленная на подавление T , включают анти-CD25 MAB и MAB против CCR4. Агонисты OX40 и агонисты GITR в настоящее время исследуются. [ 76 ] [ 78 ] Терапия, нацеленная на передачу сигналов TCR, также возможна путем блокировки тирозинкиназ. Например, ингибитор тирозин-киназы дасатиниб используется для лечения хронической миелоидной лейкозы и связан с ингибированием t reg . [ 79 ]
Молекулярная характеристика
[ редактировать ]Подобно другим Т -клеткам, регуляторные Т -клетки развиваются в тимусе . Последнее исследование показывает, что регуляторные Т -клетки определяются экспрессией транскрипционного фактора семейства FOXP3 FOXP3 (FOKHEAD P3). Экспрессия FOXP3 необходима для регуляторного развития Т -клеток и, по -видимому, контролирует генетическую программу, указывающую судьбу этой клетки. [ 80 ] Подавляющее большинство регуляторных Т-клеток, экспрессирующих FOXP3, обнаружено в основном комплексе гистосовместимости (MHC) класса II, ограниченного CD 4-экспрессирующего (CD4 + ) популяция и выражает высокий уровень альфа-цепи рецептора интерлейкина-2 (CD25). В дополнение к Foxp3-экспрессирующему CD4 + CD25 + , по -видимому, также есть незначительная популяция MHC Class I CD8 + FOXP3-экспрессирующие регуляторные Т-клетки. Эти Foxp3-экспрессирующие CD8 + Т-клетки, по-видимому, не являются функциональными у здоровых людей, но индуцируются в аутоиммунных заболеваниях стимуляцией рецептора Т-клеток для подавления иммунных реакций, опосредованных IL-17. [ 81 ] В отличие от обычных Т-клеток, регуляторные Т-клетки не продуцируют IL-2 и, следовательно, являются анергическими на исходном уровне.
В исследованиях используется ряд различных методов для выявления и мониторинга клеток T Reg . Первоначально использовалась высокая экспрессия поверхностных маркеров CD25 и CD4 (CD4 + CD25 + клетки). Это проблематично, поскольку CD25 также экспрессируется на нерегуляторных Т-клетках в условиях иммунной активации, например, во время иммунного ответа на патоген. Как определено экспрессией CD4 и CD25, регуляторные Т -клетки составляют около 5–10% зрелого CD4 + Субпопуляция Т -клеток у мышей и людей, в то время как около 1–2% Т -рег может быть измерено в цельной крови. Дополнительное измерение клеточной экспрессии белка Foxp3 позволило более специфическому анализу клеток T -Reg (CD4 + CD25 + Foxp3 + клетки). Тем не менее, FOXP3 также временно экспрессируется в активированных человеческих эффекторных Т -клетках, что усложняет правильный анализ T REG с использованием CD4, CD25 и FOXP3 в качестве маркеров у людей. Следовательно, комбинация поверхностных маркеров золотого стандарта для определений в пределах неактивированного CD3 + CD4 + Т-клетки представляют собой высокую экспрессию CD25 в сочетании с отсутствующей или низкой экспрессией поверхностного белка CD127 (IL-7RA). Если жизнеспособные ячейки не требуются, то добавление FOXP3 в комбинацию CD25 и CD127 обеспечит дальнейшую строгость. Было описано несколько дополнительных маркеров, например, высокие уровни CTLA-4 (цитотоксические T-лимфоциты, связанные с молекулой-4) и GITR (глюкокортикоидный рецептор TNF) также экспрессируются на регуляторных Т-клетках, однако функциональное значение этой экспрессии остается быть определенным. Существует большой интерес к определению маркеров клеточной поверхности, которые являются уникально и специфически экспрессируются на всех регуляторных Т-клетках, экспрессирующих Foxp3. Однако до настоящего времени такая молекула не была идентифицирована.
Идентификация T -re -sef после активации клеток является сложной задачей, поскольку обычные Т -клетки будут экспрессировать CD25, временно экспрессируют FOXP3 и теряют экспрессию CD127 при активации. Было показано, что TER можно обнаружить с использованием анализа маркеров, вызванного активацией, посредством экспрессии CD39 [ 82 ] В сочетании с коэкспрессией CD25 и OX40 (CD134), которые определяют антиген-специфические клетки после 24-48H стимуляции антигеном. [ 83 ] [ 84 ]
В дополнение к поиску новых белковых маркеров, другой метод для анализа и мониторинга T -рег в литературе был описан -клеток. Этот метод основан на анализе метилирования ДНК . Только в клетках t -reg определенная область в гене Foxp3 (TSDR, T REG , но не в любом другом типе клеток, включая активированные эффекторные Т -клетки, обнаружена T -reg -специфическая децетилированная область), что позволяет контролировать клетки через ПЦР реакция или другие методы анализа на основе ДНК. [ 85 ] Взаимодействие между клетками Th17 и регуляторными Т -клетками важно при многих заболеваниях, таких как респираторные заболевания. [ 86 ]
Недавние данные свидетельствуют о том, что тучные клетки могут быть важными медиаторами t reg -зависимой периферической толерантности. [ 87 ]
Эпитопы
[ редактировать ]Регуляторные эпитопы Т -клеток («Tregitopes») были обнаружены в 2008 году и состояли из линейных последовательностей аминокислот, содержащихся в моноклональных антителах и иммуноглобулина G (IgG). С момента их открытия доказательства указывают на то, что Tregitopes могут иметь решающее значение для активации естественных регуляторных Т -клеток. [ 88 ] [ 89 ] [ 90 ]
Предполагается потенциальное применение регуляторных эпитопов Т -клеток: толеризация к трансплантатам, белковые препараты, терапия крови и диабет I типа , а также снижение иммунного ответа для лечения аллергии . [ 91 ] [ 92 ] [ 93 ] [ 94 ] [ 95 ] [ 96 ] [ 90 ]
Генетический дефицит
[ редактировать ]Генетические мутации в гене, кодирующем Foxp3, были идентифицированы как у людей, так и у мышей на основе наследственного заболевания, вызванного этими мутациями. Это заболевание предоставляет наиболее яркие доказательства того, что регуляторные Т -клетки играют критическую роль в поддержании нормальной иммунной системы. людей с мутациями в Foxp3 возникает тяжелое и быстро смертельное аутоиммунное расстройство, известное как дисрегуляция I Mmune, Ployendocrinopathy , Enteropathy У X -связанный ( IPEX ) синдром. [ 97 ] [ 98 ]
Синдром IPEX характеризуется развитием подавляющего системного аутоиммунитета в первом году жизни, что приводит к обычно наблюдаемой триаде водянистой диареи, экзематозного дерматита и эндокринопатии, чаще всего наблюдается как инсулинозависимый диабет . У большинства людей есть другие аутоиммунные явления, включая Coombs-позитивную гемолитическую анемию, аутоиммунную тромбоцитопению, аутоиммунную нейтропению и трубчатую нефропатию. Большинство пораженных мужчин умирают в течение первого года жизни либо метаболических расстройств, либо сепсиса. Аналогичное заболевание также наблюдается у спонтанной мыши Foxp3-мутанта, известной как «Scurfy».
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Bettelli E, Carrier Y, Gao W, Korn T, Strom TB, Oukka M, et al. (Май 2006 г.). «Взаимные пути развития для генерации патогенных эффектора Th17 и регуляторных Т -клеток». Природа . 441 (7090): 235–238. Bibcode : 2006natur.441..235b . doi : 10.1038/nature04753 . PMID 16648838 . S2CID 4391497 .
- ^ Jump up to: а беременный Curiel TJ (май 2007). «Трегс и переосмысление иммунотерапии рака» . Журнал клинических исследований . 117 (5): 1167–1174. doi : 10.1172/jci31202 . PMC 1857250 . PMID 17476346 .
- ^ Чен В (август 2011 г.). «Трег в иммунотерапии: возможности и проблемы» . Иммунотерапия . 3 (8): 911–914. doi : 10.2217/imt.11.79 . PMID 21843075 .
- ^ Miyara M, Gorochov G, Ehrenstein M, Musset L, Sakaguchi S, Amoura Z (октябрь 2011 г.). «Человеческий Foxp3+ регуляторные Т -клетки при системных аутоиммунных заболеваниях». Обзоры аутоиммунитета . 10 (12): 744–755. doi : 10.1016/j.autrev.2011.05.004 . PMID 21621000 .
- ^ Nosbaum A, Prevel N, Truong HA, Mehta P, Ettinger M, Scharschmidt TC, et al. (Март 2016 г.). «Режущая кромка: регуляторные Т -клетки облегчают кожное заживление ран» . Журнал иммунологии . 196 (5): 2010–2014. doi : 10.4049/jimmunol.1502139 . PMC 4761457 . PMID 26826250 .
- ^ Adeegbe do, Nishikawa H (2013). «Природные и индуцированные Т -регуляторные клетки при раке» . Границы в иммунологии . 4 : 190. doi : 10.3389/fimmu.2013.00190 . PMC 3708155 . PMID 23874336 .
- ^ Curiel TJ (апрель 2008 г.). «Регуляторные Т -клетки и лечение рака» . Текущее мнение в иммунологии . 20 (2): 241–246. doi : 10.1016/j.coi.2008.04.008 . PMC 3319305 . PMID 18508251 .
- ^ Hori S, Nomura T, Sakaguchi S (февраль 2003 г.). «Контроль регуляторного развития Т -клеток с помощью фактора транскрипции Foxp3». Наука . 299 (5609): 1057–1061. Bibcode : 2003sci ... 299.1057H . doi : 10.1126/science.1079490 . PMID 12522256 . S2CID 9697928 .
- ^ Singh B, Schwartz JA, Sandrock C, Bellemore SM, Nikoopour E (ноябрь 2013 г.). «Модуляция аутоиммунных заболеваний с помощью интерлейкина (IL) -17, продуцирующих регуляторные клетки T-Helper (TH17)» . Индийский журнал медицинских исследований . 138 (5): 591–594. PMC 3928692 . PMID 24434314 .
- ^ Шевач Э.М. (2000). «Регуляторные Т -клетки в аутоиммуните*» . Ежегодный обзор иммунологии . 18 : 423–449. doi : 10.1146/annurev.immunol.18.1.423 . PMID 10837065 . S2CID 15160752 .
- ^ Schmetterer KG, Neunkirchner A, Pickl WF (июнь 2012 г.). «Природные регуляторные Т -клетки: маркеры, механизмы и манипуляции» . FASEB Journal . 26 (6): 2253–2276. doi : 10.1096/fj.11-193672 . PMID 22362896 . S2CID 36277557 .
- ^ Сакагучи С. (2004). «Естественно, возникающие CD4+ регуляторные Т-клетки для иммунологической самостоятельности и негативного контроля иммунных реакций». Ежегодный обзор иммунологии . 22 : 531–562. doi : 10.1146/annurev.immunol.21.120601.141122 . PMID 15032588 .
- ^ Ли Мо, Руденский Ай (апрель 2016 г.). «Передача сигналов рецептора Т -клеток в контроле регуляторной дифференцировки и функции Т -клеток» . Природные обзоры. Иммунология . 16 (4): 220–233. doi : 10.1038/nri.2016.26 . PMC 4968889 . PMID 27026074 .
- ^ Jump up to: а беременный в дюймовый Santamaria JC, Borelli A, Irla M (2021-02-11). «Регуляторная гетерогенность Т -клеток в тимусе: воздействие на их функциональную активность» . Границы в иммунологии . 12 : 643153. DOI : 10.3389/fimmu.2021.643153 . PMC 7904894 . PMID 33643324 .
- ^ Owen DL, Sjaastad LE, Farrar MA (октябрь 2019). «Регуляторное развитие Т -клеток в тимусе» . Журнал иммунологии . 203 (8): 2031–2041. doi : 10.4049/jimmunol.1900662 . PMC 6910132 . PMID 31591259 .
- ^ Оуэн Д.Л., Махмуд С.А., Сжаастад Л.Е., Уильямс Дж.Б., Снуер Дж.А., Симеонов Д.Р. и др. (Февраль 2019 г.). «Регуляторные Т -клетки тимуса возникают с помощью двух различных программ развития» . Природа иммунология . 20 (2): 195–205. doi : 10.1038/s41590-018-0289-6 . PMC 6650268 . PMID 30643267 .
- ^ Jump up to: а беременный в дюймовый Tiault N, Darrigues J, Adoue V, Gros M, Binet B, Perals C, et al. (Июнь 2015 г.). «Периферические регуляторные Т -лимфоциты, рециркулирующие в тимус, подавляют развитие их предшественников». Природа иммунология . 16 (6): 628–634. doi : 10.1038/ni.3150 . PMID 25939024 . S2CID 7670443 .
- ^ Jump up to: а беременный Pandiyan P, Zheng L, Ishihara S, Reed J, Lenardo MJ (декабрь 2007 г.). «CD4+ CD25+ FOXP3+ регуляторные Т-клетки индуцируют опосредованный цитокином депривации апоптоз эффекторных CD4+ T-клеток». Природа иммунология . 8 (12): 1353–1362. doi : 10.1038/ni1536 . PMID 17982458 . S2CID 8925488 .
- ^ Cheng G, Yu A, Malek TR (май 2011 г.). «Толерантность к Т-клеткам и многофункциональная роль передачи сигналов IL-2R в Т-регуляторных клетках» . Иммунологические обзоры . 241 (1): 63–76. doi : 10.1111/j.1600-065x.2011.01004.x . PMC 3101713 . PMID 21488890 .
- ^ Kimmig S, Przybylski GK, Schmidt CA, Laurisch K, Möwes B, Radbruch A, Thiel A (март 2002 г.). «Два подмножества наивных T -хелпер -клеток с различным содержанием удара T -клеточного рецептора в периферической крови взрослого человека» . Журнал экспериментальной медицины . 195 (6): 789–794. doi : 10.1084/jem.20011756 . PMC 2193736 . PMID 11901204 .
- ^ Toker A, Engelbert D, Garg G, Polansky JK, Floess S, Miyao T, et al. (Апрель 2013). «Активное деметилирование локуса Foxp3 приводит к генерации стабильных регуляторных Т -клеток в тимусе» . Журнал иммунологии . 190 (7): 3180–3188. doi : 10.4049/jimmunol.1203473 . PMID 23420886 .
- ^ Jump up to: а беременный в дюймовый Nicolhool E, Elpaki Y, Higrich S, Shirter Bauses C, De Logel M, Falk C, et al. (Январь 2021 г.). "Рециркуляция IL-1R2 + тонкой настройки внутритимного развития Treg в воспалительных условиях » Клеточная молекулярная иммунология 18 ( ) DOI : 10.1038 . 182–193 . Трегс : / S41423-019-0352-8 . . и 1 S2CID 210913733 .
- ^ Peters VA, Joesting JJ, Freund GG (август 2013 г.). «Рецептор 2 IL-1 (IL-1R2) и его роль в иммунной регуляции» . Мозг, поведение и иммунитет . 32 : 1–8. doi : 10.1016/j.bbi.2012.11.006 . PMC 3610842 . PMID 23195532 .
- ^ Читать S, Malmström V, Powrie F (июль 2000 г.). «Цитотоксический T-лимфоцит-ассоциированный антиген 4 играет важную роль в функции CD25 (+) регуляторных клеток CD4 (+), которые контролируют воспаление кишечника» . Журнал экспериментальной медицины . 192 (2): 295–302. doi : 10.1084/jem.192.2.295 . PMC 2193261 . PMID 10899916 .
- ^ Collison LW, Workman CJ, Kuo TT, Boyd K, Wang Y, Vignali KM, et al. (Ноябрь 2007 г.). «Ингибирующий цитокин IL-35 способствует регуляторной функции Т-клеток». Природа . 450 (7169): 566–569. Bibcode : 2007natur.450..566c . doi : 10.1038/nature06306 . PMID 18033300 . S2CID 4425281 .
- ^ Annacker O, Asseman C, Read S, Powrie F (июнь 2003 г.). «Интерлейкин-10 в регуляции Т-клеточного колита». Журнал аутоиммунитета . 20 (4): 277–279. doi : 10.1016/s0896-8411 (03) 00045-3 . PMID 12791312 .
- ^ Керли Дж., Баркер Дж., Робинсон Д.С., Ллойд С.М. (декабрь 2005 г.). «Разрешение воспаления дыхательных путей и гиперреактивности после переноса in vivo CD4+ CD25+ регуляторных Т -клеток зависит от межллейукина 10 зависит» . Журнал экспериментальной медицины . 202 (11): 1539–1547. doi : 10.1084/jem.20051166 . PMC 1350743 . PMID 16314435 .
- ^ Gondek DC, Lu LF, Quezada SA, Sakaguchi S, Noelle RJ (февраль 2005 г.). «Режущая кромка: опосредованная контактом подавление CD4+ CD25+ регуляторными клетками включает в себя гранзим B-зависимый, не независимый от перфорин механизм» . Журнал иммунологии . 174 (4): 1783–1786. doi : 10.4049/jimmunol.174.4.1783 . PMID 15699103 .
- ^ Puccetti P, Grohmann U (октябрь 2007 г.). «IDO и регуляторные Т-клетки: роль обратной передачи сигналов и неканонической активации NF-Kappab». Природные обзоры. Иммунология . 7 (10): 817–823. doi : 10.1038/nri2163 . PMID 17767193 . S2CID 5544429 .
- ^ Borsellino G, Kleinewietfeld M, Di Mitri D, Sternjak A, Diamantini A, Giometto R, et al. (Август 2007 г.). «Экспрессия эктонуклеотидазы CD39 с помощью клеток Foxp3+ Treg: гидролиз внеклеточного АТФ и иммуно -подавления» . Кровь . 110 (4): 1225–1232. doi : 10.1182/blood-2006-12-064527 . PMID 17449799 .
- ^ Kobie JJ, Shah PR, Yang L, Rebhahn JA, Fowell DJ, Mosmann TR (ноябрь 2006 г.). «T-регуляторные и приподнятые невозможные CD4 T-клетки экспрессируют CD73, который подавляет эффекторные CD4 T-клетки путем преобразования 5'-аденозин-монофосфата в аденозин» . Журнал иммунологии . 177 (10): 6780–6786. doi : 10.4049/jimmunol.177.10.6780 . PMID 17082591 .
- ^ Huang CT, Workman CJ, Flies D, Pan X, Marson AL, Zhou G, et al. (Октябрь 2004). «Роль LAG-3 в регуляторных Т-клетках» . Иммунитет . 21 (4): 503–513. doi : 10.1016/j.immuni.2004.08.010 . PMID 15485628 .
- ^ Yu X, Harden K, Gonzalez LC, Francesco M, Chiang E, Irving B, et al. (Январь 2009 г.). «Поверхностный белок Tigit подавляет активацию Т -клеток, способствуя генерации зрелых иммунорегуляторных дендритных клеток». Природа иммунология . 10 (1): 48–57. doi : 10.1038/ni.1674 . PMID 19011627 . S2CID 205361984 .
- ^ Wardell CM, Macdonald KN, Levings Mk, Cook L (январь 2021 г.). «Перекрестные разговоры между регуляторными Т-клетками человека и антиген-презентативными клетками: уроки для клинических применений» . Европейский журнал иммунологии . 51 (1): 27–38. doi : 10.1002/eji.202048746 . HDL : 11343/276776 . PMID 33301176 .
- ^ Сакагучи С., Ямагучи Т., Номура Т, Оно М (май 2008 г.). «Регуляторные Т -клетки и иммунная толерантность» . Клетка . 133 (5): 775–787. doi : 10.1016/j.cell.2008.05.009 . PMID 18510923 .
- ^ Уокер Л.С., Сансом Д.М. (ноябрь 2011 г.). «Новая роль CTLA4 как клеточного экстразинного регулятора Т-клеточных ответов». Природные обзоры. Иммунология . 11 (12): 852–863. doi : 10.1038/nri3108 . PMID 22116087 . S2CID 9617595 .
- ^ Jump up to: а беременный в дюймовый Шевирев D, Tereshchenko V (2020). «Трег гетерогенность, функции и гомеостаз» . Границы в иммунологии . 10 : 3100. DOI : 10.3389/fimmu.2019.03100 . PMC 6971100 . PMID 31993063 .
- ^ Харибхай Д., Уильямс Дж. Б., Цзя С., Никерсон Д., Шмитт Э.Г., Эдвардс Б. и др. (Июль 2011). «Необходимая роль индуцированных регуляторных Т -клеток в толерантности на основе расширения разнообразия антиген -рецепторов» . Иммунитет . 35 (1): 109–122. doi : 10.1016/j.immuni.2011.03.029 . PMC 3295638 . PMID 21723159 .
- ^ Jump up to: а беременный Sun CM, Hall JA, Blank RB, Bouladoux N, Oukka M, Mora Jr, Belkaid Y (август 2007 г.). «Дендритные клетки с малой кишкой, способствующие генерации клеток Foxp3 T REG, способствуют генерации DE novo Foxp3 через ретиноевую кислоту» . Журнал экспериментальной медицины . 204 (8): 1775–1785. doi : 10.1084/jem.20070602 . PMC 2118682 . PMID 17620362 .
- ^ Mucida D, Park Y, Kim G, Turovskaya O, Scott I, Kronenberg M , Cheroutre H (июль 2007 г.). «Взаимная дифференцировка Th17 и регуляторные Т -клеток, опосредованная ретиноевой кислотой» . Наука . 317 (5835): 256–260. Bibcode : 2007sci ... 317..256m . doi : 10.1126/science.1145697 . PMID 17569825 . S2CID 24736012 .
- ^ Эркеленс М.Н., Мебиус Р.Е. (март 2017 г.). «Ретиноевая кислота и иммунный гомеостаз: баланс». Тенденции в иммунологии . 38 (3): 168–180. doi : 10.1016/j.it.2016.12.006 . PMID 28094101 .
- ^ Ziegler SF, Buckner JH (апрель 2009 г.). «FOXP3 и регулирование дифференциации Treg/TH17» . Микробы и инфекция . 11 (5): 594–598. doi : 10.1016/j.micinf.2009.04.002 . PMC 2728495 . PMID 19371792 .
- ^ GA Power, Nova-Lamper E, Scott C, Failer G, Chen YC, Becker PD, et al. (Demember 2018). "CD161, регулируемый кислотой CD161 + Регуляторные Т-клетки поддерживают восстановление раны в слизистой оболочке кишечника » . Природа иммунология . 19 (12): 1403–1414. : 10.1038 /S41590-018-0230-Z . PMC 6474659. . PMID 30397350 DOI
- ^ Coombes JL, Siddiqui KR, Arancibia-Cárcamo CV, Hall J, Sun CM, Belkaid Y, Powrie F (август 2007 г.). «Функционально специализированная популяция CD103+ DCS слизистой оболочки индуцирует регуляторные Т-клетки FOXP3+ через TGF-бета-бета и ретиноевую кислоту-зависимую механизм» . Журнал экспериментальной медицины . 204 (8): 1757–1764. doi : 10.1084/jem.20070590 . PMC 2118683 . PMID 17620361 .
- ^ Jump up to: а беременный Cook L, Reid KT, Häkkinen E, De Bie B, Tanaka S, Smyth DJ, et al. (Сентябрь 2021 г.). "Индукция стабильного человека Foxp3 + Tregs с помощью паразитов TGF-β-мимика » . Иммунология и клеточная биология . 99 (8): 833–847. : 10.1111 /IMCB.12475 . PMC 8453874. . PMID 33929751 DOI
- ^ Johnston CJ, Smyth DJ, Kodali RB, White MP, Harcus Y, Filbey KJ, et al. (Ноябрь 2017). «Структурно отличающийся TGF-β имитирует из кишечного паразита гельминта, который мощно индуцирует регуляторные Т-клетки» . Природная связь . 8 (1): 1741. Bibcode : 2017natco ... 8.1741j . doi : 10.1038/s41467-017-01886-6 . PMC 5701006 . PMID 29170498 .
- ^ Smyth DJ, Harcus Y, White MP, Gregory WF, Nahler J, Stephens I, et al. (Апрель 2018). «TGF-β имитирующие белки образуют расширенное семейство генов в мышином паразитовом гелигмосомоиде Polygyrus» . Международный журнал по паразитологии . 48 (5): 379–385. doi : 10.1016/j.ijpara.2017.12.004 . PMC 5904571 . PMID 29510118 .
- ^ White MP, Smyth DJ, Cook L, Ziegler SF, Levings MK, Maizels RM (сентябрь 2021 г.). "Паразит цитокин имитирует HP-TGM мощно повторяет регуляторное влияние TGF-β на мышиный CD4 + Т -клетки » . Иммунология и клеточная биология . 99 (8): 848–864. : 10.1111 /IMCB.12479 . PMC 9214624. . PMID 33988885 DOI
- ^ Ohnmacht C, Park JH, Cording S, Wing JB, Atarashi K, Obata Y, et al. (Август 2015). «Иммунология слизистой оболочки. Микробиота регулирует иммунитет 2 типа через RORγt⁺ T -клетки». Наука . 349 (6251): 989–993. Bibcode : 2015sci ... 349..989o . doi : 10.1126/science.aac4263 . PMID 26160380 . S2CID 2663636 .
- ^ Jump up to: а беременный в Ning X, Lei Z, Rui B, Li Y, Li M (2022-12-05). «Микробиота кишечника способствует иммунной толерантности, регулируя клетки RORγT+ Treg в пищевой аллергии» . Advanced Gut & Microbiome Research . 2022 : E8529578. doi : 10.1155/2022/8529578 .
- ^ Jump up to: а беременный Ohnmacht C, Park JH, Cording S, Wing JB, Atarashi K, Obata Y, et al. (Август 2015). «Иммунология слизистой оболочки. Микробиота регулирует иммунитет 2 типа через RORγt⁺ T -клетки». Наука . 349 (6251): 989–993. Bibcode : 2015sci ... 349..989o . doi : 10.1126/science.aac4263 . PMID 26160380 . S2CID 2663636 .
- ^ Al Nabhani Z, Dulauroy S, Marques R, Cousu C, Al Bounny S, Déjardin F, et al. (Май 2019). «Реакция отлучения от микробиоты необходима для устойчивости к иммунопатологии у взрослого». Иммунитет . 50 (5): 1276–1288.e5. doi : 10.1016/j.immuni.2019.02.014 . PMID 30902637 .
- ^ Раманан Д., Сефик Е., Гальван-Пена С., Ву М., Ян Л., Ян З. и др. (Июнь 2020 г.). «Иммунологический способ многопланоценного передачи управляет кишечной установкой Treg» . Клетка . 181 (6): 1276–1290.e13. doi : 10.1016/j.cell.2020.04.030 . PMC 7393667 . PMID 32402238 .
- ^ Абдель-Гадир А., Стивен-Виктор Э., Гербер Г.К., Новал Ривас М., Ван С., Харб Х. и др. (Июль 2019). «Микробиота терапия действует через регуляторную Т -клетку MyD88/RORγT для подавления пищевой аллергии» . Природная медицина . 25 (7): 1164–1174. doi : 10.1038/s41591-019-0461-z . PMC 6677395 . PMID 31235962 .
- ^ Ранкин Л.К., Кайзер ты, Парк Х, Улеманн А.С., Грей Д.Х., Арпайя. "Это + Клетки Treg за счет GATA3 + Клетки Treg и изменяют комменсальный метаболизм микробиоты » . Клеточные отчеты . 42 (3): 112135. DOI : 10.1016/ . PMC 10150404. . PMID 36840944 j.celrep.2023.112135
- ^ Бриттон Г.Дж., Могно I, Веннаро О, ОК. (Январь 2019). "Микробиоты от людей с воспалительным заболеванием кишечника изменяют баланс кишечника Th17 и RORγt + Регуляторные Т -клетки и обостряющий колит у мышей » . Иммунитет . 50 (1): 212–224.E4. : 10.1016 /j.immuni.2018.12.015 . PMC 6512335. . PMID 306503777 DOI
- ^ Hovhannisyan Z, Treatman J, Littman Dr, Mayer L (март 2011 г.). «Характеристика регуляторных Т-клеток, продуцирующих интерлейкин-17, в воспаленной слизистой оболочке кишечника у пациентов с воспалительными заболеваниями кишечника» . Гастроэнтерология . 140 (3): 957–965. doi : 10.1053/j.gastro.2010.12.002 . PMC 3049831 . PMID 21147109 .
- ^ Jump up to: а беременный Blatner NR, Mulcahy MF, Dennis KL, Scholtens D, Bentrem DJ, Phillips JD, et al. (Декабрь 2012 г.). «Экспрессия RORγT отмечает патогенный подмножество регуляторных Т -клеток при раке толстой кишки человека» . Научная трансляционная медицина . 4 (164): 164ra159. doi : 10.1126/scitranslmed.3004566 . PMC 3762575 . PMID 23241743 .
- ^ Rizzo A, Di Giovangiulio M, Stolfi C, Franzè E, Fehling HJ, Carsetti R, et al. (Сентябрь 2018 г.). «RORγT-экспрессирующие Tregs способствуют росту, связанного с колитом колоректального рака путем контроля IL6 в дендритных клетках». Раковая иммунология исследования . 6 (9): 1082–1092. doi : 10.1158/2326-6066.cir-17-0698 . PMID 29991500 .
- ^ Jacobse J, Li J, Rings EH, Samsom JN, Goettel JA (2021). «Кишечные регуляторные Т-клетки как специализированные иммунные клетки с ограниченными тканью в кишечном иммунном гомеостазе и заболеваниях» . Границы в иммунологии . 12 : 716499. DOI : 10.3389/fimmu.2021.716499 . PMC 8371910 . PMID 34421921 .
- ^ Луи П.П., Чо I, Али Н. (сентябрь 2020 г.). «Тканевые регуляторные Т -клетки» . Иммунология . 161 (1): 4–17. doi : 10.1111/imm.13208 . PMC 7450170 . PMID 32463116 .
- ^ Stringari LL, Covre LP, Da Silva FD, De Oliveira VL, Campana MC, Hadad DJ, et al. (Июль 2021 г.). «Увеличение CD4+ CD25HIGHFOXP3+ клеток ухудшает микробицидную активность человека in vitro против туберкулеза Mycobacterium во время скрытого и острого легочного туберкулеза» . ПЛО не пренебрегали тропическими заболеваниями . 15 (7): E0009605. doi : 10.1371/journal.pntd.0009605 . PMC 8321116 . PMID 34324509 .
- ^ Kleinman AJ, Sivanandham R, Pandrea I, Chougnet CA, Apetrei C (2018). «Регуляторные Т -клетки в качестве потенциальных мишеней для исследования лечения ВИЧ» . Границы в иммунологии . 9 : 734. DOI : 10.3389/fimmu.2018.00734 . PMC 5908895 . PMID 29706961 .
- ^ Svanandham R, Kleinman AJ, Sette P, Brocca-Cofano E, Kilapandal Venkatraman SM, Policiccchio BB, et al. (Сентябрь 2020 г.). Несооооооооооооом -тестирование влияния разницы в стратегиях регламентирования разности « Журнал вирусологии 94 (19): JVI.00533–20, JVI, JVI, JVI.00533–20V1 Doi : 10.1128/ jvi.00533-2 7495362PMC 32669326PMID S2CID 220579402
- ^ Beers DR, Zhao W, Wang J, Zhang X, Wen S, Neal D, et al. (Март 2017). «Регуляторные Т -лимфоциты пациентов с БАС являются дисфункциональными и коррелируют с скоростью прогрессирования заболевания и тяжести» . JCI Insight . 2 (5): E89530. doi : 10.1172/jci.insight.89530 . PMC 5333967 . PMID 28289705 .
- ^ Thonhoff JR, Beers DR, Zhao W, Pleitez M, Simpson EP, Berry JD, et al. (Июль 2018). «Расширенные аутологичные регуляторные инфузии Т-лимфоцитов в ALS: фаза I, исследование первого человека» . Неврология . 5 (4): E465. doi : 10.1212/nxi.0000000000000465 . PMC 5961523 . PMID 29845093 .
- ^ Цуда С., Накашима А., Шима Т., Сайто С. (2019). «Новая парадигма в роли регуляторных Т -клеток во время беременности» . Границы в иммунологии . 10 : 573. DOI : 10.3389/fimmu.2019.00573 . PMC 6443934 . PMID 30972068 .
- ^ Hu M, Eviston D, Hsu P, Mariño E, Chidgey A, Santner-Nanan B, et al. (Июль 2019). «Снижение ацетата материнской сыворотки и нарушение плода тимуса и регуляторных Т -клеток при преэклампсии» . Природная связь . 10 (1): 3031. Bibcode : 2019natco..10.3031H . doi : 10.1038/s41467-019-10703-1 . PMC 6620275 . PMID 31292453 .
- ^ Дранофф Г (декабрь 2005 г.). «Терапевтические последствия внутриопухолевых регуляторных Т -клеток». Клиническое исследование рака . 11 (23): 8226–8229. doi : 10.1158/1078-0432.ccr-05-2035 . PMID 16322278 . S2CID 18794337 .
- ^ Yang ZZ, Novak AJ, Ziesmer SC, Witzig TE, Ansell SM (октябрь 2007 г.). «B-клетки CD70+ неходжкин-лимфомы индуцируют экспрессию FOXP3 и регуляторную функцию во внутриопухолевых Т-клетках CD4+ CD25» . Кровь . 110 (7): 2537–2544. doi : 10.1182/blood-2007-03-082578 . PMC 1988926 . PMID 17615291 .
- ^ Gooden MJ, De Bock GH, Leffers N, Daemen T, Nijman HW (июнь 2011 г.). «Прогностическое влияние инфильтрирующих опухолевых лимфоцитов при раке: систематический обзор с мета-анализом» . Британский журнал рака . 105 (1): 93–103. doi : 10.1038/bjc.2011.189 . PMC 3137407 . PMID 21629244 .
- ^ Jump up to: а беременный в Олеиника К., Ниббс Р.Дж., Грэм Г.Дж., Фрейзер А.Р. (январь 2013 г.). «Подавление, подрыв и побег: роль регуляторных Т -клеток в прогрессировании рака» . Клиническая и экспериментальная иммунология . 171 (1): 36–45. doi : 10.1111/j.1365-2249.2012.04657.x . PMC 3530093 . PMID 23199321 .
- ^ Plitas G, Rudensky AY (2020-03-09). «Регуляторные Т -клетки при раке» . Ежегодный обзор биологии рака . 4 (1): 459–477. doi : 10.1146/annurev-cancerbio-030419-033428 . ISSN 2472-3428 .
- ^ Jump up to: а беременный Липпиц будет (май 2013). «Модели цитокинов у пациентов с раком: систематический обзор». Lancet. Онкология . 14 (6): E218 - E228. doi : 10.1016/s1470-2045 (12) 70582-x . PMID 23639322 .
- ^ Ezzeddini R, Somi MH, Taghihhani M, Moaddab SY, Masnadi Shirazi K, Shirmohammadi M, et al. (Февраль 2021 г.). «Ассоциация полиморфизма FOXP3 RS3761548 с концентрацией цитокинов у пациентов с аденокарциномой желудка». Цитокин . 138 : 155351. DOI : 10.1016/j.cyto.2020.155351 . PMID 33127257 . S2CID 226218796 .
- ^ Jump up to: а беременный Li C, Jiang P, Wei S, Xu X, Wang J (июль 2020 г.). «Регуляторные Т -клетки в микроокружении опухоли: новые механизмы, потенциальные терапевтические стратегии и будущие перспективы» . Молекулярный рак . 19 (1): 116. DOI : 10.1186/S12943-020-01234-1 . PMC 7367382 . PMID 32680511 .
- ^ Downs-Canner S, Berkey S, Delgoffe GM, Edwards RP, Curiel T, Odunsi K, et al. (Март 2017). "Подавляющий IL-17A + Foxp3 + и бывший 17 IL-17A отрицательный Foxp3 + Клетки T REG являются источником, ассоциированными с опухолью T- рег « , Природная связь . 8 (1): 14649. BIBCODE : ... 814649d . DOI : 10.1038/ncomms14649 . PMC 5355894. 2017Natco PMID 28290453 .
- ^ Togashi Y, Sithara K, Nishikawa H (июнь 2019 г.). «Регуляторные Т -клетки в иммуносупрессии рака - последствия для противоопухолевой терапии». Природные обзоры. Клиническая онкология . 16 (6): 356–371. doi : 10.1038/s41571-019-0175-7 . PMID 30705439 . S2CID 59526013 .
- ^ Оуу, Нишикава Х (июль 2019 г.). «Регуляторные клетки T (Treg) при раке: могут ли клетки Treg быть новой терапевтической мишенью?» Полем Наука о раке . 110 (7): 2080–2089. doi : 10.1111/cas.14069 . PMC 6609813 . PMID 31102428 .
- ^ Marson A, Kretchmer K, Frampton GM, Jacobsen ES, Polansky JK, Macisaac KD, et al. (Февраль 2007 г.). «Занятость FOXP3 и регуляция ключевых генов-мишеней во время стимуляции Т-клеток» . Природа . 445 (7130): 931–935. Bibcode : 2007natur.445..931m . doi : 10.1038/nature05478 . PMC 3008159 . PMID 17237765 .
- ^ Эллис С.Д., Макговерн Дж.Л., Ван Маурик А., Хоу Д., Эренштейн М., Нотли Калифорния (октябрь 2014 г.). «Индуцированные клетки CD8+ FOXP3+ TREG при ревматоидном артрите модулируются с помощью фосфорилирования p38 и моноцитов, экспрессирующих мембранную фактор некроза опухоли α и CD86». Артрит и ревматология . 66 (10): 2694–2705. doi : 10.1002/art.38761 . PMID 24980778 . S2CID 39984435 .
- ^ Seddiki N, Cook L, HSU DC, Phetsouphanh C, Brown K, Xu Y, et al. (Июнь 2014 г.). «Человеческий антиген-специфический CD4⁺ CD25⁺ CD134⁺ CD39⁺ Т-клетки обогащаются для регуляторных Т-клеток и составляют значительную долю ответов на воспоминание». Европейский журнал иммунологии . 44 (6): 1644–1661. doi : 10.1002/eji.201344102 . PMID 24752698 . S2CID 24012204 .
- ^ Zaunders JJ, Munier ML, Seddiki N, Pett S, IP S, Bailey M, et al. (Август 2009 г.). «Высокие уровни антиген-специфических CD4+ Т-клеток в периферической крови, выявленная стимулированной коэкспрессией CD25 и CD134 (OX40)» . Журнал иммунологии . 183 (4): 2827–2836. doi : 10.4049/jimmunol.0803548 . PMID 19635903 .
- ^ Poloni C, Schonhofer C, Ivison S, Levings MK, Steiner TS, Cook L (июль 2023 г.). «Т-клеточные маркерные анализы, вызванные активацией активации, в отношении здоровья и заболеваний» . Иммунология и клеточная биология . 101 (6): 491–503. doi : 10.1111/imcb.12636 . PMC 10952637 . PMID 36825901 . S2CID 257152898 .
- ^ Wieczorek G, Asemissen A, Model F, Turbachova I, Floess S, Liebenberg V, et al. (Январь 2009 г.). «Количественный анализ метилирования ДНК FOXP3 как новый метод подсчета регуляторных Т -клеток в периферической крови и твердой ткани». РАНКА . 69 (2): 599–608. doi : 10.1158/0008-5472.can-08-2361 . PMID 19147574 .
- ^ Агарвал А., Сингх М., Чаттерджи Б.П., Чаухан А., Чакраборти А. (2014). «Взаимодействие T -Helper 17 клеток с CD4 (+) CD25 (высоким) Foxp3 (+) Tregs в регуляции аллергической астмы у педиатрических пациентов» . Международный журнал педиатрии . 2014 : 636238. DOI : 10.1155/2014/636238 . PMC 4065696 . PMID 24995020 .
- ^ Лу Л.Ф., Линд Э.Ф., Гондек Д.К., Беннетт К.А., Глисон М.В., Пино-Лагос К. и др. (Август 2006 г.). «Тучные клетки являются важными посредниками в регуляторной толерантности к Т-клеткам». Природа . 442 (7106): 997–1002. Bibcode : 2006natur.442..997L . doi : 10.1038/nature05010 . PMID 16921386 . S2CID 686654 .
- ^ «Трегитоп: иммуномодуляционный электроинструмент» . Эпивакс . 2 августа 2016 года.
- ^ Hui DJ, Basner-tschakarjan E, Chen Y, Davidson RJ, Buchlis G, Yazicioglu M, et al. (Сентябрь 2013 г.). «Модуляция ответов CD8+ T-клеток на векторы AAV с эпитопами MHC Class II, полученным из IgG» . Молекулярная терапия . 21 (9): 1727–1737. doi : 10.1038/mt.2013.166 . PMC 3776637 . PMID 23857231 .
- ^ Jump up to: а беременный De Groot AS, Moise L, McMurry JA, Wambre E, Van Oupvelt L, Moingeon P, et al. (Октябрь 2008 г.). «Активация природных регуляторных Т-клеток IgG FC-полученным пептидом« Tregitopes » » . Кровь . 112 (8): 3303–3311. doi : 10.1182/blood-2008-02-138073 . PMC 2569179 . PMID 18660382 .
- ^ «Новая инфузия фондов NIH в размере 2,25 млн. Долл. США для Epivax 'Tregitope, предложенная« обработка парадигмы » . Жесткие биотехнологические исследования.
- ^ Su Y, Rossi R, De Groot AS, Scott DW (август 2013 г.). «Регуляторные Эпитопы Т -клеток (Tregitopes) в IgG вызывают толерантность in vivo и не имеют иммуногенности как таковой» . Журнал биологии лейкоцитов . 94 (2): 377–383. doi : 10.1189/jlb.0912441 . PMC 3714563 . PMID 23729499 .
- ^ Cousens LP, Su Y, McClaine E, Li X, Terry F, Smith R, et al. (2013). «Применение IgG-полученных природных Treg Epitopes (IgG Tregitopes) к индукции антиген-специфической толерантности в мышиной модели диабета 1 типа» . Журнал исследований диабета . 2013 : 621693. DOI : 10.1155/2013/621693 . PMC 3655598 . PMID 23710469 .
- ^ Cousens LP, Mingozzi F, Van der Marel S, Su Y, Garman R, Ferreira V, et al. (Октябрь 2012 г.). «Обучение толерантности: новые подходы к заместительной ферментативной терапии при болезни Помпа» . Вакцины и иммунотерапевтические средства . 8 (10): 1459–1464. doi : 10.4161/hv.21405 . PMC 3660767 . PMID 23095864 .
- ^ Cousens LP, Najafian N, Mingozzi F, Elyaman W, Mazer B, Moise L, et al. (Январь 2013). «Исследования in vitro и in vivo эпитопов Treg, полученных из IgG (Tregitopes): многообещающий новый инструмент для индукции толерантности и лечения аутоиммунитета» . Журнал клинической иммунологии . 33 (Suppl 1): S43 - S49. doi : 10.1007/s10875-012-9762-4 . PMC 3538121 . PMID 22941509 .
- ^ Elyaman W, Khory SJ, Scott DW, De Groot As (2011). «Потенциальное применение трегитопов в качестве иммуномодулирующих агентов при рассеянном склерозе» . Неврологии исследований International . 2011 : 256460. DOI : 10.1155/2011/256460 . PMC 3175387 . PMID 21941651 .
- ^ Онлайн -наследство Менделян в человеке ipex
- ^ Ipex at nih / uw genetsts
Внешние ссылки
[ редактировать ]- Регуляторные+Т-клетки в Национальной библиотеке медицинских предметов США (Mesh)