Мозжечок
| Мозжечок | |
|---|---|
 Положение мозжечка человека (сагиттальная проекция) | |
 Анимация мозжечка | |
| Подробности | |
| Произношение | / ˌ s ɛ r ə ˈ b ɛ l ə m / |
| Часть | Задний мозг |
| Артерия | СКА , АИКА , ПИКА |
| вена | Высший , низший |
| Идентификаторы | |
| латинский | мозжечок |
| МеШ | D002531 |
| Нейроимена | 643 |
| НейроЛекс ID | бирнлекс_1489 |
| ТА98 | A14.1.07.001 |
| ТА2 | 5788 |
| ФМА | 67944 |
| Анатомические термины нейроанатомии | |
Мозжечок по латыни ( мн. мозжечок или мозжечок ; «маленький мозг») является основной особенностью заднего мозга всех позвоночных . Хотя обычно он меньше головного мозга , у некоторых животных, таких как мормириды, он может быть такого же размера или даже больше. [1] У людей мозжечок играет важную роль в двигательном контроле и когнитивных функциях, таких как внимание и речь , а также в эмоциональном контроле , например, в регулировании реакций страха и удовольствия. [2] [3] [4] но его функции, связанные с движением, наиболее прочно установлены. Человеческий мозжечок не инициирует движение, но способствует координации , точности и точному расчету времени: он получает входные данные от сенсорных систем спинного мозга и других частей головного мозга и интегрирует эти входные данные для точной настройки двигательной активности. [5] Повреждение мозжечка приводит к нарушениям тонких движений , равновесия , позы и двигательного обучения у людей. [5]
Анатомически мозжечок человека имеет вид отдельной структуры, прикреплённой к нижней части мозга, спрятанной под полушариями головного мозга . Его корковая поверхность покрыта мелко расположенными параллельными бороздками, резко контрастирующими с широкими неправильными извилинами коры головного мозга . Эти параллельные бороздки скрывают тот факт, что кора мозжечка на самом деле представляет собой непрерывный тонкий слой ткани, плотно сложенный в виде гармошки . Внутри этого тонкого слоя находится несколько типов нейронов с очень регулярным расположением, наиболее важными из которых являются клетки Пуркинье и гранулярные клетки . Эта сложная нервная организация обеспечивает огромные возможности обработки сигналов, но почти весь выходной сигнал коры мозжечка проходит через набор небольших глубоких ядер, расположенных внутри белого вещества мозжечка. [6]
Помимо своей прямой роли в контроле движений, мозжечок необходим для нескольких типов двигательного обучения , в первую очередь для обучения адаптации к изменениям в сенсомоторных отношениях . Было разработано несколько теоретических моделей, объясняющих сенсомоторную калибровку с точки зрения синаптической пластичности в мозжечке. Эти модели основаны на наблюдениях Дэвида Марра и Джеймса Альбуса , основанных на наблюдении того, что каждая клетка Пуркинье мозжечка получает два совершенно разных типа входных сигналов: один включает тысячи слабых входных сигналов от параллельных волокон гранулярных клеток; другой — чрезвычайно сильный входной сигнал от одного карабкающегося волокна . [7] Основная концепция теории Марра-Альбуса заключается в том, что поднимающееся волокно служит «обучающим сигналом», который вызывает долговременное изменение силы входных параллельных волокон. Наблюдения за длительной депрессией в параллельных входных волокнах предоставили некоторую поддержку теориям этого типа, но их достоверность остается спорной. [8]
Структура
[ редактировать ]На уровне общей анатомии мозжечок состоит из плотно сложенного слоя коры с белым веществом под ним и заполненного жидкостью желудочка у основания. Четыре глубоких ядра мозжечка погружены в белое вещество. Каждая часть коры состоит из одного и того же небольшого набора нейронных элементов, расположенных в весьма стереотипной геометрии. На промежуточном уровне мозжечок и его вспомогательные структуры могут быть разделены на несколько сотен или тысяч независимо функционирующих модулей, называемых «микрозонами» или «микрокомпартментами».
Общая анатомия
[ редактировать ]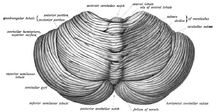
Мозжечок расположен в задней черепной ямке . Четвертый желудочек , мост и продолговатый мозг находятся впереди мозжечка. [9] Он отделен от вышележащего головного мозга слоем кожистой твердой мозговой оболочки , наметом мозжечка ; все его связи с другими частями мозга проходят через мост. Анатомы относят мозжечок к части промежуточного мозга , в который входит также мост; метэнцефалон - это верхняя часть ромбэнцефалона или «заднего мозга». Как и кора головного мозга, мозжечок разделен на два полушария мозжечка ; он также содержит узкую срединную зону ( червь ). Набор крупных складок традиционно используется для разделения общей структуры на 10 меньших «долек». Из-за большого количества крошечных гранулярных клеток мозжечок содержит больше нейронов , чем весь остальной мозг, но занимает лишь 10% от общего объема мозга. [10] Количество нейронов в мозжечке связано с количеством нейронов в неокортексе . В мозжечке примерно в 3,6 раза больше нейронов, чем в неокортексе, и это соотношение сохраняется у многих различных видов млекопитающих. [11]
Необычный внешний вид мозжечка скрывает тот факт, что большая часть его объема состоит из очень плотно сложенного слоя серого вещества : коры мозжечка . Каждый гребень или извилина в этом слое называется листком . МРТ высокого разрешения показывает, что кора мозжечка взрослого человека имеет площадь 730 квадратных см. [12] упакованы в объем размером 6 см × 5 см × 10 см. [10] Под серым веществом коры находится белое вещество , состоящее в основном из миелинизированных нервных волокон, идущих к коре и от нее. В белом веществе, которое иногда называют древом жизни (деревом жизни) из-за его разветвленного древовидного вида на поперечном сечении, находятся четыре глубоких ядра мозжечка , состоящие из серого вещества. [13]
Мозжечок с разными отделами нервной системы соединяют три парные ножки мозжечка . Это верхняя ножка мозжечка , средняя ножка мозжечка и нижняя ножка мозжечка , названные по их положению относительно червя. Верхняя ножка мозжечка в основном является выходом в кору головного мозга, неся эфферентные волокна через ядра таламуса к верхним мотонейронам коры головного мозга. Волокна берут начало из глубоких ядер мозжечка. Средняя ножка мозжечка соединена с мостом и получает всю входную информацию от моста, главным образом, от ядер моста . Входной сигнал в мост поступает из коры головного мозга и передается от ядер моста через поперечные волокна моста в мозжечок. Средняя ножка — самая крупная из трех, ее афферентные волокна сгруппированы в три отдельных пучка, подающих входы к различным частям мозжечка. Нижняя ножка мозжечка получает сигналы от афферентных волокон вестибулярных ядер, спинного мозга и покрышки. Выход из нижней ножки осуществляется по эфферентным волокнам к вестибулярным ядрам и ретикулярной формации. Весь мозжечок получает модуляторную сигнализацию от нижнего оливкового ядра через нижнюю ножку мозжечка. [6]
Подразделения
[ редактировать ]
По внешнему виду в мозжечке можно выделить три доли: переднюю долю (над первичной щелью ), заднюю долю (ниже первичной щели) и флокулонодулярную долю (ниже задней щели). Эти доли делят мозжечок от рострального до каудального (у человека сверху вниз). Однако с точки зрения функции существует более важное различие в медиальном и латеральном измерении. Оставляя в стороне флокулонодулярную долю, которая имеет различные связи и функции, мозжечок можно функционально разделить на медиальный сектор, называемый спиноцеребеллум , и более крупный латеральный сектор, называемый мозжечок . [13] Узкая полоска выступающей ткани вдоль средней линии называется червем мозжечка . ( Vermis в переводе с латыни означает «червь».) [13]
Наименьшую область, флокулонодулярную долю, часто называют вестибулоцеребеллумом . Это самая старая часть в эволюционном отношении (archicerebellum) и участвует главным образом в балансе и пространственной ориентации; его основные связи связаны с вестибулярными ядрами , хотя он также получает зрительную и другую сенсорную информацию. Повреждение этой области вызывает нарушения равновесия и походки . [13]
Медиальная зона передней и задней долей составляет спиноцеребеллум, также известный как палеоцеребеллум. Этот сектор мозжечка в основном отвечает за точную настройку движений тела и конечностей. Он получает проприоцептивную информацию от дорсальных столбов спинного мозга (включая спинно-мозжечковый тракт ) и от черепно -мозгового нерва , а также от зрительной и слуховой систем. [14] Он посылает волокна к глубоким ядрам мозжечка, которые, в свою очередь, проецируются как в кору головного мозга, так и в ствол мозга, обеспечивая тем самым модуляцию нисходящих двигательных систем. [13]
Латеральная зона, которая у человека составляет большую часть, представляет собой мозжечок, также известный как неоцеребеллум. Он получает входные данные исключительно от коры головного мозга (особенно теменной доли ) через ядра моста (формирующие кортико-понто-мозжечковые пути) и отправляет выходные данные главным образом в вентролатеральный таламус (в свою очередь связанный с двигательными областями премоторной коры и первичными двигательными путями). область коры головного мозга) и к красному ядру . [13] Существуют разногласия по поводу того, как лучше всего описать функции латерального мозжечка: считается, что он участвует в планировании предстоящих движений. [15] при оценке сенсорной информации для действий, [13] и в ряде чисто когнитивных функций, таких как определение глагола, который лучше всего соответствует определенному существительному (например, «сидеть» вместо «стул»). [16] [17] [18] [19]
Микроанатомия
[ редактировать ]В мозжечковом контуре доминирующую роль играют два типа нейронов: клетки Пуркинье и гранулярные клетки . Доминирующую роль также играют три типа аксонов : мшистые волокна и лазящие волокна (входящие в мозжечок снаружи) и параллельные волокна (являющиеся аксонами клеток-зерен). В мозжечковом контуре есть два основных пути: берущие начало от мшистых волокон и лазающих волокон, оба в конечном итоге заканчиваются в глубоких ядрах мозжечка. [10]
Мшистые волокна проецируются непосредственно в глубокие ядра, но также дают начало следующему пути: мшистые волокна → зернистые клетки → параллельные волокна → клетки Пуркинье → глубокие ядра. Восходящие волокна проецируются к клеткам Пуркинье, а также направляют коллатерали непосредственно в глубокие ядра. [10] Входы мшистого волокна и вьющегося волокна несут информацию, специфичную для волокна; мозжечок также получает дофаминергические , серотонинергические , норадренэргические и холинергические входы, которые предположительно осуществляют глобальную модуляцию. [20]
Кора мозжечка разделена на три слоя. Внизу лежит толстый зернистый слой, плотно упакованный гранулярными клетками, а также интернейронами , в основном клетками Гольджи , но также включая клетки Лугаро и униполярные клетки-щеточки . В середине находится слой Пуркинье, узкая зона, содержащая тела клеток Пуркинье и глиальных клеток Бергмана . Наверху находится молекулярный слой, который содержит уплощенные дендритные деревья клеток Пуркинье, а также огромный массив параллельных волокон, пронизывающих дендритные деревья клеток Пуркинье под прямым углом. Этот внешний слой коры мозжечка также содержит два типа тормозных интернейронов: звездчатые клетки и корзинчатые клетки . И звездчатые, и корзинчатые клетки образуют ГАМКергические синапсы на дендритах клеток Пуркинье. [10]

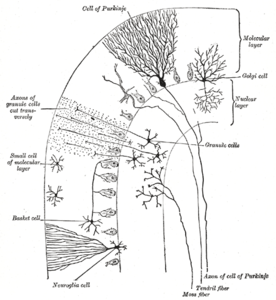
Слои коры мозжечка
[ редактировать ]Молекулярный слой
[ редактировать ]Верхний, самый внешний слой коры мозжечка — молекулярный слой. Этот слой содержит уплощенные дендритные деревья клеток Пуркинье и огромный массив параллельных волокон из зернистого слоя, которые пронизывают дендритные деревья клеток Пуркинье под прямым углом. Молекулярный слой также содержит два типа тормозных интернейронов: звездчатые клетки и корзинчатые клетки . И звездчатые, и корзинчатые клетки образуют ГАМКергические синапсы на дендритах клеток Пуркинье. [10]
Слой Пуркинье
[ редактировать ]
Клетки Пуркинье являются одними из наиболее характерных нейронов головного мозга и одним из первых обнаруженных типов — они были впервые описаны чешским анатомом Яном Евангелистой Пуркине в 1837 году. Они отличаются формой своего дендритного дерева: ветвью дендритов. очень обильно, но сильно уплощены в плоскости, перпендикулярной складкам мозжечка. Таким образом, дендриты клетки Пуркинье образуют плотную плоскую сеть, через которую под прямым углом проходят параллельные волокна. [10] Дендриты покрыты дендритными шипами , каждый из которых получает синаптический сигнал от параллельного волокна. Клетки Пуркинье получают больше синаптических входов, чем любой другой тип клеток головного мозга: по оценкам, количество шипов в одной клетке Пуркинье человека достигает 200 000. [10] Большие сферические тела клеток Пуркинье упакованы в узкий слой (толщиной в одну клетку) коры мозжечка, называемый слоем Пуркинье . После испускания коллатералей, воздействующих на близлежащие части коры, их аксоны направляются в глубокие ядра мозжечка , где они устанавливают порядка 1000 контактов каждый с несколькими типами ядерных клеток, и все это в пределах небольшого домена. Клетки Пуркинье используют ГАМК в качестве нейромедиатора и, следовательно, оказывают ингибирующее действие на свои мишени. [10]
Клетки Пуркинье образуют сердце мозжечкового контура, а их большой размер и характерный характер активности позволили относительно легко изучить закономерности их реакций на поведенческих животных с использованием методов внеклеточной регистрации. Клетки Пуркинье обычно излучают потенциалы действия с высокой скоростью даже при отсутствии синаптического входа. У бодрствующих животных типична средняя частота, составляющая около 40 Гц. Последовательности шипов представляют собой смесь так называемых простых и сложных шипов. Простой спайк представляет собой одиночный потенциал действия, за которым следует рефрактерный период длительностью около 10 мс; Сложный спайк представляет собой стереотипную последовательность потенциалов действия с очень короткими интервалами между спайками и уменьшающейся амплитудой. [22] Физиологические исследования показали, что сложные спайки (которые возникают при базовой частоте около 1 Гц и никогда не превышают 10 Гц) надежно связаны с активацией карабкающихся волокон, в то время как простые спайки возникают в результате сочетания базовой активности и параллельного входа волокон. За сложными спайками часто следует пауза в несколько сотен миллисекунд, в течение которой подавляется активность простых спайков. [23]
Специфической, узнаваемой особенностью нейронов Пуркинье является экспрессия кальбиндина . [24] Окрашивание кальбиндином головного мозга крыс после одностороннего хронического повреждения седалищного нерва позволяет предположить, что нейроны Пуркинье могут заново генерироваться во взрослом мозге, инициируя организацию новых долек мозжечка. [25]

Зернистый слой
[ редактировать ]
Зернистые клетки мозжечка , в отличие от клеток Пуркинье, относятся к числу самых маленьких нейронов головного мозга. Они также являются самыми многочисленными нейронами в мозге: у людей их общее количество составляет в среднем около 50 миллиардов, что означает, что около 3/4 нейронов мозга представляют собой зернистые клетки мозжечка. [10] Их клеточные тела упакованы в толстый слой в нижней части коры мозжечка. Зернистая клетка выделяет всего четыре-пять дендритов, каждый из которых заканчивается расширением, называемым дендритным коготком . [10] Эти расширения являются местами возбуждающего воздействия от мшистых волокон и тормозного воздействия от клеток Гольджи . [10]
Тонкие немиелинизированные аксоны гранулярных клеток поднимаются вертикально к верхнему (молекулярному) слою коры головного мозга, где они разделяются на две части, причем каждая ветвь движется горизонтально, образуя параллельное волокно ; Разделение вертикальной ветви на две горизонтальные ветви приводит к характерной Т-образной форме. Параллельное волокно человека проходит в среднем по 3 мм в каждую сторону от расщепления, общая длина около 6 мм (около 1/10 общей ширины коркового слоя). [10] По мере продвижения параллельные волокна проходят через дендритные деревья клеток Пуркинье, контактируя с одним из каждых 3–5, которые они проходят, образуя в общей сложности 80–100 синаптических связей с дендритными отростками клеток Пуркинье. [10] Клетки-гранулы используют глутамат в качестве нейромедиатора и, следовательно, оказывают возбуждающее воздействие на свои цели. [10]

Гранулярные клетки получают всю необходимую информацию из мшистых волокон, но превосходят их по численности в соотношении 200 к 1 (у людей). Таким образом, информация в состоянии активности популяции гранулярных клеток такая же, как и информация в мшистых волокнах, но перекодируется гораздо более обширным образом. Поскольку гранулярные клетки настолько малы и плотно упакованы, сложно зарегистрировать их пиковую активность у животных, ведущих себя, поэтому данных, которые можно было бы использовать в качестве основы для теоретизирования, мало. Самая популярная концепция их функции была предложена в 1969 году Дэвидом Марром , который предположил, что они могут кодировать комбинации входов мшистых волокон. Идея состоит в том, что, поскольку каждая гранулярная клетка получает входные данные только от 4–5 мшистых волокон, гранулярная клетка не будет реагировать, если активен только один из ее входных сигналов, но будет реагировать, если активны более одного. Эта комбинаторная схема кодирования потенциально позволила бы мозжечку проводить гораздо более тонкие различия между входными паттернами, чем это позволяли бы только мшистые волокна. [26]
Мшистые волокна
[ редактировать ]Мшистые волокна входят в зернистый слой из мест своего происхождения, многие из которых возникают из ядер моста , другие - из спинного мозга, вестибулярных ядер и т. Д. В человеческом мозжечке общее количество мшистых волокон оценивается в 200 миллионов. [10] Эти волокна образуют возбуждающие синапсы с клетками-зернами и клетками глубоких ядер мозжечка. Внутри зернистого слоя мшистое волокно образует ряд расширений, называемых розетками . Контакты между мшистыми волокнами и дендритами зернистых клеток происходят внутри структур, называемых клубочками . Каждый клубочек имеет в центре розетку из мшистых волокон и до 20 соприкасающихся с ним дендритных коготков зернистых клеток. Терминалы клеток Гольджи проникают в структуру и образуют тормозные синапсы на дендритах гранулярных клеток. Вся совокупность окружена оболочкой из глиальных клеток. [10] Каждое мшистое волокно дает боковые ветви к нескольким листкам мозжечка, образуя в общей сложности 20–30 розеток; таким образом, одно мшистое волокно вступает в контакт примерно с 400–600 гранулярными клетками. [10]
Восхождение волокон
[ редактировать ]Клетки Пуркинье также получают информацию от нижнего оливкового ядра на контралатеральной стороне ствола мозга через лазающие волокна . Хотя нижняя олива находится в продолговатом мозге и получает сигналы от спинного мозга, ствола мозга и коры головного мозга, ее выходные данные полностью направляются в мозжечок. Лазащее волокно отдает коллатерали к глубоким ядрам мозжечка перед тем, как войти в кору мозжечка, где оно разделяется примерно на 10 терминальных ветвей, каждая из которых передает сигнал одной клетке Пуркинье. [10] В отличие от более чем 100 000 входных сигналов от параллельных волокон, каждая клетка Пуркинье получает входные данные ровно от одного восходящего волокна; но это единственное волокно «взбирается» на дендриты клетки Пуркинье, обвивая их и образуя в общей сложности до 300 синапсов. [10] Суммарный входной сигнал настолько силен, что одиночный потенциал действия лазающего волокна способен вызвать в клетке Пуркинье расширенный комплексный спайк: вспышку из нескольких спайков подряд с уменьшающейся амплитудой, за которой следует пауза, во время которой активность подавляется. . Синапсы восходящих волокон покрывают тело клетки и проксимальные дендриты; эта зона лишена параллельных оптоволоконных входов. [10]
Лазящие волокна активируются с низкой частотой, но один потенциал действия карабкающегося волокна вызывает всплеск нескольких потенциалов действия в целевой клетке Пуркинье (сложный спайк). Контраст между параллельными волокнами и лазящими волокнами, входящими в клетки Пуркинье (более 100 000 одного типа против ровно одного другого типа), возможно, является самой провокационной особенностью анатомии мозжечка и послужил мотивом для многих теоретизирований. Фактически, функция карабкающихся волокон — самая спорная тема, касающаяся мозжечка. Существуют две школы мысли: одна вслед за Марром и Альбусом считает, что входные восходящие волокна служат в первую очередь обучающим сигналом, другая считает, что их функция заключается в непосредственном формировании выходной мощности мозжечка. Обе точки зрения подробно защищались в многочисленных публикациях. По словам одного из обзоров: «Пытаясь синтезировать различные гипотезы о функции лазающих волокон, возникает ощущение, что мы смотрим на рисунок Эшера. Кажется, что каждая точка зрения объясняет определенную совокупность результатов, но Когда пытаются свести воедино различные точки зрения, связной картины того, что делают лазающие волокна, не появляется. Для большинства исследователей лазающие волокна сигнализируют об ошибках в двигательной активности, либо в обычном порядке модуляции частоты разряда, либо в виде модуляции частоты разряда. Для других исследователей сообщение о «неожиданном событии» заключается в степени синхронности и ритмичности ансамбля среди популяции лазающих волокон». [23]
Глубокие ядра
[ редактировать ]
Глубокие ядра мозжечка представляют собой скопления серого вещества, лежащие внутри белого вещества в ядре мозжечка. Они, за небольшим исключением близлежащих вестибулярных ядер, являются единственными источниками сигнала из мозжечка. Эти ядра получают побочные проекции от мшистых и лазающих волокон, а также тормозящие сигналы от клеток Пуркинье коры мозжечка. Четыре ядра ( зубчатое , шаровидное , эмболиформное и фастигиальное ) сообщаются с разными частями головного мозга и коры мозжечка. (Шарообразное и эмболиформное ядра также называют объединенными во вставочном ядре ). Фастигиальные и вставочные ядра относятся к спиноцеребеллуму. Зубчатое ядро, которое у млекопитающих значительно крупнее остальных, образовано в виде тонкого извитого слоя серого вещества и сообщается исключительно с латеральными отделами коры мозжечка. Флоккулюс флоккулонодулярной доли — единственная часть коры мозжечка, которая не проецируется в глубокие ядра — вместо этого ее выходной сигнал поступает в вестибулярные ядра. [10]
Большинство нейронов в глубоких ядрах имеют крупные клеточные тела и сферические дендритные деревья радиусом около 400 мкм и используют глутамат в качестве нейромедиатора. Эти клетки проецируются на различные цели за пределами мозжечка. С ними смешано меньшее количество мелких клеток, которые используют ГАМК в качестве нейромедиатора и проецируются исключительно в нижнее оливковое ядро , источник лазающих волокон . Таким образом, нуклеооливарная проекция обеспечивает тормозную обратную связь , соответствующую возбуждающей проекции лазающих волокон к ядрам. Есть свидетельства того, что каждый небольшой кластер ядерных клеток проецируется на один и тот же кластер оливарных клеток, которые посылают к нему лазящие волокна; в обоих направлениях есть сильная и совпадающая топография. [10]
Когда аксон клетки Пуркинье входит в одно из глубоких ядер, он разветвляется, вступая в контакт как с большими, так и с малыми ядерными клетками, но общее количество контактирующих клеток составляет всего около 35 (у кошек). И наоборот, одна глубокая ядерная клетка получает информацию примерно от 860 клеток Пуркинье (опять же у кошек). [10]
Отделения
[ редактировать ]
С точки зрения макроанатомии кора мозжечка представляется однородным слоем ткани, а с точки зрения микроанатомии все части этого листа имеют одинаковое внутреннее строение. Однако существует ряд аспектов, в которых структура мозжечка разделена на отдельные части. Есть большие отсеки, которые обычно называют зонами ; их можно разделить на более мелкие отсеки, известные как микрозоны . [27]
Первые указания на компартментарную структуру появились в результате изучения рецептивных полей клеток в различных частях коры мозжечка. [27] Каждая часть тела соответствует определенным точкам мозжечка, но существует множество повторений базовой карты, образующих структуру, получившую название «переломная соматотопия». [28] Более четкие указания на компартментализацию получают путем иммуноокрашивания мозжечка на определенные типы белков. Самые известные из этих маркеров называются «зебрины», поскольку при окрашивании ими образуется сложный узор, напоминающий полосы на зебре. Полосы, генерируемые зебринами и другими маркерами компартментализации, ориентированы перпендикулярно складкам мозжечка, т. е. они узкие в медиолатеральном направлении, но значительно более вытянуты в продольном направлении. Разные маркеры создают разные наборы полос, ширина и длина варьируются в зависимости от местоположения, но все они имеют одинаковую общую форму. [27]
Оскарссон в конце 1970-х годов предположил, что эти корковые зоны можно разделить на более мелкие единицы, называемые микрозонами. [29] Микрозона определяется как группа клеток Пуркинье, имеющих одинаковое соматотопическое рецептивное поле. Было обнаружено, что микрозоны содержат порядка 1000 клеток Пуркинье каждая, расположенных в виде длинной узкой полоски, ориентированной перпендикулярно кортикальным складкам. [27] Таким образом, как показано на прилагаемой диаграмме, дендриты клеток Пуркинье уплощены в том же направлении, в котором простираются микрозоны, а параллельные волокна пересекают их под прямым углом. [10]
Структуру микрозон определяют не только рецептивные поля: вход лазящих волокон из нижнего ядра оливы не менее важен. Ветви лазящего волокна (обычно их около 10) обычно активируют клетки Пуркинье, принадлежащие одной микрозоне. Более того, оливарные нейроны, которые посылают лазающие волокна в одну и ту же микрозону, обычно соединяются щелевыми соединениями , которые синхронизируют их активность, в результате чего клетки Пуркинье внутри микрозоны демонстрируют коррелированную сложную импульсную активность в миллисекундном масштабе времени. [27] Кроме того, все клетки Пуркинье, принадлежащие к микрозоне, посылают свои аксоны в один и тот же небольшой кластер выходных клеток в глубоких ядрах мозжечка . [27] Наконец, аксоны корзинчатых клеток намного длиннее в продольном направлении, чем в медиолатеральном направлении, в результате чего они ограничиваются в основном одной микрозоной. [27] Следствием всей этой структуры является то, что клеточные взаимодействия внутри микрозоны гораздо сильнее, чем взаимодействия между различными микрозонами. [27]
В 2005 году Ричард Аппс и Мартин Гарвич обобщили доказательства того, что микрозоны сами по себе являются частью более крупного образования, которое они называют мультизональным микрокомплексом. Такой микрокомплекс включает в себя несколько пространственно разделенных корковых микрозон, каждая из которых проецируется на одну и ту же группу глубоких нейронов мозжечка, плюс группу связанных оливарных нейронов, которые проецируются на все включенные микрозоны, а также на глубокую ядерную область. [27]
Кровоснабжение
[ редактировать ]Мозжечок снабжается кровью из трех парных крупных артерий: верхней мозжечковой артерии (ПМА), передней нижней мозжечковой артерии (ПМАА) и задней нижней мозжечковой артерии (ЗНМА). SCA снабжает верхнюю часть мозжечка. На верхней поверхности она разделяется и разветвляется на мягкую мозговую оболочку, где ветви анастомозируют с ветвями передней и задней нижней мозжечковых артерий. AICA снабжает переднюю часть нижней поверхности мозжечка. PICA достигает нижней поверхности, где разделяется на медиальную и латеральную ветви. Медиальная ветвь продолжается назад до мозжечковой вырезки между двумя полушариями мозжечка; в то время как латеральная ветвь снабжает нижнюю поверхность мозжечка до его латеральной границы, где она анастомозирует с AICA и SCA.
Функция
[ редактировать ]Наиболее достоверные сведения о функции мозжечка были получены при изучении последствий его повреждения. У животных и людей с дисфункцией мозжечка наблюдаются прежде всего проблемы с двигательным контролем на той стороне тела, где находится поврежденная часть мозжечка. Они по-прежнему способны генерировать двигательную активность, но теряют точность, производя хаотичные, нескоординированные или неправильно рассчитанные движения. Стандартная проверка функции мозжечка заключается в том, чтобы дотянуться кончиком пальца до цели на расстоянии вытянутой руки: здоровый человек будет двигать кончиком пальца по быстрой прямой траектории, тогда как человек с повреждением мозжечка будет достигать цели медленно и беспорядочно, со многими средними движениями. -коррекция курса. Дефицит немоторных функций обнаружить труднее. Таким образом, общий вывод, сделанный десятилетия назад, заключается в том, что основная функция мозжечка — это калибровка детальной формы движения, а не инициирование движений или решение, какие движения выполнять. [13]
До 1990-х годов почти повсеместно считалось, что функция мозжечка связана исключительно с двигательной деятельностью, но новые открытия поставили эту точку зрения под сомнение. Исследования функциональной визуализации показали активацию мозжечка в отношении речи, внимания и мысленных образов; корреляционные исследования показали взаимодействие мозжечка и немоторных областей коры головного мозга; и у людей с повреждением, которое, по-видимому, ограничивается мозжечком, были обнаружены различные немоторные симптомы. [30] [31] В частности, мозжечково-когнитивно-аффективный синдром или синдром Шмахмана. [32] был описан у взрослых [33] и дети. [34] Оценки, основанные на функциональном картировании мозжечка с помощью функциональной МРТ, позволяют предположить, что более половины коры мозжечка взаимосвязано с ассоциативными зонами коры головного мозга. [35]
Кенджи Дойя утверждал, что функцию мозжечка лучше всего понимать не с точки зрения поведения, на которое он влияет, а с точки зрения нейронных вычислений, которые он выполняет; мозжечок состоит из большого числа более или менее независимых модулей, имеющих одинаковую геометрически правильную внутреннюю структуру и, следовательно, предполагается, что все они выполняют одни и те же вычисления. Если входные и выходные соединения модуля связаны с областями двигателя (а их много), то модуль будет участвовать в работе двигателя; но если связи связаны с областями, участвующими в немоторном познании, модуль покажет другие типы поведенческих коррелятов. Таким образом, мозжечок участвует в регуляции многих различных функциональных качеств, таких как привязанность, эмоции, включая эмоциональное восприятие языка тела. [36] и поведение. [37] [38] Мозжечок, как предполагает Дойя, лучше всего понимать как выбор прогнозирующих действий, основанный на «внутренних моделях» окружающей среды или устройстве контролируемого обучения , в отличие от базальных ганглиев , которые выполняют обучение с подкреплением , и коры головного мозга , которая осуществляет обучение без учителя. . [31] [39] Три десятилетия исследований мозга привели к предположению, что мозжечок генерирует оптимизированные ментальные модели и тесно взаимодействует с корой головного мозга, где обновленные внутренние модели воспринимаются как творческая интуиция («а-ха») в рабочей памяти. [40]
Принципы
[ редактировать ]Сравнительная простота и регулярность анатомии мозжечка привели к ранней надежде, что она может подразумевать аналогичную простоту вычислительных функций, как это выражено в одной из первых книг по электрофизиологии мозжечка « Мозжечок как нейрональная машина» Джона К. Экклса , Масао. Ито и Янош Сентаготаи . [41] Хотя полное понимание функции мозжечка остается неясным, по крайней мере четыре принципа были определены как важные: (1) упреждающая обработка, (2) дивергенция и конвергенция, (3) модульность и (4) пластичность.
- Прямая обработка : мозжечок отличается от большинства других частей мозга (особенно коры головного мозга) тем, что обработка сигналов почти полностью является прямой , то есть сигналы движутся через систему однонаправленно от входа к выходу с очень незначительной повторяющейся внутренней передачей. Небольшое количество рецидивов, которое действительно существует, состоит из взаимного торможения; цепи взаимного возбуждения отсутствуют. Этот режим работы с прямой связью означает, что мозжечок, в отличие от коры головного мозга, не может генерировать самоподдерживающиеся модели нейронной активности. Сигналы поступают в схему, обрабатываются каждым каскадом в последовательном порядке и затем выходят. Как писали Экклс, Ито и Сентаготаи: «Это исключение при проектировании всех возможностей реверберационных цепей нейронального возбуждения, несомненно, является большим преимуществом в работе мозжечка как компьютера, потому что то, чего требует остальная нервная система от мозжечок, по-видимому, не является каким-то выходным сигналом, выражающим работу сложных реверберационных цепей в мозжечке, а скорее быстрым и четким ответом на вход любого конкретного набора информации». [42]
- Дивергенция и конвергенция . В человеческом мозжечке информация от 200 миллионов входных волокон мшистого типа расширяется до 40 миллиардов гранулярных клеток , чьи параллельные выходные волокна затем сходятся на 15 миллионах клеток Пуркинье . [10] Из-за того, что они выстроены продольно, около 1000 клеток Пуркинье, принадлежащих к микрозоне, могут получать входные данные от 100 миллионов параллельных волокон и фокусировать свои собственные выходные данные на группе из менее чем 50 глубоких ядерных клеток. [27] Таким образом, сеть мозжечка получает скромное количество входных данных, очень тщательно их обрабатывает через свою строго структурированную внутреннюю сеть и отправляет результаты через очень ограниченное количество выходных клеток.
- Модульность : система мозжечка функционально разделена на более или менее независимые модули, число которых, вероятно, исчисляется сотнями и тысячами. Все модули имеют схожую внутреннюю структуру, но разные входы и выходы. Модуль (мультизональный микрокомпартмент в терминологии Аппса и Гарвича) состоит из небольшого скопления нейронов нижнего оливкового ядра, набора длинных узких полосок клеток Пуркинье в коре мозжечка (микрозон) и небольшого скопления нейронов. в одном из глубоких ядер мозжечка. Разные модули используют входные данные от мшистых и параллельных волокон, но в других отношениях они, по-видимому, функционируют независимо — выходные данные одного модуля, по-видимому, не оказывают существенного влияния на активность других модулей. [27]
- Пластичность . Синапсы между параллельными волокнами и клетками Пуркинье, а также синапсы между мшистыми волокнами и глубокими ядерными клетками подвержены изменению своей прочности. В одном модуле мозжечка входные сигналы от миллиарда параллельных волокон сходятся в группе из менее чем 50 глубоких ядерных клеток, и влияние каждого параллельного волокна на эти ядерные клетки можно регулировать. Такое расположение дает огромную гибкость для точной настройки взаимосвязи между входами и выходами мозжечка. [43]
Обучение
[ редактировать ]Имеются убедительные доказательства того, что мозжечок играет важную роль в некоторых типах двигательного обучения. Задачи, в которых мозжечок наиболее явно задействован, — это те, в которых необходимо внести тонкую корректировку способа выполнения действия. Однако существует много споров о том, происходит ли обучение внутри самого мозжечка или оно просто служит для передачи сигналов, способствующих обучению в других структурах мозга. [43] Большинство теорий, приписывающих обучение схемам мозжечка, основаны на идеях Дэвида Марра. [26] и Джеймс Альбус , [7] который постулировал, что лазающие волокна обеспечивают обучающий сигнал, который вызывает синаптические модификации в параллельных волокон – клеток Пуркинье . синапсе [44] Марр предположил, что восхождение на вход волокна приведет к усилению синхронно активированных параллельных входов волокна. Однако большинство последующих моделей обучения мозжечка последовали примеру Альбуса и предположили, что активность карабкающихся волокон будет сигналом ошибки и приведет к ослаблению синхронно активированных входов параллельных волокон. Некоторые из этих более поздних моделей, такие как модель адаптивного фильтра Fujita. [45] предприняли попытки понять функцию мозжечка с точки зрения теории оптимального управления .
Идея о том, что активность карабкающихся волокон действует как сигнал ошибки, рассматривалась во многих экспериментальных исследованиях, причем некоторые поддерживали ее, но другие подвергали сомнению. [23] В новаторском исследовании Гилберта и Таха, проведенном в 1977 году, клетки Пуркинье обезьян, изучающих задачу достижения цели, показали повышенную комплексную импульсную активность, которая, как известно, надежно указывает на активность входных клеток в карабкающиеся волокна, в периоды, когда производительность была плохой. [46] В нескольких исследованиях двигательного обучения кошек наблюдалась сложная импульсивная активность, когда возникало несоответствие между запланированным движением и фактически выполненным движением. Исследования вестибуло-окулярного рефлекса (который стабилизирует зрительное изображение на сетчатке при повороте головы) показали, что активность восходящих волокон указывает на «скольжение сетчатки», хотя и не совсем простым способом. [23]
Одной из наиболее широко изученных задач мозжечкового обучения является парадигма обусловливания моргания , в которой нейтральный условный стимул (УС), такой как тон или свет, неоднократно сочетается с безусловным стимулом (УС), таким как дуновение воздуха, который вызывает ответ моргания. После таких повторных представлений CS и US CS в конечном итоге вызовет моргание перед US, условный ответ или CR. Эксперименты показали, что поражения, локализованные либо в определенной части вставленного ядра (одного из глубоких ядер мозжечка), либо в нескольких определенных точках коры мозжечка, могут отменить обучение условно синхронизированной реакции моргания. Если выходы мозжечка фармакологически инактивированы, а входы и внутриклеточные цепи остаются нетронутыми, обучение происходит даже тогда, когда животное не проявляет никакой реакции, тогда как, если внутримозжечковые цепи нарушены, обучение не происходит — эти факты, взятые вместе, дают веские основания полагать, что обучение действительно происходит внутри мозжечка. [47]
Теории и вычислительные модели
[ редактировать ]
Большая база знаний об анатомическом строении и поведенческих функциях мозжечка сделала его благодатной почвой для теоретизирования — теорий о функции мозжечка, пожалуй, больше, чем о любой другой части мозга. Самое основное различие между ними проводится между «теориями обучения» и «теориями производительности», то есть теориями, которые используют синаптическую пластичность мозжечка для объяснения его роли в обучении, и теориями, которые учитывают аспекты текущего поведения на мозжечке. Основа обработки сигналов мозжечка. Некоторые теории обоих типов были сформулированы в виде математических моделей и смоделированы с помощью компьютеров. [44]
Возможно, самой ранней теорией «производительности» была гипотеза «линии задержки» Валентино Брайтенберга . Первоначальная теория, выдвинутая Брайтенбергом и Роджером Этвудом в 1958 году, предполагала, что медленное распространение сигналов по параллельным волокнам приводит к предсказуемым задержкам, которые позволяют мозжечку обнаруживать временные соотношения в пределах определенного окна. [48] Экспериментальные данные не подтвердили первоначальную форму теории, но Брайтенберг продолжал выступать за модифицированные версии. [49] Гипотезу о том, что мозжечок функционирует, по существу, как система синхронизации, также отстаивал Ричард Иври . [50] Другой влиятельной теорией «производительности» является теория тензорных сетей Пеллиониша и Ллинаса , которая предоставила продвинутую математическую формулировку идеи о том, что фундаментальные вычисления, выполняемые мозжечком, заключаются в преобразовании сенсорных координат в моторные. [51]
Почти все теории из категории «обучение» основаны на публикациях Марра и Альбуса. В статье Марра 1969 года было высказано предположение, что мозжечок — это устройство, позволяющее научиться связывать элементарные движения, кодируемые лазящими волокнами, с входными сигналами мшистых волокон, которые кодируют сенсорный контекст. [26] В 1971 году Альбус предположил, что клетки Пуркинье мозжечка функционируют как перцептрон , абстрактное обучающее устройство, работающее на основе нейронов. [7] Самое основное различие между теориями Марра и Альбуса заключается в том, что Марр предполагал, что активность карабкающихся волокон приведет к усилению синапсов параллельных волокон, тогда как Альбус предположил, что они будут ослаблены. Альбус также сформулировал свою версию в виде программного алгоритма, который он назвал CMAC (контроллер артикуляции модели мозжечка), который был протестирован в ряде приложений. [52]
Клиническое значение
[ редактировать ]

Повреждение мозжечка часто вызывает двигательные симптомы, детали которых зависят от пораженной части мозжечка и способа его повреждения. Повреждение флокулонодулярной доли может проявляться в виде потери равновесия и, в частности, в измененной, нерегулярной походке при ходьбе с широкой позицией, вызванной трудностями в балансировании. [13] Повреждение латеральной зоны обычно вызывает проблемы при умелых произвольных и запланированных движениях, что может привести к ошибкам в силе, направлении, скорости и амплитуде движений. Другие проявления включают гипотонию (снижение мышечного тонуса), дизартрию (проблемы с речевой артикуляцией), дисметрию (проблемы с оценкой расстояния или диапазона движений), дисдиадохокинезию (неспособность выполнять быстрые чередующиеся движения, такие как ходьба), нарушение контрольного рефлекса или феномен отскока и интенционный тремор (непроизвольные движения, вызванные попеременным сокращением противоположных групп мышц). [53] [54] Повреждение средней линии может нарушить движения всего тела, тогда как повреждение, локализованное латерально, с большей вероятностью нарушит тонкие движения рук или конечностей. Повреждение верхней части мозжечка обычно приводит к нарушениям походки и другим проблемам с координацией ног; повреждение нижней части чаще вызывает нескоординированные или плохо направленные движения рук и кистей, а также затруднения в скорости. [13] Этот комплекс двигательных симптомов называется атаксией .
Для выявления проблем мозжечка неврологическое обследование включает оценку походки (широкая походка указывает на атаксию), пальцевые тесты и оценку осанки. [5] Если показана дисфункция мозжечка, можно использовать магнитно-резонансную томографию для получения подробной картины любых структурных изменений, которые могут существовать. [55]
Список медицинских проблем, которые могут вызвать повреждение мозжечка, длинный, включая инсульт , кровоизлияние , отек головного мозга ( отек мозга ), опухоли , алкоголизм , физические травмы , такие как огнестрельные ранения или взрывчатые вещества, а также хронические дегенеративные состояния, такие как оливопонтоцеребеллярная атрофия . [56] [57] Некоторые формы мигренозной головной боли могут также вызывать временную дисфункцию мозжечка различной степени тяжести. [58] Инфекция может привести к повреждению мозжечка при таких состояниях, как прионные заболевания. [59] и синдром Миллера-Фишера, вариант синдрома Гийена-Барре .
Старение
[ редактировать ]Человеческий мозжечок меняется с возрастом. Эти изменения могут отличаться от изменений в других частях мозга.Мозжечок — самая молодая область мозга (и часть тела) у долгожителей согласно эпигенетическому биомаркеру тканевого возраста, известному как эпигенетические часы : он примерно на 15 лет моложе, чем ожидается у долгожителей. [60] Кроме того, паттерны экспрессии генов в мозжечке человека демонстрируют меньшие возрастные изменения, чем в коре головного мозга . [61] В некоторых исследованиях сообщалось об уменьшении количества клеток или объема тканей, но объем данных, касающихся этого вопроса, не очень велик. [62] [63]
Нарушения развития и дегенеративные нарушения
[ редактировать ]
Врожденные пороки развития, наследственные нарушения и приобретенные заболевания могут влиять на структуру мозжечка и, следовательно, на его функцию. Если причина не обратима, единственное возможное лечение — помочь людям жить со своими проблемами. [64] Визуализация мозжечка плода с помощью ультразвукового сканирования на сроке от 18 до 20 недель беременности может использоваться для выявления плода дефектов нервной трубки с чувствительностью до 99%. [65]
При нормальном развитии эндогенная передача сигналов звукового ежа стимулирует быструю пролиферацию предшественников гранулярных нейронов мозжечка (CGNP) во внешнем слое гранул (EGL). Развитие мозжечка происходит во время позднего эмбриогенеза и раннего постнатального периода, при этом пролиферация CGNP в EGL достигает пика во время раннего развития (7-й постнатальный день у мышей). [66] Когда CGNP окончательно дифференцируются в гранулярные клетки мозжечка (также называемые гранулярными нейронами мозжечка, CGN), они мигрируют во внутренний слой гранул (IGL), образуя зрелый мозжечок (к 20-му дню постнатального развития у мышей). [66] Мутации, которые аномально активируют передачу сигналов Sonic hedgehog, предрасполагают к раку мозжечка ( медуллобластоме ) у людей с синдромом Горлина и на моделях генно-инженерных мышей . [67] [68]
Врожденный порок развития или недоразвитие ( гипоплазия ) червя мозжечка характерно как для синдрома Денди-Уокера, так и для синдрома Жубера . [69] [70] весь мозжечок В очень редких случаях может отсутствовать . [71] Наследственные неврологические расстройства, болезнь Мачадо-Джозефа , атаксия-телеангиэктазия и атаксия Фридрейха вызывают прогрессирующую нейродегенерацию, связанную с потерей мозжечка. [56] [64] Врожденные пороки развития головного мозга за пределами мозжечка могут, в свою очередь, вызывать грыжу ткани мозжечка , что наблюдается при некоторых формах мальформации Арнольда-Киари . [72]
Другие состояния, тесно связанные с дегенерацией мозжечка, включают идиопатические прогрессирующие неврологические расстройства, множественную системную атрофию и синдром Рамсея Ханта I типа . [73] [74] и аутоиммунное заболевание, паранеопластическая дегенерация мозжечка , при которой опухоли в других частях тела вызывают аутоиммунный ответ, который вызывает потерю нейронов в мозжечке. [75] Атрофия мозжечка может возникнуть в результате острого дефицита витамина B1 ( тиамина ), что наблюдается при бери-бери и синдроме Вернике-Корсакова . [76] или витамина Е. дефицит [64]
Атрофия мозжечка наблюдалась при многих других неврологических заболеваниях, включая болезнь Хантингтона , рассеянный склероз , [59] эссенциальный тремор , прогрессирующая миоклонусная эпилепсия и болезнь Нимана-Пика . Атрофия мозжечка также может возникнуть в результате воздействия токсинов, включая тяжелые металлы , фармацевтические или рекреационные наркотики . [64]
Боль
[ редактировать ]Существует общее мнение, что мозжечок участвует в обработке боли. [77] [78] Мозжечок получает болевые сигналы как от нисходящих кортико-мозжечковых путей, так и от восходящих спинно-мозжечковых путей через ядра моста и нижние оливы. Часть этой информации передается в двигательную систему, вызывая сознательное двигательное избегание боли, которое оценивается в зависимости от интенсивности боли.
Считается, что эти прямые болевые воздействия, а также косвенные воздействия вызывают долгосрочное поведение избегания боли, что приводит к хроническим изменениям позы и, следовательно, к функциональному и анатомическому ремоделированию вестибулярных и проприоцептивных ядер. В результате хроническая нейропатическая боль может вызвать макроскопическое анатомическое ремоделирование заднего мозга, включая мозжечок. [25] Величина этого ремоделирования и индукция маркеров предшественников нейронов предполагают вклад взрослого нейрогенеза в эти изменения.
Сравнительная анатомия и эволюция
[ редактировать ]
Цепи мозжечка одинаковы у всех классов позвоночных . , включая рыб, рептилий, птиц и млекопитающих [79] Аналогичное строение мозга имеется и у головоногих моллюсков с хорошо развитым мозгом, например у осьминогов . [80] Это было воспринято как доказательство того, что мозжечок выполняет функции, важные для всех видов животных , имеющих мозг.
У разных видов позвоночных существуют значительные различия в размерах и форме мозжечка. У земноводных он развит слабо, а у миног и миксин мозжечок едва отличим от ствола мозга. Хотя в этих группах присутствует спиноцеребеллум, первичными структурами являются небольшие парные ядра, соответствующие вестибулоцеребеллуму. [81] Мозжечок немного крупнее у рептилий, значительно крупнее у птиц и еще крупнее у млекопитающих. Большие парные и извитые доли, обнаруженные у человека, типичны для млекопитающих, но в других группах мозжечок, как правило, представляет собой одну срединную долю и либо гладкий, либо лишь слегка бороздчатый. У млекопитающих неоцеребеллум составляет большую часть мозжечка по массе, но у других позвоночных это обычно спиноцеребеллум. [81]
Мозжечок хрящевых и костных рыб необычайно велик и сложен. По крайней мере, в одном важном отношении он отличается по внутренней структуре от мозжечка млекопитающих: мозжечок рыб не содержит дискретных глубоких ядер мозжечка . Вместо этого основной мишенью клеток Пуркинье являются клетки особого типа, распределенные по коре мозжечка, тип, не встречающийся у млекопитающих. У мормирид (семейство слабоэлектрочувствительных пресноводных рыб) мозжечок значительно крупнее остального мозга. Самая большая его часть представляет собой специальную структуру, называемую клапаном , которая имеет необычно правильную архитектуру и получает большую часть информации от электросенсорной системы. [82]
Отличительной чертой мозжечка млекопитающих является расширение боковых долей, основные взаимодействия которых происходят с неокортексом. По мере того как обезьяны превращались в человекообразных обезьян, расширение боковых долей продолжалось одновременно с расширением лобных долей неокортекса. У предков гоминид и Homo sapiens до среднего плейстоцена мозжечок продолжал расширяться, но лобные доли расширялись быстрее. Однако самый поздний период эволюции человека на самом деле мог быть связан с увеличением относительных размеров мозжечка, поскольку неокортекс несколько уменьшал свои размеры, а мозжечок расширялся. [83] Размер человеческого мозжечка по сравнению с остальным мозгом увеличивается в размерах, в то время как головной мозг уменьшается в размерах. [84] Поскольку как развитие, так и выполнение двигательных задач, зрительно-пространственных навыков и обучения происходит в мозжечке, считается, что рост мозжечка имеет некоторую форму корреляции с улучшением когнитивных способностей человека. [85] Боковые полушария мозжечка теперь в 2,7 раза больше как у человека, так и у обезьян, чем у обезьян. [84] Эти изменения в размерах мозжечка нельзя объяснить большей мышечной массой. Они показывают, что либо развитие мозжечка тесно связано с развитием остального мозга, либо что нейронная активность, происходящая в мозжечке, была важна в ходе эволюции гоминид . Из-за роли мозжечка в когнитивных функциях увеличение его размера могло сыграть роль в когнитивном расширении. [84]
Структуры, подобные мозжечку
[ редактировать ]Большинство видов позвоночных имеют мозжечок и одну или несколько мозжечковоподобных структур — областей мозга, напоминающих мозжечок с точки зрения цитоархитектуры и нейрохимии . [79] Единственная структура, подобная мозжечку, обнаруженная у млекопитающих, — это дорсальное ядро улитки (DCN), одно из двух основных сенсорных ядер, которые получают сигналы непосредственно от слухового нерва . DCN представляет собой слоистую структуру, нижний слой которой содержит гранулярные клетки, аналогичные клеткам мозжечка, дающие начало параллельным волокнам , которые поднимаются к поверхностному слою и проходят через него горизонтально. Поверхностный слой содержит набор ГАМКергических нейронов, называемых клетками колеса телеги , которые анатомически и химически напоминают клетки Пуркинье — они получают параллельные входные волокна, но не имеют никаких входных сигналов, напоминающих лазающие волокна . Выходные нейроны DCN представляют собой пирамидальные клетки . Они глутаматергические, но в некоторых отношениях напоминают клетки Пуркинье — у них есть колючие, уплощенные поверхностные дендритные деревья, которые получают параллельные входные волокна, но у них также есть базальные дендриты, которые получают входные данные от волокон слухового нерва, которые проходят через DCN в направлении под прямым углом к параллельным волокнам. DCN наиболее развит у грызунов и других мелких животных и значительно снижен у приматов. Его функция не совсем понятна; наиболее популярные предположения так или иначе связывают его с пространственным слухом. [86]
У большинства видов рыб и амфибий есть система боковой линии , которая улавливает волны давления в воде. Одна из областей мозга, которая получает первичную информацию от органа боковой линии, медиальное октаволатеральное ядро, имеет структуру, подобную мозжечку, с зернистыми клетками и параллельными волокнами. У электрочувствительных рыб сигнал от электросенсорной системы поступает в дорсальное октаволатеральное ядро, которое также имеет структуру, подобную мозжечку. У лучепёрых рыб (самая большая группа) в зрительной тектуме имеется слой — маргинальный слой, похожий на мозжечок. [79]
Все эти структуры, подобные мозжечку, по-видимому, в первую очередь связаны с сенсорикой, а не с моторикой. У всех из них есть гранулярные клетки, которые дают начало параллельным волокнам, которые соединяются с нейронами, подобными Пуркинье, с помощью модифицируемых синапсов , но ни у одного из них нет лазающих волокон, сравнимых с волокнами мозжечка — вместо этого они получают прямую информацию от периферических органов чувств. Ни у одного из них нет подтвержденной функции, но наиболее влиятельное предположение заключается в том, что они служат для преобразования сенсорных сигналов каким-то сложным способом, возможно, для компенсации изменений в позе тела. [79] Фактически, Джеймс М. Бауэр и другие утверждали, частично на основе этих структур, а частично на основе исследований мозжечка, что мозжечок сам по себе является сенсорной структурой и что он способствует моторному контролю, перемещая тело в способ управления результирующими сенсорными сигналами. [87] Несмотря на точку зрения Бауэра, существуют также убедительные доказательства того, что мозжечок напрямую влияет на двигательную активность млекопитающих. [88] [89]
История
[ редактировать ]
Описания
[ редактировать ]Даже самые ранние анатомы смогли узнать мозжечок по его характерному внешнему виду. Аристотель и Герофил (цитируется по Галену ) называли его παρεγκεφαλίς ( parenkephalis ), в отличие от ἐγκέφαλος ( энкефалос ) или собственно мозга. Подробное описание Галена — самое раннее из сохранившихся. Он предположил, что мозжечок является источником двигательных нервов. [90]
Дальнейшие значительные события произошли только в эпоху Возрождения . Везалий кратко обсудил мозжечок, а более подробно анатомия была описана Томасом Уиллисом в 1664 году. Больше анатомических работ было проведено в 18 веке, но только в начале 19 века были получены первые сведения о функции мозжечка. полученный. Луиджи Роландо в 1809 году установил ключевой вывод о том, что повреждение мозжечка приводит к двигательным нарушениям. Жан-Пьер Флурен в первой половине XIX века провел детальную экспериментальную работу, в ходе которой выяснилось, что животные с поражением мозжечка еще могут двигаться, но с потерей координации (странные движения, неловкая походка и мышечная слабость), и что восстановление после поражение может быть почти полным, если поражение не очень обширное. [91] К началу 20 века было широко признано, что основная функция мозжечка связана с контролем движений; В первой половине 20-го века появилось несколько подробных описаний клинических симптомов, связанных с заболеванием мозжечка у человека. [5]
Этимология
[ редактировать ]Название мозжечок является уменьшительным от слова cerebrum (мозг); [92] дословно это можно перевести как « маленький мозг» . Латинское название является прямым переводом древнегреческого слова παρεγκεφαλίς ( parenkephalis ), которое использовалось в трудах Аристотеля, первого известного писателя, описавшего структуру. [93] Никакое другое имя не используется в англоязычной литературе, но исторически использовались различные названия греческого или латинского происхождения, в том числе cerebrum parvum , [94] энцефалион , [95] Энкранион , [94] мозг задний [96] и парэнцефалия . [94]
См. также
[ редактировать ]Ссылки
[ редактировать ]![]() Эта статья была отправлена в WikiJournal of Medicine на внешнее академическое рецензирование в 2016 году ( отчеты рецензентов ). Обновленный контент был реинтегрирован на страницу Википедии по лицензии CC-BY-SA-3.0 ( 2016 г. ). Проверенная версия записи: Мэрион Райт; Уильям Скэггс ; Финн Аруп Нильсен; и др. (30 июня 2016 г.). «Мозжечок» (PDF) . Викижурнал медицины . 3 (1). дои : 10.15347/WJM/2016.001 . ISSN 2002-4436 . Викиданные Q44001486 .
Эта статья была отправлена в WikiJournal of Medicine на внешнее академическое рецензирование в 2016 году ( отчеты рецензентов ). Обновленный контент был реинтегрирован на страницу Википедии по лицензии CC-BY-SA-3.0 ( 2016 г. ). Проверенная версия записи: Мэрион Райт; Уильям Скэггс ; Финн Аруп Нильсен; и др. (30 июня 2016 г.). «Мозжечок» (PDF) . Викижурнал медицины . 3 (1). дои : 10.15347/WJM/2016.001 . ISSN 2002-4436 . Викиданные Q44001486 .
- ^ Ходос В. (2009). «Эволюция мозжечка». Энциклопедия неврологии . Берлин, Гейдельберг: Springer. стр. 1240–1243. дои : 10.1007/978-3-540-29678-2_3124 . ISBN 978-3-540-23735-8 .
- ^ Вольф У, Рапопорт М.Дж., Швейцер Т.А. (2009). «Оценка аффективного компонента мозжечкового когнитивно-аффективного синдрома». Журнал нейропсихиатрии и клинических нейронаук . 21 (3): 245–53. дои : 10.1176/jnp.2009.21.3.245 . ПМИД 19776302 .
- ^ Шмахманн Дж. Д., Каплан Д. (февраль 2006 г.). «Познание, эмоции и мозжечок» . Мозг . 129 (Часть 2): 290–2. дои : 10.1093/brain/awh729 . ПМИД 16434422 .
- ^ Шмахманн, Джереми Д. (2019). «Мозжечок и познание» . Письма по неврологии . 688 (688): 62–75. doi : 10.1016/j.neulet.2018.07.005 – через Elsevier Science Direct.
- ^ Перейти обратно: а б с д Fine EJ, Ионита CC, Лор Л. (декабрь 2002 г.). «История развития мозжечкового исследования». Семинары по неврологии . 22 (4): 375–84. дои : 10.1055/s-2002-36759 . ПМИД 12539058 . S2CID 260317107 .
- ^ Перейти обратно: а б Первс Д., Августин Г.Дж., Фитцпатрик Д., Холл У.К., ЛаМантия А.С., Уайт Л.Е. (2011). Нейронаука (5-е изд.). Сандерленд, Массачусетс: Синауэр. стр. 417–423. ISBN 978-0-87893-695-3 .
- ^ Перейти обратно: а б с Альбус Дж.С. (1971). «Теория функции мозжечка». Математика. Биологические науки . 10 (1–2): 25–61. CiteSeerX 10.1.1.14.7524 . дои : 10.1016/0025-5564(71)90051-4 .
- ^ Первс Д., Августин Г.Дж., Фитцпатрик Д., Холл У.К., ЛаМантия А.С., Уайт Л.Е. (2007). Нейронаука (4-е изд.). Нью-Йорк: WH Freeman. стр. 197–200. ISBN 978-0-87893-697-7 .
- ^ Стэндринг С., Борли Н.Р. и др., ред. (2008). «Глава 20». Анатомия Грея: анатомические основы клинической практики (40-е изд.). Лондон: Черчилль Ливингстон. п. 297. ИСБН 978-0-8089-2371-8 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С Ллинас Р.Р., Уолтон К.Д., Ланг Э.Дж. (2004). «Гл. 7 Мозжечок ». В Shepherd GM (ред.). Синаптическая организация мозга . Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-515955-4 .
- ^ Эркулано-Хаузель С (2010). «Координированное масштабирование коркового и мозжечкового количества нейронов» . Границы нейроанатомии . 4 : 12. дои : 10.3389/fnana.2010.00012 . ПМЦ 2839851 . ПМИД 20300467 .
- ^ Лю, Вэньцзяо; Ву, Йе; Хюинь, Кхой Мин; Ахмад, Сахар; Да, Пью-Тиан (2024). «Мультимодальный субмиллиметровый МРТ-атлас мозжечка человека» . Научные отчеты . 14 (1): 5622. Бибкод : 2024NatSR..14.5622L . дои : 10.1038/s41598-024-55412-y . ISSN 2045-2322 . ПМЦ 10920891 . ПМИД 38453991 .
- ^ Перейти обратно: а б с д и ж г час я дж Гез С., Фан С. (1985). «Мозжечок». В Кандель Э.Р., Шварц Дж.Х. (ред.). Принципы нейронауки, 2-е издание . Нью-Йорк: Эльзевир. стр. 502–522.
- ^ Снайдер Р.С., Стоуэлл А. (1 ноября 1944 г.). «Приемные области тактильной, слуховой и зрительной систем мозжечка» . Журнал нейрофизиологии . 7 (6): 331–357. дои : 10.1152/jn.1944.7.6.331 . S2CID 146700933 .
- ^ Кингсли Р.Э. (2000). Краткий текст по неврологии (2-е изд.). Липпинкотт Уильямс и Уилкинс. ISBN 978-0-683-30460-2 .
- ^ Петерсен С.Е., Фокс П.Т., Познер М.И., Минтун М., Райхл М.Е. (1989). «Позитронно-эмиссионные томографические исследования обработки отдельных слов». Журнал когнитивной нейронауки . 1 (2): 153–70. дои : 10.1162/jocn.1989.1.2.153 . ПМИД 23968463 . S2CID 35159122 .
- ^ Тимманн Д., Даум I (2007). «Вклад мозжечка в когнитивные функции: отчет о ходе работы после двух десятилетий исследований» . Мозжечок . 6 (3): 159–62. дои : 10.1080/14734220701496448 . PMID 17786810 . S2CID 25671398 .
- ^ Стрик П.Л., Дум Р.П., Фьес Дж.А. (2009). «Мозжечок и немоторные функции». Ежегодный обзор неврологии . 32 : 413–34. дои : 10.1146/annurev.neuro.31.060407.125606 . ПМИД 19555291 . S2CID 1066141 .
- ^ Бакнер Р.Л. (октябрь 2013 г.). «Мозжечок и когнитивная функция: 25 лет знаний анатомии и нейровизуализации» . Нейрон . 80 (3): 807–15. дои : 10.1016/j.neuron.2013.10.044 . ПМИД 24183029 .
- ^ Швайгофер Н., Дойя К., Курода С. (март 2004 г.). «Мозжечковая аминергическая нейромодуляция: к функциональному пониманию». Исследования мозга. Обзоры исследований мозга . 44 (2–3): 103–16. дои : 10.1016/j.brainresrev.2003.10.004 . ПМИД 15003388 . S2CID 7352039 .
- ^ Фелисола С.Дж., Накамура Й., Оно Й., Китамура К., Кикучи К., Онодера Й., Исе К., Такасе К., Сугавара А., Хаттангади Н., Рейни В.Е., Сато Ф., Сасано Х. (апрель 2014 г.). «PCP4: регулятор синтеза альдостерона в тканях надпочечников человека» . Журнал молекулярной эндокринологии . 52 (2): 159–67. дои : 10.1530/JME-13-0248 . ПМК 4103644 . ПМИД 24403568 .
- ^ Экклс Дж.К., Ллинас Р., Сасаки К. (январь 1966 г.). «Возбуждающее синаптическое действие лазающих волокон на клетки Пуркинье мозжечка» . Журнал физиологии . 182 (2): 268–96. doi : 10.1113/jphysicalol.1966.sp007824 . ПМЦ 1357472 . ПМИД 5944665 .
- ^ Перейти обратно: а б с д Симпсон Дж.И., Уайли Д.Р., Де Зеув К.И. (1996). «О восходящих оптоволоконных сигналах и их последствиях». Поведение. Наука о мозге . 19 (3): 384–398. дои : 10.1017/S0140525X00081486 .
- ^ Уитни Э.Р., Кемпер Т.Л., Розен Д.Л., Бауман М.Л., Блатт Г.Дж. (февраль 2008 г.). «Кальбиндин-D28k является более надежным маркером клеток Пуркинье человека, чем стандартные окраски по Нисслю: стереологический эксперимент». Журнал методов нейробиологии . 168 (1): 42–7. doi : 10.1016/j.jneumeth.2007.09.009 . ПМИД 17961663 . S2CID 10505177 .
- ^ Перейти обратно: а б Русанеску Г., Мао Дж. (февраль 2017 г.). «Повреждение периферических нервов вызывает нейрогенез и ремоделирование головного мозга у взрослых» . Журнал клеточной и молекулярной медицины . 21 (2): 299–314. дои : 10.1111/jcmm.12965 . ПМК 5264155 . ПМИД 27665307 .
- ^ Перейти обратно: а б с Марр Д. (июнь 1969 г.). «Теория коры мозжечка» . Журнал физиологии . 202 (2): 437–70. дои : 10.1113/jphysicalol.1969.sp008820 . ПМЦ 1351491 . ПМИД 5784296 .
- ^ Перейти обратно: а б с д и ж г час я дж к Аппс Р., Гарвич М. (апрель 2005 г.). «Анатомо-физиологические основы обработки информации мозжечком». Обзоры природы. Нейронаука . 6 (4): 297–311. дои : 10.1038/nrn1646 . ПМИД 15803161 . S2CID 10769826 .
- ^ Манни Э., Петрозини Л. (март 2004 г.). «Век соматотопии мозжечка: дискуссионное представление». Обзоры природы. Нейронаука . 5 (3): 241–9. дои : 10.1038/nrn1347 . ПМИД 14976523 . S2CID 30232749 .
- ^ Оскарссон О (1979). «Функциональные единицы мозжечково-сагиттальных зон и микрозон». Тенденции нейробиологии . 2 : 143–145. дои : 10.1016/0166-2236(79)90057-2 . S2CID 53272245 .
- ^ Рэпп Б (2001). Справочник по когнитивной нейропсихологии: что говорят недостатки о человеческом разуме . Психология Пресс. п. 481. ИСБН 978-1-84169-044-5 .
- ^ Перейти обратно: а б Дойя К. (декабрь 2000 г.). «Взаимодополняющая роль базальных ганглиев и мозжечка в обучении и контроле моторики». Современное мнение в нейробиологии . 10 (6): 732–9. дои : 10.1016/S0959-4388(00)00153-7 . ПМИД 11240282 . S2CID 10962570 .
- ^ Манто М, Мариен П (2015). «Синдром Шмахмана – выявление третьего краеугольного камня клинической аксиологии» . Мозжечок и атаксии . 2 : 2. дои : 10.1186/s40673-015-0023-1 . ПМЦ 4552302 . ПМИД 26331045 .
- ^ Шмахманн Дж.Д., Шерман Дж.К. (апрель 1998 г.). «Мозжечково-когнитивно-аффективный синдром» . Мозг . 121 (4): 561–79. дои : 10.1093/мозг/121.4.561 . ПМИД 9577385 .
- ^ Левисон Л., Кронин-Голомб А., Шмахманн Дж.Д. (май 2000 г.). «Нейропсихологические последствия резекции опухоли мозжечка у детей: мозжечковый когнитивно-аффективный синдром в детской популяции» . Мозг . 123 (5): 1041–50. дои : 10.1093/мозг/123.5.1041 . ПМИД 10775548 .
- ^ Бакнер Р.Л., Кринен Ф.М., Кастельянос А., Диас Х.К., Йео Б.Т. (ноябрь 2011 г.). «Организация мозжечка человека, оцениваемая по внутренней функциональной связности» . Журнал нейрофизиологии . 106 (5): 2322–45. дои : 10.1152/jn.00339.2011 . ПМЦ 3214121 . ПМИД 21795627 .
- ^ Эмоциональный мозжечок . Михаэль Адамашек, Марио Манто, Деннис Дж.Л.Г. Шуттер. Чам, Швейцария: Springer. 2022. ISBN 978-3-030-99550-8 . OCLC 1338132789 .
{{cite book}}: CS1 maint: другие ( ссылка ) - ^ Эрнаес-Гони П., Тирапу-Устаррос Х., Иглесиас-Фернандес Л., Луна-Ларио П. (ноябрь 2010 г.). «Участие мозжечка в регуляции привязанностей, эмоций и поведения» [Роль мозжечка в регуляции привязанностей, эмоций и поведения]. Журнал неврологии (на испанском языке). 51 (10): 597–609. дои : 10.33588/rn.5110.2010394 . ПМИД 21069639 .
- ^ Тернер Б.М., Парадизо С., Марвел CL, Пирсон Р., Болес Понто Л.Л., Хичва Р.Д., Робинсон Р.Г. (март 2007 г.). «Мозжечок и эмоциональный опыт» . Нейропсихология . 45 (6): 1331–41. doi : 10.1016/j.neuropsychologia.2006.09.023 . ПМЦ 1868674 . ПМИД 17123557 .
- ^ Дойя К. (октябрь 1999 г.). «Каковы вычисления мозжечка, базальных ганглиев и коры головного мозга?». Нейронные сети . 12 (7–8): 961–974. дои : 10.1016/S0893-6080(99)00046-5 . ПМИД 12662639 .
- ^ Манто М, Марвел С, Вандерверт Л (2022). Новая революция в психологии и нейронауках . Швейцария: Природа Спрингера. ISBN 9783031060922 .
- ^ Экклс Дж.К., Ито М., Сентаготай Дж. (1967). Мозжечок как нейронная машина . Спрингер-Верлаг.
- ^ Мозжечок как нейронная машина , с. 311
- ^ Перейти обратно: а б Бойден Э.С., Като А., Раймонд Дж.Л. (2004). «Обучение, зависящее от мозжечка: роль множественных механизмов пластичности». Ежегодный обзор неврологии . 27 : 581–609. дои : 10.1146/annurev.neuro.27.070203.144238 . ПМИД 15217344 . S2CID 1310007 .
- ^ Перейти обратно: а б Хоук Дж.К., Бэкингем Дж.Т., Барто А.Г. (1996). «Модели мозжечка и двигательного обучения» (PDF) . Поведение. Наука о мозге . 19 (3): 368–383. CiteSeerX 10.1.1.118.2997 . дои : 10.1017/S0140525X00081474 . Архивировано из оригинала (PDF) 9 августа 2017 г.
- ^ Фудзита М (1982). «Модель адаптивного фильтра мозжечка». Биологическая кибернетика . 45 (3): 195–206. дои : 10.1007/BF00336192 . ПМИД 7171642 . S2CID 3695770 .
- ^ Гилберт П.Ф., Тах В.Т. (июнь 1977 г.). «Активность клеток Пуркинье во время двигательного обучения». Исследования мозга . 128 (2): 309–28. дои : 10.1016/0006-8993(77)90997-0 . ПМИД 194656 . S2CID 40799652 .
- ^ Кристиан К.М., Томпсон РФ (2003). «Нейральные субстраты кондиционирования глазного моргания: приобретение и удержание» . Обучение и память . 10 (6): 427–55. дои : 10.1101/lm.59603 . ПМИД 14657256 .
- ^ Брайтенберг В., Этвуд Р.П. (февраль 1958 г.). «Морфологические наблюдения за корой мозжечка». Журнал сравнительной неврологии . 109 (1): 1–33. дои : 10.1002/cne.901090102 . ПМИД 13563670 . S2CID 8989536 .
- ^ Брайтенберг В., Хек Д., Султан Ф. (июнь 1997 г.). «Обнаружение и генерация последовательностей как ключ к функции мозжечка: эксперименты и теория». Поведенческие и мозговые науки . 20 (2): 229–45, обсуждение 245–77. дои : 10.1017/s0140525x9700143x . ПМИД 10096998 . S2CID 36802745 .
- ^ Иври Р.Б., Спенсер Р.М., Желязник Х.Н., Дидрихсен Дж. (декабрь 2002 г.). «Мозжечок и время событий» . Анналы Нью-Йоркской академии наук . 978 (1): 302–17. Бибкод : 2002NYASA.978..302I . дои : 10.1111/j.1749-6632.2002.tb07576.x . ПМИД 12582062 . S2CID 27237058 .
- ^ Пеллионис А., Ллинас Р. (1982). «Представление пространства-времени в мозге. Мозжечок как прогнозирующий метрический тензор пространства-времени». Нейронаука . 7 (12): 2949–70. дои : 10.1016/0306-4522(82)90224-X . ПМИД 7162624 . S2CID 20520737 .
- ^ Хорват Г (2003). «CMAC: пересмотр старой нейронной сети» (PDF) . Интеллектуальные системы управления и обработки сигналов . Проверено 24 декабря 2009 г.
- ^ Шмитц Т.Дж. (2007). «Экспертиза координации». В О'Салливан С.Б., Шмитц Т.Дж. (ред.). Физическая реабилитация . Филадельфия: Ф.А. Дэвис. стр. 193–225 . ISBN 9780803612471 .
- ^ Мариен П., Манто М (2016). Лингвистический мозжечок . Лондон, Великобритания: Академическая пресса. стр. 337–351. ISBN 978-0-12-801608-4 .
- ^ Гилман С. (март 1998 г.). «Изображение мозга. Вторая из двух частей». Медицинский журнал Новой Англии . 338 (13): 889–96. дои : 10.1056/NEJM199803263381307 . ПМИД 9516225 .
- ^ Перейти обратно: а б «Информационная страница NINDS атаксии и мозжечковой или спиноцеребеллярной дегенерации» . Национальные институты здравоохранения. 16 апреля 2014 года. Архивировано из оригинала 9 февраля 2015 года . Проверено 2 февраля 2015 г.
- ^ Юхас Д (15 января 2016 г.). «Ветераны Ирака и Афганистана демонстрируют изменения в мозге, связанные с воздействием взрыва» . Научный американец . Архивировано из оригинала 20 января 2016 года . Проверено 21 января 2016 г.
- ^ Винсент М., Хаджихани Н. (июнь 2007 г.). «Мозжечок и мигрень» . Головная боль . 47 (6): 820–33. дои : 10.1111/j.1526-4610.2006.00715.x . ПМК 3761082 . ПМИД 17578530 .
- ^ Перейти обратно: а б «Информационная страница NINDS о дегенерации мозжечка» . Национальные институты здравоохранения. 28 февраля 2014 г. Архивировано из оригинала 18 февраля 2015 г. Проверено 2 февраля 2015 г.
- ^ Хорват С., Мах В., Лу А.Т., Ву Дж.С., Чой О.В., Ясинска А.Дж., Рианчо Дж.А., Тунг С., Коулз Н.С., Браун Дж., Винтерс Х.В., Коулз Л.С. (май 2015 г.). «Мозжечок стареет медленно в соответствии с эпигенетическими часами» . Старение . 7 (5): 294–306. дои : 10.18632/aging.100742 . ПМЦ 4468311 . ПМИД 26000617 .
- ^ Фрейзер Х.Б., Хайтович П., Плоткин Дж.Б., Паабо С. , Эйзен М.Б. (сентябрь 2005 г.). «Старение и экспрессия генов в мозгу приматов» . ПЛОС Биология . 3 (9): е274. doi : 10.1371/journal.pbio.0030274 . ПМЦ 1181540 . ПМИД 16048372 .
- ^ Андерсен Б.Б., Гундерсен Х.Дж., Паккенберг Б. (ноябрь 2003 г.). «Старение мозжечка человека: стереологическое исследование». Журнал сравнительной неврологии . 466 (3): 356–65. дои : 10.1002/cne.10884 . ПМИД 14556293 . S2CID 7091227 .
- ^ Раз Н., Ганнинг-Диксон Ф., Хед Д., Уильямсон А., Акер Дж.Д. (2001). «Возрастные и половые различия в мозжечке и вентральном мосту: проспективное МРТ-исследование здоровых взрослых» (PDF) . Американский журнал нейрорадиологии . 22 (6): 1161–7. ПМЦ 7974784 . ПМИД 11415913 . Архивировано (PDF) из оригинала 17 декабря 2008 г.
- ^ Перейти обратно: а б с д Альберт Р.К., Портер Р.С., ред. (2006). Руководство Merck по диагностике и терапии (18-е изд.). Станция Уайтхаус, Нью-Джерси: Исследовательские библиотеки Merck. стр. 1886–1887.
- ^ Кэмпбелл Дж., Гилберт В.М., Николаидес К.Х., Кэмпбелл С. (август 1987 г.). «Ультразвуковой скрининг расщелины позвоночника: краниальные и мозжечковые признаки в группе высокого риска». Акушерство и гинекология . 70 (2): 247–50. ПМИД 3299184 .
- ^ Перейти обратно: а б Хаттен М.Е., Хайнц Н. (1995). «Механизмы формирования и спецификации нейронных паттернов в развивающемся мозжечке». Ежегодный обзор неврологии . 18 : 385–408. дои : 10.1146/annurev.ne.18.030195.002125 . ПМИД 7605067 .
- ^ Полкингхорн WR, Тарбелл, штат Нью-Джерси (май 2007 г.). «Медуллобластома: туморогенез, современная клиническая парадигма и усилия по улучшению стратификации риска». Природная клиническая практика. Онкология . 4 (5): 295–304. дои : 10.1038/ncponc0794 . ПМИД 17464337 . S2CID 24461280 .
- ^ Руссель М.Ф., Хаттен М.Э. (2011). Развитие мозжечка и медуллобластома . Том. 94. стр. 101-1. 235–82. дои : 10.1016/B978-0-12-380916-2.00008-5 . ISBN 9780123809162 . ПМЦ 3213765 . ПМИД 21295689 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ «Информационная страница о синдроме NINDS Жубера» . Национальные институты здравоохранения. 23 декабря 2013 года. Архивировано из оригинала 4 января 2015 года . Проверено 9 января 2015 г.
- ^ «Информационная страница NINDS Денди-Уокер» . Национальные институты здравоохранения. 14 февраля 2014 года. Архивировано из оригинала 4 января 2015 года . Проверено 9 января 2015 г.
- ^ «Информационная страница NINDS о гипоплазии мозжечка» . Национальные институты здравоохранения. 29 сентября 2011 года. Архивировано из оригинала 4 января 2015 года . Проверено 9 января 2015 г.
- ^ «Информационный бюллетень по мальформации Киари» . Национальные институты здравоохранения. 10 декабря 2014 года. Архивировано из оригинала 27 октября 2011 года . Проверено 9 января 2015 г.
- ^ «Информационная страница NINDS Dyssynergia Cerebellaris Myoclonica» . Национальные институты здравоохранения. 14 февраля 2011 года. Архивировано из оригинала 16 февраля 2015 года . Проверено 1 февраля 2015 г.
- ^ «Информационная страница NINDS оливопонтоцеребеллярной атрофии» . Национальные институты здравоохранения. 16 апреля 2014 года. Архивировано из оригинала 27 января 2012 года . Проверено 9 января 2015 г.
- ^ «Информационная страница NINDS о паранеопластических синдромах» . Национальные институты здравоохранения. 12 марта 2009 г. Архивировано из оригинала 4 января 2015 г. . Проверено 9 января 2015 г.
- ^ «Информационная страница NINDS о синдроме Вернике-Корсакова» . Национальные институты здравоохранения. 14 февраля 2007 г. Архивировано из оригинала 4 января 2015 г. Проверено 9 января 2015 г.
- ^ Моултон Э.А., Шмахманн Дж.Д., Бесерра Л., Борсук Д. (октябрь 2010 г.). «Мозжечок и боль: пассивный интегратор или активный участник?» . Обзоры исследований мозга . 65 (1): 14–27. дои : 10.1016/j.brainresrev.2010.05.005 . ПМК 2943015 . ПМИД 20553761 .
- ^ Бауманн О, Борра Р.Дж., Бауэр Дж.М., Каллен К.Е., Хабас С., Иври Р.Б., Леггио М., Маттингли Дж.Б., Молинари М., Моултон Э.А., Полин М.Г., Павлова М.А., Шмахманн Дж.Д., Соколов А.А. (апрель 2015 г.). «Консенсусный документ: роль мозжечка в процессах восприятия» . Мозжечок . 14 (2): 197–220. дои : 10.1007/s12311-014-0627-7 . ПМЦ 4346664 . ПМИД 25479821 .
- ^ Перейти обратно: а б с д Bell CC, Хан В., Sawtell NB (2008). «Структуры, подобные мозжечку, и их значение для функции мозжечка». Ежегодный обзор неврологии . 31 : 1–24. дои : 10.1146/annurev.neuro.30.051606.094225 . ПМИД 18275284 . S2CID 14536411 .
- ^ Вудхэмс PL (июль 1977 г.). «Ультраструктура аналога мозжечка у осьминога». Журнал сравнительной неврологии . 174 (2): 329–45. дои : 10.1002/cne.901740209 . ПМИД 864041 . S2CID 43112389 .
- ^ Перейти обратно: а б Ромер А.С., Парсонс Т.С. (1977). Тело позвоночного . Филадельфия: Холт-Сондерс Интернэшнл. п. 531. ИСБН 978-0-03-910284-5 .
- ^ Ши З, Чжан Ю, Мик Дж, Цяо Дж, Хан ВЗ (август 2008 г.). «Нейрональная организация уникальной специализации мозжечка: клапан мозжечка мормиридовой рыбы» . Журнал сравнительной неврологии . 509 (5): 449–73. дои : 10.1002/cne.21735 . ПМЦ 5884697 . ПМИД 18537139 .
- ^ Уивер А.Х. (март 2005 г.). «Взаимная эволюция мозжечка и неокортекса у ископаемых людей» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (10): 3576–80. Бибкод : 2005PNAS..102.3576W . дои : 10.1073/pnas.0500692102 . ПМК 553338 . ПМИД 15731345 .
- ^ Перейти обратно: а б с Шенеманн П.Т. (1 декабря 2009 г.). «Эволюция мозга и языка». Изучение языка . 59 : 162–186. дои : 10.1111/j.1467-9922.2009.00539.x . ПМИД 22230641 .
- ^ Маклеод С.Э., Зиллес К., Шлейхер А., Риллинг Дж.К., Гибсон К.Р. (апрель 2003 г.). «Расширение неоцеребеллума у Hominoidea». Журнал эволюции человека . 44 (4): 401–29. Бибкод : 2003JHumE..44..401M . дои : 10.1016/S0047-2484(03)00028-9 . ПМИД 12727461 .
- ^ Робертс П.Д., Портфорс CV (июнь 2008 г.). «Принципы проектирования сенсорной обработки в мозжечковоподобных структурах. Ранняя стадия обработки электросенсорных и слуховых объектов». Биологическая кибернетика . 98 (6): 491–507. дои : 10.1007/s00422-008-0217-1 . ПМИД 18491162 . S2CID 14393814 .
- ^ Бауэр Дж. М. (1997). «Глава 27 — сенсорика мозжечка ради моторики или моторика ради сенсорики: вид из усов крысы?». Мозжечок: от структуры к контролю . Прогресс в исследованиях мозга. Том. 114. стр. 463–96. дои : 10.1016/S0079-6123(08)63381-6 . ISBN 978-0-444-82313-7 . ПМИД 9193161 .
- ^ Хейни С.А., Ким Дж., Августин Г.Дж., Медина Дж.Ф. (февраль 2014 г.). «Точный контроль кинематики движения путем оптогенетического торможения активности клеток Пуркинье» . Журнал неврологии . 34 (6): 2321–30. doi : 10.1523/JNEUROSCI.4547-13.2014 . ПМЦ 3913874 . ПМИД 24501371 .
- ^ Виттер Л., Канто CB, Хугланд ТМ, де Груйл-младший, Де Зеув CI (2013). «Сила и время двигательных реакций, опосредованных отскоком в ядрах мозжечка после активации клеток Пуркинье» . Границы в нейронных цепях . 7 : 133. doi : 10.3389/fncir.2013.00133 . ПМЦ 3748751 . ПМИД 23970855 .
- ^ Кларк Э., компакт-диск О'Мэлли (1996). «Глава 11: Мозжечок». Человеческий мозг и спинной мозг (2-е изд.). Норман Паблишинг. п. 629. ИСБН 978-0-930405-25-0 .
- ^ Ито М. (декабрь 2002 г.). «Исторический обзор значения мозжечка и роли клеток Пуркинье в моторном обучении». Анналы Нью-Йоркской академии наук . 978 (1): 273–88. Бибкод : 2002NYASA.978..273I . дои : 10.1111/j.1749-6632.2002.tb07574.x . ПМИД 12582060 . S2CID 22860609 .
- ^ Льюис CT, Short C (1879). Латинский словарь, основанный на латинском словаре Фрейнда, изданном Эндрюсом . Оксфорд: Кларендон Пресс.
- ^ Маршалл Л.Х., Магун Х.В. (1998). Открытия в человеческом мозге. Предыстория нейронауки, структура и функции мозга . Тотова: Humana Press.
- ^ Перейти обратно: а б с Фостер Ф.Д. (1891). Иллюстрированный медицинский словарь . Нью-Йорк: Д. Эпплтон и компания.
- ^ Краус Л.А. (1844). Критико-этимологический медицинский лексикон (третье издание) . Геттинген: Издательство книжного магазина Дойерлих и Дитрих.
- ^ Шрегер CH (1805). Анатомическая синонимия. Синонимы анатомической номенклатуры . Фюрт.
Внешние ссылки
[ редактировать ]- Ллинас Р., Негрелло М.Н. (2015). «Мозжечок» . Схоларпедия . 10 (1): 4606. Бибкод : 2015SchpJ..10.4606L . doi : 10.4249/scholarpedia.4606 .
- База данных, ориентированная на клетки мозжечка
- Справочник по мозжечку и мозжечковым расстройствам – Манто М., Груол Д.Л., Шмахманн Дж., Койбучи Н., Росси Ф. (ред.) – Спрингер – Нью-Йорк
- Окрашенные изображения срезов мозга, включающие «мозжечок», в проекте BrainMaps.
- Неполный мозг мужчины раскрывает роль мозжечка в мышлении и эмоциях
- Женщина, живущая без мозжечка
- Основы мозжечка и мозжечковых расстройств . Груол Д.Л., Койбучи Н., Манто М., Молинари М., Шмахманн Д.Д., Шен Ю. (ред.). Спрингер, Нью-Йорк, 2016 г.
- Гистологические изображения мозжечка
- Мозжечок - Журнал (Springer Nature)
- Мозжечок и атаксия - журнал (BioMed Central)