Альфа-спираль

Альфа -спираль (или α-спираль ) — это последовательность аминокислот в белке, закрученная в клубок ( спираль ).
Альфа-спираль — наиболее распространенное структурное устройство структуры белков вторичной . Это также самый крайний тип локальной структуры, и именно эту локальную структуру легче всего предсказать по последовательности аминокислот.
Альфа-спираль имеет конформацию правосторонней спирали , в которой каждая N-H группа основной цепи связана водородными связями с группой C=O основной цепи аминокислоты , которая находится на четыре остатка раньше в белковой последовательности.
Другие имена
[ редактировать ]Альфа-спираль также обычно называют:
- α-спираль Полинга-Кори-Брэнсона (по именам трех ученых, описавших ее структуру)
- 3,6 13 -спираль , так как в одном кольце 3,6 аминокислот, при этом в кольце, образованном водородной связью, участвует 13 атомов (начиная с амидного водорода и заканчивая карбонильным кислородом)

Открытие
[ редактировать ]

В начале 1930-х годов Уильям Эстбери показал, что происходят резкие изменения в дифракции рентгеновских лучей влажной шерсти или волокон волос при значительном растяжении. Данные показали, что нерастянутые волокна имели спиральную молекулярную структуру с характерным повторением ≈5,1 ангстрема (0,51 нанометра ).
Первоначально Эстбери предложил структуру волокон со связанными цепями. Позже он присоединился к другим исследователям (в частности, к американскому химику Морису Хаггинсу ) и предположил, что:
- нерастянутые молекулы белка образовали спираль (которую он назвал α-формой)
- растяжение привело к раскручиванию спирали, образуя растянутое состояние (которое он назвал β-формой).
Хотя и неверные в своих деталях, модели этих форм Эстбери были верны по своей сути и соответствуют современным элементам вторичной структуры , α-спирали и β-цепи (номенклатура Эстбери была сохранена), которые были разработаны Лайнусом Полингом , Робертом Кори и Герман Брэнсон в 1951 году (см. ниже); в этой статье были показаны как правые, так и левые спирали, хотя в 1960 году кристаллическая структура миоглобина [ 1 ] показал, что наиболее распространенной является правосторонняя форма. Ганс Нейрат был первым, кто показал, что модели Эстбери не могут быть точными в деталях, поскольку они включают столкновения атомов. [ 2 ] Статья Нейрата и данные Эстбери вдохновили Г. С. Тейлора . [ 3 ] Морис Хаггинс [ 4 ] и Брэгг и сотрудники [ 5 ] предложить модели кератина , чем-то напоминающие современную α-спираль.
Двумя ключевыми достижениями в моделировании современной α-спирали были: правильная геометрия связей благодаря структуры аминокислот определению кристаллической и пептидов и предсказанию Полинга о плоских пептидных связях ; и его отказ от предположения о целом числе остатков на виток спирали. Поворотный момент наступил ранней весной 1948 года, когда Полинг простудился и лег спать. От скуки он нарисовал на полоске бумаги полипептидную цепь примерно правильных размеров и сложил ее в спираль, стараясь сохранить плоские пептидные связи. После нескольких попыток он создал модель с физически вероятными водородными связями. Затем Полинг работал с Кори и Брэнсоном, чтобы подтвердить свою модель перед публикацией. [ 6 ] В 1954 году Полинг получил свою первую Нобелевскую премию «за исследование природы химической связи и ее применение для выяснения структуры сложных веществ». [ 7 ] (например, белки), в первую очередь включая структуру α-спирали.
Структура
[ редактировать ]Геометрия и водородная связь
[ редактировать ]Аминокислоты в α-спирали расположены в виде правосторонней спиральной структуры, где каждый аминокислотный остаток соответствует повороту спирали на 100° (т. е. спираль имеет 3,6 остатков на виток) и трансляции 1,5 Å ( 0,15 нм) вдоль оси спирали. Дуниц [ 8 ] описывает, как первая статья Полинга на эту тему фактически показывает левую спираль, энантиомер истинной структуры. Короткие отрезки левой спирали иногда встречаются с большим содержанием ахиральных аминокислот глицина , но неблагоприятны для других нормальных, биологических L -аминокислот . Шаг альфа-спирали (расстояние по вертикали между последовательными витками спирали) составляет 5,4 Å (0,54 нм), что является произведением 1,5 и 3,6. Самое главное, что группа NH одной аминокислоты образует водородную связь с группой C=O аминокислоты четырьмя остатками раньше; эта повторяющаяся водородная связь i + 4 → i является наиболее заметной характеристикой α-спирали. Официальная международная номенклатура [ 9 ] [ 10 ] определяет два способа определения α-спиралей: правило 6.2 с точки зрения повторяющихся углов скручивания φ , ψ (см. ниже) и правило 6.3 с точки зрения комбинированного рисунка смолы и водородных связей. Альфа-спирали можно идентифицировать в структуре белка с помощью нескольких вычислительных методов, таких как DSSP (Определение вторичной структуры белка). [ 11 ]
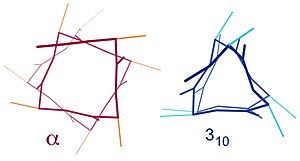
К подобным структурам относятся 3 10 спираль ( водородная связь i + 3 → i ) и π-спираль ( водородная связь i + 5 → i ). α-спираль можно охарактеризовать как спираль 3,6 13 , поскольку расстояние i + 4 добавляет к петле с Н-связями еще три атома по сравнению с более плотной спиралью 3 10 , и в среднем в одном кольце участвуют 3,6 аминокислоты. α-спираль. Нижние индексы относятся к числу атомов (включая водород) в замкнутом контуре, образованном водородной связью. [ 12 ]

Остатки в α-спиралях обычно принимают основной цепи ( φ , ψ ) двугранные углы вокруг (-60 °, -45 °), как показано на изображении справа. В более общих терминах они принимают двугранные углы так, что сумма двугранного угла ψ одного остатка и φ двугранного угла следующего остатка составляет примерно -105 °. Как следствие, α-спиральные двугранные углы, как правило, попадают на диагональную полосу на диаграмме Рамачандрана (с наклоном -1) в диапазоне от (-90 °, -15 °) до (-70 °, -35 °). . сумма двугранных углов Для сравнения: для спирали 3 10 составляет примерно −75°, а для π-спирали примерно −130°. Общая формула для угла поворота Ω, приходящегося на остаток любой полипептидной спирали с транс -изомерами, определяется уравнением [ 14 ] [ 15 ]
- 3 потому что Ω = 1 - 4 потому что 2 φ + ψ / 2
α-спираль плотно упакована; внутри спирали почти нет свободного пространства. Боковые цепи аминокислот находятся снаружи спирали и направлены примерно «вниз» (т. е. к N-концу), как ветви вечнозеленого дерева ( рождественской елки эффект ). Эта направленность иногда используется в предварительных картах электронной плотности с низким разрешением для определения направления основной цепи белка. [ 16 ]
Стабильность
[ редактировать ]Спирали, наблюдаемые в белках, могут иметь длину от четырех до более сорока остатков, но типичная спираль содержит около десяти аминокислот (около трех витков). В общем, короткие полипептиды не обладают значительной α-спиральной структурой в растворе, поскольку энтропийные затраты, связанные со сворачиванием полипептидной цепи, не компенсируются достаточным количеством стабилизирующих взаимодействий. В общем, основные водородные связи α-спиралей считаются немного более слабыми, чем те, которые обнаружены в β-листах , и легко подвергаются атаке со стороны молекул окружающей воды. Однако в более гидрофобных средах, таких как плазматическая мембрана , или в присутствии сорастворителей, таких как трифторэтанол (ТФЭ), или изолированных от растворителя в газовой фазе, [ 17 ] олигопептиды легко принимают стабильную α-спиральную структуру. Кроме того, поперечные связи могут быть включены в пептиды для конформационной стабилизации спиральных складок. Поперечные связи стабилизируют спиральное состояние, энтропийно дестабилизируя развернутое состояние и удаляя энтальпийно стабилизированные «ловушки» складки, которые конкурируют с полностью спиральным состоянием. [ 18 ] Было показано, что α-спирали более стабильны, устойчивы к мутациям и поддаются конструированию, чем β-цепи в природных белках. [ 19 ] а также в искусственно созданных белках. [ 20 ]

Визуализация
[ редактировать ]Тремя наиболее популярными способами визуализации альфа-спиральной вторичной структуры олигопептидных последовательностей являются (1) спиральное колесо , [ 21 ] (2) диаграмма Вэньсян, [ 22 ] и (3) спиральная сеть. [ 23 ] Каждый из них можно визуализировать с помощью различных пакетов программного обеспечения и веб-серверов. Чтобы создать небольшое количество диаграмм, Heliquest [ 24 ] может использоваться для винтовых колес и NetWheels [ 25 ] может использоваться для винтовых колес и винтовых сеток. Чтобы программно создать большое количество диаграмм, helixvis [ 26 ] [ 27 ] может использоваться для рисования винтовых колес и диаграмм Вэньсян на языках программирования R и Python.
Экспериментальное определение
[ редактировать ]Поскольку α-спираль определяется ее водородными связями и конформацией основной цепи, наиболее подробные экспериментальные доказательства α-спиральной структуры получены с помощью рентгеновской кристаллографии с атомным разрешением , как в примере, показанном справа. Ясно, что все карбонильные кислороды основной цепи направлены вниз (к С-концу), но слегка расширяются, а Н-связи примерно параллельны оси спирали. Белковые структуры, полученные методом ЯМР-спектроскопии, также хорошо демонстрируют спиральные структуры с характерными наблюдениями ядерного эффекта Оверхаузера (NOE) между атомами на соседних витках спирали. В некоторых случаях отдельные водородные связи можно наблюдать непосредственно как небольшую скалярную связь в ЯМР.
Существует несколько методов с более низким разрешением для определения общей спиральной структуры. ЯМР Химические сдвиги (в частности, C а , С б и C') и остаточные диполярные связи часто характерны для спиралей. спиралей в дальнем УФ-диапазоне (170–250 нм) Спектр кругового дихроизма также является своеобразным, демонстрируя выраженный двойной минимум при длинах волн около 208 и 222 нм. Инфракрасная спектроскопия используется редко, поскольку α-спиральный спектр напоминает спектр случайной катушки (хотя их можно различить, например, с помощью водородно-дейтериевого обмена ). Наконец, криоэлектронная микроскопия теперь способна различать отдельные α-спирали внутри белка, хотя их отнесение к остаткам все еще остается активной областью исследований.
Длинные гомополимеры аминокислот, если они растворимы, часто образуют спирали. Такие длинные изолированные спирали также можно обнаружить другими методами, такими как диэлектрическая релаксация , двойное лучепреломление потока и измерение константы диффузии . Более строго говоря, эти методы обнаруживают только характерную вытянутую (длинную сигару) гидродинамическую форму спирали или ее большой дипольный момент .
Склонность к аминокислотам
[ редактировать ]Различные аминокислотные последовательности имеют разную склонность к образованию α-спиральной структуры. Метионин , аланин , лейцин , глутамат и лизин незаряженный («МАЛЕК» в однобуквенном коде аминокислот ) имеют особенно высокую склонность к образованию спирали, тогда как пролин и глицин имеют плохую склонность к образованию спирали. [ 28 ] Пролин либо разрывает, либо перекручивает спираль, как потому, что он не может отдавать амидную водородную связь (не имея амидного водорода), так и потому, что его боковая цепь стерически взаимодействует с основной цепью предыдущего витка - внутри спирали это приводит к изгибу примерно на 30 градусов. ° по оси спирали. [ 12 ] Однако пролин часто рассматривают как первый остаток спирали, предположительно из-за его структурной жесткости. С другой стороны, глицин также имеет тенденцию разрушать спирали, поскольку его высокая конформационная гибкость делает энтропийно дорогостоящим принятие относительно ограниченной α-спиральной структуры.
Таблица склонностей стандартных аминокислот к альфа-спирали
[ редактировать ]Оценочные различия в изменении свободной энергии Δ(Δ G ), оцененные в ккал/моль на остаток в α-спиральной конфигурации, относительно аланина, произвольно установленного как ноль. Более высокие числа (более положительные изменения свободной энергии) менее предпочтительны. Возможны значительные отклонения от этих средних чисел в зависимости от идентичности соседних остатков.
Различия в изменении свободной энергии на остаток [ 29 ] Аминокислота 3-
письмо1-
письмоСпиральный штраф ккал/моль кДж/моль Аланин Земля А 0.00 0.00 Аргинин Арг Р 0.21 0.88 Аспарагин Асн Н 0.65 2.72 Аспарагиновая кислота Асп Д 0.69 2.89 Цистеин Цис С 0.68 2.85 Глутаминовая кислота Глу И 0.40 1.67 Глютамин Глн вопрос 0.39 1.63 Глицин Гли Г 1.00 4.18 Гистидин Его ЧАС 0.61 2.55 изолейцин С я 0.41 1.72 Лейцин Лео л 0.21 0.88 Лизин Свет К 0.26 1.09 Метионин Из М 0.24 1.00 Фенилаланин Пхе Ф 0.54 2.26 Пролин Про П 3.16 13.22 Серин Быть С 0.50 2.09 Треонин чр Т 0.66 2.76 Триптофан Трп В 0.49 2.05 Тирозин Тир И 0.53 2.22 Валин Вал V 0.61 2.55
Дипольный момент
[ редактировать ]Спираль обладает общим дипольным моментом, обусловленным совокупным действием отдельных микродиполей карбонильных групп пептидной связи, направленных вдоль оси спирали. [ 30 ] Эффекты этого макродиполя вызывают некоторые споры. α-спирали часто встречаются с N-концевым концом, связанным отрицательно заряженной группой, иногда боковой цепью аминокислоты, такой как глутамат или аспартат , или иногда фосфат-ионом. Некоторые считают, что спиральный макродиполь электростатически взаимодействует с такими группами. Другие считают, что это вводит в заблуждение, и более реалистично сказать, что потенциал водородной связи свободных групп NH на N-конце α-спирали может быть удовлетворен за счет водородной связи; это также можно рассматривать как совокупность взаимодействий между локальными микродиполями, такими как C=O···H-N . [ 31 ] [ 32 ]
Спиральные катушки
[ редактировать ]Спиральные α-спирали представляют собой высокостабильные формы, в которых две или более спиралей обвивают друг друга в структуре «суперспирали». Спиральные спирали содержат весьма характерный мотив последовательности, известный как гептадный повтор , в котором мотив повторяется каждые семь остатков последовательности ( аминокислотные остатки, а не пары оснований ДНК). Первый и особенно четвертый остатки (известные как положения a и d ) почти всегда гидрофобны ; четвертый остаток обычно представляет собой лейцин – отсюда и название структурного мотива, называемого лейциновой застежкой , который представляет собой разновидность спиральной спирали. Эти гидрофобные остатки собираются вместе внутри спиралевидного пучка. Как правило, пятый и седьмой остатки ( положения e и g ) имеют противоположные заряды и образуют солевой мостик, стабилизированный электростатическими взаимодействиями. Фиброзные белки, такие как кератин или «стебли» миозина или кинезина, часто принимают спирально-спиральную структуру, как и некоторые димеризующиеся белки. Пара витков – четырехспиральный пучок. – очень распространенный структурный мотив в белках. Например, он встречается в гормоне роста человека и некоторых разновидностях цитохрома . Белок Rop , который способствует репликации плазмиды у бактерий, представляет собой интересный случай, когда один полипептид образует спиральную спираль, а два мономера собираются в пучок из четырех спиралей.
Оформление лица
[ редактировать ]Аминокислоты, составляющие конкретную спираль, можно нанести на спиральное колесо — изображение, которое иллюстрирует ориентацию составляющих аминокислот ( см. в статье о лейциновой молнии такую диаграмму ). Часто в глобулярных белках , а также в специализированных структурах, таких как спиральные спирали и лейциновые застежки , α-спираль имеет две «грани»: одна содержит преимущественно гидрофобные аминокислоты, ориентированные внутрь белка, в гидрофобном ядре , и один, содержащий преимущественно полярные аминокислоты, ориентированные к растворителя поверхности белка, подвергающейся воздействию .
Изменения в ориентации связывания также происходят для олигопептидов с лицевой организацией. Эта закономерность особенно распространена в антимикробных пептидах , и было разработано множество моделей, чтобы описать, как это связано с их функцией. Общим для многих из них является то, что гидрофобная поверхность антимикробного пептида образует поры в плазматической мембране после связи с жирными цепями в ядре мембраны. [ 33 ] [ 34 ]
Крупномасштабные сборки
[ редактировать ]
Миоглобин и гемоглобин , первые два белка, структура которых была расшифрована с помощью рентгеновской кристаллографии , имеют очень похожие складки, примерно на 70% состоящие из α-спирали, а остальные представляют собой неповторяющиеся области или «петли», соединяющие спирали. При классификации белков по их доминантной складке база данных Структурной классификации белков поддерживает большую категорию, специально предназначенную для всех-α-белков.
Гемоглобин тогда имеет еще более масштабную четвертичную структуру , в которой функциональная молекула, связывающая кислород, состоит из четырех субъединиц.
Функциональные роли
[ редактировать ]

связывание ДНК
[ редактировать ]α-Спирали имеют особое значение в мотивах связывания ДНК , включая спираль-поворот-спираль мотивы , мотивы лейциновой молнии и мотивы цинковых пальцев . Это связано с тем удобным структурным фактом, что диаметр α-спирали составляет около 12 Å (1,2 нм), включая средний набор боковых цепей, что примерно равно ширине основной бороздки в ДНК B-формы , а также потому, что Димеры спиралей со спиральной спиралью (или лейциновой застежкой) могут легко расположить пару поверхностей взаимодействия так, чтобы они контактировали с своего рода симметричными повторами, обычными для двойной спирали ДНК. [ 35 ] Примером обоих аспектов является транскрипционный фактор Max (см. изображение слева), который использует для димеризации спиральную спираль, располагая еще одну пару спиралей для взаимодействия в двух последовательных витках большой бороздки ДНК.
Мембранный охват
[ редактировать ]α-Спирали также являются наиболее распространенным структурным элементом белка, который пересекает биологические мембраны ( трансмембранный белок ). [ 36 ] Предполагается, что спиральная структура может удовлетворять все водородные связи основной цепи внутри, не оставляя полярных групп, открытых для мембраны, если боковые цепи гидрофобны. Белки иногда закрепляются одной пронизывающей мембрану спиралью, иногда парой, а иногда пучком спиралей, наиболее классически состоящим из семи спиралей, расположенных вверх и вниз в кольце, например, для родопсинов (см. изображение справа) и другие рецепторы, связанные с G-белком (GPCR). Структурная стабильность между парами α-спиральных трансмембранных доменов зависит от консервативных мотивов мембранной межспиральной упаковки, например, мотива Glycine-xxx-Glycine (или small-xxx-small). [ 37 ]
Механические свойства
[ редактировать ]α-Спирали при осевой растягивающей деформации, характерном состоянии нагрузки, которое возникает во многих нитях и тканях, богатых альфа-спиралями, приводит к характерному трехфазному поведению касательного модуля жесткости-мягкости-жесткости. [ 38 ] Фаза I соответствует режиму малых деформаций, при котором спираль растягивается однородно, за ней следует фаза II, при которой витки альфа-спирали разрываются за счет разрыва групп Н-связей. Фаза III обычно связана с растяжением ковалентной связи с большой деформацией.
Динамические функции
[ редактировать ]Альфа-спирали в белках могут иметь низкочастотное движение, подобное аккордеону, что наблюдается с помощью рамановской спектроскопии. [ 39 ] и проанализированы с помощью модели квазиконтинуума. [ 40 ] [ 41 ] Спирали, не стабилизированные третичными взаимодействиями, демонстрируют динамическое поведение, которое в основном можно объяснить истиранием спирали с концов. [ 42 ]
Переход спираль-катушка
[ редактировать ]Гомополимеры аминокислот (таких как полилизин ) могут принимать α-спиральную структуру при низкой температуре, которая «расплавляется» при высоких температурах. Когда-то считалось, что этот переход спираль-клубок аналогичен денатурации белка . Статистическую механику этого перехода можно смоделировать с помощью элегантного метода матрицы переноса , характеризующегося двумя параметрами: склонностью к инициированию спирали и склонностью к растягиванию спирали.
В искусстве
[ редактировать ]
По крайней мере пять художников явно ссылались на α-спираль в своих работах: Джули Ньюдолл в живописи и Джулиан Восс-Андре , Батшеба Гроссман , Байрон Рубин и Майк Тыка в скульптуре.
Художница из Сан-Франциско Джули Ньюдолл, [ 43 ] имеет степень в области микробиологии со степенью в области искусства, с 1990 года специализируется на картинах, вдохновленных микроскопическими изображениями и молекулами. На ее картине «Восстание альфа-спирали» (2003) изображены человеческие фигуры, расположенные по α-спирали. По словам художника, «цветы отражают различные типы боковых цепей, которые каждая аминокислота несет миру». [ 43 ] Эта же метафора повторяется и со стороны ученого: «β-листы не демонстрируют жесткой повторяющейся регулярности, а текут изящными, извилистыми кривыми, и даже α-спираль правильнее, скорее, наподобие стебля цветка, узлы ветвления которого показывают влияние окружающей среды, истории развития и эволюции каждой части в соответствии со своей собственной идиосинкразической функцией». [ 12 ]
Юлиан Восс-Андре — скульптор немецкого происхождения, имеющий степень в области экспериментальной физики и скульптуры. С 2001 года Восс-Андре создает «белковые скульптуры». [ 44 ] основан на структуре белка, причем α-спираль является одним из его любимых объектов. Восс-Андреа создал скульптуры из α-спирали из различных материалов, включая бамбук и целые деревья. Памятник Восс-Андрее, созданный в 2004 году в честь Линуса Полинга , первооткрывателя α-спирали, выполнен из большой стальной балки, перестроенной в структуру α-спирали. Ярко-красная скульптура высотой 10 футов (3 м) стоит перед домом, где прошло детство Полинга, в Портленде, штат Орегон .
Ленточные диаграммы α-спиралей являются важным элементом кристаллических скульптур с лазерной гравировкой белковых структур, созданных художницей Батшебой Гроссман , таких как структуры инсулина , гемоглобина и ДНК-полимеразы . [ 45 ] Байрон Рубин — бывший кристаллограф белков, а теперь профессиональный скульптор по металлу белков, нуклеиновых кислот и молекул лекарств, многие из которых имеют α-спирали, такие как субтилизин , гормон роста человека и фосфолипаза А2 . [ 46 ]
Майк Тайка — вычислительный биохимик из Вашингтонского университета, работающий с Дэвидом Бейкером . Тыка занимается изготовлением скульптур из белковых молекул с 2010 года из меди и стали, включая убиквитин и тетрамер калиевых каналов . [ 47 ]
См. также
[ редактировать ]- 3 10 спираль
- Бета-лист
- Davydov soliton
- Складывание (химия)
- Ручки в дырочки, упаковка
- Пи-спираль
- Протеопедия Helices_in_Proteins
Ссылки
[ редактировать ]- ^ Кендрю Дж.К. , Дикерсон Р.Э., Страндберг Б.Е., Харт Р.Г., Дэвис Д.Р., Филлипс Д.К., Шор В.К. (февраль 1960 г.). «Структура миоглобина: трехмерный синтез Фурье с разрешением 2 Å». Природа . 185 (4711): 422–7. Бибкод : 1960Natur.185..422K . дои : 10.1038/185422a0 . ПМИД 18990802 . S2CID 4167651 .
- ^ Нейрат Х (1940). «Внутримолекулярное сворачивание полипептидных цепей в зависимости от структуры белка». Журнал физической химии . 44 (3): 296–305. дои : 10.1021/j150399a003 .
- ^ Тейлор Х.С. (1942). «Большие молекулы через атомные очки». Труды Американского философского общества . 85 (1): 1–12. JSTOR 985121 .
- ^ Хаггинс М (1943). «Строение волокнистых белков». Химические обзоры . 32 (2): 195–218. дои : 10.1021/cr60102a002 .
- ^ Брэгг В.Л. , Кендрю Дж.К. , Перуц М.Ф. (1950). «Конфигурации полипептидных цепей в кристаллических белках». Труды Лондонского королевского общества, серия A. 203 (1074): 321–?. Бибкод : 1950RSPSA.203..321B . дои : 10.1098/rspa.1950.0142 . S2CID 93804323 .
- ^ Полинг Л. , Кори Р.Б. , Брэнсон Х.Р. (апрель 1951 г.). «Строение белков; две спиральные конфигурации полипептидной цепи с водородными связями» . Труды Национальной академии наук Соединенных Штатов Америки . 37 (4): 205–11. Бибкод : 1951ПНАС...37..205П . дои : 10.1073/pnas.37.4.205 . ПМЦ 1063337 . ПМИД 14816373 .
- ^ «Нобелевская премия по химии 1954 года» .
- ^ Дуниц Дж . (2001). «Левая α-спираль Полинга». Angewandte Chemie, международное издание . 40 (22): 4167–4173. doi : 10.1002/1521-3773(20011119)40:22<4167::AID-ANIE4167>3.0.CO;2-Q . ПМИД 29712120 .
- ^ Комиссия IUPAC-IUB по биохимической номенклатуре (1970). «Сокращения и символы для описания конформации полипептидных цепей» . Журнал биологической химии . 245 (24): 6489–6497. дои : 10.1016/S0021-9258(18)62561-X .
- ^ «Конформации полипептидов 1 и 2» . www.sbcs.qmul.ac.uk. Проверено 5 ноября 2018 г.
- ^ Кабш В., Сандер С. (декабрь 1983 г.). «Словарь вторичной структуры белков: распознавание образов водородных связей и геометрических особенностей». Биополимеры . 22 (12): 2577–637. дои : 10.1002/bip.360221211 . ПМИД 6667333 . S2CID 29185760 .
- ^ Перейти обратно: а б с Ричардсон Дж.С. (1981). «Анатомия и систематика структуры белка». Достижения в области химии белков . 34 : 167–339. дои : 10.1016/S0065-3233(08)60520-3 . ISBN 9780120342341 . ПМИД 7020376 .
- ^ Ловелл С.С., Дэвис И.В., Арендалл В.Б., де Баккер П.И., Уорд Дж.М., Присант М.Г., Ричардсон Дж.С., Ричардсон Д.С. (февраль 2003 г.). «Проверка структуры с помощью геометрии Calpha: отклонение фи, пси и Cbeta». Белки . 50 (3): 437–50. дои : 10.1002/прот.10286 . ПМИД 12557186 . S2CID 8358424 .
- ^ Дикерсон Р.Э., Гейс I (1969), Структура и действие белков , Харпер, Нью-Йорк.
- ^ Зорко, Матяж (2010). «Структурная организация белков» . В Лангеле, Юло; Краватт, Бенджамин Ф .; Граслунд, Астрид; фон Хейне, Гуннар ; Земля, Зубы; Ниссен, Шерри; Зорко, Матяж (ред.). Введение в пептиды и белки . Бока-Ратон: CRC Press . стр. 100-1 36–57. ISBN 9781439882047 .
- ^ Тервиллигер TC (март 2010 г.). «Быстрое построение моделей альфа-спиралей на картах электронной плотности» . Acta Crystallographica Раздел D. 66 (Часть 3): 268–75. Бибкод : 2010AcCrD..66..268T . дои : 10.1107/S0907444910000314 . ПМЦ 2827347 . ПМИД 20179338 .
- ^ Хаджинс Р.Р., Джарролд М.Ф. (1999). «Образование спирали в несольватированных пептидах на основе аланина: спиральные мономеры и спиральные димеры». Журнал Американского химического общества . 121 (14): 3494–3501. дои : 10.1021/ja983996a .
- ^ Кучукян П.С., Ян Дж.С., Вердин Г.Л., Шахнович Е.И. (апрель 2009 г.). «Полноатомная модель стабилизации альфа-спиральной структуры пептидов углеводородными скрепками» . Журнал Американского химического общества . 131 (13): 4622–7. дои : 10.1021/ja805037p . ПМК 2735086 . ПМИД 19334772 .
- ^ Абрусан Г., Марш Дж.А. (2016). «Альфа-спирали более устойчивы к мутациям, чем бета-цепи» . PLOS Вычислительная биология . 12 (12): e1005242. Бибкод : 2016PLSCB..12E5242A . дои : 10.1371/journal.pcbi.1005242 . ПМК 5147804 . ПМИД 27935949 .
- ^ Роклин Г.Дж. и др. (2017). «Глобальный анализ сворачивания белков с использованием массово-параллельного проектирования, синтеза и тестирования» . Наука . 357 (6347): 168–175. Бибкод : 2017Sci...357..168R . дои : 10.1126/science.aan0693 . ПМЦ 5568797 . ПМИД 28706065 .
- ^ Шиффер М., Эдмундсон А.Б. (1967). «Использование спиральных колес для представления структур белков и идентификации сегментов со спиральным потенциалом» . Биофизический журнал . 7 (2): 121–135. Бибкод : 1967BpJ.....7..121S . дои : 10.1016/S0006-3495(67)86579-2 . ПМК 1368002 . ПМИД 6048867 .
- ^ Чжоу К.К., Чжан К.Т., Маджиора Г.М. (1997). «Расположение амфифильных спиралей в гетерополярных средах». Белки: структура, функции и генетика . 28 (1): 99–108. doi : 10.1002/(SICI)1097-0134(199705)28:1<99::AID-PROT10>3.0.CO;2-C . ПМИД 9144795 . S2CID 26944184 .
- ^ Даннилл П. (1968). «Использование спиральных сетевых диаграмм для представления белковых структур» . Биофизический журнал . 8 (7): 865–875. Бибкод : 1968BpJ.....8..865D . дои : 10.1016/S0006-3495(68)86525-7 . ПМЦ 1367563 . ПМИД 5699810 .
- ^ Готье Р., Дуге Д., Антонни Б., Дрин Г. (2008). «HELIQUEST: веб-сервер для просмотра последовательностей с особыми альфа-спиральными свойствами» . Биоинформатика . 24 (18): 2101–2102. doi : 10.1093/биоинформатика/btn392 . ПМИД 18662927 .
- ^ Мол А.Р., Кастро М.С., Фонтес В. (2018). «NetWheels: веб-приложение для создания высококачественных пептидных спиральных колес и чистых проекций». биоRxiv . дои : 10.1101/416347 . S2CID 92137153 .
- ^ Вадхва Р.Р., Субраманиан В., Стивенс-Трусс Р. (2018). «Визуализация альфа-спиральных пептидов в R с помощью helixvis» . Журнал программного обеспечения с открытым исходным кодом . 3 (31): 1008. Бибкод : 2018JOSS....3.1008W . дои : 10.21105/joss.01008 . S2CID 56486576 .
- ^ Субраманиан В., Вадхва Р.Р., Стивенс-Трусс Р. (2020). «Helixvis: визуализируйте альфа-спиральные пептиды в Python». ChemRxiv .
- ^ Пейс CN, Шольц Дж. М. (июль 1998 г.). «Шкала склонности к спирали, основанная на экспериментальных исследованиях пептидов и белков» . Биофизический журнал . 75 (1): 422–7. Бибкод : 1998BpJ....75..422N . дои : 10.1016/S0006-3495(98)77529-0 . ПМК 1299714 . ПМИД 9649402 .
- ^ Пейс, К. Ник; Шольц, Дж. Мартин (1998). «Шкала склонности к спирали, основанная на экспериментальных исследованиях пептидов и белков» . Биофизический журнал . 75 (1): 422–427. Бибкод : 1998BpJ....75..422N . дои : 10.1016/s0006-3495(98)77529-0 . ПМК 1299714 . ПМИД 9649402 .
- ^ Хол В.Г., ван Дуйнен П.Т., Берендсен Х.Дж. (1978). «Диполь альфа-спирали и свойства белков». Природа . 273 (5662): 443–446. Бибкод : 1978Natur.273..443H . дои : 10.1038/273443a0 . ПМИД 661956 . S2CID 4147335 .
- ^ Он JJ, Quiocho FA (октябрь 1993 г.). «Доминирующая роль локальных диполей в стабилизации некомпенсированных зарядов сульфата, секвестрированного в периплазматическом активном транспортном белке» . Белковая наука . 2 (10): 1643–7. дои : 10.1002/pro.5560021010 . ПМК 2142251 . ПМИД 8251939 .
- ^ Милнер-Уайт EJ (ноябрь 1997 г.). «Частичный заряд атома азота в пептидных связях» . Белковая наука . 6 (11): 2477–82. дои : 10.1002/pro.5560061125 . ПМК 2143592 . ПМИД 9385654 .
- ^ Кон, Эрик М.; Ширли, Дэвид Дж.; Ароцкий, Любовь; Пиччано, Анджела М.; Риджуэй, Закари; Урбан, Майкл В.; Кароне, Бенджамин Р.; Капуто, Грегори А. (04 февраля 2018 г.). «Роль катионных боковых цепей в антимикробной активности C18G» . Молекулы . 23 (2): 329. doi : 10,3390/molecules23020329 . ПМК 6017431 . ПМИД 29401708 .
- ^ Токе, Орсоля (2005). «Антимикробные пептиды: новые кандидаты в борьбе с бактериальными инфекциями» . Биополимеры . 80 (6): 717–735. дои : 10.1002/bip.20286 . ISSN 0006-3525 . ПМИД 15880793 .
- ^ Бранден и Туз, глава 10.
- ^ Бранден и Туз, глава 12.
- ^ Нэш А., Нотман Р., Диксон А.М. (2015). «Разработка de novo трансмембранных взаимодействий спираль-спираль и измерение стабильности биологической мембраны» . Biochimica et Biophysical Acta (BBA) – Биомембраны . 1848 (5): 1248–57. дои : 10.1016/j.bbamem.2015.02.020 . ПМИД 25732028 .
- ^ Акбароу Т., Чен X, Кетен С., Бюлер М.Дж. (октябрь 2007 г.). «Иерархии, множественные энергетические барьеры и надежность определяют механику разрушения альфа-спиральных и бета-листных белковых доменов» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (42): 16410–5. Бибкод : 2007PNAS..10416410A . дои : 10.1073/pnas.0705759104 . ПМК 2034213 . ПМИД 17925444 .
- ^ Художник ПК, Мошер Л.Е., Роудс С. (июль 1982 г.). «Низкочастотные моды в спектрах комбинационного рассеяния белков» . Биополимеры . 21 (7): 1469–72. дои : 10.1002/bip.360210715 . ПМИД 7115900 .
- ^ Чжоу К.С. (декабрь 1983 г.). «Идентификация низкочастотных мод в белковых молекулах» . Биохимический журнал . 215 (3): 465–9. дои : 10.1042/bj2150465 . ПМЦ 1152424 . ПМИД 6362659 .
- ^ Чжоу К.С. (май 1984 г.). «Биологические функции низкочастотных колебаний (фононов). III. Спиральные структуры и микроокружение» . Биофизический журнал . 45 (5): 881–9. Бибкод : 1984BpJ....45..881C . дои : 10.1016/S0006-3495(84)84234-4 . ПМЦ 1434967 . ПМИД 6428481 .
- ^ Фирц Б., Райнер А., Кифхабер Т. (январь 2009 г.). «Локальная конформационная динамика в альфа-спиралях, измеренная с помощью быстрого триплетного переноса» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (4): 1057–62. Бибкод : 2009PNAS..106.1057F . дои : 10.1073/pnas.0808581106 . ПМЦ 2633579 . ПМИД 19131517 .
- ^ Перейти обратно: а б «Джули Ньюдолл, вдохновленное наукой искусство, музыка, настольные игры» . www.brushwithscience.com . Проверено 06 апреля 2016 г.
- ^ Восс-Андре Дж (2005). «Белковые скульптуры: строительные блоки жизни вдохновляют искусство». Леонардо . 38 : 41–45. дои : 10.1162/leon.2005.38.1.41 . S2CID 57558522 .
- ^ Гроссман, Вирсавия. «О художнике» . Скульптура Вирсавии . Проверено 06 апреля 2016 г.
- ^ "О" . сайт молекулярной скульптуры.com . Проверено 06 апреля 2016 г.
- ^ Тыка, Майк. "О" . www.miketyka.com . Проверено 06 апреля 2016 г.
Дальнейшее чтение
[ редактировать ]- Туз Дж., Бренден С.И. (1999). Знакомство со структурой белка . Нью-Йорк: Паб Garland. ISBN 0-8153-2304-2 . .
- Айзенберг Д. (сентябрь 2003 г.). «Открытие альфа-спирали и бета-листа, основных структурных особенностей белков» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (20): 11207–10. Бибкод : 2003PNAS..10011207E . дои : 10.1073/pnas.2034522100 . ПМК 208735 . ПМИД 12966187 .
- Эстбери WT, Вудс HJ (1931). «Молекулярная масса белков». Природа . 127 (3209): 663–665. Бибкод : 1931Natur.127..663A . дои : 10.1038/127663b0 . S2CID 4133226 .
- Эстбери, WT, улица А (1931). «Рентгеновские исследования структуры волос, шерсти и родственных им волокон. I. Общие сведения» . Философские труды Лондонского королевского общества, серия A. 230 : 75–101. Бибкод : 1932РСПТА.230...75А . дои : 10.1098/rsta.1932.0003 .
- Эстбери WT (1933). «Некоторые проблемы рентгеновского анализа структуры волос животных и других белковых волокон». Пер. Фарадей Соц . 29 (140): 193–211. дои : 10.1039/tf9332900193 .
- Эстбери WT, Вудс HJ (1934). «Рентгеновские исследования структуры волос, шерсти и родственных им волокон. II. Молекулярная структура и упругие свойства кератина волос» . Философские труды Лондонского королевского общества, серия A. 232 (707–720): 333–394. Бибкод : 1934RSPTA.232..333A . дои : 10.1098/rsta.1934.0010 .
- Эстбери В.Т., Сиссон В.А. (1935). «Рентгеновские исследования структуры волос, шерсти и родственных им волокон. III. Конфигурация молекулы кератина и ее ориентация в биологической клетке» . Труды Лондонского королевского общества, серия A. 150 (871): 533–551. Бибкод : 1935RSPSA.150..533A . дои : 10.1098/rspa.1935.0121 .
- Сугета Х, Миядзава Т (1967). «Общий метод расчета спиральных параметров полимерных цепей на основе длин связей, углов связи и углов внутреннего вращения». Биополимеры . 5 (7): 673–679. дои : 10.1002/bip.1967.360050708 . S2CID 97785907 .
- Вада А (1976). «Альфа-спираль как электрический макродиполь». Достижения биофизики : 1–63. ПМИД 797240 .
- Чотия С., Левитт М., Ричардсон Д. (октябрь 1977 г.). «Строение белков: упаковка альфа-спиралей и складчатых листов» . Труды Национальной академии наук Соединенных Штатов Америки . 74 (10): 4130–4. Бибкод : 1977PNAS...74.4130C . дои : 10.1073/pnas.74.10.4130 . ПМК 431889 . ПМИД 270659 .
- Чотия С., Левитт М., Ричардсон Д. (январь 1981 г.). «Упаковка спирали в белки». Журнал молекулярной биологии . 145 (1): 215–50. дои : 10.1016/0022-2836(81)90341-7 . ПМИД 7265198 .
- Хол В.Г. (1985). «Роль диполя альфа-спирали в функции и структуре белка» . Прогресс биофизики и молекулярной биологии . 45 (3): 149–95. дои : 10.1016/0079-6107(85)90001-X . ПМИД 3892583 .
- Барлоу DJ, Торнтон JM (июнь 1988 г.). «Геометрия спирали в белках». Журнал молекулярной биологии . 201 (3): 601–19. дои : 10.1016/0022-2836(88)90641-9 . ПМИД 3418712 .
- Мурзин А.Г., Финкельштейн А.В. (декабрь 1988 г.). «Общая архитектура альфа-спиральной глобулы». Журнал молекулярной биологии . 204 (3): 749–69. дои : 10.1016/0022-2836(88)90366-X . ПМИД 3225849 .
