Корень

У сосудистых растений корни органы — это растения , которые модифицированы для обеспечения крепления растения и поступления воды и питательных веществ в тело растения, что позволяет растениям расти выше и быстрее. [1] Чаще всего они находятся ниже поверхности почвы , но корни могут быть и воздушными или аэрирующими, то есть растущими вверх над землей или особенно над водой. [2]
Функция
[ редактировать ]Основными функциями корней являются поглощение воды , питание растений и закрепление тела растения в земле. [3]
Анатомия
[ редактировать ]
Морфология корня разделена на четыре зоны: корневой чехлик, апикальную меристему , зону удлинения и волос. [4] Корневой чехлик новых корней помогает корню проникнуть в почву. Эти корневые чехлики отслаиваются по мере того, как корень проникает глубже, создавая слизистую поверхность, обеспечивающую смазку. Апикальная меристема за корневым чехликом производит новые клетки корня, которые удлиняются. Затем образуются корневые волоски, которые впитывают из почвы воду и минеральные питательные вещества. [5] Первым корнем у семенных растений является корешок , который разрастается из зародыша растения после прорастания семян.
При рассечении клетки корня располагаются следующим образом: корневые волоски , эпидермис , эпиблема , кора , эндодерма , перицикл и, наконец, сосудистая ткань в центре корня, обеспечивающая транспортировку поглощенной корнем воды в другие места корня. растение. [ нужны разъяснения ]
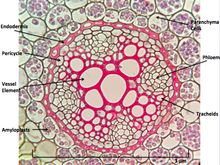
Возможно, самой поразительной характеристикой корней, которая отличает их от других органов растений, таких как стеблевые ветви и листья, является то, что корни имеют эндогенную структуру. [6] происхождения, т. е . они зарождаются и развиваются из внутреннего слоя материнской оси, например перицикла . [7] Напротив, стебли-ветви и листья экзогенны , т. е . они начинают развиваться из коры, внешнего слоя.
В ответ на концентрацию питательных веществ корни также синтезируют цитокинин , который действует как сигнал о том, насколько быстро могут расти побеги. Корни часто служат хранилищем пищи и питательных веществ. Корни большинства видов сосудистых растений вступают в симбиоз с некоторыми грибами , образуя микоризы многие другие организмы, включая бактерии . , а с корнями тесно связаны [8]

Архитектура корневой системы (RSA)
[ редактировать ]
Определение
[ редактировать ]В своей простейшей форме термин «архитектура корневой системы» (RSA) относится к пространственной конфигурации корневой системы растения. Эта система может быть чрезвычайно сложной и зависит от множества факторов, таких как вид самого растения, состав почвы и наличие питательных веществ. [9] Архитектура корня играет важную роль в обеспечении надежного снабжения питательными веществами и водой, а также фиксации и поддержки.
Конфигурация корневой системы служит для структурной поддержки растения, конкуренции с другими растениями и поглощения питательных веществ из почвы. [10] Корни растут в определенных условиях, изменение которых может затруднить рост растения. Например, корневая система, развившаяся в сухой почве, может быть не столь эффективной в затопленной почве, однако растения способны адаптироваться к другим изменениям окружающей среды, например, к сезонным изменениям. [10]
Условия и составляющие
[ редактировать ]Основными терминами, используемыми для классификации архитектуры корневой системы, являются: [11]
| Величина филиала | Количество ссылок (внешних или внутренних) |
| Топология | Схема ветвления ( Елочка , Дихотомическое , Радиальное ) |
| Длина ссылки | Расстояние между филиалами |
| Корневой угол | Радиальный угол основания бокового корня по окружности родительского корня, угол бокового корня относительно родительского корня и угол распространения всей системы. |
| Радиус связи | Диаметр корня |
Все компоненты корневой архитектуры регулируются посредством сложного взаимодействия между генетическими реакциями и реакциями на раздражители окружающей среды. Эти стимулы развития классифицируются как внутренние (генетические и пищевые влияния) или внешние (влияния окружающей среды) и интерпретируются путями передачи сигналов . [12]
Внешние факторы, влияющие на архитектуру корня, включают гравитацию, воздействие света, воды и кислорода, а также наличие или недостаток азота, фосфора, серы, алюминия и хлорида натрия. Основные гормоны (внутренние стимулы) и соответствующие пути, ответственные за развитие архитектуры корня, включают:
| Ауксин | Формирование латерального корня, сохранение апикального доминирования и образование придаточных корней. |
| Цитокинины | Цитокинины регулируют размер апикальной меристемы корня и способствуют боковому удлинению корня. |
| Этилен | Способствует формированию корончатых корней. |
| Гиббереллины | Вместе с этиленом они способствуют росту и удлинению зачатков кроны. Вместе с ауксином они способствуют удлинению корня. Гиббереллины также ингибируют зарождение зачатков боковых корней. |
Рост
[ редактировать ]
Ранний рост корня — одна из функций апикальной меристемы, расположенной вблизи кончика корня. Клетки меристемы более или менее непрерывно делятся, производя больше меристемы, клеток корневого чехлика (их приносят в жертву для защиты меристемы) и недифференцированных клеток корня. Последние становятся первичными тканями корня, сначала подвергаясь удлинению - процессу, который продвигает кончик корня вперед в среде выращивания. Постепенно эти клетки дифференцируются и созревают в специализированные клетки тканей корня. [13]
Рост из апикальных меристем известен как первичный рост , который включает в себя все удлинение. Вторичный рост включает в себя весь рост в диаметре, который является основным компонентом тканей древесных растений и многих недревесных растений. Например, запасающие корни сладкого картофеля имеют вторичный рост, но не являются деревянистыми. Вторичный рост происходит в латеральных меристемах , а именно в сосудистом камбии и пробковом камбии . Первый образует вторичную ксилему и вторичную флоэму , второй — перидерму .
У растений со вторичным ростом сосудистый камбий, берущий начало между ксилемой и флоэмой, образует цилиндр ткани вдоль стебля и корня. [ нужна ссылка ] Сосудистый камбий образует новые клетки как внутри, так и снаружи камбиевого цилиндра, причем те, которые находятся внутри, образуют вторичные клетки ксилемы, а те, которые находятся снаружи, образуют вторичные клетки флоэмы. По мере накопления вторичной ксилемы увеличивается «обхват» (боковые размеры) стебля и корня. В результате ткани за пределами вторичной флоэмы, включая эпидермис и кору головного мозга, во многих случаях имеют тенденцию выталкиваться наружу и в конечном итоге «отслаиваются» (отпадают). [ нужна ссылка ]
В этот момент пробковый камбий начинает формировать перидерму, состоящую из защитных пробковых клеток. Стенки пробковых клеток содержат утолщения суберина — внеклеточного сложного биополимера. [14] Утолщения суберина обеспечивают физический барьер, защиту от патогенов и предотвращение потери воды из окружающих тканей. Кроме того, он также способствует процессу заживления ран у растений. [15] Также предполагается, что суберин может быть компонентом апопластного барьера (присутствующего во внешних слоях клеток корней), который предотвращает попадание токсичных соединений в корень и снижает радиальную потерю кислорода (РОЛ) из аэренхимы во время переувлажнения. [16] В корнях пробковый камбий зарождается в перицикле , компоненте сосудистого цилиндра. [16]
Сосудистый камбий ежегодно производит новые слои вторичной ксилемы. [ нужна ссылка ] Сосуды ксилемы в зрелом состоянии погибают (у некоторых), но отвечают за транспорт большей части воды через сосудистую ткань в стеблях и корнях.

Корни деревьев обычно вырастают в три раза больше диаметра ветвей, из которых только половина лежит под стволом и кроной. Корни с одной стороны дерева обычно снабжают питательными веществами листву на той же стороне. Однако некоторые семейства, такие как Sapindaceae ( семейство клена ), не обнаруживают корреляции между расположением корня и тем, где корень поставляет питательные вещества растению. [17]
Регулирование
[ редактировать ]Существует корреляция между корнями, которые используют процесс восприятия растений , чтобы почувствовать свою физическую среду для роста. [18] включая ощущение света, [19] и физические барьеры. Растения также чувствуют гравитацию и реагируют через ауксиновые пути. [20] что приводит к гравитропизму . Со временем корни могут треснуть фундамент, порвать водопроводы и поднять тротуары. Исследования показали, что корни обладают способностью распознавать «свои» и «чужие» корни в одной и той же почвенной среде. [21]
Правильная среда из воздуха , минеральных питательных веществ и воды заставляет корни растений расти в любом направлении, чтобы удовлетворить потребности растения. Корни будут стесняться или сморщиваться от высыхания. [22] или другие плохие почвенные условия.
Гравитропизм заставляет корни расти вниз при прорастании — механизм роста растений, который также заставляет побеги расти вверх. [23] Различные типы корней, такие как первичные, семенные, боковые и коронковые, поддерживаются под разными гравитропными заданными углами, то есть в направлении, в котором они растут. Недавние исследования показывают, что угол наклона корней у зерновых культур, таких как ячмень и пшеница, регулируется новым геном под названием Enhanced Gravitropism 1 (EGT1). [24]
Исследования показывают, что корни растений, растущие в поисках продуктивного питания, могут чувствовать и избегать уплотнения почвы за счет диффузии газа этилена . [25]

Реакция избегания тени
[ редактировать ]Чтобы избежать тени, растения используют реакцию избегания тени. Когда растение находится под густой растительностью, присутствие поблизости другой растительности заставит растение избегать бокового роста и увеличит рост побегов вверх, а также рост корней вниз. Чтобы избежать тени, растения корректируют архитектуру своей корневой системы, в первую очередь за счет уменьшения длины и количества боковых корней, выходящих из главного корня. Эксперименты с мутантными вариантами Arabidopsis thaliana показали, что растения чувствуют соотношение красного и дальнего красного света, который поступает в растение через фоторецепторы, известные как фитохромы . [26] Листья близлежащих растений будут поглощать красный свет и отражать дальний красный свет, что приведет к снижению соотношения красного и дальнего красного света. Фитохром PhyA, который ощущает соотношение красного и дальнего красного света, локализован как в корневой системе, так и в побеговой системе растений, но в ходе экспериментов с нокаутными мутантами было обнаружено, что локализованный в корнях PhyA не ощущает соотношение света, даже напрямую. или аксиально, что приводит к изменениям в архитектуре латерального корня. [26] Вместо этого исследования показали, что локализованный в побегах PhyA является фитохромом, ответственным за эти архитектурные изменения бокового корня. Исследования также показали, что фитохром завершает эти архитектурные изменения за счет манипулирования распределением ауксина в корне растения. [26] Когда PhyA обнаруживает достаточно низкое соотношение красного и дальнего красного, phyA в побеге будет в основном находиться в активной форме. [27] В этой форме PhyA стабилизирует транскрипционный фактор HY5, предотвращая его разрушение, как это происходит, когда phyA находится в неактивной форме. Этот стабилизированный транскрипционный фактор затем может быть транспортирован к корням растения через флоэму , где он начинает индуцировать собственную транскрипцию как способ усиления своего сигнала. В корнях растения HY5 ингибирует фактор ответа на ауксин, известный как ARF19, фактор ответа, ответственный за трансляцию PIN3 и LAX3, двух хорошо известных белков, транспортирующих ауксин . [27] уровень и активность транспортеров ауксина PIN3 и LAX3. Таким образом, посредством манипуляции с ARF19 ингибируется [27] После ингибирования уровни ауксина будут низкими в областях, где обычно происходит появление боковых корней, что приводит к невозможности появления у растения зачатка бокового корня через перицикл корня . Благодаря этой сложной манипуляции с транспортом ауксина в корнях боковое появление корней будет подавлено, и вместо этого корень будет удлиняться вниз, способствуя вертикальному росту растений в попытке избежать тени. [26] [27]
Исследования арабидопсиса привели к открытию того, как работает реакция корня, опосредованная ауксином. Пытаясь выяснить роль, которую фитохром играет в развитии боковых корней, Salisbury et al. (2007) работали с Arabidopsis thaliana, выращенным на чашках с агаром. Солсбери и др. использовали растения дикого типа вместе с различными мутантами Arabidopsis с нокаутом белков и генов, чтобы наблюдать результаты, которые эти мутации оказали на архитектуру корня, присутствие белка и экспрессию генов. Для этого Солсбери и др. использовали флуоресценцию GFP вместе с другими формами как макро-, так и микроскопических изображений, чтобы наблюдать любые изменения, вызванные различными мутациями. На основании этих исследований Salisbury et al. смогли предположить, что фитохромы, расположенные в побегах, изменяют уровень ауксина в корнях, контролируя развитие боковых корней и общую архитектуру корня. [26] В экспериментах ван Гельдерена и др. (2018) они хотели выяснить, изменяется ли побег A. thaliana и влияет ли он на развитие и архитектуру корня и если да, то каким образом. Для этого они взяли растения арабидопсиса , вырастили их в агаровом геле и подвергли корни и побеги отдельным источникам света. Отсюда они изменили различные длины волн света, которые получали побег и корень растений, и зафиксировали плотность боковых корней, количество боковых корней и общую архитектуру боковых корней. Чтобы определить функцию специфических фоторецепторов, белков, генов и гормонов, они использовали различные нокаутные мутанты Arabidopsis и наблюдали возникающие в результате изменения в архитектуре боковых корней. Благодаря своим наблюдениям и различным экспериментам ван Гельдерен и др. смогли разработать механизм того, как обнаружение корнем соотношения красного и дальнего красного света влияет на развитие боковых корней. [27]
Типы
[ редактировать ]Настоящая корневая система состоит из главного корня и вторичных корней (или боковых корней ).
- диффузная корневая система: главный корень не является доминирующим; вся корневая система мочковатая и разветвляется во все стороны. Чаще всего встречается у однодольных . Основная функция мочковатого корня – закрепление растения.
Специализированный
[ редактировать ]







Корни или части корней многих видов растений стали специализированными для выполнения адаптивных целей помимо двух основных функций. [ нужны разъяснения ] , описанный во введении.
- Придаточные корни возникают вне последовательности из более обычного корневого образования ветвей главного корня и вместо этого происходят из стебля, ветвей, листьев или старых древесных корней. Они обычно встречаются у однодольных и птеридофитов, а также у многих двудольных растений , таких как клевер ( Trifolium ), плющ ( Hedera ), клубника ( Fragaria ) и ива ( Salix ). Большинство воздушных корней и ходульных корней являются придаточными. У некоторых хвойных придаточные корни могут составлять большую часть корневой системы. Образование адвентивных корней усиливается у многих видов растений во время (частичного) погружения в воду для увеличения газообмена и хранения газов, таких как кислород. [28] Различные типы придаточных корней можно классифицировать в зависимости от морфологии, динамики роста и функции. [29] [30]
- Аэрирующие корни (или коленный корень , или колено , или пневматофоры ): корни, возвышающиеся над землей, особенно над водой, как, например, у некоторых мангровых деревьев родов ( Avicennia , Sonneratia ). У некоторых растений, например у Авиценнии, прямостоячие корни имеют большое количество дыхательных пор для газообмена.
- Воздушные корни : корни полностью над землей, например, у плюща ( Хедера ) или у эпифитных орхидей . Многие воздушные корни используются для получения воды и питательных веществ непосредственно из воздуха – из туманов, росы или влажности воздуха. [31] Некоторые используют листовую систему для сбора дождя или влаги и даже хранят ее в чешуйках или карманах. Другие воздушные корни, например воздушные корни мангровых деревьев , используются для аэрации, а не для поглощения воды. Другие воздушные корни используются в основном для структуры, функционируя как опорные корни, как у кукурузы , или якорные корни, или как ствол у инжира-душителя . У некоторых эпифитов – растений, живущих над поверхностью других растений, воздушные корни служат для достижения источников воды или выхода на поверхность, а затем функционируют как обычные поверхностные корни. [31]
- Корни кроны /древесные корни : корни, которые образуются, когда ветви деревьев поддерживают маты из эпифитов и детрита, которые удерживают воду и питательные вещества в кроне. Они вырастают в эти маты, вероятно, используя доступные питательные вещества и влагу. [32]
- Сократительные корни : корни, которые тянут луковицы или клубнелуковицы однодольных растений , таких как гиацинт и лилия , а также некоторых стержневых корней , таких как одуванчик , глубже в почву, расширяясь в радиальном направлении и сжимаясь в продольном направлении. Они имеют морщинистую поверхность. [33]
- Грубые корни : корни, подвергшиеся вторичному утолщению и имеющие деревянистую структуру. Эти корни обладают некоторой способностью поглощать воду и питательные вещества, но их основная функция — транспортировка и обеспечение структуры, соединяющей тонкие корни меньшего диаметра с остальной частью растения.
- Диморфные корневые системы : корни с двумя различными формами для двух отдельных функций.
- Тонкие корни : обычно первичные корни диаметром менее 2 мм, выполняющие функцию поглощения воды и питательных веществ. Они часто сильно разветвлены и поддерживают микоризу. Эти корни могут быть недолговечными, но заменяются растением в ходе продолжающегося процесса «обновления» корней.
- Хаусториальные корни : корни растений-паразитов, которые могут поглощать воду и питательные вещества от другого растения, например, омелы ( Viscum album ) и повилики .
- Корни для размножения : корни, которые образуют придаточные почки, которые развиваются в надземные побеги, называемые присосками , из которых образуются новые растения, такие как чертополох канадский , вишня и многие другие.
- Протеоидные корни или кластерные корни: плотные скопления корешков ограниченного роста, которые развиваются в условиях низкого содержания фосфатов или железа у протейных и некоторых растений из следующих семейств Betulaceae , Casuarinaceae , Elaeagnaceae , Moraceae , Fabaceae и Myricaceae .
- Корни-ходули : придаточные опорные корни, распространенные среди мангровых зарослей . Они растут вниз из боковых ветвей, ветвясь в почве.
- Корни для хранения : корни, модифицированные для хранения пищи или воды, такие как морковь и свекла . Они включают в себя некоторые стержневые и клубневые корни.
- Структурные корни : большие корни, подвергшиеся значительному вторичному утолщению и обеспечивающие механическую поддержку древесным растениям и деревьям.
- Поверхностные корни : корни, которые разрастаются глубоко под поверхностью почвы, используя воду и легкодоступные питательные вещества. Там, где условия в поверхностных слоях почвы близки к оптимальным, рост поверхностных корней поощряется, и они обычно становятся доминирующими корнями.
- Клубневые корни : мясистые и увеличенные боковые корни для хранения пищи или воды, например, у сладкого картофеля . Тип корня хранения, отличный от стержневого корня.
- Фотосинтезирующие корни : зеленые корни, которые фотосинтезируют, обеспечивая растение сахаром. Они похожи на филлоклады . Они есть у некоторых орхидей, таких как Dendrophylax и Taeniophyllum .
- Корневые клубеньки : корни, в которых обитают азотфиксирующие почвенные бактерии. Они часто очень короткие и закругленные. Корневые клубеньки встречаются практически у всех бобовых .
- Кораллоидные корни: похожи на корневые клубеньки, они обеспечивают растение азотом. Они часто крупнее клубеньков, разветвлены, расположены на поверхности почвы или вблизи нее и содержат азотфиксирующие цианобактерии . Они встречаются только у саговников .
Глубины
[ редактировать ]
Распределение корней сосудистых растений в почве зависит от формы растения, пространственной и временной доступности воды и питательных веществ, а также физических свойств почвы. Самые глубокие корни обычно встречаются в пустынях и хвойных лесах умеренного пояса; самый мелкий в тундре, бореальных лесах и лугах умеренного пояса. Самый глубокий живой корень, находящийся на глубине не менее 60 метров (200 футов) от поверхности земли, был обнаружен во время раскопок открытого карьера в Аризоне, США. Некоторые корни могут расти так глубоко, как высота дерева. Однако большинство корней у большинства растений расположены относительно близко к поверхности, где доступность питательных веществ и аэрация более благоприятны для роста. Глубина укоренения может быть физически ограничена камнями или уплотненной почвой вблизи поверхности, а также анаэробными почвенными условиями.
Рекорды
[ редактировать ]
| Разновидность | Расположение | Максимальная глубина укоренения (м) | Ссылки [34] [35] |
|---|---|---|---|
| Боссия альбитрунка | Пустыня Калахари | 68 | Дженнингс (1974) |
| Можжевельник моносперма | Плато Колорадо | 61 | Пушка (1960) |
| Эвкалипт сп. | Австралийский лес | 61 | Дженнингс (1971) |
| Акация эриолоба | Пустыня Калахари | 60 | Дженнингс (1974) |
| Просопис juliflora | Аризонская пустыня | 53.3 | Филлипс (1963) |
Эволюционная история
[ редактировать ]Летопись окаменелостей корней — или, скорее, заполненных пустот, где корни гнили после смерти, — восходит к позднему силурийскому периоду , около 430 миллионов лет назад. [36] Их идентификация затруднена, поскольку слепки и слепки корней внешне очень похожи на норы животных. Их можно отличить по ряду признаков. [37] Эволюционное развитие корней, вероятно, произошло в результате модификации неглубоких корневищ (модифицированных горизонтальных стеблей), которые закрепляли примитивные сосудистые растения, в сочетании с развитием нитчатых выростов (называемых ризоидами ), которые закрепляли растения и проводили воду к растению из почвы. [38]
Взаимодействие с окружающей средой
[ редактировать ]
Было показано, что свет оказывает некоторое влияние на корни, но оно не изучено так тщательно, как влияние света на другие системы растений. Ранние исследования 1930-х годов показали, что свет снижает эффективность индол-3-уксусной кислоты при зарождении случайных корней. Исследования гороха в 1950-х годах показали, что образование боковых корней подавляется светом, а в начале 1960-х годов исследователи обнаружили, что в некоторых ситуациях свет может вызывать положительные гравитропные реакции. Влияние света на удлинение корней изучалось на однодольных и двудольных растениях, при этом в большинстве исследований было обнаружено, что свет ингибирует удлинение корней, как импульсное, так и непрерывное. Исследования арабидопсиса в 1990-х годах показали отрицательный фототропизм и ингибирование удлинения корневых волосков под воздействием света, воспринимаемого phyB . [39]
Некоторые растения, а именно Fabaceae , образуют корневые клубеньки , чтобы вступать в симбиотические отношения с азотфиксирующими бактериями, называемыми ризобиями . Из-за того, что для фиксации азота из атмосферы требуется большая энергия, бактерии забирают соединения углерода из растений, чтобы подпитывать этот процесс. Взамен растение забирает соединения азота, вырабатываемые бактериями из аммиака. [40]
Температура почвы является фактором, влияющим на корней зарождение и длину . На длину корня обычно более сильно влияет температура, чем на общую массу, поскольку более низкие температуры имеют тенденцию вызывать больший боковой рост, поскольку расширение вниз ограничивается более низкими температурами на уровне подпочвы. Потребности варьируются в зависимости от вида растений, но в регионах с умеренным климатом прохладные температуры могут ограничить корневую систему. Виды с прохладной температурой, такие как овес , рапс , рожь , пшеница, лучше приживаются при более низких температурах, чем летние однолетние растения , такие как кукуруза и хлопок . Исследователи обнаружили, что у таких растений, как хлопок, при более низких температурах стержневые корни становятся более широкими и короткими . Первый корень, происходящий из семени, обычно имеет более широкий диаметр, чем корневые ветви, поэтому ожидается, что корни будут меньшего диаметра, если температура увеличивает зарождение корней. Диаметр корня также уменьшается, когда корень удлиняется. [41]
Взаимодействие растений
[ редактировать ]Растения могут взаимодействовать друг с другом в окружающей среде через корневую систему. Исследования показали, что взаимодействие растений между растениями происходит между корневыми системами через почву как среду. Исследователи проверили, изменят ли растения, растущие в условиях окружающей среды, свое поведение, если соседнее растение подвергнется воздействию засухи. [42] Поскольку близлежащие растения не показали никаких изменений в устьичном отверстии, исследователи полагают, что сигнал засухи распространяется через корни и почву, а не через воздух в виде летучего химического сигнала. [43]
Взаимодействие с почвой
[ редактировать ]Почвенная микробиота способна подавлять как болезни, так и полезных корневых симбионтов (микоризные грибы легче приживаются в стерильной почве). Инокуляция почвенными бактериями может увеличить расширение междоузлий, урожайность и ускорить цветение. Миграция бактерий по корню варьируется в зависимости от естественных почвенных условий. Например, исследования показали, что корневые системы семян пшеницы, инокулированных Azotobacter, демонстрируют более высокую популяцию в почвах, благоприятных для роста Azotobacter . Некоторые исследования не смогли повысить уровень некоторых микробов (например, P. fluorescens ) в естественной почве без предварительной стерилизации. [44]
Корневая система травы полезна для уменьшения эрозии почвы , удерживая почву вместе. Многолетние травы, растущие в дикой природе на пастбищах, вносят в почву органические вещества, когда их старые корни разлагаются после нападения полезных грибов , простейших , бактерий, насекомых и червей, выделяя питательные вещества. [5]
Ученые наблюдали значительное разнообразие микробного покрова корней примерно на 10 процентах покрытых сегментов корней трехнедельного возраста. На более молодых корнях покрытие было даже низким, но даже на 3-месячных корнях покрытие составляло всего около 37%. До 1970-х годов ученые считали, что большая часть поверхности корня покрыта микроорганизмами. [5]
Усвоение питательных веществ
[ редактировать ]Исследователи, изучавшие проростки кукурузы , обнаружили, что наибольшее поглощение кальция происходит в верхушечном сегменте корня, а калия — у основания корня. Вдоль остальных сегментов корня поглощение было аналогичным. Поглощенный калий транспортируется к кончику корня и в меньшей степени к другим частям корня, а затем также к побегу и зерну. Транспорт кальция из апикального сегмента происходит медленнее, в основном он транспортируется вверх и накапливается в стебле и побеге. [45]
Исследователи обнаружили, что частичный дефицит K или P не изменил жирных кислот состав фосфатидилхолина в Brassica napus L растениях .. С другой стороны, дефицит кальция привел к заметному снижению содержания полиненасыщенных соединений, которые, как ожидается, будут иметь негативное влияние на целостность растительной мембраны , что может повлиять на некоторые свойства, такие как ее проницаемость, и которые необходимы для ионов активности поглощения . корневые оболочки. [46]
Экономическое значение
[ редактировать ]
Термин «корнеплоды» относится к любой съедобной подземной растительной структуре, но многие корнеплоды на самом деле представляют собой стебли, например клубни картофеля . Съедобные корни включают маниоку , сладкий картофель , свеклу , морковь , брюкву , репу , пастернак , редис , ямс и хрен . Специи, полученные из корней, включают сассафрас , ангелику , сарсапарель и солодку .
Сахарная свекла является важным источником сахара. Корни ямса являются источником соединений эстрогена , используемых в противозачаточных таблетках . Рыбий яд и инсектицид ротенон получают из корней Lonchocarpus spp. Важными лекарственными средствами из корней являются женьшень , аконит , ипекакуана , горечавка и резерпин . Некоторые бобовые культуры с корневыми клубеньками, фиксирующими азот, используются в качестве сидеральных культур, которые при вспашке обеспечивают азотное удобрение для других культур. Особые лысые корни кипариса , называемые коленями, продаются в качестве сувениров, подставок для ламп и вырезаются в народном искусстве. Коренные американцы использовали гибкие корни белой ели для плетения плетеных изделий.
Корни деревьев могут поднять и разрушить бетонные тротуары, а также раздавить или засорить закопанные трубы. [47] Воздушные корни инжира-душителя повредили древние майя храмы в Центральной Америке и храм Ангкор-Ват в Камбодже .
Деревья стабилизируют почву на склоне, подверженном оползням . Корневые волоски служат якорем в почве.
Вегетативное размножение растений черенками зависит от образования придаточных корней. размножаются сотни миллионов растений, черенками Ежегодно в том числе хризантемы , пуансеттии , гвоздики , декоративные кустарники и многие комнатные растения .
Корни также могут защитить окружающую среду, удерживая почву и уменьшая ее эрозию. Это особенно важно в таких местах, как песчаные дюны .

См. также
[ редактировать ]- Поглощение воды
- Кипарисовое колено
- Засушливый ризогенез
- Мочковатая корневая система
- Микориза – корневой симбиоз, при котором отдельные гифы, отходящие от мицелия гриба, заселяют корни растения-хозяина.
- Микоризная сеть
- Физиология растений
- Ризосфера - область почвы вокруг корня, находящаяся под влиянием корневых выделений и присутствующих микроорганизмов.
- Корневая обрезка
- Корневой порошок
- Столон
- Эффект Танады
- стержневой корень
Ссылки
[ редактировать ]- ^ Харли Макдональд и Донован Стивенс (3 сентября 2019 г.). Биотехнология и биология растений . ЭДТЕХ. стр. 141–. ISBN 978-1-83947-180-3 .
- ^ Нгуен, Линь Туи Ми; Хоанг, Хань Тхи; Чхве, Ынхо; Пак, Пил Сун (05 июля 2023 г.). «Распространение мангровых зарослей с различной морфологией воздушных корней на участках аккреции и эрозии в провинции Камау, Вьетнам» . Устьевые, прибрежные и шельфовые науки . 287 : 108324. Бибкод : 2023ECSS..28708324N . дои : 10.1016/j.ecss.2023.108324 . ISSN 0272-7714 .
- ^ «Части растения=Корни» . Расширение Университета Иллинойса .
- ^ Яаков Окон (24 ноября 1993 г.). Азоспириллы/Ассоциации растений . ЦРК Пресс. стр. 77–. ISBN 978-0-8493-4925-6 .
- ^ Jump up to: а б с «Садовник на заднем дворе: понимание корней растений» . Расширение сотрудничества Университета Аризоны .
- ^ Гангули Х.К., Дас К.С., Датта К.Т., Сен С. Колледж ботаники . Том. 1. Калькутта: Новое Центральное книжное агентство.
- ^ Дутта AC, Дутта TC. БОТАНИКА для студентов (6-е изд.). Издательство Оксфордского университета.
- ^ Шелдрейк, Мерлин (2020). Запутанная жизнь . Бодли Хед. п. 148. ИСБН 978-1847925206 .
- ^ Малами Дж. Э. (2005). «Внутренние и экологические пути реакции, которые регулируют архитектуру корневой системы» . Растение, клетка и окружающая среда . 28 (1): 67–77. дои : 10.1111/j.1365-3040.2005.01306.x . ПМИД 16021787 .
- ^ Jump up to: а б Колдуэлл М.М., Доусон Т.Э., Ричардс Дж.Х. (январь 1998 г.). «Гидравлический подъемник: последствия истечения воды из корней растений». Экология . 113 (2): 151–161. Бибкод : 1998Oecol.113..151C . дои : 10.1007/s004420050363 . ПМИД 28308192 . S2CID 24181646 .
- ^ Слесарь АХ (1991). «Экологическое значение архитектуры корневой системы: экономический подход». В Аткинсоне Д. (ред.). Рост корней растений: экологическая перспектива . Блэквелл. стр. 229–243.
- ^ Малами Дж. Э., Райан К. С. (ноябрь 2001 г.). «Экологическая регуляция зарождения боковых корней арабидопсиса» . Физиология растений . 127 (3): 899–909. дои : 10.1104/стр.010406 . ПМК 129261 . ПМИД 11706172 .
- ^ Рассел П.Дж., Герц П.Е., Макмиллан Б. (2013). Биология: динамическая наука . Cengage Обучение. п. 750. ИСБН 978-1-285-41534-5 . Архивировано из оригинала 21 января 2018 г. Проверено 24 апреля 2017 г.
- ^ «Суберин – обзор | Темы ScienceDirect» . www.sciencedirect.com . Проверено 31 августа 2021 г.
- ^ «Форма и функция Суберина - Марк Бернардс - Западный университет» . www.uwo.ca. Проверено 31 августа 2021 г.
- ^ Jump up to: а б Ватанабэ, Котаро; Нисиучи, Сюнсаку; Куличихин Константин; Накадзоно, Микио (2013). «Способствует ли накопление суберина в корнях растений устойчивости к заболачиванию?» . Границы в науке о растениях . 4 : 178. дои : 10.3389/fpls.2013.00178 . ISSN 1664-462X . ПМЦ 3683634 . ПМИД 23785371 .
- ^ ван ден Дриссе, Р. (1 июля 1974 г.). «Прогнозирование минерально-питательного статуса деревьев методом анализа листвы» . Ботаническое обозрение . 40 (3): 347–394. Бибкод : 1974BotRv..40..347V . дои : 10.1007/BF02860066 . ISSN 1874-9372 . S2CID 29919924 – через Springer.
- ^ Накагава И., Катагири Т., Шинозаки К., Ци Зи, Тацуми Х., Фуруичи Т. и др. (февраль 2007 г.). «Белок плазматической мембраны арабидопсиса, имеющий решающее значение для притока Ca2+ и сенсорного восприятия в корнях» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (9): 3639–44. Бибкод : 2007PNAS..104.3639N . дои : 10.1073/pnas.0607703104 . ПМК 1802001 . ПМИД 17360695 .
- ^ Механизм восприятия света УФ-В обнаружен в корнях растений , Государственный университет Сан-Франциско, 8 декабря 2008 г.
- ^ Маршан А., Каргул Дж., Мэй С.Т., Мюллер П., Дельбарр А., Перро-Рехенманн С., Беннетт М.Дж. (апрель 1999 г.). «AUX1 регулирует гравитропизм корня арабидопсиса, способствуя поглощению ауксина апикальными тканями корня» . Журнал ЭМБО . 18 (8): 2066–73. дои : 10.1093/emboj/18.8.2066 . ПМК 1171291 . ПМИД 10205161 .
- ^ Ходж А. (июнь 2009 г.). «Коренные решения» . Растение, клетка и окружающая среда . 32 (6): 628–40. дои : 10.1111/j.1365-3040.2008.01891.x . ПМИД 18811732 .
- ^ Карминати А., Веттерляйн Д., Веллер Ю., Фогель Х.Дж., Освальд С.Е. (2009). «Когда корни теряют контакт». Журнал зоны Вадосе . 8 (3): 805–809. Бибкод : 2009VZJ.....8..805C . дои : 10.2136/vzj2008.0147 . S2CID 128600212 .
- ^ Чен Р., Розен Э., Массон П.Х. (июнь 1999 г.). «Гравитропизм у высших растений» . Физиология растений . 120 (2): 343–50. дои : 10.1104/стр.120.2.343 . ПМЦ 1539215 . ПМИД 11541950 .
- ^ Фузи, Риккардо; Розиньоли, Серена; Лу, Хаоюй; Санджорджи, Джузеппе; Бовина, Риккардо; Паттем, Джейкоб К.; Боркар, Адити Н.; Ломбарди, Марко; Форестан, Кристиан; Милнер, Сара Г.; Дэвис, Джейн Л.; Лале, Аниш; Киршнер, Гвендолин К.; Сваруп, Ранджан; Тассинари, Альберто; Панди, Бипин К.; Йорк, Ларри М.; Аткинсон, Брайан С.; Старрок, Крейг Дж.; Муни, Саша Дж.; Хоххолдингер, Франк; Такер, Мэтью Р.; Химмельбах, Аксель; Штейн, Нильс; Машер, Мартин; Нагель, Керстин А.; Де Гара, Лаура; Симмондс, Джеймс; Уауи, Кристобаль; Тубероза, Роберто; Линч, Джонатан П.; Якубов Глеб Е.; Беннетт, Малкольм Дж.; Бхосале, Рахул; Сальви, Сильвио (2 августа 2022 г.). «Угол корней контролируется EGT1 в зерновых культурах с использованием антигравитропного механизма» . Труды Национальной академии наук . 119 (31): e2201350119. Бибкод : 2022PNAS..11901350F . дои : 10.1073/pnas.2201350119 . ПМЦ 9351459 . ПМИД 35881796 . S2CID 251104211 .
- ^ Панди, Бипин К.; Хуан, Гоцян; Бхосале, Рахул; Хартман, Сьон; Старрок, Крейг Дж.; Хосе, Лотти; Мартин, Оливье К.; Каради, Михал; Воесенек, Лаврентий ACJ; Люнг, Карин; Линч, Джонатан П. (15 января 2021 г.). «Корни растений ощущают уплотнение почвы за счет ограниченной диффузии этилена» . Наука . 371 (6526): 276–280. Бибкод : 2021Sci...371..276P . дои : 10.1126/science.abf3013 . hdl : 1874/418726 . ISSN 0036-8075 . ПМИД 33446554 . S2CID 231606782 .
- ^ Jump up to: а б с д и Солсбери FJ, зал A, Grierson CS, Холлидей KJ (май 2007 г.). «Фитохром координирует развитие побегов и корней арабидопсиса» . Заводской журнал . 50 (3): 429–38. дои : 10.1111/j.1365-313x.2007.03059.x . ПМИД 17419844 .
- ^ Jump up to: а б с д и ван Гельдерен К., Канг С., Паалман Р., Кеускамп Д., Хейс С., Пьерик Р. (январь 2018 г.). «Обнаружение дальнего красного света в побегах регулирует развитие боковых корней посредством фактора транскрипции HY5» . Растительная клетка . 30 (1): 101–116. дои : 10.1105/tpc.17.00771 . ПМК 5810572 . ПМИД 29321188 .
- ^ Айи, Цяоли; Цзэн, Бо; Лю, Цзяньхуэй; Ли, Сики; ван Бодегом, Питер М.; Корнелиссен, Йоханнес ХК (октябрь 2016 г.). «Поглощение кислорода придаточными корнями способствует выживанию полностью погруженных в воду наземных растений» . Анналы ботаники . 118 (4): 675–683. дои : 10.1093/aob/mcw051 . ПМК 5055620 . ПМИД 27063366 .
- ^ Линь, Чен; Огорек, Лукас Леон Перальта; Лю, Дэн; Педерсен, Оле; Заутер, Маргрет (11 января 2023 г.). «Локус количественного признака, придающий глубоководному рису устойчивость к наводнениям, регулирует образование двух различных типов водных дополнительных корней» . Новый фитолог . 238 (4): 1403–1419. дои : 10.1111/nph.18678 . ПМИД 36519256 . S2CID 254769152 .
- ^ Марич, Аида; Хартман, Сьон (11 марта 2023 г.). «Оболочка листьев обеспечивает длительную защиту от наводнений, образуя специальные придаточные корни» . Новый фитолог . 238 (4): 1337–1339. дои : 10.1111/nph.18824 . ПМИД 36905344 . S2CID 257438007 .
- ^ Jump up to: а б Новак Э.Дж., Мартин CE (1997). «Физиологические и анатомические реакции на дефицит воды у эпифита CAM Tillandsia ionantha (Bromeliaceae)» . Международный журнал наук о растениях . 158 (6): 818–826. дои : 10.1086/297495 . hdl : 1808/9858 . JSTOR 2475361 . S2CID 85888916 .
- ^ Надкарни Н.М. (ноябрь 1981 г.). «Корни полога: конвергентная эволюция в циклах питательных веществ тропических лесов». Наука . 214 (4524): 1023–4. Бибкод : 1981Sci...214.1023N . дои : 10.1126/science.214.4524.1023 . ПМИД 17808667 . S2CID 778003 .
- ^ Пютц Н. (2002). «Сократительные корни». В Вайзеле Ю., Эшеле А., Кафкафи У. (ред.). Корни растений: Скрытая половина (3-е изд.). Нью-Йорк: Марсель Деккер. стр. 975–987.
- ^ Канаделл Дж., Джексон Р.Б., Элерингер Дж.Б., Муни Х.А., Сала О.Э., Шульце Э.Д. (декабрь 1996 г.). «Максимальная глубина укоренения типов растительности в глобальном масштабе». Экология . 108 (4): 583–595. Бибкод : 1996Oecol.108..583C . дои : 10.1007/BF00329030 . ПМИД 28307789 . S2CID 2092130 .
- ^ Стоня Э.Л., Калишб П.Дж. (1 декабря 1991 г.). «О максимальной протяженности корней деревьев». Лесная экология и управление . 46 (1–2): 59–102. Бибкод : 1991ForEM..46...59S . дои : 10.1016/0378-1127(91)90245-Q .
- ^ Реталлак Г.Дж. (1986). «Ископаемая летопись почв» (PDF) . В вице-президенте Райта (ред.). Палеопочвы: их распознавание и интерпретация . Оксфорд: Блэквелл. стр. 1–57. Архивировано (PDF) из оригинала 7 января 2017 г.
- ^ Хиллиер Р., Эдвардс Д., Моррисси Л.Б. (2008). «Седиментологические данные о корневых структурах в Англо-Валлийском бассейне раннего девона (Великобритания) с предположениями об их производителях». Палеогеография, Палеоклиматология, Палеоэкология . 270 (3–4): 366–380. Бибкод : 2008PPP...270..366H . дои : 10.1016/j.palaeo.2008.01.038 .
- ^ Амрам Эшель; Том Бикман (17 апреля 2013 г.). Корни растений: скрытая половина, четвертое издание . ЦРК Пресс. стр. 1–. ISBN 978-1-4398-4649-0 .
- ^ Курата, Тецуя (1997). «Светостимулированное удлинение корня Arabidopsis thaliana». Журнал физиологии растений . 151 (3): 345–351. Бибкод : 1997JPPhy.151..346K . дои : 10.1016/S0176-1617(97)80263-5 . hdl : 2115/44841 .
- ^ Постгейт, Дж. (1998). Азотфиксация (3-е изд.). Кембридж, Великобритания: Издательство Кембриджского университета.
- ^ Лал, Ротанг (2006). Энциклопедия почвоведения . ЦРК Пресс. ISBN 978-0-8493-5054-2 .
- ^ Чамовиц, Дэниел. (21 ноября 2017 г.). Что знает растение: путеводитель по органам чувств . Фаррар, Штраус и Жиру. ISBN 9780374537128 . OCLC 1041421612 .
- ^ Фалик О, Мордок Ю, Бен-Натан Д, Вануну М, Гольдштейн О, Новоплански А (июль 2012 г.). «Реакция растений на корневую передачу сигналов стресса» . Анналы ботаники . 110 (2): 271–80. дои : 10.1093/aob/mcs045 . ПМЦ 3394639 . ПМИД 22408186 .
- ^ Боуэн Г.Д., Ровира А.Д. (1976). «Микробная колонизация корней растений». Анну. Преподобный Фитопатол . 14 : 121–144. дои : 10.1146/annurev.py.14.090176.001005 .
- ^ Корни растений и их среда обитания . Эльзевир. 1988. с. 17.
- ^ Корни растений и их среда обитания . Эльзевир. 1988. с. 25.
- ^ Захнисер, Дэвид (21 февраля 2008 г.). "Город, чтобы раздавать деньги на тротуарах?" . Лос-Анджелес Таймс . Проверено 30 марта 2023 г.
Дальнейшее чтение
[ редактировать ]- Бальдокки Д.Д., Сюй Л (октябрь 2007 г.). «Что ограничивает испарение из средиземноморских дубовых лесов - запас влаги в почве, физиологический контроль со стороны растений или потребность атмосферы?». Достижения в области водных ресурсов . 30 (10): 2113–22. Бибкод : 2007AdWR...30.2113B . дои : 10.1016/j.advwatres.2006.06.013 .
- Брундретт, MC (2002). «Коэволюция корней и микоризы наземных растений» . Новый фитолог . 154 (2): 275–304. дои : 10.1046/j.1469-8137.2002.00397.x . ПМИД 33873429 .
- Кларк Л. (2004). «Первичная корневая структура и развитие - конспекты лекций» (PDF) . Архивировано из оригинала (PDF) 3 января 2006 года.
- Член парламента Куттса (1987). «Процессы развития корневых систем деревьев». Канадский журнал лесных исследований . 17 (8): 761–767. дои : 10.1139/x87-122 .
- Рэйвен Дж. А., Эдвардс Д. (2001). «Корни: эволюционное происхождение и биогеохимическое значение». Журнал экспериментальной ботаники . 52 (Приложение 1): 381–401. дои : 10.1093/jxb/52.suppl_1.381 . ПМИД 11326045 .
- Шенк Х.Дж., Джексон Р.Б. (2002). «Глобальная биогеография корней». Экологические монографии . 72 (3): 311–328. дои : 10.2307/3100092 . JSTOR 3100092 .
- Саттон Р.Ф., Тинус Р.В. (1983). «Терминология корней и корневых систем». Лесная монография . 24 : 137.
- Филлипс WS (1963). «Глубина корней в почве». Экология . 44 (2): 424. Бибкод : 1963Ecol...44..424P . дои : 10.2307/1932198 . JSTOR 1932198 .
- Колдуэлл М.М., Доусон Т.Э., Ричардс Дж.Х. (1998). «Гидравлический подъемник: последствия истечения воды из корней растений». Экология . 113 (2): 151–161. Бибкод : 1998Oecol.113..151C . дои : 10.1007/s004420050363 . ПМИД 28308192 . S2CID 24181646 .