Корень

У сосудистых растений корни органы представляют собой растения , которые модифицированы, чтобы обеспечить закрепление для растения, и забрать воду и питательные вещества в тело растения, что позволяет растениям расти и быстрее. [ 1 ] Они чаще всего находятся ниже поверхности почвы , но корни также могут быть воздушными или аэродинамическими, то есть растущим над землей или особенно над водой. [ 2 ]
Функция
[ редактировать ]Основными функциями корней являются поглощение воды , питание растений и привязка корпуса растения на землю. [ 3 ]
Анатомия
[ редактировать ]
Морфология корней разделена на четыре зоны: корневая шапка, апикальная меристема , зона удлинения и волосы. [ 4 ] Корневая крышка новых корней помогает корню проникнуть в почву. Эти корневые шапки отключены, так как корень идет глубже, создавая слизистую поверхность, которая обеспечивает смазку. Апикальная меристема за корневой крышкой производит новые корневые клетки, которые удлиняются. Затем образуют корневые волосы, которые поглощают воду и минеральные питательные вещества из почвы. [ 5 ] Первым корнем в растениях, продуцирующих семена, является узел , который расширяется от эмбриона растений после прорастания семян.
При рассечении расположение клеток в корне - это корневые волосы , эпидермис , эпиблема , кора , эндодерма , перицикл и, наконец, сосудистая ткань в центре корня для транспортировки воды, поглощенной корнем в другие места растение. [ нужно разъяснения ]
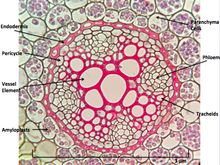
Пожалуй, самая поразительная характеристика корней, которые отличают их от других органов растений, таких как стволовые ветви и листья, заключается в том, что корни имеют эндогенные [ 6 ] Происхождение, т.е. они происходят и развиваются из внутреннего слоя материнской оси, такой как перицикл . [ 7 ] Напротив, стволовые ячейки и листья являются экзогенными , то есть они начинают развиваться из коры, внешнего слоя.
В ответ на концентрацию питательных веществ корни также синтезируют цитокинин , который действует как сигнал относительно того, как быстро могут расти побеги. Корни часто функционируют в хранении пищи и питательных веществ. Корни большинства видов сосудистых растений вступают в симбиоз с определенными грибами с образованием микоризы , а также широкий спектр других организмов, включая бактерии, также тесно связанные с корнями. [ 8 ]

Архитектура корневой системы (RSA)
[ редактировать ]
Определение
[ редактировать ]В простейшей форме термин архитектура корневой системы (RSA) относится к пространственной конфигурации корневой системы завода. Эта система может быть чрезвычайно сложной и зависит от множества факторов, таких как виды самого растения, состав почвы и наличия питательных веществ. [ 9 ] Корневая архитектура играет важную роль в обеспечении безопасного запаса питательных веществ и воды, а также якорной стоянки и поддержки.
Конфигурация корневых систем служит для структурной поддержки завода, конкурировать с другими растениями и для поглощения питательных веществ из почвы. [ 10 ] Корни растут до определенных условий, которые, если они изменены, могут препятствовать росту растения. Например, корневая система, которая развивалась в сухой почве, может быть не столь эффективной в затопленной почве, однако растения способны адаптироваться к другим изменениям в окружающей среде, таких как сезонные изменения. [ 10 ]
Условия и компоненты
[ редактировать ]Основными терминами, используемыми для классификации архитектуры корневой системы, являются: [ 11 ]
| Величина ветвления | Количество ссылок (экстерьер или интерьер) |
| Топология | Образец ветвления ( елочка , дихотомический , радиальный ) |
| Длина ссылки | Расстояние между ветвями |
| Корневой угол | Радиальный угол основания бокового корня вокруг окружности родительского корня, угол бокового корня из его родительского корня и угол, который распространяется вся система. |
| Радиус ссылки | Диаметр корня |
Все компоненты корневой архитектуры регулируются путем сложного взаимодействия между генетическими реакциями и реакциями из -за стимулов окружающей среды. Эти стимулы развития классифицируются как внутренние, генетические и пищевые влияния или внешние влияния окружающей среды и интерпретируются путями трансдукции сигнала . [ 12 ]
Внешние факторы, влияющие на корневую архитектуру, включают гравитацию, воздействие света, воду и кислород, а также доступность или отсутствие азота, фосфора, серы, алюминия и хлорида натрия. Основные гормоны (внутренние стимулы) и соответствующие пути, ответственные за развитие корневой архитектуры, включают:
| Ауксин | Боковое формирование корня, поддержание апикального доминирования и образовательного образования корней. |
| Цитокинины | Цитокинины регулируют апикальный размер меристемы корня и способствуют удлинению бокового корня. |
| Этилен | Способствует формированию корня короны. |
| Гиббеллинс | Вместе с этиленом они способствуют росту и удлинению короны. Вместе с ауксином они способствуют удлинению корня. Гиббеллины также ингибируют инициацию латерального корня. |
Рост
[ редактировать ]
Ранний рост корней является одной из функций апикальной меристемы, расположенной рядом с кончиком корня. Клетки меристемы более или менее непрерывно делятся, производя больше меристемы, клетки корневой крышки (они приносятся в жертву для защиты меристемы), и недифференцированные корневые клетки. Последние становятся первичными тканями корня, сначала проходя удлинение, процесс, который продвигает кончик корня вперед в среде для выращивания. Постепенно эти клетки дифференцируются и разразились в специализированные клетки тканей корня. [ 13 ]
Рост из апикальных меристем известен как первичный рост , который охватывает все удлинение. Вторичный рост охватывает весь рост в диаметре, основной компонент древесных тканей растений и многих не древесных растений. Например, корни для хранения сладкого картофеля имеют вторичный рост, но не являются древесными. Вторичный рост происходит в боковых меристемах , а именно в камбия сосудов и Cork Cambium . Первый образует вторичную ксилема и вторичную флоэму , в то время как вторая образует перидерму .
У растений с вторичным ростом сосудистый камбий, происходящий между ксилемой и флоэмой, образует цилиндр ткани вдоль стебля и корня. [ Цитация необходима ] Сосудистый камбий образует новые клетки как внутри, так и снаружи камбиевого цилиндра, с клетками внутри, образуя вторичные ксилемы, и на внешних образовании вторичных клеток флоэмы. По мере накопления вторичного ксилема «обхват» (боковые измерения) стебля и корня увеличивается. В результате, ткани за пределами вторичной флоэмы, включая эпидермис и кору, во многих случаях, как правило, выталкиваются наружу и в конечном итоге «отключены» (сарай). [ Цитация необходима ]
На этом этапе пробковой камбий начинает образовывать перидерму, состоящий из защитных пробковых клеток. Стены из пробковых клеток содержит утолщения саберина , который является экстраклеточным комплексным биополимером. [ 14 ] Утолщение Suberin функционирует, обеспечивая физический барьер, защиту от патогенных микроорганизмов и предотвращая потерю воды от окружающих тканей. Кроме того, это также помогает процессу заживления ран в растениях. [ 15 ] Также постулируется, что Suberin может быть компонентом апопластического барьера (присутствующего на наружных клеточных слоях корней), который предотвращает попадание токсичных соединений в корень и уменьшает потерю радиального кислорода (ROL) от Aerrenchyma во время оспаривания. [ 16 ] В корнях пробковый камбий происходит в перицикле , компоненте сосудистого цилиндра. [ 16 ]
Сосудистый камбий производит новые слои вторичного ксилема в год. [ Цитация необходима ] Ссуды ксилемы мертвы при зрелости (в некоторых случаях), но ответственны за большую часть транспорта воды через сосудистую ткань в стеблях и корнях.

Корни деревьев обычно растут до трех раз больше диаметра распространения ветви, только половина из которых лежит под стволом и навесом. Корни с одной стороны дерева обычно поставляют питательные вещества в листву на одной и той же стороне. Однако некоторые семьи, такие как Sapindaceae ( семейство Maple ), не показывают корреляции между корневым местоположением и где корень поставляет питательные вещества на растении. [ 17 ]
Регулирование
[ редактировать ]Существует корреляция корней с использованием процесса восприятия растений, чтобы ощутить их физическую среду для роста, [ 18 ] включая чувствительность света, [ 19 ] и физические барьеры. Растения также ощущают гравитацию и реагируют через ауксинские пути, [ 20 ] приводя к гравитропизму . Со временем корни могут взломать фундаменты, линии воды и поднимать тротуары. Исследования показали, что корни обладают способностью распознавать корни «я» и «не-я» в той же почвенной среде. [ 21 ]
Правильная среда воздуха , минеральных питательных веществ и воды направляет корни растений в любом направлении, чтобы удовлетворить потребности растения. Корни будут застенчивыми или удерживаются от сухого [ 22 ] или другие плохие почвенные условия.
Гравитропизм направляет корни, чтобы расти вниз при прорастании , механизм роста растений, который также приводит к росту побега. [ 23 ] Различные типы корней, такие как первичные, семенные, боковые и корона, поддерживаются под разными гравитропными углами заданными точками, то есть в направлении, в котором они растут. Недавние исследования показывают, что угол корня в зерновых культурах, таких как ячмень и пшеница, регулируется новым геном, называемым усиленным гравитропизмом 1 (EGT1). [ 24 ]
Исследования показывают, что корни растений, растущие в поисках продуктивного питания, могут ощущать и избежать уплотнения почвы посредством диффузии газового этилена . [ 25 ]

Ответ избегания тени
[ редактировать ]Чтобы избежать тени, растения используют реакцию избегания тени. Когда растение находится под плотной растительностью, присутствие другой растительности поблизости приведет к тому, что растение избегает бокового роста и испытывает увеличение восходящего побега, а также рост корня вниз. Чтобы избежать тени, растения корректируют свою корневую архитектуру, особенно путем уменьшения длины и количества боковых корней, возникающих из основного корня. Экспериментирование мутантных вариантов арабидопсиса thaliana показало, что растения чувствуют отношение красного и дальнего красного света, которое попадает в растение через фоторецепторы, известные как фитохромы . [ 26 ] Соседние листья растений поглотят красный свет и отражают дальний красный свет, что приведет к снижению коэффициента красно-красного света. Фитохром Phya, которая ощущает это соотношение красного и отдаленного света, локализуется как в корневой системе, так и в системе побега растений, но с помощью нокаутированных мутантных экспериментов было обнаружено, что локализованная корневая Phya не ощущает отношение света, независимо от того, напрямую или осевидиально, это приводит к изменениям в боковой корневой архитектуре. [ 26 ] Вместо этого исследование показало, что локализованная Phya Shoot является фитохромом, ответственным за причинение этих архитектурных изменений бокового корня. Исследования также показали, что фитохром завершает эти архитектурные изменения посредством манипуляции с распределением ауксина в корне растения. [ 26 ] Когда Phya определяется достаточно низким красно -красным соотношением, Phya в побеге будет в основном в его активной форме. [ 27 ] В этой форме Phya стабилизирует транскрипционный фактор , который, заставляющий его больше не быть ухудшенным, как это когда Phya находится в своей неактивной форме. Этот стабилизированный фактор транскрипции затем способен транспортировать в корни растения через флоэму , где он продолжает вызывать свою транскрипцию как способ усилить свой сигнал. В корнях растения функции Hy5 функционирует, чтобы ингибировать коэффициент ответа ауксина, известный как ARF19, коэффициент отклика, ответственный за трансляцию PIN3 и LAX3, два известных белка, транспортирующих ауксин . [ 27 ] Таким образом, посредством манипуляции с ARF19 уровень и активность Auxin Transporters PIN3 и LAX3 ингибируются. [ 27 ] После ингибирования уровни ауксина будут низкими в областях, где обычно происходит появление бокового корня, что приводит к неспособности растения иметь появление латерального перчарда корня через перицикл корня . С этим сложным манипуляциями с транспортом ауксина в корнях, в корнях будет ингибировано боковое появление корня, а корни вместо этого удлинятся вниз, способствуя росту вертикального растения в попытке избежать тени. [ 26 ] [ 27 ]
Исследования Arabidopsis привели к открытию того, как работает этот ауксин, опосредованный корневым ответом. В попытке выяснить роль, которую фитохром играет в развитии латерального корня, Salisbury et al. (2007) работали с Arabidopsis Thaliana, выращенной на агарских тарелках. Salisbury et al. Использовали растения дикого типа наряду с различным белком нокаутом и нокаутированными мутантами арабидопсиса, чтобы наблюдать за результатами, которые эти мутации имели по корневой архитектуре, присутствию белка и экспрессии генов. Для этого Salisbury et al. использовал флуоресценцию GFP вместе с другими формами как макро, так и микроскопических изображений для наблюдения за любыми изменениями, вызванными различными мутациями. Из этих исследований Salisbury et al. смогли теоретизировать этот побег, расположенный фитохромы изменяют уровни ауксина в корнях, контролируя развитие боковых корней и общую архитектуру корневых. [ 26 ] В экспериментах Van Gelderen et al. (2018), они хотели посмотреть, изменяется ли и как это так, влияет и влияет на развитие корней и корневую архитектуру. Чтобы сделать это, они взяли Arabidopsis растения , выращивали их в геле агара и обнажали корни и побеги, чтобы разделить источники света. Отсюда они изменили различные длина волн света, которые получали побег и корень растений, и записали боковую плотность корня, количество боковых корней и общую архитектуру боковых корней. Чтобы идентифицировать функцию специфических фоторецепторов, белков, генов и гормонов, они использовали различные мутанты нокаутирования арабидопсиса и наблюдали результирующие изменения в архитектуре боковых корней. Благодаря их наблюдениям и различным экспериментам, Van Gelderen et al. смогли разработать механизм для обнаружения корней от красного и отдаленного света, изменяет развитие латерального корня. [ 27 ]
Типы
[ редактировать ]Настоящая корневая система состоит из основных корней и вторичных корней (или боковых корней ).
- Диффузная корневая система: основной корень не является доминирующим; Вся корневая система является волокнистой и ветви во всех направлениях. Наиболее распространенный у монокотов . Основная функция фиброзного корня - закрепить растение.
Специализирован
[ редактировать ]







Корни или части корней многих видов растений стали специализированными для обслуживания адаптивных целей, кроме двух основных функций [ нужно разъяснения ] , описано во введении.
- Авентативные корни возникают вне последовательности из более обычного образования корней ветвей первичного корня, и вместо этого происходят из стебля, ветвей, листьев или старых древесных корней. Они обычно встречаются в монокотах и птеридофитах, но также во многих дикотах , таких как клевер ( трифолиум ), плющ ( хедера ), клубника ( Fragaria ) и ива ( Salix ). Большинство воздушных корней и корней снятия являются случайными. В некоторых хвойных деревьях Adventitious корни могут образовывать самую большую часть корневой системы. Авентаризация корневой формирование усиливается во многих видах растений во время (частичного) погружения, чтобы увеличить газообмен и хранение газов, таких как кислород. [ 28 ] Отдельные типы случайных корней могут быть классифицированы и зависят от морфологии, динамики роста и функции. [ 29 ] [ 30 ]
- Аэродинамические корни (или корень колена , коленное или пневматофоры ): корни, поднимающиеся над землей, особенно над водой, например, в некоторых мангровых родах ( Avicennia , Sonneratia ). На некоторых растениях, таких как Avicennia, прямое корни имеют большое количество дыхательных пор для обмена газами.
- Воздушные корни : корни полностью над землей, например, в плюще ( хедера ) или в эпифитных орхидеях . Многие воздушные корни используются для приема воды и питательных веществ непосредственно от воздуха - от туманов, росы или влажности в воздухе. [ 31 ] Некоторые полагаются на листовые системы, чтобы собрать дождь или влажность и даже хранить его в весах или карманах. Другие воздушные корни, такие как мангровые воздушные корни, используются для аэрации, а не для водопоглощения. Другие воздушные корни используются в основном для структуры, функционирующих в качестве пропагандистских корней, как в корнях кукурузы или якоря или в качестве ствола в Strangler Fig . В некоторых эпифитах - растениях, живущих над поверхностью на других растениях, воздушные корни служат для достижения источников воды или достижения поверхности, а затем функционируют как обычные поверхностные корни. [ 31 ]
- Корни навес /древесные корни : корни, которые образуются, когда ветви деревьев поддерживают коврики эпифитов и детрита, которые содержат воду и питательные вещества в навес. Они вырастут в эти коврики, вероятно, будут использовать доступные питательные вещества и влагу. [ 32 ]
- Сократительные корни : корни, которые тянут луковицы или корма монокотов , таких как гиацинт и лилия , и некоторые капля , такие как одуванчик , глубже в почве посредством расширения радиально и сжимающих продольно. У них есть морщинистая поверхность. [ 33 ]
- Грубые корни : корни, которые подвергались вторичному утолщению и имеют древесную структуру. Эти корни обладают некоторой способностью поглощать воду и питательные вещества, но их основной функцией является транспортировка и обеспечение структуры для соединения меньшего диаметра, тонких корней с остальной частью растения.
- Диморфические корневые системы : корни с двумя отличительными формами для двух отдельных функций
- Мелкие корни : обычно первичные корни <2 мм диаметром, которые имеют функцию поглощения воды и питательных веществ. Они часто сильно разветвляются и поддерживают микоризы. Эти корни могут быть недолгими, но заменяются заводом в продолжающемся процессе корневого «оборота».
- Haustorial Roots : корни паразитических растений, которые могут поглощать воду и питательные вещества из другого растения, например, в омеле ( вице -альбом ) и Доддер .
- Распространенные корни : корни, которые образуют экстенсионные бутоны, которые превращаются в надземные побеги, называются присоски , которые образуют новые растения, как в Канаде чертополох , вишни и многих других.
- Протеоидные корни или кластерные корни: плотные кластеры корнеров ограниченного роста, которые развиваются в условиях с низким фосфатом или низким содержанием железа у Proteaceae и некоторых растений из следующих семейств Betulaceae , Casuarinaceae , Elaeganaceae , Moraceae , Fabaceae и Myricaceae .
- Корни склонов : Adventious Corpes, распространенные среди мангровых лесов . Они растут из боковых ветвей, разветвив в почве.
- Корни для хранения : корни, измененные для хранения пищи или воды, таких как морковь и свекла . Они включают в себя некоторые Taproots и клубневые корни.
- Структурные корни : большие корни, которые подверглись значительному вторичному утолщению и обеспечивают механическую поддержку древесных растений и деревьев.
- Поверхностные корни : корни, которые размножаются близко под поверхностью почвы, эксплуатируют воду и легко доступны питательные вещества. Там, где условия близки к оптимальному в поверхностных слоях почвы, рост поверхностных корней способствует, и они обычно становятся доминирующими корнями.
- КУБОТЫЕ КОРНИ : Мясистые и увеличенные боковые корни для хранения пищи или воды, например, сладкий картофель . Тип корня хранения, отличный от Taproot.
- Фотосинтетические корни : корни, которые зеленые и фотосинтезируют, обеспечивая сахар для растения. Они похожи на филлоклады . У нескольких орхидей есть, такие как дендрофилакс и таениофиллум .
- Корневые узелки : корни, которые содержат азот-фиксирующие почвенные бактерии. Они часто очень короткие и округлые. Корневые узелки встречаются практически во всех бобовых .
- Кораллоидные корни: аналогично корневым узлам, они обеспечивают азот для растения. Они часто больше, чем узелки, разветвленные и расположены на поверхности почвы или рядом с ним, а также содержат азотные цианобактерии . Они найдены только в Cycads .
Глубины
[ редактировать ]
Распределение корней сосудистых растений в почве зависит от формы растения, пространственной и временной доступности воды и питательных веществ, а также физических свойств почвы. Самые глубокие корни обычно встречаются в пустынях и умеренных хвойных лесах; Самые мелкие в тундре, бореальном лесу и умеренных лугах. Самый глубокий наблюдаемый живой корень, по меньшей мере, 60 метров (200 футов) ниже поверхности земли, наблюдался во время раскопки шахты с открытым питанием в Аризоне, США. Некоторые корни могут расти так же глубоко, как дерево высокое. Большинство корней на большинстве растений, однако, обнаруживаются относительно близко к поверхности, где доступность питательных веществ и аэрация более благоприятны для роста. Глубина укоренения может быть физически ограничена породой или уплотненной почвой, близкой к поверхности или анаэробными условиями почвы.
Записи
[ редактировать ]
| Разновидность | Расположение | Максимальная глубина укоренения (м) | Ссылки [ 34 ] [ 35 ] |
|---|---|---|---|
| Albitrumone Bellia | Калахари пустыня | 68 | Дженнингс (1974) |
| Можжейпер Моноспех | Колорадское плато | 61 | Кэннон (1960) |
| Eucalyptus sp. | Австралийский лес | 61 | Дженнингс (1971) |
| Acacia erioloba | Калахари пустыня | 60 | Дженнингс (1974) |
| Просопис Юлифлора | Аризона пустыня | 53.3 | Филлипс (1963) |
Эволюционная история
[ редактировать ]Окаменечная запись корней - или, скорее, внедренных пустот, где корни сгнили после смерти - возвращаются к покойному силурийскому , около 430 миллионов лет назад. [ 36 ] Их идентификация затруднена, потому что сбросы и плесени корней настолько похожи по внешнему виду на норы животных. Они могут быть дискриминированы с помощью ряда функций. [ 37 ] Эволюционное развитие корней, вероятно, произошло из -за модификации мелких корневищ (модифицированных горизонтальных стеблей), которые закрепляли примитивные сосудистые растения в сочетании с развитием нитчатых исходов (называемых ризоидами ), которые закрепляли растения и проводили воду на растение из почвы. [ 38 ]
Экологические взаимодействия
[ редактировать ]
Было показано, что свет оказывает некоторое влияние на корни, но его не изучалось так же, как влияние света на другие системы растений. Ранние исследования в 1930-х годах показали, что свет снижает эффективность индоле-3-уксусной кислоты при случайном инициации корня. Исследования гороха в 1950 -х годах показывают, что образование латерального корня была ингибирована светом, и в начале 1960 -х исследователей обнаружили, что свет может вызвать положительные гравитропные реакции в некоторых ситуациях. Влияние света на удлинение корня было изучено для монокотидочных и двудольных растений, причем большинство исследований обнаружили, что свет ингибирует удлинение корней, будь то импульсным или непрерывным. Исследования арабидопсиса в 1990 -х годах показали негативный фототропизм и ингибирование удлинения корневых волосков при свете, чувствующих PhyB . [ 39 ]
Определенные растения, а именно Fabaceae , образуют корневые узелки , чтобы связать и сформировать симбиотическую связь с азот-фиксирующими бактериями, называемыми ризобией . Благодаря высокой энергии, необходимой для исправления азота из атмосферы, бактерии берут углеродные соединения с завода, чтобы подпитывать процесс. В свою очередь, растение берет азотные соединения, полученные из аммиака бактериями. [ 40 ]
Температура почвы является фактором, который влияет на инициацию корня и длину. Длина корня обычно больше влияет на температуру, чем общая масса, где более прохладные температуры имеют тенденцию вызывать больший боковой рост, поскольку расширение нисходящего ограничено более прохладными температурами на уровнях недр. Потребности варьируются в зависимости от видов растений, но в умеренных областях прохладные температуры могут ограничивать корневые системы. Прохладные температурные виды, такие как овес , рапс , ржаная , пшеничная плата лучше при более низких температурах, чем летние однолетние растения , такие как кукуруза и хлопок . Исследователи обнаружили, что такие растения, как хлопок, развиваются более широкие и более короткие капли при более прохладных температурах. Первый корень, исходящий из семян, обычно имеет более широкий диаметр, чем корневые ветви, поэтому ожидаются меньшие диаметры корня, если температура увеличивает инициацию корня. Диаметр корня также уменьшается, когда корень удлиняется. [ 41 ]
Взаимодействие растений
[ редактировать ]Растения могут взаимодействовать друг с другом в своей среде через их корневые системы. Исследования показали, что взаимодействие растительного завода происходит среди корневых систем через почву в качестве среды. Исследователи проверили, изменят ли растения в условиях окружающей среды их поведение, если соседнее растение подвергается воздействию условий засухи. [ 42 ] Поскольку близлежащие растения не показали изменений в устьевой апертуре, исследователи считают, что сигнал засухи распространяется по корням и почве, а не через воздух в качестве летучего химического сигнала. [ 43 ]
Почвенные взаимодействия
[ редактировать ]Микробиота почвы может подавлять как болезнь, так и полезные симбионты корня (грибы микоризы легче установить в стерильной почве). Инокуляция бактериями почвы может увеличить разгибание междоузлия, урожайность и ускорение цветения. Миграция бактерий вдоль корня варьируется в зависимости от естественных условий почвы. Например, исследования показали, что корневые системы семян пшеницы, инокулированные азотобактером, показали более высокие популяции в почвах, благоприятных для роста азотобактера . Некоторые исследования были неудачными в увеличении уровней определенных микробов (таких как P. fluorescens ) в естественной почве без предварительной стерилизации. [ 44 ]
Корневые системы травы полезны для уменьшения эрозии почвы , удерживая почву вместе. Многолетние травы, которые растут в пастбищах, вносят вклад органического вещества в почву, когда их старые корни распадаются после атаки полезных грибов , простейших , бактерий, насекомых и червей высвобождают питательные вещества. [ 5 ]
Ученые наблюдали значительное разнообразие микробного покров корней примерно в 10 процентах трехнедельных корневых сегментов. На более молодых корнях был даже низкий охват, но даже на 3-месячных корнях покрытие составляло всего около 37%. До 1970 -х годов ученые полагали, что большая часть поверхности корня была покрыта микроорганизмами. [ 5 ]
Поглощение питательных веществ
[ редактировать ]Исследователи, изучающие саженцы кукурузы , обнаружили, что поглощение кальция было наибольшим в сегменте апикального корня, а калий у основания корня. Вдоль других корневых сегментов поглощение было одинаковым. Поглощенный калий транспортируется до кончика корня и, в меньшей степени, в других частях корня, а также к побегу и зерну. Транспорт кальция из апикального сегмента медленнее, в основном транспортируется вверх и накапливается в стебле и стрельбе. [ 45 ]
Исследователи обнаружили, что частичные недостатки k или p не изменили жирных кислот состав фосфатидилхолина у Brassica napus L. растений. Дефицит кальция, с другой стороны, привел к заметному снижению полиненасыщенных соединений, которые, как ожидается, окажут негативные воздействия на целостность растительной мембраны , которые могли бы повлиять на некоторые свойства, такие как проницаемость, и необходимо для активности ионного поглощения корневые мембраны. [ 46 ]
Экономическое значение
[ редактировать ]
Термин корневой культуры относится к любой съедобной подземной структуре растений, но многие корневые культуры на самом деле являются стеблями, такими как картофельные клубни. Съедобные корни включают маниоку , сладкий картофель , свеклу , морковку , рутабагу , репу , пастернак , редьку , ямс и хрен . Специи, полученные из корней, включают сассафра , анжелику , сарсапариллу и солодку .
Сахарная свекла является важным источником сахара. Корни YAM являются источником эстрогенных соединений, используемых в противозачаточных таблетках . Рыбный яд и инсектицидный ротенон получают из корней Lonchocarpus spp. Важными лекарствами от корней являются женьшень , аконит , ipecac , gendian и reserpine . Несколько бобовых, которые имеют азотирующие корневые узелки, используются в качестве зеленых культур, которые обеспечивают азотные удобрения для других культур, когда их пахают. Специализированные лысые кипарисовые корни, называемые коленями, продаются как сувениры, основания лампы и вырезаны в народном искусстве. Коренные американцы использовали гибкие корни белой ели для башетки.
Корни деревьев могут поднимать и уничтожать бетонные тротуары и раздавить или засорить закупоренные трубы. [ 47 ] Авиационные корни рис -инжирации повредили древние майя храмы в Центральной Америке и храм Ангкор -Ват в Камбодже .
Деревья стабилизируют почву на склоне, склонном к оползням . Корневые волосы работают как якорь на почве.
Вегетативное распространение растений с помощью черенков зависит от авантюзного образования корней. Сотни миллионов растений распространяются с помощью черенков ежегодно, включая хризантемуму , пуансеттию , гвоздику , декоративные кустарники и множество комнатных растений .
Корни также могут защитить окружающую среду, удерживая почву, чтобы уменьшить эрозию почвы. Это особенно важно в таких областях, как песчаные дюны .

Смотрите также
[ редактировать ]- Поглощение воды
- Кипарисовое колено
- Засуха ризогенез
- Волокнистая корневая система
- Микориза - корневой симбиоз, при котором отдельные гифы, простирающиеся от мицелия гриба, колонизируют корни растения -хозяина.
- Микоризная сеть
- Физиология растений
- Ризосфера - область почвы вокруг корня под влиянием нынешних секреций корня и микроорганизмов
- Корневая резка
- Укоренящий порошок
- Столон
- Эффект Танады
- Тэпзот
Ссылки
[ редактировать ]- ^ Харли Макдональд и Донован Стивенс (3 сентября 2019 г.). Биотехнология и биология растений . Edtech. С. 141–. ISBN 978-1-83947-180-3 .
- ^ Нгуен, Линх Туй мой; Хоанг, Хан Т. Choi, Eunho; Парк, пил солнце (2023-07-05). «Распределение мангровых деревьев с различными морфологиями воздушных корней на участках аккреции и эрозии в провинции CA Mau, Вьетнам» . Устье, прибрежная и шельфская наука . 287 : 108324. Bibcode : 2023ecss..28708324N . doi : 10.1016/j.ecss.2023.108324 . ISSN 0272-7714 .
- ^ «Части растений = корни» . Университет Иллинойса .
- ^ Яакав Окон (24, 1993). Azispiase / Association . CRC Press. стр. 77-. ISBN 978-0-8493-4925-6 .
- ^ Jump up to: а беременный в «Садовник на заднем дворе: понимание корней растений» . Университет Аризоны кооперативное расширение .
- ^ Ganguleee HC, Das KS, Datta CT, Sen S. Botany College . Тол. 1. Калькутта: новое центральное книжное агентство.
- ^ Датта А.С., Датта Т.С. Ботаника для получения степени студентов (6 -е изд.). Издательство Оксфордского университета.
- ^ Шелдрак, Мерлин (2020). Запутанная жизнь . Бодли голова. п. 148. ISBN 978-1847925206 .
- ^ Malamy JE (2005). «Внутренние пути и экологические пути, которые регулируют архитектуру корневой системы» . Растение, клетки и окружающая среда . 28 (1): 67–77. doi : 10.1111/j.1365-3040.2005.01306.x . PMID 16021787 .
- ^ Jump up to: а беременный Caldwell MM, Dawson TE, Richards JH (январь 1998 г.). «Гидравлический подъем: последствия оттока воды от корней растений». Oecologia . 113 (2): 151–161. BIBCODE : 1998OECOL.113..151C . doi : 10.1007/s004420050363 . PMID 28308192 . S2CID 24181646 .
- ^ Fitter AH (1991). «Экологическая значимость архитектуры корневой системы: экономический подход». В Аткинсоне Д (ред.). Рост корня растений: экологическая перспектива . Блэквелл. С. 229–243.
- ^ Malamy JE, Ryan KS (ноябрь 2001 г.). «Экологическая регуляция бокового инициации корня у арабидопсиса» . Физиология растений . 127 (3): 899–909. doi : 10.1104/pp.010406 . PMC 129261 . PMID 11706172 .
- ^ Рассел П.Дж., Герц П.Е., Макмиллан Б. (2013). Биология: динамическая наука . Cengage Learning. п. 750. ISBN 978-1-285-41534-5 Полем Архивировано из оригинала 2018-01-21 . Получено 2017-04-24 .
- ^ «Suberin - обзор | темы ScienceDirect» . www.sciencedirect.com . Получено 2021-08-31 .
- ^ «Форма и функция Suberin - Марк Бернардс - Западный университет» . www.uwo.ca. Получено 2021-08-31 .
- ^ Jump up to: а беременный Ватанабе, Кохтаро; Нишючи, Шунсаку; Кулихихин, Константин; Наказоно, Микио (2013). «Способствует ли накопление Suberin в корнях растений на упущенную уплотнение?» Полем Границы в науке о растениях . 4 : 178. doi : 10.3389/fpls.2013.00178 . ISSN 1664-462x . PMC 3683634 . PMID 23785371 .
- ^ Ван ден Дрисс, Р. (1974-07-01). «Прогнозирование минерального статуса питательных веществ деревьев с помощью листового анализа» . Ботанический обзор . 40 (3): 347–394. Bibcode : 1974botrv..40..347V . doi : 10.1007/bf02860066 . ISSN 1874-9372 . S2CID 29919924 - Via Springer.
- ^ Накагава Y, Катагири Т., Шинозаки К., Ци З., Тацуми Х, Фуруичи Т. и др. (Февраль 2007 г.). «Арабидопсис плазматическая мембрана, решающий для притока CA2+ и прикосновения в корнях» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (9): 3639–44. Bibcode : 2007pnas..104.3639n . doi : 10.1073/pnas.0607703104 . PMC 1802001 . PMID 17360695 .
- ^ Механизм светодиода UV-B, обнаруженный в корнях растений , Университет штата Сан-Франциско, 8 декабря 2008 г.
- ^ Marchant A, Kargul J, May St, Muller P, Delbarre A, Perrot-Rechenmann C, Bennett MJ (апрель 1999 г.). «AUX1 регулирует корневой гравитропизм у арабидопсиса, облегчая поглощение ауксина в апикальных тканях корневых» . Embo Journal . 18 (8): 2066–73. doi : 10.1093/emboj/18.8.2066 . PMC 1171291 . PMID 10205161 .
- ^ Ходж А (июнь 2009 г.). «Корневые решения» . Растение, клетки и окружающая среда . 32 (6): 628–40. doi : 10.1111/j.1365-3040.2008.01891.x . PMID 18811732 .
- ^ Carminati A, Vetterlein D, Weller U, Vogel HJ, Oswald SE (2009). «Когда корни теряют контакт». Vadose Zone Journal . 8 (3): 805–809. Bibcode : 2009vzj ..... 8..805c . doi : 10.2136/vzj2008.0147 . S2CID 128600212 .
- ^ Чен Р., Розен Е., Массон П.Х. (июнь 1999 г.). «Гравиотропизм у высших растений» . Физиология растений . 120 (2): 343–50. doi : 10.1104/pp.120.2.343 . PMC 1539215 . PMID 11541950 .
- ^ Фуси, Риккардо; Розиньоли, Серена; Лу, Хаою; Сангиорги, Джузеппе; Бовина, Риккардо; Паттем, Джейкоб К.; Borkar, Aditi N.; Ломбарди, Марко; Форестанец, Кристиан; Милнер, Сара Г.; Дэвис, Джейн Л.; Лейл, Анеш; Киршнер, Гвендолин К.; Сваруп, Ранджан; Тассинари, Альберто; Пандей, Бипин К.; Йорк, Ларри М.; Аткинсон, Брайан С.; Старрок, Крейг Дж.; Муни, Сача Дж.; Hochholdinger, Frank; Такер, Мэтью Р.; Гиммельбах, Аксель; Стейн, Нильс; Машер, Мартин; Нагель, Керстин А.; Де Гара, Лора; Симмондс, Джеймс; Uauy, Cristobal; Tuberosa, Роберто; Линч, Джонатан П.; Якубов, Глеб Э.; Беннетт, Малкольм Дж.; Бхосале, Рахул; Сальви, Сильвио (2 августа 2022 г.). «Угол корня контролируется EGT1 в зерновых культурах, использующих антигравитропный механизм» . Труды Национальной академии наук . 119 (31): E2201350119. Bibcode : 2022pnas..11901350F . doi : 10.1073/pnas.2201350119 . PMC 9351459 . PMID 35881796 . S2CID 251104211 .
- ^ Пандей, Бипин К.; Хуан, Гоцян; Бхосале, Рахул; Хартман, Сьон; Старрок, Крейг Дж.; Хосе, Лотти; Мартин, Оливье С.; Каради, Михал; Восенек, Лауреню А.С. Лджунг, Карин; Линч, Джонатан П. (2021-01-15). «Корни растений ощущают уплотнение почвы с помощью ограниченной диффузии этилена» . Наука . 371 (6526): 276–280. Bibcode : 2021sci ... 371..276p . doi : 10.1126/science.abf3013 . HDL : 1874/418726 . ISSN 0036-8075 . PMID 33446554 . S2CID 231606782 .
- ^ Jump up to: а беременный в дюймовый и Солсбери Ф.Дж., Холл А., Грисон К.С., Хэллидей К.Дж. (май 2007). «Фитохрома координирует арабидопсис, выстрел и развитие корней» . Заводский журнал . 50 (3): 429–38. doi : 10.1111/j.1365-313X.2007.03059.x . PMID 17419844 .
- ^ Jump up to: а беременный в дюймовый и Van Gelderen K, Kang C, Paalman R, Keuskamp D, Hayes S, Pierik R (январь 2018 г.). «Обнаружение дальнего красного света в побеге регулирует развитие бокового корня через фактор транскрипции Hy5» . Растительная ячейка . 30 (1): 101–116. doi : 10.1105/tpc.17.00771 . PMC 5810572 . PMID 29321188 .
- ^ Ай, Qiaoli; Зенг, Бо; Лю, Цзяньхуи; Ли, Сики; Ван Бодегом, Питер М.; Корнелиссен, Йоханнес ХК (октябрь 2016 г.). «Абсорбция кислорода от случайных корней способствует выживанию полностью погруженных в наземные растения» . Анналы ботаники . 118 (4): 675–683. doi : 10.1093/aob/mcw051 . PMC 5055620 . PMID 27063366 .
- ^ Лин, Чен; Огорек, Лукас Леон Перальта; Лю, Дэн; Педерсен, Оле; Sauter, Маргрет (11 января 2023 г.). «Локус количественного признака, придающий устойчивость к наводнениям к глубоководному рису, регулирует образование двух разных типов водных случайных корней» . Новый фитолог . 238 (4): 1403–1419. doi : 10.1111/nph.18678 . PMID 36519256 . S2CID 254769152 .
- ^ Марич, Аида; Хартман, Сьон (11 марта 2023 г.). «Листовая оболочка способствует длительной защите от наводнения, вызывая специализированные случайные корни» . Новый фитолог . 238 (4): 1337–1339. doi : 10.1111/nph.18824 . PMID 36905344 . S2CID 257438007 .
- ^ Jump up to: а беременный Новак EJ, Martin CE (1997). «Физиологические и анатомические реакции на дефицит воды в камере Epiphyte Tillandsia Ionantha (Bromeliaceae)» . Международный журнал растительных наук . 158 (6): 818–826. doi : 10.1086/297495 . HDL : 1808/9858 . JSTOR 2475361 . S2CID 85888916 .
- ^ Nadkarni NM (ноябрь 1981). «Корни куполов: сходящаяся эволюция в циклах питательных веществ в тропических лесах». Наука . 214 (4524): 1023–4. Bibcode : 1981sci ... 214.1023n . doi : 10.1126/science.214.4524.1023 . PMID 17808667 . S2CID 778003 .
- ^ Пюц Н. (2002). «Сокращенные корни». В Waisel Y., Eshel A., Kafkafi U. (Eds.). Корни растений: скрытая половина (3 -е изд.). Нью -Йорк: Марсель Деккер. С. 975–987.
- ^ Canadell J, Jackson RB, Ehleringer JB, Mooney HA, Sala Oe, Schulze Ed (декабрь 1996 г.). «Максимальная глубина укоренения типов растительности в глобальном масштабе». Oecologia . 108 (4): 583–595. Bibcode : 1996oecol.108..583c . doi : 10.1007/bf00329030 . PMID 28307789 . S2CID 2092130 .
- ^ Stonea el, Kaliszb PJ (1 декабря 1991 г.). «В максимальной степени корней деревьев». Лесная экология и управление . 46 (1–2): 59–102. Bibcode : 1991forem..46 ... 59 с . doi : 10.1016/0378-1127 (91) 90245-q .
- ^ Retallack GJ (1986). «Окаменечные записи почв» (PDF) . В Wright VP (ред.). Палеосолы: их признание и интерпретация . Оксфорд: Блэквелл. С. 1–57. Архивировано (PDF) из оригинала 2017-01-07.
- ^ Хиллиер Р., Эдвардс Д., Моррисси Л.Б. (2008). «Седиментологические доказательства корневых структур в раннем девонском англо -Уэлшском бассейне (Великобритания) с спекуляциями на их производителях». Палеогеография, палеоклиматология, палеоэкология . 270 (3–4): 366–380. Bibcode : 2008ppp ... 270..366h . doi : 10.1016/j.palaeo.2008.01.038 .
- ^ Амрам Эшель; Том Бекман (17 апреля 2013 г.). Корни растений: скрытая половина, четвертое издание . CRC Press. С. 1-. ISBN 978-1-4398-4649-0 .
- ^ Курата, Тецуя (1997). «Светослойное удлинение корня у Arabidopsis thaliana». Журнал физиологии растений . 151 (3): 345–351. Bibcode : 1997jpphy.151..346K . doi : 10.1016/s0176-1617 (97) 80263-5 . HDL : 2115/44841 .
- ^ Postgate, J. (1998). Фиксация азота (3 -е изд.). Кембридж, Великобритания: издательство Кембриджского университета.
- ^ Лал, Раттан (2006). Энциклопедия почвенной науки . CRC Press. ISBN 978-0-8493-5054-2 .
- ^ Чамовиц, Даниэль. (21 ноября 2017 г.). Что за растение знает: полевое руководство по чувствам . Фаррар, Страус и Жиру. ISBN 9780374537128 Полем OCLC 1041421612 .
- ^ Фалик О., Мордох Ю., Бен-Натан Д., Вануну М., Гольдштейн О., Новопланский А. (июль 2012 г.). «Отзывчивость растений на общение с корневым корнем от стрессовых сигналов» . Анналы ботаники . 110 (2): 271–80. doi : 10.1093/aob/mcs045 . PMC 3394639 . PMID 22408186 .
- ^ Боуэн Г.Д., Ровира А.Д. (1976). «Микробная колонизация корней растений». Анну. Преподобный Фитопатол . 14 : 121–144. doi : 10.1146/annurev.py.14.090176.001005 .
- ^ Корни растений и их окружающая среда . Elsevier. 1988. с. 17
- ^ Корни растений и их окружающая среда . Elsevier. 1988. с. 25
- ^ Zahniser, David (2008-02-21). "Город, чтобы пройти деньги на тротуары?" Полем Лос -Анджелес Таймс . Получено 2023-03-30 .
Дальнейшее чтение
[ редактировать ]- Baldocchi DD, Xu L (октябрь 2007 г.). «Что ограничивает испарение из Средиземноморского дубового леса - поставки влаги в почве, физиологический контроль растениями или спрос на атмосферу?». Достижения в водных ресурсах . 30 (10): 2113–22. Bibcode : 2007adwr ... 30.2113b . doi : 10.1016/j.advwatres.2006.06.013 .
- Brundrett, MC (2002). "Коэволюция корней и микоризы наземных растений" . Новый фитолог . 154 (2): 275–304. doi : 10.1046/j.1469-8137.2002.00397.x . PMID 33873429 .
- Кларк Л. (2004). «Основная структура и развитие корня - заметки лекции» (PDF) . Архивировано из оригинала (PDF) 3 января 2006 года.
- Coutts MP (1987). «Процессы развития в корневых системах деревьев». Канадский журнал лесных исследований . 17 (8): 761–767. doi : 10.1139/x87-122 .
- Raven JA, Edwards D (2001). «Корни: эволюционное происхождение и биогеохимическое значение». Журнал экспериментальной ботаники . 52 (Suppl 1): 381–401. doi : 10.1093/jxb/52.suppl_1.381 . PMID 11326045 .
- Schenk HJ, Jackson RB (2002). «Глобальная биогеография корней». Экологические монографии . 72 (3): 311–328. doi : 10.2307/3100092 . JSTOR 3100092 .
- Саттон Р.Ф., Тунус Р.В. (1983). «Терминология корневой и корневой системы». Лесная научная монография . 24 : 137.
- Филлипс У.С. (1963). «Глубина корней в почве». Экология . 44 (2): 424. Bibcode : 1963ecol ... 44..424p . doi : 10.2307/1932198 . JSTOR 1932198 .
- Caldwell MM, Dawson TE, Richards JH (1998). «Гидравлический подъем: последствия оттока воды от корней растений». Oecologia . 113 (2): 151–161. BIBCODE : 1998OECOL.113..151C . doi : 10.1007/s004420050363 . PMID 28308192 . S2CID 24181646 .