Конвергентная эволюция


| Часть серии на |
| Эволюционная биология |
|---|
 |
Конвергентная эволюция - это независимая эволюция аналогичных признаков у видов разных периодов или эпох во времени. Конвергентная эволюция создает аналогичные структуры , которые имеют сходную форму или функцию, но не присутствовали в последнем общем предке этих групп. Кладистический - термин для того же явления гомоплазия . Рецидивирующая эволюция полета является классическим примером, поскольку летающие насекомые , птицы , птерозавры и летучие мыши независимо развивали полезную вместимость полета. Функционально сходные признаки, которые возникли через конвергентную эволюцию, являются аналогичными , тогда как гомологичные структуры или признаки имеют общее происхождение, но могут иметь разнородные функции. птиц, летучей мыши и птерозавра Крылья являются аналогичными структурами, но их передние конечности гомологичны, разделяя наследственное состояние, несмотря на выполнение различных функций.
Противоположностью конвергенции является дивергентная эволюция , где родственные виды эволюционируют различные признаки. Конвергентная эволюция аналогична параллельной эволюции , которая возникает, когда два независимых вида эволюционируют в одном и том же направлении и, таким образом, независимо приобретают аналогичные характеристики; Например, скользящие лягушки развивались параллельно из нескольких видов лягушки деревьев .
Многие случаи конвергентной эволюции известны у растений , в том числе повторное развитие C 4 фотосинтеза , рассеивание семян мясистыми фруктами, адаптированными для употребления животных, и плотоядного .
Обзор
[ редактировать ]
В морфологии аналогичные признаки возникают, когда разные виды живут аналогичными способами и/или аналогичной средой, и поэтому сталкиваются с одинаковыми факторами окружающей среды. Принимая подобные экологические ниши (то есть отличительный образ жизни), подобные проблемы могут привести к аналогичным решениям. [ 1 ] [ 2 ] [ 3 ] Британский анатомист Ричард Оуэн был первым, кто выявил фундаментальную разницу между аналогиями и гомологией . [ 4 ]
В биохимии физические и химические ограничения на механизмах вызвали некоторые активные расположения, такие как каталитическая триада, эволюционировать независимо в отдельных суперсемейных ферментах . [ 5 ]
В своей книге 1989 года « Прекрасная жизнь » Стивен Джей Гулд утверждал, что, если бы он мог «перемотать ленту жизни [и] те же условия, которые снова сталкивались, эволюция могла бы пройти совершенно другой курс». [ 6 ] Саймон Конвей Моррис оспаривает этот вывод, утверждая, что конвергенция является доминирующей силой в эволюции, и, учитывая, что те же самые экологические и физические ограничения на работе, жизнь неизбежно будет развиваться в направлении «оптимального» плана тела, и в какой -то момент эволюция связана Чтобы наткнуться на разум, черта, в настоящее время идентифицированная, по крайней мере, с приматами , корвидами и китообразными . [ 7 ]
Различия
[ редактировать ]Кладистика
[ редактировать ]В кладистике гомоплазия - это черта, разделяемая двумя или более таксонами по любой причине, кроме того, что они имеют общее происхождение. Таксоны, которые делятся происхождением, являются частью одной и той же клады ; Кладистика стремится организовать их в соответствии с их степенью родства, чтобы описать их филогения . Таким образом, гомопластические признаки, вызванные конвергенцией, с точки зрения кладистики, смешанные факторы, которые могут привести к неправильному анализу. [ 8 ] [ 9 ] [ 10 ] [ 11 ]
Атавизм
[ редактировать ]В некоторых случаях трудно сказать, была ли утерянная черта, а затем повторно развивалась слюне, или был ли просто отключен ген, а затем повторно включена позже. Такая повторная черта называется атавизмом . С математической точки зрения, неиспользованный ген ( селективно нейтральный ) имеет постоянно снижающуюся вероятность сохранения потенциальной функциональности с течением времени. Шкала времени этого процесса сильно варьируется в разных филогениях; У млекопитающих и птиц существует разумная вероятность остаться в геноме в потенциально функциональном состоянии в течение около 6 миллионов лет. [ 12 ]
Параллель против конвергентной эволюции
[ редактировать ]
Когда два вида схожи по конкретному характеру, эволюция определяется как параллельная, если предки также были похожи и сходились, если они не были. [ B ] Некоторые ученые утверждают, что существует континуум между параллельной и сходящейся эволюцией, [ 13 ] [ 14 ] [ 15 ] [ 16 ] В то время как другие утверждают, что, несмотря на некоторое совпадение, между ними все еще существуют важные различия. [ 17 ] [ 18 ]
Когда наследственные формы не определены или неизвестны, или диапазон рассматриваемых признаков не четко указан, различие между параллельной и сходящейся эволюцией становится более субъективным. Например, поразительный пример аналогичных плацентарных и сумчатых форм описывается Ричардом Докинсом в слепой сторожи как случай сходящейся эволюции, потому что млекопитающие на каждом континенте имели длинную эволюционную историю до исчезновения динозавров, под которыми нужно накопить актуальность различия. [ 19 ]
На молекулярном уровне
[ редактировать ]
Белки
[ редактировать ]Протеаза активные сайты
[ редактировать ]Фермология протеаз обеспечивает некоторые из самых ярких примеров конвергентной эволюции. Эти примеры отражают внутренние химические ограничения на ферменты, ведущие эволюцию для сходимости на эквивалентных решениях независимо и неоднократно. [ 5 ] [ 20 ]
Протеазы серина и цистеина используют различные аминокислотные функциональные группы (спирт или тиол) в качестве нуклеофила . Чтобы активировать этот нуклеофил, они ориентированы на кислотный и основной остаток в каталитической триаде . Химические и физические ограничения на ферментное катализ вызвали идентичные триадные композиции независимо более 20 раз в различных суперсемейных ферментах . [ 5 ]
Треониновые протеазы используют аминокислотный треонин в качестве каталитического нуклеофила . В отличие от цистеина и серина, треонин является вторичным спиртом (т.е. имеет метильную группу). Метильная группа треонина значительно ограничивает возможные ориентации триады и субстрата, как метиловые столкновения либо с основным цепью фермента, либо основом гистидина. Следовательно, большинство треониновых протеаз используют N-концевой треонин, чтобы избежать таких стерических столкновений . Несколько эволюционно независимых ферментных суперсемей с различными белковыми складками используют N-концевой остаток в качестве нуклеофила. Эта общность активного сайта , но разность белкового склада указывает на то, что активный сайт развивался слюне в этих семействах. [ 5 ] [ 21 ]
Конусная улитка и рыба инсулин
[ редактировать ]Conus Geographus производит отдельную форму инсулина , которая больше похожа на последовательности белка инсулина рыб, чем с инсулином из более близкородных моллюсков, что предполагает конвергентную эволюцию, [ 22 ] хотя с возможностью горизонтального переноса генов . [ 23 ]
Поглощение железа железа с помощью белковых переносчиков на наземных растениях и хлорофитах
[ редактировать ]Отдаленные гомологи ионных перевозчиков металлов молнии на наземных растениях и хлорофитах сходились в структуре, вероятно, займет Fe 2+ эффективно. Белки IRT1 из Arabidopsis thaliana и риса имеют чрезвычайно разные аминокислотные последовательности от Chlamydomonas IRT1 , но их трехмерные структуры схожи, что указывает на сходящуюся эволюцию. [ 24 ]
НА + К. + -Тфаза и устойчивость к насекомым к кардиотоническим стероидам
[ редактировать ]Многие примеры конвергентной эволюции существуют у насекомых с точки зрения развития сопротивления на молекулярном уровне к токсинам. Одним из хорошо охарактеризованных примеров является эволюция устойчивости к кардиотоническим стероидам (CTSS) через аминокислотные замены в четко определенных положениях α-субъединицы NA + К. + -Atpase (Атпальфа). Различия в Атпальфе были обследованы у различных видов, адаптированных CTS, охватывающих шесть заказов насекомых. [ 25 ] [ 26 ] [ 27 ] Среди 21, адаптированных CTS вида 58 (76%) из 76 аминокислотных замен на участках, участвующих в сопротивлении CTS, встречаются параллельно по меньшей мере в двух линиях. [ 27 ] 30 из этих замен (40%) встречаются только в двух участках в белке (положения 111 и 122). Адаптированные на CTS виды также регулярно развивали неофункционализированные дупликации Atpalpha с конвергентными тканеспецифическими паттернами экспрессии. [ 25 ] [ 27 ]
Нуклеиновые кислоты
[ редактировать ]Конвергенция происходит на уровне ДНК и аминокислотных последовательностей, продуцируемых путем транспортировки структурных генов в белки . Исследования обнаружили конвергенцию в аминокислотных последовательностях в эхолокации летучих мышей и дельфином; [ 28 ] среди морских млекопитающих; [ 29 ] между гигантскими и красными пандами; [ 30 ] и между тилацином и канидами. [ 31 ] Конвергенция также была обнаружена в типе некодирующей ДНК , цис-регуляторных элементов , таких как в их скоростях эволюции; Это может указывать либо на положительный отбор , либо смягченный очищающий выбор . [ 32 ] [ 33 ]
В морфологии животных
[ редактировать ]
Тела
[ редактировать ]Плавательные животные, в том числе рыбы , такие как сельди , морские млекопитающие, такие как дельфины и ихтиозавры ( мезозойских ), все сходились в одной и той же обтекаемой форме. [ 34 ] [ 35 ] Аналогичная форма и адаптация плавания даже присутствуют в моллюсках, таких как Phyllireoe . [ 36 ] Фузообразное телосложение (трубка, сужающаяся на обоих концах), принятая многими водными животными, является адаптацией, позволяющей им проходить на высокой скорости в среде высокой сопротивления . [ 37 ] Подобные формы тела находятся в уплотнениях без уха и ушах : у них все еще есть четыре ноги, но они сильно модифицированы для плавания. [ 38 ]
Комнатная фауна Австралии и плацентарные млекопитающие старого мира имеют несколько поразительно похожих форм, разработанных в двух кладах, изолированных друг от друга. [ 7 ] Тело, и особенно форма черепа тилацина ( тасманский тигр или тасманский волк) сходились с таковыми у Canidae, таких как красная лиса, vulpes vulpes . [ 39 ]
- Сближение сумчатых и плацентарных млекопитающих
-
 Red Fox Skeleton
Red Fox Skeleton -
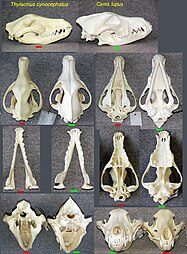 Черепа тилацина (слева), древесный волк (справа)
Черепа тилацина (слева), древесный волк (справа) -
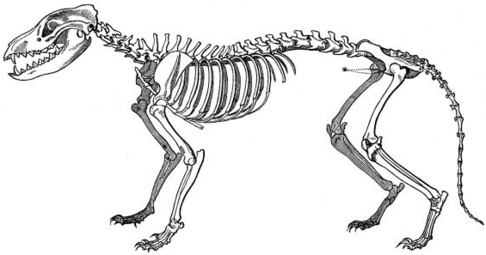 Тилациновый скелет
Тилациновый скелет



Эхолокация
[ редактировать ]В качестве сенсорной адаптации эхолокация развивалась отдельно у китообразных (дельфинов и китов) и летучих мышей, но из тех же генетических мутаций. [ 40 ]
Электрические рыбы
[ редактировать ]Gymnotiformes of Южной Америки и Mormyridae Africa независимо развивались пассивной электрорецепцией (около 119 и 110 миллионов лет назад соответственно). Примерно через 20 миллионов лет после приобретения этой способности обе группы развивали активный электрогенез , производя слабые электрические поля, чтобы помочь им обнаружить добычу. [ 41 ]
- Конвергенция слабых электрических рыб
-
 Гимнотриформная форма волны электролокации
Гимнотриформная форма волны электролокации -
 Гимнообразовательная электрическая рыба из Южной Америки
Гимнообразовательная электрическая рыба из Южной Америки -
 Мормиридная электрическая рыба Африки
Мормиридная электрическая рыба Африки -
Мормиридная электрополовация форма волны


Глаза
[ редактировать ]
Одним из самых известных примеров сходящейся эволюции являются глаз камеры головоногих (таких как кальмар и осьминог), позвоночные (включая млекопитающих) и Cnidaria (например, медуза). [ 43 ] У их последнего общего предка было больше всего простого фоторецептивного места, но ряд процессов привел к прогрессивной уточненности глаз камеры - с одним резким отличием: глаз Цефалопод «проводной» в противоположном направлении, с кровью и нервными сосудами, вступающими из Задняя часть сетчатки, а не передней, как у позвоночных. В результате у позвоночных есть слепое пятно . [ 7 ]
Полет
[ редактировать ]
У птиц и летучих мышей есть гомологичные конечности, потому что они оба в конечном итоге получены из наземных тетрапод , но их механизмы полета только аналогичны, поэтому их крылья являются примерами функциональной конвергенции. Две группы независимо развивали свои собственные средства питания. Их крылья существенно различаются в строительстве. Крыло летучих мышей - это мембрана, протянутая на четыре чрезвычайно удлиненных пальца и ноги. Аэродинамический профиль крыла птичьего крыла изготовлен из перьев , сильно прикрепленных к предплечью (локтевой кости) и сильно сплавленными костями запястья и руки ( Carpometacarpus ), с оставшимися только крошечными оставшимися оставшимися пальцами. Таким образом, в то время как крылья летучих мышей и птиц функционально сходится, они не являются анатомически сходящимися. [ 3 ] [ 44 ] Птицы и летучие мыши также имеют высокую концентрацию цереброзидов в коже их крыльев. Это повышает гибкость кожи, черту, полезную для летающих животных; Другие млекопитающие имеют гораздо более низкую концентрацию. [ 45 ] Вымершие птерозавры независимо развивали крылья из их передних и задних конечностей, в то время как у насекомых есть крылья , которые развивались отдельно от разных органов. [ 46 ]
Летающие белки и сахарные планеры очень похожи в планах их тела, с скользящими крыльями, растянутыми между их конечностями, но летающие белки - это плацентарные млекопитающие, в то время как сахарные планеры - это сумчатые, широко отделенные в линии млекопитающих от плаценлов. [ 47 ]
Humbrybird Hawk-Moths и колибри развили аналогичные схемы полета и кормления. [ 48 ]
Насекомого рта
[ редактировать ]Горничные чашки насекомых показывают много примеров сходящейся эволюции. Горничные части разных групп насекомых состоят из набора гомологичных органов, специализирующихся на потреблении диетической группы этой группы насекомых. Конвергентная эволюция многих групп насекомых, приводящих от оригинальных кусочков, к разным, более специализированным, производным типам функций. К ним относятся, например, хоботок насекомых с цветами, такими как пчелы и цветочные жуки , [ 49 ] [ 50 ] [ 51 ] Или кусочные каракули от насекомых, взлеченных кровью, таких как блохи и комары .
Противоположные большие пальцы
[ редактировать ]Противоположные большие пальцы, позволяющие схватить объекты, чаще всего ассоциируются с приматами , такими как люди и другие обезьяны, обезьяны и лемуры. Противоположные большие пальцы также развивались в гигантских пандах , но они совершенно разные по структуре, имея шесть пальцев, включая большой палец, который развивается из кости запястья, полностью отдельно от других пальцев. [ 52 ]
Приматы
[ редактировать ] Несмотря на подобное освещение цвета кожи после выхода из Африки , различные гены участвовали в европейских (левых) и восточноазиатских (справа). Несмотря на подобное освещение цвета кожи после выхода из Африки , различные гены участвовали в европейских (левых) и восточноазиатских (справа).
| ||



Конвергентная эволюция у людей включает в себя синий цвет глаз и светлый цвет кожи. [ 53 ] Когда люди мигрировали из Африки , они переехали в более северные широты с менее интенсивным солнечным светом. [ 53 ] Для них было полезно уменьшить пигментацию кожи . [ 53 ] Похоже, что было некоторое освещение цвета кожи до того, как европейские и восточноазиатские линии расходились, так как существуют некоторые генетические различия, которые являются общими для обеих групп. [ 53 ] Тем не менее, после того, как линии расходились и стали генетически изолированными, кожа обеих групп больше осветилась, и что дополнительное освещение было связано с различными генетическими изменениями. [ 53 ]
| Люди | Лемуры | ||
|---|---|---|---|
| Несмотря на сходство внешнего вида, генетическая основа голубых глаз отличается у людей и лемуров . | |||



Лемуры и люди оба являются приматами. Приматы предков были карие глаза, как это делают большинство приматов сегодня. Генетическая основа синих глаз у людей была подробно изучена, и об этом много известно. Это не тот случай, когда один ген локус отвечает, скажем, с коричневым доминирующим до синего цвета глаз . Тем не менее, один локус отвечает за около 80% вариаций. У лемуров различия между синими и карими глазами не совсем известны, но тот же генный локус не вовлечен. [ 54 ]
В растениях
[ редактировать ]
Годовой жизненный цикл
[ редактировать ]В то время как большинство видов растений многолетние , около 6% следуют ежегодному жизненному циклу, живя только в течение одного вегетационного периода. [ 55 ] Ежегодный жизненный цикл независимо появлялся в более чем 120 семействах покрытосеменных растений. [ 56 ] [ 57 ] Распространенность годовых видов увеличивается в жарких летних условиях в четырех богатых видах семейства однолетних ( Asteraceae , Brassicaceae , Fabaceae и Poaceae ), что указывает на то, что годовой жизненный цикл является адаптивным. [ 55 ] [ 58 ]
Углеродная фиксация
[ редактировать ]C 4 Фотосинтез , один из трех основных биохимических процессов с фиксированием углерода, возник независимо до 40 раз . [ 59 ] [ 60 ] Около 7600 видов растений покрытосеменных используют C 4 углеродную фиксацию, со многими монокотами, включая 46% трав, таких как кукуруза и сахарный тростник , [ 61 ] [ 62 ] и дикоты, включая несколько видов в Chenopodiaceae и Amaranthaceae . [ 63 ] [ 64 ]
Фрукты
[ редактировать ]Фрукты с широким спектром структурных происхождений сходились, чтобы стать съедобным. Яблоки представляют собой четки с пятью ковром ; Их вспомогательные ткани образуют ядро яблока, окруженное структурами извне ботанических фруктов, сосуда или гипоантия . Другие съедобные фрукты включают другие растительные ткани; [ 65 ] Мятистая часть помидора - это стены перикарпа . [ 66 ] Это подразумевает конвергентную эволюцию под селективным давлением, в этом случае конкуренция за рассеивание семян животными за счет потребления мясистых фруктов. [ 67 ]
Расездка семян муравьями ( Myrmecochory ) развивалась независимо более чем 100 раз и присутствует у более чем 11 000 видов растений. Это один из самых драматических примеров конвергентной эволюции в биологии. [ 68 ]
Плотоловый
[ редактировать ]
Carnivory эволюционировал несколько раз независимо у растений в широко отделенных группах. У трех изученных видов Cephalotus follicularis , Nepenthes alata и Sarracenia purpurea была сходимость на молекулярном уровне. Хитройные растения выделяют ферменты в пищеварительную жидкость, которую они производят. Изучая фосфатазу , гликозид -гидролазу , глюканазы , рНтазы и хитиназы , а также белок, связанный с патогенезом , и белок, связанный с тауматином , авторы обнаружили много сходящихся аминокислотных замены. Эти изменения были не в каталитических участках ферментов, а скорее на обнаженных поверхностях белков, где они могут взаимодействовать с другими компонентами клетки или пищеварительной жидкости. Авторы также обнаружили, что гомологичные гены в непрерывном растении Arabidopsis thaliana, как правило, повышают их экспрессию, когда растение подвергается стрессу, что заставляет авторов предположить, что чувствительные к стрессу белки часто кооптины [ C ] В повторяющейся эволюции плотояда. [ 69 ]
Методы вывода
[ редактировать ]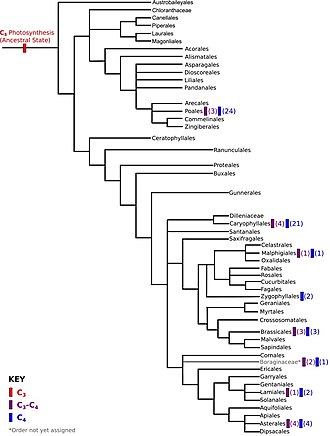
Филогенетическая реконструкция и реконструкция наследственного состояния продолжаются, предполагая, что эволюция произошла без сходимости. Однако сходящиеся паттерны могут появляться на более высоких уровнях в филогенетической реконструкции и иногда явно ищут исследователи. Методы, применяемые для вывода конвергентной эволюции, зависят от того, ожидается ли сходимость на основе процессов или на основе процессов. Конвергенция на основе рисунков является более широким термином, когда два или более линии независимо развивают паттерны сходных признаков. Конвергенция на основе процессов-это когда конвергенция обусловлена схожими силами естественного отбора . [ 70 ]
Измерения на основе шаблонов
[ редактировать ]Более ранние методы измерения конвергенции включают соотношения фенотипического и филогенетического расстояния путем моделирования эволюции с моделью эволюции черт Браун -движения вдоль филогения. [ 71 ] [ 72 ] Более поздние методы также количественно определяют силу сходимости. [ 73 ] Одним из недостатков, который следует иметь в виду, является то, что эти методы могут путать долгосрочный стаз с конвергенцией из-за фенотипического сходства. Стазис происходит, когда среди таксонов мало эволюционных изменений. [ 70 ]
Измерения на расстоянии оценивают степень сходства между линиями с течением времени. Измерения на основе частоты оценивают количество линий, которые развивались в конкретном пространстве признаков. [ 70 ]
Основанные на процессах меры
[ редактировать ]Методы, чтобы вывести модели с конвергенцией на основе процессов, подходящие для отбора для филогении и непрерывных данных о признаках, чтобы определить, действовали ли те же селективные силы на линии. Это использует процесс Ornstein -Uhlenbeck для проверки различных сценариев отбора. Другие методы зависят от априорной спецификации того, где произошли сдвиги в отборе. [ 74 ]
Смотрите также
[ редактировать ]- Неполная сортировка линии - характерно для филогенетического анализа: наличие множественных аллелей в наследственных популяциях может привести к впечатлению, что сходящаяся эволюция произошла.
- Карцинизация -эволюция ракообразных в крабовые формы
- Морфология (биология) - Изучение внешних форм и структур организмов
- Итеративная эволюция - повторная эволюция определенной черты или плана тела из одной и той же происхождения происхождения в разные моменты времени.
- Элвис Таксон - неправильная идентификация более позднего таксона, напоминающего более ранние вымершие таксон
- Размножение обратно - форма селективного размножения для воссоздания признаков вымершего вида, но геном будет отличаться от исходных видов.
- Ортогенез (в отличие от конвергентной эволюции; включает в себя телеологию)
- Непредвиденные обстоятельства (эволюционная биология) - влияние эволюционной истории на результаты
Примечания
[ редактировать ]- ^ Тем не менее, эволюционная биология развития выявила глубокую гомологию между планами тела насекомых и млекопитающих, к удивлению многих биологов.
- ^ Тем не менее, все организмы имеют общего предка более или менее недавно, поэтому вопрос о том, как далеко, чтобы выглядеть в эволюционное время и насколько похожи предки должны быть для того, чтобы параллельная эволюция имела место, не полностью разрешен в эволюционном биология.
- ^ Предыдущее существование подходящих структур было названо предварительной адаптацией или эксплуатацией .
Ссылки
[ редактировать ]- ^ Кирк, Джон Томас Осмонд (2007). Наука и уверенность . CSIRO Publishing. п. 79. ISBN 978-0-643-09391-1 Полем Архивировано из оригинала 15 февраля 2017 года . Получено 23 января 2017 года .
Эволюционная конвергенция, которая, цитируя .. Саймон Конвей Моррис. - это «повторяющаяся тенденция биологической организации приходить к тому же« решению »к конкретной« потребности ». .. «Тасманский тигр» .. выглядел и вел себя как волк и занимал подобную экологическую нишу, но на самом деле был сумчатым, а не плацентарным млекопитающим.
- ^ Рис, Дж.; Meyers, N.; Urry, L.; Каин, М.; Wasserman, S.; Minorsky, P.; Джексон, Р.; Кук, Б. (5 сентября 2011 г.). Биология Cambell, 9 -е издание . Пирсон. п. 586. ISBN 978-1-4425-3176-5 .
- ^ Jump up to: а беременный «Гомологии и аналогии» . Калифорнийский университет Беркли. Архивировано с оригинала 19 ноября 2016 года . Получено 10 января 2017 года .
- ^ Тунстад, Эрик (2009). Теория Дарвина, эволюция в течение 400 лет (на норвежском языке). Осло, Норвегия: гуманистический Форлаг. P. 978-82-92622-53-7 .
- ^ Jump up to: а беременный в дюймовый Buller, AR; Таунсенд, Калифорния (19 февраля 2013 г.). «Внутренние эволюционные ограничения на структуру протеазы, ацилирование ферментов и идентичность каталитической триады» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (8): E653–61. BIBCODE : 2013PNAS..110E.653B . doi : 10.1073/pnas.1221050110 . PMC 3581919 . PMID 23382230 .
- ^ Гулд, С.Дж. (1989). Прекрасная жизнь: сланец Берджесса и природа истории . WW Norton. С. 282–285 . ISBN 978-0-09-174271-3 .
- ^ Jump up to: а беременный в Конвей Моррис, Саймон (2005). Жизненное решение: неизбежные люди в одинокой вселенной . Издательство Кембриджского университета. С. 164, 167, 170 и 235 . ISBN 978-0-521-60325-6 Полем OCLC 156902715 .
- ^ Chirat, R.; Моултон, де; Горили, А. (2013). «Механическая основа морфогенеза и конвергентной эволюции колючих ракушек» . Труды Национальной академии наук . 110 (15): 6015–6020. Bibcode : 2013pnas..110.6015c . doi : 10.1073/pnas.1220443110 . PMC 3625336 . PMID 23530223 .
- ^ Ломолино, м; Загадка, б; Whittaker, R; Браун, Дж. (2010). Биогеография, четвертое издание . Sinauer Associates. п. 426. ISBN 978-0-87893-494-2 .
- ^ Западный Эберхард, Мэри Джейн (2003). Пластичность и эволюция развития . Издательство Оксфордского университета. С. 353–376. ISBN 978-0-19-512235-0 .
- ^ Сандерсон, Майкл Дж.; Hufford, Larry (1996). Гомоплазия: рецидив сходства в эволюции . Академическая пресса. С. 330, и Passim. ISBN 978-0-08-053411-4 Полем Архивировано из оригинала 14 февраля 2017 года . Получено 21 января 2017 года .
- ^ Коллин, Р.; Cipriani, R. (2003). «Закон Долла и повторная эволюция оболочки» . Труды Королевского общества б . 270 (1533): 2551–2555. doi : 10.1098/rspb.2003.2517 . PMC 1691546 . PMID 14728776 .
- ^ Арендт, J; Резник, D (январь 2008 г.). «Конвергенция и параллелизм пересмотрели: что мы узнали о генетике адаптации?». Тенденции в экологии и эволюции . 23 (1): 26–32. Bibcode : 2008tecoe..23 ... 26a . doi : 10.1016/j.tree.2007.09.011 . PMID 18022278 .
- ^ Уотерс, Джонатан М.; МакКаллох, Грэм А. (2021). «Изобретение колеса? Пересмотр роли потока генов, сортировки и сходимости в повторной эволюции». Молекулярная экология . 30 (17): 4162–4172. Bibcode : 2021molec..30.4162W . doi : 10.1111/mec.16018 . ISSN 1365-294X . PMID 34133810 . S2CID 235460165 .
- ^ Cerca, Хосе (октябрь 2023 г.). «Понимание естественного отбора и сходства: сходящиеся, параллельные и повторные эволюции». Молекулярная экология . 32 (20): 5451–5462. Bibcode : 2023molec..32.5451c . doi : 10.1111/mec.17132 . PMID 37724599 .
- ^ Bohutínská, Магдалена; Пейхель, Кэтрин Л. (апрель 2024 г.). «Время дивергенции формирует повторное использование гена во время повторной адаптации». Тенденции в экологии и эволюции . 39 (4): 396–407. Bibcode : 2024tecoe..39..396b . doi : 10.1016/j.tree.2023.11.007 . PMID 38155043 .
- ^ Пирс Т. (10 ноября 2011 г.). «Конвергенция и параллелизм в эволюции: нео-голландский аккаунт» . Британский журнал «Философия науки» . 63 (2): 429–448. doi : 10.1093/bjps/axr046 .
- ^ Zhang, J.; Кумар С. (1997). «Обнаружение конвергентной и параллельной эволюции на уровне аминокислотной последовательности» . Мол Биол. Эвол . 14 (5): 527–36. doi : 10.1093/oxfordjournals.molbev.a025789 . PMID 9159930 .
- ^ Докинс, Ричард (1986). Слепые часы . WW Norton. С. 100–106 . ISBN 978-0-393-31570-7 .
- ^ Додсон, Г.; Wlodawer, A. (сентябрь 1998 г.). «Каталитические триады и их родственники». Тенденции в биохимических науках . 23 (9): 347–52. doi : 10.1016/s0968-0004 (98) 01254-7 . PMID 9787641 .
- ^ Экичи, OD; Paetzel, M.; Dalbey, Re (декабрь 2008 г.). «Нетрадиционные сериновые протеазы: вариации на каталитической конфигурации Ser/Hi/ASP» . Белковая наука . 17 (12): 2023–37. doi : 10.1110/ps.035436.108 . PMC 2590910 . PMID 18824507 .
- ^ Сафави-Хемами, Хелена; Гаджьюак, Джоанна; Карант, Сантош; Робинсон, Самуил Д.; Ueberheide, Beatrix; Дуглас, Адам Д.; Шлегель, Амнон; Империал, Юлита С.; Уоткинс, Марен; Bandyopadhyay, Pradip K.; Янделл, Марк; Ли, Цин; Перселл, Энтони В.; Нортон, Рэймонд с.; Эллгаард, Ларс; Оливера, Балмиро М. (10 февраля 2015 г.). «Специализированный инсулин используется для химической войны с помощью конусных улиток с рыбами» . Труды Национальной академии наук . 112 (6): 1743–1748. Bibcode : 2015pnas..112.1743s . doi : 10.1073/pnas.1423857112 . PMC 4330763 . PMID 25605914 .
- ^ Мартин, JP; Фридович, I (1981). «Свидетельство о естественном переносе генов от пони-рыбы до его биолюминесцентного бактериального симбионта фотобактер Leiognathi -тесная связь между бактериокукраином и медным супероксид-дисмутазой рыб телеостров» . Дж. Биол. Химический 256 (12): 6080–6089. doi : 10.1016/s0021-9258 (19) 69131-3 . PMID 6787049 .
- ^ Родригес, Вендерсон Фелипе Коста; Лиссабон, Эйртон Брено П.; Лима, Джони Сор; Ricanchenevsky, Felipe Klein; Del-Bem, Luiz-Eduardo (10 января 2023 г.). «Поглощение железа железа с помощью IRT1 / Zip развивалось по крайней мере дважды в зеленых растениях» . Новый фитолог . 237 (6): 1951–1961. Doi : 10.1111/nph.18661 . PMID 36626937 .
- ^ Jump up to: а беременный Жен, Инг; Aardema, Matthew L.; Медина, Эдгар М.; Шумер, Молли; Андольфотто, Питер (28 сентября 2012 г.). «Параллельная молекулярная эволюция в сообществе травоядных» . Наука . 337 (6102): 1634–1637. Bibcode : 2012sci ... 337.1634Z . doi : 10.1126/science.12266630 . ISSN 0036-8075 . PMC 3770729 . PMID 23019645 .
- ^ Dobler, S., Dalla, S., Wagschal, V. & Agrawal, AA (2012). Конвергентная эволюция в масштабах общего пользования в адаптации насекомых к токсичным кардалидам заменами в Na, K-Atpase. Труды Национальной академии наук, 109 (32), 13040–13045. https://doi.org/10.1073/pnas.1202111109
- ^ Jump up to: а беременный в Ян, л; Ravikantharai, n; Мариньо-Пресес; Дешмукх; Wu, m; Rosentein, а; Kunte, K; Песня H; Andolfatto, P. (2019). «Предсказуемость в эволюции нечувствительности ортоптерана картенолидов» . Серия б 374 2018246 177 ) : ( . PMC 6560278 . PMID 31154978 .
- ^ Лю, Чжэнь; Ци, Фэй-Янь; Чжоу, Синь; Рен, Хай-Цин; Ши, Пэн (2014). «Параллельные сайты предполагают функциональную конвергенцию слухового гена престина среди эхолокативных млекопитающих» . Молекулярная биология и эволюция . 31 (9): 2415–2424. doi : 10.1093/molbev/msu194 . ISSN 1537-1719 . PMID 24951728 .
- ^ Фут, Эндрю Д.; Лю, Юэ; Томас, Грегг WC; Вина, Томаш; Альфёльди, Джессика; Дэн, Джиксин; Дуган, Шеннон; Элк, Корнелис Э. Ван; Хантер, Маргарет Э. (март 2015 г.). «Конвергентная эволюция геномов морских млекопитающих» . Природа генетика . 47 (3): 272–275. doi : 10.1038/ng.3198 . PMC 4644735 . PMID 25621460 .
- ^ Ху, Ибо; Wu, Qi; Ма, Шуай; MA, Tianxiao; Шан, Лей; Ван, Сяо; Ни, Юнганг; Ning, Zemin; Ян, Ли (январь 2017 г.). «Сравнительная геномика выявляет конвергентную эволюцию между бамбуковым гигантом и красными пандами» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (5): 1081–1086. Bibcode : 2017pnas..114.1081h . doi : 10.1073/pnas.1613870114 . PMC 5293045 . PMID 28096377 .
- ^ Фейгин, Чарльз Ю.; Ньютон, Аксель Х.; Происхождение, Лилия; Шмитц, Юрген; Хипсли, Христос А.; Митчелл, Кирен Дж.; Гоув, Грэм; Ламас, Бастиен; Soubrier, Жюльен (январь 2018 г.). «Геном тасманского тигра дает представление о эволюции и демографии вымершего марсупального хищника » Природа экология и эволюция 2 (1): 182–1 Doi : 10.1038/s41559-017-0417- Y 29230027PMID
- ^ Партха, Рагхавендран; Чаухан, Бхареш К; Феррейра, Зелия; Робинсон, Джозеф Д; Латроп, Кира; Нишал, Кен К.; Чикина, Мария; Кларк, Натан Л. (октябрь 2017 г.). «Подземные млекопитающие демонстрируют конвергентную регрессию в генах глаз и энхансерах, а также адаптацию к туннелированию» . элиф . 6 doi : 10.7554/elife.25884 . PMC 5643096 . PMID 29035697 .
- ^ Сактон, туберкулез; Грейсон, P; Клутье, а; Ху, Z; Лю, JS; Уилер, NE; Gardner, pp; Кларк, JA; Бейкер, AJ; Зажим, м; Эдвардс, SV (5 апреля 2019 г.). «Конвергентная регуляторная эволюция и потеря полета у палеогниных птиц» . Наука . 364 (6435): 74–78. Bibcode : 2019sci ... 364 ... 74 с . doi : 10.1126/science.aat7244 . PMID 30948549 . S2CID 96435050 .
- ^ "Как развиваются аналогии?" Полем Калифорнийский университет Беркли. Архивировано с оригинала 2 апреля 2017 года . Получено 26 января 2017 года .
- ^ Селден, Пол; Nodds, Джон (2012). Эволюция ископаемых экосистем (2 -е изд.). CRC Press. п. 133. ISBN 978-1-84076-623-3 Полем Архивировано из оригинала 15 февраля 2017 года . Получено 26 января 2017 года .
- ^ Хелм, RR (18 ноября 2015 г.). «Познакомьтесь с Phyllirepe: морским слизняком, который выглядит и плавает, как рыба» . Deep Sea News . Архивировано из оригинала 26 июля 2019 года . Получено 26 июля 2019 года .
- ^ Ballance, Лиза (2016). «Морская среда как селективная сила для вторичных морских форм» (PDF) . UCSD. Архивировано (PDF) из оригинала 2 февраля 2017 года . Получено 19 сентября 2019 года .
- ^ Ленто, GM; Хиксон, Re; Камеры, GK; Пенни Д. (1995). «Использование спектрального анализа для проверки гипотез о происхождении пехотинцев» . Молекулярная биология и эволюция . 12 (1): 28–52. doi : 10.1093/oxfordjournals.molbev.a040189 . PMID 7877495 .
- ^ Верделин Л. (1986). «Сравнение формы черепа в сумчатых и плацентарных плотоядных животных». Австралийский журнал зоологии . 34 (2): 109–117. doi : 10.1071/Zo9860109 .
- ^ Лю, Ян; Коттон, Джеймс А.; Шен, Бин; Хан, Xiuqun; Росситер, Стивен Дж.; Чжан, Шуи (1 января 2010 г.). «Конвергентная эволюция последовательности между эхолокацией летучих мышей и дельфинами» . Текущая биология . 20 (2): R53 - R54. Bibcode : 2010cbio ... 20..r53l . doi : 10.1016/j.cub.2009.11.058 . PMID 20129036 . S2CID 16117978 .
- ^ Lavoué, Sébastien; Мия, Масаки; Arnegard, Matthew E.; Салливан, Джон П.; Хопкинс, Карл Д.; Нишида, Муцуми (14 мая 2012 г.). «Сопоставимые возрасты для независимого происхождения электрогенеза у африканских и южноамериканских слабого рыб» . Plos один . 7 (5): E36287. BIBCODE : 2012PLOSO ... 736287L . doi : 10.1371/journal.pone.0036287 . PMC 3351409 . PMID 22606250 .
- ^ Робертс, MBV (1986). Биология: функциональный подход . Нельсон Торнс. п. 274. ISBN 978-0-17-448019-8 Полем Архивировано с оригинала 12 сентября 2016 года.
- ^ Козмик, Z; Рузикова, J; Джонасова, К; Matsumoto, y.; Vopalensky, P.; Козмикова, я.; Strnad, H.; Kawamura, S.; Piatigorsky, J.; PACES, v.; Vlcek, C. (1 июля 2008 г.). «С обложки: сборка глаз Cnidarian Camera-Type из компонентов, похожих на позвоночные» . Труды Национальной академии наук . 105 (26): 8989–8993. Bibcode : 2008pnas..105.8989k . doi : 10.1073/pnas.0800388105 . PMC 2449352 . PMID 18577593 .
- ^ «Растение и эволюция животных» . Университет Вайкато. Архивировано с оригинала 18 марта 2017 года . Получено 10 января 2017 года .
- ^ Бен-Хамо, Мириам; Муньоз-Гарсия, Агусти; Ларрейн, Палома; Пять, ягода; Корин, Карми; Уильямс, Джозеф Б. (июнь 2016 г.). «Кожный липидный состав крыла и хвостовых мембран летучих мышей: случай сходящейся эволюции с птицами» . Прокурор R. Soc. Беременный 283 (1833): 20160636. DOI : 10.1098/rspb.2016.0636 . PMC 4936036 . PMID 27335420 .
- ^ Александр, Дэвид Э. (2015). На крыле: насекомые, птерозавры, птицы, летучие мыши и эволюция полета животных . Издательство Оксфордского университета. п. 28. ISBN 978-0-19-999679-7 Полем Архивировано из оригинала 14 февраля 2017 года . Получено 21 января 2017 года .
- ^ «Аналогия: белки и сахарные планеры» . Калифорнийский университет Беркли. Архивировано с оригинала 27 января 2017 года . Получено 10 января 2017 года .
- ^ Эррера, Карлос М. (1992). «Схема активности и термическая биология дневного ястреба ( Macroglossum Stellatarum ) в средиземноморских летних условиях». Экологическая энтомология . 17 (1): 52–56. Bibcode : 1992coen..17 ... 52h . doi : 10.1111/j.1365-2311.1992.tb01038.x . HDL : 10261/44693 . S2CID 85320151 .
- ^ Кренн, Харальд В.; Завод, Джон Д.; Szucsich, Nikolaus U. (2005). «Ротовые части насекомых с цветочным звеном». Членистоногие структура и развитие . 34 (1): 1–40. doi : 10.1016/j.asd.2004.10.002 .
- ^ Боддер, Джулия как; Лискониг, Нора Р.; Кренн, Харальд В. (2011). «Чрезвычайно длинная неотропическая бабочка Eurybia Lycisca (Riodinidae): морфология хоботок и обработка цветов» . Членистоногие структура и развитие . 40 (2): 122–7. BIBCODE : 2011Artsd..40..122B . doi : 10.1016/j.asd.2010.11.002 . PMC 3062012 . PMID 21115131 .
- ^ Wilheelmi, Andreas P.; Krenn, Harrad W. (2012). "Coleoptera" Зоморфология 131 (4): 325–37. doi : 10.1007/ s00435-0162-0162-3 9194699S2CID
- ^ "Когда большой палец?" Полем Понимание эволюции . Архивировано с оригинала 16 октября 2015 года . Получено 14 августа 2015 года .
- ^ Jump up to: а беременный в дюймовый и Эдвардс, М.; и др. (2010). «Ассоциация полиморфизма OCA2 HIS615ARG с содержанием меланина в популяциях Восточной Азии: дальнейшие доказательства сходящейся эволюции пигментации кожи» . PLOS Genetics . 6 (3): E1000867. doi : 10.1371/journal.pgen.1000867 . PMC 2832666 . PMID 20221248 .
- ^ Мейер, WK; и др. (2013). «Конвергентная эволюция пигментации синей радужной оболочки у приматов выбрала различные молекулярные пути» . Американский журнал физической антропологии . 151 (3): 398–407. doi : 10.1002/ajpa.22280 . PMC 3746105 . PMID 23640739 .
- ^ Jump up to: а беременный Поппенвимер, Тайлер; Мейроуз, Итай; Демалах, Нив (декабрь 2023 г.). «Пересмотр глобальной биогеографии годовых и многолетних растений» . Природа . 624 (7990): 109–114. Arxiv : 2304.13101 . Bibcode : 2023natur.624..109p . doi : 10.1038/s41586-023-06644-x . ISSN 1476-4687 . PMC 10830411 . PMID 37938778 . S2CID 260332117 .
- ^ Фридман, Джаннис (2 ноября 2020 г.). «Эволюция ежегодной и многолетней истории жизни растений: экологические корреляты и генетические механизмы» . Ежегодный обзор экологии, эволюции и систематики . 51 (1): 461–481. doi : 10.1146/annurev-ecolsys-1110218-024638 . ISSN 1543-592X . S2CID 225237602 .
- ^ Hjertaas, Ane C.; Престон, Джилл С.; Кайнулайнен, Кент; Хамфрис, Элис М.; Fjellheim, Siri (2023). «Конвергентная эволюция годового синдрома жизни от многолетних предков» . Границы в науке о растениях . 13 doi : 10.3389/fpls.2022.1048656 . ISSN 1664-462x . PMC 9846227 . PMID 36684797 .
- ^ Бойко, Джеймс Д.; Хаген, Эрик Р.; Beaulieu, Джереми М.; Vasconcelos, Thais (ноябрь 2023 г.). «Эволюционные реакции стратегий истории жизни на климатическую изменчивость в цветущих растениях» . Новый фитолог . 240 (4): 1587–1600. doi : 10.1111/nph.18971 . ISSN 0028-646X . PMID 37194450 .
- ^ Уильямс, BP; Джонстон, IG; Covshoff, S.; Хибберд, JM (сентябрь 2013 г.). «Фенотипический ландшафтный вывод выявляет множественные эволюционные пути к фотосинтезу C4» . элиф . 2 : E00961. doi : 10.7554/elife.00961 . PMC 3786385 . PMID 24082995 .
- ^ Осборн, CP; Beerling, DJ (2006). «Зеленая революция природы: замечательный эволюционный рост растений C 4 » . Философские транзакции Королевского общества B: биологические науки . 361 (1465): 173–194. doi : 10.1098/rstb.2005.1737 . PMC 1626541 . PMID 16553316 .
- ^ Мудрец, Роуэн; Рассел Монсон (1999). "16" . C4 Биология растений . Elsevier. С. 551–580. ISBN 978-0-12-614440-6 .
- ^ Чжу, XG; Long, sp; Орт, доктор (2008). «Какова максимальная эффективность с каким фотосинтезом может преобразовать солнечную энергию в биомассу?» Полем Текущее мнение о биотехнологии . 19 (2): 153–159. doi : 10.1016/j.copbio.2008.02.004 . PMID 18374559 . Архивировано с оригинала 1 апреля 2019 года . Получено 29 декабря 2018 года .
- ^ Мудрец, Роуэн; Рассел Монсон (1999). "7" . C4 Биология растений . Elsevier. С. 228–229. ISBN 978-0-12-614440-6 .
- ^ Kadereit, G.; Borsch, T.; Weising, K.; Freitag, H (2003). «Филогения Amaranthaceae и Chenopodiaceae и эволюция C 4 фотосинтеза». Международный журнал растительных наук . 164 (6): 959–86. doi : 10.1086/378649 . S2CID 83564261 .
- ^ Ирландия, Хилари, с.; и др. (2013). «Apple sepallata1/2 -подобные гены контролируют развитие плодов плоти и созревание» . Заводский журнал . 73 (6): 1044–1056. doi : 10.1111/tpj.12094 . PMID 23236986 .
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Heuvelink, EP (2005). Помидоры . Каби п. 72. ISBN 978-1-84593-149-0 Полем Архивировано с оригинала 1 апреля 2019 года . Получено 17 декабря 2016 года .
- ^ Lorts, C.; Briggeman, T.; Sang, T. (2008). «Эволюция типов фруктов и рассеивания семян: филогенетический и экологический снимок» (PDF) . Журнал систематики и эволюции . 46 (3): 396–404. doi : 10.3724/sp.j.1002.2008.08039 (неактивный 12 сентября 2024 г.). Архивировано из оригинала (PDF) 18 июля 2013 года.
{{cite journal}}: CS1 Maint: doi неактивен по состоянию на сентябрь 2024 года ( ссылка ) - ^ Lengyel, S.; Гоув, объявление; Латимер, Ам; Маджер, JD; Dunn, RR (2010). «Конвергентная эволюция рассеивания семян муравьями, а также филогения и биогеография в цветущих растениях: глобальное обследование». Перспективы в экологии растений, эволюции и систематике . 12 (1): 43–55. Bibcode : 2010ppees..12 ... 43L . doi : 10.1016/j.ppees.2009.08.001 .
- ^ Фукусима, К; Клык, х; и др. (2017). «Геном растения кувшина Cephalotus выявляет генетические изменения, связанные с плотоядным» . Природа экология и эволюция . 1 (3): 0059. Bibcode : 2017natee ... 1 ... 59f . doi : 10.1038/s41559-016-0059 . PMID 28812732 .
- ^ Jump up to: а беременный в Стейтон, С. Тристан (2015). «Определение, распознавание и интерпретация конвергентной эволюции и две новые меры для количественной оценки и оценки значимости сходимости». Эволюция 69 (8): 2140–2153. doi : 10.1111/evo.12729 . PMID 26177938 . S2CID 3161530 .
- ^ Стейтон, С. Тристан (2008). «Удивительно ли конвергенция? Исследование частоты конвергенции в моделируемых наборах данных». Журнал теоретической биологии . 252 (1): 1–14. Bibcode : 2008JTHBI.252 .... 1S . doi : 10.1016/j.jtbi.2008.01.008 . PMID 18321532 .
- ^ Мушик, Мориц; Indermaur, Adrian; Зальцбургер, Уолтер (2012). «Конвергентная эволюция в адаптивном излучении цихлидных рыб» . Текущая биология . 22 (24): 2362–2368. Bibcode : 2012cbio ... 22.2362m . doi : 10.1016/j.cub.2012.10.048 . PMID 23159601 .
- ^ Арбакл, Кевин; Беннетт, Шерил М.; Скорость, Майкл П. (июль 2014 г.). «Простая мера силы конвергентной эволюции» . Методы экологии и эволюции . 5 (7): 685–693. Bibcode : 2014mecev ... 5..685a . doi : 10.1111/2041-210x.12195 .
- ^ Ингрэм, Трэвис; Малер, Д. Люк (1 мая 2013 г.). «Поверхность: обнаружение конвергентной эволюции из сравнительных данных путем подгонки моделей Ornstein-Uhlenbeck с поэтапным информационным критерием Akaike» . Методы экологии и эволюции . 4 (5): 416–425. Bibcode : 2013mecev ... 4..416i . doi : 10.1111/2041-210x.12034 . S2CID 86382470 .
Дальнейшее чтение
[ редактировать ]- Лосос, Джонатан Б. (2017). Невероятные судьбы: судьба, случайность и будущее эволюции . Риверхед книги. ISBN 978-0399184925 .
Внешние ссылки
[ редактировать ] СМИ, связанные с конвергентной эволюцией в Wikimedia Commons
СМИ, связанные с конвергентной эволюцией в Wikimedia Commons
| Базы данных управления авторитетом : национальный |
|---|