Drosophila melanogaster
| Drosophila melanogaster | |
|---|---|

| |
| Фруктовая муха питается бананом | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Членистоногие |
| Сорт: | Инсекта |
| Заказ: | Diptera |
| Семья: | Drosophilidae |
| Род: | Drosophila |
| Семья: | Второкурсник |
| Видовая группа : | Меланогастер |
| Видовой подгруппа : | Меланогастер |
| Видовой комплекс : | Меланогастер |
| Разновидность: | Д. Меланогастер
|
| Биномиальное название | |
| Drosophila melanogaster | |
| Синонимы [ 2 ] | |
|
Список | |
Drosophila melanogaster вид мухи ( насекомое порядка - это диптера) в семействе Drosophilidae . Этот вид часто называют фруктовой мухой или меньшей фруктовой мухой , или менее обычно « муха уксуса », « Помаче муха», [ А ] [ 5 ] или " банановая муха". [ 6 ] В дикой природе D. melanogaster привлекают гниющие фрукты и ферментирующие напитки и часто встречаются в садах, кухнях и пабах.
Начиная с предложения Чарльза У. Вудворта 1901 года об использовании этого вида в качестве модельного организма , [ 7 ] [ 8 ] D. Melanogaster по -прежнему широко используется для биологических исследований в области генетики , физиологии , микробного патогенеза и эволюции истории жизни . шесть нобелевских призов было присуждено По состоянию на 2017 год дрозофилистам за их работу с использованием насекомых. [ 9 ] [ 10 ]
Drosophila melanogaster обычно используется в исследованиях благодаря быстрому жизненному циклу, относительно простой генетике с четырьмя парами хромосом и большим количеством потомства на поколение. [ 11 ] Первоначально это был африканский вид, причем все неафриканские линии имели общее происхождение. [ 12 ] Его географический диапазон включает в себя все континенты, включая острова. [ 13 ] D. Melanogaster - это обычный вредитель в домах, ресторанах и в других местах, где подается еда. [ 14 ]
Мухи, принадлежащие к семейству Tephritidae, также называют «фруктовыми мухами». Это может вызвать путаницу, особенно в Средиземноморье, Австралии и Южной Африке , где середина фруктовой мухи Ceratitis Capitata является экономическим вредителем.
Этимология
[ редактировать ]Термин « drosophila », означающий «любящий рос», представляет собой современную научную адаптация латинской адаптации от греческих слов Δρόσος , drósos , « роса » и φιλία , Филия , «Любовник». Термин « melanogaster », означающий «черный-белон», происходит от древнегреческого μέλας , mélas , «black» и γᾰστήρ , gastḗr , "Belly".
Внешний вид
[ редактировать ]В отличие от людей , на секс и внешний вид фруктовых мух не влияют гормоны . [ 15 ] Внешний вид и пол фруктовых мух определяются только генетической информацией. [ 15 ]
Женские фруктовые мухи значительно больше, чем мужские фруктовые мухи, причем женщины имеют тело, которые на 30% больше, чем взрослый мужчина. [ 16 ] [ 17 ]
Фруктовые мухи дикого типа являются желто-коричневыми, с кирпичными красными глазами и поперечными черными кольцами через живот. Черные части живота являются вдохновением для названия вида ( Melanogaster = "Black Bellied" ). Кирпичный красный цвет глаз мухи дикого типа обусловлен двумя пигментами: [ 18 ] Ксантоматин, который является коричневым и получен из триптофана , и дрозоптеринов, которые являются красными и получены из гуанозин -трифосфата . [ 18 ] Они демонстрируют сексуальный диморфизм ; Женщины имеют длину около 2,5 мм (0,10 дюйма); Мужчины немного меньше. Кроме того, у мужчин есть группа колючих волосков (затяжки), окружающие воспроизводимые части, используемые для прикрепления к самке во время спаривания. Обширные изображения встречаются в Flybase . [ 19 ]

Drosophila melanogaster можно отличить от родственных видов по следующей комбинации признаков: гена ~ 1/10 диаметром глаз на величайшей вертикальной высоте; Крыло гиалиновое и с Costal Index 2.4; мужской протсар с одним рядом ~ 12 щетинок, образующих сексуальную расческу; Мужская эпандриальная задняя доля маленькая и почти треугольная; Женский брюшной тергит 6 с темной полосой, бегущей к его вентральному краю; Женский овискапт маленький, бледный, без дорсодистальной депрессии и с 12-13, похожим на колышки, внешней овисензиллы. [ 20 ] [ 21 ]
Drosophila Melanogaster Mlies может почувствовать воздушные течения с волосами на спине. Их глаза чувствительны к небольшим различиям в интенсивности света и инстинктивно улетят, когда обнаруживается тень или другое движение. [ 22 ]
Жизненный цикл и размножение
[ редактировать ]
При оптимальных условиях роста при 25 ° C (77 ° F) продолжительность жизни D. melanogaster составляет около 50 дней от яиц до смерти. [ 23 ] Период развития D. melanogaster варьируется в зависимости от температуры, как и во многих эктотермических видах. Самое короткое время развития (яйцо со взрослой), 7 дней, достигается при 28 ° C (82 ° F). [ 24 ] [ 25 ] Время развития увеличивается при более высоких температурах (11 дней при 30 ° C или 86 ° F) из -за теплового стресса. В идеальных условиях время развития при 25 ° C (77 ° F) составляет 8,5 дня, [ 24 ] [ 25 ] [ 26 ] При 18 ° C (64 ° F) это занимает 19 дней [ 24 ] [ 25 ] и при 12 ° C (54 ° F) это занимает более 50 дней. [ 24 ] [ 25 ] В переполненных условиях время развития увеличивается, [ 27 ] в то время как новые мухи меньше. [ 27 ] [ 28 ] Самки откладывают около 400 яиц (эмбрионы), примерно пять за раз, в гниющие фрукты или другой подходящий материал, такой как разлагающие грибы и потоки сока . Drosophila melanogaster - это голометаболизное насекомое, поэтому оно подвергается полной метаморфозе. Их жизненный цикл разбит на 4 этапа: эмбрион, личинка, куколка, взрослый. [ 29 ] Яйца, которые имеют длину около 0,5 мм, люк через 12–15 часов (при 25 ° C или 77 ° F). [ 24 ] [ 25 ] Полученные личинки растут около 4 дней (при 25 ° С), внося дважды (личинки второго и третьего возраста), примерно через 24 и 48 ч после вылупления. [ 24 ] [ 25 ] В течение этого времени они питаются микроорганизмом , которые разлагают фрукты, а также сахаром самого фрукта. Мать ставит фекалии на яичные мешки, чтобы установить ту же микробную композицию в кишечниках личинок, которые работали положительно для себя. [ 30 ] Затем личинки инкапсулируют в куколку и подвергаются 4-дневной метаморфозе (при 25 ° C), после чего взрослые затмевают (Emerge). [ 24 ] [ 25 ]
Drosophila Melanogaster, широко известный как Fruit Fly, был значительным модельным организмом в исследованиях эмбриональных развития. Многие из его генов, которые регулируют эмбриональное развитие и их механизмы действия, имели решающее значение для понимания фундаментальных принципов регуляции эмбрионального развития во многих многоклеточных организмах, включая людей. Вот некоторые важные гены, регулирующие эмбриональное развитие в Drosophila melanogaster и их способы действия:
Материнские гены: эти гены кодируются в женской фруктовой мух и присутствуют на ранних стадиях развития эмбрионов. Они определяют основные особенности эмбриона и раннее развитие. Например, ген, называемый Bicoid, регулирует образование переднего конца эмбриона, и его отсутствие приводит к эмбриону, в котором отсутствует голова.
Зиготические гены: эти гены активируются на более поздних стадиях развития эмбрионов, когда эмбрион фруктовой мухи начинает производить свои собственные генетические продукты. Например, ген горбуна регулирует образование сегментов в эмбрионе.
Гомеотические гены: это семейство генов регулирует сегментацию и осевое паттерн в разработке. Они действуют как регуляторные факторы, которые определяют судьбу клеток в эмбриональном развитии. Например, ген, называемый антеннапедией, регулирует образование передних конечностей в эмбрионе.
Морфогены: это молекулы, которые образуют градиенты в эмбриональном развитии и регулируют судьбу клеток в зависимости от их положения в градиенте. Например, морфоген ежа регулирует дифференцировку сегментов и идентичности сегмента у эмбриона фруктовой мухи.
Эти гены и их способы действия образуют сложную регуляторную сеть, которая направляет эмбриональное развитие Drosophila melanogaster. Они влияют на дифференцировку клеток, образование сегмента и осевое паттерн в эмбрионе, что в конечном итоге приводит к развитию полностью образованной фруктовой мухи для взрослых.
Мужчины выполняют последовательность из пяти поведенческих закономерностей для суда. Во -первых, мужчины ориентируются, играя песню ухаживания по горизонтали, расширяя и вибрируя свои крылья. Вскоре после этого мужчина позиционирует себя в задней части живота женщины в низкой позе, чтобы постучать и лизать женские гениталии. Наконец, мужчина скручивает его живот и пытается совокуптировать. Женщины могут отвергать мужчин, уходя, ударяя и вытягивая свой яйцеклетчик. [ 31 ] Копутация длится около 15–20 минут, [ 32 ] в течение которых мужчины переносят несколько сотен, очень длинные (1,76 мм) сперматозоиды в семинальной жидкости в самку. [ 33 ] Самки хранят сперму в трубчатом сосуде и в двух грибных сперматежах ; Сперматозоид из нескольких спасителей конкурирует за оплодотворение. Считается, что последнее мужское приоритет существует; Последний мужчина спарился с женскими жирами, около 80% ее потомства. Было обнаружено, что эта приоритет происходит как посредством смещения, так и из -за недостатков. [ 34 ] Смещение связано с обработкой сперматозоидов женской мухой, когда проводится множественные составы и наиболее значимы в течение первых 1–2 дня после совокупления. Смещение из семенного сосуда является более значительным, чем смещение сперматеков. [ 34 ] Спотация первого мужского сперматозоида вторым мужским сперматозоидом становится значительным через 2–7 дней после совокупления. Считается, что оригинальная жидкость второго мужчины отвечает за этот механизм неспособность (без удаления первой мужской спермы), который вступает в силу до появления оплодотворения . [ 34 ] Считается, что задержка в эффективности механизма переноса является защитным механизмом, который предотвращает мучую муху из -за вывода своей собственной спермы, если он повторяется с той же самой женской мухой. Сенсорные нейроны в матке женщины D. melanogaster реагируют на мужской белок, половой пептид , который находится в сперме. [ 35 ] Этот белок заставляет самку неохотно совести примерно через 10 дней после осеменения . Сигнальный путь, ведущий к этому изменению поведения, был определен. Сигнал отправляется в область мозга, которая является гомологом гипоталамуса и гипоталамуса, а затем контролирует сексуальное поведение и желание. [ 35 ] Гонадотропные гормоны у дрозофилы поддерживают гомеостаз и регулируют репродуктивное производство посредством циклической взаимосвязи, мало чем отличаются от эстрального цикла млекопитающих . [ 36 ] Половый пептид нарушает этот гомеостаз и резко меняет эндокринное состояние женщины, подстрекавая синтез ювенильного гормона в корпусе Аллатум. [ 37 ]
D. Melanogaster часто используется для исследований расширения жизни , например, для идентификации генов, предполагаемых для увеличения продолжительности жизни при мутировании . [ 38 ] D. Melanogaster также используется в исследованиях старения . Синдром Вернера является условием у людей, характеризующихся ускоренным старением. Это вызвано мутациями в гене , которая кодирует белок с важной ролью в восстановлении повреждения ДНК. Мутации в D. melanogaster гомологе WRN также вызывают повышенные физиологические признаки старения, такие как более короткая продолжительность жизни, более высокая заболеваемость опухоли, дегенерация мышц, снижение способности скалолазания, измененное поведение и снижение локомоторной активности. [ 39 ]
Мейоз
[ редактировать ]Мейотическая рекомбинация у D. melanogaster, по-видимому, используется для восстановления повреждения в ДНК зародышевой линии, что указывают результатам того, что мейотическая рекомбинация индуцируется ДНК, наносящими ультрафиолетовые средства ультрафиолетовым светом [ 40 ] и митомицин c . [ 41 ]
Женщины
[ редактировать ]
Женщины становятся восприимчивыми к ухаживанию за мужчинами примерно через 8–12 часов после появления. [ 42 ] специфические нейроны Было обнаружено, что у женщин влияют на поведение совокупления и выбор партнера. Одна такая группа в брюшном нервном шнуре позволяет самой мух приостановить движения ее тела для совокупления. [ 35 ] Активация этих нейронов побуждает самку прекратить движение и ориентироваться на мужчину, чтобы обеспечить монтаж. Если группа инактивирована, самка остается в движении и не совокучивается. Различные химические сигналы, такие как мужские феромоны , часто способны активировать группу. [ 35 ]
Кроме того, женщины демонстрируют копирование партнера . Когда девственным женщинам показывают другим женщинам, совокупляющимся с определенным типом мужчины, они склонны к совокупности с этим типом мужчин впоследствии, чем наивные женщины (которые не наблюдали совокупление других). Такое поведение чувствительно к условиям окружающей среды, а женщины меньше содержится в плохих погодных условиях. [ 43 ]
Мужчины
[ редактировать ]В этом разделе нужны дополнительные цитаты для проверки . ( Октябрь 2015 ) |
D. Melanogaster Males демонстрирует сильную кривую репродуктивного обучения. То есть, с сексуальным опытом, эти мухи, как правило, изменяют свое будущее поведение спаривания несколькими способами. Эти изменения включают в себя повышенную селективность для ухаживания только внутривидовой, а также сокращение времени ухаживания .
сексуально наивные мужчины D. melanogaster Известно, что тратят значительное время, ухаживая за межпотреблением, например, с D. simulans . Наивный Д. Меланогастер также попытается утомлять женщин, которые еще не являются сексуально зрелыми, и других мужчин. D. Melanogaster мужчин мало что не предпочитают женщин D. melanogaster над женщинами других видов или даже других мужских мух. Однако после того, как D. Simulans или другие мухи, неспособные к совокуплениям, отклонили достижения мужчин, мужчины D. melanogaster с гораздо меньшей вероятностью будут тратить время на неспецифически ухаживать в будущем. Эта очевидная научная модификация поведения, по -видимому, является эволюционно значимой, поскольку она позволяет мужчинам избегать инвестирования энергии в бесполезные сексуальные встречи. [ 44 ]
Кроме того, мужчины с предыдущим сексуальным опытом изменяют свой танец ухаживания при попытке спариваться с новыми женщинами - опытные мужчины тратят меньше времени на ухаживание, поэтому имеют более низкие задержки спаривания, что означает, что они способны размножаться быстрее. Это снижение латентности спаривания приводит к повышению эффективности спаривания для опытных мужчин над наивными мужчинами. [ 45 ] Эта модификация также, по -видимому, имеет очевидные эволюционные преимущества, поскольку повышение эффективности спаривания чрезвычайно важна в глазах естественного отбора .
Многоженство
[ редактировать ]Мужские и женщины D. Melanogaster выступают за многогодно (имея несколько сексуальных партнеров одновременно). [ 46 ] Как у мужчин, так и у женщин многоженство приводит к снижению вечерней активности по сравнению с девственными мухами, в большей степени у мужчин, чем у женщин. [ 46 ] Вечерняя деятельность состоит из тех, в которых мухи участвуют, кроме спаривания и поиска партнеров, таких как поиск пищи. [ 47 ] Репродуктивный успех мужчин и женщин варьируется, потому что самка должна спариваться только один раз, чтобы достичь максимальной фертильности. [ 47 ] Сокращение с несколькими партнерами не дает преимущества перед спариванием с одним партнером, поэтому женщины не проявляют разницы в вечерней активности между полигамными и моногамными людьми. [ 47 ] Для мужчин, однако, спаривание с несколькими партнерами увеличивает их репродуктивный успех, увеличивая генетическое разнообразие их потомства. [ 47 ] Это преимущество генетического разнообразия является эволюционным преимуществом, потому что это увеличивает вероятность того, что у некоторых потомков будут быть признаки, которые увеличивают их пригодность в окружающей среде.
Разница в вечерней активности между полигамными и моногамными мужскими мухами может быть объяснена ухаживанием. Для полигамных мух их репродуктивный успех увеличивается, имея потомство с несколькими партнерами, и поэтому они тратят больше времени и энергии на ухаживание с несколькими женщинами. [ 47 ] С другой стороны, моногамные мухи только судят одну женщину и тратят меньше энергии, делая это. [ 47 ] Несмотря на то, что это требует большей энергии для мух мужского пола для суда нескольких женщин, общие репродуктивные выгоды, которые он дает, сохраняет многоженство в качестве предпочтительного сексуального выбора. [ 47 ]
Механизм, который влияет на поведение ухаживания у дрозофилы, контролируется нейронами осциллятора DN1S и LND. [ 48 ] Было обнаружено, что колебание нейронов DN1 осуществляется социосексуальными взаимодействиями и связано с уменьшением вечерней активности, связанной с спариванием. [ 48 ]
Модельный организм в генетике
[ редактировать ]D. Melanogaster остается одним из наиболее изученных организмов в биологических исследованиях, особенно в области генетики и биологии развития. Это также используется в исследованиях экологического мутагенеза.
История использования в генетическом анализе
[ редактировать ]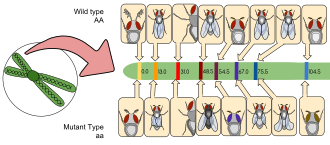
D. Melanogaster был одним из первых организмов, используемых для генетического анализа , и сегодня он является одним из наиболее широко используемых и генетически наиболее известных из всех эукариотических организмов. Все организмы используют общие генетические системы; Следовательно, понимание таких процессов, как транскрипция и репликация в плодовых мухах, помогает понять эти процессы у других эукариот, включая людей . [ 49 ]
Томас Хант Морган начал использовать фруктовые мухи в экспериментальных исследованиях наследственности в Колумбийском университете в 1910 году в лаборатории, известной как комната для мух. Комната мухи была тесно связана с восемью столами, каждая из которых занята студентами и их экспериментами. Они начали эксперименты, используя бутылки с молоком, чтобы поднять фруктовые мухи и портативные линзы для наблюдения за их признаками. Позже линзы были заменены микроскопами, которые усиливали их наблюдения. Морган и его ученики в конечном итоге выяснили многие основные принципы наследственности, в том числе наследство, связанное с полом, эпистаз , множественные аллели и картирование генов . [ 49 ]
D. Melanogaster исторически использовался в лабораториях для изучения генетики и моделей наследования. Тем не менее, D. melanogaster также имеет значение в исследованиях экологического мутагенеза, позволяя исследователям изучать эффекты специфических экологических мутагенов. [ 50 ]
Причины использования в лабораториях
[ редактировать ]
Есть много причин, по которым фруктовая муха является популярным выбором в качестве модельного организма:
- Его забота и культура требуют небольшого оборудования, пространства и затрат, даже при использовании больших культур.
- Он может быть безопасно и легко анестезирован (обычно с эфиром , газом углекислого газа , путем охлаждения или с такими продуктами, как Flynap ).
- Его морфология легко идентифицировать после анестезии.
- Он имеет короткое время поколения (около 10 дней при комнатной температуре), поэтому в течение нескольких недель можно изучить несколько поколений.
- Он имеет высокую плодовитость (женщины откладывают до 100 яиц в день и, возможно, 2000 за всю жизнь). [ 11 ]
- Мужчины и женщины легко различаются, и девственные женщины могут быть легко идентифицированы по их светло-полупрозрачному животу, способствуя генетическому пересечению.
- Зрелая личинка имеет гигантские хромосомы в слюнных железах, называемых политенными хромосомами , «Puffs», которые указывают на области транскрипции, следовательно, активность генов. Недостаточная повторная репликация рДНК происходит только 20% от ДНК по сравнению с мозгом. Сравните с 47%, меньше рДНК в яичниках саркофаги Барбата .
- Он имеет только четыре пары хромосом - три аутосома и одна пара сексуальных хромосом .
- Мужчины не показывают мейотическую рекомбинацию , облегчая генетические исследования.
- Рецессивные летальные « балансирующие хромосомы », несущие видимые генетические маркеры, могут использоваться для поддержания запасов летальных аллелей в гетерозиготном состоянии без рекомбинации из -за множественных инверсий в балансировке.
- Развитие этого организма - от оплодотворенного яйца до взрослого взрослого - хорошо понятно.
- Методы генетической трансформации были доступны с 1987 года. Один подход к вставке иностранных генов в геном Drosophila включает P -элементы. Элементы Pransposable P, также известные как транспозоны , представляют собой сегменты бактериальной ДНК, которые переносятся в геном мухи. Трансгенные мухи уже способствовали многим научным достижениям, например, моделировали такие заболевания человека, как у Паркинсона , неоплазии , ожирения и диабета .
- Его полный геном был секвенирован и впервые опубликован в 2000 году. [ 51 ]
- Сексуальная мозаика может быть легко произведена, предоставляя дополнительный инструмент для изучения развития и поведения этих мух. [ 52 ]
Генетические маркеры
[ редактировать ]
Генетические маркеры обычно используются в исследованиях Drosophila , например, в хромосомах балансировщиков или вставках P-элемента, и большинство фенотипов легко идентифицируются либо невооруженным глазом, либо под микроскопом. В списке нескольких общих маркеров ниже символ аллеля сопровождается названием затронутого гена и описание его фенотипа. (Примечание: рецессивные аллели находятся в нижнем случае, в то время как доминирующие аллели капитализируются.)
- Сай 1 : Кудрявый; Крылья изгибаются от тела, полет может быть несколько нарушен
- и 1 : Ebony; Черное тело и крылья (гетерозиготы также заметно темнее, чем дикий тип)
- Сб 1 : Щеба; Компания более короче и толще дикого типа
- В 1 : Белый ; Глазам не хватает пигментации , и кажутся белыми
- BW: Браун; Цвет глаз определяется различными пигментами вместе взятых.
- и 1 : Желтый; выглядят желтым Пигментация тела и крылья
Классические генетические мутации
[ редактировать ]Гены Drosophila традиционно названы в честь фенотипа , который они вызывают при мутировании. Например, отсутствие определенного гена у дрозофилы приведет к мутантному эмбриону, у которого не развивается сердце. Ученые, таким образом, назвали этот ген Tinman , названный в честь Оз одноименного персонажа . [ 54 ] Аналогично изменения в ген -фритюре вызывают потерю дорсальных кутикулярных волосков у Drosophila Sechellia . личинок [ 55 ] Эта система номенклатуры приводит к более широкому диапазону названий генов, чем в других организмах.
- Б: Черная - Черная мутация была обнаружена в 1910 году Томасом Хантом Морганом . [ 56 ] Черная мутация приводит к более темному телу, крыльям, венам и сегментам ноги фруктовой мухи. [ 57 ] Это происходит из-за неспособности мухи создать бета-аланин , бета-аминокислоту. [ 56 ] Фенотипическая экспрессия этой мутации варьируется в зависимости от генотипа индивидуума; Например, независимо от того, является ли образец гомозиготным или гетерозиготным, приводит к более темному или менее темному внешнему виду. [ 57 ] Эта генетическая мутация является X-связанной рецессивностью . [ 58 ]
- BW: коричневый - коричневая мутация глаз является результатом неспособности продуцировать или синтезировать пигменты птеридина (красный) из -за точечной мутации на хромосоме II. [ 59 ]
- М: Миниатюра - одна из первых записей миниатюрной мутации крыльев была также сделана Томасом Хантом Морганом в 1911 году. Он описал крылья, имеющие такую же форму, как фенотип дикого типа. Тем не менее, их миниатюрное обозначение относится к длине их крыльев, которые не растягиваются за их телом и, таким образом, особенно короче длины дикого типа. Он также отметил, что его наследство связано с полом мухи и может быть в сочетании с наследством других признаков, определенных по полу, таких как белые глаза. [ 60 ] Крылья могут также продемонстрировать другие характеристики, отклоняющиеся от крыла дикого типа, таких как более тупой и более облачный цвет. [ 61 ] Миниатюрные крылья в 1,5 раза короче, чем дикий тип, но, как полагают, имеют одинаковое количество клеток. Это связано с отсутствием полного сглаживания этими клетками, из -за чего общая структура крыла кажется короче по сравнению. Путь расширения крыла регулируется путем рецептора сигнала, где нейромормоновый бурсикон взаимодействует с его комплементарным рецептором, связанным с G-белком; Этот рецептор побуждает одну из субъединиц G-белка сигнализировать о дальнейшей активности ферментов и приводит к развитию в крыле, таких как апоптоз и рост. [ 62 ]
- SE: Sepia - Цвет глаз мутанта Sepia - это сепия , красновато -коричневый цвет. У мух дикого типа оммохромы (коричневые) и дрозоптерины (красный) придают глазам типичный красный цвет. [ 63 ] [ 64 ] Drosopterins производится по пути, который включает в себя пиримидодиазепин -синтазу , [ 65 ] который кодируется на хромосоме 3L. Ген имеет преждевременный стоп -кодон в сепии, так что мухи не могут продуцировать пиримидодиазепин -синтазу и, следовательно, без красного пигмента, так что глаза остаются сепией. [ 63 ] сепии Аллель рецессив , и, следовательно, у потомков от мух сепии и гомозиготных мух дикого типа, есть красные глаза. сепии Фенотип не зависит от пола мухи. [ 66 ]
- V: Вермилион - мутанты из вермилиона не могут производить коричневые оммохромы, оставляя красные дросоптерины, так что глаза цветные глаз (сияющий красный) по сравнению с D. melanogaster . Мутация вермилиона связана с полом и рецессивным. Ген, который является дефектом, лежит на Х -хромосоме. [ 67 ] Коричневые оммохромы синтезируются из кинуренана, который сделан из триптофана. Файлы из вермилиона не могут преобразовать триптофан в кинуренин и, таким образом, также не могут сделать оммохромы. [ 67 ] Мутанты из вермилиона живут дольше, чем мухи дикого типа. Эта более длительная продолжительность жизни может быть связана с уменьшенным количеством триптофана, преобразованного в кинуренин в мухах с вермилионом. [ 68 ]

- VG: Редиментальная - спонтанная мутация, обнаруженная в 1919 году Томасом Морганом и Кэлвином Бриджесом. Редиментарные крылья - это те, которые не полностью разработаны, и которые потеряли функцию. С момента открытия рудиментарного гена у Drosophila melanogaster было много открытий рудиментарного гена у других позвоночных и их функций в позвоночных. [ 69 ] Редиментальный ген считается одним из наиболее важных генов для формирования крыла, но когда он становится чрезмерно выраженной проблемой эктопических крыльев. [ 70 ] Редиментальный ген действует для регулирования экспрессии видальных дисков крыла в эмбрионе и действует с другими генами для регулирования развития крыльев. Мутированный рудиментарный аллель удаляет важную последовательность ДНК, необходимой для правильного развития крыльев. [ 71 ]
- W: Белый - Drosophila Melanogaster Дикий тип обычно выражает кирпичный цвет красных глаз. Мутация белого глаза в плодовых мухах вызвана отсутствием двух пигментов, связанных с красными и коричневыми цветами глаз; Перидины (красный) и Оммохромы (коричневые). [ 64 ] В январе 1910 года Томас Хант Морган впервые обнаружил белый ген и обозначил его как w . Открытие мутации белого глаза Морганом привело к началу генетических экспериментов и анализа Drosophila melanogaster. Охота в конечном итоге обнаружила, что ген последовал за аналогичной схемой наследования, связанной с мейотической сегрегацией Х -хромосомы. Он обнаружил, что ген был расположен на Х -хромосоме с этой информацией. Это привело к открытию генов, связанных с полом, а также к открытию других мутаций в Drosophila melanogaster. [ 72 ] Мутация белого глаза приводит к нескольким недостаткам у мух, таких как снижение способности скалолазания, сокращение продолжительности жизни и снижение сопротивления напряжению по сравнению с мухами дикого типа. [ 73 ] Drosophila melanogaster имеет ряд спаривающихся поведений, которые позволяют им совокуплять в данной среде и, следовательно, способствовать их физической подготовке. После того, как Морган открыл мутацию с белыми глазами, связанную с полом, исследование, проведенное Стертевантом (1915) [ 74 ] Было обнаружено, что чем больше плотность пигментации глаз, тем больше успех в спаривании для мужчин доктора Ософилы Меланогастер. [ 74 ]
- Y: Желтый - Желтый ген - это генетическая мутация , известная как DMEL \ Y в широко используемой основе данных, называемой Flybase . Эта мутация может быть легко идентифицирована с помощью атипичного желтого пигмента, наблюдаемого в кутикуле взрослых мух и кусочков рта личинки. [ 75 ] Мутация Y содержит следующие фенотипические классы: мутанты, которые показывают полную потерю пигментации от кутикулы (Y-тип) и других мутантов, которые показывают мозаичный пигментный рисунок с некоторыми областями кутикулы (дикий тип, тип Y2). [ 76 ] Роль желтого гена разнообразна и отвечает за изменения в поведении , пола-специфическое репродуктивное созревание и эпигенетическое перепрограммирование. [ 77 ] Ген Y является идеальным геном для изучения, так как ясно, когда у организма есть этот ген, что облегчает понимание передачи ДНК к потомству. [ 77 ]

Геном
[ редактировать ] D. Chromosomes D. Melanogaster для масштабирования со ссылками на мегаза-пар, ориентированных, как в Национальном центре биотехнологической информации данных , расстояния сантиморгана являются приблизительными и оцениваются из местоположения выбранных карковых локусов. | |
| NCBI Genome Id | 47 |
|---|---|
| Плуидия | диплоид |
| Количество хромосом | 8 |
| Год завершения | 2015 |
Геном D. melanogaster данных (секвенированный в 2000 году и куратор в Flybase базе [ 51 ] ) содержит четыре пары хромосом - пара x/y и три аутосома, помеченные 2, 3 и 4. Четвертая хромосома относительно очень мала и, следовательно, часто игнорируется, помимо его важного без глаз гена . D. melanogaster -секвенированный геном 139,5 миллиона пар оснований был аннотирован [ 78 ] и содержит около 15 682 генов в соответствии с высвобождением 73 ансамбля. Более 60% генома, по-видимому, является функциональной небелковой ДНК. [ 79 ] участвует в контроле экспрессии генов. Определение пола у дрозофилы возникает с помощью x: соотношение x хромосом к аутосомах, а не из -за присутствия y -хромосомы, как при определении пола человека. Хотя Y-хромосома полностью гетерохроматическая , она содержит как минимум 16 генов, многие из которых, как полагают, имеют связанные с мужчинами функции. [ 80 ]
Существует три ортолога трансферрина , все из которых резко расходятся от тех, известных в хордовых моделях. [ 81 ]
Сходство с людьми
[ редактировать ]Исследование, проведенное в июне 2001 года Национального института исследований генома человека, в сравнении с фруктовой мухой и геномом человека подсчитано, что между двумя видами сохраняются около 60% генов. [ 82 ] Около 75% известных генов заболеваний человека имеют узнаваемое соответствие в геноме плодовых мух, [ 83 ] и 50% последовательностей белка муха имеют гомологи млекопитающих [ Цитация необходима ] Полем Онлайн -база данных под названием Homophila доступна для поиска гомологов генов болезни человека у мух и наоборот. [ 84 ]
Drosophila используется в качестве генетической модели для нескольких заболеваний человека, включая нейродегенеративные расстройства Паркинсона , Хантингтон , спиноцеребеллярная атаксия и болезнь Альцгеймера . [ 85 ] Муха также используется для изучения механизмов, лежащих в основе старения и окислительного стресса , иммунитета , диабета и рака , а также злоупотребления наркотиками . [ 86 ] [ 87 ] [ 88 ]
Разработка
[ редактировать ]Жизненный цикл этого насекомого имеет четыре этапа: оплодотворенное яйцо, личинка, куколка и взрослые. [ 13 ]
Эмбриогенез у дрозофилы был тщательно изучен, поскольку его небольшой размер, короткое время поколения и большой размер выводки делают его идеальным для генетических исследований. Он также уникален среди модельных организмов в том, что расщепление происходит в синцитии .
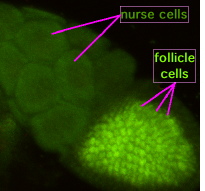
Во время оогенеза цитоплазматические мосты, называемые «кольцевыми каналами», соединяют образующий ооцит с клетками медсестры. Питательные вещества и молекулы контроля развития перемещаются из клеток медсестры в ооцит. На фигуре слева можно увидеть образующий ооцит, покрытый фолликулярными опорными клетками.
После оплодотворения ооцита ранний эмбрион (или синцитиальный эмбрион ) подвергается быстрому репликации ДНК и 13 ядерных подразделений до примерно от 5000 до 6000 ядер не накапливается в неразделенной цитоплазме эмбриона. К концу восьмого дивизии большинство ядер мигрировали на поверхность, окружая желточный мешок (оставив после себя лишь несколько ядер, которые станут ядрами желтка). После 10 -го деления полюсные клетки образуются на заднем конце эмбриона, сегрегируя зародышевую линию от синцития. Наконец, после 13 -го деления клеточные мембраны медленно инвагируют, деляя синцитик на отдельные соматические клетки. Как только этот процесс будет завершен, запускается гаструляция . [ 89 ]
Ядерное разделение в раннем эмбрионе Drosophila происходит так быстро, нет надлежащих контрольных точек, поэтому в разделении ДНК можно допустить ошибки. Чтобы обойти эту проблему, ядра, которые совершили ошибку, отделились от их центросом и попадают в центр эмбриона (желточный мешок), который не будет формировать часть мухи.
Генная сеть (транскрипционные и белковые взаимодействия), регулирующая раннее развитие эмбриона фруктовой мухи, является одной из наиболее понятных генных сетей на сегодняшний день, особенно паттерны вдоль антеропостериорских (AP) и дорсовентральных (DV) (см. В разделе морфогенез ). [ 89 ]
Эмбрион подвергается хорошо охарактеризованным морфогенетическим движениям во время гаструляции и раннего развития, включая разгибание зародышевой полосы , образование нескольких бороздок, вентральное инвагирование мезодермы , а также задней и передней инвагивации эндодермы (кишечник), а также обширная сегментация тела до, наконец вылупление от окружающей кутикулы в личинку первого возраста.
Во время развития личинок ткани, известные как имагинальные диски, растут внутри личинки. Воображаемые диски развиваются, образуя большинство структур взрослого тела, таких как голова, ноги, крылья, грудная клетка и гениталии. Клетки изображенных дисков выделяются во время эмбриогенеза и продолжают расти и делиться на стадиях личинок - в отличие от большинства других клеток личинки, которые дифференцировались для выполнения специализированных функций и растут без дальнейшего деления клеток. При метаморфозе личинка образует куколку , внутри которой личиночные ткани реабсорбируются, а изображенные ткани подвергаются обширным морфогенетическим движениям с образованием взрослых структур.
Пластичность развития
[ редактировать ]Биотические и абиотические факторы, испытываемые во время разработки, будут влиять на распределение ресурсов развития, что приводит к фенотипическим изменениям , также называемой пластичностью развития. [ 90 ] [ 91 ] Как во всех насекомых, [ 91 ] Факторы окружающей среды могут влиять на несколько аспектов развития в Drosophila melanogaster . [ 92 ] [ 93 ] Фруктовые мухи, выращенные при обработке гипоксии , снижали длину грудной клетки, в то время как гипероксия вызывает меньшие мышцы полета, что указывает на негативные эффекты развития экстремального уровня кислорода. [ 94 ] Циркадные ритмы также подвержены пластичности развития. Условия освещения во время развития влияют на повседневную активность в Drosophila melanogaster , где мухи, поднятые при постоянной темноте или свете, менее активны, чем взрослые, чем те, которые подняты в 12-часовом цикле света/темноты. [ 95 ]
Температура является одним из наиболее распространенных факторов, влияющих на развитие членистоногих . В Drosophila Melanogaster , индуцированная температурой, пластичность развития может быть полезной и/или вредной. [ 96 ] [ 97 ] Чаще всего более низкие температуры развития снижают темпы роста, что влияет на многие другие физиологические факторы. [ 98 ] Например, развитие при 25 ° C увеличивает скорость ходьбы, ширину тепловых характеристик и территориальный успех, в то время как развитие при 18 ° C увеличивает массу тела, размер крыла, все из которых привязаны к фитнесу. [ 93 ] [ 96 ] Более того, развитие при определенных низких температурах производит пропорционально большие крылья, которые улучшают полеты и репродуктивные характеристики при одинаковых низких температурах ( см. Акклимат ). [ 99 ]
Хотя определенные эффекты температуры развития, такие как размер тела, необратимы в эктотермах , другие могут быть обратимыми. [ 91 ] [ 100 ] Когда Drosophila melanogaster развивается при холодных температурах, у них будет большая толерантность к холоду, но если холодные мухи сохраняются при более теплых температурах, их устойчивость к холоду уменьшается, а теплоемкость увеличивается с течением времени. [ 100 ] [ 101 ] Поскольку насекомые, как правило, спариваются только в определенном диапазоне температур, их устойчивость к холоду/тепло является важной признаком при максимизации репродуктивной мощности. [ 102 ]
В то время как описанные выше черты, как ожидается, будут проявляться аналогично по полу, температура развития также может оказывать специфические для полов эффекты у взрослых D. melanogaster .
- На число женщин - авориоле значительно влияет температура развития у D. melanogaster. [ 103 ] На размер яиц также влияет температура развития и усугубляется, когда оба родителя развиваются при теплых температурах ( см. Материнский эффект ). [ 96 ] При стрессовых температурах эти структуры будут развиваться до меньших конечных размеров и уменьшать репродуктивный выход самки. [ 103 ] [ 96 ] Ранняя плодовитость (общее количество яиц, отложенных в первые 10 дней после экземпляра ) максимизируется при выращенном при 25 ° C (против 17 ° C и 29 ° C) независимо от температуры взрослых. [ 104 ] В широком спектре температур развития женщины, как правило, имеют большую теплостойкость, чем мужчины. [ 105 ]
- У мужчин- стрессовые температуры развития будут вызывать бесплодия у D. melanogaster мужчин ; Хотя верхняя температура может быть увеличена путем поддержания штаммов при высоких температурах ( см. Акклиматику ). [ 97 ] Мужская бесплодия может быть обратимой, если взрослые возвращаются к оптимальной температуре после развития при стрессовых температурах. [ 106 ] Мухи -мухи меньше и более успешны при защите участков пищи/яйцеклазей при выращивании при 25 ° С против 18 ° С; Таким образом, меньшие мужчины будут иметь увеличение успеха спаривания и репродуктивного производства. [ 93 ]
Определение пола
[ редактировать ]У мухи дрозофилы есть как х, так и y хромосомы, а также аутосомы . В отличие от людей, Y -хромосома не придает мужественности; Скорее, он кодирует гены, необходимые для изготовления спермы. Вместо этого пол определяется соотношением х хромосом к аутосомах. [ 107 ] Кроме того, каждая клетка «решает», быть ли мужчина или женщина независимо от остальной части организма, что приводит к случайному появлению гинандроморфов .
| X хромосомы | Аутосомы | Соотношение X: а | Секс |
|---|---|---|---|
| XXXX | Аааа | 1 | Нормальная женщина |
| XXX | Ааа | 1 | Нормальная женщина |
| Xxy | Аа | 1 | Нормальная женщина |
| Xxyy | Аа | 1 | Нормальная женщина |
| XX | Аа | 1 | Нормальная женщина |
| XY | Аа | 0.50 | Нормальный мужчина |
| Х | Аа | 0.50 | Нормальный мужчина (стерильный) |
| XXX | Аа | 1.50 | Метафемале |
| XXXX | Ааа | 1.33 | Метафемале |
| XX | Ааа | 0.66 | Интерсекс |
| Х | Ааа | 0.33 | Метамале |
Три основных гена участвуют в определении пола дрозофилы . Это сексуальные , без сестринских и невозмутимые . Deadpan -это аутосомный ген, который ингибирует пола-митальный , в то время как без сестры переносится на Х-хромосоме и ингибирует действие невозмута . Клетка AAX имеет в два раза больше невозмутимой , чем без сестрин , так что половые митальные будут препятствовать, создавая мужчину. Тем не менее, ячейка AAXX будет производить достаточно без сестры , чтобы ингибировать действие DeadPan , что позволит по полу-митаю транскрибированному гену , чтобы создать женщину.
Позже, контроль Deadpan и Sisterless исчезает, и то, что становится важным, является формой гена -пола-митального . Вторичный промотор вызывает транскрипцию как у мужчин, так и у женщин. Анализ кДНК показал , что различные формы экспрессируются у мужчин и женщин. полов-митальный Было показано, что влияет на сплайсинг собственной мРНК . третий экзон У мужчин включен , который кодирует стоп -кодон , вызывая изготовленную усеченную форму. В женской версии присутствие пола-летальных причин этого экзона пропустить; Остальные семь аминокислот продуцируются в виде полной пептидной цепи, что снова дает разницу между мужчинами и женщинами. [ 108 ]
Наличие или отсутствие функциональных полов-летальных белков теперь влияют на транскрипцию другого белка, известного как DoubleSex. В отсутствие пола-митала у DoubleSex будет удален четвертый экзон и будет переведен на экзон 6 (DSX-M [ALE]), в то время как в его присутствии четвертый экзон, который кодирует стоп-кодон, будет создавать усеченную версию белка (DSX-F [EMALE]). DSX-F вызывает транскрипцию белков желтка 1 и 2 в соматических клетках, которые будут перекачены в ооцит при его производстве.
Иммунитет
[ редактировать ]Иммунная система D. melanogaster можно разделить на два ответа: гуморальный и клеточный опосредованный. Первый является системным ответом, опосредованным в значительной степени через пути платы и IMD , которые являются параллельными системами для обнаружения микробов. Другие пути, включая пути реакции на стресс, Jak-Stat и P38 , передача сигналов питания через Foxo и передачу сигналов гибели клеток JNK, участвуют в ключевых физиологических реакциях на инфекцию. D. Melanogaster имеет орган, называемый « жировым телом », который аналогичен печени человека. Жирный организм является основным секреторным органом и производит ключевые иммунные молекулы при инфекции, такие как сериновые протеазы и антимикробные пептиды (AMP). Усильника секретируются в гемолимфе и связывают инфекционные бактерии и грибы, убивая их, образуя поры в клеточных стенках или ингибируя внутриклеточные процессы. Клеточный иммунный ответ вместо этого относится к прямой активности клеток крови (гемоцитов) у дрозофилы , которые аналогичны моноцитам/макрофагам млекопитающих. Гемоциты также обладают значительной ролью в опосредовании гуморальных иммунных реакций, таких как реакция меланизации . [ 109 ]
Иммунный ответ на инфекцию может включать до 2423 генов или 13,7% генома. Хотя транскрипционная реакция мухи на микробную проблему очень специфична для отдельных патогенов, Drosophila дифференциально экспрессирует основную группу из 252 генов при инфекции большинством бактерий. Эта основная группа генов связана с генной онтологической категориями, такими как антимикробная реакция, стресс-реакция, секреция, нейроноподобная, размножение и метаболизм. [ 110 ] [ 111 ] Drosophila также обладает несколькими иммунными механизмами для формирования микробиоты и предотвращения чрезмерных иммунных реакций при обнаружении микробных стимулов. Например, секретируемые PGRP с амидазой активностью и разлагают иммуностимулирующие PGN DAP PGN, чтобы блокировать активацию IMD. [ 112 ]
В отличие от млекопитающих, у дрозофилы есть врожденный иммунитет , но не имеет адаптивного иммунного ответа. Тем не менее, основные элементы этого врожденного иммунного ответа сохраняются между людьми и плодовыми мухами. В результате фруктовая муха предлагает полезную модель врожденного иммунитета для распутывания генетических взаимодействий передачи сигналов и эффекторной функции, поскольку мухи не должны бороться с вмешательством адаптивных иммунных механизмов, которые могут путать результаты. Различные генетические инструменты, протоколы и анализы делают Drosophila классической моделью для изучения врожденной иммунной системы , [ 113 ] который даже включал иммунные исследования на международной космической станции. [ 114 ]
Сигнализация Jak-Stat
[ редактировать ]Многочисленные элементы сигнального пути Drosophila Jak-Stat несут прямую гомологию для генов пути Jak-Stat человека . Передача сигналов JAK-STAT индуцируется на различных организмальных напряжениях, таких как тепловой стресс, обезвоживание или инфекция. Индукция JAK-STAT приводит к производству ряда белков реакции на стресс, включая тиоэфирные белки (TEPS), [ 115 ] Туруэнт, [ 116 ] и предполагаемый антимикробный пептид листерицин. [ 117 ] Механизмы, посредством которых многие из этих белков все еще находятся под следствием. Например, TEP, по-видимому, способствуют фагоцитозу грамположительных бактерий и индукции платного пути. Как следствие, мухи, лишенные TEP, подвержены инфекции с помощью проблем с платным путем. [ 115 ]
Клеточный ответ на инфекцию
[ редактировать ]Циркулирующие гемоциты являются ключевыми регуляторами инфекции. Это было продемонстрировано как с помощью генетических инструментов для генерации мух, отсутствующих гемоциты, либо посредством инъекционных микрогарных шариков или липидных капель, которые насыщают способность гемоцитов к фагоцитозу вторичной инфекции. [ 118 ] [ 119 ] Мухи, обработанные как это, не смогут фагоцитозных бактерий при инфекции и соответственно восприимчивы к инфекции. [ 120 ] Эти гемоциты происходят из двух волн гематопоэза , которые возникают в раннем эмбрионе и одну, возникающие во время развития от личинки к взрослому. [ 121 ] Однако гемоциты Drosophila не обновляются в течение срока службы взрослых, и поэтому у мухи есть конечное количество гемоцитов, которые уменьшаются в течение его срока службы. [ 122 ] Гемоциты также участвуют в регуляции событий клеточного цикла и апоптоза аберрантной ткани (например, раковых клеток) путем продуцирования EIGER, сигнальной молекулы фактора некроза опухоли , которая способствует передаче сигналов JNK , в конечном итоге гибель клеток и апоптоза. [ 123 ]
Поведенческая генетика и нейробиология
[ редактировать ]В 1971 году Рон Конопка и Сеймур Бензер опубликовали «Часовые мутанты дрозофилы Меланогастер », статью, описывающую первые мутации , которые повлияли на поведение животного. Мухи дикого типа показывают ритм активности с частотой около дня (24 часа). Они обнаружили мутантов с более быстрыми и более медленными ритмами, а также разбитыми ритмами - свети, которые движутся и отдыхают случайными всплесками. Работа в течение следующих 30 лет показала, что эти мутации (и другие, подобные им), влияют на группу генов и их продукты, которые образуют биохимические или биологические часы . Эти часы обнаруживаются в широком диапазоне клеток мухи, но в клетках, несущих часы, которые контролируют активность, представляют собой несколько десятков нейронов в центральном мозге мухи.
С тех пор Бензер и другие использовали поведенческие экраны для выделения генов, участвующих в зрении, обонянии, прослушивании, обучении/памяти, ухаживании, боли и других процессах, таких как долговечность.
После новаторской работы Альфреда Генри Стуртеванта [ 124 ] и другие, Бензер и коллеги [ 52 ] использовал сексуальную мозаику для разработки новой техники картирования судьбы . Этот метод позволил назначить конкретную характеристику определенному анатомическому расположению. Например, этот метод показал, что поведение ухаживания мужского пола контролируется мозгом. [ 52 ] Картирование судьбы мозаики также дало первое указание на существование феромонов у этого вида. [ 125 ] женщины Мужчины различают конспецифических мужчин и женщин и прямых постоянных ухаживаний, преимущественно к женщинам благодаря специфическому сексуальному феромону, который в основном производится тергитами .
Первые мутанты обучения и памяти ( Dunce , Rutabaga и т. Д.) Были выделены Уильямом «Чип» Куинн, в то время как в лаборатории Бенцера и, в конечном итоге, было показано, что он кодирует компоненты внутриклеточного сигнального пути, включающего циклический AMP , протеинкиназу A и транскрипцию фактор, известный как CREB. Было показано, что эти молекулы также участвуют в синаптической пластичности при аплизии и млекопитающих. [ 126 ]
Нобелевская премия по физиологии или медицине на 2017 год была присуждена Джеффри С. Холл, Майклу Росбаш, Майклу В. Янгу за их работы с использованием плодовых мух в понимании «молекулярных механизмов, контролирующих циркадный ритм ». [ 127 ]
Мужские мухи поют женщинам во время ухаживания, используя свои крылья для создания звука, и некоторые генетики сексуального поведения были охарактеризованы. В частности, бесплодный ген имеет несколько разных форм сплайсинга, а мужские мухи, экспрессирующие женские формы сплайсинга, имеют женское поведение и наоборот. Каналы TRP Nompc , Nanchung и Unactive выражаются в чувствительных к звуке нейронам органов Джонстона и участвуют в трансдукции звука. [ 128 ] [ 129 ] Мутирующий ген гендерблок , также известный как CG6070, изменяет сексуальное поведение дрозофилы , поворачивая бисексуалы . [ 130 ]
[Файл: фруктовые макрогианты B.Jpg | Thumb | 281x281px]]]
Мухи используют модифицированную версию фильтров Bloom для обнаружения новизны запахов, с дополнительными функциями, включая сходство нового запаха с тем, что у ранее опытных примеров, и время, прошедшее с предыдущего опыта того же запаха. [ 131 ]
Агрессия
[ редактировать ]Как и в большинстве насекомых, агрессивное поведение между мужчинами, обычно возникающих в присутствии ухаживания за женщиной и при борьбе за ресурсы. Такое поведение часто включает в себя поднятие крыльев и ног в сторону противника и атаку всего тела. [ 132 ] Таким образом, это часто вызывает повреждение крыла, что уменьшает их пригодность, удаляя их способность летать и спариваться. [ 133 ]
Акустическая связь
[ редактировать ]Для того чтобы агрессия произошла, мужские мухи производят звуки, чтобы сообщить о своих намерениях. Исследование 2017 года показало, что песни, способствующие агрессии, содержат импульсы, возникающие с более длинными интервалами. [ 134 ] Секвенирование РНК от мутантов FLY, демонстрирующих чрезмерно агрессивное поведение, обнаружив более 50 связанных с слуховыми генами (важно для переходных рецепторных потенциалов , CA 2+ Сигнализация и потенциалы механорецептора ) будут активированы в нейронах AB, расположенных в органе Джонстона . [ 134 ] Кроме того, уровни агрессии были снижены, когда эти гены были выбиты через РНК -интерференцию . [ 134 ] Это означает основную роль слуха как сенсорного модальности в общении агрессии.
Передача сигналов феромона
[ редактировать ]Помимо слуха, еще одной сенсорной модальностью, которая регулирует агрессию, является передача сигналов феромонов , которая работает через обонятельную систему или вкусовую систему в зависимости от феромона. [ 135 ] Примером является CVA , анти-афродизиак-феромон, используемый мужчинами для маркировки женщин после совокупления и сдерживания других мужчин от спаривания. [ 136 ] Этот специфичный для мужчин феромон вызывает увеличение агрессии мужского мужского мужчины, когда обнаруживается вкусовой системой другого мужчины . [ 135 ] Однако, вставив мутацию, которая делает мухи безответственными к CVA, не было видно агрессивного поведения. [ 137 ] Это показывает, как существует множество модальностей для продвижения агрессии у мух.
Конкуренция за еду
[ редактировать ]В частности, при борьбе за пищу агрессию происходит в зависимости от количества доступных продуктов питания и не зависит от каких -либо социальных взаимодействий между мужчинами. [ 138 ] В частности, было обнаружено, что сахароза стимулирует нейроны пожимающих рецепторов, что было необходимо для стимулирования агрессии. [ 138 ] Однако, как только количество пищи становится больше определенного количества, конкуренция между мужчинами снижается. [ 138 ] Возможно, это связано с чрезмерным воображением продовольственных ресурсов. В более широком масштабе пища была обнаружена, определяющая границы территории, поскольку мухи были более агрессивными при физическом периметре пищи.
Эффект лишения сна
[ редактировать ]Однако, как и большинство поведения, требующих возбуждения и бодрствования, было обнаружено, что агрессия нарушена через лишение сна . В частности, это происходит благодаря нарушению передачи сигналов осьупамина и дофамина , которые являются важными путями для регуляции возбуждения у насекомых. [ 139 ] [ 140 ] Из-за снижения агрессии было обнаружено, что мужские мухи, лишенные сна, находятся в невыгодном положении при спаривании по сравнению с нормальными мухами. [ 140 ] Однако, когда агонисты октопамина вводили на этих лишенных сна мухах, показано, что уровни агрессии увеличивались, а сексуальная подготовка впоследствии восстанавливалась. [ 140 ] Следовательно, этот вывод подразумевает важность сна в агрессии между мужчинами.
Зрение
[ редактировать ]
Составной глаз фруктовой мухи содержит 760 единичных глаз или Ommatidia , и является одним из самых продвинутых среди насекомых. Каждый омматидий содержит восемь фоторецепторных клеток (R1-8), поддерживает клетки, пигментные клетки и роговицу. Мухи дикого типа имеют красноватые пигментные клетки, которые служат для поглощения избыточного синего света, чтобы муха не была ослеплена окружающим светом. Гены цвета глаз регулируют клеточный везикулярный транспорт. Ферменты, необходимые для синтеза пигмента, затем транспортируются в пигментную гранулу клетки, которая содержит молекулы предшественника пигмента. [ 64 ]


Каждая фоторецепторная клетка состоит из двух основных секций, тела клеток и рабдоды . Клеточное тело содержит ядро , в то время как рабдомера длиной 100 мкм состоит из зубной щетки стопок мембраны, называемых микроворками . Каждый Microvillus имеет длину 1–2 мкм и около 60 нм . диаметром [ 141 ] Мембрана Rhabdomere наполнена около 100 миллионами молекул опсина , визуальным белком, который поглощает свет. Другие визуальные белки также плотно упакованы в микровортинг, оставляя мало места для цитоплазмы .
Опсины и спектральная чувствительность
[ редактировать ]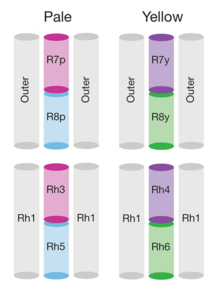
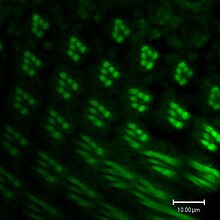
Геном дрозофилы кодирует семь опсинов, [ 143 ] Пять из них выражены в Оматидии глаза. Фоторецепторные клетки R1-R6 экспрессируют опсин RH1, [ 144 ] который поглощает максимально синий свет (около 480 нм), [ 145 ] [ 146 ] [ 147 ] Однако клетки R1-R6 охватывают более широкий спектр спектра, чем опсин, позволил бы из-за сенсибилизирующего пигмента [ 148 ] [ 149 ] Это добавляет два максимума чувствительности в УФ-диапазоне (355 и 370 нм). [ 147 ] Клетки R7 поставляются в двух типах с желтыми и бледными рабдорами (R7Y и R7P). [ 150 ] [ 151 ] Бледные клетки R7P экспрессируют опсин RH3, [ 152 ] [ 153 ] что максимально поглощает ультрафиолетовое освещение (345 нм). [ 154 ] Клетки R7P строго соединены с клетками R8P, которые экспрессируют RH5, [ 153 ] которые максимально поглощают фиолетовый свет (437 нм). [ 147 ] Другое, желтые клетки R7Y экспрессируют сине-поглощающий скрининговой пигмент [ 150 ] и Opsin RH4, [ 155 ] что максимально поглощает ультрафиолетовое освещение (375 нм). [ 154 ] Клетки R7Y строго сочетаются с клетками R8y, которые экспрессируют RH6, [ 156 ] которые максимально поглощают ультрафиолетовое освещение (508 нм). [ 147 ] В подмножестве Omatidia клетки R7 и R8 экспрессируют Opsin RH3. [ 153 ]
Однако эти максимумы поглощения опсинов, где измерены у мух с белыми глазами без скрининга пигментов (RH3-RH6), [ 154 ] [ 147 ] или из изолированного опсина напрямую (RH1). [ 145 ] Эти пигменты уменьшают свет, который достигает опсинов в зависимости от длины волны. Таким образом, у полностью пигментированных мух эффективные максимумы поглощения опсинов отличаются и, таким образом, также чувствительность их фоторецепторных клеток. С пигментом с помощью скрининга Opsin RH3 - короткая волна, смещенная с 345 нм [ B ] до 330 нм и RH4 от 375 нм до 355 нм. Присутствует ли скрининг пигмент, не имеет практической разницы для Opsin RH5 (435 нм и 437 нм), в то время как Opsin R6 - длинная волна, сдвинутая на 92 нм с 508 нм до 600 нм. [ 142 ]
Кроме того, из опсинов глаз у Drosophila есть еще два опсина: окелли экспрессируют Opsin RH2, [ 157 ] [ 158 ] который максимально поглощает фиолетовый свет (~ 420 нм). [ 158 ] И Opsin RH7, который максимально поглощает ультрафиолетовое освещение (350 нм) необычайно длинным хвостом длины волны до 500 нм. Длинный хвост исчезает, если лизин в положении 90 заменяется глутаминовой кислотой . Затем этот мутант поглощает максимально фиолетовый свет (450 нм). [ 159 ] Опсин RH7 входит в ритм Drosophila дневному к кардиостимулятора . циклу в центральных нейронах криптохромный [ 160 ]
Каждый дрозофила опсин связывает каротиноидный хромофор 11-цис-3-гидроксиретинал через лизин. [ 161 ] [ 162 ] Этот лизин сохраняется почти во всех опсинах, лишь несколько опсинов потеряли его во время эволюции . [ 163 ] Опсины без него не чувствительны к свету. [ 164 ] [ 165 ] [ 166 ] В частности, Drosophila Opsins RH1, RH4 и RH7 функционируют не только как фоторецепторы , но и как хеморецепторы для аристололовой кислоты . Эти опсины все еще имеют лизин, как и другие опсины. Однако, если он заменяется аргинином в RH1, то RH1 теряет чувствительность к свету, но все же реагирует на аристололовую кислоту. Таким образом, лизин не нужен для RH1, чтобы функционировать как хеморецептор. [ 165 ]
-
![Спектральная чувствительность Drosophila melanogaster Opsins в белых глазах. Чувствительность RH3 -R6 смоделирована с шаблонами опсина и оценками чувствительности от Salcedo et al. (1999). [147] Opsin RH1 (перекрашенный из Salcedo et al. [147]) имеет характерную форму, поскольку он связан с ультрафиолетовым пигментом.](//upload.wikimedia.org/wikipedia/commons/thumb/f/fc/Spectral_sensitivities_of_Drosophila_melanogaster_opsins_in_white_eyed_flies.png/750px-Spectral_sensitivities_of_Drosophila_melanogaster_opsins_in_white_eyed_flies.png) Спектральная чувствительность Drosophila melanogaster Opsins в белых глазах. Чувствительность RH3 -R6 смоделирована с шаблонами опсина и оценками чувствительности от Salcedo et al. (1999). [ 147 ] Opsin RH1 (перекрашенный из Salcedo et al. [ 147 ] ) имеет характерную форму, так как она связана с ультрафиолетовым пигментом.
Спектральная чувствительность Drosophila melanogaster Opsins в белых глазах. Чувствительность RH3 -R6 смоделирована с шаблонами опсина и оценками чувствительности от Salcedo et al. (1999). [ 147 ] Opsin RH1 (перекрашенный из Salcedo et al. [ 147 ] ) имеет характерную форму, так как она связана с ультрафиолетовым пигментом. -
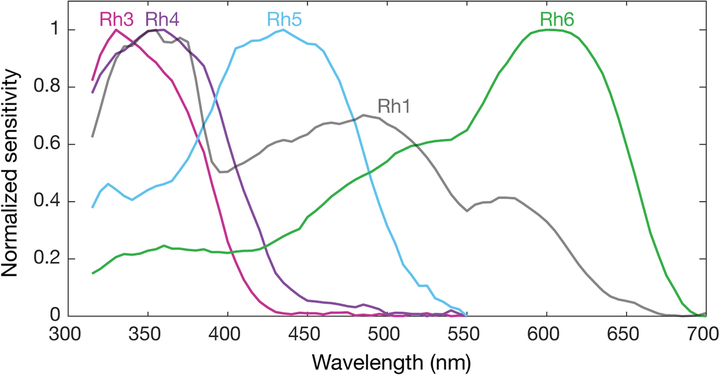 Нормализованные средние кривые спектральной чувствительности Drosophila melanogaster Opsins RH1, RH3, RH4, RH5 и RH6, измеренные в их нативных клетках фоторецептора в красных глазных мухах с помощью скрининга пигмента. Каждая спектральная кривая - это среднее из шести мух.
Нормализованные средние кривые спектральной чувствительности Drosophila melanogaster Opsins RH1, RH3, RH4, RH5 и RH6, измеренные в их нативных клетках фоторецептора в красных глазных мухах с помощью скрининга пигмента. Каждая спектральная кривая - это среднее из шести мух.
![Спектральная чувствительность Drosophila melanogaster Opsins в белых глазах. Чувствительность RH3 -R6 смоделирована с шаблонами опсина и оценками чувствительности от Salcedo et al. (1999). [147] Opsin RH1 (перекрашенный из Salcedo et al. [147]) имеет характерную форму, поскольку он связан с ультрафиолетовым пигментом.](http://upload.wikimedia.org/wikipedia/commons/thumb/f/fc/Spectral_sensitivities_of_Drosophila_melanogaster_opsins_in_white_eyed_flies.png/750px-Spectral_sensitivities_of_Drosophila_melanogaster_opsins_in_white_eyed_flies.png)

Фототрансдукция
[ редактировать ]Как и в зрении позвоночных , визуальная трансдукция у беспозвоночных происходит по пути G-белка. Однако у позвоночных G -белок является трансдуцином , в то время как G -белок у беспозвоночных представляет собой GQ (DGQ у дрозофилы ). Когда родопсин (RH) поглощает фотон света, его хромофор, 11-цис-3-гидроксиретинальный, изомерируется на All-Trans-3-гидроксиретинальный. RH претерпевает конформационное изменение в своей активной форме, метарходопсин. Метарходопсин активирует GQ, который, в свою очередь, активирует фосфолипазу Cβ (PLCβ), известный как NORPA. [ 167 ]
PLCβ гидролизует фосфатидилинозитол (4,5) -бисфосфат (PIP 2 ), фосфолипид, обнаруженный в клеточной мембране , в растворимый иннозитол трихосфат (IP 3 ) и диацилглицерин (DAG), который остается в клеточной мембране. DAG, производное от истощения DAG или PIP 2 вызывают кальциево -селективный ионный канал , известный как потенциал переходного рецептора (TRP) для открытия, а кальций и натрия в клетку. [ 168 ] IP 3 Считается, что IP 3 связывается с рецепторами в подчиненных цистернае, расширении эндоплазматической ретикулумы и вызывает высвобождение кальция, но этот процесс, по -видимому, не является важным для нормального зрения. [ 167 ]
Кальций связывается с белками, такими как кальмодулин (CAM) и специфичная для глаз протеинкиназа C (PKC), известная как INAC. Эти белки взаимодействуют с другими белками и, как было показано, необходимы для отключения светового ответа. Кроме того, белки, называемые аррестинами, связывают метарходопсин и предотвращают активацию большего количества GQ. Обмен натрия, известный как Calx, выкачивает кальций из клетки. Он использует внутренний градиент натрия для экспорта кальция при стехиометрии 3 NA + / 1 ок ++ . [ 169 ]
TRP, INAC и PLC образуют сигнальный комплекс, связывая белок каркасов, называемый INAD. INAD содержит пять связывающих доменов, называемых доменными белками PDZ , которые специфически связывают C -концевые белки -мишени. Разрушение комплекса по мутациям либо в доменах PDZ, либо в целевых белках снижает эффективность передачи сигналов. Например, нарушение взаимодействия между INAC, протеинкиназой C и INAD приводит к задержке инактивации светового ответа .
В отличие от метарходопсина позвоночных, метарходопсин беспозвоночного может быть преобразован обратно в родопсин, поглощая фотон оранжевого света (580 нм).
Приблизительно две трети мозга дрозофилы посвящены визуальной обработке. [ 170 ] Хотя пространственное разрешение их зрения значительно хуже, чем у людей, их временное разрешение примерно в 10 раз лучше.
Груминг
[ редактировать ]Известно, что Drosophila демонстрируют поведение груминга, которое выполняется предсказуемо. Drosophila постоянно начинает последовательность груминга, используя их передние ноги, чтобы очистить глаза, затем голову и антенны. Используя свои задние ноги, Drosophila продолжает ухаживать за животом, и, наконец, крылья и грудная клетка. На протяжении всей этой последовательности Drosophila периодически втирает ноги, чтобы избавиться от избыточной пыли и мусора, которые накапливаются во время процесса ухода. [ 171 ]
Было показано, что поведение груминга выполняется в иерархии подавления. Это означает, что поведение груминга, которое возникает в начале последовательности, предотвращают те, которые появляются позже в последовательности одновременно, поскольку последовательность груминга состоит из взаимно исключительного поведения. [ 172 ] [ 173 ] Эта иерархия не мешает Drosophila вернуться к поведению груминга, которые уже были доступны в последовательности груминга. [ 172 ] Считается, что порядок поведения груминга в иерархии подавления связан с приоритетом очистки определенной части тела. Например, глаза и антенны, вероятно, выполняются на ранних стадиях в последовательности груминга, чтобы предотвратить вмешательство мусора в функцию сенсорных органов D. melanogaster . [ 172 ] [ 173 ]
Ходьба
[ редактировать ]
Как и многие другие гексаподные насекомые, Drosophila обычно ходит, используя походку для штатива . [ 175 ] Это означает, что три ноги развернулись вместе, в то время как остальные три остаются неподвижными или в стойке. В частности, средняя нога движется в фазе с контралатеральными передним и задним ногами. Тем не менее, вдоль континуума существует изменчивость вокруг конфигурации штатива, что означает, что мухи не демонстрируют различные переходы между различными походками. [ 176 ] При быстрых скоростях ходьбы конфигурация ходьбы в основном штатива (3 ножки в стойке), но на более медленных скоростях ходьбы мухи с большей вероятностью имеют четыре (тетрапод) или пять ног в стойке (волна). [ 177 ] [ 178 ] Эти переходы могут помочь оптимизировать статическую стабильность . [ 179 ] Поскольку мухи настолько малы, инерционные силы незначительны по сравнению с упругими силами их мышц и суставов или вязкими силами окружающего воздуха. [ 180 ]
Полет
[ редактировать ]Мухи летают по прямым последовательностям движения, вкрапленных быстрыми поворотами, называемыми саккадами. [ 181 ] Во время этих поворотов муха способна вращать 90 ° менее чем на 50 миллисекунд. [ 181 ]
Характеристики полета Drosophila могут преобладать вязкостью воздуха , а не инерция тела мухи, но противоположный случай с инерцией, как доминирующая сила. [ 181 ] Тем не менее, последующая работа показала, что, хотя вязкое воздействие на тело насекомых во время полета может быть незначительным, аэродинамические силы на самих крыльях фактически вызывают сознание фруктовых мух. [ 182 ]
Коннектика
[ редактировать ]Drosophila - это одно из немногих животных ( C. elegans подробные нейронные цепи ( разъединение - это другое), где доступны ).
Высокий разъединок, на уровне мозговых компартментов и взаимосвязанных трактов нейронов, существует для полного мозга мухи. [ 183 ] Версия этого доступна онлайн. [ 184 ]

Подробные соединения на уровне цепи существуют для пластинки [ 185 ] [ 186 ] и медулла [ 187 ] Колонна, как в визуальной системе фруктовой мухи, так и альфа -доля грибного тела. [ 188 ]
В мае 2017 года статья, опубликованная в Biorxiv, представила стек изображений электронной микроскопии всего взрослого взрослого мозга самки при синаптическом разрешении. Объем доступен для редкого отслеживания выбранных цепей. [ 189 ] [ 190 ] С тех пор было собрано несколько наборов данных, в том числе плотный разъем половины центрального мозга Drosophila в 2020 году, [ 191 ] [ 192 ] и плотный разъединок всего взрослого нервного шнура для взрослых в 2021 году. [ 193 ] Как правило, эти наборы данных получают путем разделения ткани (например, мозга) на тонкие срезы (по порядку десяти или сотен нанометров). Затем каждый раздел отображается с использованием электронного микроскопа, и эти изображения сшивают и выровнены вместе, чтобы создать 3D -объем изображения. Методы, используемые при реконструкции и начальном анализе таких наборов данных. [ 194 ] Из -за достижений в глубоком обучении автоматизированные методы сегментации изображения сделали крупномасштабную реконструкцию, обеспечивая плотные реконструкции всех нейритов в объеме. [ 195 ] Кроме того, разрешение электронной микроскопии освещает ультраструктурные различия между нейронами, а также расположение отдельных синапсов, тем самым обеспечивая диаграмму подключения синаптической связности между всеми нейритами в данном наборе набора данных.
В 2023 году была опубликована полная карта личиночного мозга дрозофилы на уровне синапса, а также анализ его архитектуры. Личиночный мозг состоит из 3016 нейронов и 548 000 синаптических мест, [ 196 ] Принимая во внимание, что взрослый мозг имеет около 150 000 нейронов и 150 миллионов синапсов.
Заблуждения
[ редактировать ]Drosophila иногда называют вредителем из -за его тенденции жить в поселениях человека, где обнаруживается ферментирование фруктов. Мухи могут собираться в домах, ресторанах, магазинах и других местах. [ 14 ] Название и поведение этого вида мухи привели к неправильному представлению о том, что это риск биологической безопасности в Австралии и в других местах. В то время как другие виды «фруктовой мухи» представляют риск, D. Melanogaster привлекает фрукты, которые уже гниют, а не выводят фрукты гнить. [ 197 ] [ 198 ]
Смотрите также
[ редактировать ]- Испытание на животных на беспозвоночных
- Поведение питания у насекомых (измерение)
- Фруктовые мухи в космосе
- Генетически модифицированное насекомое
- Гинандроморфизм
- Jetlag Gene
- Список баз данных Drosophila
- Spaetzle (гены)
- Время летит как стрела; фрукты летают как банан
- Трансгенез
- Рвиграф - еще один широко используемый модельный организм в научных исследованиях
Примечания
[ редактировать ]- ^ «Уксусная муха» предпочтительнее нескольких недавних публикаций как более точное описание, чем «фруктовая муха». [ 3 ] [ 4 ] [ 5 ]
- ^ Sharkey et al. [ 142 ] Дайте максимум поглощения RH3 как 334 нм в их разделе результатов. Однако в разделе «Введение» и «Материал и методы» они дают его как 345 нм. Для обоих значений они цитируют Feiler et al., Которые сообщили только о 345 нм. [ 154 ] Поэтому, кажется, это ошибка, и они, вероятно, имели в виду, что там 345 нм тоже.
Ссылки
[ редактировать ]- ^ Meigen JW (1830). Систематическое описание хорошо известных европейских двойных насекомых. (Том 6) (PDF) (на немецком языке). Schulz-Wundermann. Архивировано из оригинала (PDF) 9 февраля 2012 года.
- ^ «Drosophila (Shophophora) Melanogaster Meigen, 1830» . Каталог жизни . Виды 2000 : Лейден, Нидерланды . Получено 2 мая 2024 года .
- ^ «Дрозофила | род насекомых» . Энциклопедия Британская . Получено 30 октября 2021 года .
- ^ «Уксус летит» . Penn State Extension . Получено 30 октября 2021 года .
- ^ Jump up to: а беременный Зеленый ММ (сентябрь 2002 г.). «Это действительно не фруктовая муха» . Генетика . 162 (1): 1–3. doi : 10.1093/Genetics/162.1.1 . PMC 1462251 . PMID 12242218 .
- ^ Schilthuizen, Menno (28 апреля 2015 г.). «Химический коктейль спермы может похитить мозг приятеля» . Обнаружить . Получено 11 сентября 2023 года .
Даже в эякуляции скромного бананового муха Drosophila melanogaster исследователи идентифицировали не менее 133 различных видов белков.
- ^ Биография Нобелевской премии Моргана, упоминающая CW Woodworth
- ^ Холден Б (1 января 2015 г.). Чарльз В. Вудворт: замечательная жизнь первого энтомолога UC (1 -е изд.). Брайан Холден издательство. С. 135–137. ISBN 978-0-9864105-3-6 .
- ^ «Нобелевские призы» . Хранитель . 7 октября 2017 года.
- ^ "Fruitfly-Researchgate" .
- ^ Jump up to: а беременный Sang JH, Reeve EC (23 июня 2001 г.). «Drosophila melanogaster: фруктовая муха» . Энциклопедия генетики . США: Фицрой Дирборн, Издатели, I. с. 157. ISBN 978-1-884964-34-3 Полем Получено 1 июля 2009 г.
- ^ Baudry E, Viginier B, Veuille M (август 2004 г.). «Неафриканские популяции Drosophila melanogaster имеют уникальное происхождение» . Молекулярная биология и эволюция . 21 (8): 1482–91. doi : 10.1093/molbev/msh089 . PMID 15014160 .
- ^ Jump up to: а беременный Маркоу Т.А. (июнь 2015 г.). «Секретная жизнь дрозофилы летит» . элиф . 4 doi : 10.7554/elife.06793 . PMC 4454838 . PMID 26041333 .
- ^ Jump up to: а беременный «Мухи уксуса, виды Drosophila , семейство: Drosophilidae » . Кафедра энтомологии, Колледж сельскохозяйственных наук, Университет штата Пенсильвания. 2017 . Получено 20 июля 2017 года .
- ^ Jump up to: а беременный Келли, Дарси Б.; Байер, Эмили А. (2021). «Сексуальный диморфизм: нейронные схемы переключаются в мозгу дрозофилы». Текущая биология . 31 (6). Elsevier BV: R297 - R298. doi : 10.1016/j.cub.2021.02.026 . ISSN 0960-9822 .
- ^ Палома Альварес-Рендон, Жессика; Мануэль Мурильо-Малдонадо, Хуан; Рафаэль Рисго-Эсковар, Хуан (2023). «Сигнальный путь инсулина через столетие после его открытия: половой диморфизм при передаче сигналов инсулина». Общая и сравнительная эндокринология . 330 . Elsevier BV: 114146. DOI : 10.1016/j.ygcen.2022.114146 . ISSN 0016-6480 . «... Взрослые женщины на 30 % больше мужчин; эти различия происходят в течение жизни личинок».
- ^ Коули, де; Атчли, WR (1 июня 1988 г.). «Количественная генетика Drosophila melanogaster. II. Наследие и генетические корреляции между полами для черт головы и грудной клетки» . Генетика . 119 (2). Издательство Оксфордского университета (OUP): 421–433. doi : 10.1093/Genetics/119.2.421 . ISSN 1943-2631 . PMC 1203424 .
- ^ Jump up to: а беременный Ewart GD, Howells AJ (1 января 1998 г.). «Переносчики ABC, участвующие в транспортировке предшественников пигмента глаза у Drosophila melanogaster». Транспортеры ABC: биохимические, клеточные и молекулярные аспекты . Методы в фермере. Тол. 292. Академическая пресса. С. 213–24. doi : 10.1016/s0076-6879 (98) 92017-1 . ISBN 978-0-12-182193-7 Полем PMID 9711556 .
- ^ «Flybase: база данных генов и геномов дрозофилы» . Генетическое общество Америки. 2009. Архивировано из оригинала 15 августа 2009 года . Получено 11 августа 2009 г.
- ^ Юдзуки, Чавен; Тидон, Розана (2020). «Идантификация для видов дрозофилидных видов (Ditpter, drosophilidae) экзоцич в неотропическую область и встречается в Бразилии » Revista brasileira de entomology 64 (1). Doi : 10.1590/1806-9665-Rent- 2019-1 ISSN 1806-9665 S2CID 21570766
- ^ Миллер, я; Маршалл, SA; Grimaldi, DA (2017). «Обзор видов Drosophila (Diptera: Drosophilidae) и родов Drosophilidae северо -восточной Северной Америки» . Канадский журнал идентификации членистоногих . 31 doi : 10.3752/cjai.2017.31 .
- ^ «Дрозофила Меланогастер» . Интернет -разнообразие животных. 2000. Архивировано с оригинала 30 ноября 2014 года . Получено 11 августа 2009 г.
- ^ Линфорд Н.Дж., Билгир С., Ро Дж, Плетчер С.Д. (январь 2013 г.). «Измерение продолжительности жизни у Drosophila melanogaster» . Журнал визуализированных экспериментов (71). doi : 10.3791/50068 . PMC 3582515 . PMID 23328955 .
- ^ Jump up to: а беременный в дюймовый и фон глин Ashburner M , Thompson JN (1978). «Лабораторная культура дрозофилы ». В Ashburner M, Wright TRF (ред.). Генетика и биология дрозофилы . Тол. 2A. Академическая пресса. 1–81.
- ^ Jump up to: а беременный в дюймовый и фон глин Ashburner M , Golc KG, Hawley RS (2005). Drosophila: лабораторный справочник (2 -е изд.). Cold Spring Harbor Laboratory Press. С. 162–4. ISBN 978-0-87969-706-8 .
- ^ Bloomington Drosophila Stock Center в Университете Индианы : основные методы культивирования Drosophila Archived 2006-09-01 на The Wayback Machine
- ^ Jump up to: а беременный Chiang HC, Hodson AC (1950). «Аналитическое исследование роста популяции в Drosophila melanogaster ». Экологические монографии . 20 (3): 173–206. Bibcode : 1950ecom ... 20..173c . doi : 10.2307/1948580 . JSTOR 1948580 .
- ^ Баккер К (1961). «Анализ факторов, которые определяют успех в конкуренции за пищу среди личинок Drosophila melanogaster ». Архив Néerlandaises de Zoologie . 14 (2): 200–281. doi : 10.1163/0365551661x00061 . S2CID 85129022 .
- ^ Fernández-Moreno MA, Farr CL, Kaguni LS, Garesse R (2007). «Drosophila melanogaster как модельная система для изучения митохондриальной биологии». Митохондрия . Методы в молекулярной биологии. Тол. 372. С. 33–49. doi : 10.1007/978-1-59745-365-3_3 . ISBN 978-1-58829-667-2 Полем PMC 4876951 . PMID 18314716 .
- ^ Blum JE, Fischer CN, Miles J, Handelsman J (ноябрь 2013). «Частое пополнение поддерживает полезный микробиом Drosophila melanogaster» . Мбио . 4 (6): E00860-13. doi : 10.1128/mbio.00860-13 . PMC 3892787 . PMID 24194543 .
- ^ Кук Р., Коннолли К (1973). «Ответы отказа от женского Drosophila melanogaster: их онтогенция, причинность и влияние на поведение ухаживания мужчины». Поведение . 44 (1/2): 142–166. doi : 10.1163/156853973x00364 . JSTOR 4533484 . S2CID 85393769 .
- ^ Houot B, Svetec N, Godoy-Herrera R, Ferveur JF (июль 2010 г.). «Влияние лабораторного акклиматизации на изменение связанных с размножением признаков у Drosophila melanogaster» . Журнал экспериментальной биологии . 213 (Pt 13): 2322–31. doi : 10.1242/jeb.041566 . PMID 20543131 .
- ^ Гилберт С.Ф. (2006). «9: оплодотворение у дрозофилы» . Биология развития (8 -е изд.). Sinauer Associates. ISBN 978-0-87893-250-4 Полем Архивировано из оригинала 7 февраля 2007 года.
- ^ Jump up to: а беременный в Прайс CS, Dyer KA, Coyne JA (июль 1999 г.). «Конкуренция сперматозоидов между мужчинами дрозофилы включает в себя как смещение, так и недееспособное». Природа . 400 (6743): 449–52. Bibcode : 1999natur.400..449p . doi : 10.1038/22755 . PMID 10440373 . S2CID 4393369 .
- ^ Jump up to: а беременный в дюймовый «Исследование фруктовой мухи может показать, что происходит в мозге женского пола во время ухаживания, спариваемости» . Получено 5 октября 2014 года .
- ^ Meiselman M, Lee SS, Tran RT, Dai H, Ding Y, Rivera-Perez C, et al. (Май 2017). «Дрозофила Меланогастер» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (19): E3849 - E3858. doi : 10.1073/pnas.1620760114 . PMC 5441734 . PMID 28439025 .
- ^ Мошицки П., Флейшманн I, Чаймов Н., Саудан П., Клаузер С., Кубли Э., Эпплбаум С.В. (1996). «Секс-пептид активирует биосинтез ювенильного гормона в Drosophila melanogaster corpus allatum». Архивы биохимии насекомых и физиологии . 32 (3–4): 363–74. doi : 10.1002/(SICI) 1520-6327 (1996) 32: 3/4 <363 :: AID-Arch9> 3.0.co; 2-t . PMID 8756302 .
- ^ Carnes Mu, Campbell T, Huang W, Butler DG, Carbone MA, Duncan LH, et al. (2015). «Геномная основа отложенного старения у Drosophila melanogaster» . Plos один . 10 (9): E0138569. BIBCODE : 2015PLOSO..1038569C . doi : 10.1371/journal.pone.0138569 . PMC 4574564 . PMID 26378456 .
- ^ Cassidy D, Epinyy DG, Salameh C, Zhou LT, Salomon RN, Schirmer AE и др. (Ноябрь 2019). «Свидетельство о преждевременном старении в модели синдрома Вернера Дрозофилы» . Экспериментальная геронтология . 127 : 110733. DOI : 10.1016/j.exger.2019.110733 . PMC 6935377 . PMID 31518666 .
- ^ Prudhommeau C, Proust J. УФ -облучение топольских клеток эмбрионов Drosophila melanogaster. V. Изучение мейотической рекомбинации у женщин с хромосомами различной структуры. Mutat Res. 1974 апрель; 23 (1): 63-6. PMID 4209047
- ^ Schewe MJ, Suzuki DT, Erasmus U. Генетические эффекты митомицина C у Drosophila melanogaster. II Индуцированная мейотическая рекомбинация. Mutat Res. 1971 июля; 12 (3): 269-79. doi: 10.1016/0027-5107 (71) 90015-7. PMID 5563942
- ^ Питник С. (1996). «Инвестиции в яички и стоимость изготовления длинных сперматозоидов в Drosophila ». Американский натуралист . 148 : 57–80. doi : 10.1086/285911 . S2CID 83654824 .
- ^ Dagaeff AC, Pocheville A, Nöbel S, Loyau A, Isabel G, Danchin E (2016). «Копирование Drosophila Mate коррелирует с атмосферным давлением в ситуации с скоростью обучения» . Поведение животных . 121 : 163–174. doi : 10.1016/j.anbehav.2016.08.022 .
- ^ Dukas R (2004). «Мужские фруктовые мухи учатся избегать межвидового ухаживания» . Поведенческая экология . 15 (4): 695–698. doi : 10.1093/beheco/arh068 .
- ^ Салим С., Рагглс П.Х., Эбботт В.К., Карни Г.Е. (2014). «Сексуальный опыт усиливает Drosophila melanogaster мужского поведения спаривания и успеха» . Plos один . 9 (5): E96639. Bibcode : 2014ploso ... 996639S . doi : 10.1371/journal.pone.0096639 . PMC 4013029 . PMID 24805129 .
- ^ Jump up to: а беременный фон Хаартман Л. (1951). «Последовательное многоженство». Поведение . 3 (1): 256–273. doi : 10.1163/156853951x00296 .
- ^ Jump up to: а беременный в дюймовый и фон глин Vartak VR, Varma V, Sharma VK (февраль 2015 г.). «Влияние многоженства на ритм активности/отдых мужских плодовых мух Drosophila melanogaster». Умирайте натуральный . 102 (1–2): 1252. Bibcode : 2015scina.102 .... 3V . doi : 10.1007/s00114-014-1252-5 . PMID 25604736 . S2CID 7529509 .
- ^ Jump up to: а беременный Бейтман AJ (декабрь 1948 г.). «Внутрисексуальный отбор у дрозофилы» . Наследственность . 2 (ст. 3): 349–68. doi : 10.1038/hdy.1948.21 . PMID 18103134 .
- ^ Jump up to: а беременный Пирс Б.А. (2004). Генетика: концептуальный подход (2 -е изд.). Freeman WH ISBN 978-0-7167-8881-2 .
- ^ Килби Б.Дж., Макдональд DJ, Auerbach C, Sobels FH, Vogel EW (июнь 1981 г.). «Использование Drosophila melanogaster в тестах на экологические мутагены». Мутационные исследования . 85 (3): 141–6. doi : 10.1016/0165-1161 (81) 90029-7 . PMID 6790982 .
- ^ Jump up to: а беременный Адамс MD; Celniker SE ; Холт Р.А.; Эванс КА; Gocayne JD; Верность pg; и др. (Март 2000 г.). "Последовательность геноза Drosophila melanogaster" Наука . 287 (5461): 2185–9 Бибкод : 2000SCI. Полем Citeserx 10.1.1.1.549.8639 Doi : 10.1126/ science.287.5461.2 10731132PMID
- ^ Jump up to: а беременный в Hotta Y, Benzer S (декабрь 1972 г.). «Картирование поведения у Drosophila Mosaics». Природа . 240 (5383): 527–35. Bibcode : 1972natur.240..527h . doi : 10.1038/240527a0 . PMID 4568399 . S2CID 4181921 .
- ^ Meiers S (2018). «Использование новых технологий секвенирования ДНК для изучения геномных перестроек» . archiv.ub.uni-heidelberg.de . doi : 10.11588/heidok.00024506 . Получено 28 июня 2021 года .
- ^ Azpiazu N, Frasch M (июль 1993 г.). «Tinman и Pagpipe: два гена Homeo Box, которые определяют судьбы клеток в дорсальной мезодерме дрозофилы» . Гены и развитие . 7 (7b): 1325–40. doi : 10.1101/gad.7.7b.1325 . PMID 8101173 .
- ^ Stern DL, Frankel N (декабрь 2013 г.). «Структура и эволюция цис-регуляторных областей: история о фриенбаби» . Философские транзакции Королевского общества Лондона. Серия B, биологические науки . 368 (1632): 20130028. DOI : 10.1098/rstb.2013.0028 . PMC 3826501 . PMID 24218640 .
- ^ Jump up to: а беременный Phillips AM, Smart R, Strauss R, Brembs B, Kelly Le (май 2005). «Drosophila Black Enigma: молекулярная и поведенческая характеристика аллеля мутанта Black1» (PDF) . Ген . 351 : 131–42. doi : 10.1016/j.gene.2005.03.013 . PMID 15878647 .
- ^ Jump up to: а беременный «Отчет гена Flybase: DMEL \ B» . Flybase.org . Получено 26 марта 2019 года .
- ^ Sherald AF (сентябрь 1981). «Межгенное подавление черной мутации Drosophila melanogaster». Молекулярная и общая генетика . 183 (1): 102–6. doi : 10.1007/bf00270146 . PMID 6799739 . S2CID 1210971 .
- ^ Shoup Jr (май 1966 г.). «Развитие пигментных гранул в глазах дикого типа и мутанта Drosophila melanogaster» . Журнал клеточной биологии . 29 (2): 223–49. doi : 10.1083/jcb.29.2.223 . PMC 2106902 . PMID 5961338 .
- ^ Morgan TH (март 1911 г.). «Происхождение девяти мутаций крыла у дрозофилы». Наука . 33 (848): 496–9. Bibcode : 1911sci .... 33..496m . doi : 10.1126/science.33.848.496 . JSTOR 1638587 . PMID 17774436 .
- ^ «Отчет гена Flybase: DMEL \ M» . Flybase.org . Получено 26 марта 2019 года .
- ^ Bilousov OO, Katanaev VL, Demydov SV, Kozeretska IA (март -апрель 2013). «Понижающая регуляция миниатюрного гена не воспроизводит фенотипы миниатюрной потери функции в крыле Drosophila melanogaster». Tsitologiia I Genetika . 47 (2): 77–81. PMID 23745366 .
- ^ Jump up to: а беременный Ким Дж, Су Х, Ким С., Ким К, Ан С., Йим Дж (сентябрь 2006 г.). «Идентификация и характеристики структурного гена для мутантной сепии цветового цвета дрозофилы, кодирующей PDA-синтазы, члена глютатион-глютатион-глютатион-глютатион» . Биохимический журнал . 398 (3): 451–60. doi : 10.1042/bj20060424 . PMC 1559464 . PMID 16712527 .
- ^ Jump up to: а беременный в Грант П., Мага Т., Лошаков А., Сингхал Р., Вали А., Нванкво Дж. И др. (Октябрь 2016). «Обращаясь к генам торговли: идентификация четырех мутаций цвета глаз у дрозофилы» . G3 . 6 (10): 3185–3196. doi : 10.1534/g3.116.032508 . PMC 5068940 . PMID 27558665 .
- ^ Wiederrecht GJ, Brown Gm (1984). «Очистка и свойства ферментов от Drosophila melanogaster, которые катализируют преобразование дигидроптерина трихосфата в предшественник пиримидодиазепина дрозоптеринов» . Дж. Биол. Химический 259 (22): 14121–7. doi : 10.1016/s0021-9258 (18) 89865-9 . PMID 6438092 .
- ^ «Паттерны наследования у Drosophila melanogaster» . Получено 26 марта 2019 года .
- ^ Jump up to: а беременный Зеленый ММ (апрель 1952 г.). «Мутант изоаллели в локусе вермилиона в Drosophila melanogaster» . Труды Национальной академии наук Соединенных Штатов Америки . 38 (4): 300–5. Bibcode : 1952pnas ... 38..300G . doi : 10.1073/pnas.38.4.300 . PMC 1063551 . PMID 16589094 .
- ^ Oxenkrug GF (январь 2010 г.). «Расширенная продолжительность жизни мутантов с нарушением кинуренана» с нарушением формирования кинуренана » . Журнал нейронной передачи . 117 (1): 23–26. doi : 10.1007/s00702-009-0341-7 . PMC 3013506 . PMID 19941150 .
- ^ Саймон Э., Фаушэ С., Зидер А., Тезе Н., Тьебод П (июль 2016 г.). «От рудимента до рудимента: ген Drosophila, который взял крыло». Гены развития и эволюция . 226 (4): 297–315. doi : 10.1007/s00427-016-0546-3 . PMID 27116603 . S2CID 16651247 .
- ^ Томоясу Y, Ohde T, Clark-Hachtel C (14 марта 2017 г.). «Что серийные гомологи могут рассказать нам о происхождении крыльев насекомых» . F1000Research . 6 : 268. doi : 10.12688/f1000research.10285.1 . PMC 5357031 . PMID 28357056 .
- ^ Williams JA, Bell JB, Carroll SB (декабрь 1991 г.). «Контроль развития Drosophila Wing и Haltere с помощью продукта ядерного рудиментального гена» . Гены и развитие . 5 (12b): 2481–95. doi : 10.1101/gad.5.12b.2481 . PMID 1752439 .
- ^ Зеленый ММ (январь 2010 г.). «2010: столетие генетики дрозофилы через призму белого гена» . Генетика . 184 (1): 3–7. doi : 10.1534/Genetics.109.110015 . PMC 2815926 . PMID 20061564 .
- ^ Ferreiro MJ, Pérez C, Marchesano M, Ruiz S, Caputi A, Aguilera P, et al. (2018). «Rosophila melanogaster White Mutant W11118 подвергается дегенерации сетчатки» . Границы в нейробиологии . 11 : 732. DOI : 10.3389/fnins.2017.00732 . PMC 5758589 . PMID 29354028 .
- ^ Jump up to: а беременный Сяо С., Цю С., Робертсон Р.М. (август 2017 г.). «Белый ген контролирует успех из совокупления у Drosophila melanogaster» . Научные отчеты . 7 (1): 7712. BIBCODE : 2017NATSR ... 7.7712X . doi : 10.1038/s41598-017-08155-y . PMC 5550479 . PMID 28794482 .
- ^ "Джин: dmel \ y" . Flybase.org . Консорциум Flybase . Получено 26 марта 2019 года .
- ^ Wittkopp PJ, True Jr, Carroll SB (апрель 2002 г.). «Взаимные функции белков с желтым и черным деревом дрозофилы в развитии и эволюции пигментных узоров». Разработка . 129 (8): 1849–58. doi : 10.1242/dev.129.8.1849 . PMID 11934851 .
- ^ Jump up to: а беременный Biessmann H (ноябрь 1985). «Молекулярный анализ области желтого гена (Y) Drosophila melanogaster» . Труды Национальной академии наук Соединенных Штатов Америки . 82 (21): 7369–73. Bibcode : 1985pnas ... 82.7369b . doi : 10.1073/pnas.82.21.7369 . PMC 391346 . PMID 3933004 .
- ^ «NCBI (Национальный центр биотехнологической информации) База данных генома» . Получено 30 ноября 2011 года .
- ^ Halligan DL, Keightley Pd (июль 2006 г.). «Вездесущие селективные ограничения в геноме Drosophila выявлены при сравнении межвидового по всей геноме» . Исследование генома . 16 (7): 875–84. doi : 10.1101/gr.5022906 . PMC 1484454 . PMID 16751341 .
- ^ Carvalho AB (декабрь 2002 г.). «Происхождение и эволюция дрозофилы y хромосомы». Текущее мнение в области генетики и развития . 12 (6): 664–8. doi : 10.1016/s0959-437x (02) 00356-8 . PMID 12433579 .
- ^ Габальдон Т., Кунин Э.В. (май 2013 г.). «Функциональные и эволюционные последствия ортологии генов» . Природные обзоры. Генетика . 14 (5). Природная портфель : 360–366. doi : 10.1038/nrg3456 . PMC 5877793 . PMID 23552219 .
- ^ «Фон для сравнительного геномного анализа» . Национальный институт исследований генома человека. Декабрь 2002 года.
- ^ Reiter LT, Potocki L, Chien S, Gribskov M, Bier E (июнь 2001 г.). «Систематический анализ последовательностей генов, связанных с болезнью человека, у Drosophila melanogaster» . Исследование генома . 11 (6): 1114–25. doi : 10.1101/gr.169101 . PMC 311089 . PMID 11381037 .
- ^ Chien S, Reiter LT, Bier E, Gribskov M (январь 2002 г.). «Гомофила: ген болезни человека связана с дрозофилой» . Исследование нуклеиновых кислот . 30 (1): 149–51. doi : 10.1093/nar/30.1.149 . PMC 99119 . PMID 11752278 .
- ^ Jaiswal M, Sandoval H, Zhang K, Bayat V, Bellen HJ (2012). «Исследовающие механизмы, лежащие в основе нейродегенеративных заболеваний человека у дрозофилы» . Ежегодный обзор генетики . 46 : 371–96. doi : 10.1146/annurev-genet-1110711-155456 . PMC 3663445 . PMID 22974305 .
- ^ Выберите Л (2017). Мух модели человеческих болезней. Том 121 текущих тем по биологии развития . Академическая пресса. ISBN 978-0-12-802905-3 .
- ^ Бухон Н., Сильверман Н., Черри С (декабрь 2014 г.). «Иммунитет у Drosophila melanogaster-от микробного распознавания физиологии цельноорганизма» . Природные обзоры. Иммунология . 14 (12): 796–810. doi : 10.1038/nri3763 . PMC 6190593 . PMID 25421701 .
- ^ Kaun KR, Devineni Av, Heberlein U (июнь 2012 г.). «Drosophila melanogaster как модель для изучения наркомании» . Человеческая генетика . 131 (6): 959–75. doi : 10.1007/s00439-012-1146-6 . PMC 3351628 . PMID 22350798 .
- ^ Jump up to: а беременный Weigmann K, Klapper R, Strasser T, Rickert C, Technau G, Jäckle H, et al. (Июнь 2003 г.). «Flymove-новый способ взглянуть на разработку Drosophila». Тенденции в генетике . 19 (6): 310–1. doi : 10.1016/s0168-9525 (03) 00050-7 . PMID 12801722 .
- ^ Западный Эберхард MJ (май 2005 г.). «Пластичность развития и происхождение различий в видах» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (Suppl 1): 6543–9. Bibcode : 2005pnas..102.6543W . doi : 10.1073/pnas.0501844102 . PMC 1131862 . PMID 15851679 .
- ^ Jump up to: а беременный в Abram PK, Boivin G, Moiroux J, Brodeur J (ноябрь 2017). «Поведенческое воздействие температуры на эктотермических животных: объединение тепловой физиологии и поведенческой пластичности». Биологические обзоры Кембриджского философского общества . 92 (4): 1859–1876. doi : 10.1111/brv.12312 . PMID 28980433 . S2CID 9099834 .
- ^ Gibert P, Huey RB, Gilchrist GW (январь 2001 г.). «Локомоторная эффективность Drosophila melanogaster: взаимодействие между температурами развития и взрослых, возрастом и географией» . Эволюция; Международный журнал органической эволюции . 55 (1): 205–9. doi : 10.1111/j.0014-3820.2001.tb01286.x . PMID 11263741 . S2CID 2991855 .
- ^ Jump up to: а беременный в Zamudio KR, Huey RB, Crill WD (1995). «Больше не всегда лучше: размер тела, температура развития и родительская температура и мужской территориальный успех у Drosophila melanogaster». Поведение животных . 49 (3): 671–677. doi : 10.1016/0003-3472 (95) 80200-2 . ISSN 0003-3472 . S2CID 9124942 .
- ^ Харрисон Дж. Ф., Уотерс Дж.С., Биддульф Т.А., Ковачевич А., Клок С.Дж., Соча Дж.Дж. (апрель 2018). «Пластичность и стабильность развития в сети трахеи, поставляющие летной мышцы Drosophila в ответ на выращивание уровня кислорода» . Журнал физиологии насекомых . Пределы дыхательной функции: внешние и внутренние ограничения на газообмен насекомых. 106 (Pt 3): 189–198. Bibcode : 2018jinsp.106..189H . doi : 10.1016/j.jinsphys.2017.09.006 . PMID 28927826 .
- ^ Шиба V, Чандрашекаран М.К., Джоши А., Шарма В.К. (январь 2002 г.). «Пластичность развития локомоторной активности ритм Drosophila melanogaster». Журнал физиологии насекомых . 48 (1): 25–32. Bibcode : 2002jinsp..48 ... 25 с . doi : 10.1016/s0022-1910 (01) 00139-1 . PMID 12770129 .
- ^ Jump up to: а беременный в дюймовый Crill WD, Huey RB, Gilchrist GW (июнь 1996 г.). «Внутри и между поколением влияние температуры на морфологию и физиологию Drosophila melanogaster». Эволюция; Международный журнал органической эволюции . 50 (3): 1205–1218. doi : 10.2307/2410661 . JSTOR 2410661 . PMID 28565273 .
- ^ Jump up to: а беременный Дэвид -младший, Арарипе Л.О., Чакир М., Легаут Х, Лемос Б., Петави Г. и др. (Июль 2005 г.). «Мужская бесплодия при экстремальных температурах: значительное, но заброшенное явление для понимания климатических адаптаций Drosophila» . Журнал эволюционной биологии . 18 (4): 838–46. doi : 10.1111/j.1420-9101.2005.00914.x . PMID 16033555 . S2CID 23847613 .
- ^ French V, Feast M, Partridge L (ноябрь 1998 г.). «Размер тела и размер клеток у дрозофилы: реакция развития на температуру». Журнал физиологии насекомых . 44 (11): 1081–1089. Bibcode : 1998jinsp..44.1081f . doi : 10.1016/s0022-1910 (98) 00061-4 . PMID 12770407 .
- ^ Frazier MR, Harrison JF, Kirkton SD, Roberts SP (июль 2008 г.). «Холодное выращивание улучшает производительность холодного полета у Drosophila за счет изменений в морфологии крыла» . Журнал экспериментальной биологии . 211 (Pt 13): 2116–22. doi : 10.1242/jeb.019422 . PMID 18552301 .
- ^ Jump up to: а беременный Slotsbo S, Schou MF, Kristensen TN, Loeschcke V, Sørensen JG (сентябрь 2016 г.). «Обратимость тепла и холодной пластичности развития асимметрична и имеет длительные последствия для тепловой толерантности для взрослых» . Журнал экспериментальной биологии . 219 (Pt 17): 2726–32. doi : 10.1242/jeb.143750 . PMID 27353229 .
- ^ Gilchrist GW, Huey RB (январь 2001 г.). «Влияние родительской температуры и температуры развития на термическую зависимость пригодности у Drosophila melanogaster» . Эволюция; Международный журнал органической эволюции . 55 (1): 209–14. doi : 10.1111/j.0014-3820.2001.tb01287.x . PMID 11263742 . S2CID 1329035 .
- ^ Остин CJ, Moehring AJ (май 2013). «Оптимальный температурный диапазон пластиковых видов, Drosophila Simulans» . Журнал экологии животных . 82 (3): 663–72. Bibcode : 2013, Janec..82..663a . doi : 10.1111/1365-2656.12041 . PMID 23360477 .
- ^ Jump up to: а беременный Hodin J, Riddiford LM (октябрь 2000 г.). «Различные механизмы лежат в основе фенотипической пластичности и межвидовых изменений для репродуктивного характера у дрозофилидов (Insecta: Diptera)» . Эволюция; Международный журнал органической эволюции . 54 (5): 1638–53. doi : 10.1111/j.0014-3820.2000.tb00708.x . PMID 11108591 . S2CID 6875815 .
- ^ Klepsatel P, Girish TN, Dircksen H, Gáliková M (май 2019). «Drosophila максимизируется по оптимальной температуре развития» . Журнал экспериментальной биологии . 222 (Pt 10): Jeb202184. doi : 10.1242/jeb.202184 . PMID 31064855 .
- ^ Schou MF, Kristensen TN, Pedersen A, Karlsson BG, Loesckke V, Malmendal A (февраль 2017 г.). «Метаболическая и функциональная характеристика эффектов температуры развития у Drosophila melanogaster» . Американский журнал физиологии. Нормативно -правовая, интегративная и сравнительная физиология . 312 (2): R211 - R222. doi : 10.1152/ajpregu.00268.2016 . PMC 5336569 . PMID 27927623 .
- ^ Кохет Y, Дэвид Дж (январь 1978 г.). «Контроль репродуктивного потенциала взрослых с помощью преимагинальных термических условий: исследование в Drosophila melanogaster». Oecologia . 36 (3): 295–306. Bibcode : 1978oecol..36..295c . doi : 10.1007/bf00348055 . PMID 28309916 . S2CID 12465060 .
- ^ Rideout EJ, Narsaiya MS, Grewal SS (декабрь 2015 г.). «Трансформер определения пола регулирует различия в мужчинах-жене-жене в размере тела дрозофилы» . PLOS Genetics . 11 (12): E1005683. doi : 10.1371/journal.pgen.1005683 . PMC 4692505 . PMID 26710087 .
- ^ Гилберт С.Ф. (2000). Биология развития (6 -е изд.). Sunderland (MA): Sinauer Associates; 2000. ISBN 978-0-87893-243-6 .
- ^ Lemaitre B, Hoffmann J (2007). «Защита хозяина Drosophila melanogaster» (PDF) . Ежегодный обзор иммунологии . 25 : 697–743. doi : 10.1146/annurev.immunol.25.022106.141615 . PMID 17201680 .
- ^ Troha K, IM JH, Revah J, Lazzaro BP, Buchon N (февраль 2018 г.). «Сравнительная транскриптомика показывает Кребу как новый регулятор толерантности к инфекции у D. melanogaster» . PLO -патогены . 14 (2): E1006847. doi : 10.1371/journal.ppat.1006847 . PMC 5812652 . PMID 29394281 .
- ^ Де Грегорио Е., Спеллман П.Т., Цу П., Рубин Г.М., Лемитр Б (июнь 2002 г.). «Пути платы и IMD являются основными регуляторами иммунного ответа у дрозофилы» . Embo Journal . 21 (11): 2568–79. doi : 10.1093/emboj/21.11.2568 . PMC 126042 . PMID 12032070 .
- ^ Paredes JC, Welchman DP, Poidevin M, Lemaitre B (ноябрь 2011 г.). «Отрицательная регуляция амидазой PGRP формирует антибактериальный ответ Drosophila и защищает муху от безобидной инфекции» (PDF) . Иммунитет . 35 (5): 770–9. doi : 10.1016/j.immuni.2011.09.018 . PMID 22118526 .
- ^ Троха К, Бухон Н. (сентябрь 2019 г.). «Методы изучения врожденного иммунитета у Drosophila melanogaster». Wiley междисциплинарные обзоры. Биология развития . 8 (5): E344. doi : 10.1002/wdev.344 . PMID 30993906 . S2CID 119527642 .
- ^ Гилберт Р., Торрес М., Клеменс Р., Хатели С., Хосамани Р., Уэйд В., Бхаттачарья С. (февраль 2020 г.). «Модель инфекции Drosophila melanogaster» . NPJ микрогравитация . 6 (1): 4. doi : 10.1038/s41526-019-0091-2 . PMC 7000411 . PMID 32047838 .
- ^ Jump up to: а беременный Dostálová A, Rommelaere S, Poidevin M, Lemaitre B (сентябрь 2017 г.). «Тиоэфирные белки регулируют платный путь и играют роль в защите дрозофилы от микробных патогенов и паразитоидных ос» . BMC Biology . 15 (1): 79. DOI : 10.1186/S12915-017-0408-0 . PMC 5584532 . PMID 28874153 .
- ^ Шринивасан Н., Гордон О., Аренс С., Франц А., Деддуш С., Чакраварти П. и др. (Ноябрь 2016). «Дрозофила Меланогастер» . элиф . 5 doi : 10.7554/elife.19662 . PMC 5138034 . PMID 27871362 .
- ^ Goto A, Yano T, Terashima J, Iwashita S, Oshima Y, Kurata S (май 2010 г.). «Совместная регуляция индукции нового антибактериального листерицина белком распознавания пептидогликана и пути Jak-Stat» . Журнал биологической химии . 285 (21): 15731–8. doi : 10.1074/jbc.m109.082115 . PMC 2871439 . PMID 20348097 .
- ^ Wang L, Kounatidis I, Ligoxygakis P (январь 2014 г.). «Drosophila как модель для изучения роли клеток крови в воспалении, врожденного иммунитета и рака» . Границы в клеточной и инфекционной микробиологии . 3 : 113. DOI : 10.3389/fcimb.2013.00113 . PMC 3885817 . PMID 24409421 .
- ^ Neyen C, Bretscher AJ, Binggeli O, Lemaitre B (июнь 2014 г.). «Методы изучения иммунитета Drosophila» (PDF) . Методы 68 (1): 116–28. doi : 10.1016/j.ymeth.2014.02.023 . PMID 24631888 .
- ^ Hashimoto Y, Tabuchi Y, Sakurai K, Kutsuna M, Kurokawa K, Awasaki T, et al. (Декабрь 2009 г.). «Идентификация липотеховой кислоты в качестве лиганда для драпировки при фагоцитозе стафилококка гемоцитов дрозофилы» . Журнал иммунологии . 183 (11): 7451–60. doi : 10.4049/jimmunol.0901032 . PMID 19890048 .
- ^ Хольц А., Боссингер Б., Страссер Т., Яннинг В., Клаппер Р. (октябрь 2003 г.). «Два происхождения гемоцитов у дрозофилы» . Разработка . 130 (20): 4955–62. doi : 10.1242/dev.00702 . PMID 12930778 .
- ^ Санчес Бош П., Махиджани К., Хербосо Л., Голд К.С., Багински Р., Вудкок К.Дж. и др. (Декабрь 2019). «У взрослого у взрослой у дрозофилы гематопоэз, но полагаются на резервуар клеток клетки в дыхательном эпителии, чтобы передать сигналы инфекции в окружающие ткани» . Ячейка развития . 51 (6): 787–803.e5. doi : 10.1016/j.devcel.2019.10.017 . PMC 7263735 . PMID 31735669 .
- ^ Parvy JP, Yu Y, Dostalova A, Kondo S, Kurjan A, Bulet P, et al. (July 2019). "Drosophila" . eLife . 8 : e45061. doi : 10.7554/eLife.45061 . PMC 6667213 . PMID 31358113 .
- ^ Sturtvant AH (1929). «Кларет -мутант типа симулянцев дрозофилы: исследование элиминации хромосом и клеточной линии». Журнал научной зоологии . 135 : 323–356.
- ^ Nissani M (май 1975). «Новый поведенческий биоанализа для анализа сексуального влечения и феромонов у насекомых» . Журнал экспериментальной зоологии . 192 (2): 271–5. Bibcode : 1975jez ... 192..271n . doi : 10.1002/jez.1401920217 . PMID 805823 .
- ^ Хан Ф.А. (2011). Биотехнологические основы . CRC Press. п. 213. ISBN 978-1-4398-2009-4 .
- ^ «Нобелевская премия 2017 года по физиологии или медицине совместно с Джеффри С. Холлом, Майклом Росбашем и Майклом В. Янгом за их открытия молекулярных механизмов, контролирующих циркадный ритм» . Nobelprize.org. 2 октября 2017 года . Получено 5 октября 2017 года .
- ^ Lehnert BP, Baker AE, Gaudry Q, Chiang AS, Wilson RI (январь 2013 г.). «Отличительные роли каналов TRP в слуховой трансдукции и амплификации у дрозофилы» . Нейрон . 77 (1): 115–28. doi : 10.1016/j.neuron.2012.11.030 . PMC 3811118 . PMID 23312520 .
- ^ Zhang W, Yan Z, Jan Ly, Jan Yn (август 2013 г.). «Звуковая реакция, опосредованная каналами TRP Nompc, Nanhung и неактивно в ходотональных органах личинок дрозофилы» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (33): 13612–7. Bibcode : 2013pnas..11013612Z . doi : 10.1073/pnas.1312477110 . PMC 3746866 . PMID 23898199 .
- ^ "Гомосексуализм включался и выключается в плодовых мухах"
- ^ Dasgupta S, Sheehan TC, Stevens CF, Navlakha S (декабрь 2018 г.). «Нейронная структура данных для обнаружения новизны» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (51): 13093–13098. Bibcode : 2018pnas..11513093d . doi : 10.1073/pnas.1814448115 . PMC 6304992 . PMID 30509984 .
- ^ Zwarts L, Versteven M, Callaerts P (1 января 2012 г.). «Генетика и нейробиология агрессии у дрозофилы» . Летать . 6 (1): 35–48. doi : 10.4161/fly.19249 . PMC 3365836 . PMID 22513455 .
- ^ Дэвис С.М., Томас Ал, Лю Л., Кэмпбелл И.М., Дирик Х.А. (январь 2018 г.). «Дрозофила с использованием экрана для повреждения крыла» . Генетика . 208 (1): 273–282. doi : 10.1534/Genetics.117.300292 . PMC 5753862 . PMID 29109180 .
- ^ Jump up to: а беременный в Versteven M, Vanden Broeck L, Geurten B, Zwarts L, Decraecker L, Beelen M, et al. (Февраль 2017 г.). "Дрозофила агрессия" . Труды Национальной академии наук Соединенных Штатов Америки . 114 (8): 1958–1963. doi : 10.1073/pnas.1605946114 . PMC 5338383 . PMID 28115690 .
- ^ Jump up to: а беременный Сенгупта С., Смит Д.П. (2014). «Как Drosophila обнаруживает летучие феромоны: передача сигналов, схемы и поведение». В Mucignat-Caretta C (ред.). Нейробиология химической связи . Границы в нейробиологии. CRC Press/Taylor & Francis. ISBN 978-1-4665-5341-5 Полем PMID 24830032 . Получено 30 мая 2019 года .
- ^ Laturney M, Billeter JC (август 2016 г.). «Женщины Drosophila melanogaster восстанавливают свою привлекательность после спаривания, удаляя мужские анти-афродизиак-феромоны» . Природная связь . 7 (1): 12322. Bibcode : 2016natco ... 712322L . doi : 10.1038/ncomms12322 . PMC 4976142 . PMID 27484362 .
- ^ Wang L, Han X, Mehren J, Hiroi M, Billeter JC, Miyamoto T, et al. (Июнь 2011 г.). «Иерархическая хемосенсорная регуляция социальных взаимодействий мужского мужского мужчин у дрозофилы» . Nature Neuroscience . 14 (6): 757–62. doi : 10.1038/nn.2800 . PMC 3102769 . PMID 21516101 .
- ^ Jump up to: а беременный в Lim RS, Eyjólfsdóttir E, Shin E, Perona P, DJ Anderson (27 августа 2014 г.). «Как пища контролирует агрессию у дрозофилы» . Plos один . 9 (8): E105626. BIBCODE : 2014PLOSO ... 9J5626L . doi : 10.1371/journal.pone.0105626 . PMC 4146546 . PMID 25162609 .
- ^ Эрион Р., Диангело младший, Крокер А., Сегал А (сентябрь 2012 г.). «Взаимодействие между сном и метаболизмом у дрозофилы с измененной передачей сигналов осьупамина» . Журнал биологической химии . 287 (39): 32406–14. doi : 10.1074/jbc.m112.360875 . PMC 3463357 . PMID 22829591 .
- ^ Jump up to: а беременный в Kayser MS, Mainwaring B, Yue Z, Sehgal A (июль 2015 г.). Гриффит LC (ред.). «Лишение сна подавляет агрессию у дрозофилы» . элиф . 4 : E07643. doi : 10.7554/elife.07643 . PMC 4515473 . PMID 26216041 .
- ^ Hardie RC, Raghu P (сентябрь 2001 г.). «Визуальная трансдукция у дрозофилы». Природа . 413 (6852): 186–93. Bibcode : 2001natur.413..186H . doi : 10.1038/35093002 . PMID 11557987 . S2CID 4415605 .
- ^ Jump up to: а беременный в дюймовый Шарки К.Р., Бланко Дж., Лейбовиц М.М., Пинто-Бенито Д., Вардлл Т.Дж. (октябрь 2020 г.). «Спектральная чувствительность фоторецепторов дрозофилы» . Научные отчеты . 10 (1): 18242. BibCode : 202020NATSR..1018242S . doi : 10.1038/s41598-020-74742-1 . PMC 7588446 . PMID 33106518 . S2CID 215551298 .
 Материал был скопирован и адаптирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован и адаптирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Feuda R, Goulty M, Zadra N, Gasparetti T, Rosato E, Pisani D, et al. (Август 2021 г.). «Филогеномика генов опсина в Diptera выявляет специфичные для линии события и контрастную эволюционную динамику у анофелей и дрозофила» . Биология и эволюция генома . 13 (8): EVAB170. doi : 10.1093/gbe/evab170 . PMC 8369074 . PMID 34270718 .
- ^ Харрис В.А., Старк У.С., Уокер Дж.А. (апрель 1976 г.). «Генетическое рассечение фоторецепторной системы в составном глазах Drosophila melanogaster» . Журнал физиологии . 256 (2): 415–439. doi : 10.1113/jphysiol.1976.sp011331 . PMC 1309314 . PMID 16992509 . S2CID 45888026 .
- ^ Jump up to: а беременный Ostroy SE, Wilson M, Pak WL (август 1974 г.). «Drosophila Rhodopsin: фотохимия, экстракция и различия в фототрансдукционном мутанте NORP AP12». Биохимическая и биофизическая исследовательская коммуникация . 59 (3): 960–966. doi : 10.1016/s0006-291x (74) 80073-2 . PMID 4213042 .
- ^ Ostroy SE (ноябрь 1978 г.). «Характеристики родопсина дрозофилы у мутантов трансдукции зрения дикого типа и Norpa» . Журнал общей физиологии . 72 (5): 717–732. doi : 10.1085/jgp.72.5.717 . PMC 2228556 . PMID 105082 . S2CID 5802525 .
- ^ Jump up to: а беременный в дюймовый и фон глин Сальседо Э., Хубер А., Хенрих С., Чедвелл Л.В., Чоу В.Х., Полсен Р., Бритт С.Г. (декабрь 1999 г.). «Голубое и зеленого поглощающего визуальные пигменты дрозофилы: эктопическая экспрессия и физиологическая характеристика родопсинов фоторецепторов R8 и родопсинов Rh5 и Rh6» . Журнал нейробиологии . 19 (24): 10716–10726. doi : 10.1523/jneurosci.19-24-10716.1999 . PMC 6784940 . PMID 10594055 . S2CID 17575850 .
- ^ Kirschfeld K, Franceschini N, Minke B (сентябрь 1977 г.). «Свидетельство чувствительного пигмента у фоторецепторов мух». Природа . 269 (5627): 386–390. Bibcode : 1977natur.269..386k . doi : 10.1038/269386a0 . PMID 909585 . S2CID 28890008 .
- ^ Minke B, Kirschfeld K (май 1979 г.). «Вклад сенсибилизирующего пигмента в спектры фоточувствительности мухового родопсина и метарходопсина» . Журнал общей физиологии . 73 (5): 517–540. doi : 10.1085/jgp.73.5.517 . PMC 2215190 . PMID 458418 . S2CID 12451748 .
- ^ Jump up to: а беременный Kirschfeld K, Feiler R, Franceschini N (1978). «Фотостабильный пигмент в рабдоде фоторецепторов мухи № 7». Журнал сравнительной физиологии а . 125 (3): 275–284. doi : 10.1007/bf00656606 . S2CID 40233531 .
- ^ Kirschfeld K, Franceschini N (июнь 1977 г.). «Фотостабильные пигменты в мембране фоторецепторов и их возможная роль». Биофизика структуры и механизма . 3 (2): 191–194. doi : 10.1007/bf00535818 . PMID 890056 . S2CID 5846094 .
- ^ Zuker CS, Montell C, Jones K, Laverty T, Rubin GM (май 1987). «Ген родопсина, экспрессируемый в фоторецепторной клетке R7 глаза Drosophila: гомологии с другими молекулами, трансдуцирующими сигнал» . Журнал нейробиологии . 7 (5): 1550–1557. doi : 10.1523/jneurosci.07-05-01550.1987 . PMC 6568820 . PMID 2437266 . S2CID 1490332 .
- ^ Jump up to: а беременный в Chou WH, Hall KJ, Wilson DB, Wideman CL, Townson SM, Chadwell LV, Britt SG (декабрь 1996 г.). «Идентификация нового Drosophila opsin выявляет специфические паттерны фоторецепторных клеток R7 и R8» . Нейрон . 17 (6): 1101–1115. doi : 10.1016/s0896-6273 (00) 80243-3 . PMID 8982159 . S2CID 18294965 .
- ^ Jump up to: а беременный в дюймовый Feiler R, Bjornson R, Kirschfeld K, Mismer D, Rubin GM, Smith DP, et al. (Октябрь 1992). «Эктопическая экспрессия ультрафиолетовых-раодопсинов в синих фоторецепторных клетках дрозофилы: визуальная физиология и фотохимия трансгенных животных» . Журнал нейробиологии . 12 (10): 3862–3868. doi : 10.1523/jneurosci.12-10-03862.1992 . PMC 6575971 . PMID 1403087 .
- ^ Монтелл С., Джонс К., Зукер С., Рубин Г (май 1987). «Второй ген опсина, экспрессируемый в чувствительных к ультрафиолетовым фоторецепторным клеткам R7 Drosophila melanogaster» . Журнал нейробиологии . 7 (5): 1558–1566. doi : 10.1523/jneurosci.07-05-01558.1987 . PMC 6568825 . PMID 2952772 . S2CID 17003459 .
- ^ Хубер А., Шульц С., Бентроп Дж., Гроэлл С., Вольфрум У, Паулсен Р. (апрель 1997 г.). «Молекулярное клонирование родопсина Drosophila RH6: визуальный пигмент подмножества фоторецепторных клеток R8». Письма Febs . 406 (1–2): 6–10. doi : 10.1016/s0014-5793 (97) 00210-x . PMID 9109375 . S2CID 18368117 .
- ^ Поллок JA, Benzer S (июнь 1988 г.). «Локализация транскрипта четырех генов опсина в трех визуальных органах дрозофилы; RH2 специфична для глаз». Природа . 333 (6175): 779–782. Bibcode : 1988natur.333..779p . doi : 10.1038/333779A0 . PMID 2968518 . S2CID 4303934 .
- ^ Jump up to: а беременный Feiler R, Harris WA, Kirschfeld K, Wehrhahn C, Zuker CS (июнь 1988 г.). «Целевая мисекспрессия гена Drosophila Opsin приводит к измененной визуальной функции». Природа . 333 (6175): 737–741. Bibcode : 1988natur.333..737f . doi : 10.1038/333737A0 . PMID 2455230 . S2CID 4248264 .
- ^ Сакаи К, Цуцуи К, Ямашита Т., Ивабе Н., Такахаши К, Вада А., Шичида Y (август 2017 г.). «Drosophila melanogaster Rhodopsin RH7 представляет собой датчик света с ультрафиолетом с визицией с чрезвычайно широким спектром поглощения» . Научные отчеты . 7 (1): 7349. Bibcode : 2017Natsr ... 7.7349S . doi : 10.1038/s41598-017-07461-9 . PMC 5544684 . PMID 28779161 . S2CID 3276084 .
- ^ Ni JD, Baik LS, Holmes TC, Montell C (май 2017). «Родопсин в мозге функционирует в циркадной фотоэлементной расцвете у дрозофилы» . Природа . 545 (7654): 340–344. Bibcode : 2017natur.545..340n . doi : 10.1038/nature22325 . PMC 5476302 . PMID 28489826 . S2CID 4468254 .
- ^ Vogt K (1 февраля 1984 г.). «Хромофор визуального пигмента в некоторых порядках насекомых» . Журнал естественных исследований c . 39 (1–2): 196–197. Doi : 10.1515/Znc-1984-1-236 . S2CID 88980658 .
- ^ Vogt K, Kirschfeld K (апрель 1984 г.). «Химическая идентичность хромофоров визуального пигмента мухи». Naturwissenschaften . 71 (4): 211–213. Bibcode : 1984nw ..... 71..211v . doi : 10.1007/bf00490436 . S2CID 24205801 .
- ^ Гюманн М., Портер М.Л., Бок М.Дж (август 2022). «Глуопсины: опсины без лизина, связывающего сетчатки» . Ячейки 11 (15): 2441. doi : 10.3390/cells11152441 . PMC 9368030 . PMID 35954284 .
- ^ Katana R, Guan C, Zanini D, Larsen ME, Giraldo D, Geurten BR, et al. (Сентябрь 2019). «Независимая от хромофора роли апопротеинов опсина у механорецепторов Drosophila» . Текущая биология . 29 (17): 2961–2969.e4. BIBCODE : 2019CBIO ... 29E2961K . doi : 10.1016/j.cub.2019.07.036 . PMID 31447373 . S2CID 201420079 .
- ^ Jump up to: а беременный Leung NY, Thakur DP, Gurav AS, Kim Sh, Di Pizio A, Niv MY, Montell C (апрель 2020 г.). «Функции опсинов при вкусе дрозофилы» . Текущая биология . 30 (8): 1367–1379.e6. Bibcode : 2020CBIO ... 30E1367L . doi : 10.1016/j.cub.2020.01.068 . PMC 7252503 . PMID 32243853 .
- ^ Kumbalasiri T, Rollag MD, Isoldi MC, Castrucci AM, Provencio I (март 2007 г.). «Меланопсин запускает высвобождение внутренних магазинов кальция в ответ на свет». Фотохимия и фотобиология . 83 (2): 273–279. doi : 10.1562/2006-07-11-ra-964 . PMID 16961436 . S2CID 23060331 .
- ^ Jump up to: а беременный Raghu P, Colley NJ, Webel R, James T, Hasan G, Danin M, et al. (Май 2000). «Нормальная фототрансдукция у фоторецепторов дрозофилы, в которых отсутствует ген рецептора Insp (3)». Молекулярные и клеточные нейронауки . 15 (5): 429–45. doi : 10.1006/mcne.2000.0846 . PMID 10833300 . S2CID 23861204 .
- ^ Hardie RC, Juusola M (октябрь 2015 г.). «Фототрансдукция у дрозофилы» . Современное мнение о нейробиологии . 34 : 37–45. doi : 10.1016/j.conb.2015.01.008 . PMID 25638280 . S2CID 140206989 .
- ^ Wang T, Xu H, Oberwinkler J, Gu Y, Hardie RC, Montell C (февраль 2005 г.). «Функции света, адаптация и выживание ячейки NA+/CA2+ CALX CALX» . Нейрон . 45 (3): 367–78. doi : 10.1016/j.neuron.2004.12.046 . PMID 15694324 .
- ^ Рейн К, Зёклер М., Мадер М.Т., Грюбель С., Гейзенберг М (февраль 2002 г.). «Стандартный мозг дрозофилы» . Текущая биология . 12 (3): 227–31. Bibcode : 2002cbio ... 12..227r . doi : 10.1016/s0960-9822 (02) 00656-5 . PMID 11839276 . S2CID 15785406 .
- ^ Докинс Р., Докинс М. (1976). «Иерархическая организация и постуральная облегчение: правила для ухода за мухами». Поведение животных . 24 (4): 739–755. doi : 10.1016/s0003-3472 (76) 80003-6 . S2CID 53186674 .
- ^ Jump up to: а беременный в Дэвис WJ (1979). «Поведенческие иерархии». Тенденции в нейронауках . 2 (2): 5–7. doi : 10.1016/0166-2236 (79) 90003-1 . S2CID 53180462 .
- ^ Jump up to: а беременный Семена А.М., Равабар П., Чунг П., Хэмпель С., Мидгли Ф.М., Менш Б.Д., Симпсон Дж. Х. (август 2014 г.). «Иерархия подавления среди конкурирующих моторных программ движет последовательным грумингом у дрозофилы» . элиф . 3 : E02951. doi : 10.7554/elife.02951 . PMC 4136539 . PMID 25139955 .
- ^ Матис А., Мамиданна П., Кьюри К.М., Абэ Т., Мурти В.Н., Матис М.В., Бетге М (сентябрь 2018 г.). «Deeplabcut: без маркерная поза оценка пользовательских частей тела с глубоким обучением». Nature Neuroscience . 21 (9): 1281–1289. doi : 10.1038/s41593-018-0209-y . PMID 30127430 . S2CID 4748395 .
- ^ Штраус Р., Гейзенберг М (август 1990 г.). «Координация ног во время прямой ходьбы и поворота в Drosophila melanogaster». Журнал сравнительной физиологии а . 167 (3): 403–12. doi : 10.1007/bf00192575 . PMID 2121965 . S2CID 12965869 .
- ^ Deangelis BD, Zavatone-Veth Ja, Clark Da (июнь 2019). «Дрозофила» . элиф . 8 doi : 10.7554/elife.46409 . PMC 6598772 . PMID 31250807 .
- ^ Wosnitza A, Bockemühl T, Dübbert M, Scholz H, Büschges A (февраль 2013 г.). «Координация между ногами в контроле скорости ходьбы у дрозофилы» . Журнал экспериментальной биологии . 216 (Pt 3): 480–91. doi : 10.1242/jeb.078139 . PMID 23038731 .
- ^ Mendes CS, Bartos I, Akay T, Márka S, Mann RS (январь 2013 г.). «Количественная оценка параметров походки в свободной ходьбе дикого типа и чувствительности, лишенной Drosophila melanogaster» . элиф . 2 : E00231. doi : 10.7554/elife.00231 . PMC 3545443 . PMID 23326642 .
- ^ Zczecinski NS, Bockemühl T, Chockley AS, Büschges A (ноябрь 2018 г.). «Дрозофила» . Журнал экспериментальной биологии . 221 (Pt 22): JEB189142. doi : 10.1242/jeb.189142 . PMID 30274987 .
- ^ Hooper SL (май 2012 г.). «Размер тела и нервный контроль движения» . Текущая биология . 22 (9): R318-22. Bibcode : 2012cbio ... 22.r318h . doi : 10.1016/j.cub.2012.02.048 . PMID 22575473 .
- ^ Jump up to: а беременный в Fry Sn, Sayaman R, Dickinson MH (апрель 2003 г.). «Аэродинамика маневров свободного полета у дрозофилы» (PDF) . Наука . 300 (5618): 495–8. Bibcode : 2003sci ... 300..495f . doi : 10.1126/science.1081944 . PMID 12702878 . S2CID 40952385 . Архивировано из оригинала (PDF) 24 сентября 2015 года.
- ^ Hesselberg T, Lehmann FO (декабрь 2007 г.). «Поворот поведение зависит от демпфирования трения в фруктовой мух дрозофила» . Журнал экспериментальной биологии . 210 (Pt 24): 4319–34. doi : 10.1242/jeb.010389 . PMID 18055621 .
- ^ Chiang AS, Lin Cy, Chuang CC, Chang HM, Sieh CH, Yeh CW, et al. (Январь 2011). «Трехмерная реконструкция сетей проводки в масштабах мозга у дрозофилы при разрешении с одним клетками» . Текущая биология . 21 (1): 1–11. Bibcode : 2011cbio ... 21 .... 1c . doi : 10.1016/j.cub.2010.11.056 . PMID 21129968 . S2CID 17155338 .
- ^ «FlyCircuit - база данных нейронов мозга Drosophila» . Получено 30 августа 2013 года .
- ^ Meinertzhagen IA, O'Neil SD (март 1991 г.). «Синаптическая организация столбчатых элементов в пластинке дикого типа в Drosophila melanogaster». Журнал сравнительной неврологии . 305 (2): 232–63. doi : 10.1002/cne.903050206 . PMID 1902848 . S2CID 35301798 .
- ^ Rivera-Alba M, Vitaladevuni SN, Mishchenko Y, Mischenko Y, Lu Z, Takemura Sy, et al. (Декабрь 2011 г.). «Экономика проводки и исключение объема определяют размещение нейронов в мозге дрозофилы» . Текущая биология . 21 (23): 2000–5. Bibcode : 2011cbio ... 21.2000r . doi : 10.1016/j.cub.2011.10.022 . PMC 3244492 . PMID 22119527 .
- ^ Takemura Sy, Bharioke A, Lu Z, Nern A, Vitaladevuni S, Rivlin PK, et al. (Август 2013). «Визуальная схема обнаружения движения, предложенная Drosophila Connectomics» . Природа . 500 (7461): 175–81. Bibcode : 2013natur.500..175t . doi : 10.1038/nature12450 . PMC 3799980 . PMID 23925240 .
- ^ Takemura Sy, Aso Y, Hige T, Wong A, Lu Z, Xu CS, et al. (Июль 2017). «Дрозофила мозг» . элиф . 6 : E26975. doi : 10.7554/elife.26975 . PMC 5550281 . PMID 28718765 .
- ^ «Целый фруктовый мух, изображенный с помощью электронной микроскопии» . Ученый журнал . Получено 15 июля 2018 года .
- ^ Zheng Z, Lauritzen JS, Perlman E, Robinson CG, Nichols M, Milkie D, et al. (Июль 2018). «Полный объем электронной микроскопии мозга взрослого дрозофилы меланогастера» . Клетка . 174 (3): 730–743.E22. Biorxiv 10.1101/140905 . doi : 10.1016/j.cell.2018.06.019 . PMC 6063995 . PMID 30033368 .
- ^ Xu CS, Januszewski M, Lu Z, Takemura SY, Hayworth K, Huang G, Shinomiya K, Maitin-Shepard J, Ackerman D, Berg S, Blakely T, et al. (2020). «Коннективы взрослого дрозофилы центрального мозга». Biorxiv . Лаборатория Cold Spring Harbour: 2020.01.21.911859. doi : 10.1101/2020.01.21.911859 . S2CID 213140797 .
- ^ «Инструменты анализа для разъединения» . HHMI.
- ^ Фелпс, Джаспер С.; Хильдебранд, Дэвид Грант Колберн; Грэм, Бретт Дж.; Куан, Аарон Т.; Томас, Логан А.; Нгуен, Три М.; Буманн, Джулия; Азеведо, Энтони В.; Устой, Энн; Agrawal, Sweta; Лю, Мингуан; Shanny, Brendan L.; Функе, Ян; Tuthill, John C.; Ли, Вэй-Чунг Аллен (2 февраля 2021 г.). «Реконструкция моторных схем управления у взрослых дрозофилы с использованием автоматизированной проникновенной электронной микроскопии» . Клетка . 184 (3): 759–774.E18. doi : 10.1016/j.cell.2020.12.013 . PMC 8312698 . PMID 33400916 .
- ^ Scheffer LK, Xu CS, Januszewski M, Lu Z, Takemura SY, Hayworth KJ, Huang G, Shinomiya K, Maitlin-Shepard J, Berg S, Clements J, et al. (2020). «Коннектика и анализ центрального мозга взрослого дрозофилы » . Biorxiv . 9 Холодная весенняя гавань. doi : 10.1101/2020.04.07.030213 . PMC 7546738 . PMID 32880371 . S2CID 215790785 .
- ^ Попович, Сергия; Макрина, Томас; Кемниц, Нико; Кастро, Мануэль; Нехоран, Барак; Цзя, Чжэнь; Бэ, Дж. Александр; Митчелл, Эрик; Му, Шан; Траутман, Эрик Т.; Саалфельд, Стефан; Ли, Кай; Сын, Себастьян (27 марта 2022 г.). «Petaskale Pipeline для точного выравнивания изображений из электронной микроскопии последовательной серии». Biorxiv 10.1101/2022.03.25.485816 .
- ^ Обмотка, Майкл; Педиго, Бенджамин Д.; Барнс, Кристофер Л.; Патсолика, Хизер Г.; Парк, Янгзер; Казимирс, Том; Фушики, Акира; Андраде, Ингрид В.; Хандельвал, Авинаш; Вальдес-Алман, Хавьер; Ли, Фэн; Рандель, Надин; Барсотти, Элизабет; Коррейя, Ана; Феттер, Ричард Д. (10 марта 2023 г.). «Коннектика мозга насекомых» . Наука . 379 (6636): EADD9330. doi : 10.1126/science.add9330 . PMC 7614541 . PMID 36893230 .
- ^ «Не вредители» . ЗДОРОВЬЯ ЗДОРОВЬЯ Австралия . Получено 19 сентября 2017 года .
- ^ McEvey S (5 февраля 2014 г.). «Фруктовые мухи: случай ошибочной идентичности» . Австралийский музей . Получено 19 сентября 2017 года .
Дальнейшее чтение
[ редактировать ]- Колер Р.Е. (1994). Лорды мухи: генетика Drosophila и экспериментальная жизнь . Чикаго: Университет Чикагской Прессы. ISBN 978-0-226-45063-6 .
- Гилберт С.Ф. (2000). Биология развития (6 -е изд.). Sunderland (MA): Sinauer Associates; 2000. ISBN 978-0-87893-243-6 .
- Perrimon N, Bonini NM, Dhillon P (март 2016 г.). «Фруктовые мухи на линии фронта: трансляционное воздействие дрозофилы» . Модели заболевания и механизмы . 9 (3): 229–31. doi : 10.1242/dmm.024810 . PMC 4833334 . PMID 26935101 .
- Хендерсон М (8 апреля 2010 г.). "Row Over Fruit Fly Drosophila melanogaster name Bugs Ученые" . Время . Австралийский . Получено 19 сентября 2017 года .
Внешние ссылки
[ редактировать ]- «Быстрое и простое введение в Drosophila melanogaster » . Виртуальная библиотека Drosophila .
- «Центр ресурсов геномики Drosophila» - собирает, поддерживает и распределяет Drosophila . клоны ДНК и клеточные линии
- «Блумингтон Drosophila Stock Center» - собирает, поддерживает и распределяет Drosophila melanogaster для исследования штаммы
- «Flybase - база данных генов и геномов Drosophila» .
- "NCBI MAP Viewer - Drosophila melanogaster " .
- «Виртуальная библиотека Drosophila» .
- «Проект генома Беркли Drosophila » .
- "Flymove" . - видео ресурсы для Drosophila разработки
- « Номенклатура Drosophila - Навидение генов» . Архивировано из оригинала 8 октября 2011 года.
- Посмотреть геном Fruitfly на ансамбле
- Посмотреть сборку генома DM6 в браузере генома UCSC .
- Манчестер-флайс-для общественного архивирования 2015-05-13 на машине Wayback от Университета Манчестера
- Веб-сайт Droso4schools с учебными ресурсами о Drosophila
- Часть 1 образовательных видео «Маленькая муха: большое влияние», объясняющая историю и важность модельного организма Дрозофила .
- Часть 2 образовательных видео «Маленькая муха: большое влияние», объясняющее, как исследования проводится в Drosophila .
- «Внутри лаборатории Fly» - Broadcast от WGBH и PBS , в серии программ «Любопытно» , январь 2008 года.
- «Как муха обнаруживает яд», архивировав 2013-01-13 на archive.today --whyfiles.org в статье описывается, как фруктовая муха на вкус с личинкой, убивающей химическое вещество в пище.
| Базы данных управления авторитетом : национальный |
|---|