Старение
Старение ( / s ɪ ˈ n ɛ s ə n s / ) или биологическое старение - это постепенное ухудшение функциональных характеристик в живых организмах. Старение целого организма включает в себя увеличение показателей смертности или снижение плодовитости организма с увеличением возраста, по крайней мере, в более поздней части жизненного цикла . [ 1 ] [ 2 ] Однако полученные последствия старения могут быть отложены. Открытие 1934 года о том, что ограничение калорий может продлевать продолжительность жизни на 50% у крыс, существование видов, имеющих незначительное старение , и существование потенциально бессмертных организмов, таких как члены рода , мотивировали исследование задержки старения и, следовательно, возрастных заболеваний . Редкие человеческие мутации могут вызвать ускоренные болезни старения .
Факторы окружающей среды могут влиять на старение - например, сверхэкспонирование ультрафиолетового излучения ускоряет старение кожи . Различные части тела могут возрастать с разными скоростями и отчетливо, включая мозг , сердечно -сосудистую систему и мышцы. Точно так же функции могут отчетливо снижаться со старением, включая управление движением и память . Два организма одного и того же вида также могут возрастать с различными показателями, что делает биологическое старение и хронологическое старение различными концепциями.
Определение и характеристики
[ редактировать ]Организмальное старение - это старение целых организмов. Актуарное старение может быть определено как увеличение смертности или снижение плодовитости с возрастом. говорит Закон о смертности Гомперца-Макехэма , что возрастный компонент уровня смертности возрастает в геометрической прогрессии с возрастом.
Старение характеризуется снижением способности реагировать на стресс, повышение гомеостатического дисбаланса и повышенный риск заболеваний, связанных с старением, включая рак и болезни сердца . Старение было определено как «прогрессирующее ухудшение физиологической функции, внутренний возрастный процесс потери жизнеспособности и повышения уязвимости». [ 3 ]
В 2013 году группа ученых определила девять признаков старения , которые распространены между организмами с акцентом на млекопитающих:
- геномная нестабильность ,
- истощение теломер ,
- эпигенетические изменения,
- потеря протеостаза ,
- дерегулированное зондирование питательных веществ ,
- митохондриальная дисфункция,
- клеточное старение,
- истощение стволовых клеток,
- Измененная межклеточная связь [ 4 ]
В обновлении Decadal были добавлены три отличительного признака, на общую сумму 12 предлагаемых признаков:
- инвалидов Макроавтофагия
- Хроническое воспаление
- Дисбиоз [ 5 ]
Окружающая среда вызывает повреждение на различных уровнях, например, повреждение ДНК и повреждение тканей и клеток кислородными радикалами (широко известными как свободные радикалы ), и часть этого повреждения не восстанавливается и, таким образом, накапливается со временем. [ 6 ] Клонирование из соматических клеток, а не зародышевых клеток, может начать жизнь с более высокой начальной нагрузки повреждения. Долли Овцы умерли молодым от заразного заболевания легких, но данные о всей популяции клонированных людей будут необходимы для измерения показателей смертности и количественной оценки старения. [ Цитация необходима ]
Эволюционный теоретик Джордж Уильямс написал: «Примечательно, что после, казалось бы, чудесного подвига морфогенеза , сложный метазоан должен быть неспособен выполнить гораздо более простую задачу - просто поддерживать то, что уже сформировано». [ 7 ]
Различия среди видов
[ редактировать ]Различные скорости, с которой смертность увеличивается с возрастом, соответствуют различной максимальной продолжительности жизни среди видов . Например, мышь пожилой в 3 года, человек - пожилой в 80 лет, [ 8 ] И Деревья Гинкго показывают небольшое влияние возраста даже в 667 лет. [ 9 ]
Почти все организмы Senesce, в том числе бактерии , которые имеют асимметрию между «матерью» и «дочерними» клетками при делении клеток , при этом материнская клетка испытывает старение, в то время как дочь омолажена. [ 10 ] [ 11 ] В некоторых группах наблюдается незначительное старение , такие как род Hydra . [ 12 ] Планарийные плоские черви «по -видимому, безграничная регенеративная способность теломер , способствующую популяции высокопролиферативных стволовых клеток взрослых ». [ 13 ] Эти планарии не являются биологически бессмертными , а скорее их смерть медленно увеличивается с возрастом. Организмы, которые, как считается, являются биологически бессмертными, в одном случае станут турритопсисом Dohrnii , также известным как «бессмертная медуза», из -за его способности вернуться к своей молодежи, когда он подвергается стрессу во время взросления. [ 14 ] остается репродуктивная система Наблюдается, что турритопсиса . гонады нетронутой, и даже существуют [ 15 ]
Некоторые виды демонстрируют «негативное старение», в котором способность размножения увеличивается или является стабильной, а смертность падает с возрастом, в результате чего преимущества увеличения размера тела во время старения. [ 16 ]
Теории старения
[ редактировать ]Этот раздел нуждается в расширении . Вы можете помочь, добавив к этому . ( Март 2023 г. ) |
Более 300 различных теорий были установлены для объяснения природы (механизмов) и причин (причин природного появления или факторов) старения. [ 17 ] [ Дополнительные цитаты (ы) необходимы ] Хорошие теории объяснят прошлые наблюдения и предсказывают результаты будущих экспериментов. Некоторые из теорий могут дополнять друг друга, перекрывать, противоречить или не исключать различные другие теории. [ Цитация необходима ]
Теории старения делятся на две широкие категории, эволюционные теории старения и механистические теории старения. Эволюционные теории старения в первую очередь объясняют, почему происходит старение, [ 18 ] но не заботитесь о молекулярном механизме (и), который управляет процессом. Все эволюционные теории старения опираются на основные механизмы, которые сила естественного отбора снижается с возрастом. [ 19 ] [ 20 ] Механистические теории старения можно разделить на теории, которые предлагают старения, запрограммированы, и теории накопления повреждений, то есть те, которые предлагают, чтобы старение было вызвано конкретными молекулярными изменениями, происходящими с течением времени.
Эволюционные теории старения
[ редактировать ]Антагонистическая плейотропия
[ редактировать ]Одна теория была предложена Джорджем С. Уильямсом [ 7 ] и включает антагонистическую плейотропию . Один ген может повлиять на множество признаков. Некоторые черты, которые увеличивают физическую форму в раннем возрасте, также могут иметь негативные последствия в дальнейшей жизни. Но поскольку гораздо больше людей живы в молодых возрастах, чем в старых возрастах, даже небольшие положительные эффекты могут быть сильно выбраны, а большие негативные последствия позже могут быть очень слабо отобраны. Уильямс предложил следующий пример: возможно, коды генов для осаждения кальция в костях, что способствует выживанию несовершеннолетних и, следовательно, будет пользоваться естественным отбором; Однако этот же ген способствует осаждению кальция в артериях, вызывая негативные атеросклеротические эффекты в старости. Таким образом, вредные биологические изменения в старости могут быть результатом отбора для плейотропных генов, которые полезны в начале жизни, но впоследствии вредны. В этом случае давление отбора относительно высокое, когда репродуктивная ценность Фишера высока и относительно низкая, когда репродуктивная ценность Фишера низкая.
Рак против клеточного стажира теория старения
[ редактировать ]Пожилые клетки в многоклеточном организме могут быть очищены от конкуренции между клетками, но это увеличивает риск развития рака. Это приводит к неизбежной дилемме между двумя возможностями - накоплением физиологически бесполезных стареющих клеток и раком - обе из которых приводят к увеличению показателей смертности с возрастом. [ 2 ]
Одноразовая сома
[ редактировать ]Одноразовая теория старения сома была предложена Томасом Кирквудом в 1977 году. [ 1 ] [ 21 ] Теория предполагает, что старение происходит из -за стратегии, в которой человек инвестирует только в поддержание сомы до тех пор, пока у него есть реалистичный шанс на выживание. [ 22 ] Вид, который более эффективно использует ресурсы, будет жить дольше и, следовательно, сможет передать генетическую информацию следующему поколению. Требования воспроизводства высоки, поэтому меньше усилий инвестируется в восстановление и поддержание соматических клеток по сравнению с клетками зародышевой линии , чтобы сосредоточиться на размножении и выживании видов. [ 23 ]
Запрограммированные теории старения
[ редактировать ]Программированные теории старения предполагают, что старение является адаптивным, обычно вызывает выбор для эволюции или отбора группы .
Теория цикла репродуктивных клеток предполагает, что старение регулируется изменениями гормональной передачи сигналов в течение продолжительности жизни. [ 24 ]
Теории накопления ущерба
[ редактировать ]Теория свободного радикала старения
[ редактировать ]Одна из самых выдающихся теорий старения была впервые предложена Харманом в 1956 году. [ 25 ] Это утверждает, что свободные радикалы, продуцируемые растворенным кислородом, радиацией, клеточным дыханием и другими источниками, вызывают повреждение молекулярных машин в клетке и постепенно имит их. Это также известно как окислительный стресс .
Существуют существенные доказательства, подтверждающие эту теорию. Старые животные имеют большее количество окисленных белков, ДНК и липидов, чем их более молодые коллеги. [ 26 ] [ 27 ]
Химическое повреждение
[ редактировать ]
Одной из самых ранних теорий старения была скорость живой гипотезы, описанная Рэймондом Перл в 1928 году. [ 28 ] (На основе более ранней работы Макса Рубнера ), которая утверждает, что быстрый базальный скорость метаболизма соответствует короткой максимальной продолжительности жизни .
Несмотря на то, что может быть некоторая достоверность к идее, что для различных типов конкретных повреждений, подробно описанных ниже, являются побочными продуктами метаболизма , при прочих равных условиях быстрый метаболизм может снизить продолжительность жизни, в целом эта теория не объясняет различия в Срок службы либо внутри, или между видами. Калорически ограниченные животные обрабатывают столько или более калорий на грамм массы тела, как и их ad коллеги, питающиеся [ Цитация необходима ] Точно так же скорость метаболизма является плохим предиктором продолжительности жизни для птиц, летучих мышей и других видов, которые, как предполагается, имеют снижение смертности от хищничества и, следовательно, развили длительные сроки продолжительности даже при наличии очень высоких показателей метаболизма. [ 29 ] В анализе 2007 года было показано, что, когда используются современные статистические методы для исправления эффектов размера тела и филогения , скорость метаболизма не коррелирует с долговечностью у млекопитающих или птиц. [ 30 ]
Что касается специфических типов химического повреждения, вызванного метаболизмом, предполагается, что повреждение долгоживущих биополимеров , таких как структурные белки или ДНК , вызванные повсеместными химическими агентами в организме, таких как кислород и сахар , частично ответственны за старение. Полем Повреждение может включать в себя поломку биополимерных цепей, сшивание биополимеров или химическое привязанность неестественных заместителей ( гаптентов ) к биополимерам. [ Цитация необходима ] При нормальных аэробных условиях приблизительно 4% кислорода, метаболизируемого митохондриями , в супероксид -ион, который впоследствии может быть преобразован в перекись водорода , гидроксильный радикал и, конечном превращаются в итоге Свободные радикалы, способные нанести ущерб структурным белкам и ДНК. [ 6 ] Некоторые ионы металлов , найденные в организме, такие как медь и железо , могут участвовать в процессе. (При болезни Уилсона , наследственный дефект , который заставляет организм сохранять медь, некоторые симптомы напоминают ускоренное старение.) Эти процессы, называемые окислительным стрессом, связаны с потенциальными преимуществами диетических полифенольных антиоксидантов , например, в кофе , [ 31 ] и чай . [ 32 ] Однако их обычно положительное влияние на продолжительность жизни при потреблении умеренной [ 33 ] [ 34 ] [ 35 ] также были объяснены влиянием на аутофагию , [ 36 ] метаболизм глюкозы [ 37 ] и ampk . [ 38 ]
Сахара, такие как глюкоза и фруктоза, могут реагировать с определенными аминокислотами, такими как лизин и аргинин и некоторые основания ДНК, такие как гуанин, для получения аддуктов сахара, в процессе, называемом гликированием . Эти аддукты могут дальнейшие перестройки с образованием реактивных видов, которые затем могут сшивать структурные белки или ДНК с аналогичными биополимерами или другими биомолекулами, такими как неструктурные белки. У людей с диабетом , у которых повышен уровень сахара в крови , развиваются расстройства, связанные с старением, намного раньше, чем население в целом, но могут задержать такие расстройства, строгий контроль над уровнем сахара в крови. Существуют доказательства того, что повреждение сахара связано с повреждением окислителя в процессе, называемом гликоксидированием .
Свободные радикалы могут повредить белки, липиды или ДНК . Гликирование в основном повреждает белки. Поврежденные белки и липиды накапливаются в лизосомах в виде липофусцина . Химическое повреждение структурных белков может привести к потере функции; Например, повреждение коллагена стен кровеносного сосуда может привести к жесткости сосудов и, таким образом, гипертонии , а также утолщению стенки сосуда и образованию реактивной ткани ( атеросклероз ); Подобные процессы в почке могут привести к почечной недостаточности . Повреждение ферментов снижает функциональность клеток. липидов Перекисное окисление внутренней митохондриальной мембраны уменьшает электрический потенциал и способность генерировать энергию. Вероятно, не случайно, что почти все так называемые « ускоренные болезни старения » обусловлены дефектными ферментами репарации ДНК . [ 39 ] [ 40 ]
Считается, что влияние алкоголя на старение может быть частично объяснено активацией алкоголя оси HPA , которая стимулирует секрецию глюкокортикоидов , долгосрочное воздействие, которое вызывает симптомы старения. [ 41 ]
Повреждение ДНК
[ редактировать ]Повреждение ДНК было предложено в обзоре 2021 года, чтобы стать основной причиной старения из -за механистической связи повреждения ДНК практически во всех аспектах фенотипа старения. [ 42 ] Было обнаружено, что более медленная скорость накопления повреждения ДНК , измеренная с помощью маркера повреждения ДНК гамма H2AX в лейкоцитах, коррелирует с более длительными сроками службы в сравнении с дельфинами , козами , оленями , американскими фламинго и грифонами . [ 43 ] изменения , вызванные повреждением ДНК Эпигенетические , такие как метилирование ДНК и многие модификации гистонов , по-видимому, имеют особое значение для процесса старения. [ 42 ] Доказательства теории о том, что повреждение ДНК является фундаментальной причиной старения, было впервые рассмотрено в 1981 году. [ 44 ]
Накопление мутации
[ редактировать ]Естественный отбор может поддерживать смертельные и вредные аллели , если их последствия ощущаются после воспроизведения. Генетик JBS Haldane задавался вопросом, почему доминирующая мутация, которая вызывает болезнь Хантингтона, осталась в популяции, и почему естественный отбор не исключил его. Начало этого неврологического заболевания составляет (в среднем) в возрасте 45 лет и неизменно смертельно фатально в течение 10–20 лет. Халдейн предположил, что в предыстории человека лишь немногие выжили до 45 лет. Поскольку немногие были живы в старшем возрасте, и их вклад в следующее поколение был небольшим по сравнению с большими когортами более молодых возрастных групп, сила отбора против такого позднего действия вредные мутации были соответственно небольшими. Следовательно, генетическая нагрузка вредных мутаций поздних действий может быть существенной при балансе мутации-выбора . Эта концепция стала известна как Selection Shadow . [ 45 ]
Питер Медавар формализовал это наблюдение в своей теории накопления мутации старения. [ 46 ] [ 47 ] «Сила естественного отбора ослабляется с возрастом - даже в теоретически бессмертной популяции, при условии, что она подвергается воздействию реальных опасностей смертности. Если генетическая катастрофа ... происходит достаточно поздно в индивидуальной жизни, его последствия могут быть совершенно не важны ". Возраст-независимые опасности, такие как хищничество, болезнь и несчастные случаи, называемые « внешней смертностью », означают, что даже население с незначительным старением будет иметь меньше людей, живых в старших возрастных группах.
Другой урон
[ редактировать ]Исследование пришло к выводу, что ретровирусы в человеческих геномах могут разбудиться из бездействующих состояний и способствовать старению, которое может быть заблокировано нейтрализующими антителами , облегчая «клеточное старение и дегенерацию ткани и, в некоторой степени, организмальное старение». [ 48 ]
Теории стволовых клеток старения
[ редактировать ]Теория старения стволовых клеток постулирует, что процесс старения является результатом неспособности различных типов стволовых клеток продолжать воспроизвести ткани организма способных с помощью функциональных дифференцированных клеток, поддерживать оригинальную функцию ткани (или органа ). Повреждение и накопление ошибок в генетическом материале всегда являются проблемой для систем независимо от возраста. Количество стволовых клеток у молодых людей намного выше, чем пожилые люди, и, таким образом, создает лучший и более эффективный механизм замены у молодых, противоречащих старому. Другими словами, старение не является вопросом увеличения ущерба, а вопрос о неспособности заменить его из -за уменьшения числа стволовых клеток. Стволовые клетки уменьшаются в количестве и имеют тенденцию терять способность дифференцироваться в потомства или лимфоидные линии и миелоидные линии.
Поддержание динамического баланса пулов стволовых клеток требует нескольких условий. Сбалансирование пролиферации и покоящегося наряду с самонаводом ( см. Нишу ) и самообновлением гематопоэтических стволовых клеток предпочитают элементы поддержания пула стволовых клеток, в то время как дифференциация, мобилизация и старение являются вредными элементами. Эти вредные эффекты в конечном итоге вызовут апоптоз .
Есть также несколько проблем, когда речь идет о терапевтическом использовании стволовых клеток и их способности пополнять органы и ткани. Во-первых, разные клетки могут иметь разные продолжительности жизни, даже если они происходят из одних и тех же стволовых клеток ( см. Т-клетки и эритроциты ), что означает, что старение может происходить по-разному в клетках, которые имеют более длительные сроки, в отличие от тех, у которых более короткие срок службы. Кроме того, постоянные усилия по замене соматических клеток могут вызвать истощение стволовых клеток. [ 49 ]- Повышение гематопоэтического стволовых клеток
- Повышение разнообразия гематопоэтических стволовых клеток старение
- Гематопоэтическая потеря мозаики y хромосомы Y
Биомаркеры старения
[ редактировать ]могут быть лучше предсказаны Если разные люди стареют в разных показателях, то биомаркеры в хронологическом возрасте. [ 58 ] [ 59 ] Однако седые волос , [ 60 ] Старение лица , кожные морщины и другие общие изменения, наблюдаемые при старении, не являются лучшими показателями будущей функциональности, чем хронологический возраст. Биогеронтологи продолжали усилия по поиску и проверке биомаркеров старения, но успех до сих пор был ограничен.
Уровни CD4 и CD8 T-клеток памяти и наивных Т-клеток использовались для получения хороших прогнозов ожидаемой продолжительности жизни мышей среднего возраста. [ 61 ]
Старение часов
[ редактировать ]Есть интерес к эпигенетическим часам как биомаркеру старения, основанный на его способности предсказывать хронологический возраст человека. [ 62 ] Основная биохимия крови и количество клеток также может использоваться для точного прогнозирования хронологического возраста. [ 63 ] Также возможно предсказать хронологический возраст человека с использованием транскриптомных часов старения. [ 64 ]
Существует исследование и разработка дальнейших биомаркеров, систем обнаружения и программных систем для измерения биологического возраста различных тканей или систем или в целом. Например, программное обеспечение глубокого обучения (DL) с использованием анатомических магнитно -резонансных изображений оценило возраст мозга с относительно высокой точностью, включая обнаружение ранних признаков болезни Альцгеймера и различные нейроанатомические паттерны неврологического старения, [ 65 ] человека и сообщалось о инструменте DL, чтобы рассчитать воспалительный возраст на основе моделей системного возрастного воспаления. [ 66 ]
Стареющие часы были использованы для оценки воздействия вмешательств на людей, включая комбинированную терапию . [ 67 ] [ Дополнительные цитаты (ы) необходимы ] Бывшие работы по старению для выявления и оценки вмешательств долголетия представляют собой основную цель в исследованиях биологии старения. Тем не менее, достижение этой цели требует преодоления многочисленных проблем и выполнения дополнительных шагов проверки. [ 68 ] [ 69 ]
Генетические детерминанты старения
[ редактировать ]Ряд генетических компонентов старения был идентифицирован с использованием модельных организмов, начиная от простых почкуенных дрожжей Saccharomyces cerevisiae до червей, таких как Caenorhabditis elegans и плодовые мухи ( Drosophila melanogaster ). Изучение этих организмов выявило наличие как минимум двух консервативных путей старения.
Экспрессия генов несовершенно контролируется, и вполне возможно, что случайные флуктуации в уровнях экспрессии многих генов способствуют процессу старения, как это было предложено в исследовании таких генов у дрожжей. [ 70 ] Отдельные клетки, которые являются генетически идентичными, тем не менее, могут иметь существенно разные реакции на внешние стимулы и заметно различные продолжительности жизни, что указывает на то, что эпигенетические факторы играют важную роль в экспрессии и старении генов , а также генетических факторах. Есть исследования эпигенетики старения .
Способность восстанавливать ДНК-разрывы снижается при старении у мышей [ 71 ] и люди. [ 72 ]
Набор редких наследственных ( генетических ) расстройств, каждый из которых называется прогерией , был известен в течение некоторого времени. У больных проявляются симптомы, напоминающие ускоренное старение , включая морщинистую кожу . Причина синдрома прогнома Хатчинсона -Гилфорда была зарегистрирована в журнале Nature в мае 2003 года. [ 73 ] Этот отчет предполагает, что повреждение ДНК , а не окислительное стресс , является причиной этой формы ускоренного старения.
Исследование показывает, что старение может сдвинуть активность в сторону коротких генов или более короткой длины транскрипта и что это может противостоять вмешательствам. [ 74 ]
Healthspans и старение в обществе
[ редактировать ]

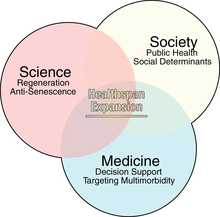
HealthSpan в целом можно определить как период жизни, который является здоровым , например, без значительных заболеваний [ 76 ] или снижение способностей (например, чувств, мышц , выносливости и познания ).
Биологическое старение или LHG поставляется с большим бременем для общества, включая потенциально растущие расходы на здравоохранение (также в зависимости от типов и затрат на лечение ). [ 75 ] [ 79 ] Это, наряду с глобальным качеством жизни или благополучием , подчеркивает важность расширения здоровья. [ 75 ]
Многие меры, которые могут продлевать продолжительность жизни, могут одновременно расширять здоровье, хотя и не обязательно так, что указывает на то, что «продолжительность жизни больше не может быть единственным параметром интереса» в связанных исследованиях. [ 80 ] В то время как недавнее увеличение ожидаемой продолжительности жизни не сопровождалось «параллельным» расширением здоровья. [ 75 ] Осведомленность о концепции и вопросах отставания HealthSpan с 2017 года. [ 76 ] Ученые отметили, что « [C] Гронические заболевания старения растут и наносят невыразимые затраты на качество жизни человека». [ 79 ]
Вмешательства
[ редактировать ]Расширение жизни -это концепция продления жизни человека , либо скромно, либо через улучшения медицины, либо резко за счет увеличения максимальной продолжительности жизни за пределами его общепризнанного биологического предела в размере около 125 лет . [ 81 ] Несколько исследователей в этом районе, наряду с «жизненными экспертистами», « бессмертными » или « долгосветными » (те, кто хочет сами достичь более длительной жизни), постулируют, что будущие прорывы в омоложении ткани , стволовых клетках , регенеративной медицине , молекулярном восстановлении, геном . Терапия , фармацевтические препараты и замена органов (например, с искусственными органами или ксенотрансплантациями ) в конечном итоге позволит людям иметь неопределенную продолжительность жизни путем полного омоложения здоровым молодому состоянию (агеразия [ 82 ] ) Этические последствия, если продление жизни становится возможным, обсуждаются биоэтиками .
Продажа предполагаемых антивозрастных продуктов, таких как добавки и замена гормонов, является прибыльной глобальной промышленностью. Например, отрасль, которая способствует использованию гормонов в качестве лечения для потребителей, чтобы замедлить или обратить вспять процесс старения на рынке США, принесла около 50 миллиардов долларов дохода в год в 2009 году. [ 83 ] Использование таких гормональных продуктов не было доказано, что является эффективным или безопасным. [ 83 ] [ 84 ] [ 85 ] [ 86 ]Смотрите также
[ редактировать ]- Антивозрастное движение
- Антимускариника
- Деменция
- Репарация ДНК
- Гериатрия
- Геронтология
- Гомеостатическая способность
- Бессмертие
- Индекс тем, связанных с продлением жизни
- Митоорезис
- Старость
- Феноптоз
- Старение растения
- Запрограммированная гибель клеток
- Стратегии для инженерного незначительного старения (Sens)
- Сублетальный урон
- Трансгенерационный дизайн
- Временная шкала исследований старения
Ссылки
[ редактировать ]- ^ Jump up to: а беременный Кирквуд Т.Б. (1977). «Эволюция старения» . Природа . 270 (5635): 301–4. Bibcode : 1977natur.270..301k . doi : 10.1038/270301A0 . PMID 593350 . S2CID 492012 . Архивировано из оригинала 15 ноября 2023 года . Получено 31 января 2022 года .
- ^ Jump up to: а беременный Нельсон П., Масел Дж (декабрь 2017 г.). «Межклеточная конкуренция и неизбежность многоклеточного старения» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (49): 12982–7. Bibcode : 2017pnas..11412982N . doi : 10.1073/pnas.16188544114 . PMC 5724245 . PMID 29087299 .
- ^ «Старение и геронтология Глоссарий» . Архивировано из оригинала 19 октября 2019 года . Получено 26 февраля 2011 года .
- ^ López-Otín C, Blasco MA, Partridge L, et al. (Июнь 2013 г.). «Признаки старения» . Клетка . 153 (6): 1194–217. doi : 10.1016/j.cell.2013.05.039 . PMC 3836174 . PMID 23746838 .
- ^ Parridge L, et. (19 января 2023 г.). Расширяющаяся вселенная " Смеситель 186 (2): 243–2 doi : 10.1016/j.cell . PMID 3699349 . S2CID 25394876 .
- ^ Jump up to: а беременный Холмс Г.Е., Бернштейн С., Бернштейн Х (сентябрь 1992). «Окислительный и другие повреждения ДНК в качестве основы старения: обзор». Мутат. Резерв 275 (3–6): 305–15. doi : 10.1016/0921-8734 (92) 90034-м . PMID 1383772 .
- ^ Jump up to: а беременный Уильямс GC (1957). «Плейотропия, естественный отбор и эволюция старения». Эволюция 11 (4): 398–411. doi : 10.2307/2406060 . JSTOR 2406060 .
- ^ Остад С.Н. (февраль 2009 г.). «Сравнительная биология старения» . Журналы геронтологии. Серия A, биологические науки и медицинские науки . 64 (2): 199–201. doi : 10.1093/gerona/gln060 . PMC 2655036 . PMID 19223603 .
- ^ Wang L, Cui J, Jin B, et al. (Январь 2020 г.). «Анализ мультифейтура сосудистых камбиальных клеток показывает механизмы долголетия в старых билоба -деревьях Гинкго » . Труды Национальной академии наук Соединенных Штатов Америки . 117 (4): 2201–10. Bibcode : 2020pnas..117.2201W . doi : 10.1073/pnas.1916548117 . PMC 6995005 . PMID 31932448 .
- ^ Ackermann M, Stearns SC, Jenal U (июнь 2003 г.). «Старение в бактерии с асимметричным делением». Наука . 300 (5627): 1920. doi : 10.1126/science.1083532 . PMID 12817142 . S2CID 34770745 .
- ^ Стюарт Э.Дж., Мэдден Р., Пол Г. и др. (Февраль 2005 г.). «Старение и смерть в организме, который воспроизводит морфологически симметричное разделение» . PLOS Биология . 3 (2): E45. doi : 10.1371/journal.pbio.0030045 . PMC 546039 . PMID 15685293 .
- ^ Данко М.Дж., Козловски Дж., Шайбл Р (октябрь 2015 г.). «Разрушение явления не высшего звезда в Гидре» . Журнал теоретической биологии . 382 : 137–49. Bibcode : 2015jthbi.382..137d . doi : 10.1016/j.jtbi.2015.06.043 . PMID 26163368 .
- ^ Tan TC, Rahman R, Jaber-Hijazi F, et al. (Март 2012 г.). «Поддержание теломер и активность теломеразы дифференциально регулируется в бесполых и сексуальных червях» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (11): 4209–14. BIBCODE : 2012PNAS..109.4209T . doi : 10.1073/pnas.1118885109 . PMC 3306686 . PMID 22371573 .
- ^ Лизенкова А.А., Григоренко А.П., Тяжелова ТВ и др. (Февраль 2017 г.). «Полный митохондриальный геном и эволюционный анализ турритопсиса Dohrnii,« бессмертной »медузы с обратимым жизненным циклом» . Молекулярная филогенетика и эволюция . 107 : 232–8. Bibcode : 2017molpe.107..232L . doi : 10.1016/j.ympev.2016.11.007 . PMID 27845203 .
- ^ Piraino S, Boero F, Aeschbach B, et al. (Июнь 1996 г.). «Изменение жизненного цикла: преобразование медуз в полипы и трансдифференцировку клеток в турритопсисе nutriculaula (Cnidaria, гидрозоа)». Биологический бюллетень . 190 (3): 302–312. doi : 10.2307/1543022 . JSTOR 1543022 . PMID 29227703 .
- ^ Vaupel JW, Baudisch A, Dölling M, et al. (Июнь 2004 г.). «Случай негативного старения». Теоретическая биология населения . 65 (4): 339–51. Bibcode : 2004tpbio..65..339W . doi : 10.1016/j.tpb.2003.12.003 . PMID 15136009 .
- ^ Vineyard J, Drugs C, Michael J (2007). "Теории старения " iubmb Жизнь 59 (4–5): 249–54. два 10.1080/15216540601178067: 17505961PMID
- ^ Кирквуд Т.Б., Остад С.Н. (2000). "Почему мы стареем?" Полем Природа . 408 (6809): 233–8. Bibcode : 2000natur.408..233K . doi : 10.1038/35041682 . PMID 11089980 . S2CID 2579770 . Архивировано из оригинала 15 ноября 2023 года . Получено 31 января 2022 года .
- ^ Medawar PB (1952). Нерешенная проблема биологии . Опубликовано для колледжа HK Lewis . OCLC 869293719 .
- ^ Роуз М.Р. (1991). Эволюционная биология старения . Издательство Оксфордского университета. ISBN 1-4237-6520-6 Полем OCLC 228167629 . Архивировано из оригинала 15 ноября 2023 года . Получено 31 января 2022 года .
- ^ Кирквуд Т. (2006). Время нашей жизни: наука о старении человека . Издательство Оксфордского университета. ISBN 978-0-19-802939-7 Полем OCLC 437175125 . Архивировано из оригинала 15 ноября 2023 года . Получено 31 января 2022 года .
- ^ Hammers M, Richardson DS, Burke T, et al. (Сентябрь 2013 г.). «Влияние репродуктивных инвестиций и условий экологических условий в раннем возрасте на старение: поддержка одноразовой гипотезы сома». Журнал эволюционной биологии . 26 (9): 1999–2007. doi : 10.1111/jeb.12204 . HDL : 11370/9CC6749C-F67D-40AB-A253-A06650C32102 . PMID 23961923 . S2CID 46466320 .
- ^ Kirkwood TB, Rose MR (апрель 1991 г.). «Эволюция старения: позднее выживание принесло в жертву ради размножения». Философские транзакции Королевского общества Лондона. Серия B, биологические науки . 332 (1262): 15–24. BIBCODE : 1991RSPTB.332 ... 15K . doi : 10.1098/rstb.1991.0028 . PMID 1677205 .
- ^ Atwood CS, Bowen RL (2011). «Теория старения репродуктивного клеток: обновление». Экспериментальная геронтология . 46 (2–3): 100–7. doi : 10.1016/j.exger.2010.09.007 . PMID 20851172 . S2CID 20998909 .
- ^ Харман Д. (июль 1956 г.). «Старение: теория, основанная на свободной радикальной и радиационной химии». Журнал геронтологии . 11 (3): 298–300. doi : 10.1093/geronj/11.3.298 . HDL : 2027/MDP.39015086547422 . PMID 13332224 .
- ^ Stadtman ER (август 1992). «Окисление белка и старение» . Наука . 257 (5074): 1220–4. Bibcode : 1992sci ... 257.1220S . doi : 10.1126/science.1355616 . PMID 1355616 . Архивировано из оригинала 31 июля 2021 года . Получено 21 июля 2021 года .
- ^ Sohal RS, Agarwal S, Dubey A, et al. (Август 1993). «Окислительное повреждение белка связано с продолжительностью домохозяйств» . Труды Национальной академии наук Соединенных Штатов Америки . 90 (15): 7255–9. Bibcode : 1993pnas ... 90,7255S . doi : 10.1073/pnas.90.15.7255 . PMC 47115 . PMID 8346242 .
- ^ Жемчужина (1928). Уровень жизни, являющийся отчетом о некоторых экспериментальных исследованиях биологии продолжительности жизни . Нью -Йорк: Альфред А. Кнопф. [ страница необходима ]
- ^ Brunet-Rossinni AK, Austad SN (2004). «Старение исследований летучих мышей: обзор». Биогеронтология . 5 (4): 211–22. doi : 10.1023/b: bgen.0000038022.65024.d8 . PMID 15314271 . S2CID 227558811 .
- ^ De Magalháes JP, Costa J, Church GM (февраль 2007 г.). «Анализ взаимосвязи между метаболизмом, графиками развития и долговечностью с использованием филогенетических независимых контрастов» . Журналы геронтологии. Серия A, биологические науки и медицинские науки . 62 (2): 149–60. Citeseerx 10.1.1.596.2815 . doi : 10.1093/gerona/62.2.149 . PMC 2288695 . PMID 17339640 .
- ^ Freedman ND, Park Y, Abnet CC, et al. (Май 2012 г.). «Ассоциация питья кофе с общей и конкретной смертностью» . Новая Англия Журнал медицины . 366 (20): 1891–904. doi : 10.1056/nejmoa1112010 . PMC 3439152 . PMID 22591295 .
- ^ Ян Ю, Чан С.В., Ху М. и др. (2011). «Влияние некоторых общих компонентов пищи на сердечно -сосудистые заболевания» . ISRN Cardiology . 2011 : 397136. DOI : 10.5402/2011/397136 . PMC 3262529 . PMID 22347642 .
- ^ Пул Р., Кеннеди О.Дж., Родерик П. и др. (22 ноября 2017 г.). «Потребление кофе и здоровье: Обзор обзора метаанализов множества результатов в отношении здоровья» . BMJ . 359 : J5024. doi : 10.1136/bmj.j5024 . PMC 5696634 . PMID 29167102 .
- ^ О'Киф Дж. Х., Диниколантонио Дж.Дж., Лави К.Дж. (1 мая 2018 года). «Кофе для кардиозащиты и долговечности». Прогресс в сердечно -сосудистых заболеваниях . 61 (1): 38–42. doi : 10.1016/j.pcad.2018.02.002 . PMID 29474816 .
- ^ Grosso G, Godos J, Galvano F, et al. (21 августа 2017 г.). «Кофе, кофеин и результаты в отношении здоровья: обзор зонтика». Ежегодный обзор питания . 37 (1): 131–156. doi : 10.1146/annurev-nutr-071816-064941 . PMID 28826374 .
- ^ Dirks-Naylor AJ (15 декабря 2015 г.). «Преимущества кофе на скелетных мышцах». Жизненные науки . 143 : 182–6. doi : 10.1016/j.lfs.2015.11.005 . PMID 26546720 .
- ^ Reis Ce, Dórea JG, Da Costa Th (1 июля 2019 г.). «Влияние потребления кофе на метаболизм глюкозы: систематический обзор клинических испытаний» . Журнал традиционной и дополнительной медицины . 9 (3): 184–191. doi : 10.1016/j.jtcme.2018.01.001 . PMC 6544578 . PMID 31193893 .
- ^ Loureiro LM, Reis Ce, Costa Th (1 мая 2018 года). «Влияние кофейных компонентов на восстановление мышечного гликогена: систематический обзор». Международный журнал спортивного питания и метаболизма физических упражнений . 28 (3): 284–293. doi : 10.1123/ijsnem.2017-0342 . PMID 29345166 .
- ^ Bernstein H, Payne CM, Bernstein C, et al. (2008). «Рак и старение как последствия не поврежденного повреждения ДНК». Полем В Kimura H, Suzuki A (ред.). Новое исследование повреждения ДНК . Nova Science Publishers. С. 1–47. ISBN 978-1604565812 Полем Архивировано из оригинала 15 ноября 2023 года . Получено 4 февраля 2016 года .
- ^ Pan MR, Li K, Lin Sy, et al. (Май 2016 г.). «Соединение точек: от повреждения ДНК и восстановления до старения» . Международный журнал молекулярных наук . 17 (5): 685. doi : 10.3390/ijms17050685 . PMC 4881511 . PMID 27164092 .
- ^ Спенсер Р.Л., Хатчисон К.Е. (1999). «Алкоголь, старение и реакция на стресс» (PDF) . Исследование алкоголя и здоровье . 23 (4): 272–83. PMC 6760387 . PMID 10890824 . Архивировано из оригинала (PDF) 11 декабря 2018 года . Получено 8 апреля 2008 года .
- ^ Jump up to: а беременный Schumacher B, Pothof J, Vijg J, et al. (Апрель 2021 г.). «Центральная роль повреждения ДНК в процессе старения» . Природа . 592 (7856): 695–703. Bibcode : 2021natur.592..695s . doi : 10.1038/s41586-021-03307-7 . PMC 9844150 . PMID 33911272 .
- ^ Whittemore K, Martínez-Nevado E, Blasco MA (ноябрь 2019). «Более медленные скорости накопления повреждения ДНК в лейкоцитах коррелируют с более длительным сроком службы у нескольких видов птиц и млекопитающих» . Старение (Олбани Нью -Йорк) . 11 (21): 9829–45. doi : 10.18632/старение.102430 . PMC 6874430 . PMID 31730540 .
- ^ Генслер Х.Л., Бернштейн Х (сентябрь 1981 г.). «Повреждение ДНК как основная причина старения» . Ежеквартальный обзор биологии . 56 (3): 279–303. doi : 10.1086/412317 . PMID 7031747 . S2CID 20822805 .
- ^ Фабиан Д., Флэтт Т (2011). «Эволюция старения» (PDF) . Природное образование .
- ^ Medawar PB (1946). «Старость и естественная смерть». Современный квартал . 1 : 30–56.
- ^ Медавар 1952 [ страница необходима ]
- ^ Liu X, Liu Z, Wu Z, et al. (19 января 2023 г.). «Воскресение эндогенных ретровирусов во время старения усиливает старение» . Клетка . 186 (2): 287–304.E26. doi : 10.1016/j.cell.2022.12.017 . PMID 36610399 . S2CID 232060038 .
- Экспертное объяснение исследования: «Старение и ретровирусы» . Наука . Архивировано из оригинала 17 февраля 2023 года . Получено 17 февраля 2023 года .
- ^ Смит Дж., А., Даниэль Р. «Стволовые клетки и старение: проблема курицы или яйца?». Старение и болезнь . 2012 Jun, Vol. 3, номер 3; 260–268.
- ^ Махла Р.С. (2016). «Стволовые клетки применение в регенеративной медицине и тройке болезни» . Международный журнал клеточной биологии . 2016 (7): 19. doi : 10.1155/2016/6940283 . PMC 4969512 . PMID 27516776 .
- ^ Rossi DJ, Bryder D, Seita J, et al. (2007). «Недостатки в восстановлении повреждения ДНК ограничивают функцию гематопоэтических стволовых клеток с возрастом». Природа . 447 (7145): 725–9. Bibcode : 2007natur.447..725r . doi : 10.1038/nature05862 . PMID 17554309 . S2CID 4416445 .
- ^ Jump up to: а беременный Beerman I, Seita J, Inlay MA, et al. (2014). «Огромные гематопоэтические стволовые клетки накапливают повреждение ДНК во время старения, которое восстанавливается при входе в клеточный цикл» . Клеточная стволовая клетка . 15 (1): 37–50. doi : 10.1016/j.stem.2014.04.016 . PMC 4082747 . PMID 24813857 .
- ^ Jump up to: а беременный Nijnik A, Woodbine L, Marchetti C, et al. (2007). «Репарация ДНК ограничивает гематопоэтические стволовые клетки во время старения». Природа . 447 (7145): 686–90. Bibcode : 2007natur.447..686n . doi : 10.1038/nature05875 . PMID 17554302 . S2CID 4332976 .
- ^ «Исследования могут показать, почему люди могут внезапно стать слабыми в возрасте 70 лет» . Хранитель . 1 июня 2022 года . Получено 18 июля 2022 года .
- ^ Mitchell E, Spencer Chapman M, Williams N, et al. (Июнь 2022 г.). «Клональная динамика гематопоэза на протяжении всей жизни» . Природа . 606 (7913): 343–350. Bibcode : 2022nater.606..343M . doi : 10.1038/s41586-022-04786-y . ISSN 1476-4687 . PMC 9177428 . PMID 35650442 .
- ^ Колата Г (14 июля 2022 года). «По мере того, как Y -хромосомы исчезают с возрастом, риски сердца могут расти» . New York Times . Получено 21 августа 2022 года .
- ^ Sano S, Horitani K, Ogawa H, et al. (15 июля 2022 г.). «Гемопоэтическая потеря Y -хромосомы приводит к фиброзу сердца и смертности от сердечной недостаточности» . Наука . 377 (6603): 292–297. Bibcode : 2022sci ... 377..292S . doi : 10.1126/science.abn3100 . ISSN 0036-8075 . PMC 9437978 . PMID 35857592 .
- ^ Gasmi A, Chirumbolo S, Peana M, et al. (17 сентября 2020 г.). «Биомаркеры старения во время старения в качестве возможных предупреждений для использования профилактических мер». Текущая лекарственная химия . 28 (8): 1471–88. doi : 10.2174/0929867327999200917150652 . PMID 32942969 . S2CID 221789280 .
- ^ Бейкер Г.Т., Спротт Р.Л. (1988). «Биомаркеры старения» . Экспериментальная геронтология . 23 (4–5): 223–39. doi : 10.1016/0531-5565 (88) 90025-3 . PMID 3058488 . S2CID 31039588 . Архивировано из оригинала 24 октября 2021 года . Получено 12 июля 2019 года .
- ^ Van Neste D, Tobin DJ (2004). «Цикл волос и пигментация волос: динамические взаимодействия и изменения, связанные со старением». Микрон . 35 (3): 193–200. doi : 10.1016/j.micron.2003.11.006 . PMID 15036274 .
- ^ Миллер Р.А. (апрель 2001 г.). «Биомаркеры старения: прогнозирование долговечности с использованием чувствительных к возрасту определений Т-клеток в генно-гетерогенной популяции мышей среднего возраста» . Журналы геронтологии. Серия A, биологические науки и медицинские науки . 56 (4): B180-6. doi : 10.1093/gerona/56.4.b180 . PMC 7537444 . PMID 11283189 .
- ^ Naue J (октябрь 2023 г.). «Вывод хронологического возраста от ДНК: использование понимания возрастного метилирования ДНК для применения судебной ДНК» . Гены и геномика . 45 (10): 1239–1261. doi : 10.1007/s13258-023-01392-8 . PMC 10504122 . PMID 37253906 .
- ^ Путин Е., Мамошина П., Алипер А. и соавт. (Май 2016 г.). «Глубокие биомаркеры старения человека: применение глубоких нейронных сетей к развитию биомаркеров» . Старение . 8 (5): 1021–33. doi : 10.18632/старение.100968 . PMC 4931851 . PMID 27191382 .
- ^ Peters MJ, Joehanes R, Pilling LC, et al. (Октябрь 2015). «Транскрипционный ландшафт возраста в периферической крови человека» . Природная связь . 6 : 8570. Bibcode : 2015natco ... 6.8570. Полем doi : 10.1038/ncomms9570 . PMC 4639797 . PMID 26490707 .
- ^ Инь С, Имс П., Ченг М. и др. (10 января 2023 г.). «Анатомически интерпретируемое глубокое обучение мозгового возраста отражает домен-специфическую когнитивную нарушение» . Труды Национальной академии наук . 120 (2): E2214634120. Bibcode : 2023pnas..12014634Y . doi : 10.1073/pnas.2214634120 . PMC 9926270 . PMID 36595679 .
- Пресс -релиз университета: «Сколько лет вашему мозгу, правда? Анализ с AI, точно отражает риск снижения когнитивных когнитивных средств и болезни Альцгеймера» . Университет Южной Калифорнии через Medicalxpress.com . Архивировано из оригинала 17 февраля 2023 года . Получено 17 февраля 2023 года .
- Новостная статья об исследовании: «ИИ может определить истинный возраст мозга» . Deutschlandfunk Nova (на немецком языке). Архивировано из оригинала 17 февраля 2023 года . Получено 17 февраля 2023 года .
- ^ Sayed N, Huang Y, Nguyen K, et al. (Июль 2021 г.). «Воспалительные часы старения (IAGE), основанные на глубоких отслеживаниях обучения, мультиморбидность, иммунозистенную, слабость и сердечно -сосудистые старения» . Старение природы . 1 (7): 598–615. doi : 10.1038/s43587-021-00082-y . PMC 8654267 . PMID 34888528 .
- Новостная статья об исследовании: «Инструмент, который рассчитывает возраст иммунной системы, может предсказать хрупкость и болезнь» . Новый Атлас . 13 июля 2021 года. Архивировано с оригинала 26 июля 2021 года . Получено 26 июля 2021 года .
- ^ Fitzgerald KN, Hodges R, Hanes D, et al. (2021). «Потенциальное изменение эпигенетического возраста с использованием диеты и вмешательства в образ жизни: пилотное рандомизированное клиническое исследование» . Старение . 13 (7): 9419–32. doi : 10.18632/старение.202913 . PMC 8064200 . PMID 33844651 . Архивировано из оригинала 2 июня 2021 года . Получено 28 июня 2021 года .
- ^ Moqri M, Herzog C, Poganik JR, et al. (Август 2023). «Биомаркеры старения для идентификации и оценки вмешательств долголетия» . Клетка . 186 (18): 3758–75. doi : 10.1016/j.cell.2023.08.003 . PMC 11088934 . PMID 37657418 .
- ^ Moqri M, Herzog C, Poganik JR, et al. (Февраль 2024 г.). «Валидация биомаркеров старения» . Природная медицина . 30 (2): 360–372. doi : 10.1038/s41591-023-02784-9 . ISSN 1546-170x . PMC 11090477 . PMID 38355974 .
- ^ Райли Дж., Перейра-Смит Ом (2006). «Микрофлюидическое устройство для анализа экспрессии отдельных генов у Saccharomyces cerevisiae». Дрожжи . 23 (14–15): 1065–73. doi : 10.1002/да.1412 . PMID 17083143 . S2CID 31356425 .
- ^ Vaidya A, Mao Z, Tian X, et al. (Июль 2014). «Мыши с нокаутом репортеров демонстрируют, что восстановление ДНК не гомологичным концом соединяется с снижением с возрастом» . PLOS GENET . 10 (7): E1004511. doi : 10.1371/journal.pgen.1004511 . PMC 4102425 . PMID 25033455 .
- ^ Li Z, Zhang W, Chen Y, et al. (Ноябрь 2016). «Нарушение двойной разрывы ДНК способствует возрастному повышению нестабильности геномной нестабильности у людей» . Клеточная гибель отличается . 23 (11): 1765–77. doi : 10.1038/cdd.2016.65 . PMC 5071568 . PMID 27391797 .
- ^ Mounkes LC, Kozlov S, Hernandez L, et al. (Май 2003 г.). «Синдром прогероидов у мышей вызван дефектами в ламинах A-типа» . Природа . 423 (6937): 298–301. Bibcode : 2003natur.423..298m . doi : 10.1038/nature01631 . PMID 12748643 . S2CID 4360055 . Архивировано из оригинала 30 мая 2022 года . Получено 21 июля 2021 года - через Zenodo.
- ^ Stoeger T, Grant RA, McQuattie-Pimentel AC, et al. (Декабрь 2022 г.). «Старение связано с системным дисбалансом транскриптома, ассоциированного с длиной» . Старение природы . 2 (12): 1191–1206. doi : 10.1038/s43587-022-00317-6 . PMC 10154227 . PMID 37118543 .
- Пресс -релиз университета: «Старение обусловлено несбалансированными генами, обнаруживает анализ ИИ множественных видов» . Северо -западный университет . 9 декабря 2022 года. Архивировано с оригинала 2 февраля 2023 года . Получено 18 января 2023 года - через Phys.org.
- Новостная статья об исследовании: Квон Д (6 января 2023 г.). «Старение связано с большей активностью в коротких генах, чем в длинных генах» . Scientific American . Архивировано из оригинала 17 января 2023 года . Получено 18 января 2023 года .
- ^ Jump up to: а беременный в дюймовый и фон Гармани А., Ямада С., Терзич А (23 сентября 2021 г.). «Longevity Leap: Enge the Healthspan Gap» . NPJ Регенеративная медицина . 6 (1): 57. doi : 10.1038/s41536-021-00169-5 . PMC 8460831 . PMID 34556664 .
- Некоммерческий пресс-релиз больницы: Граги S (7 октября 2021 года). «Регенеративный сброс для старения» Центра регенеративной биотерапевтической среды » . Клиника Майо . Архивировано с оригинала 1 марта 2023 года . Получено 1 марта 2023 года .
- ^ Jump up to: а беременный Петерсон Т (30 мая 2017 г.). «Healthspan важнее продолжительности жизни, так почему бы об этом не знает больше людей?» Полем Институт общественного здравоохранения . Вашингтонский университет в Сент -Луисе. Харви А. Фридман Центр старения. Архивировано с оригинала 1 марта 2023 года . Получено 1 марта 2023 года .
- ^ Гармани А., Ямада С., Терзич А (сентябрь 2021 г.). «Longevity Leap: Enge the Healthspan Gap» . NPJ Регенеративная медицина . 6 (1): 57. doi : 10.1038/s41536-021-00169-5 . PMC 8460831 . PMID 34556664 .
- ^ Фаррелли С (ноябрь 2022 г.). «Старение, равенство и здоровье человека» . HEC Forum . 36 (2): 187–205. doi : 10.1007/s10730-022-09499-3 . PMC 9644010 . PMID 36348214 .
- ^ Jump up to: а беременный Хансен М., Кеннеди Б.К. (1 августа 2016 г.). "Значит ли продолжительность продолжительности жизни дольше здоровья?" Полем Тенденции в клеточной биологии . 26 (8): 565–8. doi : 10.1016/j.tcb.2016.05.002 . PMC 4969078 . PMID 27238421 .
- ^ Бансал А., Чжу Л.Дж., Йен К. и др. (20 января 2015 г.). «Необвисывающая продолжительность жизни и здоровье у мутантов с долговечностью Caenorhabditis elegans» . Труды Национальной академии наук . 112 (3): E277-86. Bibcode : 2015pnas..112e.277b . doi : 10.1073/pnas.1412192112 . PMC 4311797 . PMID 25561524 .
- ^ Тернер Б.С. (2009). Можем ли мы жить вечно? Социологическое и моральное исследование . Гимн Пресс. п. 3
- ^ «Агеразия» . Оксфордский английский словарь (онлайн изд.). Издательство Оксфордского университета . в учреждении или (Требуется членство участвующее учреждение .)
- ^ Jump up to: а беременный Джапсен Б (15 июня 2009 г.). «AMA сообщает о вопросах науки о использовании гормонов в качестве антивозрастного лечения» . Чикаго Трибьюн . Получено 17 июля 2009 года .
- ^ Холлидей Р (апрель 2009 г.). «Чрезвычайное высокомерие антивозрастной медицины». Биогеронтология . 10 (2): 223–228. doi : 10.1007/s10522-008-9170-6 . PMID 18726707 . S2CID 764136 .
- ^ Олшанский С.Дж., Хейфлик Л., Карнс Б.А. (август 2002 г.). «Заявление о положении о старении человека» . Журналы геронтологии. Серия A, биологические науки и медицинские науки . 57 (8): B292 - B297. Citeseerx 10.1.1.541.3004 . doi : 10.1093/gerona/57.8.b292 . PMID 12145354 .
- ^ Warner H, Anderson J, Austad S, et al. (Ноябрь 2005 г.). "Наука факт и повестка дня Sens. Чего мы можем разумно ожидать от исследований по старе?" Полем Embo сообщает . 6 (11): 1006–1008. doi : 10.1038/sj.embor.7400555 . PMC 1371037 . PMID 16264422 .
{{cite journal}}: Cs1 maint: переопределенная настройка ( ссылка )