Старение
Старение ( / s ɪ ˈ n ɛ s ə n s / ) или биологическое старение — постепенное ухудшение функциональных характеристик живых организмов. Старение всего организма предполагает увеличение смертности или снижение плодовитости организма с возрастом, по крайней мере, на более позднем этапе жизненного цикла . [ 1 ] [ 2 ] Однако последствия старения могут быть отсрочены. Открытие 1934 года о том, что ограничение калорий может увеличить продолжительность жизни на 50% у крыс, существование видов, имеющих незначительное старение , и существование потенциально бессмертных организмов, таких как представители рода Hydra, побудили исследования по замедлению старения и, следовательно, возрастных заболеваний . Редкие человеческие мутации могут вызывать болезни ускоренного старения .
Факторы окружающей среды могут влиять на старение – например, чрезмерное воздействие ультрафиолетового излучения ускоряет старение кожи . Различные части тела могут стареть с разной скоростью и по-разному, включая мозг , сердечно-сосудистую систему и мышцы. Аналогичным образом, с возрастом могут заметно ухудшаться функции, включая контроль движений и память . Два организма одного и того же вида также могут стареть с разной скоростью, что делает биологическое старение и хронологическое старение разными понятиями.
Определение и характеристики
[ редактировать ]Старение организма – это старение целого организма. Актуарное старение можно определить как увеличение смертности или снижение плодовитости с возрастом. гласит Закон смертности Гомпертца-Мейкхэма , что возрастная составляющая уровня смертности увеличивается с возрастом экспоненциально.
Старение характеризуется снижением способности реагировать на стресс, увеличением гомеостатического дисбаланса и повышенным риском заболеваний, связанных со старением, включая рак и болезни сердца . Старение определяется как «прогрессирующее ухудшение физиологических функций, естественный возрастной процесс потери жизнеспособности и увеличения уязвимости». [ 3 ]
В 2013 году группа ученых определила девять признаков старения , общих для организмов, уделив особое внимание млекопитающим:
- геномная нестабильность ,
- истощение теломер ,
- эпигенетические изменения,
- потеря протеостаза ,
- дерегулированное восприятие питательных веществ ,
- митохондриальная дисфункция,
- клеточное старение,
- истощение стволовых клеток,
- измененное межклеточное общение [ 4 ]
В десятилетнем обновлении были добавлены три клейма, всего 12 предлагаемых клейм:
Окружающая среда вызывает повреждения на различных уровнях, например, повреждение ДНК и повреждение тканей и клеток кислородными радикалами (широко известными как свободные радикалы ), причем некоторые из этих повреждений не восстанавливаются и, таким образом, накапливаются со временем. [ 6 ] Клонирование из соматических клеток, а не из зародышевых клеток, может начать жизнь с более высокой начальной нагрузкой повреждений. Овца Долли умерла молодой от заразной болезни легких, но для измерения уровня смертности и количественной оценки старения потребуются данные обо всей популяции клонированных особей. [ нужна ссылка ]
Теоретик-эволюционист Джордж Уильямс писал: «Примечательно, что после, казалось бы, чудесного морфогенеза сложное многоклеточное животное оказывается неспособным выполнять гораздо более простую задачу — просто поддерживать то, что уже сформировалось». [ 7 ]
Различия между видами
[ редактировать ]Разная скорость увеличения смертности с возрастом соответствует разной максимальной продолжительности жизни у разных видов . Например, мышь пожила на 3 года, человек пожил на 80 лет, [ 8 ] а деревья гинкго мало влияют на возраст даже в 667 лет. [ 9 ]
Почти все организмы стареют, включая бактерии , которые имеют асимметрию между «материнскими» и «дочерними» клетками при делении клеток , при этом материнская клетка стареет, а дочерняя омолаживается. [ 10 ] [ 11 ] , наблюдается незначительное старение В некоторых группах, например, в роде Hydra . [ 12 ] Планарийные плоские черви обладают «очевидно безграничной способностью к регенерации теломер , подпитываемой популяцией высокопролиферативных взрослых стволовых клеток ». [ 13 ] Эти планарии не биологически бессмертны , но уровень их смертности медленно увеличивается с возрастом. Организмами, которые считаются биологически бессмертными, являются, в частности, Turritopsis dohrnii , также известный как «бессмертная медуза», из-за его способности возвращаться к молодости, когда он подвергается стрессу во взрослом возрасте. [ 14 ] Репродуктивная система остается интактной и даже у Turritopsis dohrnii существуют гонады. [ 15 ]
Некоторые виды демонстрируют «негативное старение», при котором репродуктивная способность увеличивается или остается стабильной, а смертность снижается с возрастом в результате преимуществ увеличения размера тела во время старения. [ 16 ]
Теории старения
[ редактировать ]Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( март 2023 г. ) |
Было выдвинуто более 300 различных теорий, объясняющих природу (механизмы) и причины (причины естественного возникновения или факторы) старения. [ 17 ] [ необходимы дополнительные ссылки ] Хорошие теории могли бы объяснить прошлые наблюдения и предсказать результаты будущих экспериментов. Некоторые теории могут дополнять друг друга, пересекаться, противоречить или не исключать различные другие теории. [ нужна ссылка ]
Теории старения делятся на две большие категории: эволюционные теории старения и механистические теории старения. Эволюционные теории старения в первую очередь объясняют, почему происходит старение. [ 18 ] но не интересуйтесь молекулярными механизмами, которые управляют этим процессом. Все эволюционные теории старения основываются на основных механизмах ослабления силы естественного отбора с возрастом. [ 19 ] [ 20 ] Механистические теории старения можно разделить на теории, которые предполагают, что старение запрограммировано, и теории накопления повреждений, то есть те, которые предполагают, что старение вызвано специфическими молекулярными изменениями, происходящими с течением времени.
Эволюционные теории старения
[ редактировать ]Антагонистическая плейотропия
[ редактировать ]Одна теория была предложена Джорджем К. Уильямсом. [ 7 ] и включает антагонистическую плейотропию . Один ген может влиять на несколько признаков. Некоторые черты, которые улучшают физическую форму в раннем возрасте, могут также иметь негативные последствия в более позднем возрасте. Но поскольку в молодом возрасте живёт гораздо больше людей, чем в пожилом, даже небольшие положительные эффекты на раннем этапе могут быть тщательно отобраны, а против крупных отрицательных эффектов на более позднем этапе отбор может быть очень слабым. Уильямс предложил следующий пример: возможно, ген кодирует отложение кальция в костях, что способствует выживанию молоди и, следовательно, будет одобрено естественным отбором; однако этот же ген способствует отложению кальция в артериях, вызывая негативные атеросклеротические эффекты в пожилом возрасте. Таким образом, вредные биологические изменения в пожилом возрасте могут быть результатом отбора плейотропных генов, которые полезны в раннем возрасте, но вредны в дальнейшем. В этом случае давление отбора относительно велико, когда репродуктивная ценность Фишера высока, и относительно низко, когда репродуктивная ценность Фишера низка.
Теория компромисса между раком и клеточным старением
[ редактировать ]Стареющие клетки в многоклеточном организме могут быть очищены за счет конкуренции между клетками, но это увеличивает риск развития рака. Это приводит к неизбежной дилемме между двумя возможностями — накоплением физиологически бесполезных стареющих клеток и раком — обе из которых приводят к увеличению уровня смертности с возрастом. [ 2 ]
Одноразовая сома
[ редактировать ]Теория старения одноразовой сомы была предложена Томасом Кирквудом в 1977 году. [ 1 ] [ 21 ] Теория предполагает, что старение происходит из-за стратегии, согласно которой человек инвестирует в поддержание сомы только до тех пор, пока у нее есть реальный шанс на выживание. [ 22 ] Вид, который более эффективно использует ресурсы, будет жить дольше и, следовательно, сможет передать генетическую информацию следующему поколению. Требования к воспроизводству высоки, поэтому на восстановление и поддержание соматических клеток вкладывается меньше усилий по сравнению с клетками зародышевой линии , чтобы сосредоточиться на воспроизводстве и выживании вида. [ 23 ]
Теории запрограммированного старения
[ редактировать ]Программированные теории старения предполагают, что старение является адаптивным и обычно требует отбора на эволюционность или группового отбора .
Теория цикла репродуктивных клеток предполагает, что старение регулируется изменениями гормональной сигнализации на протяжении всей жизни. [ 24 ]
Теории накопления ущерба
[ редактировать ]Свободнорадикальная теория старения
[ редактировать ]Одна из наиболее известных теорий старения была впервые предложена Харманом в 1956 году. [ 25 ] В нем утверждается, что свободные радикалы, вырабатываемые растворенным кислородом, радиацией, клеточным дыханием и другими источниками, повреждают молекулярные механизмы в клетке и постепенно изнашивают их. Это также известно как окислительный стресс .
Существуют существенные доказательства, подтверждающие эту теорию. Старые животные имеют большее количество окисленных белков, ДНК и липидов, чем их более молодые собратья. [ 26 ] [ 27 ]
Химическое повреждение
[ редактировать ]Этот раздел может быть слишком длинным для удобного чтения и навигации . ( март 2023 г. ) |

Одной из самых ранних теорий старения была гипотеза уровня жизни, описанная Рэймондом Перлом в 1928 году. [ 28 ] (на основе более ранней работы Макса Рубнера ), в которой говорится, что высокая скорость основного обмена соответствует короткой максимальной продолжительности жизни .
Хотя может быть некоторая обоснованность идеи о том, что при различных типах специфических повреждений, описанных ниже, которые являются побочными продуктами метаболизма , при прочих равных условиях быстрый метаболизм может сократить продолжительность жизни, в целом эта теория не объясняет адекватно различий в Продолжительность жизни внутри вида или между видами. Животные с ограниченной калорийностью перерабатывают столько же или больше калорий на грамм массы тела, что и их сородичи, которых кормили вволю , но при этом демонстрируют значительно большую продолжительность жизни. [ нужна ссылка ] Точно так же скорость метаболизма является плохим предиктором продолжительности жизни для птиц, летучих мышей и других видов, у которых, как предполагается, снижена смертность от хищничества, и поэтому они развили большую продолжительность жизни даже при наличии очень высокой скорости метаболизма. [ 29 ] В анализе 2007 года было показано, что при использовании современных статистических методов поправки на влияние размера тела и филогении скорость метаболизма не коррелирует с продолжительностью жизни млекопитающих или птиц. [ 30 ]
Что касается конкретных типов химических повреждений, вызванных метаболизмом, предполагается, что повреждение долгоживущих биополимеров , таких как структурные белки или ДНК , вызванное повсеместно распространенными химическими агентами в организме, такими как кислород и сахара , частично ответственно за старение. . Повреждение может включать разрыв биополимерных цепей, сшивание биополимеров или химическое присоединение неприродных заместителей ( гаптенов ) к биополимерам. [ нужна ссылка ] В нормальных аэробных условиях примерно 4% кислорода, метаболизируемого митохондриями , превращается в ион супероксида , который впоследствии может превращаться в перекись водорода , гидроксильный радикал и, в конечном итоге, в другие реакционноспособные соединения, включая другие пероксиды и синглетный кислород , которые, в свою очередь, могут генерировать свободные радикалы, способные повреждать структурные белки и ДНК. [ 6 ] некоторых металлов, ионы находящихся в организме, таких как медь и железо В этом процессе могут участвовать . (При болезни Вильсона , наследственном дефекте , который заставляет организм удерживать медь, некоторые симптомы напоминают ускоренное старение.) Эти процессы, называемые окислительным стрессом , связаны с потенциальной пользой пищевых полифенольных антиоксидантов , например, в кофе , [ 31 ] и чай . [ 32 ] Однако их обычно положительное влияние на продолжительность жизни при умеренном потреблении. [ 33 ] [ 34 ] [ 35 ] также объясняются влиянием на аутофагию , [ 36 ] метаболизм глюкозы [ 37 ] и АМПК . [ 38 ]
Сахара, такие как глюкоза и фруктоза, могут вступать в реакцию с определенными аминокислотами , такими как лизин и аргинин , а также с определенными основаниями ДНК, такими как гуанин , с образованием сахарных аддуктов в процессе, называемом гликированием . Эти аддукты могут далее перегруппировываться с образованием реактивных частиц, которые затем могут сшивать структурные белки или ДНК с аналогичными биополимерами или другими биомолекулами, такими как неструктурные белки. У людей с диабетом , у которых повышен уровень сахара в крови , расстройства, связанные со старением, развиваются гораздо раньше, чем у населения в целом, но они могут отсрочить возникновение таких расстройств за счет строгого контроля уровня сахара в крови. Есть доказательства того, что повреждение сахара связано с повреждением оксидантов в процессе, называемом гликооксидированием .
Свободные радикалы могут повредить белки, липиды или ДНК . Гликирование в основном повреждает белки. Поврежденные белки и липиды накапливаются в лизосомах в виде липофусцина . Химическое повреждение структурных белков может привести к потере функции; например, повреждение коллагена стенок кровеносных сосудов может привести к жесткости стенок сосудов и, таким образом, к гипертонии , а также к утолщению стенок сосудов и образованию реактивной ткани ( атеросклерозу ); подобные процессы в почках могут привести к почечной недостаточности . Повреждение ферментов снижает функциональность клеток. липидов Перекисное окисление внутренней мембраны митохондрий снижает электрический потенциал и способность вырабатывать энергию. Вероятно, не случайно, что почти все так называемые « болезни ускоренного старения » возникают из-за дефектов ферментов репарации ДНК . [ 39 ] [ 40 ]
Считается, что влияние алкоголя на старение можно частично объяснить активацией алкоголем оси HPA , которая стимулирует секрецию глюкокортикоидов , длительное воздействие которых вызывает симптомы старения. [ 41 ]
повреждение ДНК
[ редактировать ]В обзоре 2021 года было предложено считать повреждение ДНК основной причиной старения из-за механистической связи повреждения ДНК почти со всеми аспектами фенотипа старения. [ 42 ] Было обнаружено, что более медленная скорость накопления повреждений ДНК , измеренная с помощью маркера повреждения ДНК гамма H2AX в лейкоцитах, коррелирует с более продолжительной продолжительностью жизни по сравнению с дельфинами , козами , северными оленями , американскими фламинго и белоголовыми стервятниками . [ 43 ] изменения , вызванные повреждением ДНК Эпигенетические , такие как метилирование ДНК и многие модификации гистонов , по-видимому, имеют особое значение для процесса старения. [ 42 ] Доказательства теории о том, что повреждение ДНК является фундаментальной причиной старения, были впервые рассмотрены в 1981 году. [ 44 ]
Накопление мутаций
[ редактировать ]Естественный отбор может поддерживать смертельные и вредные аллели , если их последствия ощущаются после размножения. Генетик Дж.Б.С. Холдейн задавался вопросом, почему доминантная мутация, вызывающая болезнь Хантингтона, сохранилась в популяции и почему естественный отбор не устранил ее. Начало этого неврологического заболевания приходится (в среднем) на возраст 45 лет и неизменно приводит к летальному исходу в течение 10–20 лет. Холдейн предположил, что в предыстории человечества лишь немногие доживали до 45 лет. вредные мутации были соответственно небольшими. Следовательно, генетическая нагрузка поздних вредных мутаций может быть значительной при балансе мутации и отбора . Эта концепция стала известна как тень выбора . [ 45 ]
Питер Медавар формализовал это наблюдение в своей накоплении мутаций . теории старения, основанной на [ 46 ] [ 47 ] «Сила естественного отбора ослабевает с возрастом — даже в теоретически бессмертной популяции, при условии, что она подвергается реальной угрозе смертности. Если генетическая катастрофа… происходит достаточно поздно в жизни индивидуума, ее последствия могут быть совершенно незначительными. ". Независящие от возраста опасности, такие как хищничество, болезни и несчастные случаи, называемые « внешней смертностью », означают, что даже в популяции с незначительным старением будет меньше живых особей в старших возрастных группах.
Прочие повреждения
[ редактировать ]Исследование пришло к выводу, что ретровирусы в геномах человека могут пробуждаться из спящих состояний и способствовать старению, которое можно блокировать нейтрализующими антителами , облегчая «клеточное старение и дегенерацию тканей и, в некоторой степени, старение организма». [ 48 ]
Теории старения, основанные на стволовых клетках
[ редактировать ]Теория старения стволовых клеток постулирует, что процесс старения является результатом неспособности различных типов стволовых клеток продолжать пополнять ткани организма способными функционально дифференцированными клетками, этой ткани (или органа поддерживать исходную функцию ). Накопление повреждений и ошибок в генетическом материале всегда является проблемой для систем, независимо от возраста. Количество стволовых клеток у молодых людей намного выше, чем у пожилых людей, и, таким образом, у молодых людей создается лучший и более эффективный механизм замены, в отличие от пожилых. Другими словами, старение — это не вопрос увеличения повреждений, а вопрос неспособности заменить их из-за уменьшения количества стволовых клеток. Количество стволовых клеток уменьшается и они имеют тенденцию терять способность дифференцироваться в потомство или лимфоидные линии и миелоидные .
Поддержание динамического баланса пулов стволовых клеток требует нескольких условий. Балансирование пролиферации и покоя наряду с хомингом ( см. нишу ) и самообновлением гемопоэтических стволовых клеток являются благоприятными элементами поддержания пула стволовых клеток, тогда как дифференцировка, мобилизация и старение являются вредными элементами. Эти вредные эффекты в конечном итоге вызовут апоптоз .
Существует также несколько проблем, когда речь идет о терапевтическом использовании стволовых клеток и их способности пополнять запасы органов и тканей. Во-первых, разные клетки могут иметь разную продолжительность жизни, даже если они происходят из одних и тех же стволовых клеток ( см. Т-клетки и эритроциты ), а это означает, что старение может происходить по-разному в клетках с большей продолжительностью жизни, в отличие от клеток с более короткой продолжительностью жизни. Кроме того, постоянные усилия по замене соматических клеток могут привести к истощению стволовых клеток. [ 49 ]- Старение гемопоэтических стволовых клеток
- Старение разнообразия гемопоэтических стволовых клеток
- Гематопоэтическая мозаичная потеря хромосомы Y
Биомаркеры старения
[ редактировать ]Если разные люди стареют с разной скоростью, то плодовитость, смертность и функциональные способности лучше предсказать с помощью биомаркеров , чем с помощью хронологического возраста. [ 58 ] [ 59 ] Однако седина , [ 60 ] Старение лица , морщины на коже и другие общие изменения, наблюдаемые с возрастом, не являются лучшими показателями будущей функциональности, чем хронологический возраст. Биогеронтологи продолжают попытки найти и подтвердить биомаркеры старения, но успех пока ограничен.
Уровни CD4 и CD8 Т-клеток памяти и наивных Т-клеток использовались для получения хороших прогнозов ожидаемой продолжительности жизни мышей среднего возраста. [ 61 ]
Стареющие часы
[ редактировать ]Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( март 2023 г. ) |
Существует интерес к эпигенетическим часам как биомаркеру старения, основанному на их способности предсказывать хронологический возраст человека. [ 62 ] Базовую биохимию крови и подсчет клеток также можно использовать для точного прогнозирования хронологического возраста. [ 63 ] Также возможно предсказать хронологический возраст человека с помощью транскриптомных часов старения. [ 64 ]
Ведутся исследования и разработки дополнительных биомаркеров, систем обнаружения и программного обеспечения для измерения биологического возраста различных тканей или систем или в целом. Например, программное обеспечение глубокого обучения (DL), использующее анатомические магнитно-резонансные изображения, оценивало возраст мозга с относительно высокой точностью, включая обнаружение ранних признаков болезни Альцгеймера и различных нейроанатомических моделей неврологического старения. [ 65 ] человека Сообщалось, что инструмент DL позволяет рассчитать воспалительный возраст на основе закономерностей системного возрастного воспаления. [ 66 ]
Часы старения использовались для оценки воздействия вмешательств на людей, включая комбинированную терапию . [ 67 ] [ необходимы дополнительные ссылки ] Использование часов старения для выявления и оценки мер по продлению жизни представляет собой фундаментальную цель исследований в области биологии старения. Однако достижение этой цели требует преодоления многочисленных проблем и реализации дополнительных шагов проверки. [ 68 ] [ 69 ]
Генетические детерминанты старения
[ редактировать ]Ряд генетических компонентов старения был идентифицирован с использованием модельных организмов, от простых почкующихся дрожжей Saccharomyces cerevisiae до червей, таких как Caenorhabditis elegans , и плодовых мух ( Drosophila melanogaster ). Изучение этих организмов выявило наличие как минимум двух консервативных путей старения.
Экспрессия генов несовершенно контролируется, и возможно, что случайные колебания уровней экспрессии многих генов способствуют процессу старения, как показывают исследования таких генов у дрожжей. [ 70 ] Отдельные клетки, которые генетически идентичны, тем не менее могут иметь существенно разные реакции на внешние раздражители и заметно разную продолжительность жизни, что указывает на то, что эпигенетические факторы играют важную роль в экспрессии генов и старении, а также генетические факторы. Существуют исследования по эпигенетике старения .
Способность восстанавливать двухцепочечные разрывы ДНК у мышей снижается с возрастом [ 71 ] и люди. [ 72 ]
ряд редких наследственных ( генетических ) заболеваний, каждое из которых называется прогерией В течение некоторого времени известен . У больных проявляются симптомы, напоминающие ускоренное старение , включая морщинистую кожу . Причина синдрома прогерии Хатчинсона-Гилфорда была описана в журнале Nature в мае 2003 года. [ 73 ] В этом отчете предполагается, что повреждение ДНК , а не окислительный стресс причиной этой формы ускоренного старения является .
Исследование показывает, что старение может смещать активность в сторону более коротких генов или более короткой длины транскриптов, и этому можно противостоять с помощью вмешательств. [ 74 ]
Продолжительность здоровья и старение в обществе
[ редактировать ]

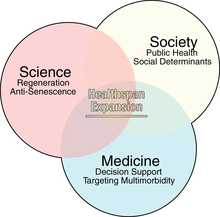
Продолжительность здоровья в широком смысле можно определить как период жизни, в течение которого человек здоров , например, не имеет серьезных заболеваний. [ 76 ] или снижение способностей (например, чувств, мышц , выносливости и познания ).
Биологическое старение или LHG влечет за собой большое бремя затрат для общества, включая потенциально растущие расходы на здравоохранение (также в зависимости от типов и стоимости лечения ). [ 75 ] [ 79 ] Это, наряду с глобальным качеством жизни и благополучием , подчеркивает важность увеличения продолжительности здоровья. [ 75 ]
Многие меры, которые могут продлить продолжительность жизни, могут одновременно также продлить продолжительность здоровья, хотя это не обязательно так, что указывает на то, что «продолжительность жизни больше не может быть единственным параметром, представляющим интерес» в соответствующих исследованиях. [ 80 ] Хотя недавнее увеличение продолжительности жизни не сопровождалось «параллельным» увеличением продолжительности жизни, [ 75 ] Осведомленность о концепции и вопросах продолжительности здоровья отстает по состоянию на 2017 год. [ 76 ] Ученые отметили, что « хронические заболевания старения растут и наносят неисчислимый ущерб качеству жизни человека». [ 79 ]
Вмешательства
[ редактировать ]Продление жизни — это концепция увеличения продолжительности жизни человека либо скромно за счет усовершенствований в медицине, либо резко за счет увеличения максимальной продолжительности жизни сверх общепринятого биологического предела, составляющего около 125 лет . [ 81 ] Некоторые исследователи в этой области, наряду с «сторонниками продления жизни», « имморталистами » или « долгожителями » (теми, кто сам хочет добиться более продолжительной жизни), постулируют, что будущие прорывы в омоложении тканей , стволовых клетках , регенеративной медицине , молекулярном восстановлении, генном терапия , фармацевтические препараты и замена органов (например, искусственными органами или ксенотрансплантациями ) в конечном итоге позволят людям иметь неопределенную продолжительность жизни за счет полного омоложения до здорового молодого состояния (агеразия). [ 82 ] ). Этические последствия, если продление жизни станет возможным, обсуждаются биоэтиками .
Продажа предполагаемых антивозрастных продуктов, таких как добавки и заменители гормонов, является прибыльной мировой индустрией. Например, индустрия, которая пропагандирует использование гормонов в качестве лечения для потребителей, чтобы замедлить или обратить вспять процесс старения на рынке США, в 2009 году принесла около 50 миллиардов долларов дохода в год. [ 83 ] Эффективность и безопасность использования таких гормональных препаратов не доказана. [ 83 ] [ 84 ] [ 85 ] [ 86 ]См. также
[ редактировать ]- Антивозрастное движение
- Антимускариновые средства
- Деменция
- восстановление ДНК
- Гериатрия
- Геронтология
- Гомеостатическая способность
- Бессмертие
- Указатель тем, связанных с продлением жизни
- Митогормезис
- Старость
- Феноптоз
- Старение растений
- Запрограммированная гибель клеток
- Стратегии искусственного незначительного старения (SENS)
- Сублетальные повреждения
- Трансгенерационный дизайн
- Хронология исследований старения
Ссылки
[ редактировать ]- ^ Jump up to: а б Кирквуд ТБ (1977). «Эволюция старения» . Природа . 270 (5635): 301–4. Бибкод : 1977Natur.270..301K . дои : 10.1038/270301a0 . ПМИД 593350 . S2CID 492012 . Архивировано из оригинала 15 ноября 2023 года . Проверено 31 января 2022 г.
- ^ Jump up to: а б Нельсон П., Мэйсел Дж. (декабрь 2017 г.). «Межклеточная конкуренция и неизбежность многоклеточного старения» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (49): 12982–7. Бибкод : 2017PNAS..11412982N . дои : 10.1073/pnas.1618854114 . ПМЦ 5724245 . ПМИД 29087299 .
- ^ «Старение и геронтологический словарь» . Архивировано из оригинала 19 октября 2019 года . Проверено 26 февраля 2011 г.
- ^ Лопес-Отин С., Бласко М.А., Партридж Л. и др. (июнь 2013 г.). «Признаки старения» . Клетка . 153 (6): 1194–217. дои : 10.1016/j.cell.2013.05.039 . ПМЦ 3836174 . ПМИД 23746838 .
- ^ Лопес-Отин С., Бласко М.А., Партридж Л. и др. (19 января 2023 г.). «Признаки старения: расширяющаяся Вселенная» . Клетка . 186 (2): 243–278. дои : 10.1016/j.cell.2022.11.001 . ПМИД 36599349 . S2CID 255394876 .
- ^ Jump up to: а б Холмс Г.Е., Бернштейн С., Бернштейн Х. (сентябрь 1992 г.). «Окислительные и другие повреждения ДНК как основа старения: обзор». Мутат. Рез . 275 (3–6): 305–15. дои : 10.1016/0921-8734(92)90034-м . ПМИД 1383772 .
- ^ Jump up to: а б Уильямс Г.К. (1957). «Плейотропия, естественный отбор и эволюция старения». Эволюция . 11 (4): 398–411. дои : 10.2307/2406060 . JSTOR 2406060 .
- ^ Аустад СН (февраль 2009 г.). «Сравнительная биология старения» . Журналы геронтологии. Серия А, Биологические и медицинские науки . 64 (2): 199–201. дои : 10.1093/gerona/gln060 . ПМК 2655036 . ПМИД 19223603 .
- ^ Ван Л., Цуй Дж., Цзинь Б. и др. (январь 2020 г.). «Многофункциональный анализ сосудистых камбиальных клеток выявляет механизмы долголетия старых деревьев гинкго двулопастного » . Труды Национальной академии наук Соединенных Штатов Америки . 117 (4): 2201–10. Бибкод : 2020PNAS..117.2201W . дои : 10.1073/pnas.1916548117 . ПМК 6995005 . ПМИД 31932448 .
- ^ Акерманн М., Стернс СК, Дженал У (июнь 2003 г.). «Старение бактерии с асимметричным делением». Наука . 300 (5627): 1920. doi : 10.1126/science.1083532 . ПМИД 12817142 . S2CID 34770745 .
- ^ Стюарт Э.Дж., Мэдден Р., Пол Дж. и др. (февраль 2005 г.). «Старение и смерть организма, размножающегося морфологически симметричным делением» . ПЛОС Биология . 3 (2): е45. doi : 10.1371/journal.pbio.0030045 . ПМК 546039 . ПМИД 15685293 .
- ^ Данько М.Ю., Козловски Ю., Шайбле Р. (октябрь 2015 г.). «Раскрытие феномена отсутствия старения на Гидре» . Журнал теоретической биологии . 382 : 137–49. Бибкод : 2015JThBi.382..137D . дои : 10.1016/j.jtbi.2015.06.043 . ПМИД 26163368 .
- ^ Тан Т.С., Рахман Р., Джабер-Хиджази Ф. и др. (март 2012 г.). «Поддержание теломер и активность теломеразы по-разному регулируются у бесполых и половых червей» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (11): 4209–14. Бибкод : 2012PNAS..109.4209T . дои : 10.1073/pnas.1118885109 . ПМК 3306686 . ПМИД 22371573 .
- ^ Лисенкова А.А., Григоренко А.П., Тяжелова Т.В. и др. (февраль 2017 г.). «Полный митохондриальный геном и эволюционный анализ Turritopsis dohrnii, «бессмертной» медузы с обратимым жизненным циклом» . Молекулярная филогенетика и эволюция . 107 : 232–8. Бибкод : 2017МолПЭ.107..232Л . дои : 10.1016/j.ympev.2016.11.007 . ПМИД 27845203 .
- ^ Пираино С., Боэро Ф., Эшбах Б. и др. (июнь 1996 г.). «Обращение жизненного цикла: медузы превращаются в полипы и трансдифференцировка клеток у Turritopsis nutricula (Cnidaria, Hydrozoa)». Биологический вестник . 190 (3): 302–312. дои : 10.2307/1543022 . JSTOR 1543022 . ПМИД 29227703 .
- ^ Ваупель Дж.В., Баудиш А., Доллинг М. и др. (июнь 2004 г.). «Дело об отрицательном старении». Теоретическая популяционная биология . 65 (4): 339–51. Бибкод : 2004TPBio..65..339W . дои : 10.1016/j.tpb.2003.12.003 . ПМИД 15136009 .
- ^ Винья Х, Боррас С, Микель Дж (2007). «Теории старения» . ИУБМБ Жизнь . 59 (4–5): 249–54. дои : 10.1080/15216540601178067 . ПМИД 17505961 .
- ^ Кирквуд Т.Б., Аустад С.Н. (2000). «Почему мы стареем?» . Природа . 408 (6809): 233–8. Бибкод : 2000Natur.408..233K . дои : 10.1038/35041682 . ПМИД 11089980 . S2CID 2579770 . Архивировано из оригинала 15 ноября 2023 года . Проверено 31 января 2022 г.
- ^ Медавар П.Б. (1952). Нерешенная проблема биологии . Опубликовано для колледжа Х. К. Льюисом . OCLC 869293719 .
- ^ Роуз М.Р. (1991). Эволюционная биология старения . Издательство Оксфордского университета. ISBN 1-4237-6520-6 . OCLC 228167629 . Архивировано из оригинала 15 ноября 2023 года . Проверено 31 января 2022 г.
- ^ Кирквуд Т. (2006). Время нашей жизни: наука о старении человека . Издательство Оксфордского университета. ISBN 978-0-19-802939-7 . OCLC 437175125 . Архивировано из оригинала 15 ноября 2023 года . Проверено 31 января 2022 г.
- ^ Хаммерс М., Ричардсон Д.С., Берк Т. и др. (сентябрь 2013 г.). «Влияние репродуктивных инвестиций и условий окружающей среды в раннем возрасте на старение: поддержка гипотезы одноразовой сомы». Журнал эволюционной биологии . 26 (9): 1999–2007. дои : 10.1111/jeb.12204 . hdl : 11370/9cc6749c-f67d-40ab-a253-a06650c32102 . ПМИД 23961923 . S2CID 46466320 .
- ^ Кирквуд Т.Б., Роуз М.Р. (апрель 1991 г.). «Эволюция старения: позднее выживание приносится в жертву воспроизводству». Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 332 (1262): 15–24. Бибкод : 1991РСПТБ.332...15К . дои : 10.1098/rstb.1991.0028 . ПМИД 1677205 .
- ^ Этвуд CS, Боуэн Р.Л. (2011). «Теория старения репродуктивно-клеточного цикла: обновление». Экспериментальная геронтология . 46 (2–3): 100–7. дои : 10.1016/j.exger.2010.09.007 . ПМИД 20851172 . S2CID 20998909 .
- ^ Харман Д. (июль 1956 г.). «Старение: теория, основанная на свободнорадикальной и радиационной химии». Журнал геронтологии . 11 (3): 298–300. дои : 10.1093/geronj/11.3.298 . hdl : 2027/mdp.39015086547422 . ПМИД 13332224 .
- ^ Штадтман Э.Р. (август 1992 г.). «Окисление белков и старение» . Наука . 257 (5074): 1220–4. Бибкод : 1992Sci...257.1220S . дои : 10.1126/science.1355616 . ПМИД 1355616 . Архивировано из оригинала 31 июля 2021 года . Проверено 21 июля 2021 г.
- ^ Сохал Р.С., Агарвал С., Дубей А. и др. (август 1993 г.). «Окислительное повреждение белков связано с продолжительностью жизни комнатных мух» . Труды Национальной академии наук Соединенных Штатов Америки . 90 (15): 7255–9. Бибкод : 1993PNAS...90.7255S . дои : 10.1073/pnas.90.15.7255 . ПМЦ 47115 . ПМИД 8346242 .
- ^ Перл Р. (1928). Скорость жизни как отчет о некоторых экспериментальных исследованиях биологии продолжительности жизни . Нью-Йорк: Альфред А. Кнопф. [ нужна страница ]
- ^ Брюне-Россинни АК, Аустад СН (2004). «Исследования старения летучих мышей: обзор». Биогеронтология . 5 (4): 211–22. дои : 10.1023/B:BGEN.0000038022.65024.d8 . ПМИД 15314271 . S2CID 22755811 .
- ^ де Магальяйнс ЖП, Коста Дж, генеральный директор Черча (февраль 2007 г.). «Анализ взаимосвязи между метаболизмом, графиками развития и долголетием с использованием независимых филогенетических контрастов» . Журналы геронтологии. Серия А, Биологические и медицинские науки . 62 (2): 149–60. CiteSeerX 10.1.1.596.2815 . дои : 10.1093/герона/62.2.149 . ПМК 2288695 . ПМИД 17339640 .
- ^ Фридман Н.Д., Парк Ю., Абнет К.С. и др. (май 2012 г.). «Связь употребления кофе с общей смертностью и смертностью от конкретных причин» . Медицинский журнал Новой Англии . 366 (20): 1891–904. дои : 10.1056/NEJMoa1112010 . ПМК 3439152 . ПМИД 22591295 .
- ^ Ян Ю, Чан СВ, Ху М и др. (2011). «Влияние некоторых распространенных пищевых компонентов на сердечно-сосудистые заболевания» . ISRN Кардиология . 2011 : 397136. doi : 10.5402/2011/397136 . ПМЦ 3262529 . ПМИД 22347642 .
- ^ Пул Р., Кеннеди О.Дж., Родерик П. и др. (22 ноября 2017 г.). «Потребление кофе и здоровье: общий обзор метаанализа многочисленных последствий для здоровья» . БМЖ . 359 : j5024. дои : 10.1136/bmj.j5024 . ПМЦ 5696634 . ПМИД 29167102 .
- ^ О'Киф Дж.Х., ДиНиколантонио Дж.Дж., Лави СиДж. (1 мая 2018 г.). «Кофе для кардиозащиты и долголетия». Прогресс в сердечно-сосудистых заболеваниях . 61 (1): 38–42. дои : 10.1016/j.pcad.2018.02.002 . ПМИД 29474816 .
- ^ Гроссо Дж., Годос Дж., Гальвано Ф. и др. (21 августа 2017 г.). «Кофе, кофеин и последствия для здоровья: общий обзор». Ежегодный обзор питания . 37 (1): 131–156. doi : 10.1146/annurev-nutr-071816-064941 . ПМИД 28826374 .
- ^ Диркс-Нейлор Эй Джей (15 декабря 2015 г.). «Польза кофе для скелетных мышц». Науки о жизни . 143 : 182–6. дои : 10.1016/j.lfs.2015.11.005 . ПМИД 26546720 .
- ^ Рейс CE, Дореа Ж.Г., да Коста TH (1 июля 2019 г.). «Влияние потребления кофе на метаболизм глюкозы: систематический обзор клинических испытаний» . Журнал традиционной и дополнительной медицины . 9 (3): 184–191. дои : 10.1016/j.jtcme.2018.01.001 . ПМК 6544578 . ПМИД 31193893 .
- ^ Лоурейро Л.М., Рейс CE, Коста TH (1 мая 2018 г.). «Влияние компонентов кофе на восстановление мышечного гликогена: систематический обзор». Международный журнал спортивного питания и метаболизма при физических нагрузках . 28 (3): 284–293. дои : 10.1123/ijsnem.2017-0342 . ПМИД 29345166 .
- ^ Бернштейн Х., Пейн СМ, Бернштейн С. и др. (2008). «Рак и старение как последствия невосстановленного повреждения ДНК». . Кимура Х, Сузуки А (ред.). Новое исследование повреждения ДНК . Издательство Nova Science. стр. 1–47. ISBN 978-1604565812 . Архивировано из оригинала 15 ноября 2023 года . Проверено 4 февраля 2016 г.
- ^ Пан М.Р., Ли К., Лин С.Ю. и др. (май 2016 г.). «Соединяя точки: от повреждения и восстановления ДНК до старения» . Международный журнал молекулярных наук . 17 (5): 685. doi : 10.3390/ijms17050685 . ПМЦ 4881511 . ПМИД 27164092 .
- ^ Спенсер Р.Л., Хатчисон К.Э. (1999). «Алкоголь, старение и реакция на стресс» (PDF) . Исследования алкоголя и здоровье . 23 (4): 272–83. ПМК 6760387 . ПМИД 10890824 . Архивировано из оригинала (PDF) 11 декабря 2018 года . Проверено 8 апреля 2008 г.
- ^ Jump up to: а б Шумахер Б., Потхоф Дж., Вийг Дж. и др. (апрель 2021 г.). «Центральная роль повреждения ДНК в процессе старения» . Природа . 592 (7856): 695–703. Бибкод : 2021Natur.592..695S . дои : 10.1038/s41586-021-03307-7 . ПМЦ 9844150 . ПМИД 33911272 .
- ^ Уиттемор К., Мартинес-Невадо Э., Бласко М.А. (ноябрь 2019 г.). «Более медленные темпы накопления повреждений ДНК в лейкоцитах коррелируют с большей продолжительностью жизни у некоторых видов птиц и млекопитающих» . Старение (Олбани, штат Нью-Йорк) . 11 (21): 9829–45. дои : 10.18632/aging.102430 . ПМК 6874430 . ПМИД 31730540 .
- ^ Генслер Х.Л., Бернштейн Х. (сентябрь 1981 г.). «Повреждение ДНК как основная причина старения» . Ежеквартальный обзор биологии . 56 (3): 279–303. дои : 10.1086/412317 . ПМИД 7031747 . S2CID 20822805 .
- ^ Фабиан Д., Флэтт Т. (2011). «Эволюция старения» (PDF) . Природное образование .
- ^ Медавар ПБ (1946). «Старость и естественная смерть». Современный ежеквартальный журнал . 1 :30–56.
- ^ Медавар 1952 г. [ нужна страница ]
- ^ Лю X, Лю Z, Ву Z и др. (19 января 2023 г.). «Возрождение эндогенных ретровирусов во время старения усиливает старение» . Клетка . 186 (2): 287–304.e26. дои : 10.1016/j.cell.2022.12.017 . ПМИД 36610399 . S2CID 232060038 .
- Экспертное объяснение исследования: «Старение и ретровирусы» . Наука . Архивировано из оригинала 17 февраля 2023 года . Проверено 17 февраля 2023 г.
- ^ Смит Дж., А., Дэниел Р. «Стволовые клетки и старение: проблема курицы или яйца?». Старение и болезни . Июнь 2012 г., Том. 3, номер 3; 260–268.
- ^ Махла РС (2016). «Применение стволовых клеток в регенеративной медицине и лечении заболеваний» . Международный журнал клеточной биологии . 2016 (7): 19. doi : 10.1155/2016/6940283 . ПМЦ 4969512 . ПМИД 27516776 .
- ^ Росси Дж., Брайдер Д., Сейта Дж. и др. (2007). «Недостаток восстановления повреждений ДНК ограничивает функцию гемопоэтических стволовых клеток с возрастом». Природа . 447 (7145): 725–9. Бибкод : 2007Natur.447..725R . дои : 10.1038/nature05862 . ПМИД 17554309 . S2CID 4416445 .
- ^ Jump up to: а б Бирман И., Сейта Дж., Инлей М.А. и др. (2014). «Покоящиеся гемопоэтические стволовые клетки накапливают повреждения ДНК во время старения, которые восстанавливаются при входе в клеточный цикл» . Клеточная стволовая клетка . 15 (1): 37–50. дои : 10.1016/j.stem.2014.04.016 . ПМЦ 4082747 . ПМИД 24813857 .
- ^ Jump up to: а б Нижник А., Вудбайн Л., Маркетти С. и др. (2007). «Репарация ДНК ограничивает гемопоэтические стволовые клетки во время старения». Природа . 447 (7145): 686–90. Бибкод : 2007Natur.447..686N . дои : 10.1038/nature05875 . ПМИД 17554302 . S2CID 4332976 .
- ^ «Исследования могут раскрыть, почему люди в возрасте 70 лет могут внезапно стать слабыми» . Хранитель . 1 июня 2022 г. Проверено 18 июля 2022 г.
- ^ Митчелл Э., Спенсер Чепмен М., Уильямс Н. и др. (июнь 2022 г.). «Клональная динамика кроветворения на протяжении всей жизни человека» . Природа . 606 (7913): 343–350. Бибкод : 2022Natur.606..343M . дои : 10.1038/s41586-022-04786-y . ISSN 1476-4687 . ПМЦ 9177428 . ПМИД 35650442 .
- ^ Колата Г (14 июля 2022 г.). «Поскольку Y-хромосомы исчезают с возрастом, риск сердечно-сосудистых заболеваний может возрастать» . Нью-Йорк Таймс . Проверено 21 августа 2022 г.
- ^ Сано С., Хоритани К., Огава Х. и др. (15 июля 2022 г.). «Гематопоэтическая потеря Y-хромосомы приводит к сердечному фиброзу и смертности от сердечной недостаточности» . Наука . 377 (6603): 292–297. Бибкод : 2022Sci...377..292S . дои : 10.1126/science.abn3100 . ISSN 0036-8075 . ПМЦ 9437978 . ПМИД 35857592 .
- ^ Гасми А., Чирумболо С., Пеана М. и др. (17 сентября 2020 г.). «Биомаркеры старения во время старения как возможные предупреждения о необходимости использования профилактических мер». Современная медицинская химия . 28 (8): 1471–88. дои : 10.2174/0929867327999200917150652 . ПМИД 32942969 . S2CID 221789280 .
- ^ Бейкер Г.Т., Спротт Р.Л. (1988). «Биомаркеры старения» . Экспериментальная геронтология . 23 (4–5): 223–39. дои : 10.1016/0531-5565(88)90025-3 . ПМИД 3058488 . S2CID 31039588 . Архивировано из оригинала 24 октября 2021 года . Проверено 12 июля 2019 г.
- ^ Ван Несте Д., Тобин DJ (2004). «Цикл роста волос и пигментация волос: динамические взаимодействия и изменения, связанные со старением». Микрон . 35 (3): 193–200. дои : 10.1016/j.micron.2003.11.006 . ПМИД 15036274 .
- ^ Миллер Р.А. (апрель 2001 г.). «Биомаркеры старения: прогнозирование продолжительности жизни с использованием определения возрастной субпопуляции Т-клеток в генетически гетерогенной популяции мышей среднего возраста» . Журналы геронтологии. Серия А, Биологические и медицинские науки . 56 (4): Б180-6. дои : 10.1093/gerona/56.4.b180 . ПМЦ 7537444 . ПМИД 11283189 .
- ^ Науэ Дж (октябрь 2023 г.). «Определение хронологического возраста ДНК: использование данных о возрастном метилировании ДНК для судебно-медицинской экспертизы ДНК» . Гены и геномика . 45 (10): 1239–1261. дои : 10.1007/s13258-023-01392-8 . ПМЦ 10504122 .
- ^ Путин Э., Мамошина П., Алипер А. и др. (май 2016 г.). «Глубокие биомаркеры старения человека: применение глубоких нейронных сетей для разработки биомаркеров» . Старение . 8 (5): 1021–33. дои : 10.18632/aging.100968 . ПМЦ 4931851 . ПМИД 27191382 .
- ^ Петерс М.Дж., Джоханес Р., Пиллинг Л.С. и др. (октябрь 2015 г.). «Транскрипционный ландшафт возраста в периферической крови человека» . Природные коммуникации . 6 : 8570. Бибкод : 2015NatCo...6.8570. . дои : 10.1038/ncomms9570 . ПМЦ 4639797 . ПМИД 26490707 .
- ^ Инь С., Иммс П., Ченг М. и др. (10 января 2023 г.). «Анатомически интерпретируемое глубокое изучение возраста мозга фиксирует когнитивные нарушения, специфичные для конкретной области» . Труды Национальной академии наук . 120 (2): e2214634120. Бибкод : 2023PNAS..12014634Y . дои : 10.1073/pnas.2214634120 . ПМЦ 9926270 . ПМИД 36595679 .
- Пресс-релиз университета: «Сколько на самом деле лет вашему мозгу? Анализ с помощью искусственного интеллекта точно отражает риск снижения когнитивных функций и болезни Альцгеймера» . Университет Южной Калифорнии (medicalxpress.com) . Архивировано из оригинала 17 февраля 2023 года . Проверено 17 февраля 2023 г.
- Новостная статья об исследовании: «ИИ может определить истинный возраст мозга» . Deutschlandfunk Nova (на немецком языке). Архивировано из оригинала 17 февраля 2023 года . Проверено 17 февраля 2023 г.
- ^ Сайед Н., Хуанг Й., Нгуен К. и др. (июль 2021 г.). «Часы воспалительного старения (iAge), основанные на глубоком обучении, отслеживают мультиморбидность, старение иммунитета, слабость и старение сердечно-сосудистой системы» . Природное старение . 1 (7): 598–615. дои : 10.1038/s43587-021-00082-y . ПМЦ 8654267 . PMID 34888528 .
- Новостная статья об исследовании: «Инструмент, вычисляющий возраст иммунной системы, может предсказать слабость и болезни» . Новый Атлас . 13 июля 2021 года. Архивировано из оригинала 26 июля 2021 года . Проверено 26 июля 2021 г.
- ^ Фицджеральд К.Н., Ходжес Р., Хейнс Д. и др. (2021). «Потенциальное изменение эпигенетического возраста с помощью диеты и изменения образа жизни: пилотное рандомизированное клиническое исследование» . Старение . 13 (7): 9419–32. дои : 10.18632/aging.202913 . ПМК 8064200 . ПМИД 33844651 . Архивировано из оригинала 2 июня 2021 года . Проверено 28 июня 2021 г.
- ^ Мокри М., Херцог С., Поганик Дж.Р. и др. (август 2023 г.). «Биомаркеры старения для выявления и оценки мер по продлению жизни» . Клетка . 186 (18): 3758–75. дои : 10.1016/j.cell.2023.08.003 . ПМЦ 11088934 . ПМИД 37657418 .
- ^ Мокри М., Херцог С., Поганик Дж.Р. и др. (февраль 2024 г.). «Валидация биомаркеров старения» . Природная медицина . 30 (2): 360–372. дои : 10.1038/s41591-023-02784-9 . ISSN 1546-170Х . ПМК 11090477 . ПМИД 38355974 .
- ^ Райли Дж., Перейра-Смит О.М. (2006). «Микрофлюидное устройство для анализа экспрессии генов в отдельных клетках Saccharomyces cerevisiae». Дрожжи . 23 (14–15): 1065–73. дои : 10.1002/да.1412 . ПМИД 17083143 . S2CID 31356425 .
- ^ Вайдья А., Мао З., Тянь Х. и др. (июль 2014 г.). «Мыши-репортёры с нокаутом демонстрируют, что репарация ДНК за счёт негомологичного соединения концов снижается с возрастом» . ПЛОС Генет . 10 (7): e1004511. дои : 10.1371/journal.pgen.1004511 . ПМК 4102425 . ПМИД 25033455 .
- ^ Ли З, Чжан В, Чен Ю и др. (ноябрь 2016 г.). «Нарушение репарации двухцепочечных разрывов ДНК способствует возрастному росту геномной нестабильности у людей» . Гибель клеток отличается . 23 (11): 1765–77. дои : 10.1038/cdd.2016.65 . ПМК 5071568 . ПМИД 27391797 .
- ^ Мункес Л.С., Козлов С., Эрнандес Л. и др. (май 2003 г.). «Прогероидный синдром у мышей вызван дефектами ламинов А-типа» . Природа . 423 (6937): 298–301. Бибкод : 2003Natur.423..298M . дои : 10.1038/nature01631 . ПМИД 12748643 . S2CID 4360055 . Архивировано из оригинала 30 мая 2022 года . Проверено 21 июля 2021 г. - через Zenodo.
- ^ Стогер Т., Грант Р.А., Маккуатти-Пиментел А.С. и др. (декабрь 2022 г.). «Старение связано с системным дисбалансом транскриптома, связанным с длиной» . Природное старение . 2 (12): 1191–1206. дои : 10.1038/s43587-022-00317-6 . ПМЦ 10154227 . ПМИД 37118543 .
- Пресс-релиз университета: «Старение обусловлено несбалансированными генами, показал анализ ИИ нескольких видов» . Северо-Западный университет . 9 декабря 2022 года. Архивировано из оригинала 2 февраля 2023 года . Проверено 18 января 2023 г. - через phys.org.
- Новостная статья об исследовании: Квон Д. (6 января 2023 г.). «Старение связано с большей активностью коротких генов, чем длинных» . Научный американец . Архивировано из оригинала 17 января 2023 года . Проверено 18 января 2023 г.
- ^ Jump up to: а б с д и ж Гармани А, Ямада С, Терзич А (23 сентября 2021 г.). «Скачок долголетия: обратите внимание на разницу в продолжительности жизни» . npj Регенеративная медицина . 6 (1): 57. дои : 10.1038/s41536-021-00169-5 . ПМЦ 8460831 . ПМИД 34556664 .
- Пресс-релиз некоммерческой больницы: Баклс С. (7 октября 2021 г.). «Регенеративная перезагрузка старения» Центр регенеративной биотерапии . Клиника Мэйо . Архивировано из оригинала 1 марта 2023 года . Проверено 1 марта 2023 г.
- ^ Jump up to: а б Петерсон Т. (30 мая 2017 г.). «Продолжительность здоровья важнее продолжительности жизни, так почему же больше людей об этом не знает?» . Институт общественного здравоохранения . Вашингтонский университет в Сент-Луисе. Центр старения Харви А. Фридмана. Архивировано из оригинала 1 марта 2023 года . Проверено 1 марта 2023 г.
- ^ Гармани А., Ямада С., Терзич А. (сентябрь 2021 г.). «Скачок долголетия: обратите внимание на разницу в продолжительности жизни» . npj Регенеративная медицина . 6 (1): 57. дои : 10.1038/s41536-021-00169-5 . ПМЦ 8460831 . ПМИД 34556664 .
- ^ Фаррелли С. (ноябрь 2022 г.). «Старение, равенство и продолжительность здоровья человека» . Форум ХЭК . 36 (2): 187–205. дои : 10.1007/s10730-022-09499-3 . ПМЦ 9644010 . ПМИД 36348214 .
- ^ Jump up to: а б Хансен М., Кеннеди Б.К. (1 августа 2016 г.). «Означает ли более продолжительная продолжительность жизни более продолжительная продолжительность здоровья?» . Тенденции в клеточной биологии . 26 (8): 565–8. дои : 10.1016/j.tcb.2016.05.002 . ПМЦ 4969078 . ПМИД 27238421 .
- ^ Бансал А., Чжу Л.Дж., Йен К. и др. (20 января 2015 г.). «Развязка продолжительности жизни и продолжительности здоровья у мутантов долголетия Caenorhabditis elegans» . Труды Национальной академии наук . 112 (3): Е277-86. Бибкод : 2015PNAS..112E.277B . дои : 10.1073/pnas.1412192112 . ПМК 4311797 . ПМИД 25561524 .
- ^ Тернер Б.С. (2009). Можем ли мы жить вечно? Социологическое и моральное исследование . Гимн Пресс. п. 3.
- ^ «агеразия» . Оксфордский словарь английского языка (онлайн-изд.). Издательство Оксфордского университета . (Требуется подписка или членство участвующей организации .)
- ^ Jump up to: а б Япсен Б. (15 июня 2009 г.). «Отчет АМА ставит под сомнение науку об использовании гормонов в качестве средства против старения» . Чикаго Трибьюн . Проверено 17 июля 2009 г.
- ^ Холлидей Р. (апрель 2009 г.). «Чрезвычайное высокомерие антивозрастной медицины». Биогеронтология . 10 (2): 223–228. дои : 10.1007/s10522-008-9170-6 . ПМИД 18726707 . S2CID 764136 .
- ^ Ольшанский С.Дж., Хейфлик Л., Карнс Б.А. (август 2002 г.). «Заявление о позиции по вопросу старения человека» . Журналы геронтологии. Серия А, Биологические и медицинские науки . 57 (8): В292–В297. CiteSeerX 10.1.1.541.3004 . дои : 10.1093/gerona/57.8.B292 . ПМИД 12145354 .
- ^ Уорнер Х., Андерсон Дж., Остад С. и др. (ноябрь 2005 г.). «Научный факт и программа SENS. Чего мы можем разумно ожидать от исследований старения?» . Отчеты ЭМБО . 6 (11): 1006–1008. дои : 10.1038/sj.embor.7400555 . ПМК 1371037 . ПМИД 16264422 .
{{cite journal}}: CS1 maint: переопределенная настройка ( ссылка )