Бета-лактамазы
| Сериновая бета-лактамаза | |||
|---|---|---|---|
 Структура Streptomyces albus бета-лактамазы | |||
| Идентификаторы | |||
| Символ | β-лактамазный домен | ||
| Пфам | PF00144 | ||
| Пфам Клан | CL0013 | ||
| ИнтерПро | ИПР001466 | ||
| PROSITE | PS00146 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 56601 / SCOPe / СУПФАМ | ||
| |||
| Металло-бета-лактамазы | |||
|---|---|---|---|
 Гомотетрамер металло-бета-лактамазы L1, Stenotropomonas | |||
| Идентификаторы | |||
| Символ | ? | ||
| Пфам | PF00753 | ||
| Пфам Клан | CL0381 | ||
| ИнтерПро | ИПР001279 | ||
| |||
| β-лактамаза | |||
|---|---|---|---|
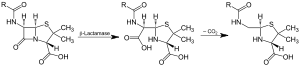 Действие β-лактамазы и декарбоксилирование интермедиата | |||
| Идентификаторы | |||
| Номер ЕС. | 3.5.2.6 | ||
| Номер CAS. | 9073-60-3 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||


Бета-лактамазы ( β-лактамазы ) — это ферменты ( КФ 3.5.2.6 ), продуцируемые бактериями , которые обеспечивают множественную устойчивость к бета-лактамным антибиотикам, таким как пенициллины , цефалоспорины , цефамицины , монобактамы и карбапенемы ( эртапенем ), хотя карбапенемы относительно устойчивы к бета-лактамазы. Бета-лактамазы обеспечивают устойчивость к антибиотикам, разрушая структуру антибиотиков . Все эти антибиотики имеют общий элемент в своей молекулярной структуре: четырехатомное кольцо, известное как бета-лактамное (β-лактамное) кольцо. В результате гидролиза фермент лактамаза разрывает кольцо β-лактама, деактивируя антибактериальные свойства молекулы.
Бета-лактамазы, продуцируемые грамотрицательными бактериями, обычно секретируются, особенно при наличии в окружающей среде антибиотиков. [ 1 ]
Структура
[ редактировать ]Структура сериновой β-лактамазы (SBL) Streptomyces представлена 1BSG . Альфа-бета-складка ( InterPro : IPR012338 ) напоминает складку DD-транспептидазы , из которой, как полагают, произошел фермент. β-лактамные антибиотики связываются с DD-транспептидазами, ингибируя биосинтез клеточной стенки бактерий. Сериновые β-лактамазы по сходству последовательностей группируются на типы A, C и D.
Другой тип бета-лактамаз относится к типу металло («тип B»). Металло-бета-лактамазы (MBL) нуждаются в ионах металлов (1 или 2 Zn 2+ ионы [ 2 ] ) на их активном сайте из-за их каталитической активности. [ 3 ] Структура металло-бета-лактамазы Нью-Дели 1 представлена 6C89 . Он напоминает РНКазу Z , из которой, как полагают, он произошел.
Механизм действия
[ редактировать ]Два типа бета-лактамаз действуют на основе двух основных механизмов раскрытия β-лактамного кольца. [ 2 ]
SBL по структуре и механизму сходны с β-лактамными целевыми пенициллин-связывающими белками (PBP), которые необходимы для построения и модификации клеточной стенки. И SBL, и PBP ковалентно изменяют остаток серина в активном центре. Разница между PBP и SBL заключается в том, что последний генерирует свободный фермент и неактивный антибиотик за счет очень быстрого гидролиза промежуточного ацил-фермента. [ нужна ссылка ]
MBL используют Zn 2+ ионы для активации молекулы воды в месте связывания для гидролиза β-лактамного кольца. Хелаторы цинка недавно были исследованы как ингибиторы металло-β-лактамаз, поскольку они часто способны восстанавливать чувствительность к карбапенемам. [ 4 ]
Пенициллиназа
[ редактировать ]Пенициллиназа представляет собой особый тип β-лактамазы, проявляющий специфичность к пенициллинам , опять же за счет гидролиза β -лактамного кольца. Молекулярные массы различных пенициллиназ имеют тенденцию группироваться около 50 килодальтон.
Пенициллиназа была первой идентифицированной β-лактамазой. Впервые он был выделен Абрахамом и Чейном в 1940 году из кишечной палочки (грамотрицательной) еще до того, как пенициллин начал применяться в клинической практике. [ 5 ] но продукция пенициллиназы быстро распространилась на бактерии, которые ранее ее не производили или производили очень редко. Были разработаны устойчивые к пенициллиназе бета-лактамы, такие как метициллин , но в настоящее время широко распространена устойчивость даже к ним.
Резистентность грамотрицательных бактерий
[ редактировать ]Этот раздел может потребовать очистки Википедии , чтобы соответствовать стандартам качества . Конкретная проблема заключается в следующем: запутанное сочетание структурных и функциональных классификаций; нужен пояснительный параграф о том, что это за классы. ( сентябрь 2021 г. ) |
Среди грамотрицательных бактерий серьезную озабоченность вызывает появление устойчивости к цефалоспоринам расширенного спектра действия. Первоначально он появился у ограниченного числа видов бактерий ( E. cloacae , C. freundii , S. marcescens и P. aeruginosa ), которые могли мутировать с гиперпродукцией хромосомной β-лактамазы класса C. Несколько лет спустя у видов бактерий, не продуцирующих в природе ферменты AmpC ( K. pneumoniae , Salmonella spp., P. mirabilis ), появилась устойчивость за счет продукции ESBL TEM- или SHV-типа (бета-лактамаз расширенного спектра). Характерно, что такая устойчивость включала оксиимино- (например , цефтизоксим , цефотаксим , цефтриаксон и цефтазидим , а также оксииминомонобактам азтреонам ), но не 7-альфа-метоксицефалоспорины ( цефамицины ; другими словами, цефокситин и цефотетан ). ; блокировался такими ингибиторами, как клавуланат , сульбактам или тазобактам, и не включал карбапенемы и темоцилин . Хромосомно-опосредованные β-лактамазы AmpC представляют новую угрозу, поскольку они придают устойчивость к 7-альфа-метоксицефалоспоринам. цефамицины ), такие как цефокситин или цефотетан , но на них не влияют коммерчески доступные ингибиторы β-лактамаз и могут у штаммов с потерей поринов внешней мембраны обеспечивать устойчивость к карбапенемам. [ 6 ]
Бета-лактамазы расширенного спектра действия (ESBL)
[ редактировать ]Члены этого семейства обычно экспрессируют β-лактамазы (например, TEM-3, TEM-4, [ 7 ] и ШВ-2 [ 8 ] ), которые придают устойчивость к цефалоспоринам расширенного спектра (расширенного спектра действия). В середине 1980-х годов была обнаружена новая группа ферментов — β-лактамазы расширенного спектра действия (ESBL) (впервые обнаружены в 1979 году). [ 9 ] Распространенность бактерий, продуцирующих БЛРС, постепенно увеличивается в больницах неотложной помощи. [ 10 ] Распространенность среди населения в целом варьируется в зависимости от страны, например, примерно 6% в Германии. [ 11 ] и Франция, [ 12 ] 13% в Саудовской Аравии, [ 13 ] и 63% в Египте. [ 14 ] ESBL представляют собой бета-лактамазы, которые гидролизуют цефалоспорины расширенного спектра действия с оксиимино боковой цепью. К этим цефалоспоринам относятся цефотаксим , цефтриаксон и цефтазидим , а также оксиимино-монобактам азтреонам . Таким образом, ESBL придают множественную устойчивость к этим антибиотикам и родственным оксиимино-бета-лактамам. В типичных обстоятельствах они происходят от генов TEM-1, TEM-2 или SHV-1 в результате мутаций, которые изменяют конфигурацию аминокислот вокруг активного сайта этих β-лактамаз. Более широкий набор β-лактамных антибиотиков подвержен гидролизу этими ферментами. В последнее время описывается все большее число ESBL, не принадлежащих к линии TEM или SHV. [ 15 ] ESBL часто кодируются плазмидами. Плазмиды, ответственные за продукцию ESBL, часто несут гены, кодирующие устойчивость к другим классам лекарств (например, аминогликозидам). Таким образом, возможности применения антибиотиков при лечении микроорганизмов, продуцирующих БЛРС, крайне ограничены. Карбапенемы являются препаратами выбора при серьезных инфекциях, вызванных микроорганизмами, продуцирующими БЛРС, однако эртапенему ). недавно появились сообщения об изолятах, устойчивых к карбапенемам (в первую очередь к [ 16 ] Организмы, продуцирующие ESBL, могут оказаться чувствительными к некоторым цефалоспоринам расширенного спектра действия . Однако лечение такими антибиотиками связано с высокой частотой неудач. [ нужна ссылка ]
Типы
[ редактировать ]Бета-лактамазы ТЕМ (класс А)
[ редактировать ]ТЕМ-1 является наиболее часто встречающейся бета-лактамазой у грамотрицательных бактерий. До 90% устойчивости E. coli к ампициллину обусловлено выработкой ТЕМ-1. [ 17 ] Также ответственен за резистентность к ампициллину и пенициллину, которая наблюдается у H. influenzae и N. gonorrhoeae во все большем количестве. Хотя бета-лактамазы ТЕМ-типа чаще всего обнаруживаются в E. coli и K. pneumoniae , они все чаще обнаруживаются и у других видов грамотрицательных бактерий. Аминокислотные замены, ответственные за фенотип бета-лактамазы расширенного спектра (ESBL), группируются вокруг активного центра фермента и изменяют его конфигурацию, обеспечивая доступ к субстратам оксиимино-бета-лактама. Открытие активного центра для субстратов бета-лактамов также обычно повышает чувствительность фермента к ингибиторам β-лактамаз, таким как клавулановая кислота. Одиночные аминокислотные замены в положениях 104, 164, 238 и 240 приводят к фенотипу ESBL, но ESBL с самым широким спектром обычно имеют более одной аминокислотной замены. На основании различных комбинаций изменений в настоящее время описано 140 ферментов ТЕМ-типа. TEM-10, TEM-12 и TEM-26 являются одними из наиболее распространенных в США. [ 18 ] [ 19 ] [ 20 ] Термин ТЕМ происходит от имени афинского пациента (Темониера), у которого в 1963 году был выделен изолят. [ 21 ]
Бета-лактамазы SHV (класс А)
[ редактировать ]SHV-1 на 68 процентов разделяет аминокислоты с TEM-1 и имеет аналогичную общую структуру. Бета-лактамазы SHV-1 чаще всего обнаруживаются у K. pneumoniae и ответственны за до 20% плазмидно-опосредованной устойчивости к ампициллину у этого вида. ESBL этого семейства также имеют аминокислотные изменения вокруг активного центра, чаще всего в положениях 238 или 238 и 240. Известно более 60 разновидностей SHV. ШВ-5 и ШВ-12 являются одними из наиболее распространенных. [ 18 ] Инициалы означают «переменная сульфгидрильного реагента». [ 22 ]
Бета-лактамазы CTX-M (класс А)
[ редактировать ]Эти ферменты получили свое название из-за их большей активности в отношении цефотаксима, чем других субстратов оксиимино-бета-лактамов (например, цефтазидима , цефтриаксона или цефепима ). Они не возникают в результате мутаций, а представляют собой примеры приобретения плазмидами генов бета-лактамаз, обычно встречающихся в хромосомах видов Kluyvera , группы редко патогенных комменсальных организмов. Эти ферменты не очень тесно связаны с бета-лактамазами TEM или SHV, поскольку они лишь примерно на 40% идентичны этим двум обычно выделяемым бета-лактамазам. Более 172 [ 23 ] Ферменты СТХ-М в настоящее время известны. Несмотря на свое название, некоторые из них более активны в отношении цефтазидима , чем цефотаксима . Они широко описаны среди видов Enterobacteriaceae , главным образом E. coli и K. pneumoniae. Обнаруженные в 1980-х годах, они с начала 2000-х годов распространились и в настоящее время являются преобладающим типом БЛРС в мире. Обычно их объединяют в пять групп на основе гомологии секвенирования; CTX-M-1, CTX-M-2, CTX-M-8, CTX-M-9 и CTX-M-25. CTX-M-15 (принадлежит к кластеру CTX-M-1) является наиболее распространенным геном CTX-M. [ 24 ] Было обнаружено , что пример бета-лактамазы CTX-M-15 вместе с IS Ecp1 транспонирован в хромосому Klebsiella pneumoniae ATCC BAA-2146. [ 25 ] Инициалы означают «Цефотаксим-Мюнхен». [ 26 ]
Бета-лактамазы OXA (класс D)
[ редактировать ]Бета-лактамазы OXA долгое время считались менее распространенной, но также опосредованной плазмидами разновидностью бета-лактамаз, которая могла гидролизовать оксациллин и родственные ему антистафилококковые пенициллины. Эти бета-лактамазы отличаются от ферментов TEM и SHV тем, что они принадлежат к молекулярному классу D и функциональной группе 2d. Бета-лактамазы OXA-типа придают устойчивость к ампициллину и цефалотину и характеризуются высокой гидролитической активностью в отношении оксациллина и клоксациллина , а также тем фактом, что они плохо ингибируются клавулановой кислотой . Аминокислотные замены в ферментах OXA также могут приводить к фенотипу ESBL. В то время как большинство ESBL были обнаружены у E. coli , K. pneumoniae и других Enterobacteriaceae , ESBL OXA-типа были обнаружены главным образом у P. aeruginosa . ESBL OXA-типа были обнаружены в основном у изолятов Pseudomonas aeruginosa из Турции и Франции. Семейство бета-лактамаз OXA изначально было создано как фенотипическая, а не генотипическая группа для нескольких бета-лактамаз, которые имели специфический профиль гидролиза. Следовательно, среди некоторых членов этого семейства гомология последовательностей составляет всего 20%. Однако недавние дополнения к этому семейству демонстрируют некоторую степень гомологии с одним или несколькими существующими членами семейства бета-лактамаз OXA. Некоторые из них придают устойчивость преимущественно к цефтазидиму, но OXA-17 придает большую устойчивость к цефотаксиму и цефепиму, чем к цефтазидиму.
Другие
[ редактировать ]Другие плазмид-опосредованные ESBL, такие как бета-лактамазы PER, VEB, GES и IBC, были описаны, но они встречаются редко и обнаруживаются главным образом у P. aeruginosa и в ограниченном числе географических участков. PER-1 в изолятах в Турции, Франции и Италии; ВЭБ-1 и ВЭБ-2 у штаммов Юго-Восточной Азии; и GES-1, GES-2 и IBC-2 в изолятах из Южной Африки, Франции и Греции. PER-1 также распространен у мультирезистентных видов Acinetobacter в Корее и Турции. Некоторые из этих ферментов также обнаружены у Enterobacteriaceae, тогда как другие необычные ESBL (такие как BES-1, IBC-1, SFO-1 и TLA-1) обнаружены только у Enterobacteriaceae.
Уход
[ редактировать ]Хотя раньше организмы, продуцирующие БЛРС, были связаны с больницами и учреждениями ухода, в настоящее время эти организмы все чаще встречаются в обществе. CTX-M-15-положительная кишечная палочка является причиной внебольничных инфекций мочевыводящих путей в Великобритании. [ 27 ] и имеют тенденцию быть устойчивыми ко всем пероральным β-лактамным антибиотикам, а также к хинолонам и сульфонамидам . Варианты лечения могут включать нитрофурантоин , фосфомицин , мециллинам и хлорамфеникол . В отчаянии эртапенема или гентамицина можно также использовать инъекции один раз в день.
Ингибитор-резистентные β-лактамазы
[ редактировать ]Хотя устойчивые к ингибиторам β-лактамазы не являются ESBL, их часто обсуждают с ESBL, поскольку они также являются производными классических ферментов типа TEM или SHV. Этим ферментам сначала было присвоено обозначение IRT для устойчивой к ингибитору TEM β-лактамазы; однако впоследствии все они были переименованы в числовые обозначения TEM. Существует по крайней мере 19 различных устойчивых к ингибиторам TEM β-лактамаз. Устойчивые к ингибиторам TEM β-лактамазы были обнаружены главным образом в клинических изолятах E. coli , а также в некоторых штаммах K. pneumoniae , Klebsiella oxytoca , P. mirabilis и Citrobacter freundii . Хотя устойчивые к ингибиторам варианты ТЕМ устойчивы к ингибированию клавулановой кислотой и сульбактамом , тем самым проявляя клиническую устойчивость к комбинациям бета-лактам-ингибитор лактамаз амоксициллин - клавуланат ( ко-амоксиклав ), тикарциллин - клавуланат ( ко-тикарклав ) и ампициллин/сульбактам , они обычно остаются чувствительными к ингибированию тазобактамом , а затем и комбинацией пиперациллин/тазобактам , [ нужна ссылка ] хотя сопротивление было описано. Это уже не преимущественно европейская эпидемиология, она часто встречается в северных частях Америки, и ее следует проверять при комплексных ИМП. [ 19 ]
β-лактамазы AmpC-типа (класс C)
[ редактировать ]β-лактамазы типа AmpC обычно выделяют из грамотрицательных бактерий, устойчивых к цефалоспоринам расширенного спектра действия. β-лактамазы AmpC (также называемые классом C или группой 1) обычно кодируются на хромосоме многих грамотрицательных бактерий, включая виды Citrobacter , Serratia и Enterobacter , где их экспрессия обычно индуцируется ; он также может встречаться на Escherichia coli , но обычно не индуцируется, хотя может иметь гиперэкспрессию. β-лактамазы типа AmpC также могут переноситься на плазмидах. [ 6 ] β-лактамазы AmpC, в отличие от ESBL, гидролизуют цефалоспорины широкого и расширенного спектра действия (цефамицины, а также оксиимино-β-лактамы), но обычно не ингибируются ингибиторами β-лактамаз клавулановой кислотой и тазобактамом , тогда как авибактам может сохранять ингибирующее действие. активность против этого класса β-лактамаз. [ 28 ] β-лактамазные организмы типа AmpC часто клинически группируются по аббревиатуре «SPACE»: Serratia , Pseudomonas или Proteus , Acinetobacter , Citrobacter и Enterobacter .
Карбапенемазы
[ редактировать ]Карбапенемы известны своей стабильностью к β-лактамазам AmpC и β-лактамазам расширенного спектра действия. Карбапенемазы представляют собой разнообразную группу β-лактамаз, которые активны не только против оксииминоцефалоспоринов и цефамицинов, но также и против карбапенемов. Азтреонам стабилен к металло-β-лактамазам. но многие производители IMP и VIM устойчивы благодаря другим механизмам. Ранее считалось, что карбапенемазы происходят только из классов A, B и D, но была описана карбапенемаза класса C.
Карбапенемазы IMP-типа (металло-β-лактамазы) (класс В)
[ редактировать ]Плазмидно-опосредованные карбапенемазы IMP-типа (IMP означает «активные на имипенеме»), 19 разновидностей которых в настоящее время известны, были установлены в Японии в 1990-х годах как у кишечных грамотрицательных организмов, так и у Pseudomonas и Acinetobacter видов . Ферменты ИМФ медленно распространялись в другие страны Дальнего Востока, о них сообщалось в Европе в 1997 году, а также они были обнаружены в Канаде и Бразилии.
VIM (металло-β-лактамаза, кодируемая веронским интеграном) (класс B)
[ редактировать ]Второе растущее семейство карбапенемаз, семейство VIM, было зарегистрировано в Италии в 1999 году и в настоящее время включает 10 членов, которые имеют широкое географическое распространение в Европе, Южной Америке и на Дальнем Востоке и были обнаружены в Соединенных Штатах. VIM-1 был обнаружен у P. aeruginosa в Италии в 1996 г.; с тех пор ВИМ-2 — ныне преобладающий вариант — неоднократно обнаруживался в Европе и на Дальнем Востоке; ВИМ-3 и -4 являются второстепенными вариантами ВИМ-2 и -1 соответственно.
Разнообразие аминокислотных последовательностей составляет до 10% в семействе VIM, 15% в семействе IMP и 70% между VIM и IMP. Тем не менее ферменты обоих семейств схожи. Оба связаны с интеграном, иногда внутри плазмид. Оба гидролизуют все β-лактамы, кроме монобактамов, и уклоняются от всех ингибиторов β-лактамов. Ферменты VIM являются одними из наиболее широко распространенных MBL: зарегистрировано >40 вариантов VIM. Биохимические и биофизические исследования показали, что варианты VIM имеют лишь небольшие различия в своих кинетических параметрах, но существенные различия в их термической стабильности и профилях ингибирования. [ 29 ]
OXA (оксациллиназа) группа β-лактамаз (класс D)
[ редактировать ]Группа OXA β-лактамаз встречается преимущественно у видов Acinetobacter и разделена на два кластера. OXA-карбапенемазы очень медленно гидролизуют карбапенемы in vitro , а высокие значения МИК, наблюдаемые у некоторых хозяев Acinetobacter (>64 мг/л), могут отражать вторичные механизмы. В клинических изолятах они иногда дополняются дополнительными механизмами устойчивости, такими как непроницаемость или отток. OXA-карбапенемазы также имеют тенденцию иметь пониженную гидролитическую эффективность по отношению к пенициллинам и цефалоспоринам. [ 30 ]
КПК ( карбапенемаза K. pneumoniae ) (класс А)
[ редактировать ]Некоторые ферменты класса А, в основном плазмид-опосредованные ферменты KPC, также являются эффективными карбапенемазами. Известны десять вариантов, от KPC-2 до KPC-11, которые отличаются одной или двумя аминокислотными заменами (KPC-1 был повторно секвенирован в 2008 году и оказался на 100% гомологичен опубликованным последовательностям KPC-2). КРС-1 был обнаружен в Северной Каролине, КРС-2 в Балтиморе и КРС-3 в Нью-Йорке. Они имеют лишь 45% гомологии с ферментами SME и NMC/IMI и, в отличие от них, могут кодироваться самотрансмиссивными плазмидами.
По состоянию на февраль 2009 г. [update] ) класса А Карбапенемаза Klebsiella pneumoniae ( KPC во всем мире была наиболее распространенной карбапенемазой и впервые была обнаружена в 1996 году в Северной Каролине , США. [ 31 ] В публикации 2010 года указывалось, что KPC, производящие Enterobacteriaceae, становятся обычным явлением в Соединенных Штатах. [ 32 ]
CMY (класс С)
[ редактировать ]Первая карбапенемаза класса С была описана в 2006 году и выделена из вирулентного штамма Enterobacter aerogenes . [ 33 ] Он переносится плазмидой pYMG-1 и поэтому может передаваться другим бактериальным штаммам. [ 34 ]
SME (ферменты Serratia marcescens), IMI (IMIпенем-гидролизующая β-лактамаза), NMC и CcrA
[ редактировать ]В целом они не имеют клинического значения.
CcrA (CfiA). Его ген встречается ок. 1–3% изолятов B. fragilis , но меньшее количество продуцирует фермент, поскольку экспрессия требует соответствующей миграции инсерционной последовательности. CcrA был известен до появления имипенема, и производители в последующем не продемонстрировали незначительного увеличения его количества.
NDM-1 (Нью-Дели металло-β-лактамаза) (класс B)
[ редактировать ]Первоначально описанный в Нью-Дели в 2009 году, этот ген в настоящее время широко распространен у Escherichia coli и Klebsiella pneumoniae из Индии и Пакистана. По состоянию на середину 2010 года бактерии, несущие NDM-1, были завезены в другие страны (включая США и Великобританию), скорее всего, из-за большого количества туристов, путешествующих по миру, которые могли подхватить штамм из окружающей среды. поскольку штаммы, содержащие ген NDM-1, были обнаружены в пробах окружающей среды в Индии. [ 35 ] У NDM есть несколько вариантов, имеющих разные свойства. [ 29 ]
Лечение ESBL/AmpC/карбапенемаз
[ редактировать ]Общий обзор
[ редактировать ]Как правило, изолят считается продуцентом БЛРС, если он обнаруживает in vitro чувствительность к цефамицинам ( цефокситин , цефотетан ), но устойчивость к цефалоспоринам третьего поколения и азтреонаму . Более того, следует подозревать эти штаммы, когда лечение этими агентами грамотрицательных инфекций не дает результатов, несмотря на сообщения о чувствительности in vitro . После обнаружения штамма, продуцирующего БЛРС, лаборатория должна сообщить о его «резистентности» ко всем пенициллинам, цефалоспоринам и азтреонаму, даже если он проверен (in vitro) как чувствительный. [ нужна ссылка ] Сопутствующая устойчивость к аминогликозидам и триметоприму - сульфаметоксазолу , а также высокая частота сосуществования устойчивости к фторхинолонам создают проблемы. Ингибиторы бета-лактамаз, такие как клавуланат , сульбактам и тазобактам , in vitro ингибируют большинство ESBL, но на клиническую эффективность комбинаций бета-лактамов/ингибиторов бета-лактамаз нельзя постоянно полагаться при лечении. Цефамицины ( цефокситин и цефотетан ) не гидролизуются большинством ESBL, но гидролизуются ассоциированной β-лактамазой типа AmpC. Кроме того, комбинации ингибиторов β-лактамов и β-лактамаз могут быть неэффективны против организмов, продуцирующих β-лактамазу типа AmpC. Иногда эти штаммы снижают экспрессию белков внешней мембраны, что делает их устойчивыми к цефамицинам. Исследования in vivo дали неоднозначные результаты в отношении K. pneumoniae , продуцирующего ESBL . ( Цефепим , цефалоспорин четвертого поколения, продемонстрировал стабильность in vitro в присутствии многих штаммов ESBL/AmpC.) В настоящее время карбапенемы в целом считаются предпочтительным средством для лечения инфекций, вызванных микроорганизмами, продуцирующими ESBL. Карбапенемы устойчивы к гидролизу, опосредованному ESBL, и проявляют превосходную активность in vitro против штаммов Enterobacteriaceae, экспрессирующих ESBL. [ нужна ссылка ]
По генам
[ редактировать ]ESBL
[ редактировать ]Штаммы, продуцирующие только ESBL, чувствительны к цефамицинам и карбапенемам in vitro и практически не проявляют инокуляционного эффекта от этих агентов.
Для организмов, продуцирующих ESBL типа TEM и SHV , очевидная in vitro чувствительность к цефепиму и пиперациллину/тазобактаму является обычным явлением, но оба препарата проявляют инокулятивный эффект со снижением чувствительности по мере увеличения размера инокулята с 10 5 до 10 7 организмы.
Штаммы с некоторыми ESBL типа CTX-M и OXA -типа устойчивы к цефепиму при тестировании, несмотря на использование стандартного инокулята.
Ингибитор-резистентные β-лактамазы
[ редактировать ]Хотя устойчивые к ингибиторам варианты ТЕМ устойчивы к ингибированию клавулановой кислотой и сульбактамом , тем самым демонстрируя клиническую устойчивость к комбинациям бета-лактам-ингибитор бета-лактамаз амоксициллин - клавуланат ( ко-амоксиклав ), тикарциллин - клавуланат и ампициллин/сульбактам , они остаются чувствительными к ингибированию тазобактамом , а затем и комбинацией пиперациллин/тазобактам .
АмпК
[ редактировать ]Штаммы, продуцирующие AmpC, обычно устойчивы к оксиимино-бета-лактамам и цефамицинам и чувствительны к карбапенемам ; однако пониженная экспрессия поринов также может сделать такой штамм устойчивым к карбапенемам.
Карбапенемазы
[ редактировать ]Штаммы с карбапенемазами IMP-, VIM- и OXA -типов обычно остаются восприимчивыми. Устойчивость к небета-лактамным антибиотикам часто встречается у штаммов, продуцирующих любой из этих ферментов, поэтому альтернативные варианты небета-лактамной терапии необходимо определять путем прямого тестирования чувствительности. устойчивость к фторхинолонам и аминогликозидам Особенно высока .
По видам
[ редактировать ]Эшерихия коли или клебсиелла
[ редактировать ]При инфекциях, вызванных видами Escherichia coli или Klebsiella , продуцирующими ESBL , лечение имипенемом или меропенемом было связано с лучшими результатами с точки зрения выживаемости и бактериологического клиренса. Цефепим и пиперациллин/тазобактам оказались менее успешными. Цефтриаксон , цефотаксим и цефтазидим оказывались неэффективными еще чаще, несмотря на чувствительность организма к антибиотику in vitro . В нескольких сообщениях документально подтверждена неэффективность терапии цефамицином из-за резистентности из-за потери поринов. Некоторые пациенты ответили на терапию аминогликозидами или хинолонами , но в недавнем сравнении ципрофлоксацина и имипенема при бактериемии, вызванной ESBL-продуцирующим K. pneumoniae , имипенем дал лучший результат.
синегнойная палочка
[ редактировать ], было немного Pseudomonas aeruginosa Клинических исследований по определению оптимальной терапии инфекций, вызванных БЛРС, продуцирующими штаммы .
Использование в качестве фармацевтического средства
[ редактировать ]В 1957 году, на фоне беспокойства по поводу аллергических реакций на пенициллинсодержащие антибиотики, бета-лактамазы продавались в качестве антидота под торговой маркой нейтрапен. [ 36 ] Было высказано предположение, что расщепление пенициллина ферментом поможет устранить аллергическую реакцию. [ 37 ] Хотя он не был полезен при остром анафилактическом шоке, он показал положительные результаты в случаях крапивницы и болей в суставах, предположительно вызванных аллергией на пенициллин. [ 38 ] [ 39 ] Его использование было предложено в педиатрических случаях, когда аллергия на пенициллин была обнаружена при введении полиомиелитной вакцины, в которой пенициллин использовался в качестве консерванта. [ 40 ] Однако у некоторых пациентов возникла аллергия на нейтрапен. [ 41 ] [ 42 ] Больница Олбани исключила его из своего формуляра в 1960 году, всего через два года после добавления, сославшись на неиспользование. [ 43 ] Некоторые исследователи продолжали использовать его в экспериментах по резистентности к пенициллину даже в 1972 году. [ 44 ] В 1997 году компания 3M Pharmaceuticals добровольно отозвала его с американского рынка. [ 45 ]
Обнаружение
[ редактировать ]Ферментативную активность бета-лактамаз можно обнаружить с помощью нитроцефина , хромогенного субстрата цефалоспорина , который меняет цвет с желтого на красный при гидролизе, опосредованном бета-лактамазой. [ 46 ]
Скрининг бета-лактамаз расширенного спектра (ESBL) можно проводить с использованием дисковой диффузии. Используются диски с цефподоксимом, цефтазидимом, азтреонамом, цефотаксимом и/или цефтриаксоном. [ 47 ]
Эволюция
[ редактировать ]Бета-лактамазы — древние бактериальные ферменты. Металло-β-лактамазы («класс B») структурно схожи с РНКазой Z и, возможно, произошли от нее. Предполагается, что из трех подклассов B1, B2 и B3 B1 и B2 возникли около одного миллиарда лет назад , тогда как B3, по-видимому, возник независимо, возможно, до расхождения грамположительных и грамотрицательных эубактерий около двух миллиардов лет назад. много лет назад. [ 48 ] PNGM-1 (Металло-β-лактамаза-1 Папуа-Новой Гвинеи) обладает активностью как металло-β-лактамазы (MBL), так и тРНКазы Z, что позволяет предположить, что PNGM-1, как полагают, произошел от тРНКазы Z и что MBL B3 Активность PNGM-1 является беспорядочной деятельностью, и считается, что MBL подкласса B3 развились благодаря активности PNGM-1. [ 49 ] Подклассы B1 и B3 получили дальнейшее подразделение. [ 50 ]
Сериновые бета-лактамазы (классы A, C и D), по-видимому, произошли от DD-транспептидаз , которые представляют собой пенициллин-связывающие белки, участвующие в биосинтезе клеточной стенки и, как таковые, являются одной из основных мишеней бета-лактамных антибиотиков. [ 51 ] Эти три класса демонстрируют необнаружимое сходство последовательностей друг с другом, но их все же можно сравнивать с использованием структурной гомологии. Группы A и D являются сестринскими таксонами, а группа C разошлась раньше A и D. [ 52 ] Эти ферменты на основе серина, как и беталактамазы группы B, имеют древнее происхождение и, как предполагается, появились около двух миллиардов лет назад. [ 53 ]
В частности, предполагается, что группа OXA (класса D) эволюционировала на хромосомах и перемещалась в плазмиды по крайней мере в двух отдельных случаях. [ 54 ]
Этимология
[ редактировать ]«β» ( бета ) относится к азота положению на втором атоме углерода в кольце. Лактам представляет собой смесь лактона (от латинского Lactis — молоко , поскольку молочную кислоту выделяли из простокваши) и амида . Суффикс -аза , обозначающий фермент, происходит от диастазы (от греческого диастаза , «разделение»), первого фермента, открытого в 1833 году Пайеном и Персосом. [ 55 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Ной ХК (июнь 1969 г.). «Влияние расположения бета-лактамаз в Escherichia coli на синергию пенициллина» . Прикладная микробиология . 17 (6): 783–6. дои : 10.1128/АЕМ.17.6.783-786.1969 . ПМЦ 377810 . ПМИД 4894721 .
- ^ Перейти обратно: а б Ротондо CM, Райт Г.Д. (октябрь 2017 г.). «Ингибиторы металло-β-лактамаз». Современное мнение в микробиологии . 39 : 96–105. дои : 10.1016/j.mib.2017.10.026 . ПМИД 29154026 .
- ^ Ши С., Чэнь Дж., Кан Х., Шэнь Х., Лао Х., Чжэн Х. (август 2019 г.). «Подходы к открытию ингибиторов металло-β-лактамаз: обзор». Химическая биология и дизайн лекарств . 94 (2): 1427–1440. дои : 10.1111/cbdd.13526 . ПМИД 30925023 . S2CID 85566136 .
- ^ Принсипе Л., Веккьо Г., Шихан Г., Кавана К., Моррони Г., Виаджи В. и др. (1 октября 2020 г.). «Хелаторы цинка как адъюванты карбапенемов для бактерий, продуцирующих металло-β-лактамазу: оценка in vitro и in vivo» . Микробная лекарственная устойчивость . 26 (10): 1133–1143. дои : 10.1089/mdr.2020.0037 . ISSN 1076-6294 . ПМИД 32364820 . S2CID 218504647 .
- ^ Авраам Э.П., Чейн Э (1940). «Фермент бактерий, способный разрушать пенициллин» . Природа . 46 (3713): 837. Бибкод : 1940Natur.146..837A . дои : 10.1038/146837a0 . S2CID 4070796 .
- ^ Перейти обратно: а б Филиппон А., Арлет Г., Джейкоби Г.А. (январь 2002 г.). «Плазмидно-определяемые бета-лактамазы типа AmpC» . Антимикробные средства и химиотерапия . 46 (1): 1–11. doi : 10.1128/AAC.46.1.1-11.2002 . ПМК 126993 . ПМИД 11751104 .
- ^ «Бета-лактамазы класса А Эмблера: ТЕА» . База данных по бета-лактамазам (BLDB . Архивировано из оригинала 11 февраля 2022 г. Проверено 11 февраля 2022 г. ).
- ^ «Бета-лактамазы класса А по Амблеру: SHV» . База данных по бета-лактамазам (BLDB) . Архивировано из оригинала 11 февраля 2022 года . Проверено 11 февраля 2022 г.
- ^ Сандерс CC, Сандерс WE (июнь 1979 г.). «Появление устойчивости к цефамандолу: возможная роль цефокситин-индуцируемых бета-лактамаз» . Антимикробные средства и химиотерапия . 15 (6): 792–797. дои : 10.1128/AAC.15.6.792 . ПМК 352760 . ПМИД 314270 .
- ^ Спадафино Дж.Т., Коэн Б., Лю Дж., Ларсон Э. (2014). «Временные тенденции и факторы риска развития Escherichia coli, продуцирующей бета-лактамазы расширенного спектра действия, у взрослых с катетер-ассоциированными инфекциями мочевыводящих путей» . Устойчивость к противомикробным препаратам и инфекционный контроль . 3 (1): 39. дои : 10.1186/s13756-014-0039-y . ПМК 4306238 . ПМИД 25625011 .
- ^ Симанзик С., Хилленбранд Дж., Стаселович Л., Грей Дж.К., Фридрих А.В., Пульц М. и др. (декабрь 2021 г.). «Новые данные об основных факторах риска ректального носительства энтеробактерий, продуцирующих бета-лактамазы расширенного спектра, среди населения в Нижней Саксонии, Германия» . Журнал прикладной микробиологии . 132 (4): 3256–3264. дои : 10.1111/jam.15399 . ПМИД 34856042 . S2CID 244854840 .
- ^ Николя-Шаноин М.Х., Грусон С., Бялек-Давенет С., Бертран Х, Томас-Жан Ф., Берт Ф. и др. (март 2013 г.). «10-кратное увеличение (2006-11) количества здоровых людей с фекальным носительством Escherichia coli, продуцирующего β-лактамазы расширенного спектра действия, в парижском центре обследования». Журнал антимикробной химиотерапии . 68 (3): 562–568. дои : 10.1093/jac/dks429 . ПМИД 23143897 .
- ^ Кадер А.А., Камат К.А. (2009). «Фекальное носительство бактерий, продуцирующих бета-лактамазы расширенного спектра действия, в обществе». Восточно-Средиземноморский журнал здравоохранения . 15 (6): 1365–1370. ПМИД 20218126 .
- ^ Вальверде А., Гриль Ф., Кок ТМ, Пинтадо В., Бакеро Ф., Кантон Р. и др. (август 2008 г.). «Высокий уровень колонизации кишечника микроорганизмами, продуцирующими бета-лактамазы расширенного спектра действия, при бытовых контактах с инфицированными внебольничными пациентами» . Журнал клинической микробиологии . 46 (8): 2796–2799. дои : 10.1128/JCM.01008-08 . ПМК 2519510 . ПМИД 18562591 .
- ^ Эмери CL, Уэймут, Лос-Анджелес (август 1997 г.). «Обнаружение и клиническое значение бета-лактамаз расширенного спектра в медицинском центре третичного уровня» . Журнал клинической микробиологии . 35 (8): 2061–2067. doi : 10.1128/JCM.35.8.2061-2067.1997 . ПМК 229903 . ПМИД 9230382 .
- ^ Грундманн Х., Ливермор Д.М., Гиске К.Г., Кантон Р., Россолини Г.М., Кампос Дж. и др. (ноябрь 2010 г.). «Нечувствительные к карбапенемам энтеробактерии в Европе: выводы совещания национальных экспертов» . Евронаблюдение . 15 (46). doi : 10.2807/ese.15.46.19711-en . hdl : 10400.18/206 . ПМИД 21144429 .
- ^ Кукси Р., Свенсон Дж., Кларк Н., Гей Э., Торнсберри С. (май 1990 г.). «Схемы и механизмы резистентности к бета-лактамам среди изолятов Escherichia coli из больниц США» . Антимикробные средства и химиотерапия . 34 (5): 739–45. дои : 10.1128/AAC.34.5.739 . ПМК 171683 . ПМИД 2193616 .
- ^ Перейти обратно: а б Патерсон Д.Л., Худжер К.М., Худжер А.М., Йейзер Б., Бономо М.Д., Райс Л.Б. и др. (ноябрь 2003 г.). «Бета-лактамазы расширенного спектра в изолятах кровотока Klebsiella pneumoniae из семи стран: доминирование и широкая распространенность бета-лактамаз SHV- и CTX-M-типов» . Антимикробные средства и химиотерапия . 47 (11): 3554–60. doi : 10.1128/AAC.47.11.3554-3560.2003 . ПМЦ 253771 . ПМИД 14576117 .
- ^ Перейти обратно: а б Брэдфорд, Пенсильвания (октябрь 2001 г.). «Бета-лактамазы расширенного спектра действия в 21 веке: характеристика, эпидемиология и выявление этой важной угрозы устойчивости» . Обзоры клинической микробиологии . 14 (4): 933–51, оглавление. дои : 10.1128/CMR.14.4.933-951.2001 . ПМК 89009 . ПМИД 11585791 .
- ^ Джейкоби Г.А., Муньос-Прайс Л.С. (январь 2005 г.). «Новые бета-лактамазы». Медицинский журнал Новой Англии . 352 (4): 380–91. дои : 10.1056/NEJMra041359 . ПМИД 15673804 .
- ^ Руис Дж (2018). «Этимология: ТЭМ» . Новые инфекционные заболевания . 24 (4): 709. doi : 10.3201/eid2404.et2404 . ПМЦ 5875283 .
- ^ Лиакопулос А., Мевиус Д., Чеккарелли Д. (5 сентября 2016 г.). «Обзор β-лактамаз расширенного спектра SHV: игнорируемые, но повсеместно распространенные» . Границы микробиологии . 7 : 1374. дои : 10.3389/fmicb.2016.01374 . ПМК 5011133 . ПМИД 27656166 .
- ^ Рамадан А.А., Абдельазиз Н.А., Амин М.А., Азиз Р.К. (март 2019 г.). «Новые варианты blaCTX-M и корреляции генотипа и фенотипа среди клинических изолятов Escherichia coli, продуцирующих бета-лактамазы расширенного спектра» . Научные отчеты . 9 (1): 4224. Бибкод : 2019НатСР...9.4224Р . дои : 10.1038/s41598-019-39730-0 . ПМК 6414621 . ПМИД 30862858 . S2CID 75136447 .
- ^ Кастанейра М. (3 сентября 2021 г.). «Бетамазы расширенного спектра действия: обновленная информация об их характеристиках, эпидемиологии и обнаружении» . Журнал антимикробной химиотерапии . 3 (3): dlab092. doi : 10.1093/jacamr/dlab092 . ПМЦ 8284625 . ПМИД 34286272 .
- ^ Хадсон СМ, Бент З.В., Мигер Р.Дж., Уильямс К.П. (7 июня 2014 г.). «Детерминанты резистентности и мобильные генетические элементы штамма Klebsiella pneumoniae, кодирующего NDM-1» . ПЛОС ОДИН . 9 (6): e99209. Бибкод : 2014PLoSO...999209H . дои : 10.1371/journal.pone.0099209 . ПМК 4048246 . ПМИД 24905728 .
- ^ Кантон Р., Гонсалес-Альба Х.М., Галан Х.К. (2012). «Ферменты СТХ-М: происхождение и распространение» . Границы микробиологии . 3 : 110. дои : 10.3389/fmicb.2012.00110 . ISSN 1664-302X . ПМК 3316993 . ПМИД 22485109 .
- ^ Вудфорд Н., Уорд Э., Кауфман М.Э. и др. «Молекулярная характеристика изолятов Escherichia coli , продуцирующих β-лактамазу расширенного спектра CTX-M-15 (ESBL) в Соединенном Королевстве» (PDF) . Агентство по охране здоровья. Архивировано из оригинала (PDF) 15 июня 2007 года . Проверено 19 ноября 2006 г.
- ^ «Клинический обзор, NDA 206494, цефтазидим-авибактам» (PDF) . Управление по контролю за продуктами и лекарствами (FDA). 18 февраля 2015 г. Архивировано (PDF) из оригинала 28 февраля 2017 г. . Проверено 14 ноября 2023 г.
- ^ Перейти обратно: а б Макена А., Дюзгюн А.О., Брем Дж., Макдонаф М.А., Рыдзик А.М., Аббуд М.И. и др. (декабрь 2015 г.). «Сравнение вариантов металло-β-лактамаз (VIM), переносимых интеграном Вероны, выявляет различия в профилях стабильности и ингибирования» . Антимикробные средства и химиотерапия . 60 (3): 1377–84. дои : 10.1128/AAC.01768-15 . ПМЦ 4775916 . ПМИД 26666919 .
- ^ Сантильяна Э., Бесейру А., Бу Дж., Ромеро А. (март 2007 г.). «Кристаллическая структура карбапенемазы OXA-24 позволяет лучше понять механизм гидролиза карбапенема» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (13): 5354–9. Бибкод : 2007PNAS..104.5354S . дои : 10.1073/pnas.0607557104 . ПМЦ 1838445 . ПМИД 17374723 .
- ^ Нордманн П., Кузон Дж., Наас Т. (апрель 2009 г.). «Реальная угроза бактерий, продуцирующих карбапенемазы Klebsiella pneumoniae». «Ланцет». Инфекционные болезни . 9 (4): 228–36. дои : 10.1016/S1473-3099(09)70054-4 . ПМИД 19324295 .
- ^ Кузон Дж., Наас Т., Нордманн П. (февраль 2010 г.). «[Карбапенемазы KPC: что поставлено на карту в клинической микробиологии?]». Pathologie-Biologie (на французском языке). 58 (1): 39–45. дои : 10.1016/j.patbio.2009.07.026 . ПМИД 19854586 .
- ^ Ким Дж.И., Юнг Х.И., Ан Ю.Дж., Ли Дж.Х., Ким С.Дж., Чон Ш. и др. (май 2006 г.). «Структурная основа расширенного спектра субстратов CMY-10, бета-лактамазы класса C, кодируемой плазмидой» . Молекулярная микробиология . 60 (4): 907–16. дои : 10.1111/j.1365-2958.2006.05146.x . ПМИД 16677302 . S2CID 44982704 .
- ^ Ли Дж.Х., Юнг Х.И., Юнг Дж.Х., Пак Дж.С., Ан Дж.Б., Чжон Ш. и др. (2004). «Распространение переносимой бета-лактамазы типа AmpC (CMY-10) в корейской больнице». Микробная лекарственная устойчивость . 10 (3): 224–30. дои : 10.1089/mdr.2004.10.224 . ПМИД 15383166 .
- ^ Уолш Т.Р., Уикс Дж., Ливермор Д.М., Толеман М.А. (май 2011 г.). «Распространение NDM-1-положительных бактерий в окружающей среде Нью-Дели и его последствия для здоровья человека: исследование распространенности в окружающей среде». «Ланцет». Инфекционные болезни . 11 (5): 355–62. дои : 10.1016/S1473-3099(11)70059-7 . ПМИД 21478057 .
- ^ «НОВЫЕ ПРЕПАРАТЫ БОРЬБЫ С ПЕНИЦИЛЛИНОМ; Эксперт по антибиотикам считает, что нейтрапен эффективен при инъекционных побочных эффектах. РОСТ АЛЛЕРГИИ, ИЗУЧЕННЫЙ В США Помощник сообщает об увеличении реакции на пенициллин и подобные вещества, серьезные реакции. Несколько серьезных реакций (опубликовано в 1957 г.)» . Нью-Йорк Таймс . 4 октября 1957 г. ISSN 0362-4331 . Архивировано из оригинала 31 января 2022 года . Проверено 24 декабря 2020 г.
- ^ Хайман А.Л. (февраль 1959 г.). «Анафилактический шок после терапии пенициллиназой». Журнал Американской медицинской ассоциации . 169 (6): 593–4. дои : 10.1001/jama.1959.73000230003011a . ПМИД 13620512 .
- ^ Фридлендер С. (апрель 1959 г.). «Пенициллиназа в лечении аллергических реакций на пенициллин». Журнал аллергии . 30 (2): 181–7. дои : 10.1016/0021-8707(59)90087-5 . ПМИД 13630649 .
- ^ Американская академия педиатрии (1 октября 1958 г.). «Новая концепция лечения реакций пенициллина: использование пенициллиназы» . Педиатрия . 22 (4): 658. doi : 10.1542/peds.22.4.658 . ISSN 0031-4005 . S2CID 245066458 . Архивировано из оригинала 28 сентября 2018 года . Проверено 24 декабря 2020 г.
- ^ Циммерман MC (август 1958 г.). «Подтвержденная пенициллиназой аллергия на пенициллин в вакцине против полиомиелита». Журнал Американской медицинской ассоциации . 167 (15): 1807–9. дои : 10.1001/jama.1958.02990320001001 . ПМИД 13563181 .
- ^ Вайс Р.К., Крепеа С.Б. (июль 1959 г.). «Развитие чувствительности к пенициллиназе после ее использования в реакции на пенициллин». Журнал аллергии . 30 (4): 337–41. дои : 10.1016/0021-8707(59)90041-3 . ПМИД 13664435 .
- ^ Комитет Сената Конгресса США по судебной власти (1961 г.). Антимонопольный закон о фармацевтической промышленности: слушания в Подкомитете по антимонопольному законодательству и монополии Комитета судебной власти Сената США, Восемьдесят седьмой Конгресс, первая [-вторая] сессия, в соответствии с рез. 52 по S. 1552, законопроект о внесении поправок и дополнений в антимонопольное законодательство в отношении производства и распространения наркотиков и для других целей . Типография правительства США.
- ^ Комитет Сената Конгресса США по судебному подкомитету по антимонопольному законодательству и монополиям (1957 г.). Администрируемые цены: продолжение] п.25. Регулируемые цены в фармацевтической промышленности (Антибиотики – Приложение А). 1961. С. 14201-15329 . Типография правительства США.
- ^ Линдстрем Э.Б., Нордстрем К. (февраль 1972 г.). «Автоматизированный метод определения пенициллинов, цефалоспоринов и пенициллиназ» . Антимикробные средства и химиотерапия . 1 (2): 100–6. дои : 10.1128/aac.1.2.100 . ПМК 444176 . ПМИД 4618452 .
- ^ «Федеральный реестр, том 62, выпуск 229 (пятница, 28 ноября 1997 г.)» . www.govinfo.gov . Архивировано из оригинала 23 октября 2023 года . Проверено 24 декабря 2020 г.
- ^ О'Каллаган CH, Моррис А., Кирби С.М., Шинглер А.Х. (апрель 1972 г.). «Новый метод обнаружения бета-лактамаз с использованием хромогенного цефалоспоринового субстрата» . Антимикробные средства и химиотерапия . 1 (4): 283–8. дои : 10.1128/AAC.1.4.283 . ПМК 444209 . ПМИД 4208895 .
- ^ Рават Д., Наир Д. (2010). «β-лактамазы расширенного спектра действия у грамотрицательных бактерий» . Журнал глобальных инфекционных заболеваний . 2 (3): 263–274. дои : 10.4103/0974-777X.68531 . ISSN 0974-777X . ПМЦ 2946684 . ПМИД 20927289 .
- ^ Холл Б.Г., Салипанте С.Дж., Барлоу М. (июль 2004 г.). «Независимое происхождение металло-бета-лактамаз подгруппы Bl + B2 и подгруппы B3». Журнал молекулярной эволюции . 59 (1): 133–41. дои : 10.1007/s00239-003-2572-9 . ПМИД 15383916 . S2CID 30833168 .
- ^ Ли Дж.Х., Такахаши М., Чон Дж.Х., Кан Л.В., Секи М., Пак К.С. и др. (2019). «Двойная активность PNGM-1 указывает на эволюционное происхождение металло- β- лактамаз подкласса B3: молекулярное и эволюционное исследование» . Новые микробы и инфекции . 8 (1): 1688–1700. дои : 10.1080/22221751.2019.1692638 . ПМК 6882493 . ПМИД 31749408 .
- ^ Берглунд Ф., Джоннинг А., Ларссон Д.Г., Кристианссон Э. (январь 2021 г.). «Обновленная филогения металло-β-лактамаз». Журнал антимикробной химиотерапии . 76 (1): 117–123. дои : 10.1093/jac/dkaa392 . ПМИД 33005957 .
- ^ ИнтерПро : IPR012338
- ^ Холл Б.Г., Барлоу М. (сентябрь 2003 г.). «Структурная филогения сериновых бета-лактамаз». Журнал молекулярной эволюции . 57 (3): 255–60. дои : 10.1007/s00239-003-2473-y . ПМИД 14629035 . S2CID 187389 .
- ^ Холл Б.Г., Барлоу М. (апрель 2004 г.). «Эволюция сериновых бета-лактамаз: прошлое, настоящее и будущее». Обновления по лекарственной устойчивости . 7 (2): 111–23. дои : 10.1016/j.drup.2004.02.003 . ПМИД 15158767 .
- ^ Барлоу М., Холл Б.Г. (сентябрь 2002 г.). «Филогенетический анализ показывает, что гены бета-лактамазы OXA присутствуют в плазмидах миллионы лет». Журнал молекулярной эволюции . 55 (3): 314–21. Бибкод : 2002JMolE..55..314B . дои : 10.1007/s00239-002-2328-y . ПМИД 12187384 . S2CID 8679468 .
- ^ «Этимология: β-лактамаза» . Новые инфекционные заболевания . 22 (9): 1689–1631. 2016. дои : 10.3201/eid2209.ET2209 . ПМЦ 4994330 .
Дальнейшее чтение
[ редактировать ]- Сава Т., Коогучи К., Морияма К. (декабрь 2020 г.). «Молекулярное разнообразие β-лактамаз и карбапенемаз расширенного спектра действия и устойчивость к противомикробным препаратам» . Журнал интенсивной терапии . 8 (1): 13. дои : 10.1186/s40560-020-0429-6 . ПМК 6988205 . ПМИД 32015881 .
- Филиппон А., Слама П., Дени П., Лабиа Р. (январь 2016 г.). «Структурная классификация β-лактамаз класса А, широко разнообразного семейства ферментов» . Обзоры клинической микробиологии . 29 (1): 29–57. дои : 10.1128/CMR.00019-15 . ПМЦ 4771212 . ПМИД 26511485 .
- Юн Э.Дж., Чон Ш. (март 2021 г.). «β-лактамазы класса D». Журнал антимикробной химиотерапии . 76 (4): 836–864. дои : 10.1093/jac/dkaa513 . ПМИД 33382875 .
Внешние ссылки
[ редактировать ]- База данных бета-лактамаз
- Бета-лактамазы Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)