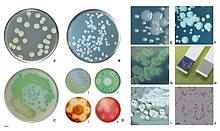
Видовое название aeruginosa - это латинское слово, означающее verdigris («медная ржавчина») и относящееся к сине-зеленому цвету лабораторных культур этого вида. Этот сине-зеленый пигмент представляет собой комбинацию двух вторичных метаболитов P. aeruginosa , пиоцианина (синего) и пиовердина (зеленого), которые придают культурам характерный сине-зеленый цвет. [ 7 ] Другое утверждение 1956 года заключается в том, что aeruginosa может происходить от греческого префикса ae, означающего «старый или престарелый», а суффикс Ruginosa означает морщинистый или неровный. [ 8 ]
Названия пиоцианин и пиовердин происходят от греческого слова «pyo- », что означает «гной». [ 9 ] цианин , что означает «синий», [ 10 ] и вердин , что означает «зеленый». [ нужна ссылка ] Следовательно, термин «гнойноциановые бактерии» относится конкретно к «голубому гною», характерному для инфекции P. aeruginosa . Пиовердин в отсутствие пиоцианина имеет флуоресцентно-желтый цвет. [ нужна ссылка ]
Окрашенные по Граму бактерии P. aeruginosa (розово-красные палочки)
Геном открытых Pseudomonas aeruginosa состоит из относительно большой кольцевой хромосомы (5,5–6,8 Мб), несущей от 5500 до 6000 рамок считывания , а иногда и плазмид различных размеров в зависимости от штамма. [ 11 ] Сравнение 389 геномов разных штаммов P. aeruginosa показало, что только 17,5% являются общими. Эта часть генома представляет собой основной геном P. aeruginosa . [ 12 ]
напряжение:
ВРФПА04
C3719
ПАО1
ПА14
PACS2
Размер хромосомы (п.н.)
6,818,030
6,222,097
6,264,404
6,537,648
6,492,423
ORF
5,939
5,578
5,571
5,905
5,676
Чувствительность P.aeruginosa к антибиотикам.
В сравнительном геномном исследовании (2020 г.) проанализировано 494 полных генома рода Pseudomonas , из которых 189 — штаммы P. aeruginosa . [ 13 ] Исследование показало, что их количество белка и содержание GC варьировались от 5500 до 7352 (в среднем: 6192) и от 65,6 до 66,9% (в среднем: 66,1%) соответственно. [ 13 ] Этот сравнительный анализ дополнительно выявил 1811 ядерных белков aeruginosa, которые составляют более 30% протеома. Более высокий процент ядерных белков aeruginosa в этом последнем анализе можно частично объяснить использованием полных геномов. Хотя P. aeruginosa является очень четко определенным монофилетическим видом как с филогеномической точки зрения, так и с точки зрения значений ANIm, он удивительно разнообразен с точки зрения содержания белка, тем самым обнаруживая очень динамичный дополнительный протеом, согласно нескольким анализам. [ 13 ] [ 14 ] [ 15 ] [ 16 ] Похоже, что в среднем промышленные штаммы имеют самые большие геномы, за ними следуют штаммы из окружающей среды, а затем клинические изоляты. [ 13 ] [ 17 ] В том же сравнительном исследовании (494 штамма Pseudomonas , из которых 189 относятся к P. aeruginosa ) было выявлено, что 41 из 1811 коровых белков P. aeruginosa присутствовали только у этого вида, а не у каких-либо других представителей рода, причем 26 (из 41 ) помечены как гипотетические. Кроме того, еще 19 ортологичных белковых групп присутствуют как минимум у 188/189 штаммов P. aeruginosa и отсутствуют у всех остальных штаммов рода. [ нужна ссылка ]
Популяцию P. aeruginosa можно разделить на три основные линии, генетически характеризующиеся модельными штаммами PAO1, PA14 и более дивергентными PA7. [ 18 ]
Хотя P. aeruginosa обычно считается условно-патогенным патогеном, некоторые широко распространенные клоны, по-видимому, стали более специализированными патогенами, особенно у пациентов с муковисцидозом, включая эпидемический штамм Liverpool (LES), который встречается главным образом в Великобритании. [ 19 ] ДК2 в Дании, [ 20 ] и AUST-02 в Австралии (также ранее известный как AES-2 и P2). [ 21 ] Существует также клон, который часто поражает репродуктивные пути лошадей. [ 22 ] [ 23 ]
P. aeruginosa является факультативным анаэробом , так как хорошо приспособлен к размножению в условиях частичного или полного истощения кислорода. Этот организм может достигать анаэробного роста, используя нитраты или нитриты в качестве терминальных акцепторов электронов . Когда кислород, нитрат и нитрит отсутствуют, он способен ферментировать аргинин и пируват путем фосфорилирования на уровне субстрата . [ 24 ] Кроме того, феназины, продуцируемые P. aeruginosa, могут действовать как переносчики электронов, способствуя выживанию клеток на глубине биопленок. [ 25 ] Адаптация к микроаэробной или анаэробной среде необходима для определенного образа жизни P. aeruginosa , например, во время легочной инфекции при муковисцидозе и первичной цилиарной дискинезии , когда толстые слои легочной слизи и продуцируемый бактериями альгинат, окружающие слизистые бактериальные клетки, могут ограничивать диффузию кислород. Рост P. aeruginosa в организме человека может протекать бессимптомно до тех пор, пока бактерии не образуют биопленку, которая подавляет иммунную систему. Эти биопленки обнаруживаются в легких людей с муковисцидозом и первичной цилиарной дискинезией и могут оказаться смертельными. [ 26 ] [ 27 ] [ 28 ] [ 29 ] [ 30 ] [ 31 ] [ чрезмерное цитирование ]
P. aeruginosa использует железо как источник питательных веществ для роста . Однако железо труднодоступно, поскольку оно редко встречается в окружающей среде. Железо обычно находится в практически нерастворимой трехвалентной форме. [ 32 ] Кроме того, чрезмерно высокие уровни железа могут быть токсичными для P. aeruginosa . Чтобы преодолеть это и регулировать правильное потребление железа, P. aeruginosa использует сидерофоры — секретируемые молекулы, связывающие и транспортирующие железо. [ 33 ] Однако эти комплексы железо-сидерофор не являются специфичными. Бактерия, продуцирующая сидерофоры, не обязательно получает прямую выгоду от потребления железа. Скорее, все члены клеточной популяции с одинаковой вероятностью имеют доступ к комплексам железо-сидерофор. Членов клеточной популяции, которые могут эффективно производить эти сидерофоры, обычно называют кооператорами; членов, которые практически не производят сидерофоров, часто называют мошенниками. Исследования показали, что когда сотрудничающие и мошенники сближаются, у сотрудничающих снижается приспособленность, а у мошенников — повышается. [ 34 ] Величина изменения физической подготовки увеличивается с увеличением ограничения по железу. [ 35 ] По мере повышения физической подготовки мошенники могут превзойти сотрудничающих; это приводит к общему снижению приспособленности группы из-за отсутствия достаточного производства сидерофоров. Эти наблюдения позволяют предположить, что сочетание сотрудничающих и мошенников может снизить вирулентность P. aeruginosa . [ 34 ]
LigD образуют подсемейство ДНК-лигаз . Все они имеют LigDom/лигазный домен, но многие бактериальные LigD также имеют отдельные полимеразные домены/PolDoms и нуклеазные домены/NucDoms. В случае P. aeruginosa домены нуклеазы являются N-концевыми , а домены полимеразы - C-концевыми , являясь расширениями единственного центрального домена лигазы. [ 36 ]
Это наиболее частая причина инфекций ожоговых травм и наружного уха ( наружный отит ), а также наиболее частый колонизатор медицинских устройств (например, катетеров ). Pseudomonas может передаваться через загрязненное и неправильно очищенное оборудование или через руки медицинских работников. [ 38 ] Pseudomonas в редких случаях может вызывать внебольничную пневмонию . [ 39 ] а также вентилятор -ассоциированные пневмонии, которые являются одним из наиболее распространенных агентов, выделенных в нескольких исследованиях. [ 40 ] Пиоцианин является фактором вирулентности бактерий и, как известно, вызывает смерть C. elegans в результате окислительного стресса . Однако салициловая кислота может ингибировать выработку пиоцианина. [ 41 ] Каждая десятая внутрибольничная инфекция вызвана Pseudomonas. [ нужна ссылка ] . Пациенты с муковисцидозом также предрасположены к P. aeruginosa заражению легких из-за функциональной потери движения ионов хлора через клеточные мембраны в результате мутации . [ 42 ] P. aeruginosa также может быть частой причиной «сыпи в горячей ванне» ( дерматита ), вызванной отсутствием надлежащего периодического внимания к качеству воды. Поскольку эти бактерии процветают во влажной среде, например, в гидромассажных ваннах и бассейнах, они могут вызвать сыпь на коже или ухо пловца. [ 38 ] Pseudomonas также является частой причиной послеоперационной инфекции у пациентов, перенесших радиальную кератотомию . Этот организм также связан с поражением кожи гангренозной эктимой . P. aeruginosa часто вызывает остеомиелит с колотыми ранами стопы, который, как полагают, возникает в результате прямой инокуляции P. aeruginosa через пенопластовую прокладку, находящуюся в теннисных туфлях, при этом пациенты с диабетом подвергаются более высокому риску.
Сравнительный геномный анализ 494 полных геномов Pseudomonas , включая 189 полных геномов P. aeruginosa , выявил несколько белков, которые являются общими для подавляющего большинства штаммов P. aeruginosa , но не наблюдаются в других проанализированных Pseudomonas . геномах [ 13 ] Известно, что эти специфичные для aeruginosa коровые белки, такие как CntL, CntM, PlcB, Acp1, MucE, SrfA, Tse1, Tsi2, Tse3 и EsrC , играют важную роль в патогенности этого вида. [ 13 ]
Феназины — окислительно-восстановительные пигменты, продуцируемые P. aeruginosa . Эти пигменты участвуют в ощущении кворума , вирулентности и приобретении железа. [ 46 ] P. aeruginosa продуцирует несколько пигментов, все из которых производятся путем биосинтеза: феназин-1-карбоксамид (PCA), 1-гидроксифеназин, бетаин 5-метилфеназин-1-карбоновой кислоты, пиоцианин и аэругинозин А. В биосинтезе феназина участвуют два почти идентичных оперона. : phzA1B1C1D1E1F1G1 и phzA2B2C2D2E2F2G2 . [ 47 ] [ 48 ] [ 49 ] Ферменты, кодируемые этими оперонами, преобразуют хоризмовую кислоту в PCA. Продукты трех ключевых генов, phzH , phzM и phzS, затем преобразуют PCA в другие феназины, упомянутые выше. Хотя биосинтез феназина хорошо изучен, остаются вопросы относительно окончательной структуры коричневого феназинпиомеланина . [ нужна ссылка ]
При угнетении биосинтеза пиоцианина P. aeruginosa наблюдается снижение патогенности in vitro . Это предполагает, что пиоцианин в основном ответственен за начальную колонизацию P. aeruginosa in vivo . [ 49 ]
при низком фосфатов уровне P. aeruginosa активируется из доброкачественного симбионта, экспрессируя смертельные токсины внутри кишечного тракта и серьезно повреждая или убивая хозяина, что можно смягчить, вводя избыток фосфата вместо антибиотиков. Было обнаружено, что [ 50 ]
P. aeruginosa — условно-патогенный микроорганизм, способный координировать экспрессию генов, чтобы конкурировать с другими видами за питательные вещества или колонизацию. Регулирование экспрессии генов может происходить посредством межклеточной коммуникации или чувства кворума (QS) посредством производства небольших молекул, называемых аутоиндукторами , которые высвобождаются во внешнюю среду. Эти сигналы при достижении определенных концентраций, коррелирующих с плотностью клеток определенной популяции, активируют соответствующие регуляторы, тем самым изменяя экспрессию генов и координируя поведение. P. aeruginosa использует пять взаимосвязанных систем QS – las, rhl, pqs, iqs и pch – каждая из которых продуцирует уникальные сигнальные молекулы. [ 61 ] Системы las и rhl отвечают за активацию многочисленных QS-контролируемых генов, система pqs участвует в передаче сигналов хинолонов, а система iqs играет важную роль в межклеточной коммуникации. [ 62 ] QS у P. aeruginosa организован иерархически. На вершине сигнальной иерархии находится las-система, поскольку las-регулятор инициирует регуляторную систему QS путем активации транскрипции ряда других регуляторов, таких как rhl. Итак, система las определяет иерархический каскад QS от las к регуляронам rhl. [ 63 ] Обнаружение этих молекул указывает на то, что P. aeruginosa растет в виде биопленки в легких пациентов с муковисцидозом. [ 64 ] Однако влияние QS и особенно las-систем на патогенность P. aeruginosa неясно. Исследования показали, что мутанты с дефицитом lasR связаны с более тяжелыми исходами у пациентов с муковисцидозом. [ 65 ] и обнаруживаются у 63% хронически инфицированных пациентов с муковисцидозом, несмотря на нарушение активности QS. [ 66 ]
контролирует экспрессию ряда факторов вирулентности Известно, что QS иерархически , включая пигмент пиоцианин. Однако хотя las-система инициирует регуляцию экспрессии генов, ее отсутствие не приводит к утрате факторов вирулентности. Недавно было продемонстрировано, что система rhl частично контролирует las-специфичные факторы, такие как протеолитические ферменты, ответственные за эластолитическую и стафилолитическую активность, но отсроченным образом. Итак, las является прямым и косвенным регулятором QS-контролируемых генов. [ 62 ] Другая форма регуляции генов , которая позволяет бактериям быстро адаптироваться к изменениям окружающей среды, — это передача сигналов окружающей среды. Недавние исследования показали, что анаэробиоз может существенно влиять на основной регуляторный контур QS. Эта важная связь между QS и анаэробиозом оказывает существенное влияние на продукцию факторов вирулентности этого организма. [ 67 ] Чеснок экспериментально блокирует ощущение кворума у P. aeruginosa . [ 68 ]
Как и у большинства грамотрицательных бактерий, P. aeruginosa образование биопленок регулируется одной молекулой: циклическим ди-ГМФ . При низкой концентрации циклического ди-ГМФ P. aeruginosa ведет свободноплавающий образ жизни. Но когда уровень циклического ди-ГМФ увеличивается, P. aeruginosa начинает образовывать сидячие сообщества на поверхностях. Внутриклеточная концентрация циклического ди-ГМФ увеличивается в течение нескольких секунд, когда P. aeruginosa касается поверхности ( например : камня, пластика, тканей хозяина...). [ 69 ] Это активирует выработку клейких пилей , которые служат «якорями» для стабилизации прикрепления P. aeruginosa к поверхности. На более поздних стадиях бактерии начнут необратимо прикрепляться, образуя прочную клейкую матрицу. В то же время циклический ди-ГМФ подавляет синтез жгутикового аппарата, препятствуя P. aeruginosa плаванию . При подавлении биопленки становятся менее прилипшими и их легче лечить.
Матрикс биопленки P. aeruginosa состоит из нуклеиновых кислот, аминокислот, углеводов и различных ионов. Он механически и химически защищает P. aeruginosa от агрессии со стороны иммунной системы и некоторых токсичных соединений. [ 70 ] Матрица биопленки P. aeruginosa состоит из трех типов сахарных полимеров (или «экзополисахаридов»), называемых PSL, PEL и альгината. [ 71 ] Какие экзополисахариды продуцируются, зависит от штамма. [ 72 ]
синтеза полисахарида Оперон и циклический ди-ГМФ образуют петлю положительной обратной связи. Этот 15-генный оперон отвечает за взаимодействия между клетками и клеточной поверхностью, необходимые для клеточной коммуникации.
ПЭЛ представляет собой катионный экзополисахарид, который сшивает внеклеточную ДНК в матриксе биопленки P. aeruginosa . [ 73 ]
При определенных сигналах или стрессах P. aeruginosa изменяет программу биопленки и отделяется. Недавние исследования показали, что диспергированные клетки биопленок P. aeruginosa имеют более низкие уровни циклического ди-ГМФ и отличаются по физиологии от клеток планктона и биопленок. [ 74 ] [ 75 ] с уникальной динамикой и подвижностью населения. [ 76 ] Установлено, что такие диспергированные клетки обладают высокой вирулентностью против макрофагов и C. elegans , но очень чувствительны к стрессу железа по сравнению с планктонными клетками. [ 74 ]
Биопленки P. aeruginosa могут вызывать хронические оппортунистические инфекции , которые представляют собой серьезную проблему для медицинской помощи в промышленно развитых странах, особенно для пациентов с ослабленным иммунитетом и пожилых людей. Их часто невозможно эффективно лечить с помощью традиционной антибиотикотерапии . Биопленки служат для защиты этих бактерий от неблагоприятных факторов окружающей среды, включая компоненты иммунной системы хозяина в дополнение к антибиотикам. P. aeruginosa может вызывать внутрибольничные инфекции и считается модельным организмом для изучения бактерий, устойчивых к антибиотикам. Исследователи считают важным узнать больше о молекулярных механизмах, которые вызывают переход от планктонного роста к фенотипу биопленки, а также о роли QS в устойчивых к лечению бактериях, таких как P. aeruginosa . Это должно способствовать улучшению клинического ведения хронически инфицированных пациентов и должно привести к разработке новых лекарств. [ 67 ]
Ученые изучают возможную генетическую основу устойчивости P. aeruginosa к антибиотикам, таким как тобрамицин . Одним из локусов, идентифицированных как важный генетический детерминант устойчивости у этого вида, является ndvB , который кодирует периплазматические глюканы , которые могут взаимодействовать с антибиотиками и вызывать их секвестрацию в периплазме. Эти результаты свидетельствуют о том, что за бактериальной устойчивостью к антибиотикам существует генетическая основа, а не биопленка, просто действующая как диффузионный барьер для антибиотика. [ 77 ]
Производство пиоцианина, водорастворимого зеленого пигмента P. aeruginosa (левая трубка)
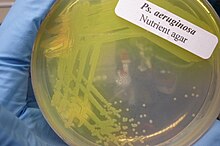
В зависимости от характера инфекции отбирают соответствующий образец и отправляют его в бактериологическую лабораторию для идентификации. Как и в случае большинства бактериологических образцов, проводится окраска по Граму , которая может выявить грамотрицательные палочки и/или лейкоциты . P. aeruginosa образует на бактериологических средах колонии с характерным запахом «винограда» или «свежей лепешки». В смешанных культурах его можно выделить в виде прозрачных колоний на агаре МакКонки (поскольку он не ферментирует лактозу ), тест на оксидазу будет положительным . Подтверждающие тесты включают образование сине-зеленого пигмента пиоцианина на цетримидном агаре и рост при 42 °C. Наклон TSI часто используется для того, чтобы отличить неферментирующие виды Pseudomonas от кишечных патогенов в образцах фекалий. [ нужна ссылка ]
Когда P. aeruginosa выделяется из обычно стерильного участка (крови, костей, глубоких скоплений), он обычно считается опасным и почти всегда требует лечения. [ 78 ] [ 79 ] Однако P. aeruginosa часто выделяется из нестерильных мест (мазков изо рта, мокроты и т. д.), и в этих обстоятельствах это может представлять собой колонизацию, а не инфекцию. Поэтому выделение P. aeruginosa из нестерильных образцов следует интерпретировать с осторожностью и с микробиологом перед началом лечения следует проконсультироваться или врачом-инфекционистом/фармацевтом. Зачастую никакого лечения не требуется. [ нужна ссылка ]
Идентификация P. aeruginosa может быть осложнена тем фактом, что отдельные изоляты часто лишены подвижности. Сама морфология колонии также имеет несколько разновидностей. Основные два типа: большие, гладкие, с плоским краем и приподнятым центром, и маленькие, шероховатые и выпуклые. [ 83 ] Также может быть обнаружен третий тип, мукоид. Большую колонию обычно можно найти в клинальных условиях, а маленькую - в природе. [ 83 ] Третий, однако, присутствует в биологических условиях и был обнаружен в дыхательных путях и мочевыводящих путях. [ 83 ] Более того, мутации в гене lasR радикально изменяют морфологию колоний и обычно приводят к невозможности гидролизовать или гемолизировать желатин. [ нужна ссылка ]
В определенных условиях P. aeruginosa может выделять различные пигменты, в том числе пиоцианин (синий), пиовердин (желтый и флуоресцентный ), пиорубин (красный) и пиомеланин (коричневый). Их можно использовать для идентификации организма. [ 84 ]
Pseudomonas aeruginosa под УФ-освещением Флуоресценция
Многие P. aeruginosa изоляты устойчивы к широкому спектру антибиотиков и могут демонстрировать дополнительную устойчивость после безуспешного лечения. Обычно лечение можно назначать в соответствии с лабораторной чувствительностью, а не выбирать антибиотик эмпирически . Если назначение антибиотиков начинается эмпирически, необходимо приложить все усилия для получения культуральных исследований (до введения первой дозы антибиотика), а выбор используемого антибиотика следует пересмотреть, когда будут получены результаты посева.
Из-за широко распространенной резистентности ко многим распространенным антибиотикам первого ряда карбапенемы , полимиксины и, в последнее время, тигециклин препаратами выбора считались ; однако также сообщалось об устойчивости к этим препаратам. Несмотря на это, они все еще используются в районах, где еще не зарегистрировано сопротивление. Рекомендуется использовать ингибиторы β-лактамаз, такие как сульбактам, в сочетании с антибиотиками для усиления противомикробного действия даже при наличии определенного уровня резистентности. После тщательного тестирования чувствительности к противомикробным препаратам комбинированная терапия оказалась лучшим методом лечения P. aeruginosa с множественной лекарственной устойчивостью . Некоторые антибиотики нового поколения, которые, как сообщается, активны против P. aeruginosa, включают дорипенем, цефтобипрол и цефтаролин. Однако для стандартизации требуются дополнительные клинические испытания. Поэтому исследования по открытию новых антибиотиков и препаратов против P. aeruginosa очень необходимы.
Антибиотики, которые могут обладать активностью в отношении P. aeruginosa включает:
Поскольку фторхинолоны являются одним из немногих классов антибиотиков, широко эффективных против P. aeruginosa , в некоторых больницах их использование строго ограничивается во избежание развития резистентных штаммов. В редких случаях, когда инфекция поверхностная и ограниченная (например, инфекции уха или ногтей), местно гентамицин или колистин. можно использовать [ нужна ссылка ] .
При псевдомонозных раневых инфекциях уксусная кислота в концентрации от 0,5% до 5% может быть эффективным бактериостатическим средством, устраняющим бактерии из раны. Обычно после орошения физиологическим раствором на рану накладывают стерильную марлю, пропитанную уксусной кислотой. Перевязка производилась один раз в день. Pseudomonas обычно устраняется в 90% случаев через 10–14 дней лечения. [ 87 ]
Одной из наиболее тревожных характеристик P. aeruginosa является ее низкая чувствительность к антибиотикам, что объясняется согласованным действием насосов оттока нескольких лекарств с хромосомно-кодируемыми генами устойчивости к антибиотикам, т.е. генами, которые кодируют белки, которые служат ферментами для расщепления антибиотиков. Примерами таких генов являются:
Конкретные гены и ферменты, участвующие в устойчивости к антибиотикам, могут различаться у разных штаммов. [ 101 ] [ 102 ] P. aeruginosa TG523 содержит гены, которые, по прогнозам, обладают антибактериальной активностью, а также гены, участвующие в вирулентности. [ 103 ]
Еще одной особенностью, способствующей устойчивости P. aeruginosa к антибиотикам , является низкая проницаемость клеточных оболочек бактерий. [ 104 ] В дополнение к этой внутренней устойчивости у P. aeruginosa легко развивается приобретенная устойчивость либо в результате мутации в хромосомно-кодируемых генах, либо в результате горизонтального переноса генов , детерминант устойчивости к антибиотикам. Развитие множественной лекарственной устойчивости изолятов P. aeruginosa требует нескольких различных генетических событий, включая приобретение различных мутаций и/или горизонтальный перенос генов устойчивости к антибиотикам. Гипермутация способствует отбору обусловленной мутациями устойчивости к антибиотикам у штаммов P. aeruginosa , вызывающих хронические инфекции, тогда как кластеризация нескольких различных генов устойчивости к антибиотикам в интегронах способствует согласованному приобретению детерминант устойчивости к антибиотикам. Некоторые недавние исследования показали, что фенотипическая устойчивость, связанная с образованием биопленок или появлением вариантов небольших колоний, может иметь важное значение в ответе популяций P. aeruginosa на лечение антибиотиками. [ 67 ]
Было обнаружено, что механизмы, лежащие в основе устойчивости к антибиотикам, включают выработку ферментов, разрушающих или инактивирующих антибиотики, белков внешней мембраны для вытеснения антибиотиков и мутаций для изменения мишеней антибиотиков. Наличие ферментов, расщепляющих антибиотики, таких как β-лактамазы расширенного спектра действия, таких как PER-1, PER-2 и VEB-1, цефалоспориназы AmpC, карбапенемазы, такие как сериновые оксациллиназы, металло-b-лактамазы, карбапенемазы OXA-типа и аминогликозиды. Сообщалось, среди прочего, о модифицирующих ферментах. P. aeruginosa также может модифицировать мишени действия антибиотиков: например, метилирование 16S рРНК для предотвращения связывания аминогликозидов и модификации ДНК или топоизомеразы для защиты ее от действия хинолонов. P. aeruginosa Сообщалось также, что обладает системами оттока нескольких лекарств, которые придают устойчивость к ряду классов антибиотиков, а семейство MexAB-OprM (семейство резистентности-узлевания-разделения ( RND )) считается наиболее важным. [ 105 ] . Важным фактором, связанным с устойчивостью к антибиотикам, является снижение вирулентности резистентного штамма. Такие результаты были получены в случае штаммов, устойчивых к рифампицину и колистину, у которых было зарегистрировано снижение инфекционной способности, чувства кворума и подвижности. [ 106 ]
Мутации ДНК-гиразы обычно связаны с устойчивостью P. aeruginosa к антибиотикам . Эти мутации в сочетании с другими обеспечивают высокую устойчивость, не препятствуя выживанию. Кроме того, гены, участвующие в передаче сигналов циклического ди-ГМФ, могут способствовать устойчивости. Когда P. aeruginosa выращивают в условиях in vitro, имитирующих легкие пациента с муковисцидозом, эти гены неоднократно мутируют. [ 107 ]
Было показано, что две малые РНК , Sr0161 и ErsA , взаимодействуют с мРНК, кодирующей основной порин OprD, ответственный за поглощение карбапенемовых антибиотиков в периплазму . МРНК связываются с 5'UTR oprD , вызывая повышение устойчивости бактерий к меропенему . Другая мРНК, Sr006 , может положительно регулировать (посттранскрипционно) экспрессию PagL, фермента, ответственного за деацилирование липида А. Это снижает провоспалительные свойства липида А. [ 108 ] Более того, аналогично процессу, обнаруженному у сальмонеллы , [ 109 ] Регуляция экспрессии PagL с помощью Sr006 может способствовать к полимиксину B. устойчивости [ 108 ]
Пробиотическая профилактика может предотвратить колонизацию и задержать начало инфекции Pseudomonas в условиях отделения интенсивной терапии. [ 110 ] [ нужен неосновной источник ] иммунопрофилактика против Pseudomonas . В настоящее время изучается [ 111 ]
Риск заражения P. aeruginosa можно снизить, избегая посещения бассейнов, гидромассажных ванн и других водоемов со стоячей водой; регулярная дезинфекция и/или замена оборудования, которое регулярно подвергается воздействию влаги (например, оборудования и растворов для контактных линз); и часто мыть руки (что также защищает от многих других патогенов). Однако даже самые лучшие меры гигиены не могут полностью защитить человека от P. aeruginosa, учитывая, насколько распространена P. aeruginosa в окружающей среде. [ 112 ]
Фаготерапия против P. aeruginosa исследовалась как возможный эффективный метод лечения, который можно сочетать с антибиотиками, не имеет противопоказаний и имеет минимальные побочные эффекты. Фаги производятся в виде стерильной жидкости, пригодной для приема внутрь, применения и т. д. [ 113 ]
О фаготерапии против ушных инфекций, вызванных P. aeruginosa, сообщалось в журнале Clinical Otolaryngology в августе 2009 года. [ 114 ] По состоянию на 2024 год [update] исследования по этой теме продолжаются. [ 115 ]
В 2013 году Жоао Ксавьер описал эксперимент, в котором P. aeruginosa , подвергаясь повторяющимся циклам условий, в которых ему приходилось роиться, чтобы добыть пищу, развил способность «гиперроиться» со скоростью на 25% быстрее, чем базовые организмы, развивая множественные жгутики , тогда как базовый организм имеет один жгутик. [ 116 ] Этот результат был примечателен в области экспериментальной эволюции тем, что он был очень повторяем. [ 117 ] P. aeruginosa изучалась на предмет использования в биоремедиации и при переработке полиэтилена в твердых бытовых отходах . [ 118 ]
этой бактерии Исследования системной биологии привели к разработке метаболических моделей в масштабе генома, которые позволяют компьютерное моделирование и прогнозирование скорости роста бактерий в различных условиях, включая их свойства вирулентности. [ 119 ] [ 120 ]
^ Кидд Т.Дж., Гибсон Дж.С., Мосс С., Грир Р.М., Кобболд Р.Н., Райт Дж.Д. и др. (май 2011 г.). «Клональный комплекс Pseudomonas aeruginosa у лошадей». Ветеринарная микробиология . 149 (3–4): 508–512. дои : 10.1016/j.vetmic.2010.11.030 . ПМИД 21183294 .
^ Шоберт М., Ян Д. (декабрь 2010 г.). «Анаэробная физиология Pseudomonas aeruginosa в легких при муковисцидозе». Международный журнал медицинской микробиологии . 300 (8): 549–556. дои : 10.1016/j.ijmm.2010.08.007 . ПМИД 20951638 .
^ «Псевдомонас аэругиноза» . Интернет-учебник по бактериологии Тодара . 4 июня 2004 г. Архивировано из оригинала 9 октября 2006 г. Проверено 9 сентября 2011 г. - через Textbookofbacteriology.net.
^ Файн М.Дж., Смит М.А., Карсон К.А., Мутха С.С., Санки С.С., Вайсфельд Л.А. и др. (январь 1996 г.). «Прогноз и исходы больных внебольничной пневмонией. Метаанализ». ДЖАМА . 275 (2): 134–141. doi : 10.1001/jama.275.2.134 (неактивен 10 июля 2024 г.). ПМИД 8531309 . {{cite journal}}: CS1 maint: DOI неактивен по состоянию на июль 2024 г. ( ссылка )
^ Чуа С.Л., Хультквист Л.Д., Юань М., Рыбтке М., Нильсен Т.Е., Гивсков М. и др. (август 2015 г.). «Получение in vitro и in vivo и характеристика клеток, диспергированных в биопленках Pseudomonas aeruginosa, с помощью манипуляций с c-ди-GMP». Протоколы природы . 10 (8): 1165–1180. дои : 10.1038/нпрот.2015.067 . HDL : 10356/84100 . ПМИД 26158442 . S2CID 20235088 .
^ Иглевски Б.Х. (1996). «Псевдомонада». В бароне С. и др. (ред.). Медицинская микробиология барона (4-е изд.). Медицинский филиал Техасского университета. ISBN 978-0-9631172-1-2 .
^ Анзай Ю, Ким Х, Пак Джи, Вакабаяши Х, Ояизу Х (июль 2000 г.). «Филогенетическая принадлежность псевдомонад на основе последовательности 16S рРНК». Международный журнал систематической и эволюционной микробиологии . 50 (Часть 4): 1563–1589. дои : 10.1099/00207713-50-4-1563 . ПМИД 10939664 .
^ Кинг Э.О., Уорд М.К., Рэйни Д.Е. (август 1954 г.). «Две простые среды для демонстрации пиоцианина и флуоресцина». Журнал лабораторной и клинической медицины . 44 (2): 301–307. ПМИД 13184240 .
^ Стрибих Р.К., Смарт CE, Гунасекера Т.С., Мюллер С.С., Штробель Э.М., МакНиколс Б.В. и др. (сентябрь 2014 г.). «Характеристика профилей разложения углеводородов дизельного топлива F-76 и авиационного топлива Jet-A Pseudomonas aeruginosa и Marinobactercarbonoclasticus». Международная биопорча и биодеградация . 93 : 33–43. Бибкод : 2014IBiBi..93...33S . дои : 10.1016/j.ibiod.2014.04.024 .
^ Хираката И., Ямагути Т., Накано М., Изумикава К., Мине М., Аоки С. и др. (июль 2003 г.). «Клинические и бактериологические характеристики Pseudomonas aeruginosa, продуцирующих металло-бета-лактамазы IMP-типа». Клин Инфекционный Дис . 37 (1): 26–32. дои : 10.1086/375594 . ПМИД 12830405 .
Arc.Ask3.Ru Номер скриншота №: 7194545bc58c6a37abe4049f78600403__1723716900 URL1:https://arc.ask3.ru/arc/aa/71/03/7194545bc58c6a37abe4049f78600403.html Заголовок, (Title) документа по адресу, URL1: Pseudomonas aeruginosa - Wikipedia
Данный printscreen веб страницы (снимок веб страницы, скриншот веб страницы), визуально-программная копия документа расположенного по адресу URL1 и сохраненная в файл, имеет: квалифицированную, усовершенствованную (подтверждены: метки времени, валидность сертификата), открепленную ЭЦП (приложена к данному файлу), что может быть использовано для подтверждения содержания и факта существования документа в этот момент времени. Права на данный скриншот принадлежат администрации Ask3.ru, использование в качестве доказательства только с письменного разрешения правообладателя скриншота. Администрация Ask3.ru не несет ответственности за информацию размещенную на данном скриншоте. Права на прочие зарегистрированные элементы любого права, изображенные на снимках принадлежат их владельцам. Качество перевода предоставляется как есть. Любые претензии, иски не могут быть предъявлены. Если вы не согласны с любым пунктом перечисленным выше, вы не можете использовать данный сайт и информация размещенную на нем (сайте/странице), немедленно покиньте данный сайт. В случае нарушения любого пункта перечисленного выше, штраф 55! (Пятьдесят пять факториал, Денежную единицу (имеющую самостоятельную стоимость) можете выбрать самостоятельно, выплаичвается товарами в течение 7 дней с момента нарушения.)