Регуляция экспрессии генов
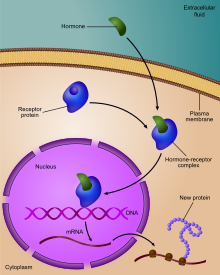
Регуляция экспрессии генов или регуляции генов , [ 1 ] Включает в себя широкий спектр механизмов, которые используются клетками для увеличения или уменьшения производства специфических генных продуктов ( белок или РНК ). Сложные программы экспрессии генов широко наблюдаются в биологии, например, для того, чтобы вызвать пути развития, реагирование на стимулы окружающей среды или адаптироваться к новым источникам пищи. Практически любая этап экспрессии генов может быть модулирована, от транскрипционной инициации , до обработки РНК и посттрансляционной модификации белка. Часто один генный регулятор контролирует другой и т. Д., В генной регуляторной сети .
Регуляция генов необходима для вирусов , прокариот и эукариот , поскольку она увеличивает универсальность и адаптивность организма , позволяя клетке экспрессировать белок при необходимости. Хотя еще в 1951 году Барбара МакКлинток показала взаимодействие между двумя генетическими локусами, активатором ( AC ) и диссоциатором ( DS ), в формировании цветов семян кукурузы, первое открытие системы регуляции генов широко считается идентификацией в 1961 году. из LAC оперона , обнаруженного François Jacob и Jacques Monod , в которых некоторые ферменты, участвующие в метаболизме лактозы, экспрессируются E. coli только в присутствии лактозы и отсутствия глюкозы.
В многоклеточных организмах регуляция генов движет клеточной дифференцировкой и морфогенез в эмбрионе, что приводит к созданию различных типов клеток, которые обладают различными профилями экспрессии генов из одной и той же последовательности генома . Хотя это не объясняет, как возникла регуляция генов, эволюционные биологи включают ее как частичное объяснение того, как работает эволюция на молекулярном уровне , и она является центральным в науке о биологии эволюционной развития («Evo-Devo»).
Регулируемые стадии экспрессии генов
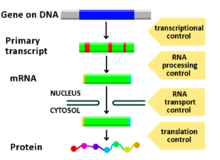
[ редактировать ]Любая этап экспрессии генов может быть модулирован, от передачи сигналов до транскрипции до посттрансляционной модификации белка. Ниже приведен список стадий, на которых регулируется экспрессия генов, где наиболее широко используемой точкой является инициация транскрипции, первая стадия в транскрипции: [ Цитация необходима ]
- Сигнальная трансдукция
- Хроматин , ремоделирование хроматина , домены хроматина
- Транскрипция
- Пост-транскрипционная модификация
- Транспорт РНК
- Перевод
- деградация мРНК
Модификация ДНК
[ редактировать ]
У эукариот из больших областей ДНК может зависеть от его структуры хроматина , которая может быть изменена в результате модификаций гистонов , направленных метилированием ДНК , NCRNA или ДНК-связывающего белка . Следовательно, эти модификации могут вверх или вниз регулировать экспрессию гена. Некоторые из этих модификаций, которые регулируют экспрессию генов, наследуются и называются эпигенетической регуляцией . [ Цитация необходима ]
Структурный
[ редактировать ]Транскрипция ДНК продиктована ее структурой. В целом, плотность его упаковки указывает на частоту транскрипции. Октамерные белковые комплексы, называемые гистонами, вместе с сегментом ДНК -раны вокруг восьми гистоновых белков (вместе, называемых нуклеосомой), ответственны за количество супертистирования ДНК, и эти комплексы могут быть временно изменены такими процессами, как фосфорилирование или более постоянно навсегда. Модифицируется такими процессами, как метилирование . Такие модификации считаются ответственными за более или менее постоянные изменения в уровнях экспрессии генов. [ 2 ]
Химический
[ редактировать ]Метилирование ДНК является распространенным методом молчания генов. ДНК обычно метилируется метилтрансферазой ферментами на цитозиновых нуклеотидах в последовательности динуклеотидов CPG (также называемого « островками CPG », когда он плотно кластерирован). Анализ паттерна метилирования в данной области ДНК (которая может быть промотором) может быть достигнут с помощью метода, называемого бисульфитовым отображением. Метилированные остатки цитозина не изменяются из -за лечения, тогда как неметилированные из них изменяются на урацил. Различия анализируются с помощью секвенирования ДНК или методов, разработанных для количественной оценки SNP, таких как пиросеквенирование ( биотаж ) или Massarray ( Sequeenom ), измеряя относительные количества C/T в CG -динуклеотиде. Считается, что аномальные паттерны метилирования участвуют в онкогенезе. [ 3 ]
Ацетилирование гистона также является важным процессом в транскрипции. Ферменты гистонцетилтрансферазы (HAT), такие как CREB-связывающий белок, также диссоциации ДНК из гистонового комплекса, позволяя пройти транскрипцию. Часто метилирование ДНК и деацетилирование гистонов работают вместе при молчании генов . Комбинация двух, по -видимому, является сигналом для упаковки ДНК более плотно, снижая экспрессию генов. [ Цитация необходима ]
Регулирование транскрипции
[ редактировать ]
Регуляция транскрипции, таким образом, контролирует, когда происходит транскрипция и сколько РНК создается. Транскрипция гена с помощью РНК -полимеразы может регулироваться несколькими механизмами. Факторы специфичности изменяют специфичность РНК -полимеразы для данного промотора или набора промоторов, что делает ее более или менее вероятной связываться с ними (то есть сигма -факторы , используемые в прокариотической транскрипции ). Репрессоры связываются с оператором , кодирующие последовательности на цепи ДНК, которые близки или перекрывают или перекрывают область промотора, препятствуя прогрессу РНК -полимеразы вдоль цепи, что препятствует экспрессии гена. Образ справа демонстрирует регулирование репрессором в опероне LAC. Общие факторы транскрипции позиционируют РНК-полимеразу в начале последовательности кодирования белка, а затем высвобождают полимеразу для транскрибирования мРНК. Активаторы усиливают взаимодействие между РНК -полимеразой и конкретным промотором , поощряя экспрессию гена. Активаторы делают это, увеличивая притяжение РНК -полимеразы для промотора посредством взаимодействия с субъединицами РНК -полимеразы или косвенно, изменяя структуру ДНК. Энхансеры - это сайты на спирали ДНК, которые связаны активаторами, чтобы зацикливаться на ДНК, приводящей специфический промотор в комплекс инициации. Усилители гораздо чаще встречаются у эукариот, чем прокариоты, где существует только несколько примеров (на сегодняшний день). [ 4 ] Глушители представляют собой области последовательностей ДНК, которые, когда они связаны определенными факторами транскрипции, могут заставить замолчать экспрессию гена.
Регуляция РНК
[ редактировать ]РНК может быть важным регулятором генной активности, например, микроРНК (miRNA), антисмысловой РНК или длинной некодирующей РНК (LNCRNA). LNCRNAs отличаются от мРНК в том смысле, что они указали субклеточные местоположения и функции. Сначала они были обнаружены в ядре и хроматине , а локализации и функции теперь очень разнообразны. Некоторые по -прежнему находятся в хроматине, где они взаимодействуют с белками. В то время как эта lncrna в конечном итоге влияет на экспрессию генов при заболеваниях нейрональных заболеваний, таких как Паркинсон , Хантингтон и болезнь Альцгеймера , другие, такие как PNCTR (богатые пиримидином некодирующие транскрипторы), играют роль в раке легких . Учитывая их роль в заболеваниях, LNCRNAS являются потенциальными биомаркерами и могут быть полезными мишенями для лекарств или генной терапии , хотя еще нет одобренных лекарств, которые Targert Lncrnas. Количество LNCRNAS в геноме человека остается плохо определено, но некоторые оценки варьируются от 16 000 до 100 000 генов LNC. [ 5 ]
Эпигенетическая регуляция генов
[ редактировать ]
Эпигенетика относится к модификации генов, которые не меняют последовательность ДНК или РНК. Эпигенетические модификации также являются ключевым фактором в влиянии на экспрессию генов . Они встречаются на геномной ДНК и гистонах , а их химические модификации регулируют экспрессию генов более эффективным образом. Существует несколько модификаций ДНК (обычно метилирование ) и более 100 модификаций РНК в клетках млекопитающих ». Эти модификации приводят к измененному связыванию белка с ДНК и изменению стабильности РНК и эффективности трансляции . [ 6 ]
Особые случаи в биологии и болезнях человека
[ редактировать ]Регуляция транскрипции при раке
[ редактировать ]У позвоночных большинство генных промоторов содержат остров CPG с многочисленными участками CPG . [ 7 ] Когда многие из участков промотора гена метилированы, ген замолчал. [ 8 ] Колоректальный рак обычно имеет от 3 до 6 мутаций драйвера и от 33 до 66 мутаций с автостопом или пассажирами. [ 9 ] Тем не менее, транскрипционное молчание может иметь большее значение, чем мутация при выборе прогрессирования к раку. Например, при колоректальном раке от 600 до 800 генов транскрипционно молчат метилированием острова CPG (см. Регуляцию транскрипции при раке ). Репрессия транскрипции при раке также может возникать другими эпигенетическими механизмами, такими как измененная экспрессия микроРНК . [ 10 ] При раке молочной железы транскрипционная репрессия BRCA1 может происходить чаще с помощью чрезмерной экспрессированной микроРНК-182, чем гиперметилированием промотора BRCA1 (см. Низкую экспрессию BRCA1 при раке молочной железы и яичников ).
Регуляция транскрипции в зависимости
[ редактировать ]Одной из кардинальных особенностей зависимости является ее устойчивость. По-видимому, постоянные поведенческие изменения связаны с длительными изменениями, возникающими в результате эпигенетических изменений, влияющих на экспрессию генов, в определенных областях мозга. [ 11 ] Напрашивания злоупотребления вызывают три типа эпигенетических изменений в мозге. Это (1) ацетилирование гистонов и метилирование гистонов , (2) метилирование ДНК в CPG и (3) эпигенетическое подавление или активацию микроРНК сайтах . [ 11 ] [ 12 ] (См. Эпигенетику зависимости кокаина для некоторых деталей.)
Хроническое потребление никотина у мышей изменяет эпигенетический контроль клеток головного мозга посредством ацетилирования гистонов . Это увеличивает экспрессию в мозге белка FOSB, важного в зависимости. [ 13 ] Привыкание к сигарете также изучалась примерно у 16 000 человек, в том числе никогда не курящих, нынешних курильщиков и тех, кто бросил курить на срок до 30 лет. [ 14 ] В клетках крови более 18 000 сайтов CPG (из примерно 450 000 проанализированных сайтов CPG в геноме) часто изменяли метилирование среди нынешних курильщиков. Эти сайты CPG происходили в более чем 7000 генах или примерно трети известных человеческих генов. Большинство дифференциально метилированных сайтов CPG вернулись на уровень никогда некурящих в течение пяти лет после прекращения курения. Тем не менее, 2568 CPG среди 942 генов оставались дифференциально метилированными у прежних и никогда не курильщиков. Такие оставшиеся эпигенетические изменения можно рассматривать как «молекулярные шрамы» [ 12 ] Это может повлиять на экспрессию генов.
В моделях грызунов, наркотики, в том числе кокаин, [ 15 ] метамфетамин, [ 16 ] [ 17 ] алкоголь [ 18 ] и табачный дымовой продукт, [ 19 ] Все вызывает повреждение ДНК в мозге. Во время восстановления повреждений ДНК некоторые индивидуальные события восстановления могут изменить метилирование ДНК и/или ацетилирования или метилирования гистонов на участках повреждения и, таким образом, могут способствовать оставлению эпигенетического рубца на хроматине. [ 20 ]
Такие эпигенетические шрамы, вероятно, способствуют постоянным эпигенетическим изменениям, обнаруженным в зависимости.
Регуляция транскрипции в обучении и памяти
[ редактировать ]
У млекопитающих метилирование цитозина (см. Рисунок) в ДНК является основным регуляторным посредником. Метилированные цитозины в основном встречаются в динуклеотидных последовательностях, где цитозин сопровождается гуанином, сайтом CPG . Общее количество участков CPG в геноме человека составляет приблизительно 28 миллионов. [ 21 ] И обычно около 70% всех сайтов CPG имеют метилированный цитозин. [ 22 ]

У крысы болезненный опыт обучения, контекстуальная кондиционирование страха , может привести к страшной памяти на протяжении всей жизни после одного тренировочного события. [ 23 ] Метилирование цитозина изменяется в промоторных областях около 9,17% всех генов в ДНК нейронов гиппокампа крысы, которая подвергалась краткому образованию страха . [ 24 ] Гиппокамп - это место, где изначально хранятся новые воспоминания.
Метилирование CPGS в промоторной области гена репрессирует транскрипцию [ 25 ] в то время как метилирование CPG в организме гена увеличивает экспрессию. [ 26 ] Тет -ферменты играют центральную роль в деметилировании метилированных цитозинов. Деметилирование CPGs в геном промоторе с помощью активности фермента TET увеличивает транскрипцию гена. [ 27 ]
Когда контекстуальное обусловленность страха применяется к крысе, более 5000 дифференциально метилированных областей (DMR) (по 500 нуклеотидам каждый) встречаются в нервном геноме гиппокампа крысы как один час, так и через 24 часа после кондиционирования в гиппокампе. [ 24 ] Это приводит к повышению регуляции около 500 генов (часто из-за деметилирования сайтов CPG в промоторной области) и около 1000 генов, которые будут понижены (часто из-за недавно образованного 5-метилцитозина в участках CPG в промоторной области). Паттерн индуцированных и репрессированных генов в нейронах, по -видимому, обеспечивает молекулярную основу для формирования первой переходной памяти этого тренировочного события в гиппокампе мозга крысы. [ 24 ]
Пост-транскрипционное регулирование
[ редактировать ]После того, как ДНК транскрибируется и образуется мРНК, должна быть какая -то регуляция о том, сколько мРНК транслируется в белки. Клетки делают это путем модуляции уплотнения, сплайсинга, добавления поли (a) хвоста, специфических для последовательностей скоростей ядерного экспорта и, в нескольких контекстах, секвестрации транскрипта РНК. Эти процессы происходят у эукариот, но не у прокариот. Эта модуляция является результатом белка или транскрипта, который, в свою очередь, регулируется и может иметь сродство к определенным последовательностям.
Три первичных нетранслируемых регионов и микроРНК
[ редактировать ]Три первичных нетранслируемых областей (3'-UTR) мессенджевых РНК (мРНК) часто содержат регуляторные последовательности, которые посттранскрипционно влияют на экспрессию генов. [ 28 ] Такие 3'-UTR часто содержат как сайты связывания для микроРНК (miRNAs), так и для регуляторных белков. Связываясь со специфическими сайтами в пределах 3'-UTR, miRNAs могут снижать экспрессию генов различных мРНК, либо ингибируя трансляцию или непосредственно вызывая деградацию транскрипта. 3'-UTR также могут иметь области глушителя, которые связывают репрессорные белки, которые ингибируют экспрессию мРНК.
3'-UTR часто содержит элементы ответа miRNA (MRES) . MRE - это последовательности, с которыми связывают miRNAs. Это распространенные мотивы в пределах 3'-UTR. Среди всех регулирующих мотивов в пределах 3'-UTR (например, включая области глушителя) MRE составляют около половины мотивов.
По состоянию на 2014 год, веб -сайт mirbase , [ 29 ] Архив последовательностей и аннотаций miRNA, перечисляется 28 645 записей у 233 биологических видов. Из них 1881 микроРНК были в аннотированных локусах miRNA человека. Предполагалось, что miRNAs будут иметь в среднем около четырехсот мРНК мишени (влияя на экспрессию нескольких сотен генов). [ 30 ] Фрейдман и соавт. [ 30 ] Оценить, что> 45 000 сайтов мишеней miRNA в пределах мРНК человека 3'-UTR сохраняются выше фоновых уровней, и> 60% генов, кодирующих белок человека, подвергались селективному давлению, чтобы поддерживать сопряжение с miRNAs.
Прямые эксперименты показывают, что одна miRNA может снизить стабильность сотен уникальных мРНК. [ 31 ] Другие эксперименты показывают, что одна miRNA может подавлять производство сотен белков, но эта репрессия часто является относительно легкой (менее чем в 2 раза). [ 32 ] [ 33 ]
Эффекты дисрегуляции miRNA экспрессии генов, по -видимому, важны при раке. [ 34 ] Например, при раке желудочно-кишечного тракта бумага 2015 года идентифицировала девять miRNAs как эпигенетически измененные и эффективные в пониженных ферментах репарации ДНК. [ 35 ]
Эффекты диспрегуляции miRNA экспрессии генов также, по -видимому, также являются важными при нейропсихиатрических расстройствах, таких как шизофрения , биполярное расстройство , серьезное депрессивное расстройство , болезнь Паркинсона , болезнь Альцгеймера и расстройства спектра аутизма . [ 36 ] [ 37 ] [ 38 ]
Регулирование перевода
[ редактировать ]Перевод мРНК также может контролироваться рядом механизмов, в основном на уровне инициации. Рекрутирование небольшой рибосомной субъединицы действительно может быть модулирована с помощью вторичной структуры мРНК, антисмыслового связывания РНК или связывания белка. Как у прокариот, так и у эукариот существует большое количество белков, связывающих РНК, которые часто направляются на их целевую последовательность по вторичной структуре транскрипта, которая может изменяться в зависимости от определенных условий, таких как температура или присутствие лиганда (аптамер) Полем Некоторые транскрипты действуют как рибозимы и саморегулируют их экспрессию.
Примеры регуляции генов
[ редактировать ]- Индукция фермента - это процесс, в котором молекула (например, препарат) индуцирует (то есть инициирует или усиливает) экспрессию фермента.
- Индукция белков теплового шока в фруктовой мух Drosophila melanogaster .
- Оперон LAC является интересным примером того, как можно регулировать экспрессию генов.
- Вирусы, несмотря на наличие только нескольких генов, обладают механизмами для регуляции экспрессии своих генов, как правило, в ранней и поздней фазе с использованием коллинеарных систем, регулируемых анти-концевыми ( Lambda Phage ) или модуляторами сплайсинга ( ВИЧ ).
- GAL4 является активатором транскрипции, который контролирует экспрессию GAL1, GAL7 и GAL10 (все из которых код для метаболизма галактозы у дрожжей). Система GAL4/UAS использовалась в различных организмах по различным филам для изучения экспрессии генов. [ 39 ]
Биология развития
[ редактировать ]Большое количество изученных систем регулирования поступает из биологии развития . Примеры включают:
- Колинеарность кластера генов Hox с их вложенным передне-задним тщательностью
- Генерация рисунка руки (цифры - междигиты): градиент звукового хеджиха (секретированный индуцирующий фактор) из зоны поляризующей активности в конечности, которая создает градиент активного глина, который активирует гремлин, который ингибирует BMPS также секретируется в конечность приводит к формированию чередующейся картины активности в результате этой системы реакции -диффузии .
- Сомитогенез представляет собой создание сегментов (сомитов) из равномерной ткани (до-ссомитская мезодерма ). Они образуются последовательно от передней к задней. Это достигается в амниотах, возможно, с помощью двух противоположных градиентов: ретиноевой кислоты в передней (волновой фронте) и Wnt и FGF в задней части, в сочетании с колебательной картиной (часы сегментации), состоящий из FGF + Notch и Wnt в антипхазе. [ 40 ]
- Определение пола в соме дрозофилы требует восприятия соотношения аутосомных генов к генам, кодируемым половым хромосомой , что приводит к производству серийного фактора сплайсинга у женщин, что приводит к женской изоформе дублеса. [ 41 ]
Схема
[ редактировать ]Повышение регуляции и понижающая регуляция
[ редактировать ]Повышенная регуляция-это процесс, который возникает в ячейке, запускаемой сигналом (происходящим внутренним или внешним по отношению к клетке), что приводит к повышению экспрессии одного или нескольких генов, и в результате белки, кодируемые этими генами. И наоборот, понижающая регуляция-это процесс, приводящий к снижению гена и соответствующей экспрессии белка.
- активация Например, наблюдается , когда клетка дефицит в каком-то рецепторе. В этом случае больше рецепторного белка синтезируется и транспортируется в мембрану клетки, и, таким образом, чувствительность клетки возвращается к нормальному восстановлению гомеостаза .
- Понижающая регуляция происходит, например, когда клетка чрезмерно стимулируется нейротрансмиттером , гормоном или препаратом в течение длительного периода времени, а экспрессия рецепторного белка снижается для защиты клетки (см. Также тахифилактику ).
Индуцируемые и репрессируемые системы
[ редактировать ]
Регуляция генов может быть обобщена реакцией соответствующей системы:
- Индуцируемые системы - индуцируемая система отключена, если не существует наличия некоторой молекулы (называемой индуктором), которая позволяет экспрессировать генов. Говорят, что молекула «индуцирует выражение». Способ, которым это происходит, зависит от механизмов контроля, а также различий между прокариотическими и эукариотическими клетками.
- Репрессируемые системы - репрессируемая система находится, за исключением присутствия некоторой молекулы (называемой корепрессором), которая подавляет экспрессию генов. Говорят, что молекула «подавляет выражение». Способ, которым это происходит, зависит от механизмов контроля, а также различий между прокариотическими и эукариотическими клетками.
Система GAL4/UAS является примером как индуцибельной, так и репрессируемой системы. GAL4 связывает последовательность активации вверх по течению (UAS), чтобы активировать транскрипцию кассеты GAL1/GAL7/GAL10. С другой стороны, ответ MIG1 на присутствие глюкозы может ингибировать GAL4 и, следовательно, остановить экспрессию кассеты GAL1/GAL7/GAL10. [ 42 ]
Теоретические схемы
[ редактировать ]- Репрессор/индуктор: активация датчика приводит к изменению экспрессии гена
- отрицательная обратная связь: генный продукт прямо или косвенно подавляет собственное производство, что может привести к
- поддержание уровней транскриптов постоянным/пропорциональным фактору
- ингибирование реакций, в сочетании с петлей положительной обратной связи
- Создание генератора, используя преимущество в задержке транскрипции и транскрипции, учитывая, что мРНК и период полураспада белка короче
- положительная обратная связь: генный продукт прямо или косвенно активизирует собственное производство, что может привести к
- усиление сигнала
- Бистабильные переключатели, когда два гена ингибируют друг друга, и оба имеют положительную обратную связь
- генерация образа
Методы исследования
[ редактировать ]
В целом, в большинстве экспериментов, исследующих дифференциальную экспрессию, использовались цельные клеточные экстракты РНК, называемые стационарными уровнями, чтобы определить, какие гены изменились и сколько. Они, однако, не информативны о том, где произошла регулирование, и могут маскировать противоречивые регуляторные процессы ( см. Пост-транскрипционную регуляцию ), но она по-прежнему наиболее часто анализируется ( количественная ПЦР и микрочип ДНК ).
При изучении экспрессии генов есть несколько методов, чтобы посмотреть на различные этапы. В Eukaryotes они включают:
- Локальная среда хроматина в области может быть определена с помощью анализа чип-чипа путем стягивания РНК-полимеразы II , гистона 3 модификаций , белка-транторизаторной группы , белка поликомб-группы или любого другого ДНК-связывающего элемента, к которому доступно хорошее антитело. Полем
- Эпистатические взаимодействия могут быть исследованы с помощью синтетического генетического массива анализа
- Из-за посттранскрипционной регуляции скорости транскрипции и общие уровни РНК значительно различаются. Для измерения скорости транскрипции могут быть проведены ядерные анализы и разработаны новые высокопроизводительные методы с использованием маркировки тиола вместо радиоактивности . [ 43 ]
- Только 5% РНК -полимеризуемых в выходах ядра, [ 44 ] И не только интроны, абортивные продукты и бессмысленные транскрипты деградируют. Следовательно, различия в ядерных и цитоплазматических уровнях можно увидеть путем разделения двух фракций путем мягкого лизиса. [ 45 ]
- Альтернативный сплайсинг может быть проанализирован с помощью массива сплайсинга или с помощью массива плитки ( см. ДНК -микрочип ).
- Вся in vivo РНК комплекс в виде RNP . Количество транскриптов, связанных с специфическим белком, также может быть проанализировано с помощью Rip-Chip . Например, DCP2 будет указывать на секвестрированный белок; РИЗОСОМА -БЛИЧНЫЕ ДАВЛЕНИЯ И УДАЛЕНИЕ Транскриптов, активных в транскрипции (хотя более устаревший метод, называемый полисом , все еще популярен в некоторых лабораториях)
- Уровни белка могут быть проанализированы с помощью масс -спектрометрии , которую можно сравнить только с количественными данными ПЦР, поскольку данные микрочипа являются относительными, а не абсолютными.
- Скорость деградации РНК и белка измеряется с помощью ингибиторов транскрипции ( актиномицин D или α-аманитин ) или ингибиторы трансляции ( циклогексимид ), соответственно.
Смотрите также
[ редактировать ]- Искусственные факторы транскрипции (мелкие молекулы, которые имитируют белок транскрипции)
- Клеточная модель
- Консервативная некодирующая последовательность ДНК
- Enhancer (генетика)
- Структура гена
- Пространственно -временная экспрессия генов
- Регулятор генов глюкозилтрансферазы (RGG/SHP)
Примечания и ссылки
[ редактировать ]- ^ "Можно ли включаться и выключать гены в клетках?" Полем Генетика дома ссылка .
- ^ Bell JT, Pai AA, Pickrell JK, Gaffney DJ, Pique-regi R, Degner JF, et al. (2011). «Паттерны метилирования ДНК связаны с генетической и экспрессией генов в клеточных линиях HAPMAP» . Биология генома . 12 (1): R10. doi : 10.1186/gb-2011-12-1-r10 . PMC 3091299 . PMID 21251332 .
- ^ Vertino PM, Spillare EA, Harris CC, Baylin SB (апрель 1993 г.). «Измененные паттерны хромосомного метилирования сопровождают онкогеновую трансформацию эпителиальных клеток бронхов человека» (PDF) . РАНКА . 53 (7): 1684–9. PMID 8453642 .
- ^ Остин С., Диксон Р (июнь 1992 г.). «Прокариотический энхансер -связывающий белок NTRC обладает активностью АТФазы, которая является фосфорилированием и ДНК -зависимой» . Embo Journal . 11 (6): 2219–28. doi : 10.1002/j.1460-2075.1992.tb05281.x . PMC 556689 . PMID 1534752 .
- ^ Statello L, Guo CJ, Chen LL, Huarte M (февраль 2021 г.). «Регуляция генов с помощью длинных некодирующих РНК и его биологических функций» . Природные обзоры. Молекулярная клеточная биология . 22 (2): 96–118. doi : 10.1038/s41580-020-00315-9 . ISSN 1471-0072 . PMC 7754182 . PMID 33353982 .
- ^ Кан Р.Л., Чен Дж, Саллам Т (июль 2021 г.). «Перекрестка между эпитранскриптомными и эпигенетическими механизмами в регуляции генов» . Тенденции в генетике . 38 (2): 182–193. doi : 10.1016/j.tig.2021.06.014 . PMC 9093201 . PMID 34294427 . S2CID 236200223 .
- ^ Саксонов С., Берг П., Брутлаг Д.Л. (январь 2006 г.). «Анализ общегенома CPG-динуклеотидов в человеческом геноме отличает два различных класса промоторов» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (5): 1412–7. Bibcode : 2006pnas..103.1412S . doi : 10.1073/pnas.0510310103 . PMC 1345710 . PMID 16432200 .
- ^ Птица А (январь 2002 г.). «Паттерны метилирования ДНК и эпигенетическая память» . Гены и развитие . 16 (1): 6–21. doi : 10.1101/gad.947102 . PMID 11782440 .
- ^ Vogelstein B, Papadopoulos N, Velculescu Ve, Zhou S, Diaz LA, Kinzler KW (март 2013 г.). «Ландшафты генома рака» . Наука . 339 (6127): 1546–58. BIBCODE : 2013SCI ... 339.1546V . doi : 10.1126/science.1235122 . PMC 3749880 . PMID 23539594 .
- ^ Tessitore A, Cicciarelli G, Del Vecchio F, Gaggiano A, Verzella D, Phoristles M, et al. (2014). «МикроРНК в сети повреждения/восстановления ДНК и рак» . Международный журнал геномики . 2014 : 820248. DOI : 10.1155/2014/820248 . PMC 3926391 . PMID 24616890 .
- ^ Jump up to: а беременный Nestler EJ (январь 2014 г.). «Эпигенетические механизмы наркомании» . Нейрофармакология . 76 Pt B: 259–68. doi : 10.1016/j.neuropharm.2013.04.004 . PMC 3766384 . PMID 23643695 .
- ^ Jump up to: а беременный Робисон AJ, Nestler EJ (октябрь 2011 г.). «Транскрипционные и эпигенетические механизмы зависимости» . Природные обзоры. Нейробиология . 12 (11): 623–37. doi : 10.1038/nrn3111 . PMC 3272277 . PMID 21989194 .
- ^ Levine A, Huang Y, Drisaldi B, Griffin EA, Pollak DD, Xu S, et al. (Ноябрь 2011). «Молекулярный механизм лекарственного средства для шлюза: эпигенетические изменения, инициированные никотиновым геном экспрессии кокаином» . Научная трансляционная медицина . 3 (107): 107Ra109. doi : 10.1126/scitranslmed.3003062 . PMC 4042673 . PMID 22049069 .
- ^ Джоханес Р., Just AC, Marioni RE, Pilling LC, Reynolds LM, Mandaviya PR, et al. (Октябрь 2016). «Эпигенетические подписи курящего сигарет» . Циркуляция: сердечно -сосудистая генетика . 9 (5): 436–447. doi : 10.1161/circgenetics.116.001506 . PMC 5267325 . PMID 27651444 .
- ^ De Souza MF, Gonçales TA, Steinmetz A, Moura DJ, Saffi J, Gomez R, Barros HM (апрель 2014 г.). «Кокаин индуцирует повреждение ДНК в различных областях мозга самки крыс в разных гормональных условиях». Клиническая и экспериментальная фармакология и физиология . 41 (4): 265–9. doi : 10.1111/1440-1681.12218 . PMID 24552452 . S2CID 20849951 .
- ^ Johnson Z, Venters J, Guarraci FA, Zewail-Foote M (июнь 2015 г.). «Метамфетамин индуцирует повреждение ДНК в определенных областях мозга самки крысы». Клиническая и экспериментальная фармакология и физиология . 42 (6): 570–5. doi : 10.1111/1440-1681.12404 . PMID 25867833 . S2CID 24182756 .
- ^ Tokunaga I, Ishigami A, Kubo S, Gotohda T, Kitamura O (август 2008 г.). «Повреждение пероксидативного ДНК и апоптоз в мозге крысы, получавшего метамфетамин» . Журнал медицинского расследования . 55 (3–4): 241–5. doi : 10.2152/jmi.55.241 . PMID 18797138 .
- ^ Rulten SL, Hodder E, Ripley TL, Stephens DN, Mayne LV (июль 2008 г.). «Алкоголь индуцирует повреждение ДНК и белок анемии Fanconi D2, который применяет FANCD2 в путях ответа на повреждение ДНК в мозге» . Алкоголизм: клинические и экспериментальные исследования . 32 (7): 1186–96. doi : 10.1111/j.1530-0277.2008.00673.x . PMID 18482162 .
- ^ Adhami N, Chen Y, Martins-Green M (октябрь 2017 г.). «Биомаркеры заболевания могут быть обнаружены у мышей через 4 недели после начала воздействия уровня дыма в третьем руке, эквивалентных тем, которые обнаружены в домах курильщиков» . Клиническая наука . 131 (19): 2409–2426. doi : 10.1042/cs20171053 . PMID 28912356 .
- ^ Dabin J, Fortuny A, Polo SE (июнь 2016 г.). «Поддержание эпигенома в ответ на повреждение ДНК» . Молекулярная клетка . 62 (5): 712–27. doi : 10.1016/j.molcel.2016.04.006 . PMC 5476208 . PMID 27259203 .
- ^ Lövkvist C, Dodd IB, Sneppen K, Haerter Jo (июнь 2016 г.). «Метилирование ДНК в эпигеномах человека зависит от локальной топологии сайтов CPG» . Исследование нуклеиновых кислот . 44 (11): 5123–32. doi : 10.1093/nar/gkw124 . PMC 4914085 . PMID 26932361 .
- ^ Джаббари К., Бернарди Г (май 2004 г.). «Метилирование цитозина и CPG, TPG (CPA) и частоты TPA». Ген . 333 : 143–9. doi : 10.1016/j.gene.2004.02.043 . PMID 15177689 .
- ^ Ким Дж.Дж., Юнг М.В. (2006). «Нейронные схемы и механизмы, связанные с павловским страхом: критический обзор» . Нейробиологии и биобиоэвиральные обзоры . 30 (2): 188–202. doi : 10.1016/j.neubiorev.2005.06.005 . PMC 4342048 . PMID 16120461 .
- ^ Jump up to: а беременный в Герцог К.Г., Кеннеди А.Дж., Гэвин К.Ф., Дэй Дж.Дж., Поттит Дж.Д. (июль 2017 г.). «В зависимости от опыта эпигеномная реорганизация в гиппокампе» . Обучение и память . 24 (7): 278–288. doi : 10.1101/lm.045112.117 . PMC 5473107 . PMID 28620075 .
- ^ Вебер М., Хеллманн I, Стадлер М.Б., Рамос Л., Пябо С., Ребхан М., Шюбелер Д. (апрель 2007 г.). «Распределение, потенциал молчания и эволюционное влияние метилирования ДНК промотора в геноме человека». НАТ Генет . 39 (4): 457–66. doi : 10.1038/ng1990 . PMID 17334365 . S2CID 22446734 .
- ^ Yang X, Han H, De Carvalho DD, Lay FD, Jones PA, Liang G (октябрь 2014 г.). «Метилирование тела генов может изменить экспрессию генов и является терапевтической мишенью при раке» . Раковая клетка . 26 (4): 577–90. doi : 10.1016/j.ccr.2014.07.028 . PMC 4224113 . PMID 25263941 .
- ^ Maeder ML, Angstman JF, Richardson ME, Linder SJ, Cascio VM, Tsai SQ, Ho QH, Sander JD, Reyon D, Bernstein BE, Costello JF, Wilkinson MF, Joung JK (декабрь 2013 г.). «Целевое деметилирование ДНК и активация эндогенных генов с использованием программируемых слитых белков Tale-TET1» . НАТ Биотехнол . 31 (12): 1137–42. doi : 10.1038/nbt.2726 . PMC 3858462 . PMID 24108092 .
- ^ Ogorodnikov A, Kargapolova Y, Danckwardt S (июнь 2016 г.). «Обработка и расширение транскриптома на мРНК 3 'заканчиваются здоровьем и заболеванием: поиск правильного конца» . Pflügers Archiv . 468 (6): 993–1012. doi : 10.1007/s00424-016-1828-3 . PMC 4893057 . PMID 27220521 .
- ^ mirbase.org
- ^ Jump up to: а беременный Friedman RC, Farh KK, Burge CB, Bartel DP (январь 2009 г.). «Большинство мРНК млекопитающих являются консервативными целями микроРНК» . Исследование генома . 19 (1): 92–105. doi : 10.1101/gr.082701.108 . PMC 2612969 . PMID 18955434 .
- ^ Lim LP, Lau NC, Garrett-Engele P, Grimson A, Schelter JM, Castle J, et al. (Февраль 2005 г.). «Анализ микрочипов показывает, что некоторые микроРНК подавляют большое количество целевых мРНК». Природа . 433 (7027): 769–73. Bibcode : 2005natur.433..769L . doi : 10.1038/nature03315 . PMID 15685193 . S2CID 4430576 .
- ^ Selbach M, Schwanhäusser B, Thierfelder N, Fang Z, Khanin R, Rajewsky N (сентябрь 2008 г.). «Широко распространенные изменения в синтезе белка, вызванные микроРНК». Природа . 455 (7209): 58–63. Bibcode : 2008natur.455 ... 58 с . doi : 10.1038/nature07228 . PMID 18668040 . S2CID 4429008 .
- ^ Baek D, Villén J, Shin C, Camargo FD, Gygi SP, Bartel DP (сентябрь 2008 г.). «Влияние микроРНК на выход белка» . Природа . 455 (7209): 64–71. Bibcode : 2008natur.455 ... 64b . doi : 10.1038/nature07242 . PMC 2745094 . PMID 18668037 .
- ^ Palmer EI, из Campos SG, Campos M, De Souza NC, ID Warrior, Carvalho AL, Marques MM (июль 2011 г.). «Механизмы и роль микронной дерегуляции в начале и прогрессировании рака» . Биология генетика и молекулярная . 34 (3): 363–70. Doi : 10.1590/s1415-47572011000300001 . PMC 3168173 . PMID 21931505 .
- ^ Бернштейн С, Бернштейн Х (май 2015). «Эпигенетическое восстановление репарации ДНК при прогрессировании на желудочно -кишечный рак» . Всемирный журнал желудочно -кишечной онкологии . 7 (5): 30–46. doi : 10.4251/wjgo.v7.i5.30 . PMC 4434036 . PMID 25987950 .
- ^ Maffioletti E, Tardito D, Gennarelli M, Bocchio-Chiavetto L (2014). «Микро -шпионы от мозга до периферии: новые подсказки из исследований микроРНК при нейропсихиатрических расстройствах» . Границы в клеточной нейробиологии . 8 : 75. DOI : 10.3389/fncel.2014.00075 . PMC 3949217 . PMID 24653674 .
- ^ Mellios N, Sur M (2012). «Новая роль микроРНК в расстройствах шизофрении и аутистического спектра» . Границы в психиатрии . 3 : 39. doi : 10.3389/fpsyt.2012.00039 . PMC 3336189 . PMID 22539927 .
- ^ Geaghan M, Cairns MJ (август 2015 г.). «МикроРНК и посттранскрипционная дисрегуляция в психиатрии» . Биологическая психиатрия . 78 (4): 231–9. doi : 10.1016/j.biopsych.2014.12.009 . HDL : 1959.13/1335073 . PMID 25636176 .
- ^ Барнетт Дж.А. (июль 2004 г.). «История исследований в дрожжах 7: ферментативная адаптация и регулирование» . Дрожжи . 21 (9): 703–46. doi : 10.1002/да.1113 . PMID 15282797 . S2CID 36606279 .
- ^ Dequéant ML, Pourquié O (май 2008 г.). «Сегментарная паттерна эмбриональной оси эмбриональной оси позвоночных». Природные обзоры. Генетика . 9 (5): 370–82. doi : 10.1038/nrg2320 . PMID 18414404 . S2CID 2526914 .
- ^ Гилберт С.Ф. (2003). Биология развития, 7 -е изд., Sunderland, Mass: Sinauer Associates, 65–6. ISBN 0-87893-258-5 .
- ^ Нехлин Джо, Карлберг М., Ронн Х (ноябрь 1991 г.). «Контроль генов дрожжей GAL с помощью Mig1 Represress: транскрипционный каскад в реакции глюкозы» . Embo Journal . 10 (11): 3373–7. doi : 10.1002/j.1460-2075.1991.tb04901.x . PMC 453065 . PMID 1915298 .
- ^ Chadle C, Fan J, Cho-Chung YS, Werner T, Ray J, Do L, et al. (Май 2005 г.). «Контроль экспрессии генов во время активации Т -клеток: альтернативная регуляция транскрипции мРНК и стабильность мРНК» . BMC Genomics . 6 : 75. DOI : 10.1186/1471-2164-6-75 . PMC 1156890 . PMID 15907206 .
- ^ Джексон Д.А., Помбо А, Иборра Ф. (февраль 2000 г.). «Баланс для транскрипции: анализ метаболизма ядерной РНК в клетках млекопитающих» . FASEB Journal . 14 (2): 242–54. doi : 10.1096/fasebj.14.2.242 . PMID 10657981 . S2CID 23518786 .
- ^ Schwanekamp JA, Sartor MA, Karyala S, Halbleib D, Medvedovic M, Tomlinson CR (2006). «Анализ по всему геному показывает, что ядерные и цитоплазматические уровни РНК по-разному влияют диоксин». Biochimica et Biophysica Acta (BBA) - структура и экспрессия гена . 1759 (8–9): 388–402. doi : 10.1016/j.bbaexp.2006.07.005 . PMID 16962184 .
Библиография
[ редактировать ]- Латчман, Дэвид С. (2005). Регуляция гена: эукариотическая перспектива . Психология пресса. ISBN 978-0-415-36510-9 .
Внешние ссылки
[ редактировать ]- База данных о транскрипционных факторах транскрипции и платформы по регулированию транскрипции растений и платформы анализа
- Регуляция экспрессии генов (сетка) в Национальной медицинской библиотеке Медицинской библиотеки США (Mesh)
- ChipBase Открытая база данных для декодирования транскрипционных регуляторных сетей некодирующих РНК и генов, кодирующих белок, из данных CHIP-seq.
| Базы данных управления авторитетом : национальный |
|---|