Белок оболочки коронавируса
| Белок-конверт | |||
|---|---|---|---|
 Модель внешнего строения вириона SARS-CoV-2 [1] ● Синий: конверт. ● Бирюзовый: спайковый гликопротеин (S). ● Ярко-розовый: белки оболочки (E). ● Зеленый: мембранные белки (М). ● Оранжевый: гликаны. | |||
| Идентификаторы | |||
| Символ | CoV_E | ||
| Пфам | PF02723 | ||
| ИнтерПро | ИПР003873 | ||
| PROSITE | ПС51926 | ||
| |||
Белок оболочки (E) является самым маленьким и наименее хорошо охарактеризованным из четырех основных структурных белков, обнаруженных в коронавируса вирионах . [2] [3] [4] Это интегральный мембранный белок длиной менее 110 аминокислотных остатков ; [2] В SARS-CoV-2 , возбудителе Covid-19 , белок Е имеет длину 75 остатков. [5] Хотя это не обязательно необходимо для репликации вируса , отсутствие белка Е может привести к аномальной сборке вирусных капсидов или снижению репликации. [2] [3] E — многофункциональный белок [6] и, в дополнение к своей роли структурного белка в вирусном капсиде , считается, что он участвует в сборке вируса, вероятно, действует как виропорин и участвует в вирусном патогенезе . [2] [5]
Структура
[ редактировать ]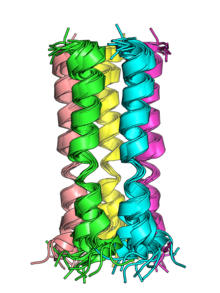
Белок Е состоит из короткой гидрофильной N-концевой области, гидрофобного спирального трансмембранного домена и несколько гидрофильной С-концевой области. У SARS-CoV и SARS-CoV-2 С-концевая область содержит PDZ-связывающий мотив (PBM). [2] [5] Эта особенность, по-видимому, сохраняется только в группах альфа- и бета -коронавирусов, но не в гамма-группах . [2] В бета- и гамма-группах консервативный остаток пролина обнаружен в С-концевой области, вероятно, участвующий в нацеливании белка на аппарат Гольджи . [2]
Трансмембранные спирали белков E SARS-CoV и SARS-CoV-2 могут олигомеризоваться , и, как было показано in vitro, они образуют пентамерные структуры с центральными порами, которые служат катион -селективными ионными каналами . [5] белка E обоих вирусов Пентамеры были структурно охарактеризованы с помощью спектроскопии ядерного магнитного резонанса . [5] [7]
Топология мембраны белка E была изучена у ряда коронавирусов с противоречивыми результатами; ориентация белка в мембране может быть переменной. [3] Совокупность данных предполагает, что наиболее распространенной ориентацией является С-конец, обращенный к цитоплазме . [8] Исследования белка E SARS-CoV-2 согласуются с этой ориентацией. [5] [9]
Посттрансляционные модификации
[ редактировать ]У некоторых, но не у всех коронавирусов белок Е посттрансляционно модифицируется путем пальмитоилирования консервативных остатков цистеина . [2] [8] В белке E SARS-CoV гликозилирования , который может влиять на топологию мембраны; наблюдался один сайт [8] однако функциональное значение гликозилирования E неясно. [2] Также было описано убиквитинирование SARS-CoV E, хотя его функциональное значение также неизвестно. [2]
Выражение и локализация
[ редактировать ]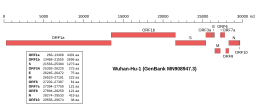 Геномная организация изолята Wuhan-Hu-1, самого раннего секвенированного образца SARS-CoV-2, с указанием местоположения гена E. | |
| NCBI Идентификатор генома | 86693 |
|---|---|
| Размер генома | 29 903 базы |
| Год завершения | 2020 |
| Геномный браузер ( UCSC ) | |
Белок Е экспрессируется в большом количестве в инфицированных клетках. Однако лишь небольшое количество общего продуцируемого белка Е обнаруживается в собранных вирионах . [2] [4] Белок Е локализован в эндоплазматическом ретикулуме , аппарате Гольджи и промежуточном компартменте эндоплазматической сети-Гольджи (ERGIC), внутриклеточном компартменте, который дает начало вирусной оболочке коронавируса . [2] [5]
Функция
[ редактировать ]Существенность
[ редактировать ]Исследования различных коронавирусов пришли к разным выводам о том, важен ли E для репликации вируса. для некоторых коронавирусов, включая MERS-CoV , E необходим. Сообщается, что [10] В других, включая мышиный коронавирус [11] и SARS-CoV, E не является существенным, хотя его отсутствие снижает титр вируса , [12] в некоторых случаях путем внесения дефектов распространения или возникновения аномальной морфологии капсида. [2]
Вирионы и сборка вируса
[ редактировать ]
Белок Е обнаруживается в собранных вирионах, где он образует межбелковые взаимодействия с мембранным белком коронавируса (М), наиболее распространенным из четырех структурных белков, содержащихся в вирусном капсиде . [2] [4] Взаимодействие между Е и М происходит через соответствующие С-концы на цитоплазматической стороне мембраны. [2] У большинства коронавирусов Е и М достаточно для образования вирусоподобных частиц . [2] [4] хотя сообщалось, что SARS-CoV зависит от N. также [14] Имеются убедительные доказательства того, что E участвует в индукции искривления мембраны для создания типичного сферического вириона коронавируса. [2] [15] Вполне вероятно, что E участвует в почковании или расщеплении вируса , хотя его роль в этом процессе недостаточно изучена. [2] [4] [15]
Виропорин
[ редактировать ]
В пентамерном состоянии E образует катион -селективные ионные каналы и, вероятно, действует как виропорин . [5] Исследования ЯМР показывают, что виропорин представляет открытую конформацию при низком pH или в присутствии ионов кальция, тогда как закрытая конформация предпочтительнее при основном pH. [16] Структура ЯМР показывает гидрофобные ворота на уровне лейцина 28 в середине поры. Считается, что прохождению ионов через ворота способствуют полярные остатки на С-конце. [17]
Утечка катионов может нарушить ионный гомеостаз , изменить проницаемость мембран и модулировать pH в клетке-хозяине, что может способствовать высвобождению вируса. [2] [4]
Роль белка Е как виропорина, по-видимому, участвует в патогенезе и может быть связана с активацией воспалительной сомы . [3] [18] При SARS-CoV мутации, которые нарушают функцию ионного канала E, приводят к ослаблению патогенеза на животных моделях, несмотря на незначительное влияние на рост вируса. [10]
Взаимодействие с белками хозяина
[ редактировать ]
Белково-белковые взаимодействия между E и белками в клетке-хозяине лучше всего описаны при SARS-CoV и происходят через мотив связывания C-концевого домена PDZ . Сообщается, что белок E SARS-CoV взаимодействует с пятью белками клетки-хозяина: Bcl-xL , PALS1 , синтенином , субъединицей натрий/калийной (Na+/K+) АТФазы α-1 и стоматином . [2] Взаимодействие с PALS1 может быть связано с патогенезом через возникающее в результате нарушение плотных контактов . [3] [10] Это взаимодействие также было выявлено в SARS-CoV-2 . [19]
Эволюция и сохранение
[ редактировать ]Последовательность белка Е недостаточно консервативна в разных родах коронавирусов: идентичность последовательностей достигает менее 30%. [12] В лабораторных экспериментах с вирусом гепатита мышей замена белков Е из разных коронавирусов, даже из разных групп, может привести к образованию жизнеспособных вирусов, что позволяет предположить, что в функциональных белках Е допускается значительное разнообразие последовательностей. [20] Белок E SARS-CoV-2 очень похож на белок SARS-CoV, с тремя заменами и одной делецией . [4] Исследование последовательностей SARS-CoV-2 показывает, что белок E развивается относительно медленно по сравнению с другими структурными белками. [21] Консервативная природа белка оболочки среди вариантов SARS-CoV и SARS-CoV-2 привела к тому, что его стали исследовать в качестве потенциальной мишени для разработки универсальной вакцины против коронавируса . [22] [23]
Ссылки
[ редактировать ]- ^ Солодовников Алексей; Архипова Валерия (29 июля 2021 г.). « Достоверно красиво: как мы сделали 3D-модель SARS-CoV-2». Н+1 . Архивировано из оригинала 30 июля 2021 г. Проверено 30 июля 2021 г.
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т Шуман Д., Филдинг, Британская Колумбия (май 2019 г.). «Белок оболочки коронавируса: современные знания» . Вирусологический журнал . 16 (1): 69. дои : 10.1186/s12985-019-1182-0 . ПМЦ 6537279 . ПМИД 31133031 .
- ^ Jump up to: а б с д и Шуман Д., Филдинг BC (03.09.2020). «Есть ли связь между патогенным белком оболочки коронавируса человека и иммунопатологией? Обзор литературы» . Границы микробиологии . 11 : 2086. doi : 10.3389/fmicb.2020.02086 . ПМЦ 7496634 . ПМИД 33013759 .
- ^ Jump up to: а б с д и ж г час Цао Ю, Ян Р., Ли И, Чжан В, Сунь Дж, Ван В, Мэн Икс (июнь 2021 г.). «Характеристика белка E SARS-CoV-2: последовательность, структура, виропорин и ингибиторы» . Белковая наука . 30 (6): 1114–1130. дои : 10.1002/pro.4075 . ПМЦ 8138525 . ПМИД 33813796 .
- ^ Jump up to: а б с д и ж г час я Мандала В.С., Маккей М.Дж., Щербаков А.А., Дрегни А.Дж., Колокурис А., Хонг М. (декабрь 2020 г.). «Структура и связывание лекарственного средства трансмембранного домена оболочечного белка SARS-CoV-2 в липидных бислоях» . Структурная и молекулярная биология природы . 27 (12): 1202–1208. дои : 10.1038/s41594-020-00536-8 . ПМЦ 7718435 . ПМИД 33177698 .
- ^ Лю Д.С., Юань Ц., Ляо Ю. (август 2007 г.). «Белок оболочки коронавируса: небольшой мембранный белок с множеством функций» . Клеточные и молекулярные науки о жизни . 64 (16): 2043–2048. дои : 10.1007/s00018-007-7103-1 . ПМК 7079843 . ПМИД 17530462 .
- ^ Сурья В., Ли Ю., Торрес Дж. (июнь 2018 г.). «Структурная модель Е-канала коронавируса SARS в мицеллах ЛМПГ» . Biochimica et Biophysical Acta (BBA) – Биомембраны . 1860 (6): 1309–1317. дои : 10.1016/j.bbamem.2018.02.017 . ПМК 7094280 . ПМИД 29474890 .
- ^ Jump up to: а б с Фунг Т.С., Лю Д.С. (июнь 2018 г.). «Посттрансляционные модификации белков коронавируса: роль и функции» . Будущая вирусология . 13 (6): 405–430. дои : 10.2217/fvl-2018-0008 . ПМК 7080180 . ПМИД 32201497 .
- ^ Дуарт Дж., Гарсиа-Муррия М.Дж., Грау Б., Акоста-Касерес Х.М., Мартинес-Хиль Л., Мингарро I (сентябрь 2020 г.). «Топология оболочечного белка SARS-CoV-2 в мембранах эукариот» . Открытая биология . 10 (9): 200209. doi : 10.1098/rsob.200209 . ПМЦ 7536074 . ПМИД 32898469 .
- ^ Jump up to: а б с ДеДиего М.Л., Грансон-Торрес Дж.Л., Хименес-Гуардино Х.М., Руле-Нава Х.А., Честнат-Родригес С., Фернандес-Дельгадо Р. и др. (декабрь 2014 г.). «Гены вирулентности коронавируса с упором на ген оболочки SARS-CoV» . Вирусные исследования 194 : 124–137. doi : 10.1016/j.virusres.2014.07.024 . ПМК 4261026 . ПМИД 25093995 .
- ^ Куо Л., Masters PS (апрель 2003 г.). «Белок Е с маленькой оболочкой не важен для репликации мышиного коронавируса» . Журнал вирусологии . 77 (8): 4597–4608. doi : 10.1128/JVI.77.8.4597-4608.2003 . ПМК 152126 . ПМИД 12663766 .
- ^ Jump up to: а б Рух Т.Р., Мачамер CE (март 2012 г.). «Белок Е коронавируса: сборка и не только» . Вирусы . 4 (3): 363–382. дои : 10.3390/v4030363 . ПМК 3347032 . ПМИД 22590676 .
- ^ Гудселл Д.С., Фойгт М., Зардецки С., Берли С.К. (август 2020 г.). «Интегративная иллюстрация работы с коронавирусом» . ПЛОС Биология . 18 (8): e3000815. дои : 10.1371/journal.pbio.3000815 . ПМЦ 7433897 . ПМИД 32760062 .
- ^ Сиу Ю.Л., Теох К.Т., Ло Дж., Чан С.М., Кьен Ф., Эскриу Н. и др. (ноябрь 2008 г.). «Структурные белки M, E и N коронавируса тяжелого острого респираторного синдрома необходимы для эффективной сборки, транспортировки и высвобождения вирусоподобных частиц» . Журнал вирусологии . 82 (22): 11318–11330. дои : 10.1128/JVI.01052-08 . ПМЦ 2573274 . ПМИД 18753196 .
- ^ Jump up to: а б Дж. Алсаади Э.А., Джонс М.М. (апрель 2019 г.). «Мембраносвязывающие белки коронавирусов» . Будущая вирусология . 14 (4): 275–286. дои : 10.2217/fvl-2018-0144 . ПМК 7079996 . ПМИД 32201500 .
- ^ Медейрос-Сильва Дж., Сомберг Н.Х., Ван Х.К., Маккей М.Дж., Мандала В.С., Дрегни А.Дж., Хонг М. (апрель 2022 г.). «РН- и кальций-зависимая ароматическая сеть в белке оболочки SARS-CoV-2» . Журнал Американского химического общества . 144 (15): 6839–6850. дои : 10.1021/jacs.2c00973 . ПМЦ 9188436 . ПМИД 35380805 .
- ^ Медейрос-Сильва Дж., Дрегни А.Дж., Сомберг Н.Х., Дуан П., Хонг М. (октябрь 2023 г.). «Атомная структура открытого виропорина SARS-CoV-2 E» . Достижения науки . 9 (41): eadi9007. дои : 10.1126/sciadv.adi9007 . ПМЦ 10575589 . ПМИД 37831764 .
- ^ Ньето-Торрес Х.Л., ДеДьего М.Л., Вердиа-Багена С., Хименес-Гуарденьо Х.М., Регла-Нава Х.А., Фернандес-Дельгадо Р. и др. (май 2014 г.). «Активность ионных каналов белка оболочки коронавируса при тяжелом остром респираторном синдроме способствует приспособленности и патогенезу вируса» . ПЛОС Патогены . 10 (5): e1004077. дои : 10.1371/journal.ppat.1004077 . ПМК 4006877 . ПМИД 24788150 .
- ^ Jump up to: а б Чай Дж., Цай Ю., Панг С., Ван Л., МакСвини С., Шанклин Дж., Лю К. (июнь 2021 г.). «Структурная основа распознавания белка оболочки SARS-CoV-2 белка PALS1 клеточного соединения человека» . Природные коммуникации . 12 (1): 3433. Бибкод : 2021NatCo..12.3433C . дои : 10.1038/s41467-021-23533-x . ПМЦ 8187709 . ПМИД 34103506 .
- ^ Куо Л., Херст К.Р., Мастерс PS (март 2007 г.). «Исключительная гибкость в требованиях к последовательности для функции белка малой оболочки коронавируса» . Журнал вирусологии . 81 (5): 2249–2262. дои : 10.1128/JVI.01577-06 . ПМК 1865940 . ПМИД 17182690 .
- ^ Рахман М.С., Хок М.Н., Ислам М.Р., Ислам И., Мишу И.Д., Рахаман М.М. и др. (март 2021 г.). «Мутационная информация о белке оболочки SARS-CoV-2» . Джин сообщает . 22 : 100997. doi : 10.1016/j.genrep.2020.100997 . ПМЦ 7723457 . ПМИД 33319124 .
- ^ Бхаттачарья С., Банерджи А., Рэй С. (март 2021 г.). «Разработка новой целевой вакцины против SARS-CoV2 с использованием белка оболочки (E): исследование, основанное на эволюционном, молекулярном моделировании и стыковке» . Международный журнал биологических макромолекул . 172 : 74–81. doi : 10.1016/j.ijbiomac.2020.12.192 . ПМЦ 7833863 . ПМИД 33385461 .
- ^ Чен Дж., Дэн Ю., Хуан Б., Хань Д., Ван В., Хуан М. и др. (24 февраля 2022 г.). «ДНК-вакцины, экспрессирующие белки оболочки и мембраны, обеспечивают частичную защиту от SARS-CoV-2 у мышей» . Границы в иммунологии . 13 : 827605. дои : 10.3389/fimmu.2022.827605 . ПМЦ 8907653 . ПМИД 35281016 .
