Cytochrome C -оксидаза субъединица I
| COX1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | COX1 , кодируемая митохондриально цитохромом С оксидаза I, COI, MTCO1, Основная субъединица цитохрома с оксидазы, CO I, цитохрома C -оксидаза IBUNIT I | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Омим : 516030 ; MGI : 102504 ; Гомологен : 5016 ; GeneCards : Cox1 ; OMA : COX1 - Ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викидид | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||


| Cytochrome C -оксидаза субъединица I | |||
|---|---|---|---|
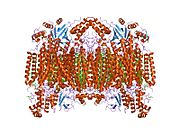
 Структура 13-субъединичной окисленной цитохрома С оксидазы. [ 5 ] | |||
| Идентификаторы | |||
| Символ | COX1 или COI | ||
| Pfam | PF00115 | ||
| InterPro | IPR000883 | ||
| PROSITE | PDOC00074 | ||
| Краткое содержание | 1occ / scope / supfam | ||
| TCDB | 3.D.4 | ||
| OPM Суперсемейство | 4 | ||
| OPM белок | 1v55 | ||
| CDD | CD01663 | ||
| |||

Цитохром С оксидаза I ( COX1 ), также известная как кодируемая митохондриально цитохромом С оксидаза I ( MT-CO1 ), представляет собой белок , который кодируется MT-CO1 геном у эукариотов . [ 6 ] Ген также называется COX1 , CO1 или COI . [ 7 ] Цитохром С оксидаза I является основной субъединицей комплекса цитохрома С оксидазы . У людей мутации в MT-CO1 были связаны с наследственной оптической невропатией Лебера (LHON), приобретенной идиопатической сидеобластической анемией , дефицитом комплекса IV , колоректальным раком , сенсорной глухотой и рецидивирующей миоглобинурией . [ 8 ] [ 9 ] [ 10 ]
Структура
[ редактировать ]У людей ген MT -CO1 расположен от нуклеотидных пар 5904 до 7444 на Гуанина тяжелой (H) секции (H) мтДНК . Ген -продукт представляет собой белок 57 кДа, состоящий из 513 аминокислот . [ 11 ] [ 12 ]
Функция
[ редактировать ]Субъединица I (CO1 или MT-CO1) цитохрома C (CO1 или MT-CO1) является одной из трех кодируемых субъединиц митохондриальной ДНК (MTDNA) (MT-CO1, MT-CO2 , MT-CO3 ) цитохрома-оксидазы , также известной как комплекс IV . Цитохром С оксидаза ( EC 1.9.3.1 ) является ключевым ферментом в аэробном метаболизме. Это третий и последний фермент цепи транспорта электронов митохондриальной окислительной фосфорилирования . [ 6 ]
Протонные накачивающие гемо-коппер оксидазы представляют терминальные, энергетические ферменты дыхательных цепей у прокариот и эукариот . Бинуклеарный центр Cub-Heme A3 (или Heme O), связанный с самой большой субъединицей I цитохрома C и убиквинол оксидаз ( EC 1.10.3.10 ), непосредственно участвует в связи между восстановлением диоксигена и протонной насоной. [ 13 ] [ 14 ] Некоторые терминальные оксидазы генерируют трансмембранную протонную градиент через плазматическую мембрану (прокариоты) или митохондриальную внутреннюю мембрану (эукариоты).
Ферментный комплекс состоит из 3-4 субъединиц (прокариот) до 13 полипептидов (млекопитающих), из которых только каталитическая субъединица (эквивалентная субъединице млекопитающих I (COI)) находится во всех респираторных оксидазах млекопитающих. Присутствие биметаллического центра (образованного хем с высокой точкой и медью B), а также хем с низким путем, оба лигировались со шестью консервативными остатками гистидина вблизи внешней стороны четырех трансмембранных пролетов внутри COI, являются общими для всех членов семьи. Полем [ 15 ] [ 16 ] [ 17 ] В отличие от эукариот, дыхательная цепь прокариот разветвлена до нескольких терминальных оксидаз. Ферментные комплексы варьируются по составу гема и меди, типа субстрата и аффинности субстрата. Различные дыхательные оксидазы позволяют клеткам настраивать свои дыхательные системы в соответствии с различными условиями роста окружающей среды. [ 13 ]
Было показано, что эубактериальная хинол оксидаза была получена из цитохрома С оксидазы в грамположительных бактериях и что архебактериальная хинол оксидаза имеет независимое происхождение. Значительное количество доказательств свидетельствует о том, что псевдомонадота (также известная как протеобактерии или фиолетовые бактерии) приобретали хинол оксидазу посредством бокового переноса гена из грамположительных бактерий . [ 13 ]
Связанный азот-оксидредуктаза ( EC 1.7.99.7 ) существует у денитрифицирующих видов архей и эубактерий и является гетеродимером цитохромов B и C. Метосульфат феназина может действовать как акцептор. Было высказано предположение, что цитохрома С оксидаза каталитические субъединицы эволюционировали из древних оксид -редуктаз оксида азота, которые могли бы уменьшить как азот, так и кислород. [ 18 ] [ 19 ]
Клиническое значение
[ редактировать ]Мутации в этом гене у людей связаны с наследственной зрительной невропатией Лебера (LHON), приобретенной идиопатической сидеобластической анемией , комплекса IV дефицитом , колоректальным раком , сенсоринеральной глухотой и рецидивирующей миоглобинурией . [ 8 ] [ 9 ] [ 10 ]
Наследственная оптическая невропатия Лебера (Лхон)
[ редактировать ]Лхон, коррелированный с мутациями в MT-CO1 , характеризуется дисфункцией зрительного нерва , вызывая подостленную или острая потеря центрального зрения . Некоторые пациенты могут демонстрировать неврологические или сердечные дефекты проводимости. Поскольку это заболевание является результатом мутаций митохондриальной ДНК, влияющих на комплексы дыхательной цепи , оно унаследовано матерински . [ 20 ] [ 9 ] [ 10 ]
Приобретенная идиопатическая сидеробластическая анемия
[ редактировать ]MT-CO1 может быть вовлечен в разработку приобретенной идиопатической сидеобластической анемии. ДНК могут вызывать дисфункцию дыхательной цепи, предотвращая восстановление железа железа железа , до железного что необходимо для окончательного стадии митохондриального биосинтеза гема Мутации в митохондриальной . Результатом является накопление железа в митохондриях и недостаточное производство гема. [ 21 ] [ 22 ] [ 9 ] [ 10 ]
Дефицит митохондриального комплекса IV (MT-C4D)
[ редактировать ]Мутации в этом гене могут вызывать дефицит митохондриального комплекса IV, заболевания митохондриальной дыхательной цепи, демонстрирующей широкий спектр клинических проявлений, начиная от изолированной миопатии до тяжелой мультисистемной болезни, затрагивающей множественные органы и ткани. Симптомы могут включать дисфункцию печени и гепатомегалию , гипотонию , мышечную слабость , непереносимость физических упражнений , отсроченное моторное развитие , умственную замедление , задержку развития и гипертрофическую кардиомиопатию . У некоторых пациентов гипертрофическая кардиомиопатия является смертельной на стадии новорожденных . Другие пострадавшие люди могут проявить болезнь Ли . [ 23 ] [ 24 ] [ 9 ] [ 10 ]
Колоректальный рак (CRC)
[ редактировать ]Мутации MT-CO1 играют роль в колоректальном раке, очень сложном заболевании, демонстрирующем злокачественные поражения во внутренних стенках толстой кишки и прямой кишки . Многочисленные такие генетические изменения часто связаны с прогрессированием аденомы или предраченочных поражений для инвазивной аденокарциномы . Давний язвенной колит , полипы толстой кишки и семейный анамнез являются факторами риска колоректального рака. [ 25 ] [ 26 ] [ 9 ] [ 10 ]
Рецидивирующая миоглобинурия митохондрия (RM-MT)
[ редактировать ]RM-MT-это заболевание, которое характеризуется рецидивирующими атаками рабдомиолиза (некроз или дезинтеграция скелетных мышц), связанных с мышечной болью и слабостью, непереносимостью физических упражнений, низкой мышечной способностью для окислительного фосфорилирования и с последующей экскременты миоглобина в моче. Это было связано с митохондриальной миопатией. Мутация G5920A и гетероплазматическая бессмысленная мутация G6708A были связаны с дефицитом Кокса и RM-MT. [ 27 ] [ 28 ] [ 9 ] [ 10 ]
Глухота, сенсориневральная, митохондриальная (DFNM)
[ редактировать ]DFNM является формой несиндромальной глухоты с материнским наследством . Пострадавшие люди проявляют прогрессирующую, постлингальную, сенсонирусную потерю слуха с высокими частотами. Мутация A1555G была связана с этим заболеванием. [ 29 ] [ 9 ] [ 10 ]
Подсемейства
[ редактировать ]- Цитохром С оксидаза CBB3-тип, субъединица I InterPro : IPR004677
- Цитохром o убихинол оксидаза, субъединица I InterPro : IPR014207
- Цитохром AA3 Кинол оксидаза, субъединица I InterPro : IPR014233
- Цитохрома С оксидаза, субъединица I бактериального типа InterPro : IPR014241
Использовать в штрих -кодировании ДНК
[ редактировать ]MT-CO1 -это ген, который часто используется в качестве штрих-кода ДНК для идентификации видов животных. Последовательность гена MT-CO1 подходит для этой роли, потому что его скорость мутаций , как правило, достаточно быстрая, чтобы различать тесно связанные виды , а также потому, что его последовательность сохраняется среди конспецифических. В отличие от первичного возражения, поднятого скептиками о том, что различия в последовательности MT-CO1 слишком малы, чтобы их можно было обнаружить, между близкими родственными видами, как правило, более 2% дивергенции последовательностей обычно обнаруживается между близкородственными видами животных, [ 30 ] предполагая, что штрих -код эффективен для большинства животных. Однако в большинстве, если не во всех семенах , скорость эволюции MT-CO1 очень медленная. Также было высказано предположение, что MT-CO1 может быть лучшим геном для штрих-кодирования ДНК почвенных грибов, чем его (ген чаще всего используется для микологического штрих-кодирования). [ 31 ]
MT-COI (= CCOI) в криптах толстой кишки
[ редактировать ]
Белок MT-COI, также известный как CCOI, обычно экспрессируется на высоком уровне в цитоплазме криптов толстой кишки человека большой кишки (толстой кишки). Тем не менее, MT-COI часто теряется в криптах толстой кишки с возрастом у людей и также часто отсутствует в полевых дефектах, которые приводят к раку толстой кишки, а также в частях рака толстой кишки. [ 32 ]
Эпителиальная внутренняя поверхность толстой кишки акцентирована инвагинациями, криптами толстой кишки. Крипты толстой кишки имеют форму пробирок с толстой стенкой с микроскопическими толстыми стенками с центральным отверстием вниз по длине трубки ( просвет склепа ). Четыре тканевые срезы показаны на изображении на этом разделе, две вырезанные по длинным осям крипт и два вырезанных параллельно длинным осям.
Большинство криптов толстой кишки человека на изображениях имеют высокую экспрессию коричневого окрашенного MT-COI. Тем не менее, в некоторых криптах толстой кишки у всех клеток не хватает MT-COI, и они кажутся в основном белыми, а их основным цветом является голубое окрашивание ядрам на внешних стенках криптов. Greaves et al. [ 33 ] показали, что недостатки MT-COI в криптах толстой кишки обусловлены мутациями в гене MT-COI. Как видно на панели B, часть стволовых клеток трех крипт, по-видимому, имеет мутацию в MT-COI, так что от 40% до 50% клеток, возникающих из этих стволовых клеток, образуют белый сегмент в зоне поперечного разреза Полем
У людей процент кишечников, дефицитных для MT-COI, составляет менее 1% до 40 лет, но затем линейно увеличивается с возрастом. [ 32 ] В среднем процент криптов толстой кишки, дефицитной для MT-COI, достигает 18% у женщин и 23% у мужчин в возрасте 80–84 лет. [ 32 ] Опухоли толстой кишки часто возникают в области криптов, содержащих большой кластер (до 410) криптов с дефицитом MT-COI. При раке толстой кишки до 80% опухолевых клеток может быть дефицит при MT-COI. [ 32 ]
Как видно на панелях C и D, крипты составляют около 75 до 110 клеток. Средняя окружность крипта составляет 23 ячейки. [ 34 ] Основываясь на этих измерениях, крипты имеют между 1725 и 2530 клетками. Другой отчет дал диапазон от 1500 до 4900 клеток на крипту толстой кишки. [ 35 ]
Появление частых криптов с почти полной потерей MT-COI в их 1700-5000 клетках предполагает процесс естественного отбора. Тем не менее, было также показано, что дефицит в конкретном склепе из -за начальной мутации митохондриальной ДНК может иногда происходить посредством стохастического процесса. [ 36 ] [ 37 ] Тем не менее, частое возникновение дефицита MT-COI во многих криптах в эпителии толстой кишки указывает на то, что отсутствие MT-COI, вероятно, дает избирательное преимущество.
MT-COI кодируется митохондриальной хромосомой . Существует множество копий хромосомы в большинстве митохондрий, обычно от 2 до 6 на митохондрион. [ 38 ] [ 39 ] [ 40 ] Если мутация возникает в MT-COI в одной хромосоме митохондриона, может быть случайная сегрегация хромосом во время деления митохондрий для создания новых митохондрий. Это может привести к митохондриону, прежде всего, или исключительно исключительно MT-COI-мутированных хромосом.
Митохондрион с в значительной степени MT-COI-мутированные хромосомы должен был иметь положительное смещение отбора , чтобы часто становиться основным типом митохондриона в клетке (клетка с гомоплазмией с дефицитом MT-COI ). В зависимости от типа клеток насчитывается от 100 до 700 митохондрий, в зависимости от типа ячейки. [ 39 ] [ 40 ] Кроме того, существует довольно быстрый оборот митохондрий, так что митохондрион с хромосомами с мутированными MT-COI и положительным смещением отбора может вскоре стать основным типом митохондриона в клетке. Средний период полураспада митохондрий у крыс, в зависимости от типа клеток, составляет от 9 до 24 дней, [ 41 ] и у мышей около 2 дней. [ 42 ] У людей вполне вероятно, что половина жизни митохондрий также в течение нескольких дней до недель.
Стволовая клетка у основания крипта толстой кишки, которая была в значительной степени с дефицитом MT-COI, может конкурировать с остальными 4 или 5 стволовыми клетками, чтобы захватить нишу стволовых клеток. Если это произойдет, то крипта толстой кишки будет дефицит в MT-COI во всех 1700-5000 ячеек, как указано для некоторых криптов на панелях A, B и D изображения.
Крипты толстой кишки могут воспроизводиться путем деления, как видно на панели C, где склеп делятся, чтобы сформировать два крипта, и на панели B, где, по крайней мере, один склеп, кажется, делятся. Большинство криптов с дефицитом в MT-COI находятся в кластерах криптов (клонов криптов) с двумя или более скрипкой с дефицитом MT-COI, прилегающими друг к другу (см. Панель D). [ 32 ] Это иллюстрирует, что часто возникают клоны дефицитных склепов, и, следовательно, существует положительное селективное смещение, которое позволило им распространяться в эпителии толстой кишки человека.
Неясно, почему дефицит MT-COI должен иметь положительный селективный уклон. Одно предложение [ 32 ] является тем, что дефицит MT-COI в митохондрионе приводит к более низкой реактивной выработке кислорода (и меньшему окислительному повреждению), и это обеспечивает селективное преимущество в конкуренции с другими митохондриями в одной и той же клетке для генерации гомоплазма для дефицита MT-COI. Другим предположением было то, что клетки с дефицитом цитохрома С оксидазы устойчивы к апоптозу и, следовательно, с большей вероятностью выживают. Связь MT-COI с апоптозом возникает из-за того, что активный цитохром С оксидаза окисляет цитохром С, который затем активирует про-каспазу 9, что приводит к апоптозу. [ 43 ] Эти два фактора могут способствовать частому возникновению криптов с дефицитом MT-COI с возрастом или во время канцерогенеза в толстой кишке человека.
Взаимодействия
[ редактировать ]цитохрома с оксидазы) кодированный белок взаимодействует В рамках комплекса и SMIM20 / Mitrac7 , кодируемый белок взаимодействует с с COA3 COA3 и SMIM20 / Mitrac7 . Это взаимодействие с SMIM20 стабилизирует недавно синтезированный MT-CO1 и предотвращает его преждевременный оборот . [ 44 ] Кроме того, он взаимодействует с TMEM177 в зависимости от COX20 . [ 45 ] [ 9 ] [ 10 ]
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в GRCH38: Ensembl Release 89: ENSG00000198804 - Ensembl , май 2017 г.
- ^ Jump up to: а беременный в GRCM38: Ensembl Release 89: Ensmusg00000064351 - Ensembl , май 2017 г.
- ^ «Человеческая PubMed ссылка:» . Национальный центр информации о биотехнологии, Национальная медицина США .
- ^ «Мышь Pubmed ссылка:» . Национальный центр информации о биотехнологии, Национальная медицина США .
- ^ Цукихара Т., Аояма Х., Ямашита Е., Томизаки Т., Ямагучи Х., Синдзава-Ито К. и др. (Май 1996). «Вся структура 13-субъединичной окисленной цитохрома С оксидазы при 2,8 А». Наука . 272 (5265): 1136–1144. Bibcode : 1996sci ... 272.1136t . doi : 10.1126/science.272.5265.1136 . PMID 8638158 . S2CID 20860573 .
- ^ Jump up to: а беременный «Ген Entrez: субъединица цитохрома С оксидазы I» .
- ^ Косакьян А., Хегер Т.Дж., Леандер Б.С., Тодоров М., Митчелл Е.А., Лара Е (май 2012 г.). «Ши штрих -кодировки небелидных испытательных амеб (амебозоа: arcellinida): обширное загадочное разнообразие и переопределение Hyalospheniidae Schultze» (PDF) . Профист . 163 (3): 415–434. doi : 10.1016/j.protis.2011.10.003 . PMID 22130576 .
- ^ Jump up to: а беременный Онлайн -наследство Менделян в Человеке (Омим): 516030
- ^ Jump up to: а беременный в дюймовый и фон глин час я «MT -CO1 - Цитохрома C -оксидаза субъединица 1 - Homo Sapiens (Human) - ген и белок MT -CO1» . www.uniprot.org . Получено 2018-08-13 .
 Эта статья включает в себя текст, доступный по лицензии CC по 4,0 .
Эта статья включает в себя текст, доступный по лицензии CC по 4,0 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я «Uniprot: универсальная база знаний белка» . Исследование нуклеиновых кислот . 45 (D1): D158 - D169. Январь 2017 года. DOI : 10.1093/nar/gkw1099 . PMC 5210571 . PMID 27899622 .
- ^ Zong NC, Li H, Li H, Lam MP, Jimenez RC, Kim CS, et al. (Октябрь 2013). «Интеграция биологии и медицины сердечного протеома специализированной базой знаний» . Исследование циркуляции . 113 (9): 1043–1053. doi : 10.1161/circresaha.113.301151 . PMC 4076475 . PMID 23965338 .
- ^ «MT -CO1 - цитохрома C -оксидаза субъединица 1» . Сердечный органеллерный белок Атлас База знаний (copakb) . [ Постоянная мертвая ссылка ]
- ^ Jump up to: а беременный в García-Horsman JA, Barquera B, Rumbley J, Ma J, Gennis RB (сентябрь 1994 г.). «Суперсемейство дыхательных оксидаз гем-коллеги» . Журнал бактериологии . 176 (18): 5587–5600. doi : 10.1128/jb.176.18.5587-5600.1994 . PMC 196760 . PMID 8083153 .
- ^ Papa S, Capitanio N, Glaser P, Villani G (май 1994). «Протонный насос гем-коллеги оксидазы». Cell Biology International . 18 (5): 345–355. doi : 10.1006/cbir.1994.1084 . PMID 8049679 . S2CID 36428993 .
- ^ Castresana J, Lübben M, Saraste M, Higgins DG (июнь 1994 г.). «Эволюция цитохромоксидазы, фермента, старше атмосферного кислорода» . Embo Journal . 13 (11): 2516–2525. doi : 10.1002/j.1460-2075.1994.tb06541.x . PMC 395125 . PMID 8013452 .
- ^ Капальди Р.А., Малатеста Ф., Дарли-Усмар Вм (июль 1983 г.). «Структура цитохрома С оксидазы». Biochimica et Biophysica Acta (BBA) - обзоры биоэнергетики . 726 (2): 135–148. doi : 10.1016/0304-4173 (83) 90003-4 . PMID 6307356 .
- ^ Холм Л., Сарасте М., Виксстрем М (сентябрь 1987 г.). «Структурные модели окислительно -восстановительных центров в цитохромоксидазе» . Embo Journal . 6 (9): 2819–2823. doi : 10.1002/j.1460-2075.1987.tb02578.x . PMC 553708 . PMID 2824194 .
- ^ Saraste M, Castresana J (март 1994 г.). «Цитохромсидаза эволюционировала путем возиться с денитрификационными ферментами» . Письма Febs . 341 (1): 1–4. doi : 10.1016/0014-5793 (94) 80228-9 . PMID 8137905 . S2CID 1248917 .
- ^ Chen J, Strous M (февраль 2013 г.). «Денитрификация и аэробное дыхание, гибридные электронные транспортные цепочки и коэволюция» . Biochimica et Biophysica Acta (BBA) - Биоэнергетика . 1827 (2): 136–144. doi : 10.1016/j.bbabio.2012.10.002 . PMID 23044391 .
- ^ Браун М.Д., Ян С.К., Торнс I, Торрони А., Лотт М.Т., Уоллес, округ Колумбия (август 1992 г.). «Вариант митохондриальной ДНК, идентифицированный у пациентов с наследственной невропатией в области зрителей, который расширяет аминокислотную последовательность субъединицы I -оксидазы цитохрома С» . Американский журнал человеческой генетики . 51 (2): 378–385. PMC 1682694 . PMID 1322638 .
- ^ Gattermann N, Retzlaff S, Wang YL, Hofhaus G, Heinisch J, Aul C, Schneider W (декабрь 1997 г.). «Гетероплазматические точечные мутации митохондриальной ДНК, влияющей на субъединицу I цитохрома с оксидазы у двух пациентов с приобретенной идиопатической сидеобластической анемией» . Кровь . 90 (12): 4961–4972. doi : 10.1182/blood.v90.12.4961 . PMID 9389715 .
- ^ Bröker S, Meunier B, Rich P, Gattermann N, Hofhaus G (ноябрь 1998). «Мутации мтДНК, связанные с сидеробластной анемией, вызывают дефект митохондриальной цитохрома с оксидазы». Европейский журнал биохимии . 258 (1): 132–138. doi : 10.1046/j.1432-1327.1998.2580132.x . PMID 9851701 .
- ^ Varlamov DA, Kudin AP, Vielhaber S, Schröder R, Sassen R, Becker A, et al. (Август 2002). «Метаболические последствия новой миссенс -мутации гена MtDna Co I» . Молекулярная генетика человека . 11 (16): 1797–1805. doi : 10.1093/hmg/11.16.1797 . PMID 12140182 .
- ^ Люсиоли С., Хоффмайер К., Карройццо Р., Тесса А., Людвиг Б., Санторелли Ф.М. (март 2006 г.). «Введение новой мутации мтДНК человека в ген Paracoccus denitrificans Cox I объясняет функциональный дефицит у пациента». Нейрогентика . 7 (1): 51–57. doi : 10.1007/s10048-005-0015-z . PMID 16284789 . S2CID 21304246 .
- ^ Greaves LC, Preston SL, Tadrous PJ, Taylor RW, Barron MJ, Oukrif D, et al. (Январь 2006 г.). «Митохондриальные ДНК -мутации установлены в стволовых клетках толстой кишки человека, а мутированные клоны расширяются путем деления крипты» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (3): 714–719. Bibcode : 2006pnas..103..714G . doi : 10.1073/pnas.0505903103 . PMC 1325106 . PMID 16407113 .
- ^ Namslauer I, Brzezinski P (март 2009 г.). «Митохондриальная ДНК -мутация, связанная с раком толстой кишки, приводит к утечкам протонов в цитохроме С оксидазы» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (9): 3402–3407. Bibcode : 2009pnas..106.3402n . doi : 10.1073/pnas.0811450106 . PMC 2651238 . PMID 19218458 .
- ^ Карадимас К.Л., Гринштейн П., Сью С.М., Джозеф Дж.Т., Танджи К., Халлер Р.Г. и др. (Сентябрь 2000). «Рецидивирующая миоглобинурия из -за бессмысленной мутации в гене Кокс I митохондриальной ДНК». Неврология . 55 (5): 644–649. doi : 10.1212/wnl.55.5.644 . PMID 10980727 . S2CID 26776388 .
- ^ Kollberg G, Moslemi AR, Lindberg C, Holme E, Oldfors A (февраль 2005 г.). «Митохондриальная миопатия и рабдомиолиз, связанные с новой бессмысленной мутацией в гене, кодирующем цитохромоксидазой -субъединицу I» . Журнал невропатологии и экспериментальной неврологии . 64 (2): 123–128. doi : 10.1093/jnen/64.2.123 . PMID 15751226 .
- ^ Pandya A, Xia XJ, Erdenetungalag R, Amendola M, Landa B, Radnaabazar J, et al. (Декабрь 1999). «Гетерогенные точечные мутации в предшественнике митохондриальной тРНК Ser (UCN) сосуществуют с мутацией A1555G у глухих учащихся из Монголии» . Американский журнал человеческой генетики . 65 (6): 1803–1806. doi : 10.1086/302658 . PMC 1288397 . PMID 10577941 .
- ^ Hebert PD, Ratnasingham S, Dewaard Jr (август 2003 г.). «Жизнь штрих -кодирования: субъединица цитохрома С оксидазы 1 среди близкородственных видов» . Разбирательство. Биологические науки . 270 (Suppl 1): S96 - S99. doi : 10.1098/rsbl.2003.0025 . PMC 1698023 . PMID 12952648 .
- ^ Molitor C, Inthavong B, Sage L, Geremia RA, Mouhamadou B (январь 2010 г.). «Потенциал гена COX1 в таксономическом разрешении почвенных грибов» . Письма микробиологии FEMS . 302 (1): 76–84. doi : 10.1111/j.1574-6968.2009.01839.x . PMID 19909345 .
- ^ Jump up to: а беременный в дюймовый и фон глин Бернштейн С., Фасиста А., Нгуен Х., Зейтлин Б., Хассауна Н., Лустаунау С. и др. (Декабрь 2010). «Недостаток рака и возраста, связанный с возрастом, в цитохроме С оксидазы I» . Всемирный журнал желудочно -кишечной онкологии . 2 (12): 429–442. doi : 10.4251/wjgo.v2.i12.429 . PMC 3011097 . PMID 21191537 .
- ^ Greaves LC, Preston SL, Tadrous PJ, Taylor RW, Barron MJ, Oukrif D, et al. (Январь 2006 г.). «Митохондриальные ДНК -мутации установлены в стволовых клетках толстой кишки человека, а мутированные клоны расширяются путем деления крипты» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (3): 714–719. Bibcode : 2006pnas..103..714G . doi : 10.1073/pnas.0505903103 . PMC 1325106 . PMID 16407113 .
- ^ Бейкер А.М., Цересер Б., Мелтон С., Флетчер А.Г., Родригес-Джасто М., Тадрус П.Дж. и др. (Август 2014). «Количественная оценка эволюции крипты и стволовых клеток в нормальной и опухолевой толстой кишке» . Сотовые отчеты . 8 (4): 940–947. doi : 10.1016/j.celrep.2014.07.019 . PMC 4471679 . PMID 25127143 .
- ^ Nooteboom M, Johnson R, Taylor RW, Wright NA, Lightowlers RN, Kirkwood TB, et al. (Февраль 2010 г.). «Возрастные мутации митохондриальной ДНК приводят к небольшим, но значительным изменениям в пролиферации клеток и апоптозе в криптах толстой кишки человека» . Старение ячейки . 9 (1): 96–99. doi : 10.1111/j.1474-9726.2009.00531.x . PMC 2816353 . PMID 19878146 .
- ^ Коллер HA, Bodyak ND, Khrapko K (апрель 2002 г.). «Частые внутриклеточные клональные экспансии соматических мутаций мтДНК: значимость и механизмы». Анналы нью -йоркской академии наук . 959 (1): 434–447. Bibcode : 2002nyasa.959..434c . doi : 10.1111/j.1749-6632.2002.tb02113.x . PMID 11976216 . S2CID 40639679 .
- ^ Nekhaeva E, Bodyak ND, Kraytsberg Y, McGrath SB, Van Orsouw NJ, Pluzhnikov A, et al. (Апрель 2002 г.). «Клонально расширенные мутации точек мтДНК в изобилии в отдельных клетках тканей человека» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (8): 5521–5526. Bibcode : 2002pnas ... 99.5521n . doi : 10.1073/pnas.072670199 . PMC 122802 . PMID 11943860 .
- ^ Legros F, Malka F, Frachon P, Lombès A, Rojo M (июнь 2004 г.). «Организация и динамика человеческой митохондриальной ДНК». Журнал сотовой науки . 117 (Pt 13): 2653–2662. doi : 10.1242/jcs.01134 . PMID 15138283 . S2CID 14335558 .
- ^ Jump up to: а беременный Робин Эд, Вонг Р. (сентябрь 1988 г.). «Молекулы митохондриальной ДНК и виртуальное количество митохондрий на клетку в клетках млекопитающих». Журнал клеточной физиологии . 136 (3): 507–513. doi : 10.1002/jcp.1041360316 . PMID 3170646 . S2CID 2841036 .
- ^ Jump up to: а беременный Сато М, Куроива Т (сентябрь 1991 г.). «Организация множественных нуклеоидов и молекул ДНК в митохондриях человеческой клетки». Экспериментальные исследования клеток . 196 (1): 137–140. doi : 10.1016/0014-4827 (91) 90467-9 . PMID 1715276 .
- ^ Menzies RA, Gold PH (апрель 1971 г.). «Оборот митохондрий в различных тканях молодых взрослых и пожилых крыс» . Журнал биологической химии . 246 (8): 2425–2429. doi : 10.1016/s0021-9258 (18) 62305-1 . PMID 5553400 .
- ^ Miwa S, Lawless C, Von Zglinicki T (декабрь 2008 г.). «Митохондриальный оборот в печени быстрее in vivo и ускоряется путем диетического ограничения: применение простой динамической модели» . Старение ячейки . 7 (6): 920–923. doi : 10.1111/j.1474-9726.2008.00426.x . PMC 2659384 . PMID 18691181 .
- ^ Brown GC, Borutaite V (2008). «Регуляция апоптоза окислительно -восстановительным состоянием цитохрома С» . Biochimica et Biophysica Acta (BBA) - Биоэнергетика . 1777 (7–8): 877–881. doi : 10.1016/j.bbabio.2008.03.024 . PMID 18439415 .
- ^ Dennerlein S, Oeljeklaus S, Jans D, Hellwig C, Bareth B, Jakobs S, et al. (Сентябрь 2015). «Mitrac7 действует как специфичный для COX1 шаперон и раскрывает контрольную точку во время сборки цитохрома C оксидазы» . Сотовые отчеты . 12 (10): 1644–1655. doi : 10.1016/j.celrep.2015.08.009 . HDL : 11858/00-001M-0000-0028-466E-c . PMID 26321642 .
- ^ Lorenzi I, Oeljeklaus S, Aich A, Ronsör C, Sillegari S, Dudek J, et al. (Февраль 2018 г.). «Митохондриальный TMEM177 связан с COX20 во время биогенеза COX2 » Biochimica et Biophysica Acta (BB) - исследования молекулярных клеток 1865 (2): 323–3 Doi : 10.1016/ j.bmcr.2017.11.0 5764226PMC 29154948PMID
Дальнейшее чтение
[ редактировать ]- Торрони А., Ахилли А., Маколей В., Ричардс М., Бандельт Х.Дж. (июнь 2006 г.). «Сбор плодов человеческого дерева мтДНК». Тенденции в генетике . 22 (6): 339–345. doi : 10.1016/j.tig.2006.04.001 . PMID 16678300 .
- Bodenteich A, Mitchell LG, Polymeropoulos MH, Merril CR (май 1992). «Динуклеотид повторения в митохондриальной D-петле» человека ». Молекулярная генетика человека . 1 (2): 140. doi : 10.1093/hmg/1.2.140-A . PMID 1301157 .
- Браун М.Д., Ян С.К., Торнс I, Торрони А., Лотт М.Т., Уоллес, округ Колумбия (август 1992 г.). «Вариант митохондриальной ДНК, идентифицированный у пациентов с наследственной невропатией в области зрителей, который расширяет аминокислотную последовательность субъединицы I -оксидазы цитохрома С» . Американский журнал человеческой генетики . 51 (2): 378–385. PMC 1682694 . PMID 1322638 .
- Lu X, Walker T, Macmanus JP, Seligy VL (июль 1992 г.). «Дифференциация клеток аденокарциномы HT-29 толстой кишки человека коррелирует с повышенной экспрессией митохондриальной РНК: влияние трегалозы на рост и созревание клеток». РАНКА . 52 (13): 3718–3725. PMID 1377597 .
- Marzuki S, Noer AS, Lertrit P, Thyagarajan D, Kapsa R, Utthanaphol P, Byrne E (декабрь 1991). «Нормальные варианты митохондриальной ДНК и продуктов перевода человека: строительство базы эталонной данных». Человеческая генетика . 88 (2): 139–145. doi : 10.1007/bf00206061 . PMID 1757091 . S2CID 28048453 .
- Moraes CT, Andreetta F, Bonilla E, Shanske S, Dimauro S, Schon EA (март 1991 г.). «Компетентная репликация человеческая митохондриальная ДНК отсутствует в области промотора тяжелой цепи» . Молекулярная и клеточная биология . 11 (3): 1631–1637. doi : 10.1128/mcb.11.3.1631 . PMC 369459 . PMID 1996112 .
- Attardi G, Chomyn A, Doolittle RF, Mariottini P, Ragan CI (1987). «Семь неопознанных кадров чтения митохондриальной ДНК человека кодируют субъединицы дыхательной цепи Nadh дегидрогеназы». Симпозии Cold Spring Harbor по количественной биологии . 51 Pt 1 (1): 103–114. doi : 10.1101/sqb.1986.051.01.013 . PMID 3472707 .
- Chomyn A, Cleeter MW, Ragan CI, Riley M, Doolittle RF, Attardi G (октябрь 1986 г.). «URF6, последняя неопознанная рамка считывания мтДНК человека, кодирует субъединицу NADH дегидрогеназы». Наука . 234 (4776): 614–618. Bibcode : 1986sci ... 234..614c . doi : 10.1126/science.3764430 . PMID 3764430 .
- Chomyn A, Mariottini P, Cleeter MW, Ragan CI, Matsuno-Yagi A, Hatefi Y, et al. (1985). «Шесть неопознанных кадров чтения митохондриальной ДНК человека кодируют компоненты дыхательной цепь надхдегидрогеназы». Природа . 314 (6012): 592–597. Bibcode : 1985natur.314..592c . doi : 10.1038/314592A0 . PMID 3921850 . S2CID 32964006 .
- Сангер Ф., Коулсон А.Р., Баррелл Б.Г., Смит А.Дж., Роу Ба (октябрь 1980). «Клонирование в одноцепочечном бактериофаге как помощь для быстрого секвенирования ДНК». Журнал молекулярной биологии . 143 (2): 161–178. doi : 10.1016/0022-2836 (80) 90196-5 . PMID 6260957 .
- Монтойя Дж., Оджала Д., Аттарди Г. (апрель 1981). «Отличительные особенности 5'-концевых последовательностей человеческих митохондриальных мРНК». Природа . 290 (5806): 465–470. Bibcode : 1981natur.290..465m . doi : 10.1038/290465A0 . PMID 7219535 . S2CID 4358928 .
- Хорай С., Хаясака К., Кондо Р., Цуган К., Такахата Н. (январь 1995 г.). «Недавнее африканское происхождение современных людей, выявленное полными последовательностями гоминоидных митохондриальных ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 92 (2): 532–536. Bibcode : 1995pnas ... 92..532H . doi : 10.1073/pnas.92.2.532 . PMC 42775 . PMID 7530363 .
- Эндрюс Р.М., Кубака I, Чиннери П.Ф., Лайтуллерс Р.Н., Тернбулл Д.М., Хауэлл Н. (октябрь 1999 г.). «Реанализ и пересмотр справочной последовательности Кембриджа для митохондриальной ДНК человека» . Природа генетика . 23 (2): 147. doi : 10.1038/13779 . PMID 10508508 . S2CID 32212178 .
- Ингман М., Кассманн Х, Пябо С., Джилленстен У (декабрь 2000 г.). «Изменение митохондриального генома и происхождение современных людей». Природа . 408 (6813): 708–713. Bibcode : 2000natur.408..708i . doi : 10.1038/35047064 . PMID 11130070 . S2CID 52850476 .
- Finnilä S, Lehtonen MS, Majamaa K (июнь 2001 г.). «Пилогенетическая сеть для европейской мтДНК» . Американский журнал человеческой генетики . 68 (6): 1475–1484. Doi : 10.1086/320591 . PMC 1226134 . PMID 11349229 .
- Maca-Meyer N, González AM, Larruga JM, Flores C, Cabrera VM (2003). «Основные геномные митохондриальные линии определяют ранние экспансии человека» . BMC Genetics . 2 : 13. DOI : 10.1186/1471-2156-2-13 . PMC 55343 . PMID 11553319 .
PDB Галерея |
|---|












Эта статья включает в себя текст из Национальной медицины Соединенных Штатов , которая находится в общественном достоянии .