Базовая пара

Пара оснований ( п.н. ) — это фундаментальная единица двухцепочечных нуклеиновых кислот, состоящая из двух нуклеиновых оснований, связанных друг с другом водородными связями . Они образуют строительные блоки двойной спирали ДНК и участвуют в складчатой структуре как ДНК, так и РНК . Продиктованные определенным характером водородных связей , пары оснований «Уотсон-Крик» (или «Уотсон-Крик-Франклин») ( гуанин - цитозин и аденин - тимин ) [ 1 ] позволяют спирали ДНК сохранять регулярную спиральную структуру, которая тонко зависит от ее нуклеотидной последовательности . [ 2 ] Комплементарный закодированной характер этой парной структуры обеспечивает избыточную копию генетической информации, в каждой нити ДНК. Регулярная структура и избыточность данных, обеспечиваемая двойной спиралью ДНК, делают ДНК хорошо подходящей для хранения генетической информации, в то время как спаривание оснований между ДНК и входящими нуклеотидами обеспечивает механизм, с помощью которого ДНК-полимераза реплицирует ДНК, а РНК-полимераза транскрибирует ДНК в РНК. Многие ДНК-связывающие белки могут распознавать определенные закономерности спаривания оснований, которые идентифицируют определенные регуляторные области генов.
Внутримолекулярные пары оснований могут встречаться в составе одноцепочечных нуклеиновых кислот. Это особенно важно в молекулах РНК (например, транспортных РНК ), где пары оснований Уотсона-Крика (гуанин-цитозин и аденин- урацил ) допускают образование коротких двухцепочечных спиралей и широкий спектр взаимодействий, не относящихся к Уотсону-Крику. (например, G–U или A–A) позволяют РНК сворачиваться в широкий спектр специфических трехмерных структур . Кроме того, спаривание оснований между транспортной РНК (тРНК) и информационной РНК (мРНК) формирует основу для событий молекулярного распознавания , которые приводят к тому, что нуклеотидная последовательность мРНК транслируется в аминокислотную последовательность белков через генетический код .
Размер отдельного гена организма или всего генома часто измеряется в парах оснований, поскольку ДНК обычно двухцепочечная. Следовательно, общее количество пар оснований равно числу нуклеотидов в одной из цепей (за исключением некодирующих одноцепочечных участков теломер ). геном По оценкам , гаплоидный человека (23 хромосомы ) имеет длину около 3,2 миллиарда оснований и содержит 20 000–25 000 различных генов, кодирующих белки. [ 3 ] [ 4 ] [ 5 ] Килобаза ( кб ) — единица измерения в молекулярной биологии, равная 1000 парам оснований ДНК или РНК. [ 6 ] Общее количество пар оснований ДНК на Земле оценивается в 5,0 × 10. 37 весом 50 миллиардов тонн . [ 7 ] Для сравнения, общая масса биосферы ( триллион оценивается в 4 TtC тонн углерода ). [ 8 ]
Водородная связь и стабильность
[ редактировать ]
|

|
Водородная связь — это химическое взаимодействие, лежащее в основе описанных выше правил спаривания оснований. Соответствующее геометрическое соответствие доноров и акцепторов водородных связей позволяет стабильно образовывать только «правильные» пары. ДНК с высоким содержанием GC более стабильна, чем ДНК с низким содержанием GC. Однако важно отметить, что стэкинг-взаимодействия в первую очередь ответственны за стабилизацию двойной спиральной структуры; Вклад спаривания оснований Уотсона-Крика в глобальную структурную стабильность минимален, но его роль в специфичности, лежащей в основе комплементарности, напротив, имеет максимальное значение, поскольку это лежит в основе зависимых от матрицы процессов центральной догмы (например, репликации ДНК ). [ 9 ]
Более крупные азотистые основания , аденин и гуанин, относятся к классу химических структур с двойным кольцом, называемых пуринами ; меньшие азотистые основания, цитозин и тимин (и урацил), являются членами класса однокольцевых химических структур, называемых пиримидинами . Пурины комплементарны только пиримидинам: пары пиримидин-пиримидин энергетически невыгодны, поскольку молекулы расположены слишком далеко друг от друга для установления водородной связи; Пурин-пуриновые пары энергетически невыгодны, поскольку молекулы расположены слишком близко, что приводит к перекрывающемуся отталкиванию. Спаривание пурин-пиримидиновых оснований AT, GC или UA (в РНК) приводит к правильной структуре дуплекса. Единственными другими пурин-пиримидиновыми парами могут быть AC, GT и UG (в РНК); эти пары являются несовпадениями, поскольку структуры доноров и акцепторов водорода не совпадают. Спаривание GU с двумя водородными связями действительно встречается в РНК довольно часто (см. Пара оснований колебания ).
Парные молекулы ДНК и РНК сравнительно стабильны при комнатной температуре, но две нуклеотидные цепи разделяются при температуре выше точки плавления , которая определяется длиной молекул, степенью неправильного спаривания (если таковое имеется) и содержанием GC. Более высокое содержание GC приводит к более высоким температурам плавления; поэтому неудивительно, что геномы экстремофильных организмов, таких как Thermus thermophilus, особенно богаты GC. И наоборот, области генома, которые необходимо часто разделять — например, промоторные области часто транскрибируемых генов — сравнительно бедны GC (например, см. вставку TATA ). Содержание GC и температуру плавления также необходимо учитывать при разработке праймеров для ПЦР . реакций [ нужна ссылка ]
Примеры
[ редактировать ]Следующие последовательности ДНК иллюстрируют парные двухцепочечные структуры. По соглашению верхняя цепь записывается от 5'-конца к 3'-концу ; таким образом, нижняя цепь записывается от 3' до 5'.
- Последовательность ДНК, спаренная основаниями:
ATCGATTGAGCTCTAGCGTAGCTAACTCGAGATCGC
- Соответствующая последовательность РНК, в которой урацил заменен на тимин в цепи РНК:
AUCGAUUGAGCUCUAGCGUAGCUAACUCGAGAUCGC
Базовые аналоги и интеркалаторы
[ редактировать ]Химические аналоги нуклеотидов могут занять место собственных нуклеотидов и установить неканоническое спаривание оснований, что приводит к ошибкам (в основном точечным мутациям ) в репликации и транскрипции ДНК . Это связано с их изостерическим химическим составом. Одним из распространенных аналогов мутагенных оснований является 5-бромурацил , который напоминает тимин, но может образовывать пару оснований с гуанином в его енольной форме. [ 10 ]
Другие химические вещества, известные как интеркаляторы ДНК , встраиваются в зазор между соседними основаниями одной цепи и вызывают мутации сдвига рамки считывания , «маскируясь» под основание, заставляя механизм репликации ДНК пропускать или вставлять дополнительные нуклеотиды в интеркалированный сайт. Большинство интеркаляторов представляют собой крупные полиароматические соединения и являются известными или предполагаемыми канцерогенами . Примеры включают бромид этидия и акридин . [ 11 ] [ нужна ссылка ]
Ремонт несоответствия
[ редактировать ]Несовпадающие пары оснований могут образовываться в результате ошибок репликации ДНК и выступать в качестве промежуточных продуктов во время гомологичной рекомбинации . Процесс репарации ошибочных спариваний обычно должен распознавать и правильно восстанавливать небольшое количество неправильных пар оснований в длинной последовательности нормальных пар оснований ДНК. Для устранения несоответствий, образовавшихся во время репликации ДНК, было разработано несколько различных процессов репарации, позволяющих различать матричную цепь и вновь образованную цепь, так что удаляется только вновь вставленный неправильный нуклеотид (во избежание возникновения мутации). [ 12 ] Белки, участвующие в репарации несоответствий во время репликации ДНК, а также клиническое значение дефектов в этом процессе описаны в статье « Репарация несоответствий ДНК» . Процесс исправления ошибочных пар при рекомбинации описан в статье « Конверсия генов» .
Измерения длины
[ редактировать ]
Для описания длины молекулы D/R NA обычно используются следующие сокращения :
- bp = пара оснований - одна bp соответствует примерно 3,4 Å (340 пм ). [ 13 ] длины вдоль цепи и примерно до 618 или 643 дальтон для ДНК и РНК соответственно.
- kb (= kbp) = килограмм пары оснований = 1000 bp
- Mb (= Mbp) = пара мегаоснований = 1 000 000 bp
- Gb (= Gbp) = гига-пара оснований = 1 000 000 000 п.н.
Для одноцепочечной ДНК/РНК единицы нуклеотидов используются , сокращенно nt (или knt, Mnt, Gnt), поскольку они не спарены. Чтобы различать единицы компьютерной памяти и базы, для пар оснований можно использовать kbp, Mbp, Gbp и т. д.
Сантиморган . также часто используется для обозначения расстояния вдоль хромосомы, но количество пар оснований, которым он соответствует, широко варьируется В геноме человека сентиморган составляет около 1 миллиона пар оснований. [ 14 ] [ 15 ]
Неестественная пара оснований (UBP)
[ редактировать ]Неестественная пара оснований (UBP) — это спроектированная субъединица (или азотистое основание ) ДНК , созданная в лаборатории и не встречающаяся в природе. Были описаны последовательности ДНК, которые используют вновь созданные нуклеиновые основания для образования третьей пары оснований в дополнение к двум парам оснований, обнаруженным в природе: AT ( аденин - тимин ) и GC ( гуанин - цитозин ). Несколько исследовательских групп искали третью пару оснований ДНК, в том числе команды под руководством Стивена А. Беннера , Филиппа Марлиера , Флойда Э. Ромесберга и Ичиро Хирао . [ 16 ] Сообщалось о некоторых новых парах оснований, основанных на альтернативных водородных связях, гидрофобных взаимодействиях и координации металлов. [ 17 ] [ 18 ] [ 19 ] [ 20 ]
В 1989 году Стивен Беннер (тогда работавший в Швейцарском федеральном технологическом институте в Цюрихе) и его команда начали использовать модифицированные формы цитозина и гуанина в молекулах ДНК in vitro . [ 21 ] Нуклеотиды, кодирующие РНК и белки, были успешно реплицированы in vitro . С тех пор команда Беннера пытается создать ячейки, которые смогут создавать зарубежные базы с нуля, устраняя необходимость в сырье. [ 22 ]
В 2002 году группа Ичиро Хирао в Японии разработала неприродную пару оснований между 2-амино-8-(2-тиенил)пурином (s) и пиридин-2-оном (y), которая участвует в транскрипции и трансляции, для сайт-специфических включение нестандартных аминокислот в белки. [ 23 ] В 2006 году они создали 7-(2-тиенил)имидазо[4,5-b]пиридин (Ds) и пиррол-2-карбальдегид (Pa) в качестве третьей пары оснований для репликации и транскрипции. [ 24 ] Впоследствии Ds и 4-[3-(6-аминогексанамидо)-1-пропинил]-2-нитропиррол (Px) были обнаружены как пара с высокой точностью при ПЦР-амплификации. [ 25 ] [ 26 ] В 2013 году они применили пару Ds-Px для генерации аптамеров ДНК путем селекции in vitro (SELEX) и продемонстрировали, что расширение генетического алфавита значительно увеличивает сродство аптамеров ДНК к белкам-мишеням. [ 27 ]
В 2012 году группа американских ученых во главе с Флойдом Ромесбергом, химиком-биологом из Исследовательского института Скриппса в Сан-Диего, Калифорния, опубликовала информацию о том, что его команда разработала неестественную пару оснований (UBP). [ 19 ] Два новых искусственных нуклеотида или пары неестественных оснований (UBP) были названы d5SICS и dNaM . С технической точки зрения, эти искусственные нуклеотиды, несущие гидрофобные нуклеиновые основания , имеют два слитых ароматических кольца , которые образуют комплекс (d5SICS – dNaM) или пару оснований в ДНК. [ 22 ] [ 28 ] Его команда разработала множество шаблонов in vitro или «пробирок», содержащих неестественную пару оснований, и они подтвердили, что она эффективно и с высокой точностью воспроизводится практически во всех контекстах последовательностей с использованием современных стандартных методов in vitro , а именно ПЦР-амплификации ДНК и ПЦР. -приложения. [ 19 ] Их результаты показывают, что для ПЦР и приложений на основе ПЦР неестественная пара оснований d5SICS-dNaM функционально эквивалентна природной паре оснований, а в сочетании с двумя другими природными парами оснований, используемыми всеми организмами, A-T и G-C. , они предоставляют полностью функциональный и расширенный шестибуквенный «генетический алфавит». [ 28 ]
В 2014 году та же команда из Научно-исследовательского института Скриппса сообщила, что они синтезировали участок кольцевой ДНК, известный как плазмида, содержащая природные пары оснований TA и CG, а наиболее эффективная лаборатория UBP Ромесберга разработала и вставила его в клетки обычной бактерии. E. coli , которая успешно воспроизводила неестественные пары оснований в нескольких поколениях. [ 16 ] Трансфекция клеток E. не препятствовала росту coli и не выявила признаков потери неестественных пар оснований в результате естественных механизмов репарации ДНК . Это первый известный пример передачи живым организмом расширенного генетического кода последующим поколениям. [ 28 ] [ 29 ] Ромесберг сказал, что он и его коллеги создали 300 вариантов, чтобы усовершенствовать конструкцию нуклеотидов, которые будут достаточно стабильными и будут воспроизводиться так же легко, как и естественные, при делении клеток. Частично это было достигнуто за счет добавления вспомогательного гена водорослей , который экспрессирует транспортер нуклеотид-трифосфата , который эффективно импортирует трифосфаты как d5SICSTP, так и dNaMTP в E. coli . бактерии [ 28 ] Затем естественные пути репликации бактерий используют их для точной репликации плазмиды, содержащей d5SICS–dNaM. Другие исследователи были удивлены тем, что бактерии воспроизвели эти субъединицы ДНК, созданные человеком. [ 30 ]
Успешное включение третьей пары оснований является значительным прорывом на пути к значительному расширению числа аминокислот , которые могут кодироваться ДНК, с существующих 20 аминокислот до теоретически возможных 172, тем самым расширяя возможности живых организмов производить новые белки . [ 16 ] Искусственные нити ДНК пока ничего не кодируют, но ученые предполагают, что их можно спроектировать для производства новых белков, которые могут найти промышленное или фармацевтическое применение. [ 31 ] Эксперты заявили, что синтетическая ДНК, включающая неестественную пару оснований, повышает вероятность существования форм жизни, основанных на другом коде ДНК. [ 30 ] [ 31 ]
Неканоническое спаривание оснований
[ редактировать ]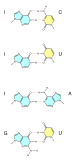
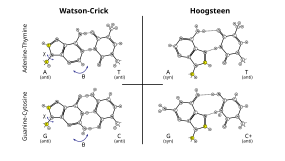
Помимо канонического спаривания, некоторые условия могут также благоприятствовать спариванию оснований с альтернативной ориентацией оснований, а также количеством и геометрией водородных связей. Эти пары сопровождаются изменениями формы локального позвоночника. [ нужна ссылка ]
Наиболее распространенным из них является колебание спаривания оснований , которое происходит между тРНК и мРНК в третьем положении оснований многих кодонов во время транскрипции. [ 33 ] и во время зарядки тРНК некоторыми тРНК-синтетазами . [ 34 ] Они также наблюдались во вторичных структурах некоторых последовательностей РНК. [ 35 ]
Кроме того, спаривание оснований Хугстина (обычно обозначаемое как A•U/T и G•C) может существовать в некоторых последовательностях ДНК (например, динуклеотидах CA и TA) в динамическом равновесии со стандартным спариванием Уотсона-Крика. [ 32 ] Они также наблюдались в некоторых комплексах белок-ДНК. [ 36 ]
Помимо этих альтернативных пар оснований, во вторичной и третичной структуре РНК наблюдается широкий спектр водородных связей основания-основания. [ 37 ] Эти связи часто необходимы для точной и сложной формы РНК, а также для ее связывания с партнерами по взаимодействию. [ 37 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Спенсер М. (10 января 1959 г.). «Стереохимия дезоксирибонуклеиновой кислоты. II. Пары оснований с водородной связью» . Акта Кристаллографика . 12 (1): 66–71. дои : 10.1107/S0365110X59000160 . ISSN 0365-110X .
- ^ Журкин В.Б., Толсторуков М.Ю., Сюй Ф, Коласанти А.В., Олсон В.К. (2005). «Последовательность-зависимая изменчивость B-ДНК». Конформация ДНК и транскрипция . стр. 18–34. дои : 10.1007/0-387-29148-2_2 . ISBN 978-0-387-25579-8 .
- ^ Моран Л.А. (24 марта 2011 г.). «Общий размер человеческого генома, скорее всего, составит ~3200 МБ» . Sandwalk.blogspot.com . Проверено 16 июля 2012 г.
- ^ «Конечная длина генома человека составляет 2,86 Гб» . Стратегическая геномика.com. 12 июня 2006 г. Проверено 16 июля 2012 г.
- ^ Международный консорциум по секвенированию генома человека (октябрь 2004 г.). «Завершение эухроматической последовательности генома человека» . Природа . 431 (7011): 931–945. Бибкод : 2004Natur.431..931H . дои : 10.1038/nature03001 . ПМИД 15496913 .
- ^ Кокберн А.Ф., Ньюкирк М.Дж., Фиртел Р.А. (декабрь 1976 г.). «Организация генов рибосомальной РНК Dictyostelium discoideum: картирование нетранскрибируемых спейсерных областей». Клетка . 9 (4, ч. 1): 605–613. дои : 10.1016/0092-8674(76)90043-X . ПМИД 1034500 . S2CID 31624366 .
- ^ Нувер Р. (18 июля 2015 г.). «Подсчитаем всю ДНК на Земле» . Нью-Йорк Таймс . Нью-Йорк. ISSN 0362-4331 . Архивировано из оригинала 1 января 2022 г. Проверено 18 июля 2015 г.
- ^ «Биосфера: разнообразие жизни» . Аспенский институт глобальных изменений . Базальт, Колорадо. Архивировано из оригинала 10 ноября 2014 г. Проверено 19 июля 2015 г.
- ^ Яковчук П., Протозанова Е., Франк-Каменецкий М.Д. (30 января 2006 г.). «Вклад укладки оснований и спаривания оснований в термическую стабильность двойной спирали ДНК» . Исследования нуклеиновых кислот . 34 (2): 564–574. дои : 10.1093/nar/gkj454 . ПМЦ 1360284 . ПМИД 16449200 .
- ^ Траутнер Т.А., Шварц М.Н., Корнберг А. (март 1962 г.). «Ферментативный синтез дезоксирибонуклеиновой кислоты. X. Влияние бромурациловых замен на репликацию» . Труды Национальной академии наук Соединенных Штатов Америки . 48 (3): 449–455. дои : 10.1073/pnas.48.3.449 . ПМК 220799 . ПМИД 13922323 .
- ^ Кребс Дж. Э., Гольдштейн Э. С., Килпатрик С. Т., Левин Б. (2018). «Гены представляют собой ДНК и кодируют РНК и полипептиды». Гены Левина XII (12-е изд.). Берлингтон, Массачусетс: Jones & Bartlett Learning. п. 12. ISBN 978-1-284-10449-3 .
Каждое мутагенное событие в присутствии акридина приводит к добавлению или удалению одной пары оснований.
- ^ Компакт-диск Патнэма (сентябрь 2021 г.). «Дискриминация цепей при восстановлении несоответствий ДНК» . Восстановление ДНК . 105 : 103161. doi : 10.1016/j.dnarep.2021.103161 . ПМЦ 8785607 . ПМИД 34171627 .
- ^ Альбертс Б., Джонсон А., Льюис Дж., Морган Д., Рафф М., Робертс К., Уолтер П. (декабрь 2014 г.). Молекулярная биология клетки (6-е изд.). Нью-Йорк/Абингдон: Garland Science, Taylor & Francisco Group. п. 177. ИСБН 978-0-8153-4432-2 .
- ^ «NIH ORDR – Глоссарий – C» . Rarediseases.info.nih.gov. Архивировано из оригинала 17 июля 2012 г. Проверено 16 июля 2012 г.
- ^ Скотт М.П., Мацудайра П., Лодиш Х., Дарнелл Дж., Зипурски Л., Кайзер К.А., Берк А., Кригер М. (2004). Молекулярно-клеточная биология (Пятое изд.). Сан-Франциско: WH Freeman. п. 396 . ISBN 978-0-7167-4366-8 .
...у человека 1 сантиморган в среднем представляет собой расстояние примерно 7,5x10 5 пары оснований.
- ^ Перейти обратно: а б с Файкс Би Джей (8 мая 2014 г.). «Жизнь создана с использованием расширенного генетического кода» . Сан-Диего Юнион Трибьюн . Архивировано из оригинала 9 мая 2014 года . Проверено 8 мая 2014 г.
- ^ Ян З., Чен Ф., Альварадо Дж.Б., Беннер С.А. (сентябрь 2011 г.). «Амплификация, мутация и секвенирование шестибуквенной синтетической генетической системы» . Журнал Американского химического общества . 133 (38): 15105–15112. дои : 10.1021/ja204910n . ПМЦ 3427765 . ПМИД 21842904 .
- ^ Ямасигэ Р., Кимото М., Такезава Ю., Сато А., Мицуи Т., Ёкояма С., Хирао И. (март 2012 г.). «Высокоспецифичные системы неприродных пар оснований в качестве третьей пары оснований для ПЦР-амплификации» . Исследования нуклеиновых кислот . 40 (6): 2793–2806. дои : 10.1093/nar/gkr1068 . ПМЦ 3315302 . ПМИД 22121213 .
- ^ Перейти обратно: а б с Малышев Д.А., Дхами К., Куах Х.Т., Лавернь Т., Ордуханян П., Торкамани А., Ромесберг Ф.Е. (июль 2012 г.). «Эффективная и независимая от последовательности репликация ДНК, содержащей третью пару оснований, создает функциональный шестибуквенный генетический алфавит» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (30): 12005–12010. Бибкод : 2012PNAS..10912005M . дои : 10.1073/pnas.1205176109 . ПМК 3409741 . ПМИД 22773812 .
- ^ Такезава Ю., Мюллер Дж., Сионоя М. (05.05.2017). «Искусственное спаривание оснований ДНК, опосредованное различными ионами металлов» . Химические письма . 46 (5): 622–633. дои : 10.1246/кл.160985 . ISSN 0366-7022 .
- ^ Свитцер С., Морони С.Э., Беннер С.А. (1989). «Ферментативное включение новой пары оснований в ДНК и РНК». Дж. Ам. хим. Соц . 111 (21): 8322–8323. дои : 10.1021/ja00203a067 .
- ^ Перейти обратно: а б Каллауэй Э (7 мая 2014 г.). «Ученые создали первый живой организм с «искусственной» ДНК» . Новости природы . Хаффингтон Пост . Проверено 8 мая 2014 г.
- ^ Хирао И., Оцуки Т., Фудзивара Т., Мицуи Т., Ёкогава Т., Окуни Т. и др. (февраль 2002 г.). «Неестественная пара оснований для включения аналогов аминокислот в белки». Природная биотехнология . 20 (2): 177–182. дои : 10.1038/nbt0202-177 . ПМИД 11821864 . S2CID 22055476 .
- ^ Хирао И., Кимото М., Мицуи Т., Фудзивара Т., Каваи Р., Сато А. и др. (сентябрь 2006 г.). «Неестественная гидрофобная система пар оснований: сайт-специфическое включение аналогов нуклеотидов в ДНК и РНК». Природные методы . 3 (9): 729–735. дои : 10.1038/nmeth915 . ПМИД 16929319 . S2CID 6494156 .
- ^ Кимото М., Каваи Р., Мицуи Т., Ёкояма С., Хирао И. (февраль 2009 г.). «Неестественная система пар оснований для эффективной ПЦР-амплификации и функционализации молекул ДНК» . Исследования нуклеиновых кислот . 37 (2): е14. дои : 10.1093/нар/gkn956 . ПМЦ 2632903 . ПМИД 19073696 .
- ^ Ямасигэ Р., Кимото М., Такезава Ю., Сато А., Мицуи Т., Ёкояма С., Хирао И. (март 2012 г.). «Высокоспецифичные системы неприродных пар оснований в качестве третьей пары оснований для ПЦР-амплификации» . Исследования нуклеиновых кислот . 40 (6): 2793–2806. дои : 10.1093/nar/gkr1068 . ПМЦ 3315302 . ПМИД 22121213 .
- ^ Кимото М., Ямасигэ Р., Мацунага К., Ёкояма С., Хирао И. (май 2013 г.). «Поколение аптамеров ДНК с высоким сродством с использованием расширенного генетического алфавита». Природная биотехнология . 31 (5): 453–457. дои : 10.1038/nbt.2556 . ПМИД 23563318 . S2CID 23329867 .
- ^ Перейти обратно: а б с д Малышев Д.А., Дхами К., Лавернь Т., Чен Т., Дай Н., Фостер Дж.М. и др. (май 2014 г.). «Полусинтетический организм с расширенным генетическим алфавитом» . Природа . 509 (7500): 385–388. Бибкод : 2014Natur.509..385M . дои : 10.1038/nature13314 . ПМК 4058825 . ПМИД 24805238 .
- ^ Образец I (7 мая 2014 г.). «Первые формы жизни, передавшие искусственную ДНК, созданную американскими учеными» . Хранитель . Проверено 8 мая 2014 г.
- ^ Перейти обратно: а б «Ученые создали первый живой организм, содержащий искусственную ДНК» . Уолл Стрит Джорнал . Фокс Ньюс. 8 мая 2014 года . Проверено 8 мая 2014 г.
- ^ Перейти обратно: а б Поллак А (7 мая 2014 г.). «Ученые добавляют буквы в алфавит ДНК, вселяя надежду и страх» . Нью-Йорк Таймс . Проверено 8 мая 2014 г.
- ^ Перейти обратно: а б Николова Е.Н., Ким Э., Уайз А.А., О'Брайен П.Дж., Андрисиоаи И., Аль-Хашими Х.М. (февраль 2011 г.). «Переходные пары оснований Хугстина в канонической дуплексной ДНК» . Природа . 470 (7335): 498–502. Бибкод : 2011Natur.470..498N . дои : 10.1038/nature09775 . ПМК 3074620 . ПМИД 21270796 .
- ^ Мерфи Ф.В., Рамакришнан В. (декабрь 2004 г.). «Структура пурин-пуриновой колебательной пары оснований в декодирующем центре рибосомы». Структурная и молекулярная биология природы . 11 (12): 1251–1252. дои : 10.1038/nsmb866 . ПМИД 15558050 . S2CID 27022506 .
- ^ Варгас-Родригес О., Мюзье-Форсайт К. (июнь 2014 г.). «Структурная биология: колебание направляет РНК на цель». Природа . 510 (7506): 480–481. дои : 10.1038/nature13502 . ПМИД 24919145 . S2CID 205239383 .
- ^ Гарг А., Хайнеманн Ю (февраль 2018 г.). «Новая форма двойной спирали РНК, основанная на G·U и C·A. + спаривание оснований колебания» . РНК . 24 (2): 209–218. : 10.1261 /rna.064048.117 . PMC 5769748. . PMID 29122970 doi
- ^ Айшима Дж., Гитти Р.К., Ной Дж.Э., Ган Х.Х., Шлик Т., Вольбергер С. (декабрь 2002 г.). «Пара оснований Хугстина, встроенная в неискаженную B-ДНК» . Исследования нуклеиновых кислот . 30 (23): 5244–5252. дои : 10.1093/nar/gkf661 . ПМЦ 137974 . ПМИД 12466549 .
- ^ Перейти обратно: а б Леонтис Н.Б., Вестхоф E (июнь 2003 г.). «Анализ мотивов РНК». Современное мнение в области структурной биологии . 13 (3): 300–308. дои : 10.1016/S0959-440X(03)00076-9 . ПМИД 12831880 .
Дальнейшее чтение
[ редактировать ]- Уотсон Дж.Д., Бейкер Т.А., Белл С.П., Ганн А., Левин М., Лосик Р. (2004). Молекулярная биология гена (5-е изд.). Пирсон Бенджамин Каммингс: CSHL Press. (См. особенно гл. 6 и 9)
- Сигел А, Сигел Х, Сигел Р.К., ред. (2012). Взаимодействие между ионами металлов и нуклеиновыми кислотами . Ионы металлов в науках о жизни. Том. 10. Спрингер. дои : 10.1007/978-94-007-2172-2 . ISBN 978-9-4007-2171-5 . S2CID 92951134 .
- Умный Г.Х., Шионоя М. (2012). «Альтернативное спаривание оснований ДНК посредством координации металлов». Взаимодействие между ионами металлов и нуклеиновыми кислотами . Ионы металлов в науках о жизни. Том. 10. С. 269–294. дои : 10.1007/978-94-007-2172-2_10 . ISBN 978-94-007-2171-5 . ПМИД 22210343 .
- Меггер Д.А., Меггер Н., Мюллер Дж. (2012). «Металло-опосредованные пары оснований в нуклеиновых кислотах с нуклеозидами пуринового и пиримидинового происхождения». Взаимодействие между ионами металлов и нуклеиновыми кислотами . Ионы металлов в науках о жизни. Том. 10. С. 295–317. дои : 10.1007/978-94-007-2172-2_11 . ISBN 978-94-007-2171-5 . ПМИД 22210344 .
Внешние ссылки
[ редактировать ]- DAN — веб-серверная версия инструмента EMBOSS для расчета температур плавления.
Виды нуклеиновых кислот |
|---|