амфибия
| Земноводные Временной диапазон:
| |
|---|---|
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Батрахоморфа |
| Сорт: | Амфибия Серый 1825 г. [ 2 ] |
| Подклассы | |
|
(неполный список)
| |




Земноводные — экзотермические , анамниотические , четырёхконечные позвоночные животные , составляющие класс Амфибии . В самом широком смысле это парафилетическая группа, охватывающая всех четвероногих, за исключением амниот (четвероногих с амниотической оболочкой , таких как современные рептилии , птицы и млекопитающие ). Все ныне живущие амфибии принадлежат к монофилетическому подклассу Lissamphibia с тремя современными отрядами : Anura ( лягушки и жабы ), Urodela ( саламандры ) и Gymnophiona ( червячие ). В ходе эволюции амфибии стали в основном полуводными , и они приспособились к обитанию в самых разных средах обитания , причем большинство видов живут в пресноводных , водно-болотных угодьях или наземных экосистемах (таких как прибрежные леса , ископаемые и даже древесные среды обитания). Их жизненный цикл обычно начинается с появления водных личинок с жабрами , известных как головастики , но некоторые виды выработали поведенческие адаптации, позволяющие обойти это.
Молодые амфибии обычно претерпевают метаморфоз из водной личиночной формы с жабрами во взрослую форму, дышащую воздухом, с легкими . Земноводные используют свою кожу в качестве вторичного дыхательного канала, а у некоторых мелких наземных саламандр и лягушек нет легких, и они полностью полагаются на свою кожу. Внешне они похожи на рептилий, таких как ящерицы , но, в отличие от рептилий и других амниот , для размножения им требуется доступ к водоемам. Земноводные с их сложными репродуктивными потребностями и проницаемой кожей часто являются экологическими индикаторами условий среды обитания; в последние десятилетия произошло резкое сокращение популяций амфибий многих видов по всему миру.
Самые ранние земноводные произошли в девонский период от тетраподоморфных саркоптеригов ( лопастных рыб с сочлененными конечности , похожими на плавниками ), у которых развились примитивные легкие, которые помогли адаптироваться к суше. Они диверсифицировались и стали экологически доминирующими в каменноугольный и пермский периоды, но позже были вытеснены в наземной среде ранними рептилиями и базальными синапсидами (предшественниками млекопитающих). Происхождение современных лисамфибий, которые впервые появились в раннем триасе, около 250 миллионов лет назад, долгое время вызывало споры. Самая популярная гипотеза состоит в том, что они, вероятно, произошли от темноспондилов , самой разнообразной группы доисторических амфибий, в пермский период. [ 4 ] Другая гипотеза состоит в том, что они произошли от лепоспондисов. [ 5 ] Четвертая группа лисамфибий, Albanerpetontidae , вымерла около 2 миллионов лет назад.
Число известных видов земноводных составляет около 8000, из которых почти 90% — лягушки. Самая маленькая амфибия (и позвоночное животное) в мире — лягушка из Новой Гвинеи ( Paedophryne amauensis ) длиной всего 7,7 мм (0,30 дюйма). ) длиной 1,8 м (5 футов 11 дюймов) Самая крупная из ныне живущих амфибий — южнокитайская гигантская саламандра ( Andrias sligoi , но она затмевается доисторическими темноспондилами, такими как мастодонзавр , длина которых могла достигать 6 м (20 футов). [ 6 ] Изучение амфибий называется батрахологией , а изучение пресмыкающихся и амфибий — герпетологией .
Классификация

Слово амфибия происходит от древнегреческого термина ἀμφίβιος ( амфибиос ), что означает «оба вида жизни», ἀμφί означает «оба вида» и βίος означает «жизнь». Первоначально этот термин использовался как общее прилагательное для животных, которые могли жить на суше или в воде, включая тюленей и выдр. [ 7 ] Традиционно к классу амфибий относятся все четвероногие позвоночные, не являющиеся амниотами. Земноводные в самом широком смысле ( sensu lato ) разделились на три подкласса , два из которых вымерли: [ 8 ]
- Подкласс Lepospondyli † (потенциально полифилетическая позднепалеозойская группа мелких форм, вероятно, более тесно связанная с амниотами, чем Lissamphibia)
- Подкласс Temnospondyli † (разнообразные позднепалеозойские и раннемезозойские классы, некоторые из которых были крупными хищниками)
- Подкласс Lissamphibia (все современные земноводные, включая лягушек, жаб, саламандр, тритонов и червяг)
- Salientia (лягушки, жабы и родственники): от раннего триаса до настоящего времени - 7360 современных видов в 53 семействах. [ 9 ] Современные выступающие ( коронная группа ) описываются именем Анура.
- Caudata ( саламандры , тритоны и родственники): от позднего триаса до настоящего времени — 764 современных вида в 9 семействах. [ 9 ] Современные (коронная группа) каудатаны описаны под названием Urodela.
- Gymnophiona ( червяги и родственники): от позднего триаса до настоящего времени - 215 современных видов в 10 семействах. [ 9 ] Название Apoda также иногда используется для обозначения червяг.
- Allocaudata† ( Albanerpetontidae ) средняя юра – ранний плейстоцен.
Эти три подкласса не включают всех вымерших земноводных. Другие вымершие группы амфибий включают Embolomeri (крупные водные хищники позднего палеозоя), Seymouriamorpha (от полуводных до наземных пермских форм, связанных с амниотами) и другие. Такие названия, как Tetrapoda и Stegocephalia, охватывают всех четвероногих класса амфибий, тогда как Reptiliomorpha или Anthracosauria по-разному используются для описания вымерших амфибий, более тесно связанных с амниотами, чем с лиссамфибиями.

Фактическое количество видов в каждой группе зависит от используемой таксономической классификации. Двумя наиболее распространенными системами являются классификация, принятая на веб-сайте AmphibiaWeb Калифорнийского университета в Беркли , и классификация герпетолога Даррела Фроста и Американского музея естественной истории , доступная в виде справочной онлайн-базы данных «Виды земноводных мира». [ 10 ] Численность указанных выше видов соответствует Фросту, а общее количество известных (живых) видов земноводных по состоянию на 31 марта 2019 г. составляет ровно 8000. [ 11 ] из них почти 90% — лягушки. [ 12 ]
При филогенетической классификации таксон Labyrinthodontia был исключен, поскольку это полипарафилетическая группа без уникальных определяющих особенностей, кроме общих примитивных характеристик . Классификация варьируется в зависимости от предпочтительной филогении автора и от того, использует ли он классификацию на основе стебля или узла . Традиционно к земноводным как классу относят всех четвероногих с личиночной стадией, а группу, включающую общих предков всех ныне живущих земноводных (лягушек, саламандр и червяг) и всех их потомков, называют Lissamphibia. Филогения палеозойских амфибий неясна, и Lissamphibia, возможно, могут относиться к вымершим группам, таким как Temnospondyli (традиционно относимых к подклассу Labyrinthodontia) или Lepospondyli, а в некоторых анализах даже к амниотам. Это означает, что сторонники филогенетической номенклатуры удалили большое количество базальных групп четвероногих амфибий девонского и каменноугольного периода, которые ранее относились к амфибиям в Линнеевскую таксономию и включил их в другие разделы кладистической таксономии . [ 2 ] Если в состав амфибий включить общего предка земноводных и амниот, то он становится парафилетической группой. [ 13 ]
Все современные земноводные включены в подкласс Lissamphibia, который обычно считают кладой — группой видов, произошедших от общего предка. Три современных отряда — это Anura (лягушки), Caudata (или Urodela, саламандры) и Gymnophiona (или Apoda, червячие). [ 14 ] Было высказано предположение, что саламандры произошли отдельно от темноспондилоподобного предка и даже что червяги являются сестринской группой продвинутых рептилиоморфных амфибий и, следовательно, амниот. [ 15 ] Хотя известны окаменелости нескольких более старых протолягушек с примитивными характеристиками, самой старой «настоящей лягушкой» с прыжковыми адаптациями является Prosalirus bitis из раннеюрской формации Кайента в Аризоне. Анатомически он очень похож на современных лягушек. [ 16 ] Самыми древними из известных червяг являются Funcusvermis gilmorei (из позднего триаса) и Eocaecilia micropodia (из ранней юры), оба из Аризоны. [ 17 ] Самая ранняя саламандра — Beiyanerpeton jianpingensis из поздней юры северо-восточного Китая. [ 18 ]
Авторы расходятся во мнениях относительно того, является ли Salientia надотрядом, включающим отряд Anura, или Anura является подотрядом Salientia. Лиссамфибии традиционно делятся на три отряда , но вымершее семейство саламандровых, Albanerpetontidae, теперь считается частью Lissamphibia наряду с надотрядом Salientia. Кроме того, Salientia включает в себя все три недавних отряда, а также триасовую протолягушку Triadobatrachus . [ 19 ]
Эволюционная история


Первые крупные группы земноводных развились в девонский период, около 370 миллионов лет назад, из лопастноперых рыб , которые были похожи на современных целакантов и двоякодышащих рыб . [ 20 ] У этих древних рыб с лопастными плавниками развились многочленистые плавники, похожие на ноги, с пальцами, которые позволяли им ползать по морскому дну. У некоторых рыб развились примитивные легкие, которые помогают им дышать воздухом, когда в стоячих водоемах девонских болот было мало кислорода. Они также могли использовать свои сильные плавники, чтобы выбраться из воды на сушу, если того требовали обстоятельства. В конце концов, их костистые плавники превратились в конечности, и они стали предками всех четвероногих , включая современных амфибий, рептилий, птиц и млекопитающих . Несмотря на способность ползать по суше, многие из этих доисторических рыб -тетраподоморфов по-прежнему проводили большую часть времени в воде. У них начали развиваться легкие, но они все еще дышали преимущественно жабрами. [ 21 ]
множество примеров видов, демонстрирующих переходные черты Было обнаружено . Ихтиостега была одной из первых примитивных амфибий с ноздрями и более эффективными легкими. У него было четыре крепких конечности, шея, хвост с плавниками и череп, очень похожий на череп лопастной рыбы Eusthenopteron . [ 20 ] Земноводные развили приспособления, которые позволили им оставаться вне воды в течение более длительного времени. Их легкие улучшились, а скелеты стали тяжелее и сильнее, способные лучше выдерживать вес тела на суше. У них появились «руки» и «ноги» с пятью и более пальцами; [ 22 ] кожа стала более способна удерживать жидкости организма и сопротивляться высыханию. [ 21 ] рыбы Кость гиомандибулы в подъязычной области за жабрами уменьшилась в размерах и превратилась в стремечко уха амфибии - приспособление, необходимое для слуха на суше. [ 23 ] Родством между земноводными и костистыми рыбами является многоскладчатое строение зубов и парные надзатылочные кости в задней части головы; ни одна из этих особенностей не встречается где-либо еще в животном мире. [ 24 ]

В конце девонского периода (360 миллионов лет назад) моря, реки и озера кишели жизнью, тогда как суша была царством ранних растений и лишена позвоночных животных. [ 24 ] хотя некоторые, такие как Ихтиостега , возможно, иногда вылезали из воды. Предполагается, что они, возможно, передвигались с помощью передних конечностей, волоча задние конечности аналогично тому, как это делает морской слон . [ 22 ] В раннем каменноугольном периоде (360–323 миллиона лет назад) климат был относительно влажным и теплым. Обширные болота заросли мхами , папоротниками , хвощами и каламитами . Дышащие воздухом членистоногие эволюционировали и вторглись на сушу, где давали пищу хищным земноводным, которые начали приспосабливаться к наземной среде. На суше не было других четвероногих, и земноводные находились на вершине пищевой цепи, а некоторые из них занимали экологические позиции, которые в настоящее время занимают крокодилы. Хотя они были оснащены конечностями и способностью дышать воздухом, большинство из них все же имели длинное суженное тело и сильный хвост. [ 24 ] Другие были верхними наземными хищниками, иногда достигавшими нескольких метров в длину и охотившимися на крупных насекомых того периода и на множество видов рыб в воде. Им все равно нужно было возвращаться в воду, чтобы отложить яйца без скорлупы, и даже у большинства современных амфибий есть полностью водная личиночная стадия с жабрами, как у их предков-рыб. Именно развитие амниотической яйцеклетки, которая предотвращает высыхание развивающегося эмбриона, позволило рептилиям размножаться на суше и привело к их доминированию в последующий период. [ 20 ]
После разрушения тропических лесов в каменноугольном периоде доминирование земноводных уступило место рептилиям. [ 25 ] а земноводные подверглись дальнейшему опустошению в результате пермско-триасового вымирания . [ 26 ] В течение триасового периода (252–201 миллион лет назад) рептилии продолжали вытеснять земноводных, что приводило к уменьшению как размера земноводных, так и их значения в биосфере . Согласно летописи окаменелостей, Lissamphibia , которая включает в себя всех современных земноводных и является единственной сохранившейся линией, возможно, ответвилась от вымерших групп Temnospondyli и Lepospondyli в какой-то период между поздним карбоном и ранним триасом. Относительная нехватка ископаемых свидетельств не позволяет точно датировать. [ 21 ] но самое последнее молекулярное исследование, основанное на мультилокусном типировании последовательностей , предполагает позднекаменноугольное/ раннепермское происхождение современных амфибий. [ 27 ]

Происхождение и эволюционные взаимоотношения между тремя основными группами амфибий являются предметом дискуссий. Молекулярная филогения 2005 года, основанная на анализе рДНК , предполагает, что саламандры и червячи более тесно связаны друг с другом, чем с лягушками. Похоже также, что расхождение трех групп произошло в палеозое или раннем мезозое (около 250 миллионов лет назад), до распада суперконтинента Пангея и вскоре после их расхождения с лопастноперыми рыбами. Кратковременность этого периода и быстрота распространения радиации могут помочь объяснить относительную нехватку окаменелостей примитивных амфибий. [ 28 ] имеются большие пробелы В летописи окаменелостей : открытие диссорофоида темноспондилового Gerobatrachus из ранней перми в Техасе в 2008 году обеспечило недостающее звено со многими характеристиками современных лягушек. [ 15 ] Молекулярный анализ показывает, что расхождение лягушки и саламандры произошло значительно раньше, чем указывают палеонтологические данные. [ 15 ] Одно исследование показало, что последний общий предок всех современных земноводных жил около 315 миллионов лет назад и что стереоспондилы- темноспондилы являются ближайшими родственниками червяг. [ 29 ] Однако большинство исследований подтверждают единое монофилетическое происхождение всех современных амфибий в составе диссорофоидных темноспондилов. [ 4 ]
Эволюционировав от рыб, амфибий пришлось совершить определенные приспособления для жизни на суше, включая необходимость разработки новых способов передвижения. В воде их двигали вперед боковые толчки хвостов, но на суше требовались совсем другие механизмы. Их позвоночник, конечности, пояса конечностей и мускулатура должны были быть достаточно сильными, чтобы поднимать их над землей для передвижения и питания. Взрослые наземные животные отказались от своих систем боковой линии и адаптировали свои сенсорные системы для получения стимулов из воздуха. Им нужно было разработать новые методы регулирования тепла своего тела, чтобы справиться с колебаниями температуры окружающей среды. Они выработали поведение, подходящее для размножения в наземной среде. Их кожа подвергалась воздействию вредных ультрафиолетовых лучей, которые ранее поглощались водой. Кожа изменилась, стала более защитной и предотвратила чрезмерную потерю воды. [ 30 ]
Характеристики
Надкласс четвероногих делится на четыре класса позвоночных животных с четырьмя конечностями. [ 31 ] Рептилии, птицы и млекопитающие относятся к амниотам, яйца которых либо откладывает, либо переносит самка и окружены несколькими оболочками, некоторые из которых непроницаемы. [ 32 ] Не имея этих мембран, земноводным для размножения требуются водоемы, хотя некоторые виды разработали различные стратегии защиты или обхода уязвимой водной личиночной стадии. [ 30 ] В море они не встречаются, за исключением одной-двух лягушек, обитающих в солоноватой воде мангровых болот ; [ 33 ] Между тем саламандра Андерсона встречается в озерах с солоноватой или соленой водой. [ 34 ] На суше земноводные обитают во влажной среде обитания из-за необходимости сохранять кожу влажной. [ 30 ]
Современные земноводные имеют упрощенную анатомию по сравнению со своими предками из-за педоморфоза , вызванного двумя эволюционными тенденциями: миниатюризацией и необычно большим геномом, что приводит к более медленной скорости роста и развития по сравнению с другими позвоночными. [ 35 ] [ 36 ] Другая причина их размера связана с их быстрым метаморфозом, который, по-видимому, развился только у предков лисамфибий; во всех других известных линиях развитие было гораздо более постепенным. Поскольку ремоделирование пищевого аппарата означает, что они не едят во время метаморфоза, метаморфоз должен идти тем быстрее, чем меньше особь, поэтому это происходит на ранней стадии, когда личинки еще малы. (Самые крупные виды саламандр не подвергаются метаморфозам.) [ 37 ] Земноводные, откладывающие яйца на суше, часто проходят весь метаморфоз внутри яйца. Анамниотическое наземное яйцо имеет диаметр менее 1 см из-за проблем с диффузией, и этот размер ограничивает объем роста после вылупления. [ 38 ]
Самая маленькая амфибия (и позвоночное животное) в мире — это лягушка -микрогилида из Новой Гвинеи ( Paedophryne amauensis ), впервые обнаруженная в 2012 году. Она имеет среднюю длину 7,7 мм (0,30 дюйма) и является частью рода, включающего четыре мировых вида амфибий. десять самых маленьких видов лягушек. [ 39 ] длиной 1,8 м (5 футов 11 дюймов). Самая крупная из ныне живущих амфибий — китайская гигантская саламандра ( Andrias davidianus ) [ 40 ] но это намного меньше, чем самая крупная амфибия, когда-либо существовавшая — вымерший 9-метровый (30 футов) Prionosuchus , крокодилоподобный темноспондил, датируемый 270 миллионами лет назад из средней перми Бразилии. [ 41 ] Самая крупная лягушка — африканская лягушка-голиаф ( Conraua goliath ), которая может достигать 32 см (13 дюймов) и весить 3 кг (6,6 фунта). [ 40 ]
Земноводные — это экзотермические (хладнокровные) позвоночные животные, которые не поддерживают температуру своего тела за счет внутренних физиологических процессов. у них Скорость метаболизма низкая, и в результате их потребности в пище и энергии ограничены. Во взрослом состоянии у них есть слезные протоки и подвижные веки, а у большинства видов есть уши, способные улавливать вибрации воздуха или земли. У них мускулистые языки, которые у многих видов могут высовываться. Современные земноводные имеют полностью окостеневшие позвонки с суставными отростками . Их ребра обычно короткие и могут срастаться с позвонками. Их черепа в основном широкие и короткие и часто не полностью окостеневшие. Их кожа содержит мало кератина и лишена чешуи, за исключением нескольких рыбьих чешуек у некоторых червяг. В коже имеется множество слизистых желез , а у некоторых видов — ядовитых желез (разновидность зернистых желез). Сердце амфибий имеет три камеры: два предсердия и один желудочек . У них есть мочевой пузырь , и азотистые продукты жизнедеятельности выводятся преимущественно в виде мочевина . Большинство земноводных откладывают яйца в воду, и у них появляются водные личинки, которые претерпевают метаморфозы и становятся наземными взрослыми особями. Земноводные дышат посредством насосного действия, при котором воздух сначала всасывается в щечно-глоточную область через ноздри. Затем они закрываются, и воздух нагнетается в легкие за счет сокращения горла. [ 42 ] Они дополняют это газообменом через кожу. [ 30 ]
Анура

Отряд Анура (от древнегреческого a(n) — «без» и Oura — «хвост») включает лягушек и жаб. У них обычно длинные задние конечности, которые складываются под ними, более короткие передние конечности, перепончатые пальцы без когтей и хвоста, большие глаза и влажная железистая кожа. [ 14 ] Представителей этого отряда с гладкой кожей обычно называют лягушками, а особей с бородавчатой кожей — жабами. С таксономической точки зрения это различие не является формальным, и из этого правила существует множество исключений. Члены семейства Bufonidae известны как «настоящие жабы». [ 43 ] Размер лягушек варьируется от 30-сантиметровой (12 дюймов) лягушки-голиафа ( Conraua goliath ) из Западной Африки. [ 44 ] до 7,7-миллиметрового (0,30 дюйма) Paedophryne amauensis , впервые описанного в Папуа-Новой Гвинее в 2012 году, который также является самым маленьким из известных позвоночных. [ 45 ] Хотя большинство видов связано с водой и влажной средой обитания, некоторые приспособлены к жизни на деревьях или в пустынях. Они встречаются по всему миру, за исключением полярных районов. [ 46 ]
Анура разделена на три подотряда, которые широко признаны научным сообществом, но взаимоотношения между некоторыми семействами остаются неясными. Будущие молекулярные исследования должны предоставить дальнейшее понимание их эволюционных взаимоотношений. [ 47 ] Подотряд Archaeobatrachia включает четыре семейства примитивных лягушек. Это Ascaphidae , Bombinatoridae , Discoglossidae и Leiopelmatidae , которые имеют мало производных особенностей и, вероятно, являются парафилетическими по отношению к другим линиям лягушек. [ 48 ] Шесть семейств в более эволюционно развитом подотряде Mesobatrachia — это ископаемые Megophryidae , Pelobatidae , Pelodytidae , Scaphiopodidae и Rhinophrynidae и обязательно водные Pipidae . Они обладают определенными характеристиками, которые занимают промежуточное положение между двумя другими подотрядами. [ 48 ] Neobatrachia на сегодняшний день является крупнейшим подотрядом и включает в себя остальные семейства современных лягушек, включая наиболее распространенные виды. Примерно 96% из более чем 5000 существующих видов лягушек являются необатрахиями. [ 49 ]
Хвостатый

( Andrias japonicus ), примитивная саламандра.
Отряд Caudata (от латинского cauda, означающего «хвост») состоит из саламандр — вытянутых, низкорослых животных, по форме больше всего напоминающих ящериц. Это симплезиоморфный признак , и они не более близки к ящерицам, чем к млекопитающим. [ 50 ] У саламандр отсутствуют когти, кожа без чешуи, гладкая или покрытая бугорками , и хвосты, обычно сплющенные из стороны в сторону и часто с плавниками. По размеру они варьируются от китайской гигантской саламандры ( Andrias davidianus ), длина которой, как сообщается, вырастает до 1,8 метра (5 футов 11 дюймов), до [ 51 ] до миниатюрного Thorius pennatulus из Мексики, длина которого редко превышает 20 мм (0,8 дюйма). [ 52 ] Саламандры имеют преимущественно лавразийское распространение и обитают на большей части голарктической области северного полушария. Семейство Plethodontidae встречается также в Центральной Америке и Южной Америке к северу от бассейна Амазонки ; [ 46 ] Южная Америка, очевидно, была захвачена из Центральной Америки примерно в начале миоцена , 23 миллиона лет назад. [ 53 ] Уродела — это название, которое иногда используется для всех существующих видов саламандр. [ 54 ] Члены нескольких семейств саламандр стали педоморфными и либо не смогли завершить метаморфоз, либо сохранили некоторые личиночные характеристики во взрослом возрасте. [ 55 ] Большинство саламандр имеют длину менее 15 см (5,9 дюйма). Они могут быть наземными или водными, и многие из них проводят часть года в каждой среде обитания. На суше они в основном проводят день, спрятавшись под камнями, бревнами или в густой растительности, а вечером и ночью выходят на поиски червей, насекомых и других беспозвоночных. [ 46 ]

( Triturus dobrogicus ), продвинутая саламандра.
Подотряд Cryptobranchoidea включает примитивных саламандр. Был найден ряд ископаемых криптожаберных, но живых видов осталось только три: китайская гигантская саламандра ( Andrias davidianus ), японская гигантская саламандра ( Andrias japonicus ) и повелитель ада ( Cryptobranchus alleganiensis ) из Северной Америки. Эти крупные амфибии сохраняют некоторые личиночные характеристики во взрослом состоянии; жаберные щели имеются, глаза открыты. Уникальной особенностью является их способность питаться путем всасывания, нажимая либо на левую, либо на правую сторону нижней челюсти. [ 56 ] Самцы выкапывают гнезда, уговаривают самок отложить в них яйцеклетки и охраняют их. Помимо дыхания легкими, они дышат через множество складок своей тонкой кожи, капилляры которой расположены близко к поверхности. [ 57 ]
Подотряд Salamandroidea содержит продвинутых саламандр. От криптожаберхид они отличаются сросшимися предсуставными костями нижней челюсти и внутренним оплодотворением. У саламандридов самец откладывает пучок спермы, сперматофор , а самка подхватывает его и вводит в свою клоаку , где сперма хранится до откладки яиц. [ 58 ] Самое крупное семейство в этой группе — Plethodontidae, безлегочные саламандры, в которое входит 60% всех видов саламандр. Семейство включает настоящих саламандр , Salamandridae а название « тритон » дано членам подсемейства Pleurodelinae . [ 14 ]
Третий подотряд, Sirenoidea , содержит четыре вида сирен, которые входят в одно семейство Sirenidae . Представители этого отряда — водные саламандры, похожие на угрей , с сильно редуцированными передними конечностями и отсутствием задних конечностей. Некоторые из их особенностей являются примитивными, тогда как другие являются производными. [ 59 ] Оплодотворение, вероятно, будет внешним, поскольку у сиренид отсутствуют клоакальные железы, используемые самцами саламандридов для производства сперматофоров, а у самок отсутствуют сперматеки для хранения спермы. Несмотря на это, яйца откладываются поодиночке, что не способствует внешнему оплодотворению. [ 58 ]
Гимнофиона

Отряд Gymnophiona (от греческого Gymnos, означающего «обнаженный», и ophis, означающего «змея») или Apoda, включает червяг. Это длинные цилиндрические животные без конечностей змеевидной или червеобразной формы. Длина взрослых особей варьируется от 8 до 75 сантиметров (от 3 до 30 дюймов), за исключением червяг Томсона ( Caecilia thompsoni ), длина которых может достигать 150 см (59 дюймов). Кожа червяг имеет большое количество поперечных складок, а у некоторых видов содержит крошечные встроенные кожные чешуйки. У него рудиментарные глаза, покрытые кожей, которые, вероятно, ограничены распознаванием различий в интенсивности света. Рядом с глазом у него также есть пара коротких щупалец , которые можно вытягивать и которые выполняют тактильные и обонятельные функции. Большинство червяг живут под землей в норах во влажной почве, в гнилой древесине и под растительными остатками, но некоторые из них ведут водный образ жизни. [ 60 ] Большинство видов откладывают яйца под землей, а когда личинки вылупляются, они направляются в соседние водоемы. Другие высиживают яйца, и личинки претерпевают метаморфозы до того, как из яиц вылупятся яйца. Некоторые виды рождают живых детенышей, питая их железистыми выделениями, пока они находятся в яйцеводе. [ 61 ] Цецилии распространены в основном в Гондване и встречаются в тропических регионах Африки, Азии, Центральной и Южной Америки. [ 62 ]
Анатомия и физиология
Кожа

Покровная структура содержит некоторые типичные характеристики , общие для наземных позвоночных, такие как наличие сильно ороговевших наружных слоев, периодически обновляющихся в процессе линьки, контролируемой гипофизом и щитовидной железой. Местные утолщения (часто называемые бородавками) являются обычным явлением, например, у жаб. Наружная часть кожи периодически сбрасывается в основном целиком, в отличие от млекопитающих и птиц, у которых она сбрасывается хлопьями. Земноводные часто поедают шелушащуюся кожу. [ 46 ] Цецилии уникальны среди амфибий тем, что имеют минерализованные кожные чешуйки, встроенные в дерму между бороздками кожи. Сходство их с чешуей костистых рыб во многом поверхностное. Ящерицы и некоторые лягушки имеют несколько схожие остеодермы, образующие костные отложения в дерме, но это пример конвергентной эволюции , когда сходные структуры возникли независимо в различных линиях позвоночных. [ 63 ]
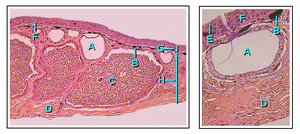
- Слизистая железа
- хроматофор
- Гранулярная ядовитая железа
- Соединительная ткань
- Роговой слой
- Переходная зона
- Эпидермис
- Дерма
Кожа земноводных проницаема для воды. Газообмен может происходить через кожу ( кожное дыхание ), что позволяет взрослым земноводным дышать, не поднимаясь на поверхность воды, и зимовать на дне водоемов. [ 46 ] Чтобы компенсировать тонкую и нежную кожу, у земноводных развились слизистые железы, главным образом на голове, спине и хвосте. Выделяемые ими выделения помогают сохранять кожу влажной. Кроме того, у большинства видов земноводных имеются зернистые железы, выделяющие неприятные или ядовитые вещества. Некоторые токсины амфибий могут быть смертельными для человека, тогда как другие оказывают незначительное воздействие. [ 64 ] Основные ядообразующие железы — паротоиды — вырабатывают нейротоксин буфотоксин и расположены за ушами жаб, вдоль спины лягушек, за глазами саламандр и на верхней поверхности червяг. [ 65 ]
Цвет кожи земноводных определяется тремя слоями пигментных клеток, называемых хроматофорами . Эти три слоя клеток состоят из меланофоров (занимающих самый глубокий слой), гуанофоров (образующих промежуточный слой и содержащих множество гранул, придающих сине-зеленый цвет) и липофоров (желтых, самый поверхностный слой). Изменение цвета у многих видов инициируется гормонами, выделяемыми гипофизом. В отличие от костистых рыб, здесь нет прямого контроля пигментных клеток со стороны нервной системы, в результате чего изменение окраски происходит медленнее, чем у рыб. Ярко окрашенная кожа обычно указывает на то, что этот вид токсичен, и является предупреждающим знаком для хищников. [ 66 ]
Скелет и передвижение

Скелетная система амфибий структурно гомологична другим четвероногим, хотя и с рядом вариаций. У всех них четыре конечности, за исключением безногих червяг и нескольких видов саламандр с редуцированными конечностями или вообще без них. Кости полые и легкие. Скелетно-мышечная система крепкая, что позволяет ей поддерживать голову и тело. Кости полностью окостеневшие , а позвонки сцепляются друг с другом посредством перекрывающихся отростков. Грудной пояс поддерживается мышцами, а хорошо развитый тазовый пояс крепится к позвоночнику парой крестцовых ребер. наклонена Подвздошная кость вперед, и тело удерживается ближе к земле, чем у млекопитающих. [ 67 ]
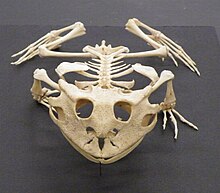
( Ceratophrys cornuta )
У большинства земноводных четыре пальца на передней лапе и пять на задней, но ни на одной из них нет когтей. У некоторых саламандр меньше пальцев, а амфиумы внешне напоминают угрей и имеют крошечные короткие ноги. Сирены — водные саламандры с короткими передними конечностями и отсутствием задних конечностей. Червячие лишены конечностей. Они роют норы наподобие дождевых червей, при этом вдоль тела перемещаются зоны мышечных сокращений. На поверхности земли или в воде они передвигаются, покачивая телом из стороны в сторону. [ 68 ]
У лягушек задние лапы крупнее передних, особенно у тех видов, которые в основном передвигаются прыжками или плаванием. У ходоков и бегунов задние конечности не такие большие, а у норных конечности преимущественно короткие и широкое туловище. Ноги имеют приспособления к образу жизни: перепонки между пальцами для плавания, широкие липкие подушечки пальцев для лазания и ороговевшие бугорки на задних лапах для рытья (лягушки обычно роют почву задом наперед). У большинства саламандр конечности короткие, более или менее одинаковой длины и выступают из тела под прямым углом. Передвижение по суше осуществляется при ходьбе, хвост часто раскачивается из стороны в сторону или используется как опора, особенно при лазании. При их обычной походке одновременно выдвигается только одна нога, как это было у их предков — лопастных рыб. [ 67 ] Некоторые саламандры рода Aneides и некоторые плетодонтиды лазают по деревьям и имеют длинные конечности, большие подушечки пальцев и цепкие хвосты. [ 58 ] У водных саламандр и головастиков лягушек хвост имеет спинные и брюшные плавники и перемещается из стороны в сторону для движения. У взрослых лягушек нет хвостов, а у червяг только очень короткие. [ 68 ]

Саламандры используют свои хвосты для защиты, а некоторые готовы выбросить их за борт, чтобы спасти свою жизнь в процессе, известном как автотомия . Некоторые виды Plethodontidae имеют слабую зону у основания хвоста и легко используют эту стратегию. Хвост часто продолжает дергаться после отделения, что может отвлечь нападавшего и позволить саламандре убежать. И хвосты, и конечности можно регенерировать. [ 69 ] Взрослые лягушки не способны отрастить конечности, но головастики могут это сделать. [ 68 ]
Кровеносная система

- Внутренние жабры, где кровь насыщается кислородом.
- Точка, в которой кровь обедняется кислородом и возвращается к сердцу по венам.
- Двухкамерное сердце
У амфибий есть ювенильная и взрослая стадии, и системы кровообращения у них разные. На стадии молоди (или головастика) кровообращение аналогично рыбьему; двухкамерное сердце перекачивает кровь через жабры, где она насыщается кислородом, и распространяется по всему телу и обратно к сердцу в одном цикле. На взрослой стадии земноводные (особенно лягушки) теряют жабры и развиваются легкие. У них есть сердце, состоящее из одного желудочка и двух предсердий. Когда желудочек начинает сокращаться, дезоксигенированная кровь перекачивается через легочную артерию в легкие. Продолжающееся сокращение затем перекачивает насыщенную кислородом кровь по всему телу. Смешение двух кровотоков сведено к минимуму благодаря анатомии камер. [ 70 ]
Нервная и сенсорная системы
Нервная система в основном такая же, как и у других позвоночных, с центральным мозгом, спинным мозгом и нервами по всему телу. Мозг земноводных относительно прост, но по своей структуре в целом такой же, как у рептилий, птиц и млекопитающих. Их мозг удлинен, за исключением червяг, и содержит обычные двигательные и сенсорные области четвероногих. [ 71 ] Считается, что шишковидное тело , которое, как известно, регулирует режим сна у людей, вырабатывает гормоны, участвующие в спячке и зимнем отдыхе у амфибий. [ 72 ]
Головастики сохраняют систему боковой линии своих предков, но она утрачена у наземных взрослых амфибий. Многие водные саламандры и некоторые червячие обладают электрорецепторами, называемыми ампулярными органами (полностью отсутствующими у бесхвостых), которые позволяют им определять местонахождение объектов вокруг себя при погружении в воду. [ 73 ] Уши у лягушек хорошо развиты. Наружного уха нет, но большая круглая барабанная перепонка лежит на поверхности головы сразу за глазом. Это вибрирует, и звук передается через единственную кость — стремечко — во внутреннее ухо. Таким образом слышны только высокочастотные звуки, такие как брачные крики, а низкочастотные шумы можно обнаружить с помощью другого механизма. [ 67 ] Во внутреннем ухе есть участок специализированных волосковых клеток, называемый papilla amphibiorum , способный улавливать более глубокие звуки. Другой особенностью, уникальной для лягушек и саламандр, является комплекс колумелла-покрышка, примыкающий к слуховой капсуле, который участвует в передаче как воздушных, так и сейсмических сигналов. [ 74 ] Уши саламандр и червяг менее развиты, чем у лягушек, поскольку они обычно не общаются друг с другом посредством звука. [ 75 ]
Глаза головастиков лишены век, но при метаморфозе роговица становится более куполообразной, хрусталик становится более плоским, развиваются веки и связанные с ними железы и протоки. [ 67 ] Глаза взрослых представляют собой усовершенствованную версию глаз беспозвоночных и стали первым шагом в развитии более развитых глаз позвоночных. Они обеспечивают цветовое зрение и глубину фокуса. В сетчатке находятся зеленые палочки, чувствительные к широкому диапазону длин волн. [ 75 ]
Пищеварительная и выделительная системы

- Правое предсердие
- Печень
- Аорта
- Яичная масса
- Двоеточие
- Левое предсердие
- желудочек
- Желудок
- Левое легкое
- Селезенка
- Тонкая кишка
- Клоака
Многие земноводные ловят добычу, высовывая удлиненный язык с липким кончиком и втягивая его обратно в рот, прежде чем схватить предмет челюстями. Некоторые используют инерционное кормление, чтобы помочь им проглотить добычу, неоднократно резко выталкивая голову вперед, заставляя пищу по инерции двигаться назад во рту . Большинство амфибий заглатывают добычу целиком, не пережевывая ее, поэтому желудки у них объемные. Короткий пищевод покрыт ресничками , которые помогают перемещать пищу в желудок, а слизь, вырабатываемая железами во рту и глотке, облегчает ее прохождение. Фермент хитиназа, вырабатываемый в желудке, помогает переваривать хитиновую кутикулу добычи членистоногих. [ 76 ]
Земноводные обладают поджелудочной железой , печенью и желчным пузырем . Печень обычно большая, с двумя долями. Его размер определяется его функцией в качестве единицы хранения гликогена и жира и может меняться в зависимости от времени года по мере накопления или использования этих запасов. Жировая ткань является еще одним важным средством хранения энергии, и это происходит в брюшной полости (во внутренних структурах, называемых жировыми телами), под кожей и, у некоторых саламандр, в хвосте. [ 77 ]
Две почки расположены дорсально, у крыши полости тела. Их работа состоит в том, чтобы фильтровать кровь от метаболических отходов и транспортировать мочу через мочеточники в мочевой пузырь, где она хранится, прежде чем периодически выходить через клоакальное отверстие. Личинки и большинство водных взрослых амфибий выделяют азот в виде аммиака в больших количествах разбавленной мочи, в то время как наземные виды, которым требуется большая потребность в сохранении воды, выделяют менее токсичный продукт - мочевину. Некоторые древесные лягушки с ограниченным доступом к воде выделяют большую часть своих метаболических отходов в виде мочевой кислоты. [ 78 ]
Мочевой пузырь
У большинства водных и полуводных земноводных имеется перепончатая кожа, позволяющая им поглощать воду непосредственно через нее. У некоторых полуводных животных также имеется проницаемая мембрана мочевого пузыря. [ 79 ] В результате у них, как правило, наблюдается высокий уровень выработки мочи, чтобы компенсировать такое высокое потребление воды, и у них в моче мало растворенных солей. Мочевой пузырь помогает таким животным удерживать соли. Некоторые водные амфибии, такие как Xenopus, не поглощают воду повторно, чтобы предотвратить чрезмерный приток воды. [ 80 ] У наземных амфибий обезвоживание приводит к снижению диуреза. [ 81 ]
У земноводных мочевой пузырь обычно очень растяжим и у некоторых наземных видов лягушек и саламандр может составлять от 20% до 50% общей массы тела. [ 81 ] Моча из почек через мочеточники поступает в мочевой пузырь и периодически выделяется из мочевого пузыря в клоаку. [ 82 ]
Дыхательная система

Легкие земноводных примитивны по сравнению с легкими амниот, имеют мало внутренних перегородок и крупных альвеол и, следовательно, имеют сравнительно медленную скорость диффузии кислорода, поступающего в кровь. Вентиляция осуществляется буккальной откачкой . [ 83 ] Однако большинство земноводных способны обмениваться газами с водой или воздухом через кожу. Чтобы обеспечить достаточное кожное дыхание , поверхность их хорошо васкуляризированной кожи должна оставаться влажной, чтобы кислород мог диффундировать с достаточно высокой скоростью. [ 76 ] Поскольку концентрация кислорода в воде увеличивается как при низких температурах, так и при высоких скоростях течения, водные амфибии в этих ситуациях могут полагаться в первую очередь на кожное дыхание, как у водяной лягушки Титикака и саламандры-повелителя ада . В воздухе, где кислород более сконцентрирован, некоторые мелкие виды могут полагаться исключительно на кожный газообмен, наиболее известный из которых — многодонтидные саламандры , у которых нет ни легких, ни жабр. Многие водные саламандры и все головастики имеют жабры на личиночной стадии, а у некоторых (например, аксолотля ) жабры сохраняются и у взрослых водных особей. [ 76 ]
Воспроизведение

Для размножения большинству земноводных требуется пресная вода , хотя некоторые откладывают яйца на суше и разработали различные способы поддержания их влажности. Некоторые из них (например, Fejervarya raja не существует . ) могут обитать в солоноватой воде, но настоящих морских амфибий [ 84 ] Однако есть сообщения об отдельных популяциях амфибий, неожиданно вторгающихся в морские воды. Так было в случае с в Черное море инвазией природного гибрида Pelophylax esculentus , о которой сообщалось в 2010 году. [ 85 ]
Однако несколько сотен видов лягушек в адаптивной радиации (например, Eleutherodactylus , Pacific Platymantis , австрало-папуасские микрогилиды и многие другие тропические лягушки) не нуждаются в воде для размножения в дикой природе . Они размножаются посредством прямого развития — экологической и эволюционной адаптации, которая позволила им быть полностью независимыми от стоячей воды. Почти все эти лягушки живут во влажных тропических лесах , и из их яиц вылупляются непосредственно миниатюрные версии взрослой особи, проходя головастика внутри яйца стадию . Репродуктивный успех многих земноводных зависит не только от количества осадков, но и от сезонности. [ 86 ]
В тропиках многие земноводные размножаются непрерывно или в любое время года. В регионах с умеренным климатом размножение в основном носит сезонный характер, обычно весной, и вызывается увеличением продолжительности дня, повышением температуры или количеством осадков. Эксперименты показали важность температуры, но провоцирующим событием, особенно в засушливых регионах, часто является шторм. У бесхвостых самцы обычно прибывают к местам размножения раньше самок, и производимый ими голосовой хор может стимулировать овуляцию у самок и эндокринную активность самцов, которые еще не являются репродуктивно активными. [ 87 ]
У червяг оплодотворение внутреннее, самец выдвигает интромиттирующий орган , phallodeum и вставление его в женскую клоаку. Парные мюллеровы железы внутри мужской клоаки выделяют жидкость, напоминающую жидкость, вырабатываемую предстательной железой млекопитающих, которая может транспортировать и питать сперму. Оплодотворение, вероятно, происходит в яйцеводе. [ 88 ]
Большинство саламандр также занимаются внутренним оплодотворением . В большинстве из них самец откладывает сперматофор, небольшой пакетик спермы на вершине студенистого конуса, на субстрат либо на суше, либо в воде. Самка подхватывает пакетик со спермой, захватывая его губами клоаки и выталкивая в отверстие. Сперматозоиды перемещаются в сперматеку в крыше клоаки, где они остаются до овуляции, которая может произойти спустя много месяцев. Ритуалы ухаживания и методы передачи сперматофора различаются у разных видов. В некоторых случаях сперматофор может быть помещен непосредственно в клоаку самки, в то время как в других женщину можно направить к сперматофору или удержать с помощью объятия, называемого амплексусом . Некоторые примитивные саламандры из семейств Sirenidae, Hynobiidae и Cryptobranchidae практикуют внешнее оплодотворение аналогично лягушкам: самка откладывает яйца в воду, а самец выделяет сперму на яйцеклетку. [ 88 ]
За некоторыми исключениями, лягушки используют наружное оплодотворение. Самец крепко обхватывает самку передними конечностями либо за руками, либо перед задними ногами, либо, в случае Epipedobates tricolor , за шею. Они остаются в амплексусе, их клоаки расположены близко друг к другу, пока самка откладывает яйца, а самец покрывает их спермой. Шероховатые брачные подушечки на руках самца помогают удерживать хватку. Часто самец собирает и удерживает икру, образуя задними лапами подобие корзины. Исключением является гранулярная ядовитая лягушка ( Oophaga granulifera ), у которой самец и самка размещают свои клоаки в непосредственной близости, глядя в противоположные стороны, а затем одновременно выделяют яйцеклетки и сперму. ( Хвостатая лягушка Ascaphus truei ) обладает внутренним оплодотворением. «Хвост» принадлежит только самцам, является продолжением клоаки и используется для оплодотворения самки. Эта лягушка живет в быстрых ручьях, и внутреннее оплодотворение предотвращает вымывание спермы до того, как произойдет оплодотворение. [ 89 ] Сперму можно хранить в пробирках, прикрепленных к яйцеводу, до следующей весны. [ 90 ]
Большинство лягушек можно отнести к категории продолжительного или взрывного размножения. Обычно длительные заводчики собираются на месте размножения, причем самцы обычно прибывают первыми, звонят и обустраивают территории. Другие самцы-спутники тихо остаются поблизости, ожидая возможности захватить территорию. Самки прилетают спорадически, происходит выбор партнера и откладка яиц. Самки уходят, и территории могут переходить из рук в руки. Появляется больше самок, и со временем сезон размножения подходит к концу. С другой стороны, взрывоопасные размножители встречаются там, где после дождя в засушливых регионах появляются временные лужи. Эти лягушки, как правило, относятся к ископаемым видам, которые появляются после проливных дождей и собираются в местах размножения. Их привлекает туда призыв первого самца, который найдет подходящее место, например лужу, образующуюся на одном и том же месте каждый сезон дождей. Собравшиеся лягушки могут кричать в унисон, и начинается бешеная активность: самцы пытаются спариться с обычно меньшим количеством самок. [ 89 ]

У саламандр и тритонов существует прямая конкуренция между самцами за внимание самок, а тщательно продуманные ухаживания позволяют удержать внимание самки достаточно долго, чтобы она заинтересовалась выбором его для спаривания . [ 91 ] Некоторые виды хранят сперму в течение длительных сезонов размножения, поскольку дополнительное время может позволить взаимодействовать с конкурирующими сперматозоидами. [ 92 ]
Однополое размножение
Однополые самки кротов-саламандр (род Ambystoma) распространены в районе Великих озер Северной Америки. [ 93 ] Эти саламандры являются старейшей известной однополой линией позвоночных, появившейся около 5 миллионов лет назад. [ 94 ] Обмен геномом иногда может происходить между однополой самкой Ambystoma и самцами симпатрических половых видов. [ 94 ]
Жизненный цикл
Большинство земноводных проходят метаморфоз — процесс значительных морфологических изменений после рождения. При типичном развитии земноводных яйца откладываются в воде, а личинки приспосабливаются к водному образу жизни. Лягушки, жабы и саламандры вылупляются из яйца в виде личинок с внешними жабрами. Метаморфоз у земноводных регулируется тироксина концентрацией в крови , стимулирующего метаморфоз, и пролактина , противодействующего действию тироксина. Конкретные события зависят от пороговых значений для разных тканей. [ 95 ] Поскольку большая часть эмбрионального развития происходит вне родительского тела, оно подвержено множеству адаптаций из-за конкретных условий окружающей среды. По этой причине у головастиков вместо зубов могут быть роговые гребни , отростки кожи или плавники, похожие на усы. Они также используют сенсорный орган боковой линии, аналогичный рыбьему. После метаморфоза эти органы становятся ненужными и будут реабсорбированы в результате контролируемой гибели клеток, называемой апоптозом . Разнообразие адаптаций земноводных к конкретным условиям окружающей среды велико, и многие открытия все еще сделаны. [ 96 ]
Яйца

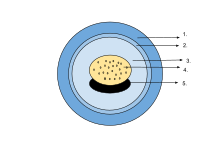
- Желейная капсула
- Желточные оболочки
- Перивителлиновая жидкость
- Желточная пробка
- Эмбрион
В яйце эмбрион находится в перивителлиновой жидкости и окружен полупроницаемыми желатиновыми капсулами, а желточная масса обеспечивает питательные вещества. Когда личинки вылупляются, капсулы растворяются под действием ферментов, выделяемых железой на кончике морды. [ 75 ] Яйца некоторых саламандр и лягушек содержат одноклеточные зеленые водоросли. Они проникают в желеобразную оболочку после откладки яиц и могут увеличить снабжение эмбриона кислородом посредством фотосинтеза. Похоже, они ускоряют развитие личинок и снижают смертность. [ 97 ] Было обнаружено, что у лесной лягушки ( Rana sylvatica ) внутренняя часть шаровидного скопления яиц на 6 ° C (11 ° F) теплее, чем окружающая среда, что является преимуществом в ее прохладной северной среде обитания. [ 98 ]
Яйца могут откладываться поодиночке, группами или длинными нитями. Местами для откладки яиц являются вода, грязь, норы, мусор, а также растения, под бревнами или камнями. [ 99 ] ( Тепличная лягушка Eleutherodactylus planirostris ) откладывает яйца небольшими группами в почву, где примерно за две недели они развиваются непосредственно в молодых лягушек без промежуточной личиночной стадии. [ 100 ] Тунгара лягушка ( Physalaemus pustulosus ) строит плавучее гнездо из пены, чтобы защитить свои яйца. Сначала строится плот, затем в центр откладываются яйца и, наконец, накладывается пенопластовая шапка. Пена обладает антимикробными свойствами. Он не содержит детергентов , а создается путем взбивания белков и лектинов, выделяемых самкой. [ 101 ] [ 102 ]
Личинки

Яйца земноводных обычно откладываются в воде, из них вылупляются свободноживущие личинки, которые завершают свое развитие в воде и позже превращаются в водных или наземных взрослых особей. У многих видов лягушек и у большинства безлегочных саламандр (Plethodontidae) происходит прямое развитие: личинки растут внутри яиц и превращаются в миниатюрных взрослых особей. Многие червячие и некоторые другие амфибии откладывают яйца на суше, а только что вылупившиеся личинки извиваются или переносятся в водоемы. Некоторые червячие, альпийская саламандра ( Salamandra atra ) и часть африканских живородящих жаб ( Nectophrynoides spp. ) являются живородящими . Их личинки питаются железистыми выделениями и развиваются внутри яйцевода самки, часто в течение длительного времени. Другие амфибии, кроме червяг, яйцеживородящие . Яйца сохраняются внутри или на теле родителей, но личинки питаются желтками своих яиц и не получают питания от взрослых особей. Личинки появляются на разных стадиях своего роста, до или после метаморфоза, в зависимости от их вида. [ 103 ] Род жаб Nectophrynoides демонстрирует все эти закономерности развития среди своих дюжины или около того представителей. [ 12 ] Личинки земноводных известны как головастики . У них толстое округлое тело с мощным мускулистым хвостом. [ 78 ]
Лягушки
В отличие от других земноводных, головастики лягушек не похожи на взрослых особей. [ 104 ] Свободноживущие личинки обычно ведут полностью водный образ жизни, но головастики некоторых видов (например, Nannophrys ceylonensis ) ведут полуназемный образ жизни и обитают среди влажных камней. [ 105 ] Головастики имеют хрящевой скелет, жабры для дыхания (сначала внешние, затем внутренние), системы боковой линии и большие хвосты, которые они используют для плавания. [ 106 ] У только что вылупившихся головастиков вскоре появляются жаберные мешки, закрывающие жабры. Эти внутренние жабры и жаберные крышки не гомологичны таковым у рыб. [ 107 ] и встречаются только у головастиков, поскольку и у саламандр, и у червяг есть только внешние жабры. [ 108 ] В сочетании с буккальной перекачкой внутренние жабры позволили головастикам вести образ жизни с фильтрующим питанием , хотя с тех пор некоторые виды выработали другие типы стратегий питания. [ 109 ] Легкие развиваются рано и используются в качестве вспомогательных органов дыхания: головастики поднимаются к поверхности воды, чтобы глотать воздух. Некоторые виды завершают свое развитие внутри яйца и вылупляются прямо в маленьких лягушек. У этих личинок нет жабр, но есть специальные участки кожи, через которые происходит дыхание. Хотя у головастиков нет настоящих зубов, у большинства видов челюсти имеют длинные параллельные ряды небольших ороговевших структур, называемых керадонтами, окруженных роговым клювом. [ 110 ] Передние ноги формируются под жаберным мешком, а задние становятся видны через несколько дней.
Йод и Т4 (чрезмерно стимулируют впечатляющий апоптоз [запрограммированную гибель клеток] клеток личиночных жабр, хвоста и плавников) также стимулируют эволюцию нервной системы, превращая водного головастика-вегетарианца в наземную плотоядную лягушку с лучшими неврологическими и зрительно-пространственными способностями. , обонятельные и когнитивные способности для охоты. [ 111 ] [ 112 ]
На самом деле головастики, развивающиеся в прудах и ручьях, обычно травоядны . Прудовые головастики, как правило, имеют глубокое тело, большие хвостовые плавники и маленькие рты; они плавают в тихих водах, питаясь растущими или рыхлыми фрагментами растительности. Обитатели ручьев обычно имеют более крупные рты, мелкое тело и хвостовые плавники; они прикрепляются к растениям и камням и питаются поверхностными пленками водорослей и бактерий. [ 113 ] Они также питаются диатомовыми водорослями , отфильтрованными из воды через жабры , и взбалтывают осадок на дне пруда, заглатывая съедобные фрагменты. У них относительно длинный кишечник спиралевидной формы, позволяющий им переваривать эту диету. [ 113 ] Некоторые виды на стадии головастика плотоядны и поедают насекомых, более мелких головастиков и рыбу. Молодняк кубинской квакши ( Osteopilus septentrionalis ) иногда может быть каннибалом : более молодые головастики нападают на более крупных и развитых головастиков, когда тот подвергается метаморфозу. [ 114 ]
При метаморфозе происходят быстрые изменения в организме, поскольку образ жизни лягушки полностью меняется. Спиралевидный рот с роговыми зубными гребнями реабсорбируется вместе со спиральной кишкой. У животного развивается большая челюсть, а его жабры исчезают вместе с жаберным мешком. Быстро растут глаза и ноги, формируется язык. С этим связаны изменения в нейронных сетях, такие как развитие стереоскопического зрения и потеря системы боковой линии. Все это может произойти примерно за сутки. Через несколько дней хвост реабсорбируется из-за необходимой для этого более высокой концентрации тироксина. [ 113 ]
Саламандры

( Амбистома макродактилум )

( Амбистома макродактилум )

( Ихтиозаура альпестрис )
При вылуплении типичная личинка саламандры имеет глаза без век, зубы как на верхней, так и на нижней челюсти, три пары перистых наружных жабр и длинный хвост со спинными и брюшными плавниками. Передние конечности могут быть частично развиты, а задние конечности рудиментарны у видов, обитающих в прудах, но могут быть более развиты у видов, которые размножаются в движущейся воде. Личинки прудового типа часто имеют пару балансиров, стержнеобразных структур по обе стороны головы, которые могут предотвратить засорение жабр осадком. [ 115 ] [ 116 ] Оба из них способны размножаться. [ 117 ] У некоторых есть личинки, которые никогда полностью не развиваются во взрослую форму — состояние, известное как неотения . [ 118 ] Неотения возникает, когда скорость роста животного очень низкая и обычно связана с неблагоприятными условиями, такими как низкая температура воды, которые могут изменить реакцию тканей на гормон тироксин. [ 119 ] а также отсутствие еды. Существует пятнадцать видов облигатных неотенических саламандр, включая виды Necturus , Proteus и Amphiuma , а также множество примеров факультативных саламандр, таких как северо-западная саламандра ( Ambystoma gracile ) и тигровая саламандра ( A. tigrinum ), которые принимают эту стратегию в соответствующих условиях окружающей среды. обстоятельства. [ 118 ]
Безлегочные саламандры семейства Plethodontidae ведут наземный образ жизни и откладывают небольшое количество непигментированных яиц скоплением среди влажной листовой подстилки. Каждое яйцо имеет большой желточный мешок, и личинка питается им, пока развивается внутри яйца, образуя полностью сформированную молодую саламандру. Самка саламандры часто высиживает яйца. У представителей рода Ensatinas было замечено, что самка обвивает их и прижимает к ним область горла, эффективно массируя их слизистым секретом. [ 120 ]
У тритонов и саламандр метаморфоз менее драматичен, чем у лягушек. Это связано с тем, что личинки уже плотоядны и продолжают питаться как хищники, когда становятся взрослыми, поэтому в их пищеварительной системе требуется мало изменений. Их легкие начинают функционировать рано, но личинки не используют их так активно, как головастики. Их жабры никогда не покрыты жаберными мешками и рассасываются непосредственно перед тем, как животные покидают воду. Другие изменения включают уменьшение размера или потерю хвостовых плавников, закрытие жаберных щелей, утолщение кожи, развитие век и некоторые изменения в строении зубов и языка. Саламандры наиболее уязвимы во время метаморфоза, поскольку скорость плавания снижается, а трансформирующиеся хвосты становятся помехой на суше. [ 121 ] Взрослые саламандры часто имеют водную фазу весной и летом и наземную фазу зимой. Для адаптации к водной фазе необходим гормон пролактин, а для адаптации к сухопутной фазе — тироксин. Наружные жабры не возвращаются в последующие водные фазы, поскольку они полностью поглощаются при первом выходе из воды. [ 115 ]
Цецилианцы

Большинство наземных червяг, откладывающих яйца, делают это в норах или влажных местах на суше вблизи водоемов. Развитие молоди Ichthyophis Glutinosus , вида из Шри-Ланки, хорошо изучено. Угреобразные личинки вылупляются из яиц и направляются в воду. У них три пары наружных красных перистых жабр, тупая голова с двумя рудиментарными глазами, система боковой линии и короткий хвост с плавниками. Они плавают, покачивая телом из стороны в сторону. В основном они активны ночью, вскоре теряют жабры и совершают вылазки на сушу. Метаморфоза постепенная. Примерно к десяти месяцам у них появилась заостренная голова с чувствующими щупальцами возле рта, они лишились глаз, системы боковой линии и хвоста. Кожа утолщается, появляются вросшие чешуйки, тело делится на сегменты. К этому времени червяк построил нору и живет на суше. [ 122 ]

У большинства видов червяг детеныши производятся путем живорождения. Типичным из них является Typhlonectes compressicauda — вид из Южной Америки. Одновременно в яйцеводе может развиваться до девяти личинок. Они удлиненной формы, имеют парные мешкообразные жабры, маленькие глаза и специальные скребущие зубы. Сначала они питаются желтками яиц, но по мере того, как этот источник питания уменьшается, они начинают царапать клетки мерцательного эпителия, выстилающие яйцевод. Это стимулирует секрецию жидкости, богатой липидами и мукопротеинами, которой они питаются вместе соскобами со стенки яйцевода. До рождения они могут увеличивать свою длину в шесть раз и составлять две пятых длины своей матери. К этому времени они претерпели метаморфозу, потеряли глаза и жабры, у них появилась более толстая кожа и ротовые щупальца, а зубы снова рассосались. Постоянный набор зубов вырастает вскоре после рождения. [ 123 ] [ 124 ]
Жабры есть только обязательно во время эмбрионального развития, а у видов, дающих потомство, потомство рождается после дегенерации жабр. У яйцекладущих червяг жабры либо рассасываются перед вылуплением, либо, у видов, которые вылупляются с еще присутствующими остатками жабр, живут недолго и оставляют после себя только жаберную щель. У видов, имеющих чешуйки под кожей, чешуя не образуется раньше во время метаморфоза. [ 125 ]
Кольчатая червяга ( Siphonops annulatus ) выработала уникальное приспособление для целей размножения. Потомство питается слоем кожи, специально развитым взрослой особью, в результате явления, известного как материнская дерматофагия. Выводок кормят порциями в течение примерно семи минут с интервалом примерно в три дня, что дает коже возможность восстановиться. Между тем было замечено, что они заглатывают жидкость, выделяемую из материнской клоаки. [ 126 ]
Родительская забота

Забота о потомстве у земноводных мало изучена, но, как правило, чем больше яиц в партии, тем менее вероятно, что родительская забота имеет место. Тем не менее, по оценкам, у 20% видов земноводных одна или обе взрослые особи играют определенную роль в уходе за молодняком. [ 127 ] Те виды, которые размножаются в небольших водоемах или других специализированных средах обитания, как правило, имеют сложные модели поведения при уходе за своим потомством. [ 128 ]
Многие лесные саламандры откладывают кладки яиц под мертвые бревна или камни на суше. Черная горная саламандра ( Desmognathus welteri ) делает это, мать высиживает яйца и охраняет их от хищников, пока эмбрионы питаются желтками яиц. Когда они полностью развиваются, они вырываются из яйцевых капсул и рассеиваются в виде молодых саламандр. [ 129 ] Самец адского мага, примитивная саламандра, выкапывает подводное гнездо и побуждает самок лежать там. Затем самец охраняет это место в течение двух или трех месяцев до вылупления яиц, используя движения тела, чтобы обмахивать яйца и увеличивать снабжение их кислородом. [ 57 ]

Самец Colostethus subpunctatus , крошечная лягушка, охраняет скопление яиц, спрятанное под камнем или бревном. Когда икра вылупляется, самец переносит на своей спине головастиков, застрявших там из-за слизистого секрета, во временный водоем, где окунается в воду, и головастики падают. [ 130 ] Самец жабы-акушерки ( Alytes obstetricans ) обматывает нити яиц вокруг своих бедер и носит их до восьми недель. Он сохраняет их влажными, а когда они готовы вылупиться, посещает пруд или канаву и выпускает головастиков. [ 131 ] Самка лягушки , насиживающей желудок ( Rheobatrachus spp. ), выращивала личинок в желудке после проглатывания яиц или птенцов; однако эта стадия никогда не наблюдалась до вымирания вида. Головастики выделяют гормон, который подавляет пищеварение у матери во время развития, потребляя очень большой запас желтка. [ 132 ] ( Сумчатая лягушка Assa darlingtoni ) откладывает яйца на землю. Когда они вылупляются, самец носит головастиков в выводковых мешочках на задних лапах. [ 133 ] Водная суринамская жаба ( Pipa pipa ) выращивает детенышей в порах на спине, где они остаются до метаморфоза. [ 134 ] Гранулярная ядовитая лягушка ( Oophaga granulifera ) типична для ряда древесных лягушек семейства древолазов семейства Dendrobatidae . Яйца откладываются на лесной подстилке, а когда они вылупляются, головастики по одному переносятся на спине взрослой особи в подходящую, заполненную водой расщелину, например, в листа или розетку бромелии пазуху . Самка регулярно посещает места выгула и откладывает в воду неоплодотворенные икринки, которые поедаются головастиками. [ 135 ]
Генетика и геномика
Земноводные выделяются среди позвоночных разнообразием хромосом и геномов. Кариотипы диплоидных ( хромосомы ) определены как минимум у 1193 (14,5%) из ≈8200 известных ( ) видов, включая 963 бесхвостых животных , 209 саламандр и 21 червяг . В целом кариотипы диплоидных амфибий характеризуются 20–26 двуплечими хромосомами. Земноводные также имеют очень большие геномы по сравнению с другими таксонами позвоночных и соответствующие различия в размере генома ( значение C : пикограммы ДНК в гаплоидных ядрах). Размеры генома лягушек колеблются от 0,95 до 11,5 пг, у саламандр от 13,89 до 120,56 пг и у червяг от 2,94 до 11,78 пг. [ 136 ]
Большие размеры генома не позволили провести полногеномное секвенирование амфибий, хотя в последнее время был опубликован ряд геномов. Проект генома Xenopus тропического размером 1,7 ГБ был первым, о котором сообщалось для амфибий в 2010 году. [ 136 ] По сравнению с некоторыми саламандрами геном этой лягушки крошечный. Например, геном мексиканского аксолотля оказался размером 32 Гб, что более чем в 10 раз превышает размер генома человека (3 ГБ). [ 137 ]
Кормление и диета

( Ambystoma gracile ) поедает червя.
За некоторыми исключениями, взрослые амфибии — хищники , питающиеся практически всем, что движется, и могут проглотить. Диета в основном состоит из мелкой добычи, которая не движется слишком быстро, например жуков, гусениц, дождевых червей и пауков. Сирены ( Siren spp. ) часто заглатывают водный растительный материал вместе с беспозвоночными, которыми они питаются. [ 138 ] а бразильская древесная лягушка ( Xenohyla truncata ) включает в свой рацион большое количество фруктов. [ 139 ] Мексиканская роющая жаба ( Rhinophrynus dorsalis ) имеет специально приспособленный язык для ловли муравьев и термитов. Он выдвигает его кончиком вперед, тогда как у других лягушек сначала выдвигается задняя часть, а их языки шарнирно закреплены спереди. [ 140 ]
Еду в основном выбирают на глаз, даже в условиях неяркой освещенности. Движение добычи вызывает реакцию кормления. Лягушек ловили на рыболовные крючки с наживкой из красной фланели, а у зеленых лягушек ( Rana clamitans ) желудки были полны семян вяза, которые они видели проплывающими мимо. [ 141 ] Жабы, саламандры и червяги также используют запах для обнаружения добычи. Эта реакция в основном вторична, поскольку было замечено, что саламандры остаются неподвижными рядом с пахучей добычей, но питаются только в том случае, если она движется. Пещерные амфибии обычно охотятся по запаху. Некоторые саламандры, похоже, научились распознавать неподвижную добычу, когда она не имеет запаха, даже в полной темноте. [ 142 ]
Земноводные обычно заглатывают пищу целиком, но могут сначала слегка пережевать ее, чтобы подчинить себе. [ 46 ] Обычно у них маленькие шарнирные зубы на ножке — особенность, уникальная для амфибий. Их основание и коронка состоят из дентина, разделенного некальцинированным слоем, и они периодически заменяются. Саламандры, червячие и некоторые лягушки имеют один или два ряда зубов в обеих челюстях, но у некоторых лягушек ( Rana spp. ) зубы отсутствуют в нижней челюсти, а у жаб ( Bufo spp. ) зубов нет. У многих земноводных имеются также сошковые зубы, прикрепленные к лицевой кости нёба. [ 143 ]

Тигровая саламандра ( Ambystoma tigrinum ) типична для лягушек и саламандр, которые прячутся под укрытием, готовые устроить засаду на неосторожных беспозвоночных. Другие амфибии, такие как Bufo spp. жабы активно ищут добычу, а аргентинская рогатая лягушка ( Ceratophrys ornata ) приманивает любознательную добычу поближе, поднимая задние лапы над спиной и вибрируя желтыми пальцами ног. [ 144 ] Среди лягушек-подстилок в Панаме лягушки, которые активно охотятся на добычу, имеют узкие рты, тонкие, часто ярко окрашенные и ядовитые, тогда как лягушки, устраивающие засады, имеют широкие рты, широкие и хорошо замаскированные. [ 145 ] Цецилии не щелкают языком, а ловят добычу, хватая ее слегка направленными назад зубами. Борьба добычи и дальнейшие движения челюстей толкают ее внутрь, и червяки обычно прячутся в свою нору. Покоренная добыча заглатывается целиком. [ 146 ]
Вылупившиеся личинки лягушки питаются желтком яйца. Когда запасы истощаются, некоторые переходят на питание бактериями, водорослевыми корками, детритом и шероховатостями подводных растений. Вода всасывается через рот, который обычно находится в нижней части головы, и проходит через жаберные пищевые ловушки между ртом и жабрами, где мелкие частицы улавливаются слизью и отфильтровываются. У других есть специализированный ротовой аппарат, состоящий из рогового клюва, окаймленного несколькими рядами губных зубов. Они соскребают и откусывают разнообразную пищу, а также взбалтывают донный осадок, отфильтровывая более крупные частицы с помощью сосочков вокруг рта. Некоторые из них, например чесночноногие жабы, имеют сильные кусающие челюсти и являются плотоядными или даже каннибалистическими. [ 147 ]
Вокализация

Крики червяг и саламандр ограничиваются случайным тихим писком, хрюканьем или шипением и мало изучены. Щелкающий звук, иногда издаваемый червягами, может быть средством ориентации, как у летучих мышей, или формой общения. Большинство саламандр считаются безголосыми , но калифорнийская гигантская саламандра ( Dicamptodon ensatus ) имеет голосовые связки и может издавать дребезжащий или лающий звук. Некоторые виды саламандр при нападении издают тихий писк или визг. [ 148 ]

Лягушки гораздо более разговорчивы, особенно во время сезона размножения, когда они используют свой голос для привлечения партнеров. Присутствие определенного вида на территории легче обнаружить по его характерному крику, чем по мимолетному взгляду на само животное. У большинства видов звук производится путем вытеснения воздуха из легких через голосовые связки в воздушный мешок или мешочки в горле или в углу рта. Он может раздуваться, как воздушный шар, и действовать как резонатор, помогая передавать звук в атмосферу или воду в моменты, когда животное погружается в воду. [ 148 ] Основная вокализация - это громкий рекламный призыв самца, который направлен как на то, чтобы побудить самку приблизиться, так и на то, чтобы отговорить других самцов от вторжения на ее территорию. Этот призыв изменяется на более тихий брачный призыв при приближении самки или на более агрессивную версию, если приближается злоумышленник-самец. Призыв сопряжен с риском привлечения хищников и требует затрат большого количества энергии. [ 149 ] Другие призывы включают призывы, исходящие от женщины в ответ на рекламный призыв, и призыв к освобождению, исходящий от мужчины или женщины во время нежелательных попыток амплексуса. Когда на лягушку нападают, она издает сигнал бедствия или испуга, часто напоминающий крик. [ 150 ] Обычно ночная кубинская древесная лягушка ( Osteopilus septentrionalis ) издает сигнал дождя, когда в дневное время идут дожди. [ 151 ]
Территориальное поведение
Мало что известно о территориальном поведении червяг, но некоторые лягушки и саламандры защищают свои участки обитания. Обычно это места кормления, размножения или укрытия. Такое поведение обычно демонстрируют самцы, хотя у некоторых видов также участвуют самки и даже молодые особи. Хотя у многих видов лягушек самки крупнее самцов, это не относится к большинству видов, у которых самцы активно участвуют в территориальной защите. Некоторые из них имеют особые приспособления, такие как увеличенные зубы для кусания или шипы на груди, руках или больших пальцах. [ 152 ]

У саламандр защита территории предполагает принятие агрессивной позиции и при необходимости нападение на злоумышленника. Это может включать в себя щелканье, преследование, а иногда и укусы, что иногда приводит к потере хвоста. Поведение красноспинных саламандр ( Plethodon cinereus ) хорошо изучено. 91% отмеченных особей, которые позже были пойманы повторно, находились в пределах метра (ярда) от места своего первоначального дневного убежища под бревном или камнем. [ 153 ] Аналогичная часть, экспериментально перемещенная на расстояние 30 метров (98 футов), вернулась на свою базу. [ 153 ] Саламандры оставляли запаховые следы на своей территории, размер которой в среднем составлял от 0,16 до 0,33 квадратных метров (от 1,7 до 3,6 квадратных футов), и иногда на них проживала пара самца и самки. [ 154 ] Они сдерживали вторжение других и очерчивали границы между соседними районами. Большая часть их поведения казалась стереотипной и не предполагала какого-либо реального контакта между людьми. Агрессивная поза заключалась в том, чтобы поднять тело над землей и пристально посмотреть на противника, который часто покорно отворачивался. Если злоумышленник упорствовал, он обычно наносил кусающий выпад либо в область хвоста, либо в носогубные бороздки. Повреждение любой из этих областей может снизить физическую форму соперника либо из-за необходимости регенерации тканей, либо из-за ухудшения его способности обнаруживать пищу. [ 153 ]
У лягушек в местах размножения часто наблюдается территориальное поведение самцов; Звонок – это одновременно и объявление о принадлежности части этого ресурса, и рекламный звонок потенциальным партнерам. В общем, более глубокий голос означает более тяжелого и сильного человека, и этого может быть достаточно, чтобы предотвратить вторжение более мелких самцов. На вокализацию тратится много энергии, и это наносит ущерб владельцу территории, который может быть заменен более сильным соперником, если он утомится. Самцы склонны терпимо относиться к владельцам соседних территорий, одновременно энергично нападая на неизвестных злоумышленников. Владельцы территорий имеют «домашнее преимущество» и обычно выигрывают в столкновении двух лягушек одинакового размера. Если угроз недостаточно, могут произойти драки грудь к груди. Методы борьбы включают в себя толчки и толчки, сдувание голосового мешка противника, захват его за голову, прыжки на спину, кусание, преследование, разбрызгивание и ныряние под воду. [ 155 ]
Защитные механизмы

У земноводных мягкие тела с тонкой кожей, у них отсутствуют когти, защитная броня или шипы. Тем не менее, они выработали различные защитные механизмы, чтобы сохранить свою жизнь. Первой линией защиты у саламандр и лягушек является вырабатываемый ими слизистый секрет. Это сохраняет их кожу влажной, делает их скользкими и трудными для захвата. Секреция часто бывает липкой, неприятной или токсичной. [ 156 ] Было замечено, что змеи зевают и открывают рот при попытке проглотить африканских когтистых лягушек ( Xenopus laevis ), что дает лягушкам возможность убежать. [ 156 ] [ 157 ] В этом отношении червяги мало изучены, но кайенские червяки ( Typhlonectes compressicauda ) производят токсичную слизь, которая убила хищную рыбу в ходе эксперимента по кормлению в Бразилии. [ 158 ] У некоторых саламандр кожа ядовита. Грубокожий тритон ( Taricha granulosa ) из Северной Америки и другие представители его рода содержат нейротоксин тетродотоксин (ТТХ), наиболее токсичное из известных небелковых веществ, почти идентичное тому, которое вырабатывается рыбой-фугу . Обращение с тритонами не причиняет вреда, но проглатывание даже самого незначительного количества кожи смертельно опасно. В ходе испытаний на кормление было обнаружено, что рыбы, лягушки, рептилии, птицы и млекопитающие восприимчивы. [ 159 ] [ 160 ] Единственными хищниками, имеющими некоторую толерантность к яду, являются определенные популяции обыкновенной подвязочной змеи ( Thamnophis sirtalis ). В местах, где сосуществуют и змеи, и саламандры, змеи выработали иммунитет благодаря генетическим изменениям и безнаказанно питаются амфибиями. [ 161 ] Коэволюция происходит, когда тритон увеличивает свои токсичные возможности с той же скоростью, что и змея, развивая свой иммунитет. [ 160 ] Некоторые лягушки и жабы токсичны: основные ядовитые железы находятся сбоку на шее и под бородавками на спине. Эти области попадают в атакующее животное, и их выделения могут иметь неприятный вкус или вызывать различные физические или неврологические симптомы. Всего из ограниченного числа исследованных видов земноводных было выделено более 200 токсинов. [ 162 ]


Ядовитые виды часто используют яркую окраску, чтобы предупредить потенциальных хищников о своей токсичности. Эти предупреждающие цвета, как правило, представляют собой красный или желтый в сочетании с черным, огненная саламандра ( Salamandra salamandra примером может служить ). Как только хищник попробует один из них, он, скорее всего, запомнит окраску, когда в следующий раз встретит подобное животное. У некоторых видов, таких как огнебрюхая жаба ( Bombina spp. ), предупреждающая окраска находится на животе, и при нападении эти животные принимают защитную позу, демонстрируя хищнику свою яркую окраску. Лягушка Allobates zaparo не ядовита, но имитирует появление других токсичных видов в своей местности - стратегия, которая может обмануть хищников. [ 164 ]
Многие земноводные ведут ночной образ жизни и днем прячутся, избегая тем самым дневных хищников, охотящихся зрением. Другие земноводные используют камуфляж , чтобы не быть обнаруженными. Они имеют различные цвета, такие как пестрый коричневый, серый и оливковый, которые сливаются с фоном. Некоторые саламандры принимают оборонительную позу при столкновении с потенциальным хищником, таким как североамериканская северная короткохвостая землеройка ( Blarina brevicauda ). Их тела корчатся, они поднимают и бьют хвостами, из-за чего хищнику трудно избежать контакта с их зернистыми железами, вырабатывающими яд. [ 165 ] Некоторые саламандры автоматически автоматизируют свои хвосты при нападении, жертвуя этой частью своей анатомии, чтобы иметь возможность спастись. Хвост может иметь перетяжку у основания, позволяющую его легко отсоединить. Хвост регенерируется позже, но затраты энергии на его замену для животного значительны. [ 69 ] Некоторые лягушки и жабы надуваются, чтобы выглядеть большими и свирепыми, а некоторые чесночноногие жабы ( Pelobates spp ) кричат и прыгают на нападавшего. [ 46 ] Гигантские саламандры рода Andrias , а также лягушки Ceratophrine и Pyxicephalus обладают острыми зубами и способны тянуть кровь при защитном укусе. Чернобрюхая саламандра ( Desmognathus Quadramulatus ) может укусить атакующую обыкновенную подвязочную змею ( Thamnophis sirtalis ) в два или три раза больше ее размера за голову, и часто ей удается убежать. [ 166 ]
Познание
У амфибий наблюдаются привыкание , ассоциативное обучение посредством классического и инструментального обучения , а также способности к различению. [ 167 ] Многие считают, что амфибии разумны и способны чувствовать такие эмоции, как тревога и страх . [ 168 ]
В одном эксперименте, когда саламандрам предлагали живых плодовых мух ( Drosophila virilis ), они выбрали большее из 1 против 2 и 2 против 3. Лягушки могут различать небольшие числа (1 против 2, 2 против 3, но не 3 против 4) и большие количества (3 против 6, 4 против 8, но не 4 против 6) добычи. Это не зависит от других характеристик, т.е. площади поверхности, объема, веса и движения, хотя различение больших чисел может основываться на площади поверхности. [ 169 ]
Сохранение

Резкое сокращение популяций амфибий, включая катастрофу популяций и массовое локализованное вымирание , отмечается с конца 1980-х годов в разных местах по всему миру, и поэтому сокращение численности земноводных воспринимается как одна из наиболее серьезных угроз глобальному биоразнообразию . [ 170 ] В 2004 году Международный союз охраны природы (МСОП) сообщил, что в настоящее время птицы, [ 171 ] темпы вымирания млекопитающих и амфибий были как минимум в 48 раз выше, чем темпы естественного вымирания, а возможно, в 1024 раза выше. В 2006 году считалось, что существует 4035 видов земноводных, жизнь которых на определенном этапе жизненного цикла зависит от воды. Из них 1356 (33,6%) считались находящимися под угрозой исчезновения, и эта цифра, вероятно, занижена, поскольку она исключает 1427 видов, по которым не было достаточно данных для оценки их статуса. [ 172 ] Считается, что здесь задействован ряд причин, в том числе разрушение и изменение среды обитания, чрезмерная эксплуатация , загрязнение, интродуцированные виды , глобальное потепление , загрязняющие вещества, нарушающие работу эндокринной системы , разрушение озонового слоя ( ультрафиолетовое излучение особенно вредно для кожи). , глаза и яйца амфибий), а также такие заболевания, как хитридиомикоз . Однако многие причины сокращения численности амфибий до сих пор плохо изучены и являются темой постоянных дискуссий. [ 173 ]

Пищевые сети и хищничество
Любое сокращение численности земноводных повлияет на характер хищничества. Утрата хищных видов, находящихся на вершине пищевой цепи, нарушит хрупкий баланс экосистемы и может привести к резкому увеличению численности условно-патогенных видов.
Хищники, питающиеся земноводными, страдают от их сокращения. Западная наземная подвязочная змея ( Thamnophis elegans ) в Калифорнии в основном ведет водный образ жизни и сильно зависит от двух видов лягушек, численность которых сокращается: йосемитской жабы ( Bufo canorus ) и горной желтоногой лягушки ( Rana muscosa ), что ставит змею будущее под угрозой. Если змеи станет мало, это повлияет на хищных птиц и других хищников, которые ею питаются. [ 174 ] Между тем, в прудах и озерах меньше лягушек означает меньше головастиков. Обычно они играют важную роль в контроле роста водорослей, а также питаются детритом , который накапливается в виде отложений на дне. Уменьшение количества головастиков может привести к чрезмерному росту водорослей, что приведет к истощению кислорода в воде, когда водоросли позже погибнут и разложатся. Тогда водные беспозвоночные и рыбы могут погибнуть, что приведет к непредсказуемым экологическим последствиям. [ 175 ]
Загрязнение и пестициды
Сокращение популяций амфибий и рептилий привело к осознанию воздействия пестицидов на рептилий и амфибий. [ 176 ] В прошлом аргумент о том, что амфибии или рептилии более восприимчивы к любому химическому загрязнению, чем любые наземно-водные позвоночные, до недавнего времени не подтверждался исследованиями. [ 176 ] Земноводные и рептилии имеют сложный жизненный цикл, живут в разных климатических и экологических зонах и более уязвимы к химическому воздействию. Некоторые пестициды, такие как органофосфаты, неоникотиноиды и карбаматы, реагируют посредством ингибирования холинэстеразы. Холинэстераза — это фермент, который вызывает гидролиз ацетилхолина, возбуждающего нейромедиатора, которого много в нервной системе. Ингибиторы АХЭ бывают обратимыми или необратимыми, а карбаматы более безопасны, чем фосфорорганические инсектициды, которые с большей вероятностью вызывают холинергическое отравление. Воздействие на рептилий пестицидов, ингибирующих AChE, может привести к нарушению нервной функции у рептилий. Нарастание этих тормозящих эффектов на двигательные функции, такие как потребление пищи и другие виды деятельности.
Стратегии сохранения и защиты
Группа специалистов по амфибиям МСОП возглавляет усилия по реализации комплексной глобальной стратегии по сохранению амфибий. [ 177 ] Amphibian Ark — это организация, созданная для реализации рекомендаций этого плана по сохранению ex-situ. Они сотрудничают с зоопарками и аквариумами по всему миру, поощряя их создавать страховые колонии для находящихся под угрозой исчезновения амфибий. [ 177 ] Одним из таких проектов является Панамский проект по спасению и сохранению амфибий, основанный на существующих природоохранных усилиях в Панаме с целью создания общенационального ответа на угрозу хитридиомикоза. [ 178 ]
Другой мерой могло бы стать прекращение эксплуатации лягушек в пищу человеку. На Ближнем Востоке растущий аппетит к поеданию лягушачьих лапок и последующий сбор их в пищу уже был связан с увеличением численности комаров и, таким образом, имеет прямые последствия для здоровья человека. [ 179 ]
См. также
- Туннель для амфибий и рептилий
- Культурные изображения земноводных
- Список амфибий
- Список родов амфибий
- Список находящихся под угрозой исчезновения рептилий и амфибий США
- Мягкопанцирная черепаха - таксономическое семейство нескольких родов черепах , способных «дышать» под водой ритмичными движениями ротовой полости, содержащей многочисленные отростки, обильно снабжаемые кровью, действующие аналогично жаберным нитям у рыб.
- Кольчатая морская змея - вид ядовитой морской змеи , которая способна дышать под водой с помощью обширной сосудистой сети на макушке головы и поглощать кислород из окружающей воды.
- Кожное дыхание
Ссылки
- ^ Марьянович, Д. (2021). «Создание калибровочной колбасы на примере повторной калибровки транскриптомного древа времени челюстных позвоночных» . Границы генетики . 12 . 521693. дои : 10.3389/fgene.2021.521693 . ПМК 8149952 . ПМИД 34054911 .
- ^ Jump up to: а б Блэкберн, округ Колумбия; Уэйк, Д.Б. (2011). «Класс Amphibia Grey, 1825. В: Чжан, Z.-Q. (Ред.) Биоразнообразие животных: план классификации более высокого уровня и обзор таксономического богатства» (PDF) . Зоотакса . 3148 : 39–55. дои : 10.11646/zootaxa.3148.1.8 . Архивировано (PDF) из оригинала 18 мая 2016 г. Проверено 29 ноября 2012 г.
- ^ Лэмб, Дженнифер Ю.; Дэвис, Мэтью П. (27 февраля 2020 г.). «Саламандры и другие земноводные светятся биофлуоресценцией» . Научные отчеты . 10 (1): 2821. Бибкод : 2020NatSR..10.2821L . дои : 10.1038/S41598-020-59528-9 . ISSN 2045-2322 . ПМК 7046780 . ПМИД 32108141 . S2CID 257031840 . Викиданные Q89930490 .
- ^ Jump up to: а б Аткинс, Джейд Б.; Рейс, Роберт Р.; Мэддин, Хиллари К. (22 марта 2019 г.). «Упрощение мозговой оболочки и происхождение лисамфибий» . ПЛОС ОДИН . 14 (3): e0213694. Бибкод : 2019PLoSO..1413694A . дои : 10.1371/journal.pone.0213694 . ISSN 1932-6203 . ПМК 6430379 . ПМИД 30901341 .
... растет консенсус в отношении того, что лиссамфибии представляют собой монофилетическую совокупность, происходящую из Temnospondyli, а точнее, из диссорофоидов амфибамид.
- ^ Марьянович, Д. и Лаурин, М. (2019) Филогения палеозойских конечностей позвоночных, подвергнутая повторной оценке посредством пересмотра и расширения крупнейшей опубликованной матрицы соответствующих данных . ПирДж :6:e5565. дои: 10.7717/peerj.5565. Электронная коллекция 2019.
- ^ «Снова в школу: темно превосходная степень» . Брайан Джи, доктор философии . Проверено 29 июня 2022 г.
- ^ Скит, Уолтер В. (1897). Краткий этимологический словарь английского языка . Кларендон Пресс. п. 39.
- ^ Бэрд, Дональд (май 1965 г.). «Палеозойские лепоспондиловые амфибии» . Интегративная и сравнительная биология . 5 (2): 287–294. дои : 10.1093/icb/5.2.287 .
- ^ Jump up to: а б с «Виды по количеству» . АмфибияВеб. Архивировано из оригинала 12 января 2021 года . Проверено 11 января 2021 г.
- ^ Фрост, Даррел (2013). «Американский музей естественной истории: виды земноводных мира 5.6, онлайн-справочник» . Американский музей естественной истории. Архивировано из оригинала 14 июня 2012 года . Проверено 24 октября 2013 г.
- ^ «Паутина-амфибия» . Архивировано из оригинала 25 ноября 2019 года . Проверено 1 апреля 2019 г.
- ^ Jump up to: а б Крамп, Марта Л. (2009). «Разнообразие и история жизни земноводных» (PDF) . Экология и охрана амфибий. Справочник по методам : 3–20. дои : 10.1093/oso/9780199541188.003.0001 . ISBN 978-0-19-954118-8 . Архивировано из оригинала (PDF) 15 июля 2011 года.
- ^ Шпеер, Б.В.; Ваггонер, Бен (1995). «Амфибия: Систематика» . Музей палеонтологии Калифорнийского университета. Архивировано из оригинала 7 марта 2013 года . Проверено 13 декабря 2012 г.
- ^ Jump up to: а б с Стеббинс и Коэн 1995 , с. 3.
- ^ Jump up to: а б с Андерсон, Дж.; Рейс, Р.; Скотт, Д.; Фрёбиш, Н.; Сумида, С. (2008). «Стеблевой батрах из ранней перми Техаса и происхождение лягушек и саламандр» . Природа . 453 (7194): 515–518. Бибкод : 2008Natur.453..515A . дои : 10.1038/nature06865 . ПМИД 18497824 . S2CID 205212809 . Архивировано из оригинала 14 июля 2021 года . Проверено 10 ноября 2016 г.
- ^ Рочек, З. (2000). «14. Мезозойские амфибии» (PDF) . В Хитволе, Х.; Кэрролл, Р.Л. (ред.). Биология амфибий: Палеонтология: эволюционная история земноводных . Том. 4. Суррей Битти и сыновья. стр. 1295–1331. ISBN 978-0-949324-87-0 . Архивировано (PDF) из оригинала 18 июля 2011 г. Проверено 29 сентября 2012 г.
- ^ Дженкинс, Фариш А. младший; Уолш, Денис М.; Кэрролл, Роберт Л. (2007). «Анатомия Eocaecilia micropodia , конечностей червяг ранней юры». Вестник Музея сравнительной зоологии . 158 (6): 285–365. doi : 10.3099/0027-4100(2007)158[285:AOEMAL]2.0.CO;2 . S2CID 86379456 .
- ^ Гаоа, Кэ-Цинь; Шубин, Нил Х. (2012). «Позднеюрский саламандроид из западного Ляонина, Китай» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (15): 5767–5772. Бибкод : 2012PNAS..109.5767G . дои : 10.1073/pnas.1009828109 . ПМЦ 3326464 . ПМИД 22411790 .
- ^ Каннателла, Дэвид (2008). «Салиентия» . Веб-проект «Древо жизни» . Архивировано из оригинала 22 апреля 2019 года . Проверено 31 августа 2012 г.
- ^ Jump up to: а б с «Эволюция земноводных» . Университет Вайкато: Эволюция растений и животных. Архивировано из оригинала 27 июня 2016 года . Проверено 30 сентября 2012 г.
- ^ Jump up to: а б с Кэрролл, Роберт Л. (1977). Халлам, Энтони (ред.). Закономерности эволюции, иллюстрированные летописями окаменелостей . Эльзевир. стр. 405–420. ISBN 978-0-444-41142-6 . Архивировано из оригинала 14 апреля 2021 года . Проверено 15 октября 2020 г.
- ^ Jump up to: а б Клак, Дженнифер А. (2006). « Ихтиостега » . Веб-проект «Древо жизни» . Архивировано из оригинала 14 июня 2020 года . Проверено 29 сентября 2012 г.
- ^ Ломбард, RE; Болт, младший (1979). «Эволюция уха четвероногих: анализ и реинтерпретация» . Биологический журнал Линнеевского общества . 11 (1): 19–76. дои : 10.1111/j.1095-8312.1979.tb00027.x . Архивировано из оригинала 4 октября 2018 года . Проверено 10 ноября 2016 г.
- ^ Jump up to: а б с Спочинска, JOI (1971). Окаменелости: исследование эволюции . Frederick Muller Ltd., стр. 120–125. ISBN 978-0-584-10093-8 .
- ^ Сахни, С.; Бентон, М.Дж. и Ферри, Пенсильвания (2010). «Связь между глобальным таксономическим разнообразием, экологическим разнообразием и распространением позвоночных на суше» . Письма по биологии . 6 (4): 544–547. дои : 10.1098/rsbl.2009.1024 . ПМК 2936204 . ПМИД 20106856 .
- ^ Сахни, С.; Бентон, MJ (2008). «Восстановление после самого глубокого массового вымирания всех времен» . Труды Королевского общества B: Биологические науки . 275 (1636): 759–65. дои : 10.1098/rspb.2007.1370 . ПМЦ 2596898 . ПМИД 18198148 .
- ^ Сан-Мауро, Д. (2010). «Многолокусная временная шкала происхождения современных земноводных» . Молекулярная филогенетика и эволюция . 56 (2): 554–561. дои : 10.1016/j.ympev.2010.04.019 . ПМИД 20399871 . Архивировано из оригинала 12 декабря 2019 года . Проверено 10 ноября 2016 г.
- ^ Сан-Мауро, Диего; Винс, Майкл; Альковендас, ВМС; Зардоя, Рафаэль; Мейер, Аксель (2005). «Первоначальное разнообразие современных амфибий предшествовало распаду Пангеи» . Американский натуралист . 165 (5): 590–599. дои : 10.1086/429523 . ПМИД 15795855 . S2CID 17021360 . Архивировано из оригинала 7 февраля . Получено 10 , ноября
- ^ «Крошечные окаменелости раскрывают предысторию самой загадочной из ныне живущих амфибий» . ScienceDaily . Архивировано из оригинала 5 ноября 2019 года . Проверено 28 июня 2017 г.
- ^ Jump up to: а б с д Дорит, Уокер и Барнс 1991 , стр. 843–859.
- ^ Лорен, Мишель (2011). «Наземные позвоночные» . Веб-проект «Древо жизни» . Архивировано из оригинала 1 марта 2011 года . Проверено 16 сентября 2012 г.
- ^ Лорен, Мишель; Готье, Жак А. (2012). «Амниота» . Веб-проект «Древо жизни» . Архивировано из оригинала 1 июня 2020 года . Проверено 16 сентября 2012 г.
- ^ Сумич, Джеймс Л.; Моррисси, Джон Ф. (2004). Введение в биологию морской жизни . Джонс и Бартлетт Обучение. п. 171. ИСБН 978-0-7637-3313-1 . Архивировано из оригинала 14 апреля 2021 года . Проверено 15 октября 2020 г.
- ^ Группа специалистов МСОП по амфибиям SSC. (2020) [версия оценки 2015 года с опечатками]. « Амбистома Андерсони » . Красный список исчезающих видов МСОП . 2015 : e.T59051A176772780. doi : 10.2305/IUCN.UK.2015-4.RLTS.T59051A176772780.en .
- ^ Уэллс, Кентвуд Д. (15 февраля 2010 г.). Экология и поведение амфибий . Издательство Чикагского университета. ISBN 9780226893334 . Архивировано из оригинала 25 июня 2020 года . Проверено 19 июня 2020 г. - через Google Книги.
- ^ Леви, Дэниел Л.; Хилд, Ребекка (20 января 2016 г.). «Проблемы биологического масштабирования и решения у амфибий» . Перспективы Колд-Спринг-Харбор в биологии . 8 (1): а019166. doi : 10.1101/cshperspect.a019166 . ПМЦ 4691792 . ПМИД 26261280 .
- ^ Шох, Райнер Р. (19 марта 2014 г.). Эволюция земноводных: жизнь ранних наземных позвоночных . Джон Уайли и сыновья. ISBN 9781118759134 . Архивировано из оригинала 25 июля 2020 года . Получено 21 июля 2020 г. - через Google Книги.
- ^ Мишель Лорен (2004). «Эволюция размеров тела, правило Коупа и происхождение амниот» . Систематическая биология . 53 (4): 594–622. дои : 10.1080/10635150490445706 . ПМИД 15371249 . Архивировано из оригинала 14 июля 2021 года . Проверено 21 июля 2020 г.
- ^ Риттмайер, Эрик Н.; Эллисон, Аллен; Грюндлер, Майкл К.; Томпсон, Деррик К.; Остин, Кристофер К. (2012). «Экологическая эволюция гильдии и открытие самого маленького позвоночного в мире» . ПЛОС ОДИН . 7 (1): e29797. Бибкод : 2012PLoSO...729797R . дои : 10.1371/journal.pone.0029797 . ПМК 3256195 . ПМИД 22253785 .
- ^ Jump up to: а б Нгуен, Брент; Каваньяро, Джон (июль 2012 г.). «Факты об амфибиях» . АмфибияВеб. Архивировано из оригинала 23 января 2014 года . Проверено 9 ноября 2012 г.
- ^ Прайс, Л.И. (1948). «Лабиринтодонтная амфибия из формации Педра-де-Фого, штат Мараньян». Бюллетень . 24 . Министерство сельского хозяйства, Национальный департамент добычи полезных ископаемых, Отдел геологии и минералогии: 7–32.
- ^ Стеббинс и Коэн 1995 , стр. 24–25.
- ^ Каннателла, Дэвид; Грейбил, Анна (2008). «Bufonidae, настоящие жабы» . Веб-проект «Древо жизни» . Архивировано из оригинала 14 июня 2020 года . Проверено 1 декабря 2012 г.
- ^ «Забавные факты о лягушках» . Американский музей естественной истории. 12 января 2010. Архивировано из оригинала 24 февраля 2013 года . Проверено 29 августа 2012 г.
- ^ Челленджер, Дэвид (12 января 2012 г.). «Самая маленькая лягушка в мире обнаружена в Папуа-Новой Гвинее» . Си-Эн-Эн. Архивировано из оригинала 20 апреля 2012 года . Проверено 29 августа 2012 г.
- ^ Jump up to: а б с д и ж г Арнольд, Николас; Овенден, Денис (2002). Рептилии и амфибии Британии и Европы . Издательство Харпер Коллинз. стр. 13–18. ISBN 978-0-00-219318-4 .
- ^ Фаивович Дж.; Хаддад, ЦФБ; Гарсия, PCA; Фрост, ДР; Кэмпбелл, Дж.А.; Уиллер, WC (2005). «Систематический обзор семейства лягушек Hylidae с особым упором на Hylinae: филогенетический анализ и пересмотр». Бюллетень Американского музея естественной истории . 294 : 1–240. CiteSeerX 10.1.1.470.2967 . doi : 10.1206/0003-0090(2005)294[0001:SROTFF]2.0.CO;2 . S2CID 83925199 .
- ^ Jump up to: а б Форд, Лос-Анджелес; Каннателла, округ Колумбия (1993). «Основные клады лягушек». Герпетологические монографии . 7 : 94–117. дои : 10.2307/1466954 . JSTOR 1466954 .
- ^ Сан-Мауро, Диего; Винс, Майкл; Альковендас, ВМС; Зардоя, Рафаэль; Мейер, Аксель (2005). «Первоначальное разнообразие современных амфибий предшествовало распаду Пангеи» . Американский натуралист . 165 (5): 590–599. дои : 10.1086/429523 . ПМИД 15795855 . S2CID 17021360 . Архивировано из оригинала 7 февраля . Получено 10 , ноября
- ^ Баум, Дэвид (2008). «Эволюция признаков на филогенетическом дереве: родство, сходство и миф об эволюционном развитии» . Природное образование. Архивировано из оригинала 8 марта 2014 года . Проверено 1 декабря 2012 г.
- ^ Спарребум, Макс (7 февраля 2000 г.). « Андриас Давидианус Китайская гигантская саламандра» . АмфибияВеб. Архивировано из оригинала 26 декабря 2012 года . Проверено 1 декабря 2012 г.
- ^ Уэйк, Дэвид Б. (8 ноября 2000 г.). « Торий пеннатулус » . АмфибияВеб. Архивировано из оригинала 15 мая 2012 года . Проверено 25 августа 2012 г.
- ^ Элмер, КР; Бонетт, РМ; Уэйк, ДБ; Лохид, Южная Каролина (4 марта 2013 г.). «Раннемиоценовое происхождение и загадочная диверсификация южноамериканских саламандр» . Эволюционная биология BMC . 13 (1): 59. Бибкод : 2013BMCEE..13...59E . дои : 10.1186/1471-2148-13-59 . ПМК 3602097 . ПМИД 23497060 .
- ^ Ларсон, А.; Диммик, В. (1993). «Филогенетические взаимоотношения семейств саламандр: анализ соответствия морфологических и молекулярных признаков». Герпетологические монографии . 7 (7): 77–93. дои : 10.2307/1466953 . JSTOR 1466953 .
- ^ Дорит, Уокер и Барнс 1991 , с. 852.
- ^ Привет, Хизер (2003). «Криптобранхиды» . Сеть разнообразия животных . Мичиганский университет. Архивировано из оригинала 21 октября 2012 года . Проверено 25 августа 2012 г.
- ^ Jump up to: а б Маясич, Дж.; Гранмезон, Д.; Филлипс, К. (1 июня 2003 г.). «Отчет об оценке статуса Восточного мага ада» (PDF) . Служба охраны рыбы и дикой природы США. Архивировано (PDF) оригинала 4 сентября 2012 г. Проверено 25 августа 2012 г.
- ^ Jump up to: а б с Уэйк, Дэвид Б. «Каудата» . Британская энциклопедия . Архивировано из оригинала 16 мая 2013 года . Проверено 25 августа 2012 г.
- ^ Коггер, Х.Г. (1998). Цвайфель, Р.Г. (ред.). Энциклопедия рептилий и амфибий . Академическая пресса. стр. 69–70. ISBN 978-0-12-178560-4 .
- ^ Стеббинс и Коэн 1995 , с. 4.
- ^ Дорит, Уокер и Барнс 1991 , с. 858.
- ^ Дуэллман, Уильям Э. «Гимнофиона» . Британская энциклопедия . Архивировано из оригинала 14 января 2012 года . Проверено 30 сентября 2012 г.
- ^ Зильберберг, Луиза; Уэйк, Марвали Х. (1990). «Строение чешуи Dermophis и Microcaecilia (Amphibia: Gymnophiona) и сравнение с кожными оссификациями других позвоночных» . Журнал морфологии . 206 (1): 25–43. дои : 10.1002/jmor.1052060104 . ПМИД 29865751 . S2CID 46929507 . Архивировано из оригинала 16 декабря 2017 года . Проверено 10 ноября 2016 г.
- ^ Институт биоразнообразия Онтарио; Хеберт, Пол Д.Н. (12 октября 2008 г.). «Морфология и размножение амфибий» . Энциклопедия Земли . Архивировано из оригинала 1 февраля 2008 года . Проверено 15 августа 2012 г.
- ^ Стеббинс и Коэн 1995 , стр. 10–11.
- ^ Спирмен, РИК (1973). Покровы: Учебник биологии кожи . Издательство Кембриджского университета. п. 81 . ISBN 978-0-521-20048-6 .
Цвет кожи земноводных.
- ^ Jump up to: а б с д Дорит, Уокер и Барнс 1991 , с. 846.
- ^ Jump up to: а б с Стеббинс и Коэн 1995 , стр. 26–36.
- ^ Jump up to: а б Бенески, Джон Т. младший (сентябрь 1989 г.). «Адаптационное значение аутотомии хвоста у саламандры Ensatina ». Журнал герпетологии . 23 (3): 322–324. дои : 10.2307/1564465 . JSTOR 1564465 .
- ^ Дорит, Уокер и Барнс 1991 , с. 306.
- ^ Гонсалес А.; Лопес, Дж. М.; Морона, Р.; Морона, Н. (2020). «Организация центральной нервной системы амфибий». В Хассе, Дж. Х. (ред.). Эволюционная нейронаука . Эльзевир Наука. п. 127. ИСБН 978-0-12-820584-6 .
- ^ Стеббинс и Коэн 1995 , с. 100.
- ^ Естественная история земноводных
- ^ Стеббинс и Коэн 1995 , с. 69.
- ^ Jump up to: а б с Дуэллман, Уильям Э.; Цуг, Джордж Р. (2012). «Амфибия» . Британская энциклопедия . Архивировано из оригинала 8 марта 2012 года . Проверено 27 марта 2012 г.
- ^ Jump up to: а б с Дорит, Уокер и Барнс 1991 , с. 847.
- ^ Стеббинс и Коэн 1995 , с. 66.
- ^ Jump up to: а б Дорит, Уокер и Барнс 1991 , с. 849.
- ^ Уракабе, Сигэхару; Шираи, Дайроку; Юаса, Сигэказу; Кимура, Генджиро; Орита, Ёсимаса; Абэ, Хироши (1976). «Сравнительное изучение влияния различных диуретиков на проницаемость мочевого пузыря жабы». Сравнительная биохимия и физиология. Часть C: Сравнительная фармакология . 53 (2): 115–119. дои : 10.1016/0306-4492(76)90063-0 . ПМИД 5237 .
- ^ Сибата, Юки; Катаяма, Идзуми; Накакура, Такаши; Огуши, Юджи; Окада, Рэйко; Танака, Сигэясу; Сузуки, Масакадзу (2015). «Молекулярная и клеточная характеристика аквапорина типа мочевого пузыря у Xenopus laevis». Общая и сравнительная эндокринология . 222 : 11–19. дои : 10.1016/j.ygcen.2014.09.001 . ПМИД 25220852 .
- ^ Jump up to: а б Лори Дж. Витт; Джанали П. Колдуэлл (25 марта 2013 г.). Герпетология: вводная биология амфибий и рептилий . Академический. п. 184. ИСБН 978-0-12-386920-3 .
- ^ Федер, Мартин Э.; Бурггрен, Уоррен В. (15 октября 1992 г.). Экологическая физиология земноводных . Издательство Чикагского университета. ISBN 978-0-226-23944-6 .
- ^ Брейнерд, Э.Л. (1999). «Новые взгляды на эволюцию механизмов вентиляции легких у позвоночных». Экспериментальная биология онлайн . 4 (2): 1–28. Бибкод : 1999EvBO....4b...1B . дои : 10.1007/s00898-999-0002-1 . S2CID 35368264 .
- ^ Хопкинс Гарет Р.; Броди Эдмунд Д. младший (2015). «Распространение земноводных в солончаковых средах обитания: обзор и эволюционная перспектива». Герпетологические монографии . 29 (1): 1–27. doi : 10.1655/HERPMONOGRAPHHS-D-14-00006 . S2CID 83659304 .
- ^ Натчев, Николай; Цанков, Николай; Джеме, Ричард (2011). «Инвазия зеленых лягушек в Черное море: экология среды обитания популяции комплекса Pelophylax esculentus (Anura, Amphibia) в районе лагуны Шабленска Тузла в Болгарии» (PDF) . Герпетологические заметки . 4 : 347–351. Архивировано из оригинала (PDF) 24 сентября 2015 года . Проверено 17 августа 2012 г.
- ^ Хоган, К. Майкл (31 июля 2010 г.). «Абиотический фактор» . Энциклопедия Земли . Национальный совет по науке и окружающей среде. Архивировано из оригинала 8 июня 2013 года . Проверено 30 сентября 2012 г.
- ^ Стеббинс и Коэн 1995 , стр. 140–141.
- ^ Jump up to: а б Дуэллман, Виллия Э.; Труб, Линда (1994). Биология амфибий . Джу Пресс. стр. 77–79. ISBN 978-0-8018-4780-6 . Архивировано из оригинала 3 февраля 2021 года . Проверено 15 октября 2020 г.
- ^ Jump up to: а б Стеббинс и Коэн 1995 , стр. 154–162.
- ^ Адамс, Майкл Дж.; Перл, Кристофер А. (2005). « Аскаф истинный » . АмфибияВеб. Архивировано из оригинала 4 ноября 2012 года . Проверено 23 ноября 2012 г.
- ^ Романо, Антонио; Бруни, Джакомо (2011). «Поведение ухаживания, брачный сезон и сексуальное вмешательство самцов Salamandrina perspicillata » . Земноводные-рептилии . 32 (1): 63–76. дои : 10.1163/017353710X541878 .
- ^ Адамс, Эрика М.; Джонс, АГ; Арнольд, SJ (2005). «Множественное отцовство в естественной популяции саламандры с длительным хранением спермы» . Молекулярная экология . 14 (6): 1803–1810. Бибкод : 2005MolEc..14.1803A . дои : 10.1111/j.1365-294X.2005.02539.x . ПМИД 15836651 . S2CID 18899524 . Архивировано из оригинала 4 октября 2018 года . Проверено 10 ноября 2016 г.
- ^ Богарт Дж. П., Би К., Фу Дж., Нобл Д. В., Недзвецки Дж. Однополые саламандры (род Ambystoma) представляют новый репродуктивный способ для эукариот. Геном. 2007 февраль;50(2):119-36. дои: 10.1139/g06-152. ПМИД 17546077
- ^ Jump up to: а б Гиббс Х.Л., Дентон Р.Д. Загадочный секс? Оценки геномного обмена у однополых кротовых саламандр (Ambystoma sp.). Мол Экол. 2016 июня;25(12):2805-15. дои: 10.1111/mec.13662. Epub, 2016 г., 21 мая. PMID 27100619.
- ^ Кикуяма, Сакаэ; Кавамура, Косуке; Танака, Сигэясу; Ямамото, Какутоши (1993). «Аспекты метаморфоза амфибий: Гормональный контроль» . Международный обзор цитологии: обзор клеточной биологии . Академическая пресса. стр. 105–126. ISBN 978-0-12-364548-7 . Архивировано из оригинала 14 апреля 2021 года . Проверено 15 октября 2020 г.
- ^ Ньюман, Роберт А. (1992). «Адаптивная пластичность в метаморфозе амфибий». Бионаука . 42 (9): 671–678. дои : 10.2307/1312173 . JSTOR 1312173 .
- ^ Гилберт, Перри В. (1942). «Наблюдения за яйцами Ambystoma maculatum с особым упором на зеленые водоросли, обнаруженные в оболочках яиц». Экология . 23 (2): 215–227. Бибкод : 1942Экол...23..215Г . дои : 10.2307/1931088 . JSTOR 1931088 .
- ^ Уолдман, Брюс; Райан, Майкл Дж. (1983). «Термические преимущества совместного отложения яичной массы у лесных лягушек ( Rana sylvatica )» . Журнал герпетологии . 17 (1): 70–72. дои : 10.2307/1563783 . JSTOR 1563783 . Архивировано из оригинала 14 июля 2021 года . Проверено 10 ноября 2016 г.
- ^ Олдер, Крейг; Труб (2002). «Амфибии». В Холлидее, Тим; Адлер, Крейг (ред.). Энциклопедия рептилий и амфибий Firefly . Книги Светлячка. п. 17. ISBN 978-1-55297-613-5 .
- ^ Мешака, Уолтер Э. младший « Eleutherodactylus planirostris » . АмфибияВеб. Архивировано из оригинала 9 июня 2013 года . Проверено 12 декабря 2012 г.
- ^ Далгетти, Лаура; Кеннеди, Малкольм В. (2010). «Строительство дома из пенопласта: архитектура пенопластового гнезда тунгарской лягушки и трехэтапный процесс строительства» . Письма по биологии . 6 (3): 293–296. дои : 10.1098/rsbl.2009.0934 . ПМЦ 2880057 . ПМИД 20106853 .
- ^ «Белки пенных гнезд лягушек» . Школа естественных наук Университета Глазго. Архивировано из оригинала 3 июня 2013 года . Проверено 24 августа 2012 г.
- ^ Стеббинс и Коэн 1995 , стр. 6–9.
- ^ Витт, Лори Дж.; Колдуэлл, Джанали П. (2013). Герпетология: вводная биология амфибий и рептилий . Академическая пресса. п. 42. ИСБН 978-0123869197 .
- ^ Янзен, Питер (10 мая 2005 г.). « Наннофрис цейлоненсис » . АмфибияВеб. Архивировано из оригинала 7 июня 2022 года . Проверено 20 июля 2012 г.
- ^ Дуэллман, МЫ; Цуг Г.Р. «Анура: от головастика до взрослой особи» . Британская энциклопедия . Архивировано из оригинала 2 ноября 2012 года . Проверено 13 июля 2012 г.
- ^ Сравнительная анатомия и биология развития позвоночных (книга по зоологии)
- ^ Биологические системы позвоночных, Том. 1 Функциональная морфология дыхательных систем позвоночных
- ^ Биология земноводных
- ^ Стеббинс и Коэн 1995 , стр. 179–181.
- ^ Вентури, Себастьяно (2011). «Эволюционное значение йода» . Современная химическая биология . 5 (3): 155–162. дои : 10.2174/187231311796765012 . ISSN 1872-3136 . Архивировано из оригинала 9 декабря 2019 года . Проверено 10 ноября 2016 г.
- ^ Вентури, Себастьяно (2014). «Йод, ПНЖК и йодолипиды в здоровье и болезнях: эволюционная перспектива». Эволюция человека . 29 (1–3): 185–205. ISSN 0393-9375 .
- ^ Jump up to: а б с Дуэллман, Уильям Э.; Цуг, Джордж Р. (2012). «Анура» . Британская энциклопедия . Архивировано из оригинала 2 ноября 2012 года . Проверено 26 марта 2012 г.
- ^ Крамп, Марта Л. (1986). «Каннибализм молодых головастиков: еще одна опасность метаморфоза». Копейя . 1986 (4): 1007–1009. дои : 10.2307/1445301 . JSTOR 1445301 .
- ^ Jump up to: а б Уэйк, Дэвид Б. (2012). «Каудата» . Британская энциклопедия . Архивировано из оригинала 16 мая 2013 года . Проверено 26 марта 2012 г.
- ^ Валентин, Барри Д.; Деннис, Дэвид М. (1964). «Сравнение системы жаберных дуг и плавников трех родов личинок саламандр: Rhyacotriton , Gyrinophilus и Ambystoma ». Копейя . 1964 (1): 196–201. дои : 10.2307/1440850 . JSTOR 1440850 .
- ^ Шаффер, Х. Брэдли (2005). « Амбистома изящная » . АмфибияВеб. Архивировано из оригинала 26 декабря 2012 года . Проверено 21 ноября 2012 г.
- ^ Jump up to: а б Дуэллман, Уильям Э.; Труб, Линда (1994). Биология амфибий . Джу Пресс. стр. 191–192. ISBN 978-0-8018-4780-6 . Архивировано из оригинала 26 января 2021 года . Проверено 15 октября 2020 г.
- ^ Киёнага, Робин Р. «Метаморфоза против неотении (педоморфоза) у саламандр (Caudata)» . Архивировано из оригинала 20 августа 2012 года . Проверено 21 ноября 2012 г.
- ^ Стеббинс и Коэн 1995 , с. 196.
- ^ Шаффер, Х. Брэдли; Остин, CC; Хьюи, РБ (1991). «Последствия метаморфозы на саламандры ( амбистомы двигательную активность )» (PDF) . Физиологическая зоология . 64 (1): 212–231. дои : 10.1086/physzool.64.1.30158520 . JSTOR 30158520 . S2CID 87191067 . Архивировано из оригинала (PDF) 10 февраля 2020 г.
- ^ Брекенридж, WR; Натанаэль, С.; Перейра, Л. (1987). «Некоторые аспекты биологии и развития Ichthyophis Glutinosus ». Журнал зоологии . 211 (3): 437–449. дои : 10.1111/jzo.1987.211.3.437 .
- ^ Уэйк, Марвали Х. (1977). «Фетальное поддержание плода и его эволюционное значение у земноводных: Gymnophiona» . Журнал герпетологии . 11 (4): 379–386. дои : 10.2307/1562719 . JSTOR 1562719 . Архивировано из оригинала 4 октября 2018 года . Проверено 10 ноября 2016 г.
- ^ Дуэллман, Уильям Э. (2012). «Гимнофиона» . Британская энциклопедия . Архивировано из оригинала 14 января 2012 года . Проверено 26 марта 2012 г.
- ^ Метаморфоза: проблема биологии развития
- ^ Уилкинсон, Марк; Купфер, Александр; Маркес-Порто, Рафаэль; Джеффкинс, Хилари; Антониацци, Марта М.; Джаред, Карлос (2008). «Сто миллионов лет питания кожей? Расширенная родительская забота о неотропических червягах (Amphibia: Gymnophiona)» . Письма по биологии . 4 (4): 358–361. дои : 10.1098/rsbl.2008.0217 . ПМК 2610157 . ПМИД 18547909 .
- ^ Крамп, Марта Л. (1996). «Родительская забота среди амфибий». Родительская забота: эволюция, механизмы и адаптивное значение . Достижения в изучении поведения. Том. 25. С. 109–144. дои : 10.1016/S0065-3454(08)60331-9 . ISBN 978-0-12-004525-9 .
- ^ Браун, Дж.Л.; Моралес, В.; Саммерс, К. (2010). «Ключевая экологическая черта привела к развитию заботы о двух родителях и моногамии у амфибий» . Американский натуралист . 175 (4): 436–446. дои : 10.1086/650727 . ПМИД 20180700 . S2CID 20270737 . Архивировано из оригинала 29 июля 2020 года . Проверено 10 ноября 2016 г.
- ^ Дорит, Уокер и Барнс 1991 , стр. 853–854.
- ^ Фандиньо, Мария Клаудия; Люддеке, Хорст; Амескита, Адольфо (1997). «Вокализация и транспортировка личинок самца Colostethus subpunctatus (Anura: Dendrobatidae)». Земноводные-рептилии . 18 (1): 39–48. дои : 10.1163/156853897X00297 .
- ^ ван дер Мейден, Ари (18 января 2010 г.). « Алитес акушеры » . АмфибияВеб. Архивировано из оригинала 25 декабря 2012 года . Проверено 29 ноября 2012 г.
- ^ Семейн, Э. (2002). « Реобатрах силус » . Сеть разнообразия животных . Зоологический музей Мичиганского университета. Архивировано из оригинала 6 июля 2012 года . Проверено 5 августа 2012 г.
- ^ Группа специалистов МСОП по амфибиям SSC. (2023). « Асса Дарлингтони » . Красный список исчезающих видов МСОП . 2023 : e.T211150139A78440087. doi : 10.2305/IUCN.UK.2023-1.RLTS.T211150139A78440087.en .
- ^ Группа специалистов МСОП по амфибиям SSC. (2023). « Пипа-пипа » . Красный список исчезающих видов МСОП . 2023 : e.T58163A85900348. doi : 10.2305/IUCN.UK.2023-1.RLTS.T58163A85900348.en .
- ^ ван Вейнгаарден, Рене; Боланьос, Федерико (1992). «Родительский уход у Dendrobates granuliferus (Anura: Dendrobatidae) с описанием головастика» . Журнал герпетологии . 26 (1): 102–105. дои : 10.2307/1565037 . JSTOR 1565037 . Архивировано из оригинала 4 октября 2018 года . Проверено 10 ноября 2016 г.
- ^ Jump up to: а б Уно, Ёсинобу (2021). «Вывод об эволюции геномов и хромосом позвоночных на основе геномного и цитогенетического анализа с использованием амфибий» . Хромосомная наука . 24 (1–2): 3–12. дои : 10.11352/scr.24.3 . Архивировано из оригинала 6 сентября 2021 года . Проверено 6 сентября 2021 г.
- ^ Новошилов, Сергей; Шлойсниг, Зигфрид; Фей, Цзи-Фэн; Даль, Андреас; Панг, Энди У.К.; Пиппель, Мартин; Винклер, Силке; Хасти, Алекс Р.; Янг, Джордж; Росцито, Джулиана Г.; Сокол, Франциско (2018). «Геном аксолотля и эволюция ключевых регуляторов тканевого образования» . Природа . 554 (7690): 50–55. Бибкод : 2018Natur.554...50N . дои : 10.1038/nature25458 . hdl : 21.11116/0000-0003-F659-4 . ISSN 0028-0836 . ПМИД 29364872 . S2CID 256770603 .
- ^ Габбард, Джесси (2000). « Сирена промежуточная : Малая Сирена» . Сеть разнообразия животных . Зоологический музей Мичиганского университета. Архивировано из оригинала 2 января 2010 года . Проверено 11 августа 2012 г.
- ^ Да Силва, HR; Де Бритто-Перейра, MC (2006). «Сколько фруктов едят фруктовоядные лягушки? Исследование рациона Xenohyla truncata (Lissamphibia: Anura: Hylidae)» . Журнал зоологии . 270 (4): 692–698. дои : 10.1111/j.1469-7998.2006.00192.x . Архивировано из оригинала 23 июня 2019 года . Проверено 10 ноября 2016 г.
- ^ Труб, Линда; Ганс, Карл (1983). «Специализация питания мексиканской роющей жабы Rhinophrynus dorsalis (Anura: Rhinophrynidae)» (PDF) . Журнал зоологии . 199 (2): 189–208. дои : 10.1111/j.1469-7998.1983.tb02090.x . hdl : 2027.42/74489 . Архивировано (PDF) из оригинала 27 июля 2020 г. Проверено 27 августа 2019 г.
- ^ Гамильтон, У.Дж. младший (1948). «Пища и пищевое поведение зеленой лягушки Rana clamitans Latreille в штате Нью-Йорк». Копейя . 1948 (3). Американское общество ихтиологов и герпетологов: 203–207. дои : 10.2307/1438455 . JSTOR 1438455 .
- ^ Стеббинс и Коэн 1995 , с. 56.
- ^ Стеббинс и Коэн 1995 , стр. 57–58.
- ^ Рэдклифф, Чарльз В.; Чизар, Дэвид; Эстеп, Карен; Мерфи, Джеймс Б.; Смит, Хобарт М. (1986). «Наблюдения за приманкой педалей и движениями педалей у лягушек-лептодактилид». Журнал герпетологии . 20 (3): 300–306. дои : 10.2307/1564496 . JSTOR 1564496 .
- ^ Тофт, Кэтрин А. (1981). «Экология питания панамских бесхвостых бесхвостых: особенности рациона и режима кормления» . Журнал герпетологии . 15 (2): 139–144. дои : 10.2307/1563372 . JSTOR 1563372 . Архивировано из оригинала 30 июля 2020 года . Проверено 10 ноября 2016 г.
- ^ Бемис, МЫ; Швенк, К.; Уэйк, Миннесота (1983). «Морфология и функция пищевого аппарата Dermophis mexicanus (Amphibia: Gymnophiona)» . Зоологический журнал Линнеевского общества . 77 (1): 75–96. дои : 10.1111/j.1096-3642.1983.tb01722.x . Архивировано из оригинала 30 июля 2020 года . Проверено 10 ноября 2016 г.
- ^ Стеббинс и Коэн 1995 , стр. 181–185.
- ^ Jump up to: а б Стеббинс и Коэн 1995 , стр. 76–77.
- ^ Салливан, Брайан К. (1992). «Половой отбор и призывное поведение американской жабы ( Bufo americanus )». Копейя . 1992 (1): 1–7. дои : 10.2307/1446530 . JSTOR 1446530 .
- ^ Толедо, LF; Хаддад, CFB (2007). «Глава 4» (PDF) . Когда лягушки кричат! Обзор защитных вокализаций ануранов (Диссертация). Институт бионаук, Сан-Паулу. Архивировано (PDF) оригинала 4 сентября 2012 г. Проверено 13 августа 2012 г.
- ^ Джонсон, Стив А. (2010). «Кубинская древесная лягушка ( Osteopilus septentrionalis ) во Флориде» . ЭДИС . Университет Флориды. Архивировано из оригинала 20 августа 2012 года . Проверено 13 августа 2012 г.
- ^ Шайн, Ричард (1979). «Половой отбор и половой диморфизм у амфибий». Копейя . 1979 (2): 297–306. дои : 10.2307/1443418 . JSTOR 1443418 .
- ^ Jump up to: а б с Гергитс, ВФ; Йегер, Р.Г. (1990). «Прикрепление к сайту красноспинной саламандры Plethodon cinereus ». Журнал герпетологии . 24 (1): 91–93. дои : 10.2307/1564297 . JSTOR 1564297 .
- ^ Каспер, Гэри С. « Plethodon cinereus » . АмфибияВеб. Архивировано из оригинала 9 июня 2013 года . Проверено 25 сентября 2012 г.
- ^ Уэллс, К.Д. (1977). «Территориальность и успех спаривания самцов зеленой лягушки ( Rana clamitans )». Экология . 58 (4): 750–762. Бибкод : 1977Ecol...58..750W . дои : 10.2307/1936211 . JSTOR 1936211 .
- ^ Jump up to: а б Бартальмус, GT; Зелински В.Дж. (1988). « Слизь кожи Xenopus вызывает дискинезию полости рта, которая способствует бегству от змей». Фармакология Биохимия и поведение . 30 (4): 957–959. дои : 10.1016/0091-3057(88)90126-8 . ПМИД 3227042 . S2CID 25434883 .
- ^ Крайон, Джон Дж. «Xenopus laevis» . АмфибияВеб. Архивировано из оригинала 9 октября 2014 года . Проверено 8 октября 2012 г.
- ^ Муди, GEE (1978). «Наблюдения за историей жизни червяг Typhlonectes compressicaudus (Dumeril и Bibron) в бассейне Амазонки» . Канадский журнал зоологии . 56 (4): 1005–1008. дои : 10.1139/z78-141 . Архивировано из оригинала 5 июля 2020 года . Проверено 10 ноября 2016 г.
- ^ Броди, Эдмунд Д. младший (1968). «Исследования кожного токсина взрослого грубокожего тритона Taricha granulosa ». Копейя . 1968 (2): 307–313. дои : 10.2307/1441757 . JSTOR 1441757 .
- ^ Jump up to: а б Ханифин, Чарльз Т.; Ёцу-Ямасита, Мари; Ясумото, Такеши; Броди, Эдмунд Д.; Броди, Эдмунд Д. младший (1999). «Токсичность опасной добычи: вариации уровней тетродотоксина внутри и среди популяций тритона Taricha granulosa » . Журнал химической экологии . 25 (9): 2161–2175. дои : 10.1023/А:1021049125805 . S2CID 543221 . Архивировано из оригинала 5 июля 2020 года . Проверено 10 ноября 2016 г.
- ^ Геффни, Шана Л.; Фудзимото, Эстер; Броди, Эдмунд Д.; Броди, Эдмунд Д. младший; Рубен, Питер К. (2005). «Эволюционная диверсификация устойчивых к ТТХ натриевых каналов во взаимодействии хищник-жертва». Природа . 434 (7034): 759–763. Бибкод : 2005Natur.434..759G . дои : 10.1038/nature03444 . ПМИД 15815629 . S2CID 4426708 .
- ^ Стеббинс и Коэн 1995 , с. 110.
- ^ Паточка, Иржи; Вульф, Кроуфф; Паломеке, Мария Виктория (1999). «Ядовитые лягушки-дротики и их токсины» . Информационный бюллетень АСА . 5 (75). ISSN 1057-9419 . Архивировано из оригинала 23 марта 2014 года . Проверено 29 января 2013 г.
- ^ Дарст, Кэтрин Р.; Каммингс, Молли Э. (9 марта 2006 г.). «Обучение хищников способствует имитации менее токсичной модели ядовитых лягушек» . Природа . 440 (7081): 208–211. Бибкод : 2006Natur.440..208D . дои : 10.1038/nature04297 . ПМИД 16525472 .
- ^ Броди, Эдмунд Д. младший; Новак, Роберт Т.; Харви, Уильям Р. (1979). «Выделения против хищников и поведение избранных саламандр против землероек». Копейя . 1979 (2): 270–274. дои : 10.2307/1443413 . JSTOR 1443413 .
- ^ Броди, ЭД-младший (1978). «Кусание и вокализация как механизмы борьбы с хищниками у наземных саламандр». Копейя . 1978 (1): 127–129. дои : 10.2307/1443832 . JSTOR 1443832 .
- ^ Хлох, А. (2010). Что помнит саламандра после зимы? (PDF) . Венский университет. Факультет наук о жизни. Архивировано (PDF) из оригинала 4 октября 2018 г. Проверено 15 ноября 2015 г.
- ^ Ламберт, Хелен; Элвин, Энджи; Д'Круз, Нил (1 февраля 2022 г.). «Лягушка в колодце: обзор научной литературы, подтверждающей разумность земноводных» . Прикладная наука о поведении животных . 247 : 105559. doi : 10.1016/j.applanim.2022.105559 . ISSN 0168-1591 . Проверено 26 мая 2023 г.
- ^ Стэнчер, Г.; Ругани, Р.; Реголин, Л.; Валлортигара, Г. (2015). «Численная дискриминация лягушек (Bombina orientalis)» . Познание животных . 18 (1): 219–229. дои : 10.1007/s10071-014-0791-7 . ПМИД 25108417 . S2CID 16499583 . Архивировано из оригинала 14 июля 2021 года . Проверено 10 ноября 2016 г.
- ^ МакКаллум, ML (2007). «Упадок или вымирание земноводных? Текущее сокращение затмевает фоновые темпы вымирания». Журнал герпетологии . 41 (3): 483–491. doi : 10.1670/0022-1511(2007)41[483:ADOECD]2.0.CO;2 . S2CID 30162903 .
- ^ «Что значит быть человеком?» . Смитсоновский национальный музей естественной истории. Архивировано из оригинала 22 ноября 2013 года . Проверено 19 ноября 2013 г.
- ^ Хукстра, Дж. М.; Мольнар, Дж.Л.; Дженнингс, М.; Ревенга, К.; Сполдинг, доктор медицины; Баучер, ТМ; Робертсон, Дж. К.; Хейбель, Ти Джей; Эллисон, К. (2010). «Число видов амфибий, находящихся под угрозой исчезновения, по пресноводным экорегионам» . Атлас глобальной охраны природы: изменения, проблемы и возможности изменить ситуацию . Охрана природы. Архивировано из оригинала 27 октября 2012 года . Проверено 5 сентября 2012 г.
- ^ «Группа специалистов по амфибиям» . Группа специалистов МСОП по амфибиям SSC. Архивировано из оригинала 22 февраля 2011 года . Проверено 30 марта 2012 г.
- ^ Дженнингс, В. Брайан; Брэдфорд, Дэвид Ф.; Джонсон, Дейл Ф. (1992). «Зависимость подвязочной змеи Thamnophis elegans от амфибий в Сьерра-Неваде в Калифорнии» . Журнал герпетологии . 26 (4): 503–505. дои : 10.2307/1565132 . JSTOR 1565132 . Архивировано из оригинала 4 октября 2018 года . Проверено 10 ноября 2016 г.
- ^ Стеббинс и Коэн 1995 , с. 249.
- ^ Jump up to: а б Холл, младший; Генри, ФПП (1992). «Обзор: Оценка воздействия пестицидов на земноводных и рептилий: состояние и потребности». Герпетологический журнал . 2 : 65–71.
- ^ Jump up to: а б «План действий по сохранению амфибий» . МСОП . Архивировано из оригинала 27 апреля 2012 года . Проверено 30 марта 2012 г.
- ^ «Панамский проект спасения и сохранения амфибий» . Ковчег-амфибия. Архивировано из оригинала 14 июня 2010 года . Проверено 30 марта 2012 г.
- ^ Режье, Генри А.; Баскервиль, Гордон, Л. (1996). «Проблемы устойчивого развития для менеджеров ресурсов» . Устойчивое восстановление региональных экосистем, деградированных в результате эксплуататорского освоения . Издательство ДИАНА. стр. 36–38. ISBN 978-0-7881-4699-2 . Архивировано из оригинала 14 апреля 2021 года . Проверено 15 октября 2020 г.
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка )
Цитируемые тексты
- Дорит, РЛ; Уокер, ВФ; Барнс, Р.Д. (1991). Зоология . Издательство Колледжа Сондерса. ISBN 978-0-03-030504-7 .
- Стеббинс, Роберт С .; Коэн, Натан В. (1995). Естественная история амфибий . Издательство Принстонского университета. ISBN 978-0-691-03281-8 .
Дальнейшее чтение
- Кэрролл, Роберт Л. (1988). Палеонтология и эволюция позвоночных . У. Х. Фриман. ISBN 978-0-7167-1822-2 .
- Кэрролл, Роберт Л. (2009). Возникновение амфибий: 365 миллионов лет эволюции . Издательство Университета Джонса Хопкинса. ISBN 978-0-8018-9140-3 .
- Дуэллман, Уильям Э.; Линда Труб (1994). Биология амфибий . Издательство Университета Джонса Хопкинса. ISBN 978-0-8018-4780-6 .
- Дуэллман, Уильям Э., Берг, Барбара (1962), Типовые образцы амфибий и рептилий в Музее естественной истории Канзасского университета.
- Фрост, Даррел Р.; Грант, Таран; Фаивович, Хулиан; Бейн, Рауль Х.; Хаас, Александр; Хаддад, Селио ФБ; Де Са, Рафаэль О.; Ченнинг, Алан; Уилкинсон, Марк; Доннеллан, Стивен С.; Раксуорси, Кристофер Дж.; Кэмпбелл, Джонатан А.; Блотто, Борис Л.; Молер, Пол; Древес, Роберт С.; Нуссбаум, Рональд А.; Линч, Джон Д.; Грин, Дэвид М.; Уиллер, Уорд К. (2006). «Древо жизни земноводных» . Бюллетень Американского музея естественной истории . 297 : 1–291. doi : 10.1206/0003-0090(2006)297[0001:TATOL]2.0.CO;2 . hdl : 2246/5781 . S2CID 86140137 .
- Паундс, Дж. Алан; Бустаманте, Мартин Р.; Колома, Луис А.; Невестка Джейми А.; Фогден, Майкл П.Л.; Фостер, Пру Н.; Ла Марка, Энрике; Мастерс, Карен Л.; Мерино-Витери, Андрес; Пушендорф, Роберт; Рон, Джеймс Р.; Санчес-Азофейфа, Дж. Артуро; И все же Кристофер Дж.; Янг, Брюс Э. (2006). «Широкое вымирание земноводных из-за эпидемических заболеваний, вызванных глобальным потеплением» . Природа . 439 (7073): 161–167. Бибкод : 2006Natur.439..161A . дои : 10.1038/nature04246 . ПМИД 16407945 . S2CID 4430672 .
- Стюарт, Саймон Н.; Шансон, Дженис С.; Кокс, Нил А.; Янг, Брюс Э.; Родригес, Ана С.Л.; Фишман, Дебра Л.; Уоллер, Роберт В. (2004). «Состояние и тенденции сокращения и исчезновения земноводных во всем мире». Наука . 306 (5702): 1783–1786. Бибкод : 2004Sci...306.1783S . CiteSeerX 10.1.1.225.9620 . дои : 10.1126/science.1103538 . ПМИД 15486254 . S2CID 86238651 .
- Стюарт, С.Н.; Хоффманн, М.; Шансон, Дж.С.; Кокс, Северная Каролина; Берридж, Р.Дж.; Рамани, П.; Янг, Б.Е., ред. (2008). Угрожаемые амфибии мира . Опубликовано Lynx Edicions совместно с МСОП (Всемирным союзом охраны природы) , Conservation International и NatureServe . ISBN 978-84-96553-41-5 . Архивировано из оригинала 30 октября 2014 года . Проверено 30 октября 2014 г.
Внешние ссылки
- Земноводные – AnimalSpot.net
- ArchéoZooThèque: рисунки скелетов амфибий : доступны в векторном формате, формате изображений и PDF.
- Группа специалистов по амфибиям
- Ковчег-амфибия
- АмфибияВеб
- Глобальная оценка амфибий. Архивировано 12 апреля 2011 г. в Wayback Machine.
- Вокализации амфибий на архивных звукозаписях, заархивированных 26 мая 2010 г. в Wayback Machine.