Лист



- Апекс
- Средняя вена (первичная вена)
- Вторичная вена.
- Ламина.
- Край листа
- Черешок
- Бутон
- Корень

Внизу: скунсовая капуста, Symplocarpus foetidus (простой лист).
- Апекс
- Первичная вена
- Вторичная вена
- пластинка
- Край листа
- Рачис
Лист Leaves ( мн.ч .: ) — главный придаток стебля сосудистого растения . [ 1 ] обычно располагаются сбоку над землей и специализируются на фотосинтезе . Листья вместе называются листвой , например «осенняя листва». [ 2 ] [ 3 ] в то время как листья, стебель, цветы и плоды вместе образуют систему побегов . [ 4 ] В большинстве листьев первичной фотосинтетической тканью является палисадный мезофилл , расположенный на верхней стороне пластинки или пластинки листа. [ 1 ] но у некоторых видов, включая зрелую листву эвкалипта , [ 5 ] палисадный мезофилл присутствует с обеих сторон, и листья называются изобилатеральными. Большинство листьев уплощены и имеют отчетливые верхнюю ( адаксиальную ) и нижнюю ( абаксиальную ) поверхности, различающиеся по цвету, опушению, количеству устьиц (пор, поглощающих и выводящих газы), количеству и строению эпикутикулярного воска и другим признакам. Листья в основном имеют зеленый цвет из-за присутствия соединения под названием хлорофилл , которое необходимо для фотосинтеза, поскольку оно поглощает световую энергию солнца . Лист с более светлыми или белыми пятнами или краями называется пестрым листом .
Листья могут иметь самую разную форму, размер, текстуру и цвет. Широкие плоские листья со сложным жилкованием цветковых растений известны как мегафиллы , а виды, несущие их, большинство - как широколистные или мегафилловые растения, к которым также относятся акрогимноспермовые и папоротники . У плаунов различного эволюционного происхождения листья простые (только с одной жилкой) и известны как микрофиллы . [ 6 ] Некоторые листья, например чешуи луковицы , не находятся над землей. У многих водных видов листья погружены в воду. Суккулентные растения часто имеют толстые сочные листья, но некоторые листья лишены основной фотосинтетической функции и могут погибнуть по мере созревания, как у некоторых катафиллов и колючек . Более того, некоторые виды листообразных структур, обнаруженных у сосудистых растений, не полностью им гомологичны. Примеры включают уплощенные стебли растений, называемые филлокладами и кладодами , а также уплощенные стебли листьев, называемые филлодиями , которые отличаются от листьев как по своей структуре, так и по происхождению. [ 3 ] [ 7 ] Некоторые структуры несосудистых растений выглядят и функционируют так же, как листья. включают филлиды мхов . и печеночников Примеры
Общие характеристики
[ редактировать ]Листья — важнейшие органы большинства сосудистых растений. [ 8 ] Зеленые растения автотрофны , то есть они не получают пищу от других живых существ, а вместо этого создают себе пищу посредством фотосинтеза . Они улавливают энергию солнечного света и используют ее для производства простых сахаров , таких как глюкоза и сахароза , из углекислого газа и воды. Затем сахара сохраняются в виде крахмала , который далее перерабатывается путем химического синтеза в более сложные органические молекулы, такие как белки или целлюлоза , основной структурный материал в стенках растительных клеток, или метаболизируется посредством клеточного дыхания , чтобы обеспечить химическую энергию для запуска клеточных процессов. Листья всасывают воду из земли в транспирационном потоке через сосудистую проводящую систему, известную как ксилема , и получают углекислый газ из атмосферы путем диффузии через отверстия, называемые устьицами, во внешнем покровном слое листа ( эпидермисе ), при этом листья ориентированы так, чтобы максимизировать их воздействие солнечного света. После синтеза сахара его необходимо транспортировать в места активного роста, например, в побеги растений. и корни . Сосудистые растения транспортируют сахарозу в специальной ткани, называемой флоэмой . Флоэма и ксилема параллельны друг другу, но транспорт веществ обычно происходит в противоположных направлениях. Внутри листа эти сосудистые системы разветвляются (разветвляются), образуя вены, которые снабжают как можно большую часть листа, гарантируя, что клетки, осуществляющие фотосинтез, находятся близко к транспортной системе. [ 9 ]
Обычно листья широкие, плоские и тонкие (сплющенные в дорзивентральном направлении), что увеличивает площадь поверхности, непосредственно подвергающейся воздействию света, и позволяет свету проникать в ткани и достигать хлоропластов , способствуя тем самым фотосинтезу. Они расположены на растении так, чтобы максимально эффективно освещать свои поверхности, не затеняя друг друга, но есть много исключений и сложностей. Например, растения, приспособленные к ветреным условиям, могут иметь свисающие листья, как, например, у многих ив и эвкалиптов . Плоская или ламинарная форма также максимизирует тепловой контакт с окружающим воздухом , способствуя охлаждению. Функционально, помимо осуществления фотосинтеза, лист является основным местом транспирации , обеспечивая энергию, необходимую для вытягивания потока транспирации вверх от корней и гуттации .
Многие хвойные деревья имеют тонкие игольчатые или чешуйчатые листья, что может быть полезно в холодном климате с частым снегом и морозами. [ 10 ] Они интерпретируются как производные от крупнолистных листьев их девонских предков. [ 6 ] Некоторые формы листьев приспособлены модулировать количество поглощаемого ими света, чтобы избежать или смягчить чрезмерное тепло, повреждение ультрафиолетом или высыхание, или пожертвовать эффективностью поглощения света в пользу защиты от травоядных. Для ксерофитов основным ограничением является не световой поток или интенсивность , а засуха. [ 11 ] Некоторые оконные растения, такие как виды Fenestraria и некоторые виды Haworthia , такие как Haworthia tesselata и Haworthia truncata, являются примерами ксерофитов. [ 12 ] и Bulbine mesembryanthemoides . [ 13 ]
Листья также выполняют функцию хранения химической энергии и воды (особенно у суккулентов ) и могут стать специализированными органами, выполняющими другие функции, например, усики гороха и других бобовых, защитные колючки кактусов и ловушки для насекомых у плотоядных растений, таких как непентес и саррацения . [ 14 ] Листья являются основными структурными единицами, из которых построены шишки у голосеменных растений (каждая чешуя шишки представляет собой модифицированный лист мегафилла, известный как спорофилл). [ 6 ] : 408 и из чего построены цветы цветковых растений . [ 6 ] : 445

Внутренняя организация большинства видов листьев эволюционировала таким образом, чтобы максимизировать воздействие на фотосинтетические органеллы , хлоропласты , света и увеличить поглощение углекислого газа , одновременно контролируя потерю воды. Их поверхности водонепроницаемы кутикулой растения , а газообмен между клетками мезофилла и атмосферой контролируется мельчайшими (длина и ширина измеряются десятками мкм) отверстиями, называемыми устьицами , которые открываются или закрываются, регулируя скорость обмена углекислого газа (CO 2 ) . ), кислорода (О 2 ) и водяного пара во внутреннее межклеточное пространство и из него. Открытие устьица контролируется тургорным давлением в паре замыкающих клеток , окружающих устьичное отверстие. В любом квадратном сантиметре листа растения может быть от 1000 до 100 000 устьиц. [ 15 ]


Форма и структура листьев значительно различаются от вида к виду растений, во многом в зависимости от их адаптации к климату и доступному свету, а также от других факторов, таких как выпас животных (например, олени), наличие питательных веществ и экологическая конкуренция со стороны других растений. . Значительные изменения в типе листьев происходят и внутри видов, например, по мере взросления растения; Например, виды эвкалипта в зрелом возрасте обычно имеют изобилатеральные свисающие листья и доминируют над своими соседями; однако такие деревья, как правило, имеют прямостоячие или горизонтальные дорзивентральные листья в качестве сеянцев, когда их рост ограничен доступным светом. [ 16 ] Другие факторы включают необходимость сбалансировать потерю воды при высокой температуре и низкой влажности с необходимостью поглощать углекислый газ из атмосферы. У большинства растений листья также являются основным органом, ответственным за транспирацию и гуттацию (капельки жидкости, образующиеся по краям листьев).
Листья также могут хранить пищу и воду и соответствующим образом модифицируются для выполнения этих функций, например, в листьях суккулентных растений и в чешуях луковиц . Концентрация фотосинтетических структур в листьях требует, чтобы они были богаче белками , минералами и сахарами, чем, скажем, ткани древесного стебля. Соответственно, листья занимают важное место в рационе многих животных .

Соответственно, листья представляют собой тяжелые инвестиции со стороны растений, несущих их, а их сохранение или утилизация являются предметом тщательно продуманных стратегий борьбы с давлением вредителей, сезонными условиями и защитными мерами, такими как рост шипов и образование фитолитов . лигнины , дубильные вещества и яды .
Листопадные растения в регионах с холодным или умеренным климатом обычно сбрасывают листья осенью, тогда как в районах с суровым засушливым сезоном некоторые растения могут сбрасывать листья до окончания засушливого сезона. В любом случае можно ожидать, что сброшенные листья внесут сохраненные питательные вещества в почву, куда они падают.

Напротив, многие другие несезонные растения, такие как пальмы и хвойные деревья, сохраняют листья в течение длительного времени; Вельвичия сохраняет два основных листа на протяжении всей жизни, которая может превышать тысячу лет.
Листовидные органы мохообразных (например, мхов и печеночников ), известные как филлиды , сильно отличаются морфологически от листьев сосудистых растений . В большинстве случаев у них отсутствует сосудистая ткань, они имеют толщину всего в одну клетку и не имеют кутикулы , устьиц или внутренней системы межклеточных пространств. (Филлиды семейства мхов Polytrichaceae являются заметным исключением.) Филлиды мохообразных присутствуют только на гаметофитах , тогда как листья сосудистых растений присутствуют только на спорофитах . В дальнейшем они могут развиваться либо в вегетативные, либо в репродуктивные структуры. [ 14 ]
Простые васкуляризированные листья ( микрофиллы ), такие как листья раннедевонского плауновидного Baragwanathia , сначала развились как энации, продолжения стебля. Настоящие листья или эвфиллы более крупных размеров и с более сложным жилкованием не получили широкого распространения в других группах до девона , к этому времени концентрация углекислого газа в атмосфере значительно снизилась. Это произошло независимо в нескольких отдельных линиях сосудистых растений, у прогимноспермовых , таких как Archaeopteris , у Sphenopsida , папоротников , а позже у голосеменных и покрытосеменных . Эуфиллы также называют макрофиллами или мегафиллами (большие листья). [ 6 ]
Морфология
[ редактировать ]

Структурно законченный лист покрытосеменных растений состоит из черешка (черешка листа), пластинки (листовой пластинки), прилистников (небольших структур, расположенных по обе стороны от основания черешка) и влагалища. Не каждый вид производит листья со всеми этими структурными компонентами. называется ножкой Проксимальный стебель или черешок у папоротников . Пластинка — это расширенная плоская часть листа, содержащая хлоропласты . Оболочка представляет собой структуру, обычно у основания, которая полностью или частично обхватывает стебель над узлом, к которому прикрепляется лист. Листовые влагалища обычно встречаются у Poaceae (травы) и Apiaceae (зонтичные). Между влагалищем и пластинкой может располагаться псевдопетиол — черешковоподобная структура. Псевдопетиолы встречаются у некоторых однодольных растений, включая бананы , пальмы и бамбук . [ 18 ] Прилистники могут быть заметными (например, у бобов и роз ), вскоре опадать или быть неочевидными, как у Moraceae , или вообще отсутствовать, как у Magnoliaceae . Черешок может отсутствовать (петиолатный) или пластинчатая пластинка может быть не пластинчатой (сплюснутой). Черешок механически связывает лист с растением и обеспечивает путь для переноса воды и сахаров в лист и обратно. Пластинка обычно является местом большей части фотосинтеза. Верхний ( адаксиальный ) угол между листом и стеблем известен как пазуха листа. Часто это место расположения бутона . Расположенные там структуры называются «подмышечными».
Внешние характеристики листьев, такие как форма, край, волоски, черешок, а также наличие прилистников и желез, часто важны для идентификации растений на уровне семейства, рода или вида , и ботаники разработали богатую терминологию для описания характеристик листьев. Листья почти всегда имеют детерминированный рост. Они вырастают до определенного рисунка и формы, а затем останавливаются. Другие части растений, такие как стебли или корни, имеют недетерминированный рост и обычно продолжают расти, пока у них есть для этого ресурсы.
Тип листа обычно характерен для вида (мономорфный), хотя некоторые виды образуют более одного типа листа (диморфный или полиморфный ). Самые длинные листья у пальмы рафия , R. regalis , могут достигать 25 м (82 фута) в длину и 3 м (9,8 фута) в ширину. [ 19 ] Терминология, связанная с описанием морфологии листьев, представлена в иллюстрированном виде в Wikibooks .

Листья, расположенные у основания и лежащие на земле, называются распростертыми .
Основные типы листьев
[ редактировать ]
многолетние растения Говорят, что , листья которых сбрасывают ежегодно, имеют лиственные листья, а листья, сохраняющиеся до зимы, — вечнозеленые . Листья, прикрепленные к стеблю черешками (так называемые черешки ), называются черешковыми, а если они прикреплены непосредственно к стеблю без черешка, они называются сидячими. [ 20 ]
- У папоротников есть листья .
- Листья хвойных деревьев обычно имеют игольчатую, шиловидную или чешуйчатую форму, обычно они вечнозеленые, но иногда могут быть листопадными. Обычно они имеют одну жилку.
- Листья цветковых растений (покрытосеменных): стандартная форма включает прилистники , черешок и пластинку .
- Ликофиты имеют микрофиллы .
- Листья- оболочки встречаются у большинства трав и многих других однодольных растений.
- Другие специализированные листья включают листья Непентеса , кувшинчатого растения.
Листья двудольных имеют пластинки с перистым жилкованием (где основные жилки расходятся от одной большой средней жилки и между ними имеются более мелкие соединительные сети). Реже листовые пластинки двудольных могут иметь пальчатое жилкование (несколько крупных жилок, расходящихся от черешка к краям листа). Наконец, у некоторых наблюдается параллельное жилкование. [ 20 ]
Листья однодольных растений в умеренном климате обычно имеют узкие лезвия и обычно параллельное жилкование, сближающееся на кончиках или краях листьев. Некоторые также имеют перистое жилкование. [ 20 ]
Композиция на стебле
[ редактировать ]Расположение листьев на стебле известно как филлотаксис . [ 21 ] В природе встречается большое разнообразие филлотактических узоров:


- Альтернативный
- Один лист, ветка или часть цветка прикрепляется к каждой точке или узлу стебля, а листья в большей или меньшей степени чередуются вдоль стебля.
- Базальный
- Возникает из основания растения.
- Каулин
- Крепится к воздушному стеблю.
- Противоположный
- Два листа, ветки или части цветка прикрепляются к каждой точке или узлу стебля. Прикрепления листьев попарно расположены в каждом узле.
- перекуссировать
- Противоположное расположение, при котором каждая последующая пара повернута на 90° относительно предыдущей.
- Мутовчатая , или мутовчатая
- Три или более листьев, ветвей или частей цветка прикрепляются к каждой точке или узлу стебля. Как и в случае с противоположными листьями, последующие мутовки могут быть перекрещенными, а могут и не быть перекрещенными, повернутыми на половину угла между листьями в мутовке (т. е. последовательные мутовки из трех повернуты на 60°, из четырех мутовок повернуты на 45° и т. д.). Противоположные листья могут выглядеть мутовчатыми на кончике стебля. Псевдовертициллат описывает расположение, которое только кажется закрученным, но на самом деле это не так.
- Розулат
- Листья образуют розетку .
- Строки
- Термин disticous буквально означает два ряда . Листья в этом расположении могут быть очередными или противоположными по расположению. Термин 2-ранговый эквивалентен. термины тристих и тетрастих Иногда встречаются . Например, «листья» (на самом деле микрофиллы ) у большинства видов Selaginella тетрастихозные, но не перекрещивающиеся.
В простейших математических моделях филлотаксиса вершина стебля изображается в виде круга. Каждый новый узел формируется на вершине и поворачивается на постоянный угол относительно предыдущего узла. Этот угол называется углом расхождения . Количество листьев, вырастающих из узла, зависит от вида растения. Когда из каждого узла вырастает по одному листу и стебель держится прямо, листья образуют спираль .
Угол расхождения часто представляют как долю полного оборота вокруг стержня. Доля вращения 1/2 (угол расхождения 180°) приводит к альтернативному расположению, как, например, у Gasteria или веерообразного алоэ Kumara plicatilis . Доли поворота 1/3 (углы расхождения 120°) встречаются у бука и орешника . У дуба и абрикоса — на 2/5, у подсолнечника, тополя и груши — на 3/8, а у ивы и миндаля — на 5/13. [ 22 ] Эти мероприятия носят периодический характер. Знаменатель совершенных дроби вращения указывает количество створок за один период, а числитель указывает количество полных оборотов или круговоротов, за один период. Например:
- 180° (или 1 ⁄ 2 ): два листа в одном круге (чередующиеся листья)
- 120° (или 1 ⁄ 3 ): три листа в одном круге.
- 144° (или 2 ⁄ 5 ): пять листьев в двух кругах.
- 135° (или 3 ⁄ 8 ): восемь листьев в трёх кругах.
Большинство углов расхождения связаны с последовательностью чисел Фибоначчи F n . Эта последовательность начинается с 1, 1, 2, 3, 5, 8, 13; каждое слагаемое представляет собой сумму двух предыдущих. Дроби вращения часто представляют собой частное F n / F n + 2 числа Фибоначчи на два члена, находящихся позже в последовательности. Так обстоит дело с дробями 1/2, 1/3, 2/5, 3/8 и 5/13. Отношение между последовательными числами Фибоначчи стремится к золотому сечению φ = (1 + √5)/2 . Когда круг разделен на две дуги, длины которых находятся в соотношении 1:φ , угол, образованный меньшей дугой, является золотым углом , который равен 1/φ. 2 × 360° ≈ 137,5° . Из-за этого многие углы расхождения составляют примерно 137,5° .
У растений, у которых из каждого узла вырастает пара противоположных листьев, листья образуют двойную спираль. Если узлы не вращаются (доля вращения равна нулю, а угол расхождения 0°), две спирали становятся парой параллельных линий, создавая неоднородное расположение, как у клена или оливковых деревьев. Чаще встречается перекрёстный узор, при котором каждый узел поворачивается на 1/4 (90°), как у травяного базилика . Листья трехстворчатых растений, таких как Nerium oleander, образуют тройную спираль.
Листья некоторых растений не образуют спиралей. У некоторых растений угол расхождения меняется по мере роста растения. [ 23 ] У orixate phyllotaxis, названного в честь Orixa japonica , угол расхождения не является постоянным. Вместо этого он является периодическим и следует последовательности 180°, 90°, 180°, 270°. [ 24 ]
Отделения клинка
[ редактировать ]
Учитывая способ разделения пластинки (листья), можно описать две основные формы листьев. имеет Простой лист цельную пластинку. Однако лист может быть рассечен с образованием долей, но промежутки между долями не доходят до главной жилки. Сложный лист имеет полностью разделенную пластинку, причем каждый листок пластинки разделен по главной или вторичной жилке. Листочки могут иметь черешки и прилистники, эквиваленты черешков и прилистников листьев. Поскольку каждый листочек может выглядеть как простой лист, важно определить, где находится черешок, чтобы определить сложный лист. Сложные листья характерны для некоторых семейств высших растений, например Fabaceae . Средняя жилка сложного листа или вайи , если она есть, называется рахисом .
- Пальчатое соединение
- Все листочки имеют общую точку прикрепления на конце черешка, расходящуюся, как пальцы руки; например, Cannabis (конопля) и Aesculus (каштаны).
- Перистое соединение
- Листочки располагаются по обе стороны от главной оси или рахиса .
- Нечетный перистый
- С брошюрой по терминалу; например, Fraxinus (ясень).
- Даже перистый
- Отсутствие терминальной брошюры; например, Swietenia (красное дерево). Особый тип перистых — двулистный , листья которого состоят только из двух листочков; например, Гименея .
- Двуперистое соединение
- Листья разделены дважды: листочки (технически « подлисточки ») расположены вдоль вторичной оси, которая является одной из нескольких, отходящих от ости. Каждый листочек называется пиннулой . Группа перьев на каждой вторичной жилке образует ушную раковину ; например, Альбиция (шелковое дерево).
- Тройчатые (или тройчатые)
- Перистый лист всего с тремя листочками; например, Trifolium (клевер), Laburnum (лабурнум) и некоторые виды токсикодендронов (например, ядовитый плющ ).
- Поверхностные шпильки
- Перисто-рассеченный до центральной жилки, но листочки не совсем отдельные; например, Polypodium , некоторые Sorbus (белые лучи). У листьев с перистыми жилками центральная жилка известна как средняя жилка .
Характеристики черешка
[ редактировать ]
Листья, имеющие черешок (черешок), называются черешковыми .
Сидячие (эпетиолетные) листья не имеют черешка, а пластинка прикрепляется непосредственно к стеблю. Подчерешковые листья почти черешковые или имеют очень короткий черешок и могут казаться сидячими.
У смыкающихся или нисходящих листьев пластинка частично окружает стебель.
Когда основание листа полностью окружает стебель, листья называются продырявленными , например, у Eupatorium perfoliatum .
У пельтатных листьев черешок прикрепляется к пластинке внутри края пластинки.
У некоторых видов акации , таких как дерево коа ( Acacia koa ), черешки расширены или расширены и функционируют как листовые пластинки; они называются филлодами . На кончике филлодии могут быть или не быть нормальные перистые листья.
Прилистник , присутствующий на листьях многих двудольных растений , представляет собой придаток с каждой стороны у основания черешка, напоминающий небольшой лист. Прилистники могут быть прочными и не опадать (оговоренный лист, например, у роз и фасоли ) или опадать по мере расширения листа, оставляя на ветке рубец от прилистника (эксстипулятивный лист). Расположение, расположение и структура прилистников называется «стипуляцией».
- Свободный, боковой
- Как в Гибискусе .
- сросшийся
- Сросшиеся с основанием черешка, как у Rosa .
- Охреат
- С охряными или чешуйчатыми прилистниками, как у Polygonaceae ; например, ревень .
- Окружение основания черешка
- Интерпетиолярный
- Между черешками двух противоположных листьев, как у Rubiaceae .
- Внутрипетиолярный
- Между черешком и стягивающим стеблем, как у Malpighiaceae .
Вены
[ редактировать ]


Жилки (иногда называемые нервами) представляют собой одну из наиболее заметных особенностей листьев. Жилки в листе представляют собой сосудистую структуру органа, проходящую в лист через черешок и обеспечивающую транспортировку воды и питательных веществ между листом и стеблем, а также играют решающую роль в поддержании водного статуса листа и фотосинтетической способности. Они также играют роль в механической поддержке листа. [ 25 ] [ 26 ] Внутри пластинки листа, хотя у некоторых сосудистых растений имеется только одна жилка, у большинства эта сосудистая сеть обычно делится (разветвляется) по множеству рисунков (жилкование) и образует цилиндрические пучки, обычно лежащие в срединной плоскости мезофилла . между двумя слоями эпидермиса . [ 27 ] Этот образец часто специфичен для таксонов, из которых покрытосеменные имеют два основных типа: параллельные и сетчатые (сетчатые). В целом параллельное жилкование характерно для однодольных, а сетчатое более характерно для эвдикотов и магнолиид (« двудольных »), хотя есть и немало исключений. [ 28 ] [ 27 ] [ 29 ]
Жилку или жилки, входящие в лист от черешка, называют первичными жилками или жилками первого порядка. Вены, отходящие от них, относятся к венам вторичного или второго порядка. Эти первичные и вторичные вены считаются основными венами или венами низшего порядка, хотя некоторые авторы включают третий порядок. [ 30 ] Каждое последующее разветвление последовательно нумеруется, и это жилы более высокого порядка, причем каждое разветвление связано с более узким диаметром жилы. [ 31 ]
В листьях с параллельными жилками первичные жилки идут параллельно и на равном расстоянии друг от друга на большей части длины листа, а затем сходятся или сливаются (анастомозируют) к верхушке. Обычно эти первичные жилки соединяются множеством более мелких второстепенных жилок, но могут заканчиваться очень тонкими жилками в мезофилле. Мелкие жилки более характерны для покрытосеменных растений, которые могут иметь до четырех высших порядков. [ 30 ]
Напротив, листья с сетчатым жилкованием имеют одну (иногда больше) первичную жилку в центре листа, называемую средней жилкой или ребром, которая переходит в сосудистую сеть черешка. Вторичные жилки, также известные как жилки второго порядка или боковые жилки, отходят от средней жилки и тянутся к краям листа. Они часто заканчиваются гидатодой , секреторным органом, на краю. В свою очередь, более мелкие вены отходят от вторичных вен, известных как вены третичного или третьего (или более высокого порядка), образуя плотный сетчатый узор. Участки или островки мезофилла, лежащие между жилками высшего порядка, называются ареолами . Некоторые из самых маленьких вен (прожилок) могут иметь окончания в ареолах — процесс, известный как ареоляция. [ 31 ] Эти второстепенные жилки действуют как места обмена между мезофиллом и сосудистой системой растения. [ 26 ] Таким образом, малые жилки собирают продукты фотосинтеза (фотосинтеза) из клеток, где он происходит, а крупные отвечают за их транспортировку за пределы листа. В то же время вода транспортируется в противоположном направлении. [ 32 ] [ 28 ] [ 27 ]
Количество окончаний вен очень варьируется, как и то, заканчиваются ли вены второго порядка на краю или соединяются с другими венами. [ 29 ] Существует множество сложных вариаций узоров, образующих жилки листьев, и они имеют функциональное значение. Из них наибольшее разнообразие имеют покрытосеменные. [ 30 ] Внутри них основные жилки функционируют как опорная и распределительная сеть для листьев и коррелируют с формой листьев. Например, параллельное жилкование, обнаруженное у большинства однодольных, коррелирует с их удлиненной формой листа и широким основанием листа, в то время как сетчатое жилкование наблюдается в простых цельных листьях, тогда как пальцевидные листья обычно имеют жилкование, в котором три или более первичных жилок расходятся радиально из одной точки. . [ 33 ] [ 26 ] [ 31 ] [ 34 ]
С эволюционной точки зрения ранние таксоны, как правило, имеют дихотомическое ветвление, а сетчатые системы появляются позже. Жилки появились в пермский период (299–252 млн лет назад), до появления покрытосеменных растений в триасе (252–201 млн лет назад), в течение которого появилась иерархия жилок, обеспечивающая более высокую функцию, больший размер листьев и адаптацию к более широкому разнообразию климатических условий. . [ 30 ] Хотя это более сложная структура, ветвящиеся жилки кажутся плезиоморфными и в той или иной форме присутствовали у древних семенных растений еще 250 миллионов лет назад. Псевдосетчатое жилкование, которое на самом деле представляет собой сильно модифицированное пернипараллельное жилкование, представляет собой аутапоморфию некоторых Melanthiaceae , которые являются однодольными; например, Paris Quadrifolia (Узел истинного любовника). У листьев с сетчатым жилкованием жилки образуют каркасный матрикс, придающий листьям механическую жесткость. [ 35 ]
Изменения морфологии внутри одного растения
[ редактировать ]- Гомобластия
- Характеристика, при которой растение имеет небольшие изменения в размере, форме и характере роста листьев между ювенильной и взрослой стадиями, в отличие от;
- Гетеробластия
- Характеристика, при которой у растения наблюдаются заметные изменения размера, формы и особенностей роста листьев между ювенильной и взрослой стадиями.
Анатомия
[ редактировать ]Среднемасштабные функции
[ редактировать ]Листья обычно сильно васкуляризированы и обычно имеют сеть сосудистых пучков , содержащих ксилему , которая поставляет воду для фотосинтеза , и флоэму , которая транспортирует сахара, образующиеся в результате фотосинтеза. Многие листья покрыты трихомами (маленькими волосками), имеющими разнообразную структуру и функции.

Мелкомасштабные функции
[ редактировать ]Основными тканевыми системами являются:
- Эпидермис . , покрывающий верхнюю и нижнюю поверхности
- , Ткань мезофилла состоящая из фотосинтезирующих клеток, богатых хлоропластами . (также называемая хлоренхимой )
- Расположение вен ( сосудистой ткани )
Эти три тканевые системы обычно образуют регулярную организацию на клеточном уровне. Специализированные клетки, которые заметно отличаются от окружающих клеток и которые часто синтезируют специализированные продукты, такие как кристаллы, называются идиобластами . [ 36 ]
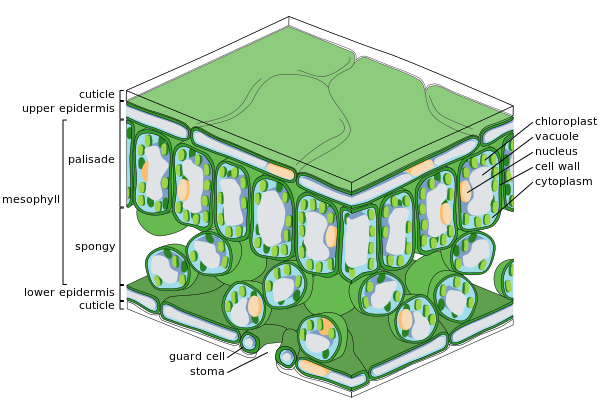
Основные ткани листа
[ редактировать ]-
 Разрез листа
Разрез листа -
 Эпидермальные клетки
Эпидермальные клетки -
 Клетки губчатого мезофилла
Клетки губчатого мезофилла



Эпидермис
[ редактировать ]
Эпидермис покрывающий – это внешний слой клеток, лист. Он покрыт восковой кутикулой , непроницаемой для жидкой воды и водяного пара и образующей границу, отделяющую внутренние клетки растения от внешнего мира. Кутикула в некоторых случаях тоньше на нижнем эпидермисе, чем на верхнем эпидермисе, и обычно толще на листьях из сухого климата по сравнению с листьями из влажного климата. [ 37 ] Эпидермис выполняет несколько функций: защиту от потери воды путем транспирации , регуляцию газообмена и секреции метаболических соединений. Большинство листьев имеют дорсовентральную анатомию: верхняя (адаксиальная) и нижняя (абаксиальная) поверхности имеют несколько разное строение и могут выполнять разные функции.
Ткань эпидермиса включает несколько дифференцированных типов клеток; эпидермальные клетки, эпидермальные волосковые клетки ( трихомы ), клетки устьичного комплекса; замыкающие клетки и вспомогательные клетки. Эпидермальные клетки являются наиболее многочисленными, крупнейшими и наименее специализированными и составляют большую часть эпидермиса. они обычно более удлиненные, У однодольных чем у двудольных .
Хлоропласты в эпидермальных клетках обычно отсутствуют, за исключением замыкающих клеток устьиц . Устьичные поры пронизывают эпидермис и с каждой стороны окружены замыкающими клетками, содержащими хлоропласты, и двумя-четырьмя вспомогательными клетками, лишенными хлоропластов, образуя специализированную группу клеток, известную как устьичный комплекс. Открытие и закрытие устьичного отверстия контролируется устьичным комплексом и регулирует обмен газов и водяных паров между наружным воздухом и внутренней частью листа. Таким образом, устьица играют важную роль в обеспечении фотосинтеза, не позволяя листу высыхать. В типичном листе устьица более многочисленны в абаксиальном (нижнем) эпидермисе, чем в адаксиальном (верхнем) эпидермисе, и более многочисленны у растений из более прохладного климата.
Мезофилл
[ редактировать ]Большая часть внутренней части листа между верхним и нижним слоями эпидермиса представляет собой паренхиму (основную ткань) или хлоренхимы, ткань называемую мезофиллом (по-гречески «средний лист»). Эта ассимиляционная ткань является основным местом фотосинтеза в растении. Продукты фотосинтеза называются «ассимилятами».
У папоротников и большинства цветковых растений мезофилл разделен на два слоя:
- Верхний палисадный слой из вертикально вытянутых клеток толщиной в одну-две клетки расположен непосредственно под адаксиальным эпидермисом, с межклеточными воздушными промежутками между ними. Его клетки содержат гораздо больше хлоропластов, чем губчатый слой. Цилиндрические клетки, хлоропласты которых расположены близко к стенкам, могут оптимально использовать свет. Небольшое разделение клеток обеспечивает максимальное поглощение углекислого газа. Солнечные листья имеют многослойный палисадный слой, тогда как теневые листья или более старые листья, расположенные ближе к почве, однослойные.
- Под слоем палисада находится губчатый слой . Клетки губчатого слоя более разветвлены и не так плотно упакованы, поэтому между ними имеются большие межклеточные воздушные пространства. Поры или устьица эпидермиса открываются в подустьичные камеры, которые соединены с межклеточными воздушными пространствами между губчатой и палисадной клеткой мезофилла, так что кислород, углекислый газ и водяной пар могут диффундировать в лист и выходить из него и получать доступ к клеткам мезофилла. при дыхании, фотосинтезе и транспирации.
Листья обычно зеленые из-за наличия хлорофилла в хлоропластах клеток мезофилла. У некоторых растений листья разного цвета из-за присутствия дополнительных пигментов, таких как каротиноиды в клетках мезофилла .
Сосудистая ткань
[ редактировать ]
Жилки сосудистую представляют собой ткань листа и расположены в губчатом слое мезофилла. Рисунок жилок называется жилкованием . У покрытосеменных жилкование обычно параллельное у однодольных образует соединительную сеть , а у широколистных растений . Когда-то их считали типичными примерами формирования паттернов посредством ветвления , но вместо этого они могут служить примером паттерна, сформированного в поле тензора напряжений . [ 38 ] [ 39 ] [ 40 ]
Вена состоит из сосудистого пучка . В основе каждого пакета лежат кластеры из двух различные типы проводящих клеток:
- Ксилема
- Клетки, которые переносят воду и минеральные вещества из корней в лист.
- Флоэма
- Клетки, которые обычно перемещают сок с растворенной сахарозой (от глюкозы до сахарозы), образующейся в результате фотосинтеза в листе, из листа.
Ксилема обычно лежит на адаксиальной стороне сосудистого пучка, а флоэма обычно лежит на абаксиальной стороне. Оба встроены в плотную ткань паренхимы, называемую оболочкой, которая обычно включает некоторую структурную ткань колленхимы.
Развитие листьев
[ редактировать ]Согласно Агнес Арбер , листья представляют собой частичные побеги. теории листа [ 41 ] происходит из зачатков листьев верхушки побега. На ранних стадиях развития они уплощены в дорзивентральном направлении как с дорсальной, так и с вентральной поверхностью. [ 14 ] Сложные листья ближе к побегам, чем простые. Исследования развития показали, что сложные листья, как и побеги, могут разветвляться в трех измерениях. [ 42 ] [ 43 ] На основе молекулярной генетики Эккардт и Баум (2010) пришли к выводу, что «в настоящее время общепринято, что сложные листья проявляют свойства как листа, так и побега». [ 44 ] Многие двудольные листья демонстрируют эндогенно обусловленную суточную ритмику роста. [ 45 ] [ 46 ] [ 47 ]
Экология
[ редактировать ]Биомеханика
[ редактировать ]Растения реагируют и адаптируются к факторам окружающей среды, таким как свет и механический стресс от ветра. Листьям необходимо поддерживать свою массу и выравниваться таким образом, чтобы оптимизировать воздействие солнца, обычно более или менее горизонтально. Однако горизонтальное выравнивание максимизирует воздействие изгибающих сил и отказов от таких стрессов, как ветер, снег, град, падающие обломки, животные и истирание окружающей листвы и растительных структур. В целом листья относительно хрупкие по сравнению с другими структурами растений, такими как стебли, ветви и корни. [ 48 ]
И листовая пластинка, и структура черешка влияют на реакцию листа на такие силы, как ветер, позволяя в определенной степени менять положение, чтобы минимизировать сопротивление и повреждение, а не сопротивление. Подобное движение листьев может также увеличить турбулентность воздуха вблизи поверхности листа, что утончает пограничный слой воздуха, непосредственно прилегающий к поверхности, увеличивая способность к газообмену и теплообмену, а также фотосинтезу. Сильные силы ветра могут привести к уменьшению количества листьев и площади поверхности, что, хотя и снижает сопротивление, влечет за собой компромисс , связанный с уменьшением фотосинтеза. Таким образом, конструкция листьев может включать компромисс между приростом углерода, терморегуляцией и потерей воды, с одной стороны, и затратами на выдерживание как статических, так и динамических нагрузок. В сосудистых растениях перпендикулярные силы распространяются на большую площадь и относительно гибки как при изгибе, так и при кручении , что обеспечивает упругую деформацию без повреждений. [ 48 ]
Прочность многих листьев зависит от гидростатической поддержки, организованной вокруг скелета сосудистой ткани, которая зависит от поддержания состояния воды в листе. И механика, и архитектура створки отражают потребность в транспортировке и поддержке. Рид и Стоукс (2006) рассматривают две основные модели: «гидростатическую» и «двутавровую» форму (см. Рис. 1). [ 48 ] Гидростатические листья, такие как Prostanthera lasianthos, большие и тонкие, поэтому может потребоваться несколько листьев, а не одиночные большие листья из-за количества жилок, необходимых для поддержки периферии больших листьев. Но большой размер листьев способствует эффективности фотосинтеза и сохранению воды, что требует дополнительных компромиссов. С другой стороны, двутавровые листья, такие как Banksia Marginata, имеют специальные структуры, придающие им жесткость. Эти двутавровые балки образуются из расширений оболочки склеренхимы , встречающихся с жесткими субэпидермальными слоями. Это смещает баланс с опоры на гидростатическое давление на структурную поддержку, что является очевидным преимуществом там, где воды относительно мало. [ 48 ] Длинные узкие листья легче сгибаются, чем яйцевидные листовые пластинки той же площади. Однодольные растения обычно имеют такие линейные листья, которые максимизируют площадь поверхности и сводят к минимуму самозатенение. В них высокая доля продольных основных жилок обеспечивает дополнительную поддержку. [ 48 ]
Взаимодействие с другими организмами
[ редактировать ]
Хотя листья не так питательны, как другие органы, такие как фрукты, они служат источником пищи для многих организмов. Лист является жизненно важным источником производства энергии для растений, и растения выработали защиту от животных, которые потребляют листья, например дубильные вещества , химические вещества, которые препятствуют перевариванию белков и имеют неприятный вкус. Животные, которые специализируются на поедании листьев, известны как фоливоры .
У некоторых видов есть загадочные приспособления, с помощью которых они используют листья, чтобы избежать хищников. Например, гусеницы некоторых листоверток создают в листе небольшой дом, складывая его на себя. Некоторые пилильщики аналогичным образом скручивают в трубочки листья своих пищевых растений. Самки Attelabidae , так называемых листоверток, откладывают яйца в листья, которые затем свертывают в качестве средства защиты. Другие травоядные животные и их хищники имитируют внешний вид листа. Рептилии, такие как некоторые хамелеоны, и насекомые, такие как некоторые кузнечики , также имитируют колебательные движения листьев на ветру, перемещаясь из стороны в сторону или вперед и назад, уклоняясь при этом от возможной угрозы.
Сезонная потеря листьев
[ редактировать ]
Листья в умеренных , бореальных и сезонно засушливых зонах могут быть сезонно лиственными (опадать или отмирать в ненастный сезон). Этот механизм сбрасывания листьев называется опадением . Когда лист сбрасывается, на ветке остается рубец. Холодной осенью они иногда меняют цвет и становятся желтыми , ярко - оранжевыми или красными , так как различные вспомогательные пигменты ( каротиноиды и ксантофиллы ) обнаруживаются, когда дерево реагирует на холод и уменьшение количества солнечного света , сокращая выработку хлорофилла. Сейчас считается, что красные антоциановые пигменты вырабатываются в листьях по мере их отмирания, возможно, для маскировки желтого оттенка, остающегося при потере хлорофилла - желтые листья, по-видимому, привлекают травоядных животных, таких как тля . [ 49 ] Оптическая маскировка хлорофилла антоцианами снижает риск фотоокислительного повреждения клеток листьев по мере их старения, что в противном случае может снизить эффективность извлечения питательных веществ из стареющих осенних листьев. [ 50 ]
Эволюционная адаптация
[ редактировать ]
В ходе эволюции листья приспособились к различным средам следующим образом: [ нужна ссылка ]
- Восковые микро- и наноструктуры на поверхности уменьшают намокание дождем и прилипание загрязнений ( см. Эффект лотоса ).
- Разделенные и сложные листья уменьшают сопротивление ветра и способствуют охлаждению.
- Волоски на поверхности листьев задерживают влагу в сухом климате и создают пограничный слой, уменьшающий потерю воды.
- Кутикулы восковых растений уменьшают потерю воды.
- Большая площадь поверхности обеспечивает большую площадь улавливания солнечного света.
- При вредном уровне солнечного света специализированные листья, непрозрачные или частично скрытые, пропускают свет через полупрозрачное окошко для фотосинтеза на внутренней поверхности листа (например, фенестрария ).
- Анатомия листьев Кранца у растений, осуществляющих C 4 фиксацию углерода
- Сочные листья хранят воду и органические кислоты для использования в фотосинтезе САМ .
- Ароматические масла , яды или феромоны, вырабатываемые железами листьев, отпугивают травоядных животных (например, эвкалипты ).
- Включения кристаллических минералов отпугивают травоядных животных (например, кремнеземные фитолиты в травах, рафиды в ароидных ).
- Лепестки привлекают опылителей.
- Шипы защищают растения от травоядных животных (например, кактусов ).
- Жгучие волосы для защиты от травоядных животных, например, у Urtica dioica и Dendrocnide moroides ( Urticaceae ).
- Специальные листья хищных растений приспособлены для захвата пищи, в основном беспозвоночных, хотя некоторые виды ловят и мелких позвоночных (см. Плотоядные растения ).
- Луковицы хранят пищу и воду (например, лук ).
- Усики позволяют растению карабкаться (например, гороху).
- Прицветники и псевдоцветки (ложные цветки) заменяют нормальные цветочные структуры, когда настоящие цветки сильно редуцируются (например, молочай , повязки у ароидных и цветочные головки у астровых ).
Терминология
[ редактировать ]
Форма
[ редактировать ]
Край (поля)
[ редактировать ]Край — или поле это внешний периметр листа. Термины взаимозаменяемы.
| Изображение | Срок | латинский | Описание |
|---|---|---|---|
| Весь | Форма Интегра |
Даже; с ровным краем; без зубьев | |
| Инфузория | ресничный | Окаймленный волосками | |
| Кренат | городчатый | Волнистозубый; зубчатый с закругленными зубами | |
| зубчатый | хохлатый | Мелко городчатый | |
| хрустящий | кудрявый | Кудрявый | |
| Зубчатый | зубчатый | Зубчатый;
могут быть крупнозубчатыми , с крупными зубами или железисто-зубчатые , имеющие зубы с железами. | |
| зубчатый | зубчатый | Мелкозубчатый | |
| Двойно зубчатый | двузубый | Каждый зуб имеет меньшие зубы | |
| Зубчатый | зубчатый | Пилообразный; с асимметричными зубами, направленными вперед | |
| Зубчатый | зубчатый | Мелко зубчатые | |
| Выемчатый | извилистый | С глубокими волнообразными углублениями; грубо городчатый | |
| лопастной | хвалил | С отступом, углубления не доходят до центра. | |
| волнистый | волнистый | С волнистым краем, более мелким, чем выемчатый. | |
| Колючий или острый | с шипами | С жесткими острыми кончиками, такими как чертополох. |
Апекс (наконечник)
[ редактировать ]| Изображение | Срок | латинский | Описание |
|---|---|---|---|
| Остроконечный | _ | Длинноконечный, переходящий в узкий, вогнуто сужающийся кончик. | |
| Острый | _ | Окончание в острой, но не продолжительной точке | |
| остроконечный | _ | С острым, удлиненным, жестким кончиком; с острием | |
| Маргинализированные | _ | Зазубренный, с неглубокой выемкой на конце. | |
| Мукронат | _ | Резкий кончик с небольшим коротким острием | |
| мукронулировать | _ | Мукронат, но с заметно миниатюрным шипом. | |
| Запретить | _ | В форме обратного сердца | |
| Тупой | _ | Закругленный или тупой | |
| Усечь | _ | Резко заканчивается с плоским концом |
База
[ редактировать ]- Остроконечный
- Подходим к острой, узкой, продолжительной точке.
- Острый
- Подходим к острой, но не продолжительной точке.
- Наушники
- Ушной формы.
- сердцевидный
- Сердцевидной формы с выемкой к плодоножке.
- Клин
- Клиновидный.
- копье
- По форме напоминает алебарду, базальные лопасти направлены наружу.
- Косой
- Косой.
- почкообразный
- Почковидной формы, но более круглые и широкие, чем длинные.
- Закругленный
- Изогнутая форма.
- Стрелять
- По форме напоминает наконечник стрелы, с острыми базальными лопастями, направленными вниз.
- Усечь
- Резко заканчивается плоским концом, который выглядит отрезанным.
Поверхность
[ редактировать ]
Поверхность листьев также является местом обитания большого количества микроорганизмов ; в этом контексте ее называют филлосферой .
- Лепидот
- Покрыты мелкими шелушащимися чешуйками.
волосатость
[ редактировать ]


«Волоски» на растениях правильно называются трихомами . Листья могут иметь несколько степеней опушения. Значения некоторых из следующих терминов могут совпадать.
- Арахноидальный, или арахноз
- Со множеством тонких, запутанных волосков, создающих впечатление паутины.
- Барбеллате
- С мелко колючими волосками (усиками).
- Бородатый
- С длинными жесткими волосами.
- Щетинистый
- С жесткими колючками, похожими на волосы.
- канесцентный
- Седой, с густым серовато-белым опушением.
- Инфузория
- По краям окаймлены короткими волосками (ресничками).
- Ресничный
- Мелкореснитчатые.
- слоеный
- С пучками мягких, шерстяных волос, которые имеют свойство стираться.
- глянцевый
- Выпадение волос с возрастом.
- Голый
- Никаких волос нету.
- Железистый
- С железой на кончике волоса.
- Хирсуте
- С довольно грубой или жесткой шерстью.
- Хиспид
- С жесткими щетинистыми волосками.
- безумный
- Мгновенно шипел.
- седой
- С тонким плотным серовато-белым опушением.
- Ланате, или ланосе
- С густыми волосами.
- Пилосе
- С мягкими, четко разделенными волосками.
- Половозрелый, или половозрелый
- С тонкими волосками.
- половозрелый
- С мягкой, короткой и прямостоячей шерстью.
- Чешуйчатый, или чешуйчатый
- Грубый на ощупь.
- серьезный
- Шелковистый вид благодаря тонким, прямым и прижатым (плотно прилегающим и плоским) волоскам.
- шелковистый
- С прижатым, мягким и прямым опушением.
- Звездчатая, или звездчатая
- С волосами звездообразной формы.
- Стригоза
- С прижатыми, острыми, прямыми и жесткими волосками.
- войлочный
- Густо опушён спутанными мягкими белыми пушистыми волосками.
- Кано-войлочный
- Между канесцентным и войлочным.
- Валяно-войлочный
- Шерстистый и спутанный с вьющимися волосками.
- Войлочная лоза
- Мелко или слегка войлочно-опушенный.
- ворсинчатый
- С длинными и мягкими волосками, обычно изогнутыми.
- Шерстистый
- С длинными, мягкими и извилистыми или спутанными волосками.
Тайминг
[ редактировать ]Жилкование
[ редактировать ]Классификация
[ редактировать ]





Описан ряд различных систем классификации рисунков жилок листьев (жилкование или жилкование). [ 29 ] начиная с Эттингсгаузена (1861 г.), [ 53 ] вместе со множеством различных описательных терминов, а терминология была описана как «грозная». [ 29 ] Одной из наиболее распространенных среди них является система Хики, первоначально разработанная для « двудольных » и использующая ряд терминов Эттингсгаузена, заимствованных из греческого языка (1973–1979): [ 54 ] [ 55 ] [ 56 ] ( см. также : Симпсон Рисунок 9.12, стр. 468) [ 29 ]
Система Хикки
[ редактировать ]- 1. Перистые (перисто-жилковые, сетчатые, перисто-сетчатые, перисто-ребристые, перисто-нервные или перисто-жилковые).
- Вены возникают перистыми (перистыми) из единственной первичной вены (средней вены) и подразделяются на вторичные прожилки, известные как вены более высокого порядка. Они, в свою очередь, образуют сложную сеть. Этот тип жилкования типичен (но не ограничивается) « двудольными » (неоднодольными покрытосеменными ). Например, Остря . Различают три подтипа перистого жилкования:
- Краспедодромус (греч. kraspedon – край, дромос – бег)
- Основные жилки доходят до края листа.
- Камптодромный
- Основные вены проходят близко к краю, но изгибаются до пересечения с краем.
- гиподромный
- Все вторичные вены отсутствуют, рудиментарны или скрыты.
Они, в свою очередь, имеют ряд дополнительных подтипов, таких как эвкамптодромный, при котором вторичные жилки изгибаются вблизи края, не соединяясь с соседними вторичными жилками.



- 2. Параллелодромные (параллельножилковые, параллельноребристые, параллельнонервные, пеннипараллельные, полосатые).
- Две или несколько первичных жилок, начинающихся рядом друг с другом у основания листа, идущих параллельно друг другу до верхушки, а затем сходящихся там. Комиссуральные вены (мелкие вены) соединяют большие параллельные вены. Типично для большинства однодольных растений , например трав . Также используются дополнительные термины маргинальный (первичные жилки достигают края) и сетчатый (сетчатый).

- 3. Кампилодромный ( campylos – изогнутый)
- Несколько первичных жилок или ветвей, берущих начало в одной точке или вблизи нее и идущих изогнутыми дугами, а затем сходящихся на вершине. Например, Майантемум .

- 4. Акродромный
- Две или более первичных или хорошо развитые вторичные жилки в сходящихся дугах к вершине, без базальной кривизны, как у кампилодромного. Могут быть базальными или супрабазальными в зависимости от происхождения, совершенными или несовершенными в зависимости от того, достигают ли они 2/3 длины вершины. Например, Miconia (базальный тип), Endlicheria (супрабазальный тип).




- 5. Актинодромный
- Три и более первичных жилок, радиально расходящихся из одной точки. Например, Arcangelisia (базальный тип), Givotia (супрабазальный тип).


- 6. Палинактодромный
- Первичные жилки с одной или несколькими точками вторичного дихотомического ветвления за пределами первичного расхождения, расположенные близко или более удаленно. Например, Платан .


Типы 4–6 аналогичным образом можно подразделить на базальные (первичные первичные волокна соединяются у основания лопасти) или супрабазальные (расходящиеся над основанием лопасти), а также совершенные или несовершенные, а также флебеллатные.
Примерно в то же время Мелвилл (1976) описал систему, применимую ко всем покрытосеменным растениям, используя латинскую и английскую терминологию. [ 57 ] В Мелвилле также было шесть подразделений в зависимости от порядка развития вен.
- Арбускулярный (arbuscularis)
- Многократное ветвление с помощью регулярной дихотомии с образованием трехмерной кустообразной структуры, состоящей из линейных сегментов (2 подкласса)
- Флабеллат (флабеллат)
- Первичные жилки прямые или лишь слегка изогнутые, веерообразно расходящиеся от основания (4 подкласса).
- Пальчатая (пальматус)
- Изогнутые первичные вены (3 подкласса)
- Перистый (перистый)
- Единственная первичная жилка, средняя жилка, вдоль которой через более или менее равные промежутки расположены прямые или дугообразные вторичные жилки (6 подклассов).
- Коллимат (коллимат)
- Многочисленные продольно-параллельные первичные жилки, отходящие от поперечной меристемы (5 подклассов).
- Конглютинат (conglutinatus)
- Произведено от сросшихся перистых листочков (3 подкласса).
Модифицированная форма системы Хики была позже включена в Смитсоновскую классификацию (1999), которая предложила семь основных типов жилкования, основанных на архитектуре первичных жилок, с добавлением Flabellate в качестве дополнительного основного типа. Затем была проведена дальнейшая классификация на основе вторичных жил с еще 12 типами, такими как;
- Брохидодромный
- Закрытая форма, в которой второстепенные ветви соединены в ряд выступающих арок, как у Хильдегардии .
- Краспедодромный
- Открытая форма с вторичными ветвями, заканчивающимися по краю зубчатыми листьями, как у Celtis .
- эвкамптодромный
- Промежуточная форма с перевернутыми вторичными ветвями, которые постепенно уменьшаются на вершине, но внутри края, и соединены промежуточными третичными жилками, а не петлями между вторичными ветвями, как у Cornus .
- кладодромный
- Вторичные ветви свободно ветвятся к краю, как у Rhus .
термины, которые использовались в качестве подтипов в исходной системе Хики. [ 58 ]





Хильдегардия мигеоди

Западные кельты

Корнус лекарственный

Русус яйцевидный
Дальнейшие описания включали жилки более высокого порядка или второстепенные жилки и узоры ареол ( см. Рабочую группу по архитектуре листьев, рисунки 28–29). [ 58 ]

- Фантастически
- От нескольких до многих одинаковых тонких базальных жилок, расходящихся радиально под небольшими углами и ветвящихся на вершине. Например, Параномус .

При анализе рисунка жилок часто учитываются порядки жилок, тип первичных жил, тип вторичных жил (основные жилы) и плотность второстепенных жилок. Ряд авторов приняли упрощенные варианты этих схем. [ 59 ] [ 29 ] В простейшем случае основные типы жилок можно разделить на три или четыре группы в зависимости от рассматриваемых подразделений растения;
- перистый
- пальмовидный
- параллельный
где пальчатая относится к множеству первичных жилок, отходящих от черешка, в отличие от ответвлений от центральной главной жилки в перистой форме, и включает оба типа засоса 4 и 5, которые сохраняются как подтипы; например, пальчато-акродромный ( см. Справочник Службы национальных парков). [ 60 ]

- Пальчато-сетчатые, пальчато-сетчатые, пальчато-жилковые, веерообразно-жилковые.
- Несколько основных жилок примерно одинакового размера расходятся от общей точки у основания листа, где прикрепляется черешок, и расходятся к краю листа. Листья с пальчатыми прожилками часто лопастные или разделенные, лопасти расходятся от общей точки. Они могут различаться по числу первичных жилок (3 и более), но всегда расходятся из общей точки. [ 61 ] например, большинство Acer (клены).

Другие системы
[ редактировать ]Альтернативно, Симпсон использует: [ 29 ]
- Хладнокровный
- Центральная средняя жилка без боковых жилок ( микрофилл ), наблюдается у несемянных трахеофитов , таких как хвощи.
- дихотомический
- Вены последовательно разветвляются на вены одинакового размера из общей точки, образуя Y-образное соединение и расходясь веером. Среди древесных растений умеренного пояса гинкго двулопастный — единственный вид с дихотомическим жилкованием. Также некоторые птеридофиты (папоротники). [ 61 ]
- Параллельно
- Первичные и вторичные жилки примерно параллельны друг другу, проходят по всей длине листа, часто соединены короткими перпендикулярными звеньями, а не образуют сети. У некоторых видов параллельные жилки соединяются у основания и вершины, например, у игольчатых вечнозеленых растений и трав. Характерно для однодольных, но исключения включают Arisaema и, как показано ниже, сетчатую. [ 61 ]
- Сетчатые (сетчатые, перистые)
- Выраженная средняя жилка, по обеим сторонам которой отходят вторичные жилки. Название происходит от конечных прожилок, которые образуют взаимосвязанную сеть, подобную узору или сети. (Первичное и вторичное жилкование можно назвать перистым, а сетчатое, как и более тонкие жилки, - сетчатым или сетчатым); большинство неоднодольных покрытосеменных, за исключением Calophyllum . Некоторые однодольные растения имеют сетчатое жилкование, в том числе Colocasia , Dioscorea и Smilax . [ 61 ]

Редуцированные микрофилловые листья (L), отходящие в мутовке от узла.

Дихотомическое жилкование
Однако эти упрощенные системы допускают дальнейшее разделение на несколько подтипов. Симпсон, [ 29 ] (и другие) [ 62 ] разделяет параллельные и сетчатые (и некоторые используют только эти два термина для покрытосеменных) [ 63 ] по числу первичных жилок (коста) следующим образом;
- Параллельно
- Penni-parallel (pinnate, pinnate parallel, unicostate parallel)
- Единственная центральная выступающая средняя жилка, отходящие от нее вторичные вены отходят перпендикулярно ей и идут параллельно друг другу по направлению к краю или кончику, но не соединяются (анастомозируют). Термин «уникостат» относится к выступающей единственной средней жилке (косте), проходящей по всей длине листа от основания до вершины. например, Zingiberales , такие как бананы и т. д.
- Пальчато-параллельный (многосостоятельный параллельный)
- Несколько одинаково выраженных первичных жилок, исходящих из одной точки у основания и идущих параллельно кончику или краю. Термин «мультикостный» относится к наличию более одной выступающей главной вены. например, «веерные» (пальчатые) пальмы (Arecaceae)
- Многоуровневая параллельная конвергенция
- Средние жилки сходятся на вершине, например, Bambusa arundinacea = B. bambos (Aracaceae), Eichornia.
- Многосостояние параллельное расхождение
- Средние жилки расходятся более или менее параллельно к краю, например, Borassus (Poaceae), веерные пальмы.
- Сетчатый (сетчатый)
- Перистолистные (жилковые, сетчатые, одностворчатые).
- Одна выступающая средняя жилка, идущая от основания к вершине, вторичные жилки, отходящие с обеих сторон по длине первичной средней жилки, идущие к краю или вершине (кончику), с сетью более мелких жилок, образующих сетку (сетку или сеть). например Mangifera , Ficus religiosa , Psidium guajava , Hibiscus rosa-sinensis , Salix alba.
- Пальчатые (многодольные сетчатые)
- Несколько первичных жилок, исходящих из одной точки и идущих от основания к вершине. например, Liquidambar styraciflua. Его можно подразделить далее;
- Мультикостный конвергентный
- Основные жилки расходятся от начала основания, затем сходятся к кончику. например , Зизифус , Смилакс , Корица
- Многосостояние расходящееся
- Все основные жилки расходятся к кончику. например , Gossypium , Cucurbita , Carica papaya , Ricinus communis
- Тройчатые (тройчато-сетчатые)
- Три первичные жилки, как указано выше, например ( см .) Ceanothus leucodermis , [ 64 ] C. tomentosus , [ 65 ] Энселия фариноза


Пальчато-параллельный

Многоуровневая параллельная конвергенция

Многосостояние параллельное расхождение
Перисто сетчатый

Пальчато-сетчатый

Многореберная пальчатая конвергенция

Многореберные пальчаторасходящиеся
Эти сложные системы мало используются в морфологическом описании таксонов, но полезны при идентификации растений. [ 29 ] хотя его критиковали за чрезмерное обременение жаргоном. [ 66 ]
Более старая и еще более простая система, используемая в некоторых растениях. [ 67 ] использует только две категории: открытую и закрытую.
- Открытый: жилки более высокого порядка имеют свободные окончания среди клеток и более характерны для неоднодольных покрытосеменных растений. Они чаще связаны с формой листьев: зубчатыми, лопастными или сложными. Их можно подразделить на;
- Перистые (перисто-жилковые) листья с главной центральной жилкой или ребром (средней жилкой), от которого отходит остальная жилковая система.
- Пальчатые, у которых три и более основных ребра поднимаются вместе у основания листа и расходятся вверх.
- Дихотомический, как у папоротников, где жилки многократно разветвляются.
- Закрытый: жилки высшего порядка соединены петлями, не заканчиваясь свободно между клетками. Обычно они имеют листья с гладкими очертаниями и характерны для однодольных растений.
- Их можно подразделить по тому, идут ли жилки параллельно, как у трав, или имеют другой рисунок.
Другие описательные термины
[ редактировать ]Существует также множество других описательных терминов, часто очень специализированных и привязанных к конкретным таксономическим группам. [ 68 ] Выраженность вен зависит от ряда особенностей. К ним относятся ширина вен, их выступание по отношению к поверхности пластинки и степень непрозрачности поверхности, которая может скрывать более мелкие вены. В связи с этим вены называются неясными , а порядок скрытых вен и верхняя, нижняя или обе поверхности уточняются дополнительно. [ 69 ] [ 61 ]
Термины, описывающие выступающие вены, включают буллезную , желобчатую , плоскую , желобчатую , вдавленную , выступающую и утопленную ( рис . 6.1 Hawthorne & Lawrence 2013). [ 66 ] [ 70 ] На разных участках листа жилки могут проявляться по-разному. Например, Pimenta Racemosa имеет желобчатую среднюю жилку на верхней поверхности, но она заметна на нижней поверхности. [ 66 ]
Описание выступания вен:
- Буллатировать
- Поверхность листа приподнята в виде ряда куполов между жилками на верхней поверхности и поэтому также с выраженными углублениями. например, Rytigynia pauciflora , [ 71 ] Винная лоза
- Направленный (каналикулирующий)
- Вены погружаются под поверхность, образуя округлый канал. Иногда путают с «желобами», потому что каналы могут функционировать как желоба для стока дождя и обеспечения высыхания, как у многих Melastomataceae . [ 72 ] например ( см .) Кисть перца (Myrtaceae), [ 73 ] Клидемия гирта (Melastomataceae).
- с желобами
- Жилки частично выступающие, гребень над поверхностью листовой пластинки, но с каналами, идущими вдоль каждой стороны, как желоба.
- Впечатленный
- Жилка образует приподнятую линию или гребень, лежащий ниже плоскости несущей ее поверхности, как бы вдавленной в нее, и часто обнажается на нижней поверхности. Ткани возле вен часто сморщиваются, придавая им впалый или рельефный вид.
- Затемнять
- Вены не видны или совсем нечеткие; если не указано, то не видно невооруженным глазом. например, Berberis gagnepainii . У этого барбериса жилки неясны только на нижней поверхности. [ 74 ]
- Видный
- Вена приподнята над окружающей поверхностью, чтобы ее можно было легко прощупать пальцем. например ( см .) Пимента кистевидная , [ 73 ] Спатифиллум каннифолиум [ 75 ]
- Утопленный
- Вена погружена под поверхность, более выражена, чем окружающие ткани, но более углублена в русле, чем при вдавленных венах. например, Калина складчатая .


Неясный (под поверхностью)

Видный

Утопленный
Описание других особенностей:
- Плинервий (плинервед)
- Более одной главной вены (нерва) у основания. Боковые вторичные жилки ветвятся от точки над основанием листа. Обычно выражается суффиксом , например, в трех- или трехслойном листе. В 3-слойном (триплиневидном) листе три основные жилки разветвляются над основанием пластинки (две вторичные жилки и главная жилка) и впоследствии идут по существу параллельно, как у Ceanothus и у Celtis . Точно так же пятижилковый (пятижилковый) лист имеет четыре второстепенные жилки и одну главную жилку. особенно заметен рисунок из 3–7 жилок У Melastomataceae . Этот термин также использовался в Vaccinieae . Этот термин использовался как синоним акродромного, пальчато-акродромного или супрабазального акродромного, и считается, что он имеет слишком широкое определение. [ 76 ] [ 76 ]
- Лестничная форма
- Вены расположены как ступеньки лестницы, особенно вены более высокого порядка.
- субмаргинальный
- Жилки идут близко к краю листа.
- Тринервед
- 2 основных базальных нерва, кроме среднего ребра
Схемы жилкования
[ редактировать ]| Изображение | Срок | Описание |
|---|---|---|
 |
дугообразный | Вторичное выгибание к вершине |
 |
дихотомический | Вены разделяются на две части |
 |
Продольный | Все вены расположены преимущественно вдоль средней вены. |
 |
Параллельно | Все вены параллельны и не пересекаются. |
 |
Перистый | Вторичные вены, отходящие от средней жилки |
 |
сетчатый | Все жилки многократно ветвятся, сетчатые. |
 |
Поворот | Жилки идут от центра листа и расходятся к краям. |
 |
поперечный | Третичные вены идут перпендикулярно оси главной вены, соединяя вторичные вены. |
Размер
[ редактировать ]Термины мегафилл , макрофилл , мезофилл , нотофилл , микрофилл , нанофилл и лептофилл используются для описания размеров листьев (в порядке убывания) в классификации, разработанной в 1934 году Кристеном К. Раункером и с тех пор модифицированной другими. [ 77 ] [ 78 ]
См. также
[ редактировать ]- Глоссарий морфологии листьев
- Глоссарий морфологии растений § Листья
- Корона (ботаника)
- Эволюционная история листьев
- Эволюционное развитие листьев
- Индекс площади листьев
- Концентрат листового белка
- Датчик листьев – устройство, измеряющее уровень влажности в листьях растений.
- Форма листа
- Вернация – прорастание листьев, а также расположение листьев в бутоне.
- гладкий лист как музыкальный инструмент
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Исав 2006 .
- ^ Главный 1953 год .
- ^ Перейти обратно: а б Маусет 2009 .
- ^ «Стреляющая система» . Словарь ботанической терминологии . Питомник искусств кактусов. nd Архивировано из оригинала 4 мая 2021 года . Проверено 4 мая 2021 г.
- ^ Джеймс и др. 1999 .
- ^ Перейти обратно: а б с д и Стюарт и Ротвелл 1993 .
- ^ Куни-Советтс и Саттлер, 1987 .
- ^ Цукая 2013 .
- ^ Фейжер 2006 .
- ^ Перселл 2016 .
- ^ Виллерт и др. 1992 .
- ^ Байер 1982 .
- ^ Марлот 1913–1932 .
- ^ Перейти обратно: а б с Симпсон 2011 , с. 356.
- ^ Крог 2010 .
- ^ Джеймс и Белл 2000 .
- ^ Хейвуд и др. 2007 .
- ^ Симпсон 2011 , стр. 356–357.
- ^ Халле 1977 .
- ^ Перейти обратно: а б с Иллюстрированная ботаника: введение в растения. Основные группы, семейства цветковых растений . Томсон Сайенс. 1984. с. 21.
- ^ Дидье Рейнхардт и Крис Кулемейер, «Филлотаксис у высших растений», в книге Майкла Т. Макмануса, Брюса Вейта, ред., Меристематические ткани в росте и развитии растений , январь 2002 г., ISBN 978-1-84127-227-6 , Уайли-Блэквелл.
- ^ Коксетер Х.С. (1961). Введение в геометрию . Уайли. п. 169.
- ^ Рейнхардт и Кулемайер, стр. 175
- ^ Ёнекура, Такааки; Ивамото, Акитоши; Фудзита, Хиронори; Сугияма, Мунетака (6 июня 2019 г.). Умулис, Дэвид (ред.). «Математические модельные исследования комплексного формирования основных и второстепенных филлотаксисных паттернов у растений с преобладающим акцентом на филлотаксис orixate» . PLOS Вычислительная биология . 15 (6): e1007044. Бибкод : 2019PLSCB..15E7044Y . дои : 10.1371/journal.pcbi.1007044 . ISSN 1553-7358 . ПМК 6553687 . ПМИД 31170142 .
- ^ Роллан-Лаган и др. 2009 .
- ^ Перейти обратно: а б с Стены 2011 .
- ^ Перейти обратно: а б с Дикисон 2000 .
- ^ Перейти обратно: а б Рудалль 2007 .
- ^ Перейти обратно: а б с д и ж г час я Симпсон 2011 , Жилкование листьев, стр. 465–468.
- ^ Перейти обратно: а б с д Сак и Скоффони, 2013 .
- ^ Перейти обратно: а б с Рот-Небельсик и др . , 2001 г.
- ^ Уэно и др. 2006 .
- ^ Рунионс и др. 2005 .
- ^ Мэсси и Мерфи 1996 , Текстура поверхностного жилкования
- ^ Багчи и др. 2016 .
- ^ Кот 2009 .
- ^ Клементс 1905 .
- ^ Кудер и др. 2002 .
- ^ Корсон и др. 2009 .
- ^ Лагуна и др. 2008 .
- ^ Арбер 1950 .
- ^ Рутисхаузер и Саттлер 1997 .
- ^ Лакруа и др. 2003 .
- ^ Экардт и Баум 2010 .
- ^ Пуаре, Ришар; Визе-Клинкенберг, Аника; Родитель Борис; Мелевчик, Майкл; Шурр, Ульрих; Тардье, Франсуа; Уолтер, Ахим (2010). «Суточная динамика роста листьев у однодольных и двудольных видов: эндогенные ритмы и температурные эффекты» . Журнал экспериментальной ботаники . 61 (6): 1751–1759. дои : 10.1093/jxb/erq049 . ISSN 1460-2431 . ПМЦ 2852670 . ПМИД 20299442 .
- ^ Мелевчик, Майкл; Фридли, Майкл; Кирхгесснер, Норберт; Уолтер, Ахим (25 июля 2013 г.). «Дневной рост листьев сои: новый метод анализа двумерного расширения листьев с высоким временным разрешением на основе подхода отслеживания маркеров (Martrack Leaf)» . Растительные методы . 9 (1): 30. дои : 10.1186/1746-4811-9-30 . hdl : 20.500.11850/76534 . ISSN 1746-4811 . ПМЦ 3750653 . ПМИД 23883317 .
- ^ Фридли, Майкл; Уолтер, Ахим (2015). «Суточная динамика роста молодых листочков сои (G lycine max) синхронна в разных положениях на растении» . Растение, клетка и окружающая среда . 38 (3): 514–524. дои : 10.1111/шт.12407 . ISSN 0140-7791 . ПМИД 25041284 .
- ^ Перейти обратно: а б с д и Рид и Стоукс 2006 .
- ^ Доринг и др. 2009 .
- ^ Фейлд и др. 2001 .
- ^ «Глоссарий Кью – определение истеричного состояния» . 3 декабря 2013. Архивировано из оригинала 3 декабря 2013 года . Проверено 12 мая 2017 г.
{{cite web}}: CS1 maint: bot: исходный статус URL неизвестен ( ссылка ) - ^ «Глоссарий Кью - определение синантуса» . 3 декабря 2013. Архивировано из оригинала 3 декабря 2013 года . Проверено 12 мая 2017 г.
{{cite web}}: CS1 maint: bot: исходный статус URL неизвестен ( ссылка ) - ^ Эттингсгаузен 1861 .
- ^ Хикки 1973 .
- ^ Хики и Вулф 1975 .
- ^ Хикки 1979 .
- ^ Мелвилл 1976 .
- ^ Перейти обратно: а б Рабочая группа по листовой архитектуре, 1999 .
- ^ Джадд и др. 2007 .
- ^ Флориссан Leaf Key 2016 .
- ^ Перейти обратно: а б с д и Клинг и др. 2005 , Жилкование листьев.
- ^ Берг 2007 .
- ^ Морфология покрытосеменных 2017 , Жилкование.
- ^ Симпсон 2017 , Ceanothus leucodermis
- ^ Симпсон 2017 , Ceanothus tomentosus
- ^ Перейти обратно: а б с Hawthorne & Lawrence 2013 , Жилкование листьев, стр. 135–136.
- ^ Каллен и др. 2011 .
- ^ Неотропики 2017 .
- ^ Оксфордский глоссарий гербариев, 2017 г ..
- ^ Оксфордский глоссарий гербариев, 2017 г. , Выдающееся положение вен.
- ^ Вердкорт и Бридсон 1991 .
- ^ Hemsley & Poole 2004 , Морфология листьев и сушка, с. 254
- ^ Перейти обратно: а б Хьюз 2017 , Пимента кистевидная
- ^ Каллен и др. 2011 , Berberis gagnepainii vol. II стр. 398
- ^ Квантлен 2015 , Spathiphyllum cannifolium
- ^ Перейти обратно: а б Педраса-Пеньалоса 2013 .
- ^ Уиттен и др. 1997 .
- ^ Уэбб, Лен (1 октября 1959 г.). «Физиономическая классификация тропических лесов Австралии». Журнал экологии . 47 (3). Британское экологическое общество: Журнал экологии Vol. 47, № 3, стр. 551–570: 555. Бибкод : 1959JEcol..47..551W . дои : 10.2307/2257290 . JSTOR 2257290 .
Библиография
[ редактировать ]Книги и главы
[ редактировать ]- Арбер, Агнес (1950). Естественная философия растительной формы . Архив Кубка . GGKEY:HCBB8RZREL4.
- Байер, МБ (1982). Новый справочник по Хавортии . Кирстенбош: Национальный ботанический сад Южной Африки . ISBN 978-0-620-05632-8 . Архивировано из оригинала 06 сентября 2023 г. Проверено 25 июля 2018 г.
- Берг, Линда (23 марта 2007 г.). Вводная ботаника: растения, люди и окружающая среда, издание для СМИ . Cengage Обучение. ISBN 978-1-111-79426-2 .
- Каллен, Джеймс; Колени, Сабина Г.; Кьюби, Х. Сюзанна Кьюби, ред. (2011) [1984–2000]. Европейская садовая флора, Цветковые растения: Руководство по идентификации растений, выращиваемых в Европе как на открытом воздухе, так и под стеклом. 5 томов (2-е изд.). Кембридж: Издательство Кембриджского университета . Архивировано из оригинала 28 декабря 2016 г. Проверено 8 марта 2017 г.
- Каттер, Э. Г. (1969). Анатомия растений, эксперимент и интерпретация. Часть 2. Органы . Лондон: Эдвард Арнольд. п. 117. ИСБН 978-0-7131-2302-9 .
- Дикисон, Уильям К. (2000). Интегративная анатомия растений . Академическая пресса . ISBN 978-0-08-050891-7 .
- Исав, Кэтрин (2006) [1953]. Эверт, Рэй Ф. (ред.). Анатомия растений Исава: меристемы, клетки и ткани тела растения: их структура, функции и развитие (3-е изд.). John Wiley & Sons Inc. Нью-Йорк: ISBN 978-0-470-04737-8 . Архивировано из оригинала 06 сентября 2023 г. Проверено 2 сентября 2017 г.
- Эттингсгаузен, К. (1861). Скелеты листьев двудольных с особым вниманием к изучению и идентификации остатков ископаемых растений . Вена: Классификация архитектуры двудольных.
- Хаупт, Артур Винг (1953). Морфология растений . МакГроу-Хилл .
- Хоторн, Уильям; Лоуренс, Анна (2013). Идентификация растений: создание удобных для пользователя полевых руководств по управлению биоразнообразием . Рутледж. ISBN 978-1-136-55972-3 .
- Хемсли, Алан Р.; Пул, Имоджен, ред. (2004). Эволюция физиологии растений . Академическая пресса . ISBN 978-0-08-047272-0 .
- Хейвуд, Вирджиния ; Браммитт, РК; Калхэм, А .; Себерг, О. (2007). Семейства цветковых растений мира . Нью-Йорк: Книги Светлячка. п. 287. ИСБН 978-1-55407-206-4 .
- Хикки, ЖЖ. Пересмотренная классификация строения двудольных листьев . стр. я 5–39. , в Меткалф и Мел (1979)
- Джадд, Уолтер С .; Кэмпбелл, Кристофер С.; Келлог, Элизабет А.; Стивенс, Питер Ф .; Донохью, Майкл Дж. (2007) [1-е изд. 1999, 2-е 2002]. Систематика растений: филогенетический подход (3-е изд.). Синауэр Ассошиэйтс. ISBN 978-0-87893-407-2 . Архивировано из оригинала 06 сентября 2023 г. Проверено 2 сентября 2017 г.
- Крог, Дэвид (2010), Биология: Путеводитель по миру природы (5-е изд.), Издательство Benjamin-Cummings Publishing Company, стр. 463, ISBN 978-0-321-61655-5 , заархивировано из оригинала 24 января 2023 г. , получено 24 мая 2016 г.
- Рабочая группа по листовой архитектуре (1999). Руководство по архитектуре листа - морфологическое описание и классификация двудольных и однодольных покрытосеменных с сетчатыми жилками (PDF) . Смитсоновский институт . ISBN 978-0-9677554-0-3 . Архивировано (PDF) из оригинала 20 октября 2016 г. Проверено 15 февраля 2017 г.
- Марлот, Рудольф (1913–1932). Флора Южной Африки: с синопическими таблицами родов высших растений. 6 томов . Кейптаун: Darter Bros. & Co. Архивировано из оригинала 6 сентября 2023 г. Проверено 27 августа 2020 г.
- Маусет, Джеймс Д. (2009). Ботаника: введение в биологию растений (4-е изд.). Садбери, Массачусетс: Издательство Jones and Bartlett. ISBN 978-0-7637-5345-0 .
- Меткалф, Чехия; Мел, Л., ред. (1979) [1957]. Анатомия двудольных: листья, стебель и древесина в связи с таксономией, с примечаниями об экономическом использовании. 2 тома (2-е изд.). Оксфорд: Кларендон Пресс. ISBN 978-0-19-854383-1 .
- Пранс, Гиллин Толми (1985). Листья: формирование, характеристики и использование сотен листьев, встречающихся во всех частях света . Фотографии Кьелла Б. Сандведа. Лондон: Темза и Гудзон. ISBN 978-0-500-54104-3 .
- Рудалл, Паула Дж. (2007). Анатомия цветковых растений: введение в строение и развитие (3-е изд.). Кембридж: Издательство Кембриджского университета . ISBN 978-0-521-69245-8 . Архивировано из оригинала 06 сентября 2023 г. Проверено 27 августа 2020 г.
- Симпсон, Майкл Г. (2011). Систематика растений . Академическая пресса. ISBN 978-0-08-051404-8 . Архивировано из оригинала 17 января 2023 г. Проверено 24 мая 2016 г.
- Стюарт, Уилсон Н; Ротвелл, Гар В. (1993) [1983]. Палеоботаника и эволюция растений (2-е изд.). Издательство Кембриджского университета . ISBN 978-0-521-38294-6 .
- Вердкур, Бернар; Бридсон, Дайан М. (1991). Флора тропической Восточной Африки - Rubiaceae Том 3 . ЦРК Пресс. ISBN 978-90-6191-357-3 .
- Уиттен, Тони; Сориатмаджа, Рохаят Эмон; Афифф, Сурая А. (1997). Экология Явы и Бали . Издательство Оксфордского университета. п. 505. ИСБН 978-962-593-072-5 . Архивировано из оригинала 06 сентября 2023 г. Проверено 27 августа 2020 г.
- Виллерт, Дитер Дж. фон; Эллер, Б.М.; Вергер, MJA; Бринкманн, Э; Иленфельдт, HD (1992). Жизненные стратегии суккулентов в пустынях: с особым упором на пустыню Намиб . Архив Кубка . ISBN 978-0-521-24468-8 .
Статьи и тезисы
[ редактировать ]- Багчи, Дебжани; Дасгупта, Авик; Гондалия, Амит Д.; Раджпут, Кишор С. (2016). «Информация из мира растений: подход к фрактальному анализу для настройки механической жесткости матрицы каркаса в тонких пленках». Передовые исследования материалов . 1141 : 57–64. дои : 10.4028/www.scientific.net/AMR.1141.57 . S2CID 138338270 .
- Клементс, Эдит Шварц (декабрь 1905 г.). «Связь структуры листа с физическими факторами» . Труды Американского микроскопического общества . 26 : 19–98. дои : 10.2307/3220956 . JSTOR 3220956 . Архивировано из оригинала 4 августа 2023 г. Проверено 6 сентября 2023 г.
- Куни-Советс, К.; Саттлер, Р. (1987). «Развитие филлоклады у Asparagaceae: пример гомеоза». Ботанический журнал Линнеевского общества . 94 (3): 327–371. дои : 10.1111/j.1095-8339.1986.tb01053.x .
- Корсон, Фрэнсис; Адда-Бедиа, Мохтар; Будауд, Арезки (2009). «Сети жилкования листьев In silico: рост и реорганизация, вызванные механическими силами» (PDF) . Журнал теоретической биологии . 259 (3): 440–448. Бибкод : 2009JThBi.259..440C . дои : 10.1016/j.jtbi.2009.05.002 . ПМИД 19446571 . S2CID 25560670 . Архивировано из оригинала (PDF) 9 декабря 2017 г.
- Кот, Г.Г. (2009). «Разнообразие и распределение идиобластов, продуцирующих кристаллы оксалата кальция, у Dieffenbachia seguine (Araceae)» . Американский журнал ботаники . 96 (7): 1245–1254. дои : 10.3732/ajb.0800276 . ПМИД 21628273 .
- Кудер, Ю.; Пошар, Л.; Аллен, К.; Адда-Бедиа, М.; Дуади, С. (1 июля 2002 г.). «Жилкование листа, сформированное в тензорном поле» (PDF) . Европейский физический журнал Б. 28 (2): 135–138. Бибкод : 2002EPJB...28..135C . дои : 10.1140/epjb/e2002-00211-1 . S2CID 51687210 . Архивировано из оригинала (PDF) 9 декабря 2017 года.
- Дёринг, Т.Ф; Арчетти, М.; Харди, Дж. (7 января 2009 г.). «Осенние листья глазами травоядных» . Труды Королевского общества B: Биологические науки . 276 (1654): 121–127. дои : 10.1098/rspb.2008.0858 . ПМК 2614250 . ПМИД 18782744 .
- Экардт, Северная Каролина; Баум, Д. (20 июля 2010 г.). «Загадка подостемад: эволюция необычной морфологии Podostemaceae» . Растительная клетка онлайн . 22 (7): 2104. doi : 10.1105/tpc.110.220711 . ПМЦ 2929115 . ПМИД 20647343 .
- Фейжье, Франсуа (14 декабря 2006 г.). Модели формирования сосудистого рисунка листьев (кандидатская диссертация) . Парижский университет VI . Архивировано (PDF) из оригинала 7 марта 2017 года . Проверено 6 марта 2017 г.
- Филд, Т.С.; Ли, Д.В.; Холбрук, Нью-Мексико (1 октября 2001 г.). «Почему листья краснеют осенью. Роль антоцианов в старении листьев кизила красно-ивового» . Физиология растений . 127 (2): 566–574. дои : 10.1104/стр.010063 . ПМЦ 125091 . ПМИД 11598230 .
- Халле, Ф. (1977). «Самый длинный лист на ладони». Принципы . 21:18 .
- Хики, Лео Дж. (1 января 1973 г.). «Классификация строения двудольных листьев» (PDF) . Американский журнал ботаники . 60 (1): 17–33. дои : 10.2307/2441319 . JSTOR 2441319 . Архивировано (PDF) из оригинала 11 августа 2017 года . Проверено 14 февраля 2017 г.
- Хики, Лео Дж.; Вулф, Джек А. (1975). «Основы филогении покрытосеменных: вегетативная морфология» . Анналы ботанического сада Миссури . 62 (3): 538–589. дои : 10.2307/2395267 . JSTOR 2395267 . Архивировано из оригинала 31 октября 2022 г. Проверено 6 сентября 2023 г.
- Ингерсолл, Эрнест (1920). . Американская энциклопедия . Том. XVII.
- Джеймс, ЮАР; Белл, Д.Т. (2000). «Влияние доступности света на структуру листьев и рост двух сортов Eucalyptus globulus ssp. globulus » (PDF) . Физиология дерева . 20 (15): 1007–1018. дои : 10.1093/treephys/20.15.1007 . ПМИД 11305455 . Архивировано (PDF) из оригинала 1 апреля 2022 г. Проверено 10 мая 2019 г.
- Лакруа, К.; Жен, Б.; Перселл-Макдональд, С. (2003). «Сравнение побегов и сложных листьев эвдикотов: динамическая морфология как альтернативный подход» . Ботанический журнал Линнеевского общества . 143 (3): 219–230. дои : 10.1046/j.1095-8339.2003.00222.x . Архивировано из оригинала 22 июня 2020 г. Проверено 8 сентября 2019 г.
- Лагуна, Мария Ф.; Бон, Штеффен; Ягла, Эдуардо А.; Борн, Филип Э. (2008). «Роль упругих напряжений в морфогенезе жилкования листьев» . PLOS Вычислительная биология . 4 (4): e1000055. arXiv : 0705.0902 . Бибкод : 2008PLSCB...4E0055L . дои : 10.1371/journal.pcbi.1000055 . ПМК 2275310 . ПМИД 18404203 .
- Мелвилл, Р. (ноябрь 1976 г.). «Терминология листовой архитектуры». Таксон . 25 (5/6): 549–561. дои : 10.2307/1220108 . JSTOR 1220108 .
- Педраса-Рокки, Паола; Салинас, Нельсон Р.; Уиллер, Уорд К. (26 апреля 2013 г.). «Схемы жилкования неотропической голубики (Vaccinieae: Ericaceae) и их филогенетическая полезность» (PDF) . Фитотаксы . 96 (1):1.doi : 10.11646 /phytotaxa.96.1.1 . Архивировано (PDF) 18 февраля. из оригинала Получено 17 февраля.
- Рид, Дж.; Стоукс, А. (1 октября 2006 г.). «Биомеханика растений в экологическом контексте» . Американский журнал ботаники . 93 (10): 1546–1565. дои : 10.3732/ajb.93.10.1546 . ПМИД 21642101 .
- Роллан-Лаган, Анн-Гаэль; Амин, Мира; Пакульска, Малгосия (январь 2009 г.). «Количественная оценка структуры жилкования листьев: двумерные карты». Заводской журнал . 57 (1): 195–205. дои : 10.1111/j.1365-313X.2008.03678.x . ПМИД 18785998 .
- Рот-Небельсик, А; Уль, Дитер; Мосбругер, Волкер; Керп, Ганс (май 2001 г.). «Эволюция и функции архитектуры жилкования листьев: обзор» . Анналы ботаники . 87 (5): 553–566. дои : 10.1006/anbo.2001.1391 .
- Рунионс, Адам; Фюрер, Мартин; Лейн, Брендан; Федерл, Павол; Роллан-Лаган, Анн-Гаэль; Прусинкевич, Пшемыслав (1 января 2005 г.). «Моделирование и визуализация узоров жилкования листьев». Доклады ACM SIGGRAPH 2005 . Том. 24. стр. 702–711. CiteSeerX 10.1.1.102.1926 . дои : 10.1145/1186822.1073251 . ISBN 9781450378253 . S2CID 2629700 .
- Рутисхаузер, Р.; Саттлер, Р. (1997). «Экспрессия побеговых процессов в развитии листьев Polemonium caeruleum ». Ботанические ежегодники по систематике . 119 :563-582.
- Сак, Лоурен; Скоффони, Кристина (июнь 2013 г.). «Жилкование листьев: строение, функции, развитие, эволюция, экология и применение в прошлом, настоящем и будущем» . Новый фитолог . 198 (4): 983–1000. дои : 10.1111/nph.12253 . ПМИД 23600478 .
- Шелли, Эй Джей; Смит, ВК; Фогельманн, TC (1998). «Онтогенетические различия в структуре мезофилла и распределении хлорофилла у Eucalyptus globulus ssp. globulus (Myrtaceae)» . Американский журнал ботаники . 86 (2): 198–207. дои : 10.2307/2656937 . JSTOR 2656937 . ПМИД 21680359 .
- Цукая, Хирокадзу (январь 2013 г.). «Развитие листьев» . Книга «Арабидопсис» . 11 : e0163. дои : 10.1199/tab.0163 . ПМЦ 3711357 . ПМИД 23864837 .
- Уэно, Осаму; Кавано, Юкико; Вакаяма, Масатака; Такеда, Томосиро (1 апреля 2006 г.). «Сосудистые системы листьев трав C 3 и C 4 : двумерный анализ» . Анналы ботаники . 97 (4): 611–621. дои : 10.1093/aob/mcl010 . ПМЦ 2803656 . ПМИД 16464879 .
- Уоллс, РЛ (25 января 2011 г.). «Модели жилок листьев покрытосеменных связаны с функциями листьев в наборе данных глобального масштаба». Американский журнал ботаники . 98 (2): 244–253. дои : 10.3732/ajb.1000154 . ПМИД 21613113 .
Веб-сайты
[ редактировать ]- Букш, Александр; Блондер, Бенджамин; Прайс, Чарльз; Винг, Скотт; Вайц, Джошуа; Дас, Абхирам (2017). «Очищенная база данных изображений листьев» . Школа биологии Технологического института Джорджии . Архивировано из оригинала 25 сентября 2014 года . Проверено 12 марта 2017 г.
- Женева, Роберт. «Лист» (PDF) . PLS 220: Введение в идентификацию растений . Университет Кентукки: факультет садоводства. Архивировано из оригинала (PDF) 15 марта 2016 г.
- Клинг, Гэри Дж.; Хайден, Лаура Л.; Поттс, Джошуа Дж. (2005). «Ботаническая терминология» . Университет Иллинойса , Урбана-Шампейн. Архивировано из оригинала 8 марта 2017 года . Проверено 7 марта 2017 г.
- де Кок, Рожье; Биффин, Эд (ноябрь 2007 г.). «The Pea Key: интерактивный ключ к австралийским бобовым с горошком» . Австралийская исследовательская группа по бобовым с цветками гороха. Архивировано из оригинала 26 февраля 2017 года . Проверено 9 марта 2017 г.
- Кранц, Лаура. «Жилочные узоры листьев» (рисунки) . Архивировано из оригинала 6 марта 2017 г. Проверено 5 марта 2017 г.
- Мэсси, Джимми Р.; Мерфи, Джеймс К. (1996). «Систематика сосудистых растений» . НК-ботнет . Университет Северной Каролины в Чапел-Хилл. Архивировано из оригинала 17 января 2016 года . Проверено 19 января 2016 г.
- "Листья" . Архивировано из оригинала 25 июля 2016 г. Проверено 19 января 2016 г. , в Мэсси и Мерфи (1996)
- Перселл, Адам (16 января 2016 г.). "Листья" . Базовая биология . Адам Перселл. Архивировано из оригинала 19 апреля 2015 года . Проверено 17 февраля 2017 г. .
- Симпсон, Майкл Г. «Растения округа Сан-Диего, Калифорния» . Колледж наук Государственного университета Сан-Диего . Архивировано из оригинала 3 марта 2017 года . Проверено 2 марта 2017 г.
- «Кнопка листьев ископаемых пластов Флориссана» (PDF) . Национальный памятник «Ископаемые пласты Флориссана» . национальных парков Служба Министерства внутренних дел США . Архивировано (PDF) из оригинала 16 февраля 2017 года . Проверено 16 февраля 2017 г.
- «База данных растений» . Факультет садоводства Квантленского политехнического университета . 2015. Архивировано из оригинала 21 сентября 2017 года . Проверено 9 марта 2017 г.
- «Морфология покрытосеменных» . TutorVista. 2017. Архивировано из оригинала 21 июня 2020 г. Проверено 9 марта 2017 г.
- Глоссарии
- Хьюз, Колин. «Виртуальный полевой гербарий» . Гербарии Оксфордского университета . Архивировано из оригинала 5 марта 2017 года . Проверено 4 марта 2017 г.
- «Характеристика растений» . Архивировано из оригинала (Глоссарий) 5 марта 2017 года . Проверено 4 марта 2017 г. , в Хьюзе (2017)
- «Словарь ботанических терминов» . Неотропики . Королевский ботанический сад, Кью. Архивировано из оригинала 21 января 2017 года . Проверено 18 февраля 2017 г.
- «Иллюстрированный словарь форм листьев» (PDF) . Центр водных и инвазивных растений, Институт пищевых и сельскохозяйственных наук , Университет Флориды. 2009. Архивировано (PDF) из оригинала 10 января 2020 года . Проверено 8 января 2020 г.
- «Листья» . Донсгарден . Архивировано из оригинала 4 февраля 2016 года . Проверено 9 января 2020 г.
Внешние ссылки
[ редактировать ]- Рендл, Альфред Бартон (1911). . Британская энциклопедия . Том. 15 (11-е изд.). п. 322-329.
- Ингерсолл, Эрнест (1920). . Американская энциклопедия . Том. XVII.