Синтаза жирной кислоты
| Синтаза жирной кислоты | |||
|---|---|---|---|
| Идентификаторы | |||
| ЕС №. | 2.3.1.85 | ||
| CAS №. | 9045-77-6 | ||
| Базы данных | |||
| Intenz | Intenz View | ||
| Бренда | Бренда вход | ||
| Расширение | Вид Nicezyme | ||
| Кегг | Кегг вход | ||
| Метатический | Метаболический путь | ||
| Напрямую | профиль | ||
| PDB Структуры | RCSB PDB PDBE PDBSUM | ||
| Джин Онтология | Друг / Quickgo | ||
| |||
| Синтаза жирной кислоты | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | жирные кислоты синтазы | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | GeneCards : [1] ; OMA : - ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викидид | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
Синтаза жирной кислоты ( FA ) [ 1 ] это фермент , который у людей кодируется FASN геном . [ 2 ] [ 3 ] [ 4 ] [ 5 ]
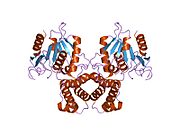
Синтаза жирных кислот представляет собой мультинзимный белок , который катализирует синтез жирных кислот . Это не единый фермент , а целая ферментативная система, состоящая из двух идентичных многофункциональных полипептидов 272 кДа , в которых субстраты передаются из одного функционального домена к следующему. [ 1 ] [ 6 ] [ 7 ] [ 8 ] [ 9 ]
Его основная функция-катализировать синтез пальмитата (C16: 0, длинноцепочечной насыщенной жирной кислоты ) из ацетил-КоА и малонил-КоА , в присутствии NADPH . [ 5 ]
Жирные кислоты синтезируются серией декарбоксилирующих реакций Claisen Condensation от ацетил-КоА и малонил-КоА . После каждого раунда удлинения группа бета -кето уменьшается до полностью насыщенной углеродной цепи последовательным действием кеторедуктазы ( KR), дегидратазы (DH) и эноилредуктазы (ER). Растущая цепь жирных кислот переносится между этими активными участками, в то время как прикрепляется ковалентно к протезной группе фосфопантетеиина ацильного белка (ACP) и высвобождается действием тиоэстеразы ( TE) при достижении длины углеродной цепи 16 (Palmitic кислота). [ 1 ]
Классы
[ редактировать ]Есть два основных класса синтаз жирных кислот.
- Системы типа используют один большой многофункциональный полипептид и являются общими как для животных , так и для грибов (хотя структурное расположение грибковых и синтезов животных различается). Система синтазы жирных кислот I типа также обнаружена в группе бактерий CMN ( Corynebacteria , Mycobacteria и Nocardia ). В этих бактериях система Fas I производит пальмитиновую кислоту и сотрудничает с системой Fas II для получения большего разнообразия липидных продуктов. [ 10 ]
- Тип II содержится в археи, бактериях и растительных пластидах и характеризуется использованием дискретных монофункциональных ферментов для синтеза жирных кислот. Ингибиторы этого пути (FASII) исследуются как возможные антибиотики . [ 11 ]
Механизм удлинения и восстановления Fas I и Fas II одинаков, как и домены ферментов FAS II в значительной степени гомологичны их доменным аналогам в полипептидах Fas I. Тем не менее, различия в организации ферментов - интегрированные в Fas I, дискретные в Fas II - дают многие важные биохимические различия. [ 12 ]
Эволюционная история синтаз жирных кислот очень сильно взаимосвязана с историей поликетид -синтаз (PKS). Поликетид -синтазы используют аналогичный механизм и гомологичные домены для получения вторичных метаболитных липидов. Кроме того, поликетид -синтазы также демонстрируют организацию типа I и типа II. Считается, что FAS I у животных возникли благодаря модификации PKS I в грибах, тогда как Fas I в грибах и группа бактерий CMN, по -видимому, возникли отдельно через слияние генов Fas II. [ 10 ]
Структура
[ редактировать ]ФАС млекопитающих состоит из гомодимера двух идентичных белковых субъединиц, в которых три каталитические домены в N-концевой сечении (-ккетоацилсинтаза (KS), малонил/ацетилтрансфераза (MAT) и дегидраза (DH)) разделяются ядро (MAT) и дегидраза (DH)) Область (известная как междомен) из 600 остатков из четырех С -концевых доменов (эноилредуктаза (ER), -ккетоацилредуктаза (KR), белка ацила -носителя (ACP) и тиоэстеразы (TE)). [ 13 ] [ 14 ] Междоменная область позволяет двум мономерным доменам сформировать димер. [ 13 ]
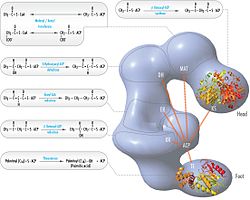
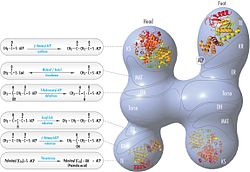
Обычная модель для организации FAS (см. Модель «головы к хвостам» справа) в значительной степени основана на наблюдениях о том, что бифункциональный реагент 1,3-дибромпропанон (DBP) способен скрещивать цистеин активного сайта тиол Домен KS в одном мономере FAS с фосфопантэцетейной протезированной группой домена ACP в другом мономере. [ 15 ] [ 16 ] Анализ комплементации димеров FA, несущих различные мутации на каждом мономере, установил, что домены KS и MAT могут сотрудничать с ACP любого мономера. [ 17 ] [ 18 ] и реинвестирование экспериментов по сшивке DBP показало, что тиол активного сайта KS может быть сшит с ACP 4'- фосфопантиином любого мономера. [ 19 ] Кроме того, недавно сообщалось, что гетеродимерный FA, содержащий только один компетентный мономер, способен к синтезу пальмитата. [ 20 ]
Вышеуказанные наблюдения казались несовместимыми с классической моделью «головы к хвостам» для организации FAS, и была предложена альтернативная модель, предсказывающая, что KS и MAT-домены обоих мономеров находятся ближе к центру димера Fas, где они может получить доступ к ACP любой субъединицы (см. Рисунок в правом верхнем углу). [ 21 ]
Рентгеновская кристаллографическая структура с низким разрешением обеих свиней (гомодимер) [ 22 ] и дрожжи Fas (гетерододекамер) [ 23 ] Наряду с разрешением электронной крио-микроскопии ~ 6 Å (крио-эм) структура FAS FAS [ 24 ] были решены.
Механизм шаттласа субстрата
[ редактировать ]Решенные структуры дрожжевых FA и FAS млекопитающих показывают две отдельные организации высококонсервативных каталитических доменов/ферментов в этой мультинциторной клеточной машине. Дрожжевые FAS имеют высокоэффективную жесткую бочкообразную структуру с 6 реакционными камерами, которые независимо синтезируют жирные кислоты, в то время как Fas млекопитающих имеет открытую гибкую структуру только с двумя реакционными камерами. Однако в обоих случаях консервативный ACP действует как мобильная область, ответственная за то, чтобы перекрывать субстраты промежуточных жирных кислот в различные каталитические участки. Первое прямое структурное понимание этого механизма шаттлата субстрата было получено с помощью крио-эм-анализа, где наблюдается ACP, связанный с различными каталитическими доменами в синтазе дрожжей в форме ствола. [ 24 ] Результаты крио-ЭМ предполагают, что связывание ACP с различными сайтами является асимметричным и стохастическим, что также указывается исследованиями компьютерной имиляции [ 25 ]
 |
 |
Регулирование
[ редактировать ]Метаболизм и гомеостаз синтазы жирной кислоты транскрипционно регулируются с помощью восходящих стимулирующих факторов ( USF1 и USF2 ) и связывающего белка -связывающего элемента стерола -1C (SREBP -1C) в ответ на кормление/инсулин у живых животных. [ 26 ] [ 27 ]
Хотя рецепторы печени X (LXR) модулируют экспрессию белка, связывающего регуляторного элемента, -1C (SREBP-1C) при питании, регуляция FAS с помощью SREBP-1C зависит от USF. [ 27 ] [ 28 ] [ 29 ] [ 30 ]
Ацил -флороглюцинолы, выделенные из папоротниковой косой, обладают ингибирующей активностью синтазы жирной кислоты. [ 31 ]
Клиническое значение
[ редактировать ]Ген FASN был исследован как возможный онкоген . [ 32 ] FAS активируется при раке молочной железы и желудка, а также является показателем плохого прогноза, и поэтому может быть стойким в качестве химиотерапевтической цели. [ 33 ] [ 34 ] [ 35 ] FAS Следовательно, ингибиторы являются активной областью исследований по обнаружению лекарств . [ 36 ] [ 37 ] [ 38 ] [ 39 ] [ 40 ]
FAS также может быть вовлечена в производство эндогенного лиганда для ядерного рецептора PPARALPHA , мишени фибразирующих препаратов для гиперлипидемии, [ 41 ] и исследуется как возможная мишень для лечения метаболического синдрома. [ 42 ] Orlistat , который является желудочно -кишечным ингибитором липазы, также ингибирует Fas и имеет потенциал в качестве лекарства от рака . [ 43 ] [ 44 ]
Было обнаружено, что в некоторых раковых клеточных линиях этот белок сливается с альфа-рецептором эстрогена (ER-альфа), на котором N-конце Fas сливается в рамке с C-конце ER-альфа. [ 5 ]
об ассоциации с лейомиомата матки . Сообщалось [ 45 ]
Смотрите также
[ редактировать ]- Открытие и развитие ингибиторов желудочно -кишечной липазы
- Синтез жирных кислот
- Метаболизм жирных кислот
- Деградация жирных кислот
- Enoyl-acyl-белок-перевозчик редуктаза
- Список нарушений метаболизма жирных кислот
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в Paiva P, Medina FE, Viegas M, Ferreira P, Neves RP, Sousa JP, Ramos MJ, Fernandes PA (2021-08-11). «Синтаза животных жировой аварии: химическая нанофабрика» . Химические обзоры . 121 (15): 9502–9553. Doi : 10.1021/acs.chemrev.1c00147 . ISSN 0009-2665 . PMID 34156235 . S2CID 235595027 .
- ^ Jayakumar A, Chirala SS, Chinault AC, Baldini A, Abu-Elheiga L, Wakil SJ (февраль 1995 г.). «Выделение и хромосомное картирование геномных клонов, кодирующих ген синтазы жирной кислоты человека». Геномика . 23 (2): 420–424. doi : 10.1006/geno.1994.1518 . PMID 7835891 .
- ^ Jayakumar A, Tai MH, Huang Wy, Al-Feel W, Hsu M, Abu-Elega L, Chirala SS, Wakil SJ (октябрь 1995). «Синтаза жирной кислоты человека: свойства и молекулярное клонирование» . Труды Национальной академии наук Соединенных Штатов Америки . 92 (19): 8695–8699. Bibcode : 1995pnas ... 92.8695j . doi : 10.1073/pnas.92.19.8695 . PMC 41033 . PMID 7567999 .
- ^ Persson B, Kallberg Y, Bray Je, Bruford E, Dellaporta SL, Favia AD, Duarte RG, Jörnvall H, Kavanagh KL, Kedishvili N, Kisiela M, Maser E, Mindnich R, Orchard S, Penning TM, Thornton JM, Adamski J, Mindnich R, Orchard S, Penning TM, Thornton JM, Adamski J, Mindnich R, Orchard S, Penning TM, Thornton JM, Adamski J, Orchard , Oppermann U (февраль 2009 г.). «Инициатива SDR (короткоцепочечная дегидрогеназа/редуктаза и родственные ферменты)» . Химико-биологические взаимодействия . 178 (1–3): 94–98. Bibcode : 2009cbi ... 178 ... 94p . doi : 10.1016/j.cbi.2008.10.040 . PMC 2896744 . PMID 19027726 .
- ^ Jump up to: а беременный в «Ген Entrez: синтаза FASN жирной кислоты» .
- ^ Alberts AW, Strauss AW, Hennessy S, Wargelos PR (октябрь 1975 г.). «Регуляция синтеза печеночной жирной кислоты синтетазы: связывание антител синтетазы жирной кислоты с полисомами» . Труды Национальной академии наук Соединенных Штатов Америки . 72 (10): 3956–3960. Bibcode : 1975pnas ... 72.3956a . doi : 10.1073/pnas.72.10.3956 . PMC 433116 . PMID 1060077 .
- ^ Ступс Дж.К., Арслан М.Дж., О.Ю., Аун К.С., Ванаман Т.С., Вакиль С.Дж. (май 1975). «Присутствие двух полипептидных цепей, содержащих синтетазу жирных кислот» . Труды Национальной академии наук Соединенных Штатов Америки . 72 (5): 1940–1944. Bibcode : 1975pnas ... 72.1940S . doi : 10.1073/pnas.72.5.1940 . PMC 432664 . PMID 1098047 .
- ^ Смит С., Агради Э., Либертини Л., Дилипан КН (апрель 1976 г.). «Специфическое высвобождение компонента тиоэстеразы мультиферментного комплекса с жирной кислотой с ограниченной трипсинизацией» . Труды Национальной академии наук Соединенных Штатов Америки . 73 (4): 1184–1188. Bibcode : 1976pnas ... 73.1184S . doi : 10.1073/pnas.73.4.1184 . PMC 430225 . PMID 1063400 .
- ^ Смит С., Витковски А., Джоши А.К. (июль 2003 г.). «Структурная и функциональная организация синтазы животных жирных кислот». Прогресс в исследованиях липидов . 42 (4): 289–317. doi : 10.1016/s0163-7827 (02) 00067-x . PMID 12689621 .
- ^ Jump up to: а беременный Дженке-Кодама Х., Сэндманн А., Мюллер Р., Диттманн Е (октябрь 2005 г.). «Эволюционные последствия бактериальных поликетовых синтаз» . Молекулярная биология и эволюция . 22 (10): 2027–2039. doi : 10.1093/molbev/msi193 . PMID 15958783 .
- ^ Фулмер Т (март 2009 г.). «Не так фэс» . Научный бизнес обмен . 2 (11): 430. doi : 10.1038/scibx.2009.430 .
- ^ Стивенс Л., Прайс Н.К. (1999). Основы фермеры: клеточная и молекулярная биология каталитических белков . Оксфорд [Оксфордшир]: издательство Оксфордского университета. ISBN 978-0-19-850229-6 .
- ^ Jump up to: а беременный Chirala SS, Jayakumar A, Gu Zw, Wakil SJ (март 2001 г.). «Синтаза жирной кислоты человека: роль междонавого в формировании каталитически активной синтазы димера» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (6): 3104–3108. Bibcode : 2001pnas ... 98.3104C . doi : 10.1073/pnas.051635998 . PMC 30614 . PMID 11248039 .
- ^ Смит С (декабрь 1994 г.). «Синтаза животных жирных кислот: один ген, один полипептид, семь ферментов» . FASEB Journal . 8 (15): 1248–1259. doi : 10.1096/fasebj.8.15.8001737 . PMID 8001737 . S2CID 22853095 .
- ^ Ступс Дж.К., Вакил С.Дж. (май 1981). «Синтетаза с жирной кислотой животных. Новое расположение бета-кетоацил-синтетазы, содержащих домены двух субъединиц» . Журнал биологической химии . 256 (10): 5128–5133. doi : 10.1016/s0021-9258 (19) 69376-2 . PMID 6112225 .
- ^ STOOPS JK, Wakil SJ (март 1982 г.). «Синтетаза с жирной кислотой животных. Идентификация остатков, включающих новое расположение бета-кетоацил-синтетазы и их роль в его холодной инактивации» . Журнал биологической химии . 257 (6): 3230–3235. doi : 10.1016/s0021-9258 (19) 81100-6 . PMID 7061475 .
- ^ Джоши А.К., Ранган против, Смит С (февраль 1998 г.). «Дифференциальная аффинная маркировка двух субъединиц гомодимерной животной синтазы жирной кислоты позволяет выделять гетеродимеры, состоящие из субъединиц, которые были независимо модифицированы» . Журнал биологической химии . 273 (9): 4937–4943. doi : 10.1074/jbc.273.9.4937 . PMID 9478938 .
- ^ Ранган В.С., Джоши А.К., Смит С. (сентябрь 2001 г.). «Картирование функциональной топологии синтазы жирной кислоты животных с помощью комплементации мутантов in vitro». Биохимия . 40 (36): 10792–18799. doi : 10.1021/bi015535z . PMID 11535054 .
- ^ Витковски А., Джоши А.К., Ранган В.С., Флик А.М., Витковска Хе, Смит С. (апрель 1999 г.). «Сшивание дибромопропанона фосфовантететеина и активных ситовых тиолов синтазы животных жирных кислот может происходить как между и внутрисубнитом. Переоценка бок по бок, антипараллельная модель субъединицы» . Журнал биологической химии . 274 (17): 11557–11563. doi : 10.1074/jbc.274.17.11557 . PMID 10206962 .
- ^ Джоши А.К., Ранган В.С., Витковски А., Смит С. (февраль 2003 г.). «Инженерная инженерия активного димера синтазы жирной кислоты животных только с одной компетентной субъединицей» . Химия и биология . 10 (2): 169–173. doi : 10.1016/s1074-5521 (03) 00023-1 . PMID 12618189 .
- ^ Asturias FJ, Chadick JZ, Cheung Ik, Stark H, Witkowski A, Joshi Ak, Smith S (март 2005 г.). «Структура и молекулярная организация синтазы жирных кислот млекопитающих». Природа структурная и молекулярная биология . 12 (3): 225–232. doi : 10.1038/nsmb899 . PMID 15711565 . S2CID 6132878 .
- ^ Maier T, Leibundgut M, Ban N (сентябрь 2008 г.). «Кристаллическая структура синтазы жирной кислоты млекопитающих». Наука . 321 (5894): 1315–1322. Bibcode : 2008Sci ... 321.1315m . doi : 10.1126/science.1161269 . PMID 18772430 . S2CID 3168991 .
- ^ Lomakin IB, Xiong Y, Steitz Ta (апрель 2007 г.). «Кристаллическая структура дрожжевой жирной кислоты синтазы, клеточной машины с восемью активными участками, работающими вместе» . Клетка . 129 (2): 319–332. doi : 10.1016/j.cell.2007.03.013 . PMID 17448991 . S2CID 8209424 .
- ^ Jump up to: а беременный Gipson P, Mills DJ, Wouts R, Grininger M, Vonck J, Kühlbrandt W (май 2010). «Прямое структурное понимание механизма подложки синтазы жирных кислот дрожжей с помощью электронной криоомкроскопии» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (20): 9164–9169. Bibcode : 2010pnas..107.9164G . doi : 10.1073/pnas.0913547107 . PMC 2889056 . PMID 20231485 .
- ^ Anselmi C, Grininger M, Gipson P, Faraldo-Gómez JD (сентябрь 2010 г.). «Механизм шаттлата субстрата белком ацил-карьера в мега-синтазе жирной кислоты». Журнал Американского химического общества . 132 (35): 12357–12364. doi : 10.1021/ja103354w . PMID 20704262 .
- ^ Paulauskis JD, Sul HS (январь 1989 г.). «Гормональная регуляция транскрипции гена синтазы мыши жирной кислоты в печени» . Журнал биологической химии . 264 (1): 574–577. doi : 10.1016/s0021-9258 (17) 31298-х . PMID 2535847 .
- ^ Jump up to: а беременный Latasa MJ, Griffin MJ, Moon YS, Kang C, Sul HS (август 2003 г.). «Занятость и функция регуляторного элемента -150 стерола и -65 E -Box в регуляции питательных веществ жирового гена синтазы у живых животных» . Молекулярная и клеточная биология . 23 (16): 5896–5907. doi : 10.1128/mcb.23.16.5896-5907.2003 . PMC 166350 . PMID 12897158 .
- ^ Griffin MJ, Wong RH, Pandya N, Sul HS (февраль 2007 г.). «Прямое взаимодействие между USF и SREBP-1C опосредует синергетическую активацию промотора жирной синтазы» . Журнал биологической химии . 282 (8): 5453–5467. doi : 10.1074/jbc.m610566200 . PMID 17197698 .
- ^ Йошикава Т., Шимано Х., Амемия-Кудо М., Яхаги Н., поспешная А.Х., Мацузака Т., Оказаки Х, Тамура Ю., Иидзука Ю., Охаши К, Осуга Дж, Харада К, Готода Т., Кимура С., Иибаши С., Ямада Н. Май 2001 г.). «Идентификация рецептора x рецептора печени Retinoid X в качестве активатора промотора гена гена 1C-связывающего белка, связывающего белок, связывающего стерол» . Молекулярная и клеточная биология . 21 (9): 2991–3000. doi : 10.1128/mcb.21.9.2991-3000.2001 . PMC 86928 . PMID 11287605 .
- ^ Repa JJ, Liang G, Ou J, Bashmakov Y, Lobaccaro JM, Shimomura I, Shan B, Brown MS, Goldstein JL, Mangelsdorf DJ (ноябрь 2000 г.). «Регуляция мышиного стерола-регуляторного элемента связывающего белка-1C (SREBP-1C) с помощью оксистерольных рецепторов, Lxralpha и Lxrbeta» . Гены и развитие . 14 (22): 2819–2830. doi : 10.1101/gad.844900 . PMC 317055 . PMID 11090130 .
- ^ Na M, Jang J, Min BS, Lee SJ, Lee MS, Kim By, Oh Wk, Ahn JS (сентябрь 2006 г.). «Ингибирующая активность жирной кислоты ацилфлорлюцинолов, выделенных из суроговой кризирхизомы». Биоорганические и лекарственные химические письма . 16 (18): 4738–4742. doi : 10.1016/j.bmcl.2006.07.018 . PMID 16870425 .
- ^ Барон А., Мигита Т., Тан Д., Лода М (январь 2004 г.). «Синтаза жирной кислоты: метаболический онкоген при раке простаты?». Журнал сотовой биохимии . 91 (1): 47–53. doi : 10.1002/jcb.10708 . PMID 14689581 . S2CID 26175683 .
- ^ Хант Д.А., Лейн Х.М., Зигмонт М.Е., Дерван П.А., Хеннигар Р.А. (2007). «Стабильность мРНК и сверхэкспрессия синтазы жирной кислоты в клеточных линиях рака молочной железы человека». Противоопухолевые исследования . 27 (1a): 27–34. PMID 17352212 .
- ^ Ганслер Т.С., Хардман В., Хант Д.А., Шаффель С., Хеннигар Р.А. (июнь 1997 г.). «Повышенная экспрессия синтазы жирных кислот (OA-519) в неоплазмах яичников предсказывает более короткую выживаемость». Человеческая патология . 28 (6): 686–692. doi : 10.1016/s0046-8177 (97) 90177-5 . PMID 9191002 .
- ^ Ezzeddini R, Taghikhani M, Somi MH, Samadi N, Rasaee, MJ (май 2019). «Клиническая важность FASN в отношении HIF-1α и SREBP-1C при аденокарциноме желудка» . Жизненные науки . 224 : 169–176. doi : 10.1016/j.lfs.2019.03.056 . PMID 30914315 . S2CID 85532042 .
- ^ «Первое исследование человека, проходящее с ингибитором синтазы жирной кислоты» . oncotherapynetwork.com. 7 апреля 2017 года. Архивировано с оригинала 15 апреля 2019 года . Получено 4 июня 2018 года .
- ^ Lu T, Schubert C, Cummings MD, Bignan G, Connolly PJ, Smans K, Ludovici D, Parker MH, Meyer C, Rocaboy C, Alexander R, Grasberger B, De Breucker S, Esser N, Fraiponts E, Gilissen R, Janssens B, Peeters D, Van Nuffel L, Vermeulen P, Bischoff J, Meerpoel L (май 2018). «Дизайн и синтез серии ингибиторов домена домена из биодоступных жирных кислот (FASN) для терапии рака». Биоорганические и лекарственные химические письма . 28 (12): 2159–2164. doi : 10.1016/j.bmcl.2018.05.014 . PMID 29779975 . S2CID 29159508 .
- ^ Hardwicke MA, Rendina AR, Williams SP, Moore ML, Wang L, Krueger JA, Plant RN, Totoritis RD, Zhang G, Briand J, Burkhart WA, Brown KK, Parrish CA (сентябрь 2014 г.). «Ингибитор синтазы жирной кислоты человека связывает β-кетоацилредуктазу в сайте кето-субстрата». Природная химическая биология . 10 (9): 774–779. doi : 10.1038/nchembio.1603 . PMID 25086508 .
- ^ Вандер Хейден М.Г., Дебердинис Р.Дж. (февраль 2017 г.). «Понимание пересечений между метаболизмом и биологией рака» . Клетка . 168 (4): 657–669. doi : 10.1016/j.cell.2016.12.039 . PMC 5329766 . PMID 28187287 .
- ^ SGRO CD (2009-01-01). Исследование междоменной области синтазы жирных кислот Caenorhabditis elegans и ее последствий в качестве лекарственной мишени (тезис диссертации). La Trobe.
- ^ Чакраварти М.В., Лодхи И.Дж., Инь Л., Малапака Р.Р., Сюй Х. Х., Турк Дж., Семенкович К.Ф. (август 2009 г.). «Выявление физиологически значимого эндогенного лиганда для Pparalpha в печени» . Клетка . 138 (3): 476–488. doi : 10.1016/j.cell.2009.05.036 . PMC 2725194 . PMID 19646743 .
- ^ Wu M, Singh SB, Wang J, Chung CC, Salituro G, Karanam BV, Lee SH, Powles M, Ellsworth KP, Lassman ME, Miller C, Myers RW, Tota MR, Zhang BB, Li C (март 2011 г.). «Антидиабетические и антистеатические эффекты ингибитора селективной жирной кислоты (FAS) Platensimycin на мышиных моделях диабета» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (13): 5378–5383. Bibcode : 2011pnas..108.5378W . doi : 10.1073/pnas.1002588108 . PMC 3069196 . PMID 21389266 .
- ^ Flavin R, Peluso S, Nguyen PL, Loda M (апрель 2010 г.). «Синтаза жирной кислоты как потенциальная терапевтическая мишень при раке» . Будущая онкология . 6 (4): 551–562. doi : 10.2217/fon.10.11 . PMC 3197858 . PMID 20373869 .
- ^ Richardson Rd, Ma G, Oyola Y, Zancanella M, Knowles LM, Cieplak P, Romo D, Smith JW (сентябрь 2008 г.). «Синтез новых бета-лактоновых ингибиторов синтазы жирных кислот» . Журнал лекарственной химии . 51 (17): 5285–5296. doi : 10.1021/jm800321h . PMC 3172131 . PMID 18710210 .
- ^ Rose L, Heath AC, Treloar PM, Rose L, Buring JE, Ridker PM, Ridker PM. "Weom-Willa-это анализ PASN . Журнал человеческой генетики 91 (4): 621–628. doi : 10.1016/jhg.2012.08.009 . PMC 3484658 . PMID 23040493 .
Дальнейшее чтение
[ редактировать ]- Wakil SJ (1989). «Синтаза жирной кислоты, опытный многофункциональный фермент». Биохимия . 28 (11): 4523–4530. doi : 10.1021/bi00437a001 . PMID 2669958 .
- Барон А., Мигита Т., Тан Д., Лода М. (2004). «Синтаза жирной кислоты: метаболический онкоген при раке простаты?». Журнал сотовой биохимии . 91 (1): 47–53. doi : 10.1002/jcb.10708 . PMID 14689581 . S2CID 26175683 .
- Летен Д. (1978). «[Вискозиметрия в клинической практике]». Медицинский предварительный . 30 (9–10): 477–482. PMID 600212 .
- Wronkowski Z (1976). «[Диагноз рака респираторной системы]». Pielȩgniarka I Połozna (12): 7–8. PMID 1044453 .
- Семенкович CF, Coleman T, Fiedorek FT (1995). «МРНК жирной кислоты человека синтазы: распределение тканей, генетическое картирование и кинетика распада после депривации глюкозы» . Журнал липидных исследований . 36 (7): 1507–1521. doi : 10.1016/s0022-2275 (20) 39738-8 . PMID 7595075 .
- Kuhajda FP, Jenner K, Wood FD, Hennigar RA, Jacobs LB, Dick JD, Pasternack GR (1994). «Синтез жирных кислот: потенциальная селективная мишень для противоопухолевой терапии» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (14): 6379–6383. Bibcode : 1994pnas ... 91.6379K . doi : 10.1073/pnas.91.14.6379 . PMC 44205 . PMID 8022791 .
- HSU MH, Chirala SS, Wakil SJ (1996). «Жирная синтаза человека . Журнал биологической химии . 271 (23): 13584–13592. doi : 10.1074/jbc.271.23.13584 . PMID 8662758 .
- Pizer ES, Kurman RJ, Pasternack GR, Kuhajda FP (1997). «Экспрессия синтазы жирных кислот тесно связана с пролиферацией и стромальной децидуализацией в велосипедной эндометрии». Международный журнал гинекологической патологии . 16 (1): 45–51. doi : 10.1097/00004347-199701000-00008 . PMID 8986532 . S2CID 45195801 .
- Jayakumar A, Chirala SS, Wakil SJ (1997). «Синтаза жирной кислоты человека: сборка рекомбинантных половинок белка белка субъединицы жирной кислоты реконструкция ферментов» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (23): 12326–12330. Bibcode : 1997pnas ... 9412326J . doi : 10.1073/pnas.94.23.12326 . PMC 24928 . PMID 9356448 .
- Кусакабе Т., Маэда М., Хоши Н., Сугино Т., Ватанабе К., Фукуда Т., Сузуки Т. (2000). «Синтаза жирных кислот экспрессируется в основном в чувствительных к гормонам клеткам или клеткам с высоким липидным метаболизмом с высоким липидным метаболизмом и в пролиферирующих клетках плода» . Журнал гистохимии и цитохимии . 48 (5): 613–622. doi : 10.1177/002215540004800505 . PMID 10769045 .
- Ye Q, Chung LW, Li S, Zhau HE (2000). «Идентификация нового транскрипта слияния Fas/ER-альфа, экспрессируемого в раковых клетках человека». Biochimica et Biophysica Acta (BBA) - структура и экспрессия гена . 1493 (3): 373–377. doi : 10.1016/s0167-4781 (00) 00202-5 . PMID 11018265 .
- Rochat-Steiner V, Becker K, Micheau O, Schneider P, Burns K, Tschopp J (2000). «FIST/HIPK3: FAS/FADD-взаимодействие сериновой/треонинкиназы, которая индуцирует фосфорилирование FADD и ингибирует Fas-опосредованную активацию киназы Jun NH (2)» . Журнал экспериментальной медицины . 192 (8): 1165–1174. doi : 10.1084/jem.192.8.1165 . PMC 2311455 . PMID 11034606 .
- Chirala SS, Jayakumar A, Gu ZW, Wakil SJ (2001). «Синтаза жирной кислоты человека: роль междонавого в формировании каталитически активной синтазы димера» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (6): 3104–3108. Bibcode : 2001pnas ... 98.3104C . doi : 10.1073/pnas.051635998 . PMC 30614 . PMID 11248039 .
- Бринк Дж., Ладтке С.Дж., Ян Сай, Гу З.В., Вакил С.Дж., Чиу В. (2002). «Четвертая структура синтазы жирных кислот человека с помощью электронной криомикроскопии» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (1): 138–143. Bibcode : 2002pnas ... 99..138b . doi : 10.1073/pnas.012589499 . PMC 117528 . PMID 11756679 .
- Джозеф С.Б., Лаффитт Б.А., Патель П.Х., Уотсон М.А., Мацукума К.Е., Уолчак Р., Коллинз Дж. Л., Осборн Т.Ф., Тонтонос П. (2002). «Прямые и косвенные механизмы регуляции экспрессии гена синтазы жирной кислоты рецепторами печени X» . Журнал биологической химии . 277 (13): 11019–11025. doi : 10.1074/jbc.m111041200 . PMID 11790787 .
- Ming D, Kong Y, Wakil SJ, Brink J, Ma J (2002). «Доменные движения в синтазе жирных кислот человека с помощью квантовой упругих деформационных модели» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (12): 7895–7899. Bibcode : 2002pnas ... 99,7895M . doi : 10.1073/pnas.112222299 . PMC 122991 . PMID 12060737 .
- Field FJ, Born E, Murthy S, Mathur SN (2003). «Полиненасыщенные жирные кислоты уменьшают экспрессию регуляторного белка-1 с стеролом в клетках Caco-2: влияние на синтез жирных кислот и транспорт триацилглицерина» . Биохимический журнал . 368 (Pt 3): 855–864. doi : 10.1042/bj20020731 . PMC 1223029 . PMID 12213084 .
Внешние ссылки
[ редактировать ]- Жирная+кислота+синтаза в Национальной библиотеке медицинской библиотеки Медицинской библиотеки США (Mesh)
- Синтез жирных кислот: Политехнический институт Ренсселера
- Синтаза жирной кислоты: RCSB PDB-молекула месяца Архивирована 2014-07-14 на машине Wayback
- 3D -электронная микроскопия структуры синтазы жирной кислоты из банка данных EM (EMDB)
- PDBE-KB предоставляет обзор всей информации о структуре, доступной в PDB для синтазы жирной кислоты человека
PDB Галерея |
|---|