Papillomaviridae
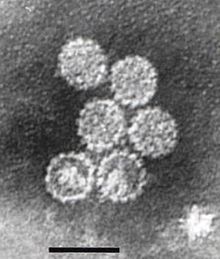
Papillomaviridae- это семейство не окруженных вирусов ДНК , члены которых известны как папилломавирусы. [ 1 ] Несколько сотен видов папилломы, традиционно называемых «типами», [ 2 ] были идентифицированы заражение всех тщательно проверенных млекопитающих, [ 2 ] но также и другие позвоночные, такие как птицы, змеи, черепахи и рыба. [ 3 ] [ 4 ] [ 5 ] Инфекция большинством типов папилломы, в зависимости от типа, является либо бессимптомной (например, большинство бета-ПВ), либо вызывает небольшие доброкачественные опухоли, известные как папилломы или бородавки (например, папилломавирус человека 1, HPV6 или HPV11). Однако папилломы, вызванные некоторыми типами, такими как папилломавирусы человека 16 и 18, несут риск стать раком . [ 6 ]
Папилломавирусы обычно считаются очень хозяином и тканями , и, как полагают, редко передаются между видами. [ 7 ] Папилломавирусы повторяются исключительно в базальном слое поверхностных тканей тела . Все известные типы папилломы заражают определенную поверхность тела, [ 2 ] Обычно эпителий кожи или слизистой оболочки гениталий, заднего прохода, рта или дыхательных путей. [ 8 ] Например, человеческий папилломавирус (ВПЧ) типа 1 имеет тенденцию заражать подошвы ног, а тип 2 типа ВПЧ - ладони рук, где они могут вызвать бородавки . Кроме того, существуют описания присутствия ДНК папилломы в крови и в мононуклеарных клетках периферической крови .
Папилломавирусы были впервые идентифицированы в начале 20 -го века, когда было показано, что кожные бородавки или папилломы могут передаваться между индивидуумами отфильтруемым инфекционным агентом. В 1935 году Фрэнсис Пейтон Рус , который ранее продемонстрировал существование вызывающего рак вирус саркомы у цыплят, продолжал показать, что папилломавирус может вызвать рак кожи у инфицированных кроликов. Это была первая демонстрация, что вирус может вызвать рак у млекопитающих.
Таксономия папилломы
[ редактировать ]
Распознается более 100 видов папилломы, [ 9 ] Хотя ICTV официально распознает меньшее число, классифицированное на 53 рода, по состоянию на 2019 год. [ 10 ] [ 11 ] [ 12 ] Все папилломавирусы (PVS) имеют сходные геномные организации, а любая пара PV содержит по меньшей мере пять гомологичных генов , хотя нуклеотидная последовательность может расходиться более чем на 50%. Филогенетические алгоритмы , которые позволяют сравнивать гомологии, привели к филогенетическим деревьям , которые имеют сходную топологию, независимо от проанализированного гена. [ 13 ]
Филогенетические исследования убедительно свидетельствуют о том, что PV обычно развиваются вместе с их видами хозяина млекопитающих и птиц, но адаптивные излучения , случайные зоонотические события и рекомбинации также могут повлиять на их диверсификацию. [ 13 ] Их основная геномная организация, по -видимому, поддерживается в течение периода, превышающего 100 миллионов лет, и эти сравнения последовательностей заложили основу для таксономии PV, которая в настоящее время официально признается Международным комитетом по таксономии вирусов . Все PV образуют семейство Papillomaviridae , которое отличается от полиомавиридов, таким образом, исключает термин Papovaridae . Основные ветви филогенетического дерева PV считаются родами , которые идентифицируются греческими буквами. Незначительные ветви считаются видами и объединяют типы PV, которые геномно различаются, не демонстрируя известных биологических различий. Эта новая таксономическая система не влияет на традиционную идентификацию и характеристику PV «типов» и их независимых изолятов с незначительными геномными различиями, называемыми «подтипами» и «вариантами», все они таксоны ниже уровня «видов». [ 14 ] Кроме того, были предложены филогенетические группировки на более высоком таксономическом уровне. [ 15 ]
Эта классификация может потребовать пересмотра в свете существования рекомбинантов вируса папилломы -полиомы. [ 16 ] Дополнительные виды также были описаны. Sparus aurata papillomavirus 1 был изолирован от рыбы. [ 17 ]
Человеческие папилломавирусы
[ редактировать ]Более 170 типов папилломы человека были полностью секвенированы. [ 18 ] Они были разделены на 5 родов: альфапилломавирус, бетапилломавирус, гаммапапилломавирус, мупапилломавирус и нупапилломавирус. Было выявлено не менее 200 дополнительных вирусов, которые ожидают секвенирования и классификации. [ Цитация необходима ]
Животные папилломавирусы
[ редактировать ]
Отдельные типы папилломы, как правило, сильно адаптированы к репликации у одного вида животных. В одном исследовании исследователи провалили кожу лба различных животных зоопарка и использовали ПЦР, чтобы усилить любую папилломавирусную ДНК, которая может присутствовать. [ 19 ] Хотя в исследовании было идентифицировано широкое разнообразие последовательностей папилломы, авторы обнаружили мало доказательств межвидовой передачи. Было обнаружено, что один зоокепер временно положительным для специфической для шимпанзе последовательность папилломы. Тем не менее, авторы отмечают, что специфическая для шимпанзе последовательность папилломавируса могла быть результатом загрязнения поверхности кожи зоокета, в отличие от продуктивной инфекции. [ Цитация необходима ]
Cottontail Rabbit Papillomavirus (CRPV) может вызвать выпуклые бородавки у своего родного хозяина, рода североамериканского кролика Sylvilagus . Эти рогоподобные бородавки могут быть первоначальной основой для городских легенд американского рогового кролика, шакалопа и европейского Вулпертингера . [ 20 ] Европейские домашние кролики (род Oryctolagus ) могут быть временно инфицированы CRPV в лабораторных условиях. Однако, поскольку европейские внутренние кролики не производят инфекционную потомство вируса потомства, они считаются случайным или «тупиковым» хозяином для CRPV. [ 21 ]
Межвидовая передача также была задокументирована для бычьего папилломавируса (BPV) типа 1. [ 22 ] В своем естественном хозяине (крупный рогатый скот) BPV-1 индуцирует большие фиброзные кожные бородавки. BPV-1 Инфекция лошадей, которые являются случайным хозяином для вируса, может привести к развитию доброкачественных опухолей, известных как саркоиды . Сельскохозяйственное значение BPV-1 стимулировало успешные усилия по разработке вакцины против вируса. [ Цитация необходима ]
В нескольких сообщениях выявили папилломавирусы у небольших грызунов, таких как сирийские хомяки , африканская крыса с мультимаматом и евразийская мыша для сбора урожая . [ 23 ] Тем не менее, известно, что папилломавирусы способны заразить лабораторных мышей . Отсутствие модели мышиной мыши для папилломавирусной инфекции стало основным ограничением для лабораторных исследований папилломавирусов. [ Цитация необходима ]
Известно, что четыре папилломавируса заражают птиц, экологические коэлбс Папилломавирус 1, Франколинус Лейкосцепс Папилломавирус 1, попугай Эритак Папилломавирус 1 и Pygoscelis Adeliae Papillomavirus 1. [ 24 ] Все эти виды имеют ген (E9) неизвестной функции, предполагая общее происхождение.
Эволюция
[ редактировать ]Считается, что эволюция папилломы является медленной по сравнению со многими другими типами вирусов, но в настоящее время нет экспериментальных измерений. Вероятно, это связано с тем, что геном папилломавируса состоит из генетически стабильной двухцепочечной ДНК, которая реплицируется с высокой точностью от механизма репликации ДНК-клетки. [ Цитация необходима ]
Считается, что папилломавирусы, как правило, совпадают с определенным видом животных-хозяина в течение многих лет, хотя существуют сильные доказательства против гипотезы о коэволюции. [ 13 ] [ 25 ] В особенно быстром примере HPV-16 немного развивался, так как население человека расширилось по всему миру и теперь варьируется в разных географических регионах таким образом, что, вероятно, отражает историю миграции человека. [ 26 ] [ 27 ] Типы Cutaneotropic HPV иногда обмениваются между членами семьи в течение всей жизни, но другие доноры также следует учитывать при вирусной передаче. [ 28 ]
Другие типы ВПЧ, такие как HPV-13, относительно мало в разных популяциях человека. Фактически, последовательность ВПЧ-13 очень напоминает папилломавирус бонобо (также известный как пигмея шимпанзе). [ 29 ] Неясно, связано ли это сходство с недавней передачей между видами или потому, что ВПЧ-13 просто очень мало изменилось за шесть или около того миллиона лет, так как люди и бонобо расходились. [ 27 ]
По оценкам, самый последний общий предок этой группы вирусов существовал 424 миллионов лет назад . [ 30 ]
Есть пять основных родов, заражающих людей (альфа, бета, гамма, му и ню). Самый последний общий предок этих родов развился 49,7 миллиона лет назад - 58,5 миллионов лет назад . [ 31 ] По оценкам, самый последний предок гамма -рода развился между 45,3 миллионами лет назад и 67,5 миллиона лет назад . [ Цитация необходима ]
Структура
[ редактировать ]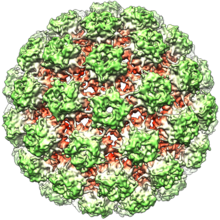
Папилломавирусы не развиты, что означает, что внешняя оболочка или капсид вируса не покрыты липидной мембраной . Один вирусный белок, известный как L1, необходим и достаточен для образования капсида нанометра 55–60, состоящего из 72 звездных капсомеров (см. Рисунок). Как и большинство неразвитых вирусов, капсид является геометрически регулярным и представляет икосаэдрическую симметрию . Самосборные вирусные частицы, состоящие из L1, являются основой успешной группы профилактических вакцин против ВПЧ , предназначенных для выявления вирус-нейтрализующих антител , которые защищают от первоначальной инфекции ВПЧ. Таким образом, папилломавирид стабильны в окружающей среде . [ Цитация необходима ]
Геном папилломы представляет собой двухцепочечную круговую молекулу ДНК ~ 8000 пар оснований длиной . Он упакован в оболочку L1 вместе с клеточными гистоновыми белками, которые служат для обертывания и конденсации ДНК. [ Цитация необходима ]
Капсид папилломы также содержит вирусный белок, известный как L2, который менее распространен. Хотя это и не ясно, как L2 расположен в вирионе, известно, что он выполняет несколько важных функций, включая облегчение упаковки вирусного генома в зарождающиеся вирионы, а также инфекционную проникновение вируса в новые клетки -хозяина. L2 представляет интерес как возможную цель для более широко защитных вакцин против ВПЧ .
Вирусный капсид состоит из 72 капсомеров, из которых 12 составляют пять координированных, а 60-шестью координированные капсомеры, расположенные на T = 7D икосаэдрической поверхностной решетке. [ 32 ]
Тканевая специфичность
[ редактировать ]Папилломавирусы повторяются исключительно в кератиноцитах . Кератиноциты образуют самые внешние слои кожи, а также некоторые поверхности слизистой оболочки , такие как внутренняя часть щеки или стены влагалища. Эти поверхностные ткани, которые известны как стратифицированные плоскоклеточные эпителии , состоят из сложенных слоев уплощенных клеток. Слои клеток образуются через процесс, известный как клеточная дифференцировка , в котором кератиноциты постепенно становятся специализированными, в конечном итоге образуя жесткую, сшитую поверхность, которая предотвращает потерю влаги и действует как барьер против патогенов. Считается, что стволовые клетки кератиноцитов, пополняемые на поверхностном слое, являются начальной мишенью продуктивных папилломавирусных инфекций. Последующие шаги в жизненном цикле вируса строго зависят от процесса дифференцировки кератиноцитов. В результате папилломавирусы могут повторять только в поверхностных тканях тела. [ Цитация необходима ]
Жизненный цикл
[ редактировать ]Заразительный вход
[ редактировать ]Папилломавирусы получают доступ к стволовым клеткам кератиноцитов через небольшие раны, известные как микротравмы, на поверхности кожи или слизистой оболочки. Взаимодействие между L1 и сульфатированными сахарами на клеточной поверхности способствует начальному прикреплению вируса. [ 33 ] [ 34 ] Затем вирус способен проникать внутрь с клеточной поверхности через взаимодействие со специфическим рецептором, вероятно, через интегран Alpha-6 Beta-4, [ 35 ] [ 36 ] и транспортируется в мембранные везикулы, называемые эндосомами . [ 37 ] [ 38 ] Капсидный белок L2 нарушает мембрану эндосомы через катионные клеточные пептид , позволяя вирусному геному избежать и движения, наряду с L2, к клеточному ядру. [ 39 ] [ 40 ] [ 41 ]
Вирусная стойкость и задержка
[ редактировать ]После успешной инфекции кератиноцита вирус экспрессирует белки E1 и E2, которые предназначены для репликации и поддержания вирусной ДНК в качестве круговой эпизоды . Вирусные онкогены E6 и E7 способствуют росту клеток путем инактивации белков -супрессоров опухоли p53 и prb . Стволовые клетки кератиноцитов в эпителиальном базальном слое могут поддерживать геномы папилломы на протяжении десятилетий. [ 8 ]
Производство вируса потомства
[ редактировать ]Текущее понимание состоит в том, что репликация вирусной ДНК, вероятно, происходит в G 2 фазе клеточного цикла и полагается на рекомбинационную репликацию, подтверждаемую механизмами ответа на повреждение ДНК (активируется белком E7) для получения геномов вируса потомства. [ 42 ] Геномы папилломы иногда интегрируются в геном-хозяина, особенно заметный с онкогенными ВПЧ, но не являются нормальной частью жизненного цикла вируса и тупиковым, который устраняет потенциал выработки вирусного потомства. [ 42 ]
Экспрессия вирусных поздних генов, L1 и L2, исключительно ограничена дифференцированием кератиноцитов во внешних слоях кожи или поверхности слизистой оболочки. Повышенная экспрессия L1 и L2 обычно коррелирует с резким увеличением количества копий вирусного генома. Поскольку внешние слои стратифицированного плоскоклеточного эпителия подвергаются относительно ограниченному наблюдению клетками иммунной системы, считается, что это ограничение вирусной экспрессии позднего гена представляет собой форму иммунной уклонения. [ Цитация необходима ]
Новые инфекционные вирусы потомства собираются в клеточном ядре . Папилломавирусы развили механизм освобождения вирионов в окружающую среду. Другие виды неразвитых вирусов животных используют активный литический процесс для убийства клетки-хозяина, позволяя высвобождать частицы вируса потомства. Часто этот литический процесс связан с воспалением , что может вызвать иммунную атаку против вируса. Папилломавирусы используют десквамацию как скрытный, невоспалительный механизм высвобождения. [ Цитация необходима ]
| Род | Детали хоста | Ткань тропизм | Детали записи | Выпустить детали | Сайт репликации | Сайт сборки | Передача инфекции |
|---|---|---|---|---|---|---|---|
| Dyoxipapillomavirus | Позвоночные | Никто | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Omikronpapillomavirus | Свинья | Эпителиальный: слизистый; Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Dyodeltapillomavirus | Позвоночные | Никто | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Омегапилломавирус | Позвоночные | Эпителиальный: слизистый; Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Нупапилломавирус | Люди | Эпителиальный: слизистый; Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Dyomupapillomavirus | Позвоночные | Никто | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Dyozetapillomavirus | Позвоночные | Никто | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Каппапапилломавирус | Кролики | Эпителиальный: слизистый; Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Upsilonpapillomavirus | Позвоночные | Эпителиальный: слизистый; Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Dyoetapillomavirus | Позвоночные | Никто | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Сигмапилломавирус | Позвоночные | Эпителиальный: слизистый; Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Lambdapillomavirus | Кошки; собак | Эпителиальный: слизистый; Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Taupapallomanavirus | Позвоночные | Эпителиальный: слизистый; Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Бетапапилломавирус | Люди | Эпителиальный: слизистый; Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Xipapillomavirus | Бычьи | Эпителиальный: слизистый; Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Dyoepsilonpapillomavirus | Позвоночные | Никто | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Thetapillomavirus | Птицы | Эпителиальный: слизистый; Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Etapapillomavirus | Птицы | Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Ропапилломавирус | Позвоночные | Эпителиальный: слизистый; Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Dyothetapillomavirus | Позвоночные | Никто | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Dyoomikronpapillomavirus | Позвоночные | Никто | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Гаммапапилломавирус | Люди | Эпителиальный: слизистый; Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Альфапилломавирус | Люди; обезьяны | Эпителиальный: слизистый; Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Секс; контакт |
| Zetapapillomavirus | Лошади | Эпителиальный: слизистый; Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Deltapillomavirus | Жвачки | Эпителиальный: слизистый; Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Dyolambdapillomavirus | Позвоночные | Никто | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Dyosigmapillomavirus | Позвоночные | Никто | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Dyorhopillomavirus | Позвоночные | Никто | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Psipapillomavirus | Позвоночные | Эпителиальный: слизистый; Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Dyokappapillomavirus | Позвоночные | Никто | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Pipapillomavirus | Хомяки | Эпителиальный: слизистый; Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Йотапапилломавирус | Грызуны | Эпителиальный: слизистый; Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Эпсилонпапилломавирус | Бычьи | Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Фипапилломавирус | Позвоночные | Эпителиальный: слизистый; Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Dyonupapillomavirus | Позвоночные | Никто | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Dyopipipillomavirus | Позвоночные | Никто | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Dyoiotapillomavirus | Позвоночные | Никто | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
| Мупапилломавирус | Люди | Эпителиальный: слизистый; Эпителиальный: кожа | Эндоцитоз клеточного рецептора | Лизис | Ядро | Ядро | Контакт |
Ассоциация с раком
[ редактировать ]Хотя некоторые типы папилломы могут вызывать рак в эпителиальных тканях, в которых они обитают, рак не является типичным результатом инфекции. Разработка раковых заболеваний, вызванных папилломой, обычно происходит в течение многих лет. Папилломавирусы были связаны с развитием рака шейки матки , рака полового члена [ 43 ] и устный рак . [ 44 ] связь с раком вульвального рака и уротелиальной карциномой с плоскоклеточной дифференцировкой у пациентов с нейрогенным мочевым пузырем. Также была отмечена [ 45 ] [ 46 ] Существует рак, вызывающий папилломавирус геном, который кодирует два небольших белка, называемые E6 и E7, которые имитируют рак, вызывающий онкогены. Они работают, что они стимулируют неестественный рост клеток и блокируют свою естественную защиту. Также они действуют на многих сигнальных белках, которые контролируют пролиферацию и апоптоз. [ 47 ]
Лабораторное исследование
[ редактировать ]Тот факт, что жизненный цикл папилломы строго требует дифференциации кератиноцитов, создал существенный барьер для изучения папилломы в лаборатории, поскольку он исключил использование обычных клеточных линий для выращивания вирусов. Поскольку инфекционные вирионы BPV-1 могут быть извлечены из больших бородавок, которые вирус индуцирует на крупном рогатый скот, он был моделью папилломавируса рабочей лошадки в течение многих лет. CRPV, папилломавирус полости кролика (ROPV) и пероральный папилломавирус собак (COPV) также широко использовались для лабораторных исследований. Как только исследователи обнаружили, что эти вирусы вызывают рак, они работали вместе, чтобы найти в ней вакцину. В настоящее время наиболее эффективный способ сделать это - имитировать вирус, который состоит из белка L1, но не имеет ДНК. По сути, наша иммунная система строит защиту от инфекций, но если эти инфекции не вызывают заболевания, их можно использовать в качестве вакцины. Вход PDB 6BT3 показывает, как антитела поверхности атакуют поверхность вируса, чтобы отключить его. [ 48 ]
Некоторые типы ВПЧ, передаваемых половым путем, были распространены с использованием мышиной системы «ксенотрансплантата», в которой человеческие клетки, инфицированные HPV, имплантируются на мышей с иммунодефицитом . Совсем недавно некоторым группам удалось выделить инфекционный ВПЧ-16 из поражений шейки матки человека. Тем не менее, выделение инфекционных вирионов с использованием этой техники является трудной, а выход инфекционного вируса очень низкий. [ Цитация необходима ]
Дифференциация кератиноцитов можно имитировать in vitro , подвергая культивируемых кератиноцитов на границу воздуха/жидкость. Адаптация таких систем «культуры плота» к изучению папилломы была значительным прорывом для in vitro . исследования жизненного цикла вируса [ 49 ] Тем не менее, системы культивирования плота относительно громоздки, а урожайность инфекционных ВПЧ может быть низким. [ 50 ]
Разработка системы на основе дрожжей, которая позволяет стабильно эпизомальной репликации ВПЧ, обеспечивает удобное, быстрое и недорогое средство для изучения нескольких аспектов жизненного цикла ВПЧ (Angeletti 2002). Например, E2-зависимая транскрипция, амплификация генома и эффективное инкапдиацию полноразмерных DNA HPV могут быть легко воссозданы у дрожжей (Angelelti 2005).
временные методы высокодоходства для производства псевдовирусов В последнее время были разработаны ВПЧ, несущих репортерные гены. Хотя псевдовирусы не подходят для изучения определенных аспектов жизненного цикла вируса, первоначальные исследования показывают, что их структура и первоначальный инфекционный вступление в клетки, вероятно, во многих отношениях схожи с подлинными папилломавирусами.
Папилломавирус человека связывается с молекулами гепарина на поверхности клеток, которые он заражает. Исследования показали, что кристалл изолированных капсомеров L1 имеет цепи гепарина, распознаваемые канавками лизинов на поверхности вируса. Также те, у кого антитела, показывают, что они могут заблокировать это признание. [ 51 ]
Генетическая организация и экспрессия генов
[ редактировать ]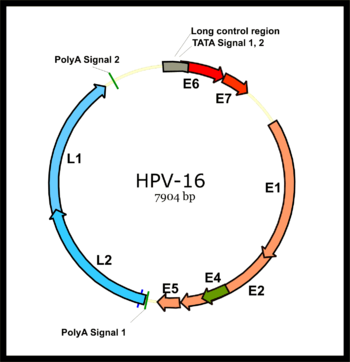
Геном папилломы делится на раннюю область (E), кодируя шесть рамков открытых считываний (ORF) (E1, E2, E4, E5, E6 и E7), которые экспрессируются сразу после первоначальной инфекции клетки -хозяина, и позднее область (L), кодирующая основной капсидный белок L1 и незначительный капсидный белок L2. Все вирусные ORF кодируются на одной пряди ДНК (см. Рисунок). Это представляет собой драматическую разницу между папилломавирусами и полиомавирусами , поскольку последний тип вируса экспрессирует его ранние и поздние гены путем двунаправленной транскрипции обеих целей ДНК. Это различие было основным фактором в установлении консенсуса о том, что папилломавирусы и полиомавирусы, вероятно, никогда не имели общего предка, несмотря на поразительные сходства в структурах их вирионов. [ Цитация необходима ]
После того, как клетка -хозяина заражена, ранний промотор HPV16 активируется и транскрибируется полицистронная первичная РНК, содержащая все шесть ранних ORF. Эта полицистронная РНК содержит три экзона и два интрона и подвергается активному сплайсингу РНК, чтобы генерировать множественные изоформы мРНК. [ 52 ] Один из сплайсированных изоформ RNA, E6*I, служит мРНК E7 для трансляции онкопротеина E7. [ 53 ] Напротив, интрон в E6 ORF, который остается нетронутым без сплайсинга, необходим для трансляции онкопротеина E6. [ 53 ] Тем не менее, вирусная ранняя транскрипция подвергается регуляции вируса E2 и высоким уровням E2, подавляют транскрипцию. Геномы ВПЧ интегрируются в геном хозяина путем нарушения E2 ORF, предотвращая репрессию E2 на E6 и E7. Таким образом, интеграция вирусного генома в геном ДНК -хозяина увеличивает экспрессию E6 и E7, чтобы способствовать клеточной пролиферации и вероятности злокачественности. [ Цитация необходима ]
Основной вирусный поздний промотор в вирусной ранней области становится активным только в дифференцированных клетках, и его активность может быть сильно повышена при репликации вирусной ДНК. Поздняя транскрипция также является полицистронной РНК, которая содержит два интрона и три экзона. Альтернативный сплайсинг РНК этого позднего транскрипта имеет важное значение для экспрессии L1 и L2 и может регулироваться РНК-цис-элементами и факторами сплайсинга хозяина. [ 52 ] [ 54 ] [ 55 ]
Техническое обсуждение функций гена папилломавируса
[ редактировать ]Гены в геноме папилломы обычно идентифицируются после сходства с другими ранее идентифицированными генами. Тем не менее, некоторые ложные открытые рамки для чтения могли быть приняты за гены просто после их положения в геноме, и не могли быть истинными генами. E3, E4, E5 и E8 Это особенно относится к определенным рамкам . [ Цитация необходима ]
E1
[ редактировать ]Кодирует белок, который связывается с вирусным происхождением репликации в длинной контрольной области вирусного генома. E1 использует АТФ для проявления активности геликазы , которая выделяет цепи ДНК, тем самым приготовляя вирусный геном для репликации с помощью клеточных факторов репликации ДНК .
E2
[ редактировать ]Белок E2 служит основным регулятором транскрипции для вирусных промоторов, расположенных в основном в области длинной контроля. Белок имеет трансактивационный домен, связанный с относительно неструктурированной областью шарнира с хорошо охарактеризованным ДНК-связывающим доменом. E2 облегчает связывание E1 с вирусным происхождением репликации. E2 также использует клеточный белок, известный как бромодомен -4 (BRD4) для привязки вирусного генома к клеточным хромосомам. [ 56 ] клетки Это привязанность к ядерной матрице обеспечивает верное распределение вирусных геномов по каждой дочерней клетке после деления клеток. Считается, что E2 служит негативным регулятором экспрессии для онкогенов E6 и E7 в латентном HPV-инфицированном базальных кератиноцитах . Генетические изменения, такие как интеграция вирусной ДНК в хромосому клеток -хозяина, которые инактивируют экспрессию E2, имеет тенденцию увеличивать экспрессию онкогенов E6 и E7, что приводит к клеточной трансформации и, возможно, дальнейшей генетической дестабилизации.
E3
[ редактировать ]Этот небольшой предполагаемый ген существует только в нескольких типах папилломы. Известно, что ген не экспрессируется как белок и, по -видимому, не выполняет никаких функций.
E4
[ редактировать ]Хотя белки E4 экспрессируются на низких уровнях во время ранней фазы вирусной инфекции, экспрессия E4 значительно увеличивается в течение поздней фазы инфекции. Другими словами, его «E» апелляция может быть чем -то вроде неправильного. В случае HPV-1 E4 может составлять до 30% общего белка на поверхности бородавки. [ 57 ] Считается, что белок E4 многих типов папилломы способствует высвобождению вириона в окружающую среду, нарушая промежуточные нити кератиноцитов цитоскелета . Вирусные мутанты, неспособные экспрессировать E4, не поддерживают репликацию высокого уровня вирусной ДНК, но пока не ясно, как E4 облегчает репликацию ДНК. Также было показано, что E4 участвует в арестовающих клетках в фазе G2 клеточного цикла .
E5
[ редактировать ]E5 - это небольшие, очень гидрофобные белки, которые дестабилизируют функцию многих мембранных белков в инфицированной клетке. [ 58 ] Белок E5 некоторых типов папилломы животных (в основном бычьего папилломавируса типа 1) функционирует как онкоген, главным образом, путем активации сигналов, способствующей развитию роста клеток рецепторов фактора роста тромбоцитов . Однако белки E5 папилломы человека, связанные с раком, по -видимому, активируют сигнальный каскад, инициированный эпидермальным фактором роста при связывании лиганда. Также было показано, что HPV16 E5 и HPV2 E5 подавляют поверхностную экспрессию основных белков комплекса гистосовместимости , которые могут предотвратить устранение инфицированных клетки с помощью T-клеток .
E6
[ редактировать ]
E6 представляет собой аминокислотный пептид 151, который включает в себя мотив типа 1 с консенсусной последовательности- (T/S)-(x)-(V/i) -coOH. [ 60 ] [ 61 ] У этого также есть два мотива цинковых пальцев . [ 60 ]
E6 представляет особый интерес, потому что он, по -видимому, играет множественную роль в клетке и взаимодействует со многими другими белками. Однако его основная роль состоит в том, чтобы опосредовать деградацию p53 , основного белка -супрессора опухоли , снижая способность клетки реагировать на повреждение ДНК . [ 62 ] [ 63 ]
Также было показано, что E6 нацелен на другие клеточные белки, тем самым изменяя несколько метаболических путей . Одной из таких мишени является NFX1-91 , который обычно подавляет продукцию теломеразы , белок, который позволяет клеткам делить неограниченное количество раз. Когда NFX1-91 деградируется E6, уровни теломеразы увеличиваются, инактивируя основной механизм, контролирующий рост клеток. [ 64 ] Кроме того, E6 может действовать как транскрипционный кофактор - в частности, активатор транскрипции - при взаимодействии с клеточным фактором транскрипции E2F1/DP1. [ 60 ]
E6 также может связываться с PDZ-доменами , короткими последовательностями, которые часто обнаруживаются в сигнальных белках. Структурный мотив E6 позволяет взаимодействовать с доменами PDZ на генах DLG (диски большие) и HDLG (Drosophila Large). [ 61 ] [ 65 ] Связывание в этих местах вызывает трансформацию белка DLG и разрушение его функции подавителя. Белки E6 также взаимодействуют с белками семейства гуанилат-киназы Maguk (мембрана. Эти белки, включая MAGI-1, MAG-2 и MAGI-3, обычно являются структурными белками и могут помочь с передачей сигналов. [ 61 ] [ 65 ] Что еще более важно, они, как полагают, связаны с действием подавления DLG. Когда комплексы E6 с доменами PDZ на белках Magi он искажает их форму и тем самым препятствует их функции. В целом, белок E6 служит для того, чтобы препятствовать нормальной активности белка таким образом, чтобы позволить клетке расти и умножаться с повышенной частотой, характерной для рака.
Поскольку экспрессия E6 строго требуется для поддержания злокачественного фенотипа при раке, вызванных HPV, это привлекательная мишень терапевтических вакцин против ВПЧ, предназначенных для искоренения установленных опухолей рака шейки матки.
E7
[ редактировать ]У большинства вирусов папилломы первичной функцией белка E7 является инактивирование членов семейства PRB белков -супрессоров опухолей. Вместе с E6 E7 служит для предотвращения гибели клеток ( апоптоз ) и способствует прогрессированию клеточного цикла , таким образом заполняя клетку для репликации вирусной ДНК. E7 также участвует в иммортализации инфицированных клеток путем активации клеточной теломеразы . Как и E6, E7 является предметом интенсивного исследовательского интереса и, как полагают, оказывает множество других эффектов на инфицированные клетки. Как и в случае с E6, продолжающаяся экспрессия E7 требуется для выживания раковых клеточных линий, таких как HELA , которые получены из HPV-индуцированных опухолей. [ 66 ]
E8
[ редактировать ]Только несколько типов папилломы кодируют короткий белок из гена E8. В случае BPV-4 (Papillomavirus genus XI ) рамка открытого считывания E8 может заменить рамку открытого считывания E6, которая отсутствует в этом роде папилломавируса. [ 67 ] Эти гены E8 химически и функционально похожи на гены E5 из некоторых папилломавирусов человека, а также называются E5/E8.
L1
[ редактировать ]L1 спонтанно сами входит в пентамерные капсомеры. Очищенные капсомеры могут продолжать образовывать капсиды, которые стабилизируются дисульфидными связями между соседними молекулами L1. Капсиды L1, собранные in vitro, являются основой профилактических вакцин против нескольких типов ВПЧ. По сравнению с другими генами папилломы, аминокислотные последовательности большинства частей L1 хорошо справляются между типами. Однако поверхностные петли L1 могут существенно различаться, даже для разных членов конкретного вида папилломы. Это, вероятно, отражает механизм уклонения нейтрализующих реакций антител, вызванных предыдущими папилломавирусными инфекциями. [ 68 ]
L2
[ редактировать ]L2 существует в окисленном состоянии в вирионе папилломы, причем два консервативных остатка цистеина образуют внутримолекулярную дисульфидную связь. [ 69 ] В дополнение к сотрудничеству с L1 для упаковки вирусной ДНК в вирион, было показано, что L2 взаимодействует с рядом клеточных белков в процессе инфекционного входа. После начального связывания вириона с клеткой L2 должен быть расщеплен клеточным протеазой фурин . [ 70 ] Вирион интернализуется, вероятно, посредством клатрин-опосредованного процесса, в эндосому, где, как полагают, кислые условия приводят к воздействию мембранно-прерывающих частей L2. [ 39 ] клеточных белков Бета -актин [ 71 ] и синтаксин -18 [ 72 ] Может также участвовать в L2-опосредованных событиях входа. После выхода эндосомы L2 и вирусного генома импортируются в ядро клетки, где они движутся в субназорный домен, известный как тело ND-10 , которое богато факторами транскрипции . [ 40 ] Небольшие части L2 хорошо справляются между различными типами папилломы, и экспериментальные вакцины, нацеленные на эти консервативные домены, могут обеспечить защиту от широкого диапазона типов ВПЧ. [ 73 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Van Doorlaer, K; Чен, Z; Бернард, Ху; Чан, PSS; Desalle, R; Dillner, J; Forslund, o; пока, t; Макбрайд, А.А; Villa, LL; Burk, Rd; Отчет ICTV, Консорциум (август 2018 г.). "Профиль таксономии вируса ICTV: Papillomaviridae " Журнал общей вирусологии 99 (8): 989–9 Doi : 10.1099/ jgv.0.0 6171710PMC 29927370PMID
- ^ Jump up to: а беременный в De Villiers EM, Fauquet C, Broker TR, Bernard Hu, Zur Hausen H (июнь 2004 г.). «Классификация папилломы» . Вирусология . 324 (1): 17–27. doi : 10.1016/j.virol.2004.03.033 . PMID 15183049 .
- ^ Herbst LH, Lenz J, Van Toslare K, Chen Z, Stacy Ba, Wellhan JF, Manire CA, Birk Rd (Janary 2009). «Парламент парламента Enmic или два парламента парламента, Cholonia Mydas Parrot 1 и парламент Caretta Parstta 1» . Viology . 383 (1): 131–5. doi : 10,1016/j.vill.2008.09,022 . PMID 18973915 .
- ^ Друри С.Е., Гоф Р.Е., Макартур С., Джессоп М. (декабрь 1998 г.). «Обнаружение герпесвирусных и папилломавирусных частиц, связанных с болезнями черепах». Ветеринарная запись . 143 (23): 639. PMID 9881444 .
- ^ Lange CE, Favrot C, Ackermann M, Gull J, Vetsch E, Tobler K (сентябрь 2011 г.). «Роман Снейк Папилломавирус не кластет с другими папилломавирусами не млекопитающих» . Virology Journal . 8 : 436. DOI : 10.1186/1743-422X-8-436 . PMC 3179961 . PMID 21910860 .
- ^ Muñoz N, Castellsagué X, De González AB, Gissmann L (август 2006 г.). «Глава 1: ВПЧ в этиологии рака человека». Вакцина . 24 Suppl 3 (3): S3/1–10. doi : 10.1016/j.vaccine.2006.05.115 . PMID 16949995 .
- ^ Мистри Н., Вибом С, Эвандер М (октябрь 2008 г.). «Кожные и слизистые папилломавирусы человека отличаются по чистому поверхностному заряду, потенциальное влияние на тропизм» . Virology Journal . 5 : 118. DOI : 10.1186/1743-422X-5-118 . PMC 2571092 . PMID 18854037 .
- ^ Jump up to: а беременный Дверь J (март 2005 г.). «Жизненный цикл папилломы». Журнал клинической вирусологии . 32 (Suppl 1): S7–15. doi : 10.1016/j.jcv.2004.12.006 . PMID 15753007 .
- ^ Kocjan BJ, Hosnjak L, Seme K, Poljak M (май 2013). «Полная последовательность генома нового человеческого бетапапилломавируса, HPV-159» . Объявления генома . 1 (3): E00298–13. doi : 10.1128/genomea.00298-13 . PMC 3668007 . PMID 23723399 .
- ^ «Таксономия вируса: выпуск 2018b» . Международный комитет по таксономии вирусов. Февраль 2019 . Получено 26 марта 2019 года .
- ^ «Таксономия вируса: выпуск 2014 года» . ICTV . Получено 15 июня 2015 года .
- ^ Бернард Ху, Берк Р.Д., Чен З., Ван Дорклаер К., Зур Хаузен Х., Де Вильерс Эм (май 2010 г.). «Классификация папилломы (PVS) на основе 189 типов PV и предложения таксономических поправок» . Вирусология . 401 (1): 70–9. doi : 10.1016/j.virol.2010.02.002 . PMC 3400342 . PMID 20206957 .
- ^ Jump up to: а беременный в Gottschling M, Stamatakis A, Nindl I, Stockfleth E, Alonso A, Bravo IG (2007). «Многочисленные эволюционные механизмы способствуют диверсификации папилломы» . Молекулярная биология и эволюция . 24 (5): 1242–58. doi : 10.1093/molbev/msm039 . PMID 17344207 .
- ^ Campo MS , ed. (2006). Исследование папилломы: от естественной истории до вакцин и за ее пределами . Caister Academic Press. ISBN 978-1-904455-04-2 . [1] .
- ^ Bravo IG, De Sanjosé Llongueras S, Gottschling M (2010). «Клиническая важность знаний об эволюции папилломы». Тенденции в микробиологии . 18 (10): 432–8. doi : 10.1016/j.tim.2010.07.008 . PMID 20739182 .
- ^ Ректор А, Ван Ранст М (октябрь 2013 г.). «Папилломавирусы животных» . Вирусология . 445 (1–2): 213–23. doi : 10.1016/j.virol.2013.05.007 . PMID 23711385 .
- ^ Лопес-Буэно А., Мавиан С., Лабелла А.М., Кастро Д., Боррего Дж.Дж., Альками А., Алехо А (октябрь 2016 г.). «Керренс иридовируса, полиомавируса и уникального члена новой группы рыбных палопломавирусов в морском лещном лещном лещном из лимфоциза, затронутым лимфоцизом », Журнал вирусологии 90 (19): 8768–7 Doi : 10.1128/ jvi.01369-1 5021401PMC PMID 274440877
- ^ Choy D, Bolatti E, Pérez GR, Prumber 2013). Полем Журнал . 94 (Pt 11): 2480–8 doi : 10.1099/ vir.05513-0 HDL : 2133/9862 . PMID 23997181 .
- ^ Антонсон А., Ханссон Б.Г. (декабрь 2002 г.). «Здоровая кожа многих видов животных содержит папилломавирусы, которые тесно связаны с их коллегами -человеком» . Журнал вирусологии . 76 (24): 12537–42. doi : 10.1128/JVI.76.24.12537-12542.2002 . PMC 136724 . PMID 12438579 .
- ^ Холлидей, Чак. «Профессор Чак Холлидея на странице www в Лафайет -колледже» . Архивировано с оригинала 2014-07-18 . Получено 2014-07-13 .
- ^ Кристенсен Н.Д. (2005). «Модельная система Cottontail Papillomavirus (CRPV) для тестирования антивирусных и иммунотерапевтических стратегий» . Антивирусная химия и химиотерапия . 16 (6): 355–62. doi : 10.1177/095632020501600602 . PMID 16331841 .
- ^ Coggins LW, MA JQ, Slater AA, Campo MS (июнь 1985 г.). «Гомологии последовательности между бычьим папилломавирусным геномами, отображаемыми новым методом гетеродуплекса с низкой струней». Вирусология . 143 (2): 603–11. doi : 10.1016/0042-6822 (85) 90398-8 . PMID 2998027 .
- ^ Ван Ранст М., Тачжи Р., Прус Дж., Берк Р.Д. (июнь 1992 г.). «Первичная структура белка E6 микромис minutus papillomavirus и Mastomys natalensis papillomavirus» . Исследование нуклеиновых кислот . 20 (11): 2889. doi : 10.1093/nar/20.11.2889 . PMC 336941 . PMID 1319576 .
- ^ Varsani A, Kraberger S, Jennings S, Porzig EL, Julian L, Massaro M, Pollard A, Ballard G, Ainley DG (июнь 2014 г.). «Новый папилломавирус в фекалиях Adélie Penguin (Pygoscelis Adeliae), отобранных в колонии мыса Крозье, Антарктида» . Журнал общей вирусологии . 95 (Pt 6): 1352–65. doi : 10.1099/vir.0.064436-0 . PMID 24686913 . S2CID 206218507 .
- ^ Gottschling M, Göker M, Stamatakis A, Bininda-Emonds или, Nindl I, Bravo IG (июль 2011 г.). «Количественная оценка филодинамических сил, способствующих эволюции папилломы» . Молекулярная биология и эволюция . 28 (7): 2101–13. doi : 10.1093/molbev/msr030 . PMID 21285031 .
- ^ Ho L, Chan Sy, Burk RD, Das BC, Fujinaga K, Icenogle JP, Kahn T, Kiviat N, Lancaster W, Mavromara Nazos P (ноябрь 1993 г.). «Генетический дрейф человеческого папилломавируса типа 16 является средством реконструкции доисторического вирусного распространения и движения древних человеческих популяций» . Журнал вирусологии . 67 (11): 6413–23. doi : 10.1128/jvi.67.11.6413-6423.1993 . PMC 238076 . PMID 8411343 .
- ^ Jump up to: а беременный Callja-Macias IE, Villa LL, Prado JC, Kalantari M, Allan B, Williamson AL, Chung LP, Collins RJ, Zuna RE, Dunn St, Chu Ty, Cubie Ha, Cuschieri K, Von Knebel-Debeberitz M, Marins CR, Санчес Г.И., Бош Фкс, Муньос Н., Бернард Ху (ноябрь 2005 г.). «Всемирное геномное разнообразие папилломавирусов человека высокого риска типов 31, 35, 52 и 58, четыре близких относительных отношениях человеческого папилломавируса типа 16 » Журнал вирусологии 79 (21): 13630–40. Doi : 10.1128/ jvi.79.21.13630-13640.2 1262609PMC 16227283PMID
- ^ Gottschling M, Göker M, Köhler A, Lehmann MD, Stockfleth E, Nindl I (2009). «Катнеотропные типы β-/γ-HPV редко используются между членами семьи» . Журнал расследования дерматологии . 129 (10): 2427–34. doi : 10.1038/jid.2009.94 . PMID 19516265 .
- ^ От Ranst M, Fuse A, Fiten P, Books E, Pfister H, Borrk Rd, октябрь G (октябрь 1992 г.). «Человеческий папиллавирус типа 13 и пигмейный христиан падилломавируса типа 1: компрессон или организации, взятые организации». Viology . 190 (2): 587–96. doi : 10,1016/0042-6822 (92) 90896-W . PMID 1325697 .
- ^ Виллемен А., Браво И.Г. (2019) Происхождение и эволюция папилломавируса (ONCO) генов и геномов. Philos Trans R Soc Lond B Biol Sci. 374 (1773): 20180303
- ^ Murahwa AT, Nindo F, Onywera H, Mering TL, Martin DP, Williamson AL (2019). BMC Genomics 20 (1): 368
- ^ Rayment I, Baker TS, Caspar DL, Murakami WT (январь 1982). «Капсидная структура вируса полиомы при 22,5 разрешении» . Природа . 295 (5845): 110–5. Bibcode : 1982natur.295..110R . doi : 10.1038/295110A0 . PMC 4144041 . PMID 6276752 .
- ^ Джойс Дж.Г., Тунг Дж. С., Прзисицки К.Т., Кук Дж.С., Леман Э.Д., Сэндс Дж.А., Янсен Ку, Келлер П.М. (февраль 1999 г.). «Основной капсидный белок L1 из рекомбинантных вирусных вирусных частиц человека типа типа 11 взаимодействует с гликозаминогликанами гепарина и клеточной поверхности на кератиноцитах человека» . Журнал биологической химии . 274 (9): 5810–22. doi : 10.1074/jbc.274.9.5810 . PMID 10026203 .
- ^ Джироглу Т., Флорин Л., Шефер Ф., Стрейк Р.Е., Сапп М. (февраль 2001 г.). «Человеческая папилломавирусная инфекция требует гепаран -сульфата клеточной поверхности» . Журнал вирусологии . 75 (3): 1565–70. doi : 10.1128/JVI.75.3.1565-1570.2001 . PMC 114064 . PMID 11152531 .
- ^ Эвандер М., Фрейзер И.Х., Пейн Е., Q YM, Хенгт К., Макмиллан На (март 1997 г.). «Идентификация интегрина Alpha6 в качестве рецептора кандидата для папилломавирусов » Журнал вирусологии 71 (3): 2449–5 Doi : 10.1128/ jvi.71.3.2449-2456.1 191355PMC 9032382PMID
- ^ McMillan NA, Payne E, Frazer IH, Evander M (сентябрь 1999 г.). «Экспрессия Alpha6 Intein придает папилломавирусное связывание на рецептор-негативные B-клеток » Вирусология 261 (2): 271–9 Doi : 10.1006/ viro.1999.9 PMID 10497112
- ^ Selinka HC, Giroglou T, Sapp M (август 2002 г.). «Анализ инфекционного пути въезда псевдовирионов папилломы 33 типа 33» . Вирусология . 299 (2): 279–287. doi : 10.1006/viro.2001.1493 . PMID 12202231 .
- ^ День PM, Lowy DR, Schiller JT (март 2003 г.). «Папилломавирусы заражают клетки через клатрин-зависимый путь» . Вирусология . 307 (1): 1–11. doi : 10.1016/s0042-6822 (02) 00143-5 . PMID 12667809 .
- ^ Jump up to: а беременный Kämper N, Day PM, Nowak T, Selinka HC, Florin L, Bolscher J, Hilbig L, Schiller JT, Sapp M (январь 2006 г.). «Мембрано-пропагандирующий пептид в капсидном белке L2 необходим для выхода геномов папилломы из эндосомов» . Журнал вирусологии . 80 (2): 759–68. doi : 10.1128/jvi.80.2.759-768.2006 . PMC 1346844 . PMID 16378978 .
- ^ Jump up to: а беременный День PM, Baker CC, Lowy DR, Schiller JT (сентябрь 2004 г.). «Создание инфекции папилломавируса усиливается экспрессией белка белка промиелоцитарного лейкоза (PML)» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (39): 14252–7. Bibcode : 2004pnas..10114252d . doi : 10.1073/pnas.0404229101 . PMC 521143 . PMID 15383670 .
- ^ Чжан, Пенгвей; Монтейро да Силва, Габриэль; Deatherage, Екатерина; Берд, Кристофер; Dimaio, Daniel (2018). «Пептид, проникающий клетками, опосредует внутриклеточную мембрану прохождения белка папилломы человека человека, чтобы вызвать ретроградный транспортировку» . Клетка . 174 (6): 1465–1476.e13. doi : 10.1016/j.cell.2018.07.031 . PMC 6128760 . PMID 30122350 .
- ^ Jump up to: а беременный Элисон А. Макбрайд (18 марта 2017 г.). «Механизмы и стратегии репликации папилломавируса». Биологическая химия . 398 (8): 919–927. doi : 10.1515/hsz-2017-0113 . ISSN 1431-6730 . PMID 28315855 . Wikidata Q39186071 .
- ^ Do HT, Koriyama C, Khan NA, Higashi M, Kato T, Le NT, Matsushita S, Kanekura T, Akiba S (январь 2013 г.). «Этиологическая роль папилломы человека при раке полового члена: исследование во Вьетнаме» . Британский журнал рака . 108 (1): 229–33. doi : 10.1038/bjc.2012.583 . PMC 3553541 . PMID 23299525 .
- ^ Гогилашвили К, Шония Н., Буркадзе Г (декабрь 2012 г.). «Роль папилломы человека в плоскоклеточном раке полости рта (обзор)». Грузинские медицинские новости (213): 32–6. PMID 23293230 .
- ^ Preti M, Rotondo JC, Holzinger D, Micheletti L, Gallio N, McKay-Chopin S, Pritaira C, Priviateira SS, Watanabe R, Ridder R, Pawlita M, Benedetto C, Tommasino M, Gheit T (2020). «Роль папилломавирусной инфекции человека в этиологии вульветного рака у итальянских женщин » Заражение агентами рак 20 : 20. doi : 10.1186/ s13027-020-00286-8 7110671PMC 32266002PMID
- ^ Толстов Ю., Хадаскик Б., Пахерник С., Хоэнфеллнер М., Дуэнсинг С. (январь 2014 г.). «Человеческие папилломавирусы в урологических злокачественных новообразованиях: критическая оценка». Урологическая онкология . 32 (1): 46.E19–27. doi : 10.1016/j.urolonc.2013.06.012 . PMID 24140249 .
- ^ «PDB101: Молекула месяца: папилломавирус человека и вакцины» . RCSB: PDB-101 . Получено 2018-05-14 .
- ^ Guan J, Bywaters SM, Brendle SA, Ashley RE, Makhov AM, Conway JF, Christensen ND, Hafenstein S (6 декабря 2017 г.). «Анализ структуры с высоким разрешением конформационного эпитопа антитела V5 на папилломавирусе человека 16» . Вирусы . 9 (12): 374. doi : 10.3390/v9120374 . PMC 5744149 . PMID 29211035 .
- ^ Meyers C, Frattini MG, Hudson JB, Laimins LA (август 1992). «Биосинтез папилломы человека из непрерывной клеточной линии при эпителиальной дифференцировке». Наука . 257 (5072): 971–3. Bibcode : 1992sci ... 257..971m . doi : 10.1126/science.1323879 . PMID 1323879 .
- ^ McLaughlin-Drubin Me, Christensen ND, Meyers C (май 2004). «Распространение, инфекция и нейтрализация подлинного вируса HPV16» . Вирусология . 322 (2): 213–9. doi : 10.1016/j.virol.2004.02.011 . PMID 15110519 .
- ^ Goodsell, DS (2018). «Человеческий папилломавирус и вакцины». RCSB Белковой банк данных . doi : 10.2210/rcsb_pdb/mom_2018_5 .
- ^ Jump up to: а беременный в Zheng ZM, Baker CC (сентябрь 2006 г.). «Структура генома папилломы, экспрессия и посттранскрипционная регуляция» . Границы в биологии . 11 : 2286–302. doi : 10.2741/1971 . PMC 1472295 . PMID 16720315 .
- ^ Jump up to: а беременный Тан С., Тао М, Маккой Дж.П., Чжэн Зм (май 2006 г.). «Онкопротеин E7 переводится из сплайсированных транскриптов E6*I в папилломе высокого риска папилломавируса типа 16 или типа 18-позитивных клеточных клеточных линий рака шейки матки посредством повторной повторной трансляции» . Журнал вирусологии . 80 (9): 4249–63. doi : 10.1128/jvi.80.9.4249-4263.2006 . PMC 1472016 . PMID 16611884 .
- ^ Wang X, Meyers C, Wang HK, Chow LT, Zheng ZM (август 2011 г.). «Строительство полной транскрипционной карты папилломы человека типа 18 типа во время продуктивной вирусной инфекции» . Журнал вирусологии . 85 (16): 8080–92. doi : 10.1128/jvi.00670-11 . PMC 3147953 . PMID 21680515 .
- ^ Цзя Р., Лю Х, Тао М., Крухлак М., Го М., Мейерс С., Бейкер К.К., Чжэн Зм (январь 2009 г.). «Контроль папилломы в раннем-до-позднем переключении дифференциально экспрессированным SRP20» . Журнал вирусологии . 83 (1): 167–80. doi : 10.1128/jvi.01719-08 . PMC 2612334 . PMID 18945760 .
- ^ McBride AA, McPhillips MG, Oliveira JG (декабрь 2004 г.). «BRD4: привязка, сегрегация и за его пределами». Тенденции в микробиологии . 12 (12): 527–9. doi : 10.1016/j.tim.2004.10.002 . PMID 15539109 .
- ^ Дверь Дж., Кэмпбелл Д., Гранд Р.Дж., Галлимор PH (февраль 1986 г.). «Идентификация продуктов гена вируса папилломы человека-1A E4» . Embo Journal . 5 (2): 355–62. doi : 10.1002/j.1460-2075.1986.tb04219.x . PMC 1166739 . PMID 3011404 .
- ^ Bravo IG, Alonso A (декабрь 2004 г.). «Слизистые папилломавирусы человека кодируют четыре различных белка E5, химия и филогения которых коррелируют со злокачественным или доброкачественным ростом» . Журнал вирусологии . 78 (24): 13613–26. doi : 10.1128/JVI.78.24.13613-13626.2004 . PMC 533923 . PMID 15564472 .
- ^ "PDB 2i0i" .
- ^ Jump up to: а беременный в Гупта С., Тахар П.П., Дегенколбе Р., Ко Ч.Х., Циммерманн Х., Ян С.М., Гуан Сим К, Хсу С.И., Бернард Ху (декабрь 2003 г.). «Белки Papillomavirus Papillomavirus 11 и 16 типа типа 11 и 16 модулируют регулятор клеточного цикла и транскрипционную кофактор Trip Br1» . Вирусология . 317 (1): 155–64. doi : 10.1016/j.virol.2003.08.008 . PMID 14675634 .
- ^ Jump up to: а беременный в Glaunsinger BA, Lee SS, Thomas M, Banks L, Javier R (ноябрь 2000 г.). «Взаимодействие PDZ-протеина Magi-1 с аденовирусом E4-ORF1 и онкопротеинами Papillomavirus E6 высокого риска» . Онкоген . 19 (46): 5270–80. doi : 10.1038/sj.onc.1203906 . PMC 3072458 . PMID 11077444 .
- ^ «IHOP Информация о гиперссылке по белкам UBE3A» . Архивировано из оригинала 2007-09-27 . Получено 2007-05-01 .
- ^ «Биохимия, Ноттингемский университет - 3,0 ферменты пути убиквитина» . Архивировано из оригинала на 2007-05-06 . Получено 2007-05-01 .
- ^ Келли М.Л., Кейгер К.Е., Ли С.Дж., Хуибрегтс Дж. М. (март 2005 г.). «Глобальные транскрипционные эффекты белка Papillomavirus E6 человека в клеточных линиях карциномы шейки матки опосредованы убиквитин -лигазой E6AP» . Журнал вирусологии . 79 (6): 3737–47. doi : 10.1128/JVI.79.6.3737-3747.2005 . PMC 1075713 . PMID 15731267 .
- ^ Jump up to: а беременный Kiyono T, Hiraiwa A, Fujita M, Hayashi Y, Akiyama T, Ishibashi M (октябрь 1997 г.). «Связывание онкопротеинов PAPILLOMAVIRUS E6 человека с высоким риском с гомологом человека Drosophila Disces крупного белка-супрессора опухоли» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (21): 11612–6. Bibcode : 1997pnas ... 9411612K . doi : 10.1073/pnas.94.21.11612 . PMC 23554 . PMID 9326658 .
- ^ Нишимура А., Накахара Т., Уэно Т., Сасаки К., Йошида С., Кё С., Хоули П.М., Сакай Х (апрель 2006 г.). «Требование E7 онкопротеина для жизнеспособности клеток HeLa» . Микробы и инфекция . 8 (4): 984–93. doi : 10.1016/j.micinf.2005.10.015 . PMID 16500131 .
- ^ Джексон М.Е., Пенни В.Д., МакКаффери Р.Е., Смит К.Т., Гриндлей Г.Дж., Кампо М.С. (1991). «Bovine Papillomavirus Bovine Bovine не имеют идентифицируемой рамки открытого считывания E6». Молекулярный канцерогенез . 4 (5): 382–7. doi : 10.1002/mc.2940040510 . PMID 1654923 . S2CID 22514962 .
- ^ Картер JJ, WIPF GC, Мадлен М.М., Шварц С.М., Каутски Л.А., Галлоуэй Д.А. (май 2006 г.). «Идентификация человеческого папилломавируса типа 16 -го типа 16 L1, необходимых для нейтрализации человеческой сыворотой» . Журнал вирусологии . 80 (10): 4664–72. doi : 10.1128/jvi.80.10.4664-4672.2006 . PMC 1472072 . PMID 16641259 .
- ^ Кампос С.К., Озбун М.А. (2009). Papavasiliou n (ред.). «Два высоко консервативных остатка цистеина в HPV16 L2 образуют внутримолекулярную дисульфидную связь и имеют решающее значение для инфекционности в кератиноцитах человека» . Plos один . 4 (2): E4463. Bibcode : 2009ploso ... 4.4463c . doi : 10.1371/journal.pone.0004463 . PMC 2636891 . PMID 19214230 .
- ^ Ричардс Р.М., Лоуи Д.Р., Шиллер Дж.Т., День Прем (январь 2006 г.). «Расщепление папилломавирусного капсидного белка, L2, в консенсусном сайте фурин, необходим для инфекции» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (5): 1522–7. Bibcode : 2006pnas..103.1522r . doi : 10.1073/pnas.0508815103 . PMC 1360554 . PMID 16432208 .
- ^ Yang R, Yutzy WH, Viscidi RP, Roden RB (апрель 2003 г.). «Взаимодействие L2 с бета-актином направляет внутриклеточный транспорт папилломы и инфекции» . Журнал биологической химии . 278 (14): 12546–53. doi : 10.1074/jbc.m208691200 . PMID 12560332 .
- ^ Боссис I, Роден Р.Б., Гамбхира Р., Ян Р., Тагая М., Хоули П.М., Менесес П.И. (июнь 2005 г.). «Взаимодействие синтаксина TSNARE 18 с папилломавирусом -малым капсидным белком опосредует инфекцию » Журнал вирусологии 79 (11): 6723–3 Doi : 10.1128/ jvi.79.11.6723-6 1112158PMC 15890910PMID
- ^ Пастрана Д.В., Гамбхира Р., Бак К.Б., Панг Й.Ю., Томпсон К.Д., Калп Т.Д., Кристенсен Н.Д., Лоуи Д.Р., Шиллер Дж.Т., Роден Р.Б. (июль 2005 г.). «Крестная нейтрализация кожных и слизистой оболочки папилломавируса с антисером в амино-кончике L2» . Вирусология . 337 (2): 365–72. doi : 10.1016/j.virol.2005.04.011 . PMID 15885736 .
Внешние ссылки
[ редактировать ]- ICTV сообщает Papillomaviridae
- Вирус : Papillomaviridae
- Национальная лаборатория Лос -Аламоса поддерживает комплексную (хотя и несколько датированную) базу данных последовательности папилломавируса . Эта полезная база данных содержит подробные описания и ссылки для различных типов папилломы.
- Короткое видео, которое показывает влияние папилломы на кожу индонезийского человека с эпидермодисплазией verruciformis , генетической неспособности защищаться от некоторых типов кожного ВПЧ.
- Лучшая суставная добавка, которая на самом деле работает для мужчин, женщин и колена де Вильерса, Э.М., Бернарда, Ху, Брокера, Т., Делиуса, Х. и Зура Хаузена, Х. Индекс вирусов - Papillomaviridae (2006). В кн.: ICTVDB-база данных универсального вируса, версия 4. Büchen-Osmond, C (Ed), Колумбийский университет, Нью-Йорк, США.
- 00.099. Papillomaviridae Описание в: ICTVDB-база данных универсальных вирусов, версия 4. Büchen-Osmond, C. (ed), Колумбийский университет, Нью-Йорк, США.
- Человеческие частицы папилломы и визуализация генома
- ICTV
| Базы данных управления авторитетом : национальный |
|---|