Лист



- Вершина
- Средний век (первичная вена)
- Вторичная вена.
- Пластинка.
- Края листья
- Черешка
- Бутон
- Корень

Внизу: капуста скунса, Symplocarpus fetidus (простая похвала)
- Вершина
- Первичная вена
- Вторичная вена
- Пластинка
- Края листья
- Рахис
Лист , ( пл.: Листья ) основным придаткой стебля сосудистого растения является [ 1 ] Обычно носится с боковым надземным и специализируется на фотосинтез . Листья коллективно называются листвой , как в «Осенней лиственности», [ 2 ] [ 3 ] в то время как листья, стебель, цветок и фрукты коллективно образуют систему побега . [ 4 ] В большинстве листьев основной фотосинтетической тканью является мезофилл палисада и расположена на верхней стороне лезвия или пластинки листа [ 1 ] но у некоторых видов, включая зрелую листвование эвкалипта , [ 5 ] Палисаде мезофилл присутствует с обеих сторон, и листья говорят, что являются изобилатеральными. Большинство листьев сплющены и имеют отчетливые верхние ( адаксиальные ) и нижние ( абаксиальные ) поверхности, которые различаются по цвету, волосатости, количеству устьиц (поры, которые потребляют и выходные газы), количество и структуру эпикутикулярного воска и другие особенности. Листья в основном зеленые цвета из -за присутствия соединения, называемого хлорофиллом , которое необходимо для фотосинтеза, поскольку он поглощает энергию света от солнца . Лист с более светлыми или белыми пятнами или краями называется пестрым листом .
Листья могут иметь много разных форм, размеров, текстур и цветов. Широкие, плоские листья со сложным веновлением цветущих растений известны как мегафиллы и виды, которые их несут, большинство, как широколистные или мегафиллические растения, которые также включают акрогимноспермы и папоротники . В ликоподах , с различным эволюционным происхождением листья просты (только с одной веной) и известны как микрофиллы . [ 6 ] Некоторые листья, такие как лампочки , не находятся над землей. У многих водных видов листья погружены в воду. Сочные растения часто имеют густые сочные листья, но некоторые листья имеют неэффективную функцию фотосинтеза и могут быть мертвы в зрелости, как у некоторых катафиллов и шипов . Кроме того, несколько видов листоподобных структур, обнаруженных в сосудистых растениях, не полностью гомологичны с ними. Примеры включают сплющенные стебли растений, называемые филлокладами и кладодами , а также сплющенные стебли листьев, называемые филлоками , которые отличаются от листьев как по своей структуре, так и в происхождении. [ 3 ] [ 7 ] Некоторые структуры не сосудистых растений выглядят и функционируют так же, как листья. включают филлиды мхов и . печеночныхвортов Примеры
Общие характеристики
[ редактировать ]Листья являются наиболее важными органами большинства сосудистых растений. [ 8 ] Зеленые растения являются автотрофными , что означает, что они не получают пищи от других живых существ, а вместо этого создают свою собственную пищу с помощью фотосинтеза . Они захватывают энергию в солнечном свете и используют ее для изготовления простых сахаров , таких как глюкоза и сахароза , из углекислого газа и воды. Затем сахара хранятся как крахмал , дополнительно обрабатывая химическим синтезом в более сложные органические молекулы, такие как белки или целлюлоза , основной структурный материал в клеточных стенках растений или метаболизируется клеточным дыханием , чтобы обеспечить химическую энергию для запуска клеточных процессов. Листья вытягивают воду из земли в потоке транспирации через систему сосудистой проводящей работы, известную как ксилем , и получают диоксид углерода из атмосферы путем диффузии через отверстия, называемые устьями во внешнем покрывающем слое листа ( эпидермис ), в то время как листья ориентированы, чтобы максимизировать максимизировать, чтобы максимизировать их воздействие на солнечный свет. Как только сахар синтезирован, его необходимо транспортировать в районы активного роста, такие как побеги растений и корни . Сосудистые растения транспортируют сахарозу в специальной ткани, называемой флоэмой . Флоэма и ксилема параллельны друг другу, но перенос материалов обычно находится в противоположных направлениях. Внутри листа эти сосудистые системы ветвь (Ramify) для образования вен, которые поставляют как можно больше листа, гарантируя, что клетки, проводящие фотосинтез, близки к транспортной системе. [ 9 ]
Обычно листья широкие, плоские и тонкие (дорсивентрально сплющенные), тем самым максимизируя площадь поверхности, непосредственно подвергаясь свету и позволяя свету проникать в ткани и достигать хлоропластов , что способствует фотосинтезу. Они расположены на растении, чтобы выявлять свои поверхности как можно эффективно, не затеняя друг друга, но есть много исключений и осложнений. Например, растения, адаптированные к ветреным условиям, могут иметь листья подтяжки , например, во многих ивах и эвкалиптах . Плоская или ламинарная форма также максимизирует тепловой контакт с окружающим воздухом , способствуя охлаждению. Функционально, в дополнение к проведению фотосинтеза, лист является основным местом транспирации , обеспечивая энергию, необходимую для вытягивания потока транспирации от корней, и гуттации .
Многие хвойные деревья имеют тонкие иглы, похожие на иглу или масштабные листья, которые могут быть полезны в холодном климате с частым снегом и заморозом. [ 10 ] Они интерпретируются как уменьшенные от мегафиллевых листьев их девонских предков. [ 6 ] Некоторые формы листьев адаптированы для модуляции количества света, которое они поглощают, чтобы избежать или смягчить чрезмерное тепло, ультрафиолетовое повреждение или высыхание, или для жертвоприношения эффективности поглощения света в пользу защиты от травоядных. Для ксерофитов основным ограничением является не поток света или интенсивность , а засуха. [ 11 ] Некоторые оконные растения, такие как виды Fenestraria и некоторые виды Haworthia , такие как Haworthia Tesselata и Haworthia Truncata, являются примерами ксерофитов. [ 12 ] и Bulbine Mesembryanthemoides . [ 13 ]
Листья также функционируют для хранения химической энергии и воды (особенно в суккулентах ) и могут стать специализированными органами, выполняющими другие функции, такие как усики гороха и других бобовых, защитные шипы кактусов и ловушки насекомых в плотоядных растениях, таких как Nepenthes и Sarracenia . [ 14 ] Листья являются фундаментальными структурными единицами, из которых конусы построены в спортивных полках (каждая шкала конуса представляет собой модифицированный мегафилл лист, известный как спорофилл) [ 6 ] : 408 и из которых цветы построены в цветущих растениях . [ 6 ] : 445

Внутренняя организация большинства видов листьев развивалась, чтобы максимизировать воздействие фотосинтетических органелл , хлоропластов , освещения и увеличения поглощения углекислого газа , одновременно контролируя потерю воды. Их поверхности гидроизоляются кутикулой растений и газовым обменом между мезофилльными клетками, а атмосфера контролируется мельчайшим (длиной и шириной, измеренной в десятках мкМ), называемых устьиками , которые открываются или близко к регулированию обмена скоростью углекиса (CO 2 ), кислород (O 2 ) и водяной пар во внутреннюю межклеточную космическую систему. Овергание в упирке контролируется давлением тургора в паре охраняемых ячеек , которые окружают апертуру устья. В любом квадратном сантиметре растения листья могут быть от 1000 до 100 000 устьев. [ 15 ]


Форма и структура листьев значительно варьируются от видов к видам растения, в зависимости от их адаптации к климату и доступному свету, а также от других факторов, таких как пастбищные животные (такие как олени), доступные питательные вещества и экологическая конкуренция со стороны других растений Полем Значительные изменения в типе листьев также происходят в видах, например, в качестве созревания растения; В качестве примера виды эвкалипта обычно имеют изобилатеральные листья свинца, когда зрелые и доминирующие в своих соседях; Тем не менее, такие деревья, как правило, имеют прямые или горизонтальные дорсивентральные листья в качестве саженцев, когда их рост ограничен доступным светом. [ 16 ] Другие факторы включают необходимость сбалансировать потерю воды при высокой температуре и низкую влажность против необходимости поглощения атмосферного углекислого газа. У большинства растений листья также являются основными органами, ответственными за транспирацию и гуттацию (шарики жидкости, образующиеся на краях листа).
Листья также могут хранить пищу и воду и соответственно модифицированы для выполнения этих функций, например, в листьях суккулентных растений и в ламповых масштабах. Концентрация фотосинтетических структур в листьях требует, чтобы они были богаче в белках , минералах и сахарах, чем, скажем, древесные ткани стебля. Соответственно, листья заметны в рационе многих животных .

Соответственно, листья представляют собой тяжелые инвестиции со стороны растений, несущих их, и их удержание или расположение являются предметом сложных стратегий для борьбы с давлением вредителей, сезонными условиями и защитными мерами, такими как рост шипов и производство фитолитов , Лигнины , танины и яды .
Лиственные растения в холодных или холодных умеренных областях обычно проливают листья осенью, тогда как в районах с тяжелым сухим сезоном некоторые растения могут сбросить листья до конца сухого сезона. В любом случае, можно ожидать, что листья сарая будут вносить свои удержанные питательные вещества в почву, где они падают.

Напротив, многие другие не сезонные растения, такие как ладони и хвойные, сохраняют свои листья на длительные периоды; Welwitschia сохраняет два основных листья на протяжении всей жизни, которые могут превышать тысячу лет.
Листоподобные органы бриофитов (например, мх и печеночных веществ ), известные как филлиды , сильно морфологически отличаются от листьев сосудистых растений . В большинстве случаев им не хватает сосудистой ткани, имеют только одну клеточную толщину и не имеют кутикулы , устьиц или внутренней системы межклеточных пространств. (Филлиды семейства Мосс Политрихасей являются заметными исключениями.) Филлиды бриофитов присутствуют только на гаметофитах , в то время как листья сосудистых растений присутствуют только на спорофитах . Они могут дополнительно развиваться в вегетативные или репродуктивные структуры. [ 14 ]
Простые, сосудистые листья ( микрофиллы ), такие как лиды раннего девонского ликопсида Барагванатии , сначала развивались в виде источников, расширения стебля. Истинные листья или эйфгиллы большего размера и с более сложным венерированием не стали широко распространенными в других группах до периода Девон , к этому времени концентрация углекислого газа в атмосфере значительно упала. Это произошло независимо в нескольких отдельных линиях сосудистых растений, у прогимносперм , таких как Archaeopteris , в Sphenopsida , папоротниках , а затем в спортивных зданиях и покрытосеменных . Euphylls также называют макрофиллами или мегафиллами (большие листья). [ 6 ]
Морфология
[ редактировать ]

Структурно полный лист покрыто ассоренности состоит из черешка (стебель листьев), пластинки (листовой лезвие), оговорки (небольшие сооружения, расположенные по обе стороны основания черешка) и оболочки. Не каждый вид производит листья со всеми этими структурными компонентами. Проксимальный стебель или черешки называют стип в папоротниках . Пластинка - это расширенный, плоский компонент листа, который содержит хлоропласты . Оболочка представляет собой структуру, обычно у основания, которая полностью или частично закрывает стебель над узлом, где прикреплен лист. Листовые ножны обычно встречаются в Poaceae (травы) и Apiaceae (Umbellifers). Между оболочкой и пластинкой может быть псевдопетиолевая , чашка, похожая на структуру. Псевдопетиолы встречаются в некоторых монокотидонах, включая бананы , ладони и бамбуки . [ 18 ] Скоры могут быть заметными (например, бобы и розы ), вскоре падают или иным образом не очевидны, как у Moraceae или вообще отсутствуют, как у Magnoliaceae . Мереола может отсутствовать (апетиолат), или лезвие не может быть ламинарным (сплюснутым). Черерьи механически связывают лист с растением и обеспечивают маршрут для переноса воды и сахаров в лист и обратно. Пластинка, как правило, является местоположением большинства фотосинтеза. Верхний ( адаксиальный ) угол между листом и стеблем известен как пазуть листа. Это часто расположение бутона . Структуры, расположенные там, называются «подмышечным».
Внешние характеристики листьев, такие как форма, края, волосы, черешки и наличие охраны и желез, часто важны для идентификации растений для семейства, рода или уровней видов , а ботаники разработали богатую терминологию для описания характеристик листьев. Листья почти всегда имеют определение роста. Они растут до определенного рисунка и формы, а затем останавливаются. Другие части растений, такие как стебли или корни, имеют неопределенный рост и обычно продолжают расти до тех пор, пока у них есть ресурсы для этого.
Тип листья обычно характерен для вида (мономорфного), хотя некоторые виды производят более одного типа листьев (диморфный или полиморфный ). Самые длинные листья - это листья Raffia Palm , R. Regalis , которые могут иметь длину до 25 м (82 фута) и шириной 3 м (9,8 фута). [ 19 ] Терминология, связанная с описанием морфологии листьев, представлена в иллюстрированной форме, в Wikibooks .

Там, где листья базальны, и лежат на земле, они называются простратными .
Основные типы листьев
[ редактировать ]
многолетние растения Говорят, что , листья которых проливаются ежегодно, имеют лиственные листья, в то время как листья, которые остаются через зиму, являются вечнозелеными растениями . Листья, прикрепленные к стеблям стеблями (известными как черешки ), называются черешками, и, если они прикреплены непосредственно к стебле без черешками, их называют сидячими. [ 20 ]
- Папоротники имеют листья .
- Листья хвойного дерева, как правило, являются иглами или AWL в форме или в масштабе, они обычно являются вечнозеленой, но иногда могут быть лиственными. Обычно у них есть одна вена.
- Цветочное растение (покрытоперстное) листья: стандартная форма включает в себя стапулы , черешка и пластинка .
- Ликофиты имеют микрофиллы .
- Листья оболочки - это тот тип, который можно найти в большинстве трав и многих других монокотов.
- Другие специализированные листья включают листья Непентеса , растения кувшина.
Листья DiCot имеют лезвия с вднацией верната (где основные вены расходятся с одной большой середины ввеина и имеют меньшие соединительные сети между ними). Реже, лезвия листьев DiCot могут иметь вденцию пальмы (несколько больших вен, расходящихся от черешка к краям листа). Наконец, некоторые экспонируют параллельное венецию. [ 20 ]
Листья монокота в умеренном климате обычно имеют узкие лезвия, и обычно параллельное венечение, сходящее на кончиках или краях листьев. У некоторых также есть вднащение. [ 20 ]
Договоренность на стебле
[ редактировать ]Расположение листьев на стебле известно как филлотаксис . [ 21 ] Большое разнообразие филлотаксических паттернов встречается в природе:


- Альтернативный
- Один лист, ветвь или цветочная часть прикрепляется в каждой точке или узле на стебле и оставляет альтернативное направление, в большей или меньшей степени, вдоль стебля.
- Базал
- Возникает из основания растения.
- Каулин
- Прикреплен к воздушному стеблу.
- Противоположный
- Два листа, ветви или части цветов прикрепляются в каждой точке или узле на стебле. Листовые вложения в паре в каждом узле.
- Отключен
- Противоположное расположение, при котором каждая последовательная пара повернута на 90 ° от предыдущего.
- Сбитый или вертилятный
- Три или более листьев, ветвей или деталей цветов прикрепляются в каждой точке или узле на стебле. Как и в случае с противоположными листьями, последовательные обороты могут быть или не быть декауссами, вращающиеся на половину угла между листьями в обороте (то есть последовательные обороты из трех вращающихся 60 °, оборотов из четырех вращающихся 45 ° и т. Д.). Противоположные листья могут появиться в восторге от кончика стебля. Pseudoverticillate описывает аранжировку, появляющуюся только с виртурой, но на самом деле не так.
- Розолат
- Листья образуют розетку .
- Ряды
- Термин, дистич , буквально означает два ряда . Листья в этом расположении могут быть альтернативными или противоположными в их прикреплении. Термин, 2-й рейтинг , эквивалентен. Термины, тристихные и тетрастишные , иногда встречаются. Например, «листья» (на самом деле микрофиллы ) большинства видов Selaginella являются тетрастишными, но не декауссами.
В простых математических моделях филлотаксиса вершина стебля представлена в виде круга. Каждый новый узел образуется на вершине, и он вращается под постоянным углом из предыдущего узла. Этот угол называется углом дивергенции . Количество листьев, которые растут из узла, зависит от видов растений. Когда один лист вырастет из каждого узла, и когда стебель удерживается прямо, листья образуют спираль .
Угол дивергенции часто представлен как доля полного вращения вокруг стебля. Фракция вращения 1/2 (угол дивергенции 180 °) дает альтернативное расположение, например, в Гастеррии или фанате-алоэ Кумара Пликатилис . Фракции вращения 1/3 (углы дивергенции 120 °) встречаются в буже и камере . Дуб и абрикос вращаются на 2/5, подсолнухи, тополь и груша на 3/8, а в иве и миндале доля - 5/13. [ 22 ] Эти меры периодические. Знаменатель сделанных за фракции вращения указывает количество листьев за один период, в то время как числитель указывает количество полных поворотов или кругов, один период. Например:
- 180 ° (или 1 ⁄ 2 ): два листа в одном круге (альтернативные листья)
- 120 ° (или 1 ⁄ 3 ): три листа в одном круге
- 144 ° (или 2 ⁄ 5 ): пять листьев в двух кругах
- 135 ° (или 3 ~ 8 ): восемь листьев в трех кругах.
Большинство углов дивергенции связаны с последовательности чисел Фибоначчи f n . Эта последовательность начинается 1, 1, 2, 3, 5, 8, 13; Каждый термин является суммой предыдущих двух. Фракции вращения часто являются коэффициентами для N / F N + 2 числа Fibonacci на два термина позже в последовательности. Это относится к фракциям 1/2, 1/3, 2/5, 3/8 и 5/13. Соотношение между последовательными числами Фибоначчи имеет тенденцию к золотому соотношению φ = (1 + √5)/2 . Когда круг делится на две дуги, длина которых находится в соотношении 1: φ , угол, образованный меньшим дугой, является золотым углом , который составляет 1/φ 2 × 360 ° ≈ 137,5 ° . Из -за этого многие углы дивергенции составляют приблизительно 137,5 ° .
У растений, где пара противоположных листьев растет из каждого узла, листья образуют двойную спираль. Если узлы не вращаются (фракция поворота нуля и угол дивергенции 0 °), две спирали становятся парой параллельных линий, создавая дифихное расположение, как у клена или оливковых деревьев. Чаще всего в декауссовом рисунке, в которой каждый узел вращается на 1/4 (90 °), как в базилике травы . Листья трехлетних растений, таких как Nerium Oleander, образуют тройную спираль.
Листья некоторых растений не образуют спирали. У некоторых растений угол дивергенции меняется по мере роста растения. [ 23 ] В Phyllotaxis Orixate, названном в честь Orixa japonica , угол дивергенции не является постоянным. Вместо этого это периодически и следует последовательности 180 °, 90 °, 180 °, 270 °. [ 24 ]
Подразделения лезвия
[ редактировать ]
Две основные формы листьев могут быть описаны с учетом того, как лопасть (пластинка) разделен. Простой лист имеет неразделенное лезвие. Тем не менее, лист может быть рассечен для образования доли, но промежутки между долями не достигают основной вены. Составной лист имеет полностью подразделенное лезвие, каждая листовка лезвия разделяется вдоль главной или вторичной вены. У листочек могут быть черешки и стойпели, эквиваленты черешками и условиями листьев. Поскольку каждая листовка может показаться простым листом, важно распознать, где происходит черепица, чтобы идентифицировать составной лист. Составные листья являются характерными для некоторых семейств более высоких растений, таких как Fabaceae . Средняя вена составного листа или листья , когда он присутствует, называется рахисом .
- Ладони
- Все листочки имеют общую точку прикрепления в конце черешка, излучаясь, как пальцы руки; Например, каннабис (конопля) и aesculus (Buckeyes).
- Пинн. Соединение
- Листочки расположены по обе стороны от основной оси или рахиса .
- Странная вершина
- С терминальной листовкой; Например, Fraxinus (пепел).
- Даже верно
- Отсутствует терминальная листовка; Например, Swietenia (красное дерево). Определенным типом ровного уровня является бифолиолат , где листья состоят только из двух листовок; Например, Hymenaea .
- Двусмысленное соединение
- Листья дважды делятся: листочки (технически « подсолины ») расположены вдоль вторичной оси, которая является одной из нескольких разветвленных от рахис. Каждая листовка называется вершиной . Группа пиннел по каждой вторичной вене образует утилизацию ; Например, Альбиция (шелковое дерево).
- Трилиат (или трифолиолат)
- Лист вершины с тремя листочками; Например, Trifolium (Clover), Laburnum (Laburnum) и некоторые виды Toxicodendron (например, ядовитый плющ ).
- Поверхность
- Пинново рассеканный в центральную вену, но с листочками не совсем отдельно; Например, Polypodium , Sorbus (белые лучи). В листьях в стиле центральная вена известна как средняя жилка .
Характеристики черешка
[ редактировать ]
Говорят , что листья, которые имеют черешка (стебель листьев), являются черешками .
Сидячие (эпотиолатные) листья не имеют черешка, а лезвие прикрепляется непосредственно к стеблу. Листья субпететалиала практически черешко или имеют чрезвычайно короткую черешка и могут казаться сидячими.
В сжатии или листьях пособия лезвие частично окружает стебель.
Когда основание листья полностью окружает стебель, говорят, что листья являются перфолированными , например, в эупатории Perfoliatum .
В листьях шерсть черешка прикрепляется к лезвию внутри края лезвия.
У некоторых видов акации , таких как дерево коа ( акация коа ), черешки расширяются или расширяются и функционируют как лезвия листьев; Они называются филлодом . Там могут или не могут быть нормальные листья листьев на кончике филлода.
Старость , представляет собой , присутствующая на листьях многих DiCotyledons придаток с каждой стороны у основания черешка, напоминающий небольшой лист. Стипулы могут быть продолжительными и не проливаться (оцениваемый лист, например, в розах и бобах ), или проливаться по мере расширения листа, оставляя шрам на стапике на веточку (лист изгнания). Ситуация, расположение и структура стапул называются «условиями».
- Бесплатный, боковой
- Как в гибискусе .
- Аднате
- Слился с базой черешка, как в Розе .
- Ochreate
- При условии, что охреа или формированные оболочки, как у Polygonaceae ; Например, ревень .
- Окружая базу черепах
- Межпетоаларный
- Между черешками двух противоположных листьев, как у Rubiaceae .
- Внутрипетиол
- Между черешками и стволом подтенка, как у Malpighiaceae .
Вены
[ редактировать ]


Вены (иногда называемые нервами) составляют одну из наиболее заметных признаков листьев. Вены в листе представляют собой сосудистую структуру органа, простирающиеся в лист через черешка и обеспечивая транспортировку воды и питательных веществ между листами и стеблем, и играют решающую роль в поддержании статуса воды листьев и фотосинтетической способности. Они также играют роль в механической поддержке листа. [ 25 ] [ 26 ] В рамках пластинки листа, в то время как некоторые сосудистые растения обладают только одной веной, в большинстве случаев в этой сосудистой сети обычно (разветвляется) в соответствии с различными паттернами (венециями) и формирует цилиндрические пучки, обычно лежащие в медианной плоскости мезофилла , между двумя слоями эпидермиса . [ 27 ] Эта схема часто специфична для таксонов, и из которых покрытосеменные имеют два основных типа, параллельные и сетчатые (необработанные). В целом, параллельное венеция типична для монокотов, в то время как сетчатые, более типичные для эвдикотов и магнолидов (« дикоты »), хотя есть много исключений. [ 28 ] [ 27 ] [ 29 ]
Вена или вены, попадающие в лист из черешка, называются первичными венами или венами первичного или первого порядка. Вены разветвляются из них вторичные или вены второго порядка. Эти первичные и вторичные вены считаются основными венами или венами более низкого порядка, хотя некоторые авторы включают третий порядок. [ 30 ] Каждое последующее ветвление последовательно пронумеровано, и это вены более высокого порядка, каждый разветвление связано с более узким диаметром вены. [ 31 ]
В параллельных вуженевых листьях первичные вены работают параллельными и равноудаленными друг к другу для большей части длины листа, а затем сходится или сливаются (анастомоза) к вершине. Обычно многие меньшие незначительные вены соединяют эти первичные вены, но могут прекращаться с очень тонкими окончаниями в мезофилле. Незначительные вены более типичны для покрытосеменных, которые могут иметь до четырех более высоких порядков. [ 30 ]
Напротив, листья с сетчатым веновлением имеют одну (иногда более) первичную вену в центре листа, называемая средней жилью или коемой, которая непрерывна с сосудистой сетью черешка. Вторичные вены, также известные как вены второго порядка или боковые вены, разветвляются от средней жилки и простираются к краям листа. Они часто заканчиваются в гидатоде , секреторном органе, на краю. В свою очередь, меньшие вены веются из вторичных вен, известных как третичные или третий порядка (или более высокий порядок), образуя плотный сетчатый паттерн. Районы или острова Мезофилла, лежащие между венами более высокого порядка, называются азеолами . Некоторые из самых маленьких вен (вены) могут иметь свои окончания в ареолях, процесс, известный как ареляция. [ 31 ] Эти незначительные вены действуют как сайты обмена между мезофиллом и сосудистой системой растения. [ 26 ] Таким образом, второстепенные вены собирают продукты фотосинтеза (фотосинтивы) из клеток, где они происходят, в то время как основные вены отвечают за его транспорт за пределами листа. В то же время вода транспортируется в противоположном направлении. [ 32 ] [ 28 ] [ 27 ]
Количество окончаний вен очень изменчиво, как и вены второго порядка на краю или связываются с другими венами. [ 29 ] Существует много сложных вариаций на модели, которые образуют вены листовых, и они имеют функциональные последствия. Из них покрытосеменные имеют наибольшее разнообразие. [ 30 ] Внутри этого основные вены функционируют как сеть поддержки и распределения для листьев и коррелируют с формой листьев. Например, параллельное венецию, обнаруженное в большинстве монокотов, коррелирует с их удлиненной формой листьев и широким основанием листьев, в то время как сетчатое венеция наблюдается в простых целых листьях, в то время как листья цифровых средств обычно имеют венецию, в котором три или более первичные вены расходятся радиально из одной точки Полем [ 33 ] [ 26 ] [ 31 ] [ 34 ]
В эволюционном плане ранние возникающие таксоны имеют тенденцию иметь дихотомическое ветвление с ретикулированными системами, появляющимися позже. Вены появились в пермском периоде (299–252 млн лет), до появления покрытосеменных в триасе (252–201 млн лет), в ходе которого иерархия вен обеспечивала более высокие функции, больший размер листьев и адаптация к более широкому разнообразию условий климатических условий Полем [ 30 ] Хотя это более сложный рисунок, ветвящиеся вены, по -видимому, являются плезиоморфными и в той или иной форме присутствовали в древних семенах до 250 миллионов лет назад. Псевдо-ретикационное веневание, которое на самом деле является высоко измененным пеннипараллельным, представляет собой аутапоморфию некоторых Melanthiaceae , которые являются монокотами; Например, Paris Quadrifolia (узел истинного любовника). В листьях с сетчатыми венециями вены образуют матрицу каркасов, придающая механическую жесткость листьям. [ 35 ]
Изменения морфологии в одном растении
[ редактировать ]- Гомобластика
- Характеристика, в которой растение имеет небольшие изменения в размере листьев, форме и привычке роста между ювенильными и взрослыми этапами, в отличие от;
- Гетеробластия
- Характеристика, в которой растение имеет заметные изменения в размере листьев, форме и привычке роста между юношескими и взрослыми этапами.
Анатомия
[ редактировать ]Средние функции
[ редактировать ]Листья, как правило, широко сосудится и обычно имеют сети сосудистых пучков , содержащих ксилемы , которые обеспечивают воду для фотосинтеза , и Phloem , который транспортирует сахар, полученные с помощью фотосинтеза. Многие листья покрыты трихомами (маленькими волосками), которые имеют разнообразные структуры и функции.

Мелкие функции
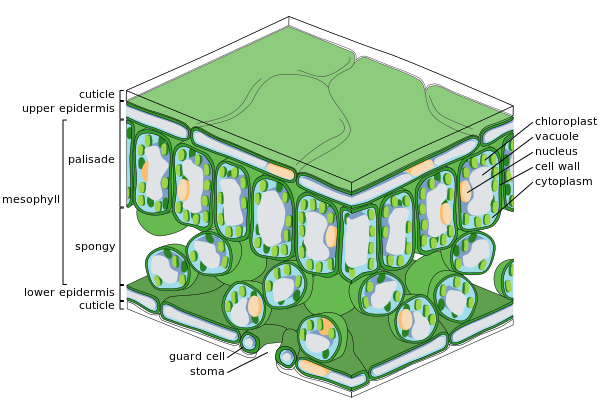
[ редактировать ]Основные присутствующие тканевые системы
- Эпидермис поверхность , который покрывает верхнюю и нижнюю
- Месофилл ткань , которая состоит из фотосинтетических клеток, богатых хлоропластами . (также называется хлорхимой )
- Расположение вен ( сосудистая ткань )
Эти три тканевые системы обычно образуют регулярную организацию в клеточной шкале. Специализированные клетки, которые заметно отличаются от окружающих клеток и часто синтезируют специализированные продукты, такие как кристаллы, называются идиоластами . [ 36 ]
Основные ткани листьев
[ редактировать ]-
 Поперечное сечение листа
Поперечное сечение листа -
 Эпидермальные клетки
Эпидермальные клетки -
 Губчатые мезофилл клетки
Губчатые мезофилл клетки



Эпидермис
[ редактировать ]
Эпидермис покрывающих является внешним слоем клеток, лист. Он покрыт восковой кутикулой , которая непроницаемой для жидкой воды и водяного пара и образует границу, отделяющую внутренние клетки растения от внешнего мира. Кутикула в некоторых случаях тоньше на нижнем эпидермисе, чем на верхнем эпидермисе, и, как правило, толще на листьях из сухого климата по сравнению с листьями из влажного климата. [ 37 ] Эпидермис выполняет несколько функций: защита от потери воды путем транспирации , регулирование газообмена и секрецию метаболических соединений. Большинство листьев показывают дорсовентуальную анатомию: верхние (адаксиальные) и нижние (абаксиальные) поверхности имеют несколько иную конструкцию и могут выполнять различные функции.
Ткань эпидермиса включает в себя несколько дифференцированных типов клеток; Эпидермальные клетки, эпидермальные волосковые клетки ( трихомы ), клетки в устьяльном комплексе; охраняемые ячейки и вспомогательные ячейки. Эпидермальные клетки являются наиболее многочисленными, крупнейшими и наименее специализированными и формируют большую часть эпидермиса. Они, как правило, более удлинены в листьях монокотов , чем в листьях дикотов .
Хлоропласты, как правило, отсутствуют в эпидермальных клетках, исключенным являются охраняющие клетки устьиц . Утомающие поры перфорируют эпидермис и окружены с каждой стороны, содержащими хлоропласт, и две-четыре вспомогательные клетки, в которых отсутствуют хлоропласты, образуя специализированную клеточную группу, известную как комплекс устьиц. Открытие и закрытие апертуры в устьице контролируется устьяльным комплексом и регулирует обмен газами и водяным парами между внешним воздухом и внутренней частью листа. Поэтому стомата играют важную роль в допуске фотосинтеза, не позволяя листьям высохнуть. В типичном листе устья более многочисленны в абаксиальном (нижнем) эпидермисе, чем адаксиальный (верхний) эпидермис и более многочисленны в растениях из холодного климата.
Мезофилл
[ редактировать ]Большая часть внутренней части листа между верхним и нижним слоями эпидермиса представляет собой паренхиму (земля) или ткань хлорхимы, называемая мезофиллом (греческий для «среднего листа»). Эта ткань ассимиляции является основным местоположением фотосинтеза на растении. Продукты фотосинтеза называются «ассимилированными».
У папоротников и большинства цветущих растений мезофилл разделен на два слоя:
- Верхний слой палисада вертикально удлиненных клеток, толщиной от одной до двух клеток, непосредственно под адаксиальным эпидермисом, с межклеточными воздушными пространствами между ними. Его клетки содержат гораздо больше хлоропластов, чем губчатый слой. Цилиндрические клетки с хлоропластами , близкими к стенкам клетки, могут использовать оптимальное преимущество света. Небольшое разделение клеток обеспечивает максимальное поглощение углекислого газа. Солнцезащитные листья имеют многослойный слой палисада, в то время как тенистые листья или более старые листья ближе к почве однослойные.
- Под палисадным слоем находится губчатый слой . Клетки губчатого слоя более разветвленные и не так плотно упакованы, так что между ними существуют большие межклеточные воздушные пространства. Поры или устья эпидермиса, открытые в подлиматальные камеры, которые связаны с межклеточными воздушными пространствами между губчатой и палисадной мезофиллевой клеткой, так что кислород, углекислый газ и водяной пары могут диффундировать в листь Во время дыхания, фотосинтез и транспирация.
Листья обычно зеленые из -за хлорофилла в хлоропластах в клетках мезофилла. Некоторые растения имеют листья разных цветов из -за присутствия вспомогательных пигментов, таких как каротиноиды в их мезофилле.
Сосудистая ткань
[ редактировать ]
Вены листа и расположены в представляют собой сосудистую ткань губчатом слое мезофилла. Образец вен называется Venation . У покрытосеменных, венеция обычно параллелен монокотидонам и образует взаимосвязанную сеть на широколистных растениях . Когда -то считалось типичным примером формирования схемы посредством последствий , но вместо этого они могут иллюстрировать образец, образованный в тензором стресса . [ 38 ] [ 39 ] [ 40 ]
Вена состоит из сосудистого пучка . В основе каждого пакета лежат кластеры двух Отдельные типы проводящих клеток:
- Ксилем
- Клетки, которые приносят воду и минералы из корней в лист.
- Флоэма
- Клетки, которые обычно перемещают сок с растворенной сахарозой (глюкоза в сахарозу), продуцируемые фотосинтезом в листе, из листа.
Ксилеми, как правило, лежит на адаксиальной стороне сосудистого пакета, а флоэма обычно лежит на абасиальной стороне. Оба встроены в плотную паренхиму, называемую оболочкой, которая обычно включает в себя некоторую структурную ткань колланхимы.
Развитие листьев
[ редактировать ]Согласно Агнес Арбера , листья являются частичными побегами, теории листьев [ 41 ] Быть полученным из зачатки листьев вершины побега. В начале развития они дорсивентально сплющены как дорсальными, так и вентральными поверхностями. [ 14 ] Составные листья ближе к побегам, чем простые листья. Исследования в области развития показали, что листья соединений, такие как побеги, могут разветвляться в трех измерениях. [ 42 ] [ 43 ] На основании молекулярной генетики, Эккардт и Баум (2010) пришли к выводу, что «теперь общепризнанно, что составные листья экспрессируют как листья, так и свойства побега». [ 44 ] Многие двудольные листья показывают эндогенно управляемую ежедневную ритмичность роста. [ 45 ] [ 46 ] [ 47 ]
Экология
[ редактировать ]Биомеханика
[ редактировать ]Растения реагируют и адаптируются к факторам окружающей среды, такими как свет и механическое напряжение от ветра. Листья должны поддерживать свою собственную массу и выровнять себя таким образом, чтобы оптимизировать их воздействие на солнце, как правило, более или менее горизонтально. Тем не менее, горизонтальное выравнивание максимизирует воздействие изгибающих сил и отказа от таких напряжений, как ветер, снег, град, падающий мусор, животные и истирание из окружающей листва и растительных сооружений. Общие листья относительно хрупкие в отношении других растительных структур, таких как стебли, ветви и корни. [ 48 ]
Как листовые лезвия, так и черепок влияют на реакцию листа на такие силы, как ветер, что позволяет определенной степени перемещения, чтобы минимизировать сопротивление и повреждение, в отличие от сопротивления. Подобное движение листьев может также увеличить турбулентность воздуха вблизи поверхности листа, которая зажигает пограничный слой воздуха, непосредственно прилегающий к поверхности, увеличивая способность к газу и теплообмену, а также фотосинтез. Сильные ветровые силы могут привести к уменьшению числа листьев и площади поверхности, которые, одновременно уменьшая сопротивление, включают обмен из также уменьшения фотосинтеза. Таким образом, конструкция листьев может включать компромисс между усилением углерода, терморегуляцией и потерей воды, с одной стороны, и стоимостью поддержания как статических, так и динамических нагрузок. У сосудистых растений перпендикулярные силы распространяются на большую площадь и являются относительно гибкими как при изгибе, так и при кручке , что позволяет упругое деформирование без повреждений. [ 48 ]
Многие листья полагаются на гидростатическую поддержку, расположенную вокруг скелета сосудистой ткани для их прочности, что зависит от поддержания статуса воды листовой воды. Как механика, так и архитектура листа отражают необходимость транспортировки и поддержки. Читайте и Стокс (2006) Рассмотрим две основные модели: форма «гидростатический» и «лист» I (см. Рис. 1). [ 48 ] Гидростатические листья, такие как в Простантере Ласиантос, являются большими и тонкими, и могут включать необходимость в нескольких листьях, довольно одиноких больших листьев из -за количества вен, необходимых для поддержки периферии больших листьев. Но большой размер листьев способствует эффективности фотосинтеза и сохранения воды, включая дальнейшие обмены. С другой стороны, листья I-балки, такие как Banksia marginata, включают специализированные конструкции, чтобы закрепить их. Эти I-балки образуются из наращивания пакетов Sclerenchyma , встречающихся закрепленных субэпидермальных слоев. Это меняет баланс от зависимости от гидростатического давления к структурной поддержке, что является очевидным преимуществом, когда вода относительно скудна. [ 48 ] Длинные узкие листья сгибаются легче, чем яйцевидные лопасти листьев той же области. Монокоты, как правило, имеют такие линейные листья, которые максимизируют площадь поверхности, с минимизацией самостирации. В этом высокая доля продольных основных вен обеспечивает дополнительную поддержку. [ 48 ]
Взаимодействие с другими организмами
[ редактировать ]
Хотя листья не так питательны, как другие органы, такие как фрукты, предоставляют источник пищи для многих организмов. Лист является жизненно важным источником производства энергии для растения, и растения развили защиту от животных, которые потребляют листья, такие как танины , химические вещества, которые препятствуют перевариванию белков и имеют неприятный вкус. Животные, специализирующиеся на употреблении листьев, известны как фоливоры .
Некоторые виды имеют загадочные адаптации, с помощью которых они используют листья, позволяющие избегать хищников. Например, гусеницы некоторых листовых мотыльков создадут небольшой дом в листе, сложив его над собой. Некоторые видели, как и листья их пищевых растений, в трубки. Женщины Attelabidae , так называемые долгоносики листьев, откладывают яйца в листья, которые затем свернуты как средства защиты. Другие травоядные и их хищники имитируют внешний вид листа. Рептилии, такие как некоторые хамелеоны, и насекомые, такие как некоторые катидиды , также имитируют колеблющиеся движения листьев на ветру, двигаясь из стороны в сторону или вперед и вперед, уклоняясь от возможной угрозы.
Сезонная потеря листьев
[ редактировать ]
Листья в умеренных , бореальных и сезонно сухих зонах могут быть сезонно лиственными (падение или умирание в течение сезона ненастных). Этот механизм для сброса листьев называется арестом . Когда лист проливается, он оставляет шрам на листьях на веточку. В холодных осени они иногда меняют цвет и становятся желтыми , ярко -оранжевыми или красными , в качестве различных вспомогательных пигментов ( каротиноидов и ксантофиллов ) выявляются, когда дерево реагирует на холодный и уменьшенный солнечный свет , ограничивая производство хлорофилла. красного антоцианина Считается, что пигменты производятся в листе, когда он умирает, возможно, чтобы замаскировать желтый оттенок, оставленный, когда хлорофилл теряется - желтые листья, по -видимому, привлекают травоядные животные, такие как тля . [ 49 ] Оптическое маскирование хлорофилла антоцианинами снижает риск фотоокисления к клеткам листьев, который в противном случае может снизить эффективность извлечения питательных веществ из сжигающих осенних листьев. [ 50 ]
Эволюционная адаптация
[ редактировать ]
В ходе эволюции листья адаптировались к разным средам следующим образом: [ Цитация необходима ]
- Восковые микро- и наноструктуры на поверхности уменьшают смачивание дождем и адгезией загрязнения ( см. Эффект лотоса ).
- Разделенные и составные листья уменьшают сопротивление ветра и способствуют охлаждению.
- Волосы на поверхности листья ловушки влажности в сухом климате и создают пограничный слой, уменьшающий потерю воды.
- Восковая кутикула растений уменьшает потерю воды.
- Большая площадь поверхности обеспечивает большую площадь для захвата солнечного света.
- На вредных уровнях солнечного света, специализированных листьев, непрозрачных или частично похороненных, допустите свет через полупрозрачное окно листьев для фотосинтеза на внутренних поверхностях листьев (например, Fenestraria ).
- Анатомия листьев Кранца у растений, которые выполняют C 4 углеродную фиксацию
- Сочные листья хранят воду и органические кислоты для использования в фотосинтезе CAM .
- Ароматические масла , яды или феромоны, произведенные листовыми железами, сдерживают травоядные животные (например, Eucalypts ).
- Включения кристаллических минералов сдерживают травоядные животные (например, фитолиты кремнезема в травах, рафиды в Araceae ).
- Лепестки привлекают опылителей.
- Колючки защищают растения от травоядных (например, кактусы ).
- Платы волос для защиты от травоядных, например, в Urtica Dioica и дендрокниде Мороиды ( Urticaceae ).
- Особые листья на плотоядных растениях адаптированы для ловушки пищи, в основном беспозвоночных добычи, хотя некоторые виды также затрагивают мелких позвоночных (см. Плотоядные растения ).
- Лампы хранят еду и воду (например, лук ).
- Усики позволяют растению подниматься (например, горох).
- Прицветники и псевдантия (ложные цветы) заменяют нормальные цветочные сооружения, когда истинные цветы значительно уменьшены (например , шпалы , шпалы в арасеаэ и цветочных головах в астереасах ).
Терминология
[ редактировать ]
Форма
[ редактировать ]
Край (поля)
[ редактировать ]Край - или край внешний периметр листа. Условия взаимозаменяемы.
| Изображение | Срок | латинский | Описание |
|---|---|---|---|
| Весь | Форма интеграция |
Даже; с гладким краем; без зуба | |
| Ресничный | Килиат | Бахрома с волосами | |
| Смягчить | Кренатус | Волнистые зубья; зубчатые с округлыми зубами | |
| сферат | Crenulatus | Мелко плен | |
| Crisped | кудрявый | Кудрявый | |
| Зубчатый | жесткий | Зубчатый;
может быть грубым зубным , имея большие зубы или железистые зубчатые зубцы , имеющие зубы, которые медведь железы | |
| Зубчатый | Denticulatus | Мелко зубчатые | |
| Вдвойной зубчик | удвоенный зуб | Каждый зуб с меньшими зубами | |
| Замораживание | Серратус | Пилообразный; с асимметричными зубами, указывающими вперед | |
| Серрута | Серрулатус | Мелко зубчатый | |
| Окрашивать | Подлежащий | С глубокими, волнообразными углублениями; Грубо размягчание | |
| Лобат | Лобатус | С отступами, когда отступления не достигают центра | |
| Внешний | undulatus | С волнистым краем, более мелким, чем синит | |
| Колючий или острый | Spiculatus | С жесткими, острыми точками, такими как чертополох |
Вершина (наконечник)
[ редактировать ]| Изображение | Срок | латинский | Описание |
|---|---|---|---|
| Закайтировать | _ | Давно установленная, продленная в узкую, сужающуюся точку вогнутым образом | |
| Острый | _ | Заканчиваться в острой, но не длительной точке | |
| Крус | _ | С острым, удлиненным, жестким наконечником; с | |
| Маргинальный | _ | С отступом, с мелкой выемкой на кончике | |
| Мукронат | _ | Внезапно с маленькой короткой точкой | |
| Мукроналат | _ | Мукронат, но с заметно уменьшенным позвоночником | |
| Согласие | _ | Обратно в форме сердца | |
| Тупой | _ | Округлый или тупой | |
| Усечь | _ | Внезапно заканчиваться плоским концом |
База
[ редактировать ]- Закайтировать
- Приходя в острую, узкую, длительную точку.
- Острый
- Приходя в острый, но не длинный пункт.
- Auciculate
- Ушная.
- Корпус
- Сердце с надписью к стеблу.
- Клин
- Клин в форме.
- Ускорить
- В форме, как алебарда и с базальными долями, указывающими наружу.
- Косой
- Наклон.
- Почка
- В форме почек, но круглая и шире, чем длинная.
- Округлый
- Изгибающаяся форма.
- Стрелец
- В форме стрелы и с острыми базальными долями, указывающими вниз.
- Усечь
- Окончательно заканчивая плоским концом, это выглядит отрезанным.
Поверхность
[ редактировать ]
Поверхность листьев также проводит большое разнообразие микроорганизмов ; В этом контексте это называется филлосферой .
- Лепидот
- Покрыто тонкими шкалами Scurfy.
Хакерс
[ редактировать ]


«Волосы» на растениях правильно называются трихомами . Листья могут показать несколько градусов волосатости. Значение нескольких из следующих терминов может перекрываться.
- Арахноидаль, или арахноза
- Со многими прекрасными, запутанными волосами, которые появляются с паутиной.
- Штанги
- С тонкими колючими волосками (шарниров).
- Бородатый
- С длинными, жесткими волосками.
- Щетично
- С жесткими волосами.
- Ясно
- Hoary с плотным серовато-белым опушением.
- Ресничный
- Незначительно бахрома с короткими волосками (реснички).
- Цилиол
- Минично оружейный.
- Стадо
- С стадами мягких, шерстяных волосков, которые, как правило, тереть.
- Гладко
- Потеря волос с возрастом.
- Голый
- Никаких волос любого рода.
- Желез
- С железой на кончике волос.
- Hirsute
- С довольно грубыми или жесткими волосками.
- Hispid
- С жесткими, щетинистыми волосами.
- Избитый
- Минично избитый.
- Хеар
- С тонким, близким серовато-белым опушением.
- Ланат, или ланоз
- С шерстяными волосами.
- Пилоз
- С мягкими, четко отделенными волосками.
- Половой или половой человек
- С прекрасными, мельчайшими волосами.
- Опушенные
- С мягкими, короткими и прямыми волосками.
- Scabrous, или Scabrid
- Грубо до прикосновения.
- Сериса
- Шелковистый вид через мелкие, прямые и прижатые (лежащие близко и плоские) волосы.
- Шелковистый
- С помощью Adpressed, Spect и Prome Occence.
- Stellate, или Stelliform
- С звездными волосками.
- Стриговый
- С придавленными, острыми, прямыми и жесткими волосками.
- Томентоза
- У плотно опушенные с матовыми, мягкими белыми шерстяными волосками.
- Кано-томеноза
- Между Canescent и Tomentose.
- Felted-Tomentose
- Шерстяные и спутанные с вьющимися волосами.
- Томонтулоза
- Подробно или только слегка Томентоза.
- Ворсинку
- С длинными и мягкими волосками, обычно изогнутыми.
- Шерстяное
- С длинными, мягкими и извилистыми или котовыми волосками.
Время
[ редактировать ]Венеция
[ редактировать ]Классификация
[ редактировать ]





Был описан ряд различных систем классификации моделей листовых вен (венеция или винация), [ 29 ] Начиная с Ettingshausen (1861), [ 53 ] вместе со многими различными описательными терминами, и терминология была описана как «грозная». [ 29 ] Одним из самых распространенных среди них является система Hickey, первоначально разработанная для « DiCotyledons » и с использованием ряда терминов Ettingshausen, полученных из греческого (1973–1979): [ 54 ] [ 55 ] [ 56 ] ( См. Также : Симпсон Рисунок 9.12, стр. 468) [ 29 ]
Hickey System
[ редактировать ]- 1. Странс (перья, сетчатая, сетка, сетка, пеннириблен, пеннинервена или пиненив)
- Вены возникают в пенсии (похожа на перьев) из одной первичной вены (средняя вен) и подразделяют во вторичные вены, известные как вены более высокого порядка. Они, в свою очередь, образуют сложную сеть. Этот тип венеции типичен для (но ни в коем случае не ограниченным) « DiCotyledons » (Non Monocotyledon Angiosperms ). Например, Остры . Есть три подтипа вднат:
- Craspedodromous (греческий: Kraspedon - Edge, Dromos - бег)
- Основные вены достигают края листа.
- Камптодром
- Основные вены простираются близко к краю, но сгибаются, прежде чем они пересекаются с краем.
- Гифодром
- Все вторичные вены отсутствуют, рудиментарны или скрыты
Они, в свою очередь, имеют ряд дополнительных подтипов, таких как eucamptodromous, где вторичные вены кривой возле края без соединения соседних вторичных вен.



- 2. Параллеломодочный (параллельный, параллельный, параллельный, выросший, пеннипараллельный, полосатый)
- Две или более первичных вен, происходящих рядом друг с другом у основания листьев и бегут параллельно друг другу с вершиной, а затем сходится туда. Комиссуральные вены (небольшие вены) соединяют основные параллельные вены. Типично для большинства монокотидонов , таких как травы . Дополнительные термины маргинальные (первичные вены достигают поля), а также используются сетчатые (сетчатые).

- 3. Campylodromous ( Campylos - кривая)
- Несколько основных вен или ветвей, происходящих в одной точке или близко к одной точке, и работают в повторяющихся арках, а затем сходились на вершине. Например, майантом .

- 4. Акродром
- Два или более первичных или хорошо развитых вторичных вен в сходящихся арках в направлении вершины, без базальной рекурвации, как в кампилодромозном. Может быть базальным или супрабазальным в зависимости от происхождения, и идеально или несовершенна в зависимости от того, достигают ли они до 2/3 пути к вершине. Например, Miconia (базальный тип), Endlicheria (супрабазальный тип).




- 5. Actinodromous
- Три или более первичных вен расходятся радиально от одной точки. Например, Arcangelisia (базальный тип), Givotia (супрабазальный тип).


- 6. Palinactodromous
- Первичные вены с одной или несколькими точками вторичного дихотомического ветвления за пределами первичной дивергенции, либо близко, или более отдаленно. Например, Платан .


Типы 4–6 могут также быть подклассифицированы как базальные (первичные, соединенные у основания лезвия) или супрабазально (расходящиеся над основанием лезвия), и совершенные или несовершенные, но также и яростные.
Примерно в то же время Мелвилл (1976) описал систему, применимую ко всем покрытосеменным, и с использованием латинской и английской терминологии. [ 57 ] У Мелвилла также было шесть дивизий, основанные на порядок, в котором развиваются вены.
- Arbuscular (arbuscularis)
- Повторное ветвление по регулярной дихотомии, чтобы привести к трехмерной пуш-подобной структуре, состоящей из линейного сегмента (2 подкласса)
- Яростно (Flabellatus)
- Первичные вены прямо или только слегка изогнутые, расходящиеся от основания вентилятором образом (4 подкласса)
- Ладонь (ладонь)
- Изогнутые первичные вены (3 подкласса)
- Pinnate (Pinnatus)
- Одиночная первичная вена, средняя жилка, вдоль прямой или изогнутой вторичные вены расположены с более или менее регулярными промежутками (6 подклассов)
- Коллимат (Collimatus)
- Многочисленные продольные параллельные первичные вены, возникающие из поперечной меристемы (5 подклассов)
- Конглютинат (Conglutinatus)
- Получен из плавных листовок с листами (3 подкласса)
Модифицированная форма системы Hickey была впоследствии включена в Смитсоновская классификация (1999), которая предложила семь основных типов венеции, основанные на архитектуре первичных вен, добавив в качестве дополнительного основного типа. Затем была сделана дальнейшая классификация на основе вторичных вен, с 12 типами, такими как;
- Брохидодромный
- Закрытая форма, в которой вторичные, объединенные в серии выдающихся арх, как в Хильдегардии .
- Craspedodromous
- Открытая форма с вторичными, заканчивающимися на краю, в зубчатых листьях, как у Селтиса .
- Eucamptodromous
- Промежуточная форма с перевернутыми вторичными, которые постепенно уменьшаются апикально, но внутри края и соединены промежуточными третичными венами, а не петлями между вторичными, как в роговице .
- Кладододром
- Вторые свободно разветвляются к краю, как в Rhus .
термины, которые использовались в качестве подтипов в оригинальной системе Hickey. [ 58 ]





Хильдегардия Мигеодий

Селтис Уэст

Cornus officinalis

Rhus oval
Дополнительные описания включали более высокий порядок или незначительные вены и модели ареолей ( см. Рабочая группа по архитектуре листа, рисунки 28–29). [ 58 ]

- Ускорено
- Несколько до многих равных тонких базальных вен расходятся радиально под низким углами и разветвляются апикально. Например, Paranomus .

Анализ паттернов вен часто учитывает заказы вену, тип первичной вены, вторичный тип вены (основные вены) и незначительную плотность вен. Ряд авторов приняли упрощенные версии этих схем. [ 59 ] [ 29 ] В самых простых типах первичных вен можно рассмотреть в трех или четырех группах в зависимости от рассматриваемых отделов растений;
- вершина
- ладонь
- параллель
где ладонь относится к нескольким первичным венам, которые излучаются из черешка, в отличие от разветвления из центральной основной вены в форме верната, и охватывает оба типа 4 и 5, которые сохраняются в виде подтипов; Например, ладонь-акродромис ( см. Guide Service Service Leaf). [ 60 ]

- Ладонь, ладонная, ладонная, вентиляционная, фанатная
- Несколько основных вен приблизительно одинакового размера расходятся от общей точки вблизи основания листа, где прикрепляется черешки и излучается к краю листа. Листья в ладони часто локали или делятся с долями, излучающимися от общей точки. Они могут варьироваться по количеству первичных вен (3 или более), но всегда излучаются от общей точки. [ 61 ] Например, большинство Acer (клена).

Другие системы
[ редактировать ]В качестве альтернативы Симпсон использует: [ 29 ]
- Незначительно
- Центральная средняя жилка без боковых вен ( микрофиллевые ), наблюдаемые в не снятых трахеофитах , таких как хвощики
- Дихотомический
- Вены последовательно разветвляются в вены одинаково размера из общей точки, образуя y -соединение, развернувшись. Среди умеренных древесных растений Ginkgo Biloba является единственным видом, демонстрирующим дихотомическое венецию. Также некоторые птерофиты (папоротники). [ 61 ]
- Параллель
- Первичные и вторичные вены, примерно параллельные друг другу, работая по длине листа, часто соединенные короткими перпендикулярными звенами, а не образуют сети. У некоторых видов параллельные вены соединяются у основания и вершины, такие как вечнозеленые растения и травы игла. Характеристика Monocotyledons, но исключения включают arisaema , и, как показано ниже, в разделе. [ 61 ]
- Сети (сетчатая, вернате)
- Выдающийся середина с вторичными венами разветвляется вдоль обеих сторон. Название происходит от окончательных вен, которые образуют взаимосвязанную сеть, как шаблон или сеть. (Первичное и вторичное венеция может быть названа верным, в то время как сеть, как более тонкие вены, называются сеткой или сетчатыми); Большинство немонокотных покрытосеменных, исключения, включая Calophyllum . Некоторые монокоты имеют сетчатое венецию, включая колоказию , диоскорею и Smilax . [ 61 ]

Уменьшенные микрофиллевые листья (L), возникающие в обороте из узла

Дихотомическое венеция
Однако эти упрощенные системы позволяют проводить дальнейшее разделение на несколько подтипов. Симпсон, [ 29 ] (и другие) [ 62 ] разделяет параллельные и сети (и некоторые используют только эти два термина для покрытосеменных) [ 63 ] На основании количества первичных вен (Коста) следующим образом;
- Параллель
- Penni-parallel (pinnate, pinnate parallel, unicostate parallel)
- Единственная центральная выдающаяся средняя жилка, вторичные вены из этого, возникают перпендикулярно к ней и бегут параллельно друг другу к краю или кончику, но не соединяются (анастомоза). Термин Unicostate относится к известности одной средней жилки (Costa), проходящей длину листа от основания до вершины. например, Zingiberales , такие как бананы и т. Д.
- Параллель ладони (многоцестатный параллель)
- Несколько одинаково заметных первичных вен, возникающих из одной точки у основания и бегают параллельно в сторону наконечника или поля. Термин «Многоцестат» относится к наличию более одной выдающейся основной вены. Например, «Фан» (ладони) пальмы (Arecaceae)
- Многоцестатный параллельный конверген
- Средние вины сходятся на вершине, EG Bambusa arundinacea = B. Bambos (Aracaceae), Eichornia
- Многоцестатный параллельный дивергентный
- параллельно полю Средние вины расходятся более или менее
- Сети (сетчатая)
- Pinnally (Veined, сетка, сетчатой однородной)
- Единственная выдающаяся средняя жилка, работающая от основания к вершине, вторичные вены, возникающие с обеих сторон вдоль длины первичной средней жилки, бегая к краю или вершине (наконечник), с сетью более мелких вен, образующих ретикулум (сетка или сеть). Например, Mangifera , Ficus religoisa , Psidium guajava , Hibiscus rosa-sinensis , Salix Alba
- Ладони (мультисетатная сетчатая)
- Более одного основных вен, возникающих из одной точки, проходящей от основания к вершине. Например, Liquidambar Styraciflua Это может быть дополнительно подразделено;
- Многоцестатный конвергентный
- Основные вены расходятся от происхождения у основания, затем сходятся к наконечникам. eg Zizyphus , Smilax , Cinnamomum
- Многоцестатный дивергентный
- Все основные вены расходятся к кончику. например , Gossypium , Cucurbita , Carica Papaya , Ricinus communis
- Красочно (с сетью тройки)
- Три первичные вены, как указано выше, например ( см. ) Ceanothus leucodermis , [ 64 ] C. tomentosus , [ 65 ] ЭНКЕЛИЯ ФАРИНОСА


Ладонь-параллель

Многоцестатный параллельный конверген

Многоцестатный параллельный дивергентный
Pinnaty STATED

Ладонно сетка

Многоцестатный конвергентный ладонь

Многоцестатная пальмовая дивергента
Эти сложные системы не используются в морфологических описаниях таксонов, но имеют полезность в идентификации растений, [ 29 ] хотя критикуют как чрезмерно обремененный жаргоном. [ 66 ]
Старая, даже более простая система, используемая во флоре [ 67 ] использует только две категории, открытые и закрытые.
- Открытые: вены более высокого порядка имеют свободные окончания между клетками и более характерны для немонокотидовых покрытосеменных. Они с большей вероятностью будут связаны с формами листьев, которые зубчат, лопастны или соединяются. Они могут быть подразделены как;
- Листья с вершиной (перьев), с главной центральной веной или ребра (средняя жилка), из которой возникает оставшаяся часть системы вены
- Ладонь, в которой три или более главных ребра поднимаются вместе у основания листа и расходятся вверх.
- Дихотомический, как у папоротников, где вены неоднократно раздвоились
- Закрыто: вены более высокого порядка соединены в петлях, не заканчивая свободно между клетками. Они, как правило, находятся в листьях с гладкими контурами и характерны для монокотидонов.
- Они могут быть подразделены на то, будут ли вены работать параллельно, как в травах или имеют другие узоры.
Другие описательные термины
[ редактировать ]Существует также много других описательных терминов, часто с очень специализированным использованием и ограниченными конкретными таксономическими группами. [ 68 ] Я заметный вид вен зависит от ряда функций. К ним относятся ширина вен, их известность в отношении поверхности пластинки и степень непрозрачности поверхности, которая может скрывать более тонкие вены. В связи с этим вены называются неясными , и порядок вен, которые скрыты, и, являются ли верхние, нижние или обе поверхности, дополнительно указаны. [ 69 ] [ 61 ]
Термины, которые описывают известность вену, включают в себя быка , направленные , плоские , желобы , впечатленные , выдающиеся и утопленные ( рис . 6.1 Hawthorne & Lawrence 2013). [ 66 ] [ 70 ] Вены могут демонстрировать различные типы известности в разных областях листа. Например, Pimenta Racemosa имеет каналу среднюю жилку на верхней поверхности, но это заметно на нижней поверхности. [ 66 ]
Описывая известность вен:
- Прозрачный
- Поверхность листьев поднята в серии куполов между венами на верхней поверхности, а также с заметными депрессиями. например, rytigynia pauciflora , [ 71 ] Vine Vinifera
- Канал (каналикулированная)
- Вены, затонувшие под поверхностью, что приводит к закругленному каналу. Иногда путают с «желобами», потому что каналы могут функционировать как желоба, чтобы дождь отбегал и позволить сушить, как во многих Melastomataceae . [ 72 ] например см .) Myrtaceaee ( [ 73 ] Клидемия Хирта (Melastomataceae).
- Желоб
- Вены частично выдающиеся, гребень над поверхностью листовой пластинки, но с каналами, бегущими вдоль каждой стороны, как желоба
- Впечатленный
- Вена образует поднятую линию или гребень, которая лежит ниже плоскости поверхности, которая носит ее, как будто нажимает в нее, и часто подвергаются воздействию на нижней поверхности. Ткань вблизи вен часто кажется, что пух
- Затемнять
- Вены не видны или вообще не ясны; Если не указано, то не видно невооруженным глазом. например, Berberis gagnepainii . В этом берберисе вены неясны только на нижней поверхности. [ 74 ]
- Видный
- Вена поднялась над окружающей поверхностью, поэтому ее легко ощущать при погладке пальцем. например ( см .) Pimenta racemosa , [ 73 ] Spathiphyllum cannifolium [ 75 ]
- Утопленный
- Вена затонула ниже поверхности, более заметна, чем окружающие ткани, но более затонувшие в канале, чем с впечатленными венами. например, viburnum plicatum .


Неясно (под поверхностью)

Видный

Утопленный
Описание других функций:
- Плинвисти (плиндерство)
- Более одной основной вены (нерв) у основания. Боковые вторичные вены разветвляются из точки над основанием листа. Обычно выражается в качестве суффикса , как в 3-плюсневом или триплинарном листе. В 3-плюсном (трипроизводимом) листе трех основных вен ветвь над основанием пластинки (две вторичные вены и основная вена) и впоследствии проводят параллельно, как в Ceanothus и в Celtis . Точно так же лист quintuplinerve (пять вещей) имеет четыре вторичные вены и основную вену. Узор с 3–7 венами особенно заметен в Melastomataceae . Термин также использовался в Vaccinieae . Термин использовался в качестве синонимы акродромного, ладони-акродромического или супрабазального акродрома, и считается слишком широко определенным. [ 76 ] [ 76 ]
- Скарариформ
- Вены расположены как ступени лестницы, особенно вены более высокого порядка
- Подводная лодка
- Вены бегут рядом с краем листьев
- Резкий
- 2 основные базальные нервы, кроме средней жилики
Диаграммы схемы венера
[ редактировать ]| Изображение | Срок | Описание |
|---|---|---|
 |
Дугообразно | Вторичное изгибание к вершине |
 |
Дихотомический | Вены распадаются на два |
 |
Продольный | Все вены выровнены в основном с средней частью |
 |
Параллель | Все вены параллельны и не пересекаются |
 |
Вершина | Вторичные вены, носится из средней жилки |
 |
Сетчатый | Все вены неоднократно разветвляются, чистый вун |
 |
Повернуть | Вены, исходящие от центра листа и излучаются по краям |
 |
Поперечный | Третичные вены, работающие перпендикулярно оси основной вены, соединяющие вторичные вены |
Размер
[ редактировать ]Термины мегафилла , макрофилла , мезофилла , нотафилла , микрофилла , нанофилла и лептифилла используются для описания размеров листьев (в порядке убывания), в классификации, разработанной в 1934 году Кристеном С. Раункиром и с тех пор модифицированные другими. [ 77 ] [ 78 ]
Смотрите также
[ редактировать ]- Глоссарий морфологии листьев
- Глоссарий морфологии растений § Листья
- Корона (ботаника)
- Эволюционная история листьев
- Эволюционное развитие листьев
- Индекс площади листьев
- Концентрат белка листьев
- Датчик листьев - устройство, которое измеряет уровень влаги в листьях растений
- Форма листьев
- Вернация - прорастание листьев, а также расположение листьев в зародыше
- Slek Leaf в качестве музыкального инструмента
Ссылки
[ редактировать ]- ^ Подпрыгнуть до: а беременный Исав 2006 .
- ^ Haupt 1953 .
- ^ Подпрыгнуть до: а беременный Mauseth 2009 .
- ^ "Система съемки" . Словарь ботанической терминологии . Кактус искусство, детская. и архивировано из оригинала 4 мая 2021 года . Получено 4 мая 2021 года .
- ^ Джеймс и др. 1999 .
- ^ Подпрыгнуть до: а беременный в дюймовый и Stewart & Rothwell 1993 .
- ^ Cooney-Sovetts & Sattler 1987 .
- ^ Tsukaya 2013 .
- ^ Feugier 2006 .
- ^ Purcell 2016 .
- ^ Willert et al 1992 .
- ^ Bayer 1982 .
- ^ Марлот 1913–1932 .
- ^ Подпрыгнуть до: а беременный в Симпсон 2011 , с. 356.
- ^ Крог 2010 .
- ^ Джеймс и Белл 2000 .
- ^ Heywood et al 2007 .
- ^ Симпсон 2011 , с. 356–357.
- ^ Нашел 1977 .
- ^ Подпрыгнуть до: а беременный в Ботаника иллюстрирована: введение в растения основные группы, цветущие семейства растений . Thomson Science. 1984. с. 21
- ^ Дидьер Рейнхардт и Крис Кулмеер, «Филлотаксис в высших растениях», в Майкл Т. Макманус, Брюс Вейт, ред., Меристематические ткани в росте и развитии растений , январь 2002 г., ISBN 978-1-84127-227-6 , Wiley-Blackwell.
- ^ Coxeter HS (1961). Введение в геометрию . Уайли. п. 169
- ^ Рейнхардт и Кулмеер, с. 175
- ^ Йонекура, Такааки; Ивамото, Акитоши; Fujita, Hironori; Sugiyama, Munetaka (2019-06-06). Умулис, Дэвид (ред.). «Математические модели исследования комплексной генерации основных и незначительных филлотактических паттернов у растений с преобладающим акцентом на филлотаксис Orixate» . PLOS Computational Biology . 15 (6): E1007044. BIBCODE : 2019PLSCB..15E7044Y . doi : 10.1371/journal.pcbi.1007044 . ISSN 1553-7358 . PMC 6553687 . PMID 31170142 .
- ^ Роллан-Лаган и др. 2009 .
- ^ Подпрыгнуть до: а беременный в Стены 2011 .
- ^ Подпрыгнуть до: а беременный в Dickison 2000 .
- ^ Подпрыгнуть до: а беременный Рудалл 2007 .
- ^ Подпрыгнуть до: а беременный в дюймовый и фон глин час я Simpson 2011 , Leaf Venation, стр. 465–468
- ^ Подпрыгнуть до: а беременный в дюймовый Sack & Scoffoni 2013 .
- ^ Подпрыгнуть до: а беременный в Рот-Небелск А.Л. 2001 .
- ^ Уэно и др. 2006 .
- ^ Runions et al 2005 .
- ^ Massey & Murphy 1996 , Surface-Venation-Texure
- ^ Bagchi et al 2016 .
- ^ Cote 2009 .
- ^ Клементс 1905 .
- ^ Couder et al 2002 .
- ^ Corson et al 2009 .
- ^ Laguna et в 2008 году .
- ^ Арбер 1950 .
- ^ Rutishauser & Sattler 1997 .
- ^ Lacroix et al 2003 .
- ^ Eckardt & Baum 2010 .
- ^ Пуар, Ричард; Wiese-Klinkenberg, Аника; Родитель, Борис; Милевчик, Майкл; Шурр, Ульрих; Тардие, Франсуа; Уолтер, Ахим (2010). «Diel по времени роста листьев у видов монокота и дикота: эндогенные ритмы и температурные эффекты» . Журнал экспериментальной ботаники . 61 (6): 1751–1759. doi : 10.1093/jxb/erq049 . ISSN 1460-2431 . PMC 2852670 . PMID 20299442 .
- ^ Милевчик, Майкл; Фридли, Майкл; Кирхжесер, Норберт; Уолтер, Ахим (2013-07-25). «Рост листьев диэля сои: новый метод для анализа двумерной экспансии листьев в высоком временном разрешении, основанном на подходе к отслеживанию маркеров (Martrack Leaf)» . Методы растения . 9 (1): 30. doi : 10.1186/1746-4811-9-30 . HDL : 20.500.11850/76534 . ISSN 1746-4811 . PMC 3750653 . PMID 23883317 .
- ^ Фридли, Майкл; Уолтер, Ахим (2015). «Схемы роста DIEL молодых листовок сои (G Lycine Max) синхронны в разных положениях на растении» . Растение, клетки и окружающая среда . 38 (3): 514–524. doi : 10.1111/pce.12407 . ISSN 0140-7791 . PMID 25041284 .
- ^ Подпрыгнуть до: а беременный в дюймовый и Читать и Stokes 2006 .
- ^ Doring et al 2009 .
- ^ Fild et al 2001 .
- ^ «Глоссарий Кью - определение искисления» . 3 декабря 2013 года. Архивировано с оригинала 3 декабря 2013 года . Получено 12 мая 2017 года .
{{cite web}}: CS1 Maint: Bot: исходный статус URL неизвестен ( ссылка ) - ^ «Глоссарий Кью - определение синанта» . 3 декабря 2013 года. Архивировано с оригинала 3 декабря 2013 года . Получено 12 мая 2017 года .
{{cite web}}: CS1 Maint: Bot: исходный статус URL неизвестен ( ссылка ) - ^ Ettingshausen 1861 .
- ^ Hickey 1973 .
- ^ Hickey & Wolfe 1975 .
- ^ Hickey 1979 .
- ^ Мелвилл 1976 .
- ^ Подпрыгнуть до: а беременный Leaf Architecture Working Group 1999 .
- ^ Джадд и др. 2007 .
- ^ Florissant Leaf Key 2016 .
- ^ Подпрыгнуть до: а беременный в дюймовый и Kling et al 2005 , Venation Leaf
- ^ Берг 2007 .
- ^ Морфология ангиосперма 2017 , венеция
- ^ Симпсон 2017 , Ланди Линксус
- ^ Симпсон 2017 , Ceanothus tomentosus
- ^ Подпрыгнуть до: а беременный в Hawthorne & Lawrence 2013 , Leaf Venation с. 135–136
- ^ Cullen et al 2011 .
- ^ Neotropikey 2017 .
- ^ Оксфорд Гербария Глоссарий 2017 .
- ^ Оксфорд Гербария Глоссарий 2017 , Вена известность
- Verd Courses & Bridson 1991 .
- ^ Hemsley & Poole 2004 , морфология листьев и сушка с. 254
- ^ Подпрыгнуть до: а беременный Hughes 2017 , Pimenta racemosa
- ^ Cullen E AL 2011 , Berberis Ghersupi Vol. II P. 398
- ^ Kwantlen 2015 , Spathiphyllum cannifolium
- ^ Подпрыгнуть до: а беременный Pedraza-Peñalosa 2013 .
- ^ Уиттен и др. 1997 .
- ^ Уэбб, Лен (1 октября 1959 г.). «Физиономическая классификация австралийских дождевых лесов». Журнал экологии . 47 (3). Британское экологическое общество: журнал экологии Vol. 47, № 3, с. 551–570: 555. Bibcode : 1959jecol..47..551w . doi : 10.2307/2257290 . JSTOR 2257290 .
Библиография
[ редактировать ]Книги и главы
[ редактировать ]- Арбер, Агнес (1950). Естественная философия растения формы . Кубок Архив . GGKEY: HCBB8RZREL4.
- Bayer, MB (1982). Новое руководство Haworthia . Кирстенбош: Национальные ботанические сады Южной Африки . ISBN 978-0-620-05632-8 Полем Архивировано из оригинала 2023-09-06 . Получено 2018-07-25 .
- Берг, Линда (23 марта 2007 г.). Вводная ботаника: растения, люди и окружающая среда, СМИ . Cengage Learning. ISBN 978-1-111-79426-2 .
- Каллен, Джеймс; Колени, Сабина Г.; Куби, Х. Сюзанна Куби, ред. (2011) [1984–2000]. Европейская садовая флора, цветущие растения: руководство по идентификации растений, выращенных в Европе, как на улице, так и под стеклом. 5 томов (2 -е изд.). Кембридж: издательство Кембриджского университета . Архивировано с оригинала 2016-12-28 . Получено 2017-03-08 .
- Каттер, например (1969). Анатомия растений, эксперимент и интерпретация, части 2 органы . Лондон: Эдвард Арнольд. п. 117. ISBN 978-0-7131-2302-9 .
- Dickison, William C. (2000). Интегративная анатомия растений . Академическая пресса . ISBN 978-0-08-050891-7 .
- Исав, Кэтрин (2006) [1953]. Evert, Ray F (ed.). Анатомия растения Исава: меристемы, клетки и ткани тела растения: их структура, функция и развитие (3 -е изд.). Нью -Йорк: John Wiley & Sons Inc. ISBN 978-0-470-04737-8 Полем Архивировано из оригинала 2023-09-06 . Получено 2017-09-02 .
- Etingshausen, C. (1861). Листовой скелет DiCotyledons с особым пейзажным видом на осмотр и определение ископаемого растения остается . Вена: классификация архитектуры DiCotyledonous.
- Haupt, Arthur Wing (1953). Морфология растений . МакГроу-Хилл .
- Хоторн, Уильям; Лоуренс, Анна (2013). Идентификация завода: создание удобных полевых руководств для управления биоразнообразием . Routledge. ISBN 978-1-136-55972-3 .
- Хемсли, Алан Р.; Пул, Имоген, ред. (2004). Эволюция физиологии растений . Академическая пресса . ISBN 978-0-08-047272-0 .
- Хейвуд, VH ; Brummitt, RK; Калхэм А .; Себерг, О. (2007). Цветлые семейства растений мира . Нью -Йорк: книги «Светлячок». п. 287. ISBN 978-1-55407-206-4 .
- Hickey, LJ. Пересмотренная классификация архитектуры двудольных листьев . с. I 5–39. , в Metcalfe & Chalk (1979)
- Джадд, Уолтер С .; Кэмпбелл, Кристофер С.; Келлогг, Элизабет А.; Стивенс, Питер Ф . ; Donoghue, Michael J. (2007) [1 -е изд. 1999, 2 -й 2002]. Систематика растений: филогенетический подход (3 -е изд.). Sinauer Associates. ISBN 978-0-87893-407-2 Полем Архивировано из оригинала 2023-09-06 . Получено 2017-09-02 .
- Крог, Дэвид (2010), Биология: руководство по миру природы (5-е изд.), Бенджамин-Каммингс издательская компания, с. 463, ISBN 978-0-321-61655-5 , архивировано с оригинала в 2023-01-24 , извлечен 2016-05-24
- Рабочая группа по архитектуре листьев (1999). Руководство по архитектуре листьев - морфологическое описание и категоризация двудольных и сетки монокотидовых покрытосеменных (PDF) . Смитсоновский институт . ISBN 978-0-9677554-0-3 Полем Архивировано (PDF) из оригинала 2016-10-20 . Получено 2017-02-15 .
- Марлот, Рудольф (1913–1932). Флора Южной Африки: с синопическими таблицами родов высших растений. 6 томов Кейптаун: Barter Bros. & Co. Архивировал оригинал 2023-09-06 . Получено 2020-08-27 .
- Маусет, Джеймс Д. (2009). Ботаника: введение в биологию растений (4 -е изд.). Садбери, штат Массачусетс: издатели Джонса и Бартлетта. ISBN 978-0-7637-5345-0 .
- Metcalfe, CR; Мел, L, ред. (1979) [1957]. Анатомия дикотидонов: листья, стебель и древесина по отношению к таксономии, с заметками об экономическом использовании. 2 тома (2 -е изд.). Оксфорд: Clarendon Press. ISBN 978-0-19-854383-1 .
- Пранс, Гиллеан Толми (1985). Листья: образование, характеристики и использование сотен листьев, найденных во всех частях света . Фотографии Кьелла Б. Сэндс. Лондон: Темза и Хадсон. ISBN 978-0-500-54104-3 .
- Рудалл, Паула Дж. (2007). Анатомия цветущих растений: введение в структуру и развитие (3 -е изд.). Кембридж: издательство Кембриджского университета . ISBN 978-0-521-69245-8 Полем Архивировано из оригинала 2023-09-06 . Получено 2020-08-27 .
- Симпсон, Майкл Дж. (2011). Систематика растений . Академическая пресса. ISBN 978-0-08-051404-8 Полем Архивировано из оригинала 2023-01-17 . Получено 2016-05-24 .
- Стюарт, Уилсон Н; Rothwell, Gar W. (1993) [1983]. Палеоботания и эволюция растений (2 -е изд.). Издательство Кембриджского университета . ISBN 978-0-521-38294-6 .
- Вердкорт, Бернард; Бридсон, Дайан М. (1991). Флора Тропической Восточной Африки - Рубиаааэ Том 3 . CRC Press. ISBN 978-90-6191-357-3 .
- Уиттен, Тони; Есть сирийцы Роиты, эммит; Affiff, Suraya A. (1997). Экология Java - Бали . Oxford Pressy Universy. п. 505. ISBN 978-962-593-072-5 Полем Архивировано из оригинала 2023-09-06 . Получено 2020-08-27 .
- Уиллерт, Дитер Дж. Фон; Эллер, Б.М.; Теплее, MJA; Бринкманн, E; Ihlenfeldt, HD (1992). Жизненные стратегии суккулентных в пустынях: с особым ссылкой на пустыню Намиб . Кубок Архив . ISBN 978-0-521-24468-8 .
Статьи и тезисы
[ редактировать ]- Багчи, Дебьяни; Дасгупта, Авик; Гондалия, Амит Д.; Раджпут, Кишор С. (2016). «Понимание из мира растений: подход фрактального анализа для настройки механической жесткости матрицы лесов в тонких пленках». Расширенные материалы исследования . 1141 : 57–64. doi : 10.4028/www.scientific.net/amr.1141.57 . S2CID 138338270 .
- Клементс, Эдит Шварц (декабрь 1905 г.). «Соотношение структуры листьев к физическим факторам» . Сделки американского микроскопического общества . 26 : 19–98. doi : 10.2307/3220956 . JSTOR 3220956 . Архивировано из оригинала 2023-08-04 . Получено 2023-09-06 .
- Cooney-Sovetts, C.; Саттлер Р. (1987). «Развитие Phylloclade в Asparagaceae: пример гомеоза». Ботанический журнал Линневого общества . 94 (3): 327–371. doi : 10.1111/j.1095-8339.1986.tb01053.x .
- Корсон, Фрэнсис; Адда-Бедиа, Мохтар; Boudaoud, Arezki (2009). «В сети венения листьев Силико: рост и реорганизация, обусловленные механическими силами» (PDF) . Журнал теоретической биологии . 259 (3): 440–448. BIBCODE : 2009JTHBI.259..440C . doi : 10.1016/j.jtbi.2009.05.002 . PMID 19446571 . S2CID 25560670 . Архивировано из оригинала (PDF) 2017-12-09.
- Cote, GG (2009). «Разнообразие и распределение идиобластов, производящих кристаллы оксалата кальция в Диффенбахии Сегине (Araceae)» . Американский журнал ботаники . 96 (7): 1245–1254. doi : 10.3732/ajb.0800276 . PMID 21628273 .
- Couder, y.; Pauchard, L.; Allain, C.; Adda-Bedia, M.; Doudy, S. (1 июля 2002 г.). «Венеция листьев, образованного в тенсиальном поле» (PDF) . Европейский физический журнал б . 28 (2): 135–138. Bibcode : 2002epjb ... 28..135c . doi : 10.1140/epjb/e2002-00211-1 . S2CID 51687210 . Архивировано из оригинала (PDF) 9 декабря 2017 года.
- Доринг, Т. Ф; Археетти, М.; Харди, Дж. (7 января 2009 г.). «Осенние листья видели глазами травоядных» . Труды Королевского общества B: Биологические науки . 276 (1654): 121–127. doi : 10.1098/rspb.2008.0858 . PMC 2614250 . PMID 18782744 .
- Экардт, На; Баум, Д. (20 июля 2010 г.). «Загадка Podostemad: эволюция необычной морфологии в Podostemaceae» . Растительная ячейка онлайн . 22 (7): 2104. DOI : 10.1105/TPC.110.220711 . PMC 2929115 . PMID 20647343 .
- Феугье, Франсуа (14 декабря 2006 г.). Модели формирования сосудистого рисунка в листьях (диссертация доктора философии) . Университет Парижа VI . Архивировано (PDF) из оригинала 7 марта 2017 года . Получено 6 марта 2017 года .
- Фейлд, Т.С.; Ли, DW; Холбрук, Нью -Йорк (1 октября 2001 г.). «Почему листья становятся красными осенью. Роль антоцианинов в сжигающих листьях рождингового кизила» . Физиология растений . 127 (2): 566–574. doi : 10.1104/pp.010063 . PMC 125091 . PMID 11598230 .
- Hallé, F. (1977). «Самый длинный лист в ладонях». Принципы . 21 : 18.
- Хикки, Лео Дж. (1 января 1973 г.). «Классификация архитектуры двудольных листьев» (PDF) . Американский журнал ботаники . 60 (1): 17–33. doi : 10.2307/2441319 . JSTOR 2441319 . Архивировано (PDF) из оригинала 11 августа 2017 года . Получено 14 февраля 2017 года .
- Hickey, Leo J.; Вулф, Джек А. (1975). «Основы филогения покрытосемперма: вегетативная морфология» . Анналы ботанического сада Миссури . 62 (3): 538–589. doi : 10.2307/2395267 . JSTOR 2395267 . Архивировано из оригинала 2022-10-31 . Получено 2023-09-06 .
- Ингерсолл, Эрнест (1920). . Энциклопедия Американа . Тол. XVII.
- Джеймс, СА; Bell, DT (2000). «Влияние доступности света на структуру листьев и рост двух эвкалиптовых Globulus ssp. Globulus provenances» (PDF) . Физиология деревьев . 20 (15): 1007–1018. doi : 10.1093/treephys/20.15.1007 . PMID 11305455 . Архивировано (PDF) из оригинала на 2022-04-01 . Получено 2019-05-10 .
- Lacroix, C.; Jeune, B.; Purcell-Macdonald, S. (2003). «Сравнение стрельбы и составных листьев в eudicots: динамическая морфология как альтернативный подход» . Ботанический журнал Линневого общества . 143 (3): 219–230. doi : 10.1046/j.1095-8339.2003.00222.x . Архивировано из оригинала 2020-06-22 . Получено 2019-09-08 .
- Лагуна, Мария Ф.; Бон, Штеффен; Jagla, Eduardo A.; Борн, Филип Э. (2008). «Роль упругих стрессов в морфогенезе венения листьев» . PLOS Computational Biology . 4 (4): E1000055. Arxiv : 0705.0902 . BIBCODE : 2008PLSCB ... 4E0055L . doi : 10.1371/journal.pcbi.1000055 . PMC 2275310 . PMID 18404203 .
- Мелвилл Р. (ноябрь 1976). «Терминология архитектуры листьев». Таксон 25 (5/6): 549–561. doi : 10.2307/1220108 . JSTOR 1220108 .
- Pedraza-Peñalosa, Paola; Салинас, Нельсон Р.; Уилер, Уорд С. (26 апреля 2013 г.). «Образцы венения неотропической черники (Vaccinieae: Ericaceae) и их сложно» (PDF ) Фитотакса 96 (1): 1. DOI : 10.11646/phytotaxa.96.1.1 . Архивировано (PDF) 18 февраля из оригинала Получено 17 февраля
- Read, J.; Стоукс А. (1 октября 2006 г.). «Биомеханика растений в экологическом контексте» . Американский журнал ботаники . 93 (10): 1546–1565. doi : 10.3732/ajb.93.10.1546 . PMID 21642101 .
- Роллан-Лаган, Энн-Гаэль; Амин, Мира; Пакулска, Малгозия (январь 2009 г.). «Количественная оценка паттернов внесения листьев: двумерные карты». Заводский журнал . 57 (1): 195–205. doi : 10.1111/j.1365-313x.2008.03678.x . PMID 18785998 .
- Рот-Небельс, а; UHL, диетор; Mosbrugger, Volker; Керп, Ганс (май 2001 г.). «Эволюция и функция архитектуры венения листовых: обзор» . Анналы ботаники . 87 (5): 553–566. doi : 10.1006/anbo.2001.1391 .
- Рунион, Адам; Фурер, Мартин; Лейн, Брендан; Федерл, Павол; Роллан-Лаган, Энн-Гаэль; Prusinkiewicz, Przemyslaw (1 января 2005 г.). «Моделирование и визуализация паттернов венения листьев». ACM Siggraph 2005 Документы . Тол. 24. С. 702–711. Citeseerx 10.1.1.102.1926 . doi : 10.1145/1186822.1073251 . ISBN 9781450378253 Полем S2CID 2629700 .
- Rutishauser, R.; Саттлер Р. (1997). «Экспрессия процессов побега в развитии листьев полемемолия Coeruleum ». Ботанические ежегодники для систематики . 119 : 563–582.
- Мешок, Лоурен; Скоффони, Кристина (июнь 2013 г.). «Венеция листьев: структура, функция, развитие, эволюция, экология и применение в прошлом, настоящем и будущем» . Новый фитолог . 198 (4): 983–1000. doi : 10.1111/nph.12253 . PMID 23600478 .
- Шелли, AJ; Смит, WK; Vogelmann, TC (1998). «Онтогенетические различия в структуре мезофилла и распределении хлорофилла в Eucalyptus Globulus ssp. Globulus (Myrtaceae)» . Американский журнал ботаники . 86 (2): 198–207. doi : 10.2307/2656937 . JSTOR 2656937 . PMID 21680359 .
- Цукайя, Хироказу (январь 2013 г.). "Развитие листьев" . Книга Arabidopsis . 11 : E0163. doi : 10.1199/tab.0163 . PMC 3711357 . PMID 23864837 .
- Уэно, Осаму; Кавано, Юкико; Вакаяма, Масатака; Такеда, Томосиро (1 апреля 2006 г.). «Листовые сосудистые системы в травах C 3 и C 4 : двумерный анализ» . Анналы ботаники . 97 (4): 611–621. doi : 10.1093/aob/mcl010 . PMC 2803656 . PMID 16464879 .
- Стены, RL (25 января 2011 г.). «Определенные паттерны листьев покрыто листьев связаны с функциями листьев в наборе данных в глобальном масштабе». Американский журнал ботаники . 98 (2): 244–253. doi : 10.3732/ajb.1000154 . PMID 21613113 .
Веб -сайты
[ редактировать ]- Бакш, Александр; Блюндер, Бенджамин; Прайс, Чарльз; Крыло, Скотт; Вейц, Джошуа; Дас, Абхирам (2017). «Очищенная база данных изображений листа» . Школа биологии, Технологический институт Джорджии . Архивировано с оригинала 25 сентября 2014 года . Получено 12 марта 2017 года .
- Geneve, Роберт. «Лист» (PDF) . PLS 220: Введение в идентификацию растений . Университет Кентукки: факультет садоводства. Архивировано из оригинала (PDF) 2016-03-15.
- Клинг, Гэри Дж.; Hayden, Laura L.; Поттс, Джошуа Дж. (2005). «Ботаническая терминология» . Университет Иллинойса , Урбана-Шампейн. Архивировано из оригинала 8 марта 2017 года . Получено 7 марта 2017 года .
- де Кок, Роджиер; Biffin, ed (ноябрь 2007 г.). «Ключ гороха: интерактивный ключ для австралийских гороховых бобовых» . Австралийская исследовательская группа для бобовых цветов. Архивировано из оригинала 26 февраля 2017 года . Получено 9 марта 2017 года .
- Кранц, Лора. «Образцы вены листьев» (рисунки) . Архивировано с оригинала 2017-03-06 . Получено 2017-03-05 .
- Massey, Jimmy R.; Мерфи, Джеймс С. (1996). «Систематика сосудистой растения» . NC Ботнет . Университет Северной Каролины в Чапел -Хилл. Архивировано с оригинала 17 января 2016 года . Получено 19 января 2016 года .
- "Листья" . Архивировано с оригинала 2016-07-25 . Получено 2016-01-19 . , в Massey & Murphy (1996)
- Перселл, Адам (16 января 2016 г.). "Листья" . Основная биология . Адам Перселл. Архивировано из оригинала 19 апреля 2015 года . Получено 17 февраля 2017 года .
- Симпсон, Майкл Дж. «Заводы округа Сан -Диего, Калифорния» . Колледж науки, Государственный университет Сан -Диего . Архивировано из оригинала 3 марта 2017 года . Получено 2 марта 2017 года .
- «Флориссанты ископаемых слоев листьев» (PDF) . Флориссант ископаемые кровати Национальный памятник . Служба национальных парков , Министерство внутренних дел США . Архивировано (PDF) из оригинала 16 февраля 2017 года . Получено 16 февраля 2017 года .
- «База данных растений» . Школа садоводства, Квантленский политехнический университет . 2015. Архивировано с оригинала 21 сентября 2017 года . Получено 9 марта 2017 года .
- «Морфология покрытоперма» . Tutorvista. 2017. Архивировано из оригинала 2020-06-21 . Получено 2017-03-09 .
- Глоссарии
- Хьюз, Колин. «Виртуальный поле гербарий» . Оксфордский университет Гербария . Архивировано с оригинала 5 марта 2017 года . Получено 4 марта 2017 года .
- «Характеристики растений» . Архивировано из оригинала (Глоссарий) 5 марта 2017 года . Получено 4 марта 2017 года . , в Хьюзе (2017)
- «Глоссарий ботанических терминов» . Neotropikey . Королевские ботанические сады, Кью. Архивировано из оригинала 21 января 2017 года . Получено 18 февраля 2017 года .
- «Иллюстрированный глоссарий листовых форм» (PDF) . Центр водных и инвазивных растений, Институт продовольственных и сельскохозяйственных наук , Университет Флориды. 2009. Архивировал (PDF) из оригинала 10 января 2020 года . Получено 8 января 2020 года .
- «Листовые шаги» . Донсгарден . Архивировано с оригинала 4 февраля 2016 года . Получено 9 января 2020 года .
Внешние ссылки
[ редактировать ]- Рендл, Альфред Бартон (1911). . Encyclopædia Britannica . Тол. 15 (11 -е изд.). п. 322-329.
- Ингерсолл, Эрнест (1920). . Энциклопедия Американа . Тол. XVII.