Папилломавирусиды
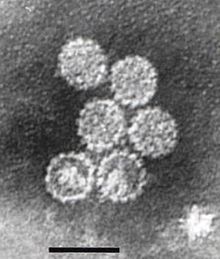
Papillomaviridae — это семейство без оболочки ДНК-вирусов , члены которого известны как папилломавирусы. [ 1 ] Несколько сотен видов папилломавирусов, традиционно называемых «типами», [ 2 ] выявлено заражение всех тщательно обследованных млекопитающих, [ 2 ] но и другие позвоночные, такие как птицы, змеи, черепахи и рыбы. [ 3 ] [ 4 ] [ 5 ] Заражение большинством типов папилломавирусов, в зависимости от типа, либо протекает бессимптомно (например, большинство бета-PV), либо вызывает небольшие доброкачественные опухоли, известные как папилломы или бородавки (например, вирус папилломы человека 1, ВПЧ6 или ВПЧ11). Однако папилломы, вызванные некоторыми типами, такими как вирусы папилломы человека 16 и 18, несут риск перерождения в рак . [ 6 ]
Папилломавирусы обычно считаются высокотропными хозяину и тканям к и редко передаются между видами. [ 7 ] Папилломавирусы реплицируются исключительно в базальном слое поверхностных тканей тела . Все известные типы папилломавирусов поражают определенную поверхность тела. [ 2 ] обычно кожа или эпителий слизистых оболочек гениталий, ануса, рта или дыхательных путей. [ 8 ] Например, вирус папилломы человека (ВПЧ) 1-го типа имеет тенденцию поражать подошвы ног, а ВПЧ 2-го типа — ладони, где они могут вызывать бородавки . Дополнительно имеются описания наличия ДНК папилломавируса в крови и в мононуклеарных клетках периферической крови .
Папилломавирусы были впервые идентифицированы в начале 20 века, когда было показано, что кожные бородавки или папилломы могут передаваться между людьми фильтрующимся инфекционным агентом. В 1935 году Фрэнсис Пейтон Роус , который ранее продемонстрировал существование вызывающего рак вируса саркомы у кур, продолжил показывать, что папилломавирус может вызывать рак кожи у инфицированных кроликов. Это была первая демонстрация того, что вирус может вызывать рак у млекопитающих.
Таксономия папилломавирусов
[ редактировать ]
Известно более 100 видов вируса папилломы. [ 9 ] хотя ICTV официально признает меньшее количество, разделенное на 53 рода, по состоянию на 2019 год. [ 10 ] [ 11 ] [ 12 ] Все папилломавирусы (ПВ) имеют сходную геномную организацию, и любая пара ПВ содержит не менее пяти гомологичных генов , хотя нуклеотидная последовательность может отличаться более чем на 50%. Филогенетические алгоритмы , позволяющие сравнивать гомологии, привели к созданию филогенетических деревьев , имеющих схожую топологию, независимо от анализируемого гена. [ 13 ]
Филогенетические исследования убедительно свидетельствуют о том, что PV обычно развиваются вместе со своими видами-млекопитающими и птицами-хозяевами, но адаптивная радиация , случайные зоонозные события и рекомбинации также могут влиять на их диверсификацию. [ 13 ] Их базовая геномная организация, по-видимому, сохранялась в течение периода, превышающего 100 миллионов лет, и эти сравнения последовательностей заложили основу для таксономии PV, которая теперь официально признана Международным комитетом по таксономии вирусов . Все PV образуют семейство Papillomaviridae , которое отличается от Polyomaviridae, что исключает термин Papovaviridae . Основными ветвями филогенетического дерева ПВ считаются роды , которые обозначаются греческими буквами. Второстепенные ветви считаются видами и объединяют типы PV, которые геномно различны, но не имеют известных биологических различий. Эта новая таксономическая система не влияет на традиционную идентификацию и характеристику «типов» PV и их независимых изолятов с незначительными геномными различиями, называемых «подтипами» и «вариантами», все из которых являются таксонами ниже уровня «вида». [ 14 ] Кроме того, были предложены филогенетические группировки на более высоком таксономическом уровне. [ 15 ]
Эта классификация может нуждаться в пересмотре в свете существования рекомбинантов вируса папилломы и полиомы. [ 16 ] Описаны и другие виды. Папилломавирус 1 Sparus aurata выделен из рыбы. [ 17 ]
Вирусы папилломы человека
[ редактировать ]Полностью секвенировано более 170 типов вируса папилломы человека. [ 18 ] Их разделили на 5 родов: Alphapapillomavirus, Betapapillomavirus, Gammapapillomavirus, Mupapillomavirus и Nupapillomavirus. Было идентифицировано по меньшей мере 200 дополнительных вирусов, ожидающих секвенирования и классификации. [ нужна ссылка ]
Папилломавирусы животных
[ редактировать ]
Отдельные типы вируса папилломы, как правило, хорошо адаптированы к репликации у одного вида животных. В одном исследовании исследователи взяли мазок с кожи лба различных животных в зоопарке и использовали ПЦР для амплификации любой ДНК папилломавируса, которая могла присутствовать. [ 19 ] Хотя в ходе исследования было идентифицировано большое разнообразие последовательностей вируса папилломы, авторы обнаружили мало доказательств межвидовой передачи. У одного смотрителя зоопарка был обнаружен временный положительный результат на последовательность папилломавируса, специфичную для шимпанзе. Однако авторы отмечают, что специфичная для шимпанзе последовательность папилломавируса могла быть результатом поверхностного загрязнения кожи смотрителя зоопарка, а не продуктивной инфекции. [ нужна ссылка ]
Вирус папилломы кролика (CRPV) может вызывать выступающие бородавки у своего родного хозяина, североамериканского кролика рода Sylvilagus . Эти роговидные бородавки могут стать исходной основой городских легенд об американском рогатом кролике шакалопе и европейском вольпертингере . [ 20 ] Европейские домашние кролики (род Oryctolagus ) могут быть транзиторно инфицированы CRPV в лабораторных условиях. Однако, поскольку европейские домашние кролики не производят вирус инфекционного потомства, они считаются случайными или «тупиковыми» хозяевами CRPV. [ 21 ]
Межвидовая передача также была зарегистрирована для вируса папилломы крупного рогатого скота (BPV) типа 1. [ 22 ] У естественного хозяина (крупного рогатого скота) BPV-1 вызывает появление крупных волокнистых кожных бородавок. Заражение BPV-1 лошадей, которые являются случайным хозяином вируса, может привести к развитию доброкачественных опухолей, известных как саркоиды . Сельскохозяйственное значение BPV-1 стимулировало успешные усилия по разработке вакцины против этого вируса. [ нужна ссылка ]
В нескольких сообщениях папилломавирусы были выявлены у более мелких грызунов, таких как сирийские хомяки , африканская многососковая крыса и евразийская промысловая мышь . [ 23 ] Однако неизвестны вирусы папилломы, способные заражать лабораторных мышей . Отсутствие поддающейся лечению мышиной модели папилломавирусной инфекции было основным ограничением для лабораторных исследований папилломавирусов. [ нужна ссылка ]
Известно, что четыре папилломавируса заражают птиц: папилломавирус Fringilla coelebs 1, папилломавирус 1 Francolinus leucoscepus, папилломавирус 1 Psittacus erithacus и папилломавирус 1 Pygoscelis adeliae. [ 24 ] У всех этих видов есть ген (E9) неизвестной функции, что позволяет предположить общее происхождение.
Эволюция
[ редактировать ]Считается, что эволюция папилломавирусов происходит медленно по сравнению со многими другими типами вирусов, но экспериментальных измерений в настоящее время нет. Вероятно, это связано с тем, что геном папилломавируса состоит из генетически стабильной двухцепочечной ДНК, которая с высокой точностью реплицируется механизмом репликации ДНК клетки-хозяина. [ нужна ссылка ]
Считается, что вирусы папилломы обычно эволюционируют совместно с определенным видом животного-хозяина в течение многих лет, хотя существуют убедительные доказательства против гипотезы коэволюции. [ 13 ] [ 25 ] В качестве особенно быстрого примера можно привести ВПЧ-16, который немного изменился по мере увеличения численности населения по всему миру и теперь различается в разных географических регионах, что, вероятно, отражает историю миграции людей. [ 26 ] [ 27 ] Кутанеотропные типы ВПЧ иногда передаются между членами семьи в течение всей жизни, но при передаче вируса следует учитывать и других доноров. [ 28 ]
Другие типы ВПЧ, такие как ВПЧ-13, относительно мало различаются в разных популяциях человека. Фактически, последовательность ВПЧ-13 очень напоминает вирус папилломы бонобо (также известный как карликовые шимпанзе). [ 29 ] Неясно, связано ли это сходство с недавней передачей вируса между видами или с тем, что ВПЧ-13 просто очень мало изменился за шесть или около того миллионов лет, прошедших с тех пор, как люди и бонобо разошлись. [ 27 ]
По оценкам, самый последний общий предок этой группы вирусов существовал 424 миллиона лет назад . [ 30 ]
Существует пять основных родов, заражающих человека (Альфа, Бета, Гамма, Му и Ню). Последний общий предок этих родов появился 49,7 миллиона лет назад – 58,5 миллиона лет назад . [ 31 ] По оценкам, самый последний предок рода гамма появился между 45,3 назад и 67,5 миллионами лет . [ нужна ссылка ]
Структура
[ редактировать ]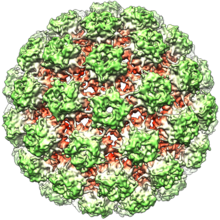
Папилломавирусы не имеют оболочки, то есть внешняя оболочка или капсид вируса не покрыта липидной мембраной . Один вирусный белок, известный как L1, необходим и достаточен для образования капсида размером 55–60 нанометров, состоящего из 72 звездчатых капсомеров (см. Рисунок). Как и у большинства вирусов без оболочки, капсид имеет геометрическую правильную форму и обладает икосаэдрической симметрией . Самособирающиеся вирусоподобные частицы, состоящие из L1, являются основой успешной группы профилактических вакцин против ВПЧ , предназначенных для выработки вируснейтрализующих антител , которые защищают от первичной инфекции ВПЧ. Таким образом, папилломавирусы устойчивы во внешней среде . [ нужна ссылка ]
Геном папилломавируса представляет собой двухцепочечную кольцевую молекулу ДНК около 8000 пар оснований длиной . Он упакован внутри оболочки L1 вместе с клеточными белками- гистонами , которые служат для обертывания и конденсации ДНК. [ нужна ссылка ]
Капсид папилломавируса также содержит вирусный белок, известный как L2, которого меньше. Хотя неясно, как L2 устроен внутри вириона, известно, что он выполняет несколько важных функций, включая облегчение упаковки вирусного генома в формирующиеся вирионы, а также инфекционное проникновение вируса в новые клетки-хозяева. L2 представляет интерес как возможная мишень для вакцин более широкого защитного действия против ВПЧ .
Вирусный капсид состоит из 72 капсомеров, из которых 12 пятикоординированных и 60 шестикоординированных, расположенных на икосаэдрической поверхностной решетке T = 7d. [ 32 ]
Специфичность ткани
[ редактировать ]Папилломавирусы реплицируются исключительно в кератиноцитах . Кератиноциты образуют внешние слои кожи, а также некоторые поверхности слизистых оболочек , например, внутреннюю часть щеки или стенки влагалища. Эти поверхностные ткани, известные как многослойный плоский эпителий , состоят из сложенных слоев уплощенных клеток. Слои клеток формируются в результате процесса, известного как клеточная дифференциация , при котором кератиноциты постепенно становятся специализированными, в конечном итоге образуя твердую сшитую поверхность, которая предотвращает потерю влаги и действует как барьер против патогенов. Менее дифференцированные стволовые клетки кератиноцитов, пополняющиеся в поверхностном слое, считаются начальной мишенью продуктивных папилломавирусных инфекций. Последующие этапы жизненного цикла вируса строго зависят от процесса дифференцировки кератиноцитов. В результате папилломавирусы могут реплицироваться только в поверхностных тканях тела. [ нужна ссылка ]
Жизненный цикл
[ редактировать ]Инфекционный вход
[ редактировать ]Папилломавирусы получают доступ к стволовым клеткам кератиноцитов через небольшие раны, известные как микротравмы, на поверхности кожи или слизистых оболочек. Взаимодействие между L1 и сульфатированными сахарами на поверхности клетки способствует начальному прикреплению вируса. [ 33 ] [ 34 ] Затем вирус может проникнуть внутрь с поверхности клетки посредством взаимодействия со специфическим рецептором, вероятно, через интегрин альфа-6 бета-4. [ 35 ] [ 36 ] и транспортируется в закрытые мембраной везикулы, называемые эндосомами . [ 37 ] [ 38 ] Капсидный белок L2 разрушает мембрану эндосомы посредством катионного пептида, проникающего в клетку , позволяя вирусному геному ускользнуть и переместиться вместе с L2 в ядро клетки. [ 39 ] [ 40 ] [ 41 ]
Персистенция и латентность вируса
[ редактировать ]После успешного заражения кератиноцита вирус экспрессирует белки E1 и E2, которые предназначены для репликации и поддержания вирусной ДНК в виде кольцевой эписомы . Вирусные онкогены E6 и E7 способствуют росту клеток, инактивируя белки-супрессоры опухолей p53 и pRb . Стволовые клетки кератиноцитов в базальном слое эпителия могут сохранять геномы папилломавируса в течение десятилетий. [ 8 ]
Производство потомства вируса
[ редактировать ]В настоящее время считается, что репликация вирусной ДНК, вероятно, происходит в G 2 фазе клеточного цикла и зависит от рекомбинационно-зависимой репликации, поддерживаемой механизмами ответа на повреждение ДНК (активируемыми белком E7) для производства потомства вирусных геномов. [ 42 ] Геномы папилломавирусов иногда интегрируются в геном хозяина, что особенно заметно при онкогенных ВПЧ, но это не является нормальной частью жизненного цикла вируса и тупиком, исключающим возможность производства вирусного потомства. [ 42 ]
Экспрессия поздних генов вируса, L1 и L2, ограничивается исключительно дифференцировкой кератиноцитов в самых внешних слоях кожи или поверхности слизистой оболочки. Повышенная экспрессия L1 и L2 обычно коррелирует с резким увеличением количества копий вирусного генома. Поскольку внешние слои многослойного плоского эпителия подлежат относительно ограниченному наблюдению со стороны клеток иммунной системы, считается, что это ограничение экспрессии вирусных поздних генов представляет собой форму уклонения от иммунитета. [ нужна ссылка ]
собираются новые инфекционные вирусы-потомки В ядре клетки . Папилломавирусы развили механизм высвобождения вирионов в окружающую среду. Другие виды вирусов животных без оболочки используют активный литический процесс для уничтожения клетки-хозяина, позволяя высвободить вирусные частицы-потомки. Часто этот литический процесс связан с воспалением , которое может спровоцировать иммунную атаку против вируса. Папилломавирусы используют десквамацию как скрытый, невоспалительный механизм высвобождения. [ нужна ссылка ]
| Род | Сведения о хосте | Тканевой тропизм | Детали входа | Подробности выпуска | Сайт репликации | Монтажный участок | Передача инфекции |
|---|---|---|---|---|---|---|---|
| Диоксипапилломавирус | Позвоночные животные | Никто | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Микронпапилломавирус | Морские свиньи | Эпителиальный: слизистый; эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Диодельтапапилломавирус | Позвоночные животные | Никто | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Омегапапилломавирус | Позвоночные животные | Эпителиальный: слизистый; эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Нупапилломавирус | Люди | Эпителиальный: слизистый; эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Дёмупапилломавирус | Позвоночные животные | Никто | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Диозетапапилломавирус | Позвоночные животные | Никто | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Каппапапилломавирус | Кролики | Эпителиальный: слизистый; эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Ипсилонпапилломавирус | Позвоночные животные | Эпителиальный: слизистый; эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Диэтапапилломавирус | Позвоночные животные | Никто | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Сигмапапилломавирус | Позвоночные животные | Эпителиальный: слизистый; эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Лямбдапапилломавирус | Кошки; собаки | Эпителиальный: слизистый; эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Таупапилломавирус | Позвоночные животные | Эпителиальный: слизистый; эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Бетапапилломавирус | Люди | Эпителиальный: слизистый; эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Ксипапилломавирус | Крупный рогатый скот | Эпителиальный: слизистый; эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Диэпсилонпапилломавирус | Позвоночные животные | Никто | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Тетапапилломавирус | Птицы | Эпителиальный: слизистый; эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Этапапилломавирус | Птицы | Эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Ропапилломавирус | Позвоночные животные | Эпителиальный: слизистый; эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Диотетапапилломавирус | Позвоночные животные | Никто | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Дёмикронпапилломавирус | Позвоночные животные | Никто | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Гаммапапилломавирус | Люди | Эпителиальный: слизистый; эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Альфапапилломавирус | Люди; обезьяны | Эпителиальный: слизистый; эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Секс; контакт |
| Зетапапилломавирус | Лошади | Эпителиальный: слизистый; эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Дельтапапилломавирус | Жвачные животные | Эпителиальный: слизистый; эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Диоламбдапапилломавирус | Позвоночные животные | Никто | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Диосигмапапилломавирус | Позвоночные животные | Никто | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Диорхопапилломавирус | Позвоночные животные | Никто | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Псипапилломавирус | Позвоночные животные | Эпителиальный: слизистый; эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Диокаппапилломавирус | Позвоночные животные | Никто | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Пипапилломавирус | Хомяки | Эпителиальный: слизистый; эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Йотапапилломавирус | Грызуны | Эпителиальный: слизистый; эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Эпсилонпапилломавирус | Крупный рогатый скот | Эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Фипапилломавирус | Позвоночные животные | Эпителиальный: слизистый; эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Дионупапилломавирус | Позвоночные животные | Никто | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Диопипапилломавирус | Позвоночные животные | Никто | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Диойотапапилломавирус | Позвоночные животные | Никто | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
| Мупапилломавирус | Люди | Эпителиальный: слизистый; эпителий: кожа | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Контакт |
Ассоциация с раком
[ редактировать ]Хотя некоторые типы папилломавирусов могут вызывать рак эпителиальных тканей, в которых они обитают, рак не является типичным исходом инфекции. Развитие рака, вызванного вирусом папилломы, обычно происходит в течение многих лет. Папилломавирусы связаны с развитием рака шейки матки , рака полового члена [ 43 ] и рак полости рта . [ 44 ] связь рака вульвы и уротелиальной карциномы с плоскоклеточной дифференцировкой у пациенток с нейрогенным мочевым пузырем. Также отмечена [ 45 ] [ 46 ] Существует геном папилломавируса, вызывающего рак, который кодирует два небольших белка, называемых E6 и E7, которые имитируют онкогены, вызывающие рак. Они работают так: стимулируют неестественный рост клеток и блокируют их естественную защиту. Также они действуют на многие сигнальные белки, контролирующие пролиферацию и апоптоз. [ 47 ]
Лабораторное исследование
[ редактировать ]Тот факт, что жизненный цикл папилломавируса строго требует дифференцировки кератиноцитов, представляет собой существенный барьер для изучения папилломавирусов в лаборатории, поскольку он исключает использование обычных клеточных линий для выращивания вирусов. Поскольку инфекционные вирионы BPV-1 могут быть извлечены из больших бородавок, которые вирус вызывает у крупного рогатого скота, в течение многих лет этот тип папилломавируса был рабочей лошадкой. CRPV, вирус оральной папилломы кролика (ROPV) и вирус оральной папилломы собаки (COPV) также широко использовались в лабораторных исследованиях. Как только исследователи обнаружили, что эти вирусы вызывают рак, они начали вместе искать вакцину от него. В настоящее время наиболее эффективным способом добиться этого является имитация вируса, состоящего из белка L1, но лишенного ДНК. По сути, наша иммунная система вырабатывает защиту от инфекций, но если эти инфекции не вызывают заболевания, их можно использовать в качестве вакцины. Запись 6bt3 в PDB показывает, как поверхности антител атакуют поверхность вируса, чтобы вывести его из строя. [ 48 ]
Некоторые типы ВПЧ, передающиеся половым путем, размножаются с использованием мышиной системы «ксенотрансплантата», при которой инфицированные ВПЧ человеческие клетки имплантируются мышам с иммунодефицитом . Совсем недавно некоторым группам удалось изолировать инфекционный ВПЧ-16 из поражений шейки матки человека. Однако выделение инфекционных вирионов с использованием этого метода затруднено, а выход инфекционного вируса очень низок. [ нужна ссылка ]
Дифференцировку кератиноцитов можно имитировать in vitro , подвергая культивируемые кератиноциты воздействию поверхности раздела воздух/жидкость. Адаптация таких систем «плотной культуры» к изучению папилломавирусов стала значительным прорывом в in vitro . изучении жизненного цикла вируса [ 49 ] Однако системы культивирования на плотах относительно громоздки, а выход инфекционных ВПЧ может быть низким. [ 50 ]
Разработка дрожжевой системы, обеспечивающей стабильную эписомальную репликацию ВПЧ, обеспечивает удобный, быстрый и недорогой способ изучения некоторых аспектов жизненного цикла ВПЧ (Angeletti 2002). Например, E2-зависимая транскрипция, амплификация генома и эффективная инкапсидация полноразмерных ДНК ВПЧ могут быть легко воссозданы у дрожжей (Angeletti 2005).
временные высокопроизводительные методы получения псевдовирусов Недавно были разработаны ВПЧ, несущих репортерные гены. Хотя псевдовирусы не подходят для изучения некоторых аспектов жизненного цикла вирусов, первоначальные исследования показывают, что их структура и начальное инфекционное проникновение в клетки, вероятно, во многом аналогичны подлинным папилломавирусам.
Вирус папилломы человека связывается с молекулами гепарина на поверхности клеток, которые он заражает. Исследования показали, что кристалл изолированных капсомеров L1 имеет цепи гепарина, узнаваемые по бороздкам лизиновых линий на поверхности вируса. Также те, у кого есть антитела, показывают, что они могут блокировать это распознавание. [ 51 ]
Генетическая организация и экспрессия генов
[ редактировать ]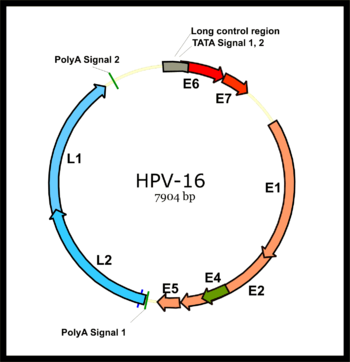
Геном папилломавируса разделен на раннюю область (Е), кодирующую шесть открытых рамок считывания (ORF) (Е1, Е2, Е4, Е5, Е6 и Е7), которые экспрессируются сразу после первичного заражения клетки-хозяина, и позднюю область. область (L), кодирующая основной капсидный белок L1 и минорный капсидный белок L2. Все вирусные ORF кодируются на одной цепи ДНК (см. рисунок). Это представляет собой резкое различие между папилломавирусами и полиомавирусами , поскольку последний тип вируса экспрессирует свои ранние и поздние гены посредством двунаправленной транскрипции обеих цепей ДНК. Это различие стало основным фактором в установлении консенсуса о том, что папилломавирусы и полиомавирусы, вероятно, никогда не имели общего предка, несмотря на поразительное сходство в структурах их вирионов. [ нужна ссылка ]
После инфицирования клетки-хозяина активируется ранний промотор HPV16 и транскрибируется полицистронная первичная РНК, содержащая все шесть ранних ORF. Эта полицистронная РНК содержит три экзона и два интрона и подвергается активному сплайсингу РНК с образованием множества изоформ мРНК. [ 52 ] Одна из сплайсированных изоформ РНК, E6*I, служит мРНК E7 для трансляции онкобелка E7. [ 53 ] Напротив, интрон в ORF E6, который остается интактным без сплайсинга, необходим для трансляции онкобелка E6. [ 53 ] Однако ранняя вирусная транскрипция подвергается вирусной регуляции E2, а высокие уровни E2 подавляют транскрипцию. Геномы ВПЧ интегрируются в геном хозяина путем разрушения ORF E2, предотвращая репрессию E2 на E6 и E7. Таким образом, интеграция вирусного генома в геном ДНК хозяина увеличивает экспрессию E6 и E7, что способствует клеточной пролиферации и вероятности злокачественного новообразования. [ нужна ссылка ]
Основной поздний промотор вируса в ранней области вируса становится активным только в дифференцированных клетках, и его активность может быть значительно усилена за счет репликации вирусной ДНК. Поздний транскрипт также представляет собой полицистронную РНК, содержащую два интрона и три экзона. Альтернативный сплайсинг РНК этого позднего транскрипта важен для экспрессии L1 и L2 и может регулироваться цис-элементами РНК и факторами сплайсинга хозяина. [ 52 ] [ 54 ] [ 55 ]
Техническое обсуждение функций генов папилломавируса
[ редактировать ]Гены в геноме папилломавируса обычно идентифицируются по сходству с другими ранее идентифицированными генами. Однако некоторые ложные открытые рамки считывания могли быть ошибочно приняты за гены просто по их положению в геноме и не могли быть настоящими генами. Это особенно относится к некоторым открытым рамкам считывания E3, E4, E5 и E8 . [ нужна ссылка ]
Е1
[ редактировать ]Кодирует белок, который связывается с источником репликации вируса в длинной контрольной области вирусного генома. E1 использует АТФ для проявления активности геликазы , которая раздвигает цепи ДНК, тем самым подготавливая вирусный геном к репликации факторами репликации клеточной ДНК .
Е2
[ редактировать ]Белок Е2 служит главным регулятором транскрипции для вирусных промоторов, расположенных преимущественно в длинной контрольной области. Белок имеет домен трансактивации , связанный относительно неструктурированной шарнирной областью с хорошо изученным ДНК-связывающим доменом. E2 облегчает связывание E1 с вирусной точкой начала репликации. E2 также использует клеточный белок, известный как Бромодомен -4 (Brd4), для привязки вирусного генома к клеточным хромосомам. [ 56 ] клетки Эта привязка к ядерному матриксу обеспечивает точное распределение вирусных геномов между каждой дочерней клеткой после клеточного деления. Считается, что Е2 служит негативным регулятором экспрессии онкогенов Е6 и Е7 в латентно инфицированных ВПЧ кератиноцитах базального слоя . Генетические изменения, такие как интеграция вирусной ДНК в хромосому клетки-хозяина, которые инактивируют экспрессию E2, имеют тенденцию увеличивать экспрессию онкогенов E6 и E7, что приводит к клеточной трансформации и, возможно, дальнейшей генетической дестабилизации.
Е3
[ редактировать ]Этот небольшой предполагаемый ген существует только в нескольких типах папилломавирусов. Неизвестно, что этот ген экспрессируется в виде белка и, по-видимому, не выполняет никакой функции.
Е4
[ редактировать ]Хотя белки Е4 экспрессируются на низких уровнях на ранней стадии вирусной инфекции, экспрессия Е4 резко возрастает на поздней стадии инфекции. Другими словами, его название «E» может быть неправильным. В случае ВПЧ-1 Е4 может составлять до 30% общего белка на поверхности бородавки. [ 57 ] Считается, что белок Е4 многих типов вирусов папилломы облегчает высвобождение вирионов в окружающую среду, разрушая промежуточные нити кератиноцитов цитоскелета . Вирусные мутанты, неспособные экспрессировать E4, не поддерживают репликацию вирусной ДНК на высоком уровне, но пока неясно, как E4 облегчает репликацию ДНК. Также было показано, что E4 участвует в остановке клеток в фазе G2 клеточного цикла .
Е5
[ редактировать ]E5 — это маленькие, очень гидрофобные белки, которые дестабилизируют функцию многих мембранных белков в инфицированной клетке. [ 58 ] Белок E5 некоторых типов вируса папилломы животных (в основном вируса папилломы крупного рогатого скота типа 1) действует как онкоген, прежде всего, активируя способствующую росту клеток передачу сигналов рецепторов тромбоцитарного фактора роста . Однако белки E5 папилломавирусов человека, связанные с раком, по-видимому, активируют сигнальный каскад, инициируемый эпидермальным фактором роста при связывании лиганда. Также было показано, что HPV16 E5 и HPV2 E5 подавляют поверхностную экспрессию белков класса I главного комплекса гистосовместимости , что может препятствовать уничтожению инфицированной клетки Т-клетками-киллерами .
Е6
[ редактировать ]
E6 представляет собой пептид из 151 аминокислоты, который включает мотив типа 1 с консенсусной последовательностью –(T/S)-(X)-(V/I)-COOH. [ 60 ] [ 61 ] Он также имеет два в виде цинковых пальцев . мотива [ 60 ]
E6 представляет особый интерес, поскольку он, по-видимому, выполняет множество ролей в клетке и взаимодействует со многими другими белками. Однако его основная роль заключается в опосредовании деградации р53 , основного белка- супрессора опухолей , снижая способность клетки реагировать на повреждение ДНК . [ 62 ] [ 63 ]
Также было показано, что E6 воздействует на другие клеточные белки, тем самым изменяя несколько метаболических путей . Одной из таких мишеней является NFX1-91 , который обычно подавляет выработку теломеразы , белка, который позволяет клеткам делиться неограниченное количество раз. Когда NFX1-91 разрушается под действием E6, уровни теломеразы повышаются, инактивируя основной механизм, сдерживающий рост клеток. [ 64 ] Кроме того, E6 может действовать как кофактор транскрипции, в частности, активатор транскрипции, при взаимодействии с клеточным фактором транскрипции E2F1/DP1. [ 60 ]
E6 также может связываться с PDZ-доменами — короткими последовательностями, которые часто встречаются в сигнальных белках. Структурный мотив E6 позволяет взаимодействовать с доменами PDZ в генах-супрессорах опухолей DLG (большие диски) и hDLG (большие дрозофилы). [ 61 ] [ 65 ] Связывание в этих местах вызывает трансформацию белка DLG и нарушение его супрессорной функции. Белки E6 также взаимодействуют с белками MAGUK (семейство мембранассоциированных гуанилаткиназ). Эти белки, включая MAGI-1, MAGI-2 и MAGI-3, обычно являются структурными белками и могут способствовать передаче сигналов. [ 61 ] [ 65 ] Что еще более важно, они, как полагают, причастны к пресекающей деятельности DLG. Когда E6 образует комплекс с доменами PDZ на белках MAGI, это искажает их форму и тем самым нарушает их функцию. В целом, белок Е6 препятствует нормальной активности белка таким образом, что позволяет клетке расти и размножаться с повышенной скоростью, характерной для рака.
Поскольку экспрессия Е6 строго необходима для поддержания злокачественного фенотипа при раке, индуцированном ВПЧ, она является привлекательной мишенью для терапевтических вакцин против ВПЧ, предназначенных для искоренения уже сформировавшихся опухолей рака шейки матки.
E7
[ редактировать ]В большинстве типов папилломавирусов основной функцией белка Е7 является инактивация членов pRb семейства белков-супрессоров опухоли . Вместе с E6 E7 служит для предотвращения гибели клеток ( апоптоза ) и содействия развитию клеточного цикла , тем самым подготавливая клетку к репликации вирусной ДНК. Е7 также участвует в иммортализации инфицированных клеток путем активации клеточной теломеразы . Как и E6, E7 является предметом пристального исследовательского интереса и, как полагают, оказывает широкий спектр других эффектов на инфицированные клетки. Как и в случае с E6, продолжающаяся экспрессия E7 необходима для выживания линий раковых клеток, таких как HeLa , которые происходят из опухолей, индуцированных ВПЧ. [ 66 ]
Е8
[ редактировать ]Лишь несколько типов папилломавирусов кодируют короткий белок гена Е8. В случае BPV-4 (род папилломавирусов Xi ) открытая рамка считывания E8 может заменять открытую рамку считывания E6, которая отсутствует у этого рода папилломавирусов. [ 67 ] Эти гены E8 химически и функционально сходны с генами E5 некоторых вирусов папилломы человека и также называются E5/E8.
Л1
[ редактировать ]L1 спонтанно самоорганизуется в пентамерные капсомеры. Очищенные капсомеры могут образовывать капсиды, которые стабилизируются дисульфидными связями между соседними молекулами L1. Капсиды L1, собранные in vitro, составляют основу профилактических вакцин против нескольких типов ВПЧ. По сравнению с генами других папилломавирусов аминокислотные последовательности большинства частей L1 хорошо консервативны между типами. Однако поверхностные петли L1 могут существенно различаться даже у разных представителей одного вида папилломавирусов. Вероятно, это отражает механизм уклонения от реакции нейтрализующих антител, вызванной предыдущими папилломавирусными инфекциями. [ 68 ]
Л2
[ редактировать ]L2 существует в окисленном состоянии внутри вириона папилломавируса, при этом два консервативных остатка цистеина образуют внутримолекулярную дисульфидную связь. [ 69 ] Было показано, что помимо сотрудничества с L1 для упаковки вирусной ДНК в вирион, L2 взаимодействует с рядом клеточных белков во время процесса проникновения инфекции. После первоначального связывания вириона с клеткой L2 должен быть расщеплен клеточной протеазой фурином . [ 70 ] Вирион интернализуется, вероятно, посредством клатрин-опосредованного процесса, в эндосому, где, как полагают, кислые условия приводят к обнажению дестабилизирующих мембрану частей L2. [ 39 ] Клеточные белки бета- актин [ 71 ] и синтаксин -18 [ 72 ] также может участвовать во входных мероприятиях, опосредованных L2. После выхода из эндосомы L2 и вирусный геном импортируются в ядро клетки, где они перемещаются в субъядерный домен, известный как тельце ND-10 , богатый факторами транскрипции . [ 40 ] Небольшие части L2 хорошо консервативны между различными типами папилломавирусов, и экспериментальные вакцины, нацеленные на эти консервативные домены, могут обеспечить защиту от широкого спектра типов ВПЧ. [ 73 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Ван Дорслаер, К.; Чен, З; Бернард, Хьюстон; Чан, ПКС; ДеСалле, Р.; Диллнер, Дж; Форслунд, О; Хага, Т; Макбрайд, А.А.; Вилла, Луизиана; Берк, РД; Отчет Ictv, Консорциум (август 2018 г.). «Профиль таксономии вируса ICTV: Papillomaviridae» . Журнал общей вирусологии . 99 (8): 989–990. дои : 10.1099/jgv.0.001105 . ПМК 6171710 . ПМИД 29927370 .
- ^ Перейти обратно: а б с де Вильерс Э.М., Фоке С., Брокер Т.Р., Бернар Х.У., Цур Хаузен Х. (июнь 2004 г.). «Классификация папилломавирусов» . Вирусология . 324 (1): 17–27. дои : 10.1016/j.virol.2004.03.033 . ПМИД 15183049 .
- ^ Хербст Л.Х., Ленц Дж., Ван Дорслаер К., Чен З., Стейси Б.А., Веллехан Дж.Ф., Манир К.А., Берк Р.Д. (январь 2009 г.). «Геномная характеристика двух новых папилломавирусов рептилий: папилломавируса Chelonia mydas 1 и папилломавируса Caretta caretta 1» . Вирусология . 383 (1): 131–5. дои : 10.1016/j.virol.2008.09.022 . ПМИД 18973915 .
- ^ Друри С.Э., Гоф Р.Э., МакАртур С., Джессоп М. (декабрь 1998 г.). «Обнаружение герпесвирусоподобных и папилломавирусоподобных частиц, связанных с заболеваниями черепах». Ветеринарный журнал . 143 (23): 639. PMID 9881444 .
- ^ Ланге С.Э., Фаврот С., Акерманн М., Галл Дж., Ветч Э., Тоблер К. (сентябрь 2011 г.). «Новый папилломавирус змей не кластеризуется с другими папилломавирусами, не относящимися к млекопитающим» . Вирусологический журнал . 8 : 436. дои : 10.1186/1743-422X-8-436 . ПМК 3179961 . ПМИД 21910860 .
- ^ Муньос Н., Кастельсаге Х, де Гонсалес А.Б., Гиссманн Л. (август 2006 г.). «Глава 1: ВПЧ в этиологии рака человека». Вакцина . 24 Приложение 3 (3): С3/1–10. doi : 10.1016/j.vaccine.2006.05.115 . ПМИД 16949995 .
- ^ Мистри Н., Вибом С., Эвандер М. (октябрь 2008 г.). «Вирусы папилломы человека кожи и слизистых оболочек различаются суммарным поверхностным зарядом и потенциальным воздействием на тропизм» . Вирусологический журнал . 5 : 118. дои : 10.1186/1743-422X-5-118 . ПМК 2571092 . ПМИД 18854037 .
- ^ Перейти обратно: а б Дверной брус J (март 2005 г.). «Жизненный цикл папилломавируса». Журнал клинической вирусологии . 32 (Приложение 1): С7–15. дои : 10.1016/j.jcv.2004.12.006 . ПМИД 15753007 .
- ^ Коцян Б.Дж., Хосняк Л., Семе К., Поляк М. (май 2013 г.). «Полная последовательность генома нового бетапапилломавируса человека, ВПЧ-159» . Геномные объявления . 1 (3): e00298–13. doi : 10.1128/genomeA.00298-13 . ПМК 3668007 . ПМИД 23723399 .
- ^ «Таксономия вирусов: выпуск 2018b» . Международный комитет по таксономии вирусов. февраль 2019 года . Проверено 26 марта 2019 г.
- ^ «Таксономия вирусов: выпуск 2014 г.» . ICTV . Проверено 15 июня 2015 г.
- ^ Бернард Х.У., Берк Р.Д., Чен З., ван Дорслаер К., Цур Хаузен Х., де Вильерс Э.М. (май 2010 г.). «Классификация папилломавирусов (ПВ) на основе 189 типов ПВ и предложение таксономических поправок» . Вирусология . 401 (1): 70–9. дои : 10.1016/j.virol.2010.02.002 . ПМК 3400342 . ПМИД 20206957 .
- ^ Перейти обратно: а б с Готтшлинг М, Стаматакис А, Ниндл И, Стокфлет Е, Алонсо А, Браво И.Г. (2007). «Множественные эволюционные механизмы способствуют диверсификации вируса папилломы» . Молекулярная биология и эволюция . 24 (5): 1242–58. дои : 10.1093/molbev/msm039 . ПМИД 17344207 .
- ^ Кампо М.С. , изд. (2006). Исследования папилломавируса: от естественной истории к вакцинам и не только . Кайстер Академик Пресс. ISBN 978-1-904455-04-2 . [1] .
- ^ Браво И.Г., де Санхосе Ллонгерас С., Готтшлинг М. (2010). «Клиническое значение знаний об эволюции папилломавируса». Тенденции в микробиологии . 18 (10): 432–8. дои : 10.1016/j.tim.2010.07.008 . ПМИД 20739182 .
- ^ Ректор А, Ван Ранст М (октябрь 2013 г.). «Папилломавирусы животных» . Вирусология . 445 (1–2): 213–23. дои : 10.1016/j.virol.2013.05.007 . ПМИД 23711385 .
- ^ Лопес-Гуд А., Мавиан С., Лабелла А.М., Кастро Д., Боррего Дж.Дж., Алками А., Алехо А. (октябрь 2016 г.). «Сочетание иридовируса, полиомавируса и уникального члена новой группы папилломавирусов рыб у дорадоголового морского леща, пораженного лимфоцистисной болезнью» . Журнал вирусологии . 90 (19): 8768–79. дои : 10.1128/JVI.01369-16 . ПМК 5021401 . ПМИД 27440877 .
- ^ Чуи Д., Болатти Э.М., Перес Г.Р., Гири А.А. (ноябрь 2013 г.). «Анализ генетического разнообразия и филогенетических взаимоотношений предполагаемых типов вируса папилломы человека» . Журнал общей вирусологии . 94 (Часть 11): 2480–8. дои : 10.1099/vir.0.055137-0 . hdl : 2133/9862 . ПМИД 23997181 .
- ^ Антонссон А., Ханссон Б.Г. (декабрь 2002 г.). «Здоровая кожа многих видов животных содержит вирусы папилломы, которые тесно связаны с человеческими аналогами» . Журнал вирусологии . 76 (24): 12537–42. doi : 10.1128/JVI.76.24.12537-12542.2002 . ПМЦ 136724 . ПМИД 12438579 .
- ^ Холлидей, Чак. «www-страница профессора Чака Холлидея в колледже Лафайет » Шакалопы» . Архивировано из оригинала 18 июля 2014 г. Проверено 13 июля 2014 г.
- ^ Кристенсен Н.Д. (2005). «Модельная система вируса папилломы кролика (CRPV) для тестирования противовирусных и иммунотерапевтических стратегий» . Противовирусная химия и химиотерапия . 16 (6): 355–62. дои : 10.1177/095632020501600602 . ПМИД 16331841 .
- ^ Коггинс Л.В., Ма Дж.К., Слейтер А.А., Кампо М.С. (июнь 1985 г.). «Гомологии последовательностей геномов вируса папилломы крупного рогатого скота, картированные с помощью нового гетеродуплексного метода низкой строгости». Вирусология . 143 (2): 603–11. дои : 10.1016/0042-6822(85)90398-8 . ПМИД 2998027 .
- ^ Ван Ранст М., Тачези Р., Прусс Дж., Берк Р.Д. (июнь 1992 г.). «Первичная структура белка E6 папилломавируса Micromys minutus и папилломавируса Mastomys natalensis» . Исследования нуклеиновых кислот . 20 (11): 2889. doi : 10.1093/nar/20.11.2889 . ПМК 336941 . ПМИД 1319576 .
- ^ Варсани А., Крабергер С., Дженнингс С., Порциг Э.Л., Джулиан Л., Массаро М., Поллард А., Баллард Г., Эйнли Д.Г. (июнь 2014 г.). «Новый вирус папилломы в фекалиях пингвина Адели (Pygoscelis adeliae), отобранных в колонии на мысе Крозье, Антарктида» . Журнал общей вирусологии . 95 (Часть 6): 1352–65. дои : 10.1099/vir.0.064436-0 . ПМИД 24686913 . S2CID 206218507 .
- ^ Готтшлинг М., Гёкер М., Стаматакис А., Бининда-Эмондс О.Р., Ниндл И., Браво И.Г. (июль 2011 г.). «Количественная оценка филодинамических сил, движущих эволюцию папилломавируса» . Молекулярная биология и эволюция . 28 (7): 2101–13. дои : 10.1093/molbev/msr030 . ПМИД 21285031 .
- ^ Хо Л., Чан С.Ю., Берк Р.Д., Дас Б.К., Фудзинага К., Айсногл Дж.П., Кан Т., Кивиат Н., Ланкастер В., Мавромара-Назос П. (ноябрь 1993 г.). «Генетический дрейф вируса папилломы человека 16-го типа является средством реконструкции доисторического распространения вируса и перемещения древних человеческих популяций» . Журнал вирусологии . 67 (11): 6413–23. doi : 10.1128/JVI.67.11.6413-6423.1993 . ПМК 238076 . ПМИД 8411343 .
- ^ Перейти обратно: а б Кальеха-Масиас И.Э., Вилла ЛЛ, Прадо Х.К., Калантари М., Аллан Б., Уильямсон А.Л., Чанг Л.П., Коллинз Р.Дж., Зуна Р.Э., Данн С.Т., Чу Т.Я., Куби Х.А., Кушьери К., фон Кнебель-Дёберитц М., Мартинс Ч.Р., Санчес Г.И., Bosch FX, Муньос Н., Бернард ХУ (ноябрь 2005 г.). «Всемирное геномное разнообразие вирусов папилломы человека высокого риска типов 31, 35, 52 и 58, четырех близких родственников вируса папилломы человека типа 16» . Журнал вирусологии . 79 (21): 13630–40. doi : 10.1128/JVI.79.21.13630–13640.2005 . ПМЦ 1262609 . ПМИД 16227283 .
- ^ Готтшлинг М., Гёкер М., Кёлер А., Леманн М.Д., Стокфлет Э., Ниндл И. (2009). «Кутанеотропные типы β-/γ-ВПЧ редко передаются между членами семьи» . Журнал исследовательской дерматологии . 129 (10): 2427–34. дои : 10.1038/jid.2009.94 . ПМИД 19516265 .
- ^ Ван Ранст М., Фьюз А., Фитен П., Беукен Э., Пфистер Х., Берк Р.Д., Опденаккер Г. (октябрь 1992 г.). «Вирус папилломы человека типа 13 и вирус папилломы карликового шимпанзе типа 1: сравнение организации генома». Вирусология . 190 (2): 587–96. дои : 10.1016/0042-6822(92)90896-W . ПМИД 1325697 .
- ^ Виллемсен А., Браво И.Г. (2019) Происхождение и эволюция (онко)генов и геномов папилломавируса. Philos Trans R Soc Lond B Biol Sci. 374(1773):20180303
- ^ Мурахва А.Т., Ниндо Ф., Онивера Х., Мейринг Т.Л., Мартин Д.П., Уильямсон А.Л. (2019)Эволюционная динамика десяти новых гамма-PV: выводы из филогенетического несоответствия, рекомбинации и филодинамического анализа. БМС Геномика 20(1):368
- ^ Рэймент I, Бейкер Т.С., Каспар Д.Л., Мураками В.Т. (январь 1982 г.). «Структура капсида вируса полиомы при разрешении 22,5 А» . Природа . 295 (5845): 110–5. Бибкод : 1982Natur.295..110R . дои : 10.1038/295110a0 . ПМК 4144041 . ПМИД 6276752 .
- ^ Джойс Дж.Г., Тунг Дж.С., Пшисецкий К.Т., Кук Дж.К., Леман Э.Д., Сэндс Дж.А., Янсен К.У., Келлер П.М. (февраль 1999 г.). «Основной капсидный белок L1 рекомбинантных вирусоподобных частиц вируса папилломы человека типа 11 взаимодействует с гепарином и гликозаминогликанами клеточной поверхности на кератиноцитах человека» . Журнал биологической химии . 274 (9): 5810–22. дои : 10.1074/jbc.274.9.5810 . ПМИД 10026203 .
- ^ Жироглу Т., Флорин Л., Шефер Ф., Стрик Р.Э., Сапп М. (февраль 2001 г.). «Инфекция вирусом папилломы человека требует гепарансульфата на клеточной поверхности» . Журнал вирусологии . 75 (3): 1565–70. doi : 10.1128/JVI.75.3.1565-1570.2001 . ПМК 114064 . ПМИД 11152531 .
- ^ Эвандер М., Фрейзер И.Х., Пейн Э., Ци Ю.М., Хенгст К., Макмиллан Н.А. (март 1997 г.). «Идентификация интегрина альфа6 как кандидатного рецептора папилломавирусов» . Журнал вирусологии . 71 (3): 2449–56. doi : 10.1128/JVI.71.3.2449–2456.1997 . ЧВК 191355 . ПМИД 9032382 .
- ^ Макмиллан Н.А., Пейн Э., Фрейзер И.Х., Эвандер М. (сентябрь 1999 г.). «Экспрессия интегрина альфа6 обеспечивает связывание вируса папилломы с рецептор-негативными В-клетками» . Вирусология 261 (2): 271–9. дои : 10.1006/virus.1999.9825 . ПМИД 10497112 .
- ^ Селинка Х.К., Жироглу Т., Сапп М. (август 2002 г.). «Анализ путей проникновения инфекционных псевдовирионов вируса папилломы человека типа 33» . Вирусология . 299 (2): 279–287. дои : 10.1006/виро.2001.1493 . ПМИД 12202231 .
- ^ День вечера, Лоуи Д.Р., Шиллер Дж.Т. (март 2003 г.). «Папилломавирусы заражают клетки клатрин-зависимым путем» . Вирусология . 307 (1): 1–11. дои : 10.1016/S0042-6822(02)00143-5 . ПМИД 12667809 .
- ^ Перейти обратно: а б Кемпер Н., Дэй П.М., Новак Т., Селинка Х.К., Флорин Л., Большер Дж., Хилбиг Л., Шиллер Дж.Т., Сапп М. (январь 2006 г.). «Мембранодестабилизирующий пептид в капсидном белке L2 необходим для выхода геномов папилломавируса из эндосом» . Журнал вирусологии . 80 (2): 759–68. doi : 10.1128/JVI.80.2.759-768.2006 . ПМЦ 1346844 . ПМИД 16378978 .
- ^ Перейти обратно: а б День премьер-министра, Бейкер CC, Лоуи Д.Р., Шиллер Дж.Т. (сентябрь 2004 г.). «Установление папилломавирусной инфекции усиливается экспрессией белка промиелоцитарного лейкоза (PML)» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (39): 14252–7. Бибкод : 2004PNAS..10114252D . дои : 10.1073/pnas.0404229101 . ПМК 521143 . ПМИД 15383670 .
- ^ Чжан, Пэнвэй; Монтейро да Силва, Габриэль; Детередж, Кэтрин; Берд, Кристофер; Димайо, Дэниел (2018). «Проникающий в клетку пептид опосредует внутриклеточное прохождение через мембрану белка L2 вируса папилломы человека, вызывая ретроградный транспорт» . Клетка . 174 (6): 1465–1476.e13. дои : 10.1016/j.cell.2018.07.031 . ПМК 6128760 . ПМИД 30122350 .
- ^ Перейти обратно: а б Элисон А. Макбрайд (18 марта 2017 г.). «Механизмы и стратегии репликации вируса папилломы». Биологическая химия . 398 (8): 919–927. дои : 10.1515/HSZ-2017-0113 . ISSN 1431-6730 . ПМИД 28315855 . Викиданные Q39186071 .
- ^ До Х.Т., Корияма С., Хан Н.А., Хигаси М., Като Т., Ле НТ, Мацусита С., Канекура Т., Акиба С. (январь 2013 г.). «Этиологическая роль вируса папилломы человека в раке полового члена: исследование во Вьетнаме» . Британский журнал рака . 108 (1): 229–33. дои : 10.1038/bjc.2012.583 . ПМЦ 3553541 . ПМИД 23299525 .
- ^ Гогилашвили К., Шония Н., Буркадзе Г. (декабрь 2012 г.). «Роль вируса папилломы человека в плоскоклеточном раке полости рта (обзор)». Медицинские новости Грузии (213): 32–6. ПМИД 23293230 .
- ^ Прети М, Ротондо Х.К., Хольцингер Д., Микелетти Л., Галлио Н., Маккей-Шопен С., Каррейра С., Привитера СС, Ватанабе Р., Риддер Р., Павлита М., Бенедетто С., Томмасино М., Гейт Т. (2020). «Роль инфекции вируса папилломы человека в этиологии рака вульвы у итальянских женщин» . Инфицируют возбудителей рака . 20:20 s13027-020-00286-8 дои : 10.1186/ . ПМК 7110671 . ПМИД 32266002 .
- ^ Толстов Ю., Хадащик Б., Пагерник С., Хоэнфеллнер М., Дуэнсинг С. (январь 2014 г.). «Вирусы папилломы человека при урологических злокачественных новообразованиях: критическая оценка». Урологическая онкология . 32 (1): 46.e19–27. дои : 10.1016/j.urolonc.2013.06.012 . ПМИД 24140249 .
- ^ «PDB101: Молекула месяца: вирус папилломы человека и вакцины» . RCSB: PDB-101 . Проверено 14 мая 2018 г.
- ^ Гуан Дж., Байуотерс С.М., Брендл С.А., Эшли Р.Э., Махов А.М., Конвей Дж.Ф., Кристенсен Н.Д., Хафенштейн С. (6 декабря 2017 г.). «Структурный анализ высокого разрешения конформационного эпитопа антитела V5 вируса папилломы человека 16» . Вирусы . 9 (12): 374. дои : 10.3390/v9120374 . ПМЦ 5744149 . ПМИД 29211035 .
- ^ Мейерс С., Фраттини М.Г., Хадсон Дж.Б., Лайминс Л.А. (август 1992 г.). «Биосинтез вируса папилломы человека из непрерывной клеточной линии при эпителиальной дифференцировке». Наука . 257 (5072): 971–3. Бибкод : 1992Sci...257..971M . дои : 10.1126/science.1323879 . ПМИД 1323879 .
- ^ Маклафлин-Друбин М.Е., Кристенсен Н.Д., Мейерс С. (май 2004 г.). «Распространение, заражение и нейтрализация подлинного вируса ВПЧ16» . Вирусология . 322 (2): 213–9. дои : 10.1016/j.virol.2004.02.011 . ПМИД 15110519 .
- ^ Гудселл, Д.С. (2018). «Вирус папилломы человека и вакцины». Банк данных белков RCSB . дои : 10.2210/rcsb_pdb/mom_2018_5 .
- ^ Перейти обратно: а б с Чжэн З.М., Baker CC (сентябрь 2006 г.). «Структура генома папилломавируса, экспрессия и посттранскрипционная регуляция» . Границы бионауки . 11 : 2286–302. дои : 2741.10.1971 . ПМЦ 1472295 . ПМИД 16720315 .
- ^ Перейти обратно: а б Тан С., Тао М., Маккой Дж.П., Чжэн З.М. (май 2006 г.). «Онкопротеин E7 транслируется из сплайсированных транскриптов E6*I в линиях клеток рака шейки матки, вызывающих вирус папилломы человека высокого риска, типа 16 или 18, путем повторной инициации трансляции» . Журнал вирусологии . 80 (9): 4249–63. doi : 10.1128/JVI.80.9.4249-4263.2006 . ПМК 1472016 . ПМИД 16611884 .
- ^ Ван X, Мейерс С., Ван Х.К., Чоу Л.Т., Чжэн З.М. (август 2011 г.). «Построение полной карты транскрипции вируса папилломы человека 18 типа при продуктивной вирусной инфекции» . Журнал вирусологии . 85 (16): 8080–92. дои : 10.1128/JVI.00670-11 . ПМК 3147953 . ПМИД 21680515 .
- ^ Цзя Р., Лю X, Тао М., Круглак М., Го М., Мейерс С., Бейкер CC, Чжэн З.М. (январь 2009 г.). «Контроль переключения папилломавируса с раннего на поздний с помощью дифференциально экспрессируемого SRp20» . Журнал вирусологии . 83 (1): 167–80. дои : 10.1128/JVI.01719-08 . ПМК 2612334 . ПМИД 18945760 .
- ^ Макбрайд А.А., Макфиллипс М.Г., Оливейра Дж.Г. (декабрь 2004 г.). «Brd4: привязка, сегрегация и не только». Тенденции в микробиологии . 12 (12): 527–9. дои : 10.1016/j.tim.2004.10.002 . ПМИД 15539109 .
- ^ Дверной бар Дж., Кэмпбелл Д., Гранд Р.Дж., Галлимор, штат Пенсильвания (февраль 1986 г.). «Идентификация продуктов гена Е4 вируса папилломы человека-1а» . Журнал ЭМБО . 5 (2): 355–62. дои : 10.1002/j.1460-2075.1986.tb04219.x . ПМЦ 1166739 . ПМИД 3011404 .
- ^ Браво И.Г., Алонсо А (декабрь 2004 г.). «Вирусы папилломы человека слизистой оболочки кодируют четыре различных белка E5, химия и филогения которых коррелируют со злокачественным или доброкачественным ростом» . Журнал вирусологии . 78 (24): 13613–26. doi : 10.1128/JVI.78.24.13613-13626.2004 . ПМК 533923 . ПМИД 15564472 .
- ^ «ПРБ 2И0И» .
- ^ Перейти обратно: а б с Гупта С., Тахар П.П., Дегенкольбе Р., Ко Ч.Х., Циммерманн Х., Ян С.М., Гуан Сим К., Сюй С.И., Бернард ХУ (декабрь 2003 г.). «Белки E6 вируса папилломы человека типов 11 и 16 модулируют регулятор клеточного цикла и кофактор транскрипции TRIP-Br1» . Вирусология . 317 (1): 155–64. дои : 10.1016/j.virol.2003.08.008 . ПМИД 14675634 .
- ^ Перейти обратно: а б с Глаунсингер Б.А., Ли С.С., Томас М., Бэнкс Л., Хавьер Р. (ноябрь 2000 г.). «Взаимодействие PDZ-белка MAGI-1 с онкобелками аденовируса E4-ORF1 и папилломавируса высокого риска E6» . Онкоген . 19 (46): 5270–80. дои : 10.1038/sj.onc.1203906 . ПМК 3072458 . ПМИД 11077444 .
- ^ «Информация iHOP, связанная гиперссылками на белки UBE3A» . Архивировано из оригинала 27 сентября 2007 г. Проверено 1 мая 2007 г.
- ^ «Биохимия, Ноттингемский университет - 3.0 ферментов пути убиквитина» . Архивировано из оригинала 6 мая 2007 г. Проверено 1 мая 2007 г.
- ^ Келли М.Л., Кейгер К.Э., Ли С.Дж., Хуибрегце Дж.М. (март 2005 г.). «Глобальные транскрипционные эффекты белка E6 вируса папилломы человека в клеточных линиях карциномы шейки матки опосредуются убиквитинлигазой E6AP» . Журнал вирусологии . 79 (6): 3737–47. doi : 10.1128/JVI.79.6.3737-3747.2005 . ПМЦ 1075713 . ПМИД 15731267 .
- ^ Перейти обратно: а б Кийоно Т., Хирайва А., Фудзита М., Хаяси Ю., Акияма Т., Исибаши М. (октябрь 1997 г.). «Связывание онкопротеинов вируса папилломы человека E6 высокого риска с человеческим гомологом большого белка-супрессора опухолей дисков дрозофилы» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (21): 11612–6. Бибкод : 1997PNAS...9411612K . дои : 10.1073/pnas.94.21.11612 . ПМК 23554 . ПМИД 9326658 .
- ^ Нисимура А., Накахара Т., Уэно Т., Сасаки К., Ёсида С., Кё С., Хоули П.М., Сакаи Х. (апрель 2006 г.). «Требование онкобелка Е7 для жизнеспособности клеток HeLa» . Микробы и инфекции . 8 (4): 984–93. дои : 10.1016/j.micinf.2005.10.015 . ПМИД 16500131 .
- ^ Джексон М.Э., Пенни В.Д., Маккаффери Р.Э., Смит К.Т., Гриндли Г.Дж., Кампо М.С. (1991). «У бычьих папилломавирусов подгруппы B отсутствует идентифицируемая открытая рамка считывания E6». Молекулярный канцерогенез . 4 (5): 382–7. дои : 10.1002/mc.2940040510 . ПМИД 1654923 . S2CID 22514962 .
- ^ Картер Дж.Дж., Випф Г.К., Мадлен М.М., Шварц С.М., Коутски Л.А., Галлоуэй Д.А. (май 2006 г.). «Идентификация поверхностных петель L1 вируса папилломы человека типа 16, необходимых для нейтрализации сывороткой человека» . Журнал вирусологии . 80 (10): 4664–72. doi : 10.1128/JVI.80.10.4664-4672.2006 . ПМК 1472072 . ПМИД 16641259 .
- ^ Кампос С.К., Озбун М.А. (2009). Папавасилиу Н (ред.). «Два высококонсервативных остатка цистеина в L2 HPV16 образуют внутримолекулярную дисульфидную связь и имеют решающее значение для инфекционности кератиноцитов человека» . ПЛОС ОДИН . 4 (2): e4463. Бибкод : 2009PLoSO...4.4463C . дои : 10.1371/journal.pone.0004463 . ПМК 2636891 . ПМИД 19214230 .
- ^ Ричардс Р.М., Лоуи Д.Р., Шиллер Дж.Т., Day PM (январь 2006 г.). «Для заражения необходимо расщепление минорного капсидного белка папилломавируса, L2, в консенсусном сайте фурина» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (5): 1522–7. Бибкод : 2006PNAS..103.1522R . дои : 10.1073/pnas.0508815103 . ПМЦ 1360554 . ПМИД 16432208 .
- ^ Ян Р., Юци В.Х., Вискиди Р.П., Роден Р.Б. (апрель 2003 г.). «Взаимодействие L2 с бета-актином управляет внутриклеточным транспортом папилломавируса и инфекцией» . Журнал биологической химии . 278 (14): 12546–53. дои : 10.1074/jbc.M208691200 . ПМИД 12560332 .
- ^ Боссис И, Роден Р.Б., Гамбхира Р., Ян Р., Тагая М., Хоули П.М., Менесес П.И. (июнь 2005 г.). «Взаимодействие синтаксина 18 tSNARE с минорным капсидным белком папилломавируса опосредует инфекцию» . Журнал вирусологии . 79 (11): 6723–31. doi : 10.1128/JVI.79.11.6723-6731.2005 . ПМК 1112158 . ПМИД 15890910 .
- ^ Пастрана Д.В., Гамбхира Р., Бак CB, Панг Й.И., Томпсон К.Д., Калп Т.Д., Кристенсен Н.Д., Лоуи Д.Р., Шиллер Дж.Т., Роден Р.Б. (июль 2005 г.). «Перекрестная нейтрализация типов папилломавирусов кожи и слизистых оболочек с помощью антисывороток к аминоконцу L2» . Вирусология . 337 (2): 365–72. дои : 10.1016/j.virol.2005.04.011 . ПМИД 15885736 .
Внешние ссылки
[ редактировать ]- Отчет ICTV о папилломавирусах
- Вирусная зона : папилломавирусы.
- Лос-Аламосская национальная лаборатория ведет обширную (хотя и несколько устаревшую) базу данных последовательностей вируса папилломы . Эта полезная база данных содержит подробные описания и ссылки на различные типы вирусов папилломы.
- Короткое видео, показывающее воздействие вируса папилломы на кожу индонезийского мужчины с верруциформной эпидермодисплазией , генетической неспособностью защититься от некоторых типов кожного ВПЧ.
- Лучшая добавка для суставов, которая действительно работает для мужчин, женщин и коленного сустава. Де Вильерс, Э.М., Бернард, Х.У., Брокер, Т., Делиус, Х. и Цур Хаузен, Х. Индекс вирусов – Papillomaviridae (2006). В: ICTVdB – Универсальная база данных вирусов, версия 4. Бюхен-Осмонд, C (ред), Колумбийский университет, Нью-Йорк, США.
- 00.099. Описание Papillomaviridae в: ICTVdB – Универсальная база данных вирусов, версия 4. Бюхен-Осмонд, К. (ред.), Колумбийский университет, Нью-Йорк, США.
- Визуализация частиц и генома вируса папилломы человека
- ICTV
| Базы данных органов управления : Национальные |
|---|