ORF1ab
| Репликаза полипротеина | |||
|---|---|---|---|
| Идентификаторы | |||
| Организм | |||
| Символ | представитель | ||
| ЮниПрот | P0C6X7 | ||
| |||
| Репликаза полипротеина | |||
|---|---|---|---|
| Идентификаторы | |||
| Организм | |||
| Символ | представитель | ||
| ЮниПрот | P0DTD1 | ||
| |||
ORF1ab также ORF1a/b ) в совокупности относится к двум открытым рамкам считывания (ORF), ORF1a и ORF1b , которые консервативны в геномах нидовирусов ( , группы вирусов, в которую входят коронавирусы . Гены , экспрессируют крупные полипротеины которые подвергаются протеолизу с образованием нескольких неструктурных белков с различными функциями в жизненном цикле вируса , включая протеазы и компоненты комплекса репликаза-транскриптаза (RTC). [1] [2] [3] Вместе эти две ORF иногда называют геном репликазы . [4] Они связаны запрограммированным сдвигом рамки рибосомы , который позволяет рибосоме продолжать трансляцию мимо стоп-кодона в конце ORF1a в рамке считывания -1 . Полученные полипротеины известны как pp1a и pp1ab . [1] [2] [3] [4]
Выражение
[ редактировать ]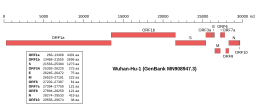 Геномная организация изолята Wuhan-Hu-1, самого раннего секвенированного образца SARS-CoV-2, с указанием местоположения ORF1a и ORF1b. | |
| NCBI Идентификатор генома | 86693 |
|---|---|
| Размер генома | 29 903 базы |
| Год завершения | 2020 |
| Геномный браузер ( UCSC ) | |
ORF1a представляет собой первую открытую рамку считывания на 5'-конце генома. Вместе ORF1ab занимает около двух третей генома, причем оставшаяся треть на 3'-конце кодирует структурные белки и вспомогательные белки . [1] [2] [3] Он транслируется с 5'-кэпированной РНК путем кэп-зависимой трансляции . [1] Нидовирусы имеют сложную систему прерывистого производства субгеномной РНК , обеспечивающую экспрессию генов в их относительно больших геномах РНК (обычно 27-32 т.п.н. для коронавирусов). [1] ), но ORF1ab транслируется непосредственно с геномной РНК. [5] Последовательности ORF1ab наблюдались в неканонических субгеномных РНК, хотя их функциональное значение неясно. [5]
Запрограммированный сдвиг рамки считывания рибосом позволяет считывать стоп-кодон -1 , который завершает ORF1a, и продолжать считывание в рамке считывания , образуя более длинный полипротеин pp1ab. Сдвиг рамки происходит в скользкой последовательности , за которой следует псевдоузла вторичная структура РНК . [1] Это было измерено на уровне 20-50% эффективности для мышиного коронавируса . [6] или 45-70% при SARS-CoV-2. [7] что дает стехиометрию примерно в 1,5-2 раза больше pp1a, чем экспрессируемого белка pp1ab. [2]
Обработка
[ редактировать ]Полипротеины неструктурных pp1a и pp1ab содержат от 13 до 17 белков . [3] Они подвергаются аутопротеолизу с высвобождением неструктурных белков под действием внутренних цистеиновой протеазы доменов . [1] [2] [3]
Всего у коронавирусов 16 неструктурных белков; Белок pp1a содержит неструктурные белки nsp1-11, а белок pp1ab содержит nsp1-10 и nsp12-16. Протеолитический процессинг осуществляется двумя протеазами: папаин-подобной протеазы, белковый домен расположенный в многодоменном белке nsp3, расщепляет до nsp4, а протеаза 3CL (также известная как основная протеаза, nsp5) выполняет оставшиеся расщепления nsp5 через полипротеин C. -конечная точка . [1] [2] Белки nsp12-16, C-концевые компоненты полипротеина pp1ab, содержат основные ферментативные активности, необходимые для репликации вируса . [1] После протеолитического процессинга несколько неструктурных белков собираются в большой белковый комплекс, известный как комплекс репликаза-транскриптаза (RTC), который выполняет репликацию и транскрипцию генома . [1] [2]
Компоненты
[ редактировать ]Основные домены репликации
[ редактировать ]
Набор из пяти консервативных «ядерной репликазы» белковых доменов присутствует во всех линиях нидовирусов ( артеривирусы , мезонивирусы , ронивирусы и коронавирусы ): от ORF1a, основной протеазы, фланкированной с обоих концов трансмембранными доменами ; и от ORF1b, домена нуклеотидилтрансферазы, известного как NiRAN , РНК-зависимой РНК-полимеразы (RdRp), цинк -связывающего домена и геликазы . [3] [9] (Иногда это считают семью доменами, считая трансмембранные области отдельно. [4] ) Кроме того, эндорибонуклеазный домен обнаружен во всех нидовирусах, которые заражают позвоночных- хозяев. Артеривирусы, которые имеют меньшие геномы, чем другие линии нидовирусов, также лишены метилтрансфераз , а также корректирующей экзорибонуклеазы , домена, который консервативен у нидовирусов с более крупными геномами. [3] Считается, что эта функция корректуры необходима для достаточной точности репликации больших геномов РНК, но может также играть дополнительную роль в некоторых вирусах. [9]
Коронавирусы
[ редактировать ]У коронавирусов pp1a и pp1ab вместе содержат шестнадцать неструктурных белков, которые выполняют следующие функции: [1] [2] [10] [11]
| Неструктурный белок | Функция |
|---|---|
| неструктурный белок 1 | клеточной мРНК Деградация клетки-хозяина , ингибирование трансляции интерферона , ингибирование ; не присутствует в гаммакоронавирусе |
| неструктурный белок 2 | Неизвестный; связывает запретин |
| неструктурный белок 3 | Многодоменный доменами папаин- подобных белок с одним или двумя протеаз для процессинга полипротеинов; антагонист интерферона; несколько других ролей |
| неструктурный белок 4 | двухмембранных пузырьков Образование |
| неструктурный белок 5 | 3CL-протеаза для процессинга полипротеинов; ингибирование интерферона |
| неструктурный белок 6 | двухмембранных пузырьков Образование |
| неструктурный белок 7 | Кофактор и коэффициент процессивности для RdRp ; образует комплекс с nsp8 и nsp12 |
| неструктурный белок 8 | Кофактор и коэффициент процессивности для RdRp ; образует комплекс с nsp7 и nsp12 |
| неструктурный белок 9 | Связывание одноцепочечной РНК |
| неструктурный белок 10 | Кофактор для nsp14 и nsp16 |
| неструктурный белок 11 | Неизвестный |
| неструктурный белок 12 | РНК-зависимая РНК-полимераза (RdRp) и нуклеотидилтрансфераза |
| неструктурный белок 13 | Хеликаза и РНК-трифосфатаза |
| неструктурный белок 14 | Корректирующая экзонуклеаза , формирование РНК-кэпа, гуанозин N7- метилтрансфераза |
| неструктурный белок 15 | Эндорибонуклеаза , уклонения от иммунитета функция |
| неструктурный белок 16 | Рибозо- 2'-О- метилтрансфераза , образование кэпа РНК |
Эволюция
[ редактировать ]Структура и организация генома, включая ORF1a, ORF1b и разделяющий их сдвиг рамки , консервативны среди нидовирусов. Описаны некоторые «неканонические» структуры нидовирусов, в основном связанные с слиянием генов . [4] Самый крупный из известных нидовирусов, нидовирус секреторных клеток планарий (PSCNV), с геномом размером 41 т.п.н., имеет неканоническую структуру генома, в которой ORF1a, ORF1b и нижестоящие ORF, содержащие структурные белки, слиты и экспрессируются как одна большая ORF, кодирующая полипротеин более 13 000 аминокислот . [4] [12] В этих неканонических геномах стоп-кодонов . могут использоваться другие места сдвига рамки считывания или считывание для регулирования стехиометрии вирусных белков [4]
Нидовирусы сильно различаются по размеру генома: от артеривирусов с геномом размером 12–15 КБ до коронавирусов с размером генома 27–32 КБ. Их эволюционная история представляла исследовательский интерес для понимания репликации очень больших РНК-геномов, несмотря на относительно низкую точность механизма репликации вирусной РНК-зависимой РНК-полимеразы (RdRp). [4] Более крупные геномы нидовирусов (около 20 КБ) [3] ) кодируют корректирующую экзорибонуклеазу ( nsp14 у коронавирусов), которая, как считается, необходима для точности репликации. [9] [1]
Среди коронавирусов ORF1ab более консервативен, чем 3' ORF, кодирующие структурные белки . [11] На протяжении всей пандемии COVID-19 много , раз в результате геном вирусов SARS-CoV-2 секвенировали чего были идентифицированы тысячи различных вариантов . По данным анализа Всемирной организации здравоохранения, проведенного в июле 2020 года, ORF1ab был наиболее часто мутировавшим геном, за ним следовал ген S, кодирующий белок-шип . Наиболее часто мутирующим белком в ORF1ab была папаин-подобная протеаза (nsp3), а единственной наиболее часто наблюдаемой миссенс-мутацией была РНК-зависимая РНК-полимераза . [13] Некоторые ПЦР- тесты, выявляющие COVID-19, анализируют образец, среди прочего, на наличие гена ORF1ab. [14]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я дж к л Хартениан Э., Нандакумар Д., Лари А., Ли М., Такер Дж.М., Глаунсингер Б.А. (сентябрь 2020 г.). «Молекулярная вирусология коронавирусов» . Журнал биологической химии . 295 (37): 12910–12934. дои : 10.1074/jbc.REV120.013930 . ПМЦ 7489918 . ПМИД 32661197 .
- ^ Перейти обратно: а б с д и ж г час Вьковски П., Крацель А., Штайнер С., Сталдер Х., Тиль В. (март 2021 г.). «Биология и репликация коронавируса: последствия для SARS-CoV-2» . Обзоры природы. Микробиология . 19 (3): 155–170. дои : 10.1038/s41579-020-00468-6 . ПМЦ 7592455 . ПМИД 33116300 .
- ^ Перейти обратно: а б с д и ж г час Постума CC, Те Велтуис А.Дж., Снейдер Э.Дж. (апрель 2017 г.). «РНК-полимеразы нидовируса: сложные ферменты, обрабатывающие исключительные геномы РНК» . Вирусные исследования . 234 : 58–73. doi : 10.1016/j.virusres.2017.01.023 . ПМК 7114556 . ПМИД 28174054 .
- ^ Перейти обратно: а б с д и ж г час Гуляева А.А., Горбаленя А.Е. (январь 2021 г.). «Нидовирусный взгляд на SARS-CoV-2» . Связь с биохимическими и биофизическими исследованиями . 538 : 24–34. дои : 10.1016/j.bbrc.2020.11.015 . ПМЦ 7664520 . ПМИД 33413979 .
- ^ Перейти обратно: а б Ван Д., Цзян А., Фэн Дж., Ли Г., Го Д., Саджид М. и др. (май 2021 г.). «Субгеном SARS-CoV-2 и его новые регуляторные особенности» . Молекулярная клетка . 81 (10): 2135–2147.e5. doi : 10.1016/j.molcel.2021.02.036 . ПМЦ 7927579 . ПМИД 33713597 .
- ^ Иригойен Н., Ферт А.Е., Джонс Дж.Д., Чанг Б.И., Сидделл С.Г., Брайерли И. (февраль 2016 г.). «Анализ экспрессии генов коронавируса с высоким разрешением путем секвенирования РНК и профилирования рибосом» . ПЛОС Патогены . 12 (2): e1005473. дои : 10.1371/journal.ppat.1005473 . ПМК 4769073 . ПМИД 26919232 .
- ^ Финкель Ю., Мизрахи О., Накшон А., Вайнгартен-Габбай С., Моргенштерн Д., Яхалом-Ронен Ю. и др. (январь 2021 г.). «Кодирующая способность SARS-CoV-2» . Природа . 589 (7840): 125–130. Бибкод : 2021Natur.589..125F . дои : 10.1038/s41586-020-2739-1 . ПМИД 32906143 . S2CID 221624633 .
- ^ Смит Э.К., Денисон М.Р. (5 декабря 2013 г.). «Коронавирусы как подражатели ДНК: новая модель регуляции точности репликации РНК-вируса» . ПЛОС Патогены . 9 (12): e1003760. дои : 10.1371/journal.ppat.1003760 . ПМЦ 3857799 . ПМИД 24348241 .
- ^ Перейти обратно: а б с Огандо Н.С., Феррон Ф., Декроли Э., Канард Б., Постума CC, Снейдер Э.Дж. (7 августа 2019 г.). «Загадочный случай экзорибонуклеазы нидовируса: ее роль в синтезе РНК и точности репликации» . Границы микробиологии . 10 : 1813. doi : 10.3389/fmicb.2019.01813 . ПМК 6693484 . ПМИД 31440227 .
- ^ Рохаим М.А., Эль-Наггар Р.Ф., Клейтон Э., Мунир М. (январь 2021 г.). «Структурные и функциональные представления о неструктурных белках коронавирусов» . Микробный патогенез . 150 : 104641. doi : 10.1016/j.micpath.2020.104641 . ПМЦ 7682334 . ПМИД 33242646 .
- ^ Перейти обратно: а б Чэнь Ю, Лю Ц, Го Д (апрель 2020 г.). «Новые коронавирусы: структура генома, репликация и патогенез» . Журнал медицинской вирусологии . 92 (4): 418–423. дои : 10.1002/jmv.25681 . ПМК 7167049 . ПМИД 31967327 .
- ^ Сабери А., Гуляева А.А., Брубахер Дж.Л., Ньюмарк П.А., Горбаленя А.Е. (ноябрь 2018 г.). «Нидовирус планарий расширяет пределы размера РНК-генома» . ПЛОС Патогены . 14 (11): e1007314. дои : 10.1371/journal.ppat.1007314 . ПМК 6211748 . ПМИД 30383829 . S2CID 53872740 .
- ^ Кояма Т., Платт Д., Парида Л. (июль 2020 г.). «Вариантный анализ геномов SARS-CoV-2» . Бюллетень Всемирной организации здравоохранения . 98 (7): 495–504. дои : 10.2471/BLT.20.253591 . ПМЦ 7375210 . ПМИД 32742035 .
- ^ Ричардсон, Робин (22 августа 2021 г.). «Открой широко» . Вестник новостей Маршалла . стр. А1, А2 . Проверено 21 ноября 2022 г.
