Рыбная масштаба

Шкала рыбы - это небольшая жесткая тарелка, которая растет из кожи рыбы. Кожа большинства челюстных рыб покрыта этими защитными масштабами , которые также могут обеспечить эффективный камуфляж с помощью отражения и окраски , а также возможных гидродинамических преимуществ. Термин «Шкала» происходит от старого французского эскале , что означает капсул или шелуху. [ 1 ]
Шкалы сильно различаются по размеру, форме, структуре и масштабу, начиная от прочных и жестких пластин до брони у рыб, таких как креветочные рыбы и ящики , до микроскопических или отсутствующих у рыб, таких как угри и рыболовные рыбы . Морфология . масштаба может быть использована для идентификации видов рыб, из которых он пришел Шкалы, возникшие в яростных остракодермах , предках для всех челюстных рыб сегодня. Большинство костных рыб покрыты циклоидными масштабами лосося и карпа , или Ctenoid Scals of Perch или ганоидными масштабами осетров и Gars . Рыбами хряща ( акулы и лучи ) покрыты плакоидными чешуйками. Вместо этого некоторые виды покрыты закусками , а другие не имеют внешнего покрытия на части или всей коже.
рыбы Рыбные чешуйки являются частью покровной системы и производятся из мезодермы дермы , что отличает их от масштабов рептилий . [ 2 ] [ 3 ] Те же гены, участвующие в развитии зубов и волос у млекопитающих, также участвуют в развитии масштаба. Плокоидные чешуйки хрящевых рыб также называют кожными зубцами и являются структурно гомологичными с зубцами позвоночных. Большинство рыб также покрыты слоем слизи или слизи, которые могут защищать от патогенных микроорганизмов, таких как бактерии, грибы и вирусы, и снижать устойчивость к поверхности, когда рыба плывет.
Thelodont Scales
[ редактировать ]
Бонистые чешуйки Телодонтов , наиболее распространенная форма ископаемой рыбы , хорошо понятны. Масштабы были сформированы и проливали на протяжении всей жизни организмов и быстро разлучены после их смерти. [ 4 ]
Кость, ткань, которая устойчива к механическому повреждению и относительно подвержена ископаемости, часто сохраняет внутреннюю деталь, что позволяет детально изучать гистологию и рост шкал. Масштабы составляют нераступирующую «корону», состоящую из дентина , с иногда эмалоидной верхней поверхностью эмалоидной эмалоидной поверхности и основой аспидина. [ 5 ] Его растущая основа сделана из бесклеточной кости, которая иногда развивалась структуры на якоре, чтобы исправить ее в стороне рыбы. [ 6 ] Кроме того, по-видимому, существует пять типов роста костей, которые могут представлять пять естественных групп в рамках телодонтов- или спектр, начиная между мета-членами конечными членами (или орто-) дентиновыми и мезодотенными тканями. [ 7 ] Каждое из пяти масштабных морфов, по -видимому, напоминает масштабы более производных групп рыб, что позволяет предположить, что группы Thelodont, возможно, были STEM -группами для последующих клад рыбы. [ 6 ]
Однако использование только морфологии шкалы для различения видов имеет некоторые ловушки. Внутри каждого организма форма масштаба сильно варьируется в зависимости от области тела, [ 8 ] С промежуточными формами, появляющимися между различными областями, и что еще хуже, морфология масштаба может даже не быть постоянной в пределах одной области. Чтобы еще больше путать вещи, морфологии масштаба не являются уникальными для таксонов и могут быть неотличимыми в одной и той же области двух разных видов. [ 9 ]
Морфология и гистология Thelodonts обеспечивают основной инструмент для количественной оценки их разнообразия и различения между видами, хотя в конечном итоге использование таких сходящихся признаков является подверженным ошибкам. Тем не менее, была предложена структура, состоящая из трех групп на основе морфологии и гистологии масштаба. [ 7 ] Сравнения с современными видами акул показали, что шкалы Thelodont были функционально аналогичны шкалам современной хрящевой рыбы и также позволили обширное сравнение между экологическими нишами. [ 10 ]
Космоидные весы
[ редактировать ]
Космоидные весы встречаются только на древних рыбах с доли , в том числе некоторых из самых ранних рыб легких (подкласса дипнои ) и в кроссоптеригии , включая живой коэлакант в модифицированной форме (см. Весы Elasmoid, ниже). Они, вероятно, были получены из слияния плакоид -ганоидных масштабов. Внутренняя часть весов изготовлена из плотной пластинчатой кости, называемой изопедином. Вдобавок к этому лежит слой губчатой или сосудистой кости, поставляемый кровеносными сосудами, за которым следует сложный дентиноподобный слой, называемый космином с поверхностным внешним покрытием витродентана . Верхняя поверхность - кератин . Космоидные масштабы увеличиваются в размере за счет роста слоя пластинчатого кости. [ 11 ]
Эламоидные масштабы
[ редактировать ]
Эламоидные чешуйки являются тонкими, имбрированными масштабами, состоящими из слоя плотной, пластинчатой коллагеновой кости, называемой изопедином, над которым представляет собой слой бугорков, обычно состоит из кости, как у Eusthenopteron . Слой дентина, который присутствовал в первой рыбе, закрепленной на доле, обычно уменьшается, как в существующем Колаканте или полностью отсутствуя, как у существующих легких и в Девонском Эустеноптероне . [ 12 ] Эламоидные весы появлялись несколько раз в течение эволюции рыбы. Они присутствуют у некоторых рыб с долиной , например, все сохранившиеся и некоторые вымершие рыбы в легких , а также в коэлакантах , которые имеют модифицированные космоидные масштабы, в которых отсутствуют космические и более тонкие, чем настоящие космоидные шкалы. Они также присутствуют в некоторых тетраподоморфах, таких как Eusthenopteron , Amiids и Teleost, чьи циклоидные и циклоидные шкалы представляют собой наименее минерализованные масштабы эламоида.
Шкалы эламоидов рыбок данио используются в лаборатории для изучения процесса минерализации кости и могут быть культивированы (сохранены) вне организма. [ 13 ] [ 14 ]
Ганоидные весы
[ редактировать ]

Ганоидные чешуйки обнаруживаются в осетрах , веслах , Gars , Bowfin и Bichirs . Они получены из космоидных масштабов и часто имеют зубчатые края. Они покрыты слоем твердого эмали-подобного дентина на месте космона , а также слоем неорганической костяной соли, называемого гануэном вместо витродленяна .
Ганойн является характерным компонентом ганоидных масштабов. Это стеклянная, часто многослойная минерализованная ткань , которая покрывает весы, а также черепные кости и ребятные лучи у некоторых неэлеостовых лучевых рыб , [ 15 ] такие как Gars , Bichirs и Coelacanths . [ 16 ] [ 17 ] Он состоит из стержня, подобных кристаллитам апатитов . [ 18 ] Ganoine является древней чертой рыб с лучами, которые находятся, например, в масштабах группы STEM Actinopteryigian cheiroleepis . [ 17 ] В то время как часто считается синапоморфным характером рыб с лучами, ганойновые или ганойноподобные ткани также обнаружены на вымершем акантодие . [ 17 ] Было предположительно, что ганойн гомологичен зубной эмале у позвоночных [ 15 ] или даже считается типом эмали. [ 18 ]
 Amblypterus striatus
|
Ганоидные чешуйки вымершей каменноугольной рыбы, Amblypterus striatus . (а) показывает внешнюю поверхность четырех шкал и (б) показывает внутреннюю поверхность двух шкал. Каждая из ганоидных масштабов Amblypterus в ромбоидальной форме имеет гребень на внутренней поверхности, который производится на одном конце в выступающий колышки, который вписывается в выемку в следующей масштабе, аналогично способу, в котором плитки скреплены на крыше дома. | 
|
Большинство ганоидных масштабов являются ромбоидальными (алмазными) и соединены суставами с колышками. Они обычно толстые и соединяются друг с другом, а не наносятся на кожу, как и другие масштабы. [ 19 ] Таким образом, ганоидные масштабы практически непроницаемы и являются отличной защитой от хищничества.
-
![Аллигатор GAR имеет жесткую громкоговорительную ганоидную масштаб в форме ромбоидальной формы. [19]](//upload.wikimedia.org/wikipedia/commons/thumb/b/bd/Alligator_gar_fish.jpg/395px-Alligator_gar_fish.jpg)
-
 Осетр раскачиваемые имеет ряды ганоидных масштабов, увеличенных в пластины брони.
Осетр раскачиваемые имеет ряды ганоидных масштабов, увеличенных в пластины брони. -
 Ганоидные масштабы на боковом сайте уменьшаются в размерах и напоминают циклоидные масштабы .
Ганоидные масштабы на боковом сайте уменьшаются в размерах и напоминают циклоидные масштабы .
![Аллигатор GAR имеет жесткую громкоговорительную ганоидную масштаб в форме ромбоидальной формы. [19]](http://upload.wikimedia.org/wikipedia/commons/thumb/b/bd/Alligator_gar_fish.jpg/395px-Alligator_gar_fish.jpg)



В осетрах масштабы значительно увеличены в пластины доспехов вдоль боковых и спины, в то время как в боковых шкалах масштабы значительно снижаются по толщине, чтобы напоминать циклоидные масштабы .
-
 Серьги, сделанные из ганоидных чешуйков аллигатора GAR
Серьги, сделанные из ганоидных чешуйков аллигатора GAR -
 Ископаемое примитивного радифина с ганоидными масштабами
Ископаемое примитивного радифина с ганоидными масштабами -




Коренные американцы и люди Карибского бассейна использовали жесткие ганоидные чешуйки аллигатора GAR для головок стрел, надолов и в качестве защиты для покрытия плугов. В текущее время ювелирные изделия изготавливаются из этих масштабов. [ 20 ]
Лептоидные масштабы
[ редактировать ]Шкалы лептоида (костного) обнаружены на костной рыбе высшего порядка, телеостах (более полученная клада рыб с лучами). Внешняя часть этих весов раздувается с костными хребтами, в то время как внутренняя часть пересекается с волокнистой соединительной тканью. Лептоидные масштабы более тонкие и полупрозрачные, чем другие типы масштабов, и не хватает закаленных эмалированных или дентиновых слоев. В отличие от ганоидных масштабов, в концентрических слоях добавляются дальнейшие масштабы по мере роста рыбы. [ 21 ]
Лептоидные масштабы перекрываются в конфигурации с головой на хвост, например, на крыше, что делает их более гибкими, чем космоидные и ганоидные масштабы. Такое расположение позволяет более плавный поток воды по телу и уменьшает сопротивление . [ 22 ] Масштабы некоторых видов демонстрируют полосы неравномерного сезонного роста, называемых аннули (единственное кольцо ). Эти полосы могут быть использованы, чтобы выдержать рыбу .
Лептоидные масштабы поставляются в двух формах: циклоид (гладкий) и Ctenoid (Combes, как). [ 23 ]
Циклоидные масштабы
[ редактировать ]Циклоидные (круглые) шкалы имеют гладкую текстуру и являются однородными, с гладким внешним краем или полем. Они наиболее распространены на рыбе с лучами мягкого плавника, такими как лосось и карп .

|

|
Азиатская Арована имеют большие циклоидные чешуйки, расположенные на рыбе в мозаике с поднятыми ребрами (слева). Сами шкалы покрыты тонким чистым рисунком (справа). [ 24 ] [ 25 ]
| |


Циклоидные (круглые) чешуйки обычно обнаруживаются на карпа-подобных или лососевом рыбах.
|





Ctenoid Scales
[ редактировать ]Ctenoid (зубчатые) чешуйки похожи на циклоидные чешуйки, за исключением того, что они имеют небольшие зубы или спиралы, называемые CTENII вдоль их внешних или задних краев. Из -за этих зубов весы имеют грубую текстуру. Они обычно встречаются на рыбах с лучами колючих плавников, таких как окутные рыбы. Эти весы почти не содержат кости, состоящая из поверхностного слоя, содержащего гидроксиапатит и карбонат кальция , и более глубокий слой, состоящий в основном из коллагена . Эмаль других типов шкалы сводится к поверхностным хребтам и CTENII.



|

|
Размер зубов на чешуйках Ctenoid может варьироваться в зависимости от положения, как эти масштабы от Cetonurus Cetonurus Crassiceps . показывают
| |
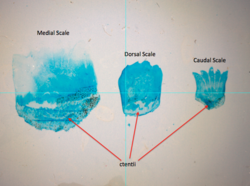

Ctenoid (зубчатые) чешуйки обычно встречаются на окутных рыбах.
|





Ctenoid Scales, аналогичные другими эпидермальными структурами, происходят из -за плакатов , а отличительная клеточная дифференцировка делает их исключительными из других структур, которые возникают из покровного положения . [ 27 ] Развитие начинается возле каудального плавника , вдоль боковой линии рыбы. [ 28 ] Процесс разработки начинается с накопления фибробластов между эпидермисом и дермой . [ 27 ] Коллагеновые фибриллы начинают организовывать себя в дермальном слое, что приводит к началу минерализации . [ 27 ] Окружность весов растет в первую очередь, после чего с последующей толщиной при перекрывании слоев минерализуется вместе. [ 27 ]
Шкалы Ctenoid могут быть дополнительно разделены на три типа:
- Cenate Scales, где рамка шкалы носит отступления и прогнозы.
- Спиноидные масштабы, где шкала несет шипы, которые непрерывны с самим шкалой.
- Истинные Ctenoid Scales, где шипы на шкале являются различными структурами.
Большинство рыб с лучами имеют чешуйчатые чешуйки. Некоторые виды столовых рыб имеют ценоидные масштабы на глазной стороне и циклоидные масштабы на слепой стороне, в то время как другие виды имеют масштаб Ctenoid у мужчин и циклоидные масштабы у женщин.
Отражение
[ редактировать ]


Многие телеострные рыбы покрыты высокими отражающими чешуйками, которые функционируют как маленькие зеркала и дают вид серебряного стекла. Отражение через серебросительство широко распространено или доминирует у рыб с открытым морем, особенно те, которые живут в первых 100 метрах. Эффект прозрачности может быть достигнут за счет серебряного серебра, чтобы сделать тело животного очень отражающим. На средней глубине в море свет наступает сверху, поэтому зеркало, ориентированное на вертикали, делает животных, таких как рыба невидимой со стороны. [ 29 ]
Морская рыба -топочка чрезвычайно уплощена в боковом направлении (от стороны в сторону), оставляя тело толщиной всего за миллиметры, а тело настолько серебристое, что напоминает алюминиевую фольгу . Зеркала состоят из микроскопических структур, аналогичных тем, которые использовались для обеспечения структурной окраски : стеки от 5 до 10 кристаллов гуанина , расположенных примерно на ¼ длина волны, чтобы конструктивно мешать и достигать почти 100 -процентного отражения. В глубоких водах, в которых живет рыба -топор, только синий свет с длиной волны 500 нанометров просачивается вниз и необходимо отражать, поэтому зеркала 125 нанометров отдельно обеспечивает хороший камуфляж. [ 29 ]
Большинство рыб в верхнем океане замаскированы серебром. У рыб, таких как сельдь , которая живет в более мелкой воде, зеркала должны отражать смесь длин волн, а рыба соответственно имеет кристаллические стеки с различными расстояниями. Еще одним осложнением для рыбы с телами, округленными по поперечному сечению, состоит в том, что зеркала будут неэффективными, если они положены на кожу, поскольку они не смогут отражать горизонтально. Общий зеркальный эффект достигается со многими небольшими отражателями, все ориентированные вертикально. [ 29 ]
Шкалы рыб с этими свойствами используются в некоторой косметике, поскольку они могут дать мерцающий эффект для макияжа и помады. [ 30 ]
Плакоидные весы
[ редактировать ]
Плокоидные (заостренные, зубные) чешуйки встречаются в хрящевых рыбах : акулы , лучи . Их также называют кожными зубцами . Шкалы плакоидов структурно гомологичны с позвоночных зубцами («зубца» транслируется как «маленький зуб»), имея центральную полость мякоти , поставляемая с кровеносными сосудами , окруженным коническим слоем дентина , все из которых сидит поверх прямоугольной базальной пластины опирается на дерму . Самый внешний слой состоит из витродулена , в значительной степени неорганической эмалевой вещество. Шкалы плакоидов не могут расти в размерах, а добавляется больше масштабов, когда рыба увеличивается в размере.
Подобные масштабы также можно найти под головой зубчатой сельдь . Количество покрытия масштаба гораздо меньше в лучах.
Предполагается, что ромбоидальные масштабы со свойствами как плакоидных, так и ганоидных масштабов существуют у современных предков рыб с челюстями: безжалостными остракодермами , а затем челюстными плакодермами .
Акула кожа
[ редактировать ]
Кожа акулы почти полностью покрыта небольшими плакоидными чешуйками. Масштабы поддерживаются шипами, которые чувствуют себя грубыми, когда поглаживаются в обратном направлении, но при сглаживании переднего движения воды создают крошечные вихри , которые уменьшают гидродинамические сопротивления и уменьшают турбулентность , что делает плавание более эффективным и более спокойным по сравнению с костями костей. Рыбы. [ 31 ] Это также выполняет роль в борьбе с планом, демонстрируя эффект лотоса . [ 32 ]
Все зубные зубцы состоят из внутренней полости мякоть с нервным и артериальным снабжением, укорененным в дерме для снабжения зубца слизью. [ 33 ] Зубцы содержат бурные структуры, которые выступают от поверхности шкалы; Под микроскопом этот бунт может выглядеть как крюк или хребты, выходящие из масштаба. Общая форма выпячивания от зубчатого зубца зависит от типа акулы и может быть обычно описана с двумя появлениями. [ 34 ] Первая - это шкала, в которой хребты расположены в боковом направлении вниз по акуле и параллельно потоку воды. Вторая форма представляет собой гладкую масштаб с тем, что выглядит как крючковая бунтика, сжащаяся с поверхности, направленной на заднюю сторону акулы. [ 34 ] Обе формы бульвара помогают в создании турбулентного пограничного слоя, заставляя ламинарный поток дальше от кожи Акул. [ 35 ]
В отличие от костной рыбы, у акул есть сложный кожный корсет, изготовленный из гибких коллагенных волокон, расположенных как спиральная сеть, окружающая их тело. Корсет работает как внешний скелет, обеспечивая привязанность к их плавающим мышцам и, таким образом, экономит энергию. [ 36 ] В зависимости от положения этих плакоидных масштабов на теле, они могут быть гибкими и могут быть пассивно установлены, что позволяет им изменить свой угол атаки. Эти масштабы также имеют буйки, которые выровнены в направлении потока, эти бугорты уменьшают силу сопротивления, действующую на кожу акулы, толкая вихрь дальше от поверхности кожи, ингибируя любой высокоскоростный поток поперечного потока. [ 37 ]
Морфология масштаба
[ редактировать ]Общая анатомия весов варьируется, но все они можно разделить на три части: корона, шея и основание. Масштабная гибкость связана с размером основания шкалы. Масштабы с более высокой гибкостью имеют меньшее основание и, таким образом, менее жестко прикреплены к Laxum Stratum. На короне быстро плавающих акул есть ряд параллельных бугортов или хребтов, которые проходят от переднего к заднему направлению. [ 38 ]
Анализируя три компонента шкалы. Можно сделать вывод, что основание зубца не вступает в контакт с какой -либо частью потока жидкости. [ 39 ] Корона и шея зубных зубьев, однако, играют ключевую роль и несут ответственность за создание турбулентных вихрей и вихрей, найденных возле поверхности кожи. [ 39 ] Поскольку зубные зубцы бывают разных форм и размеров, можно ожидать, что не все формы будут производить один и тот же тип турбулентного потока . Во время недавнего исследовательского эксперимента биомиметические образцы зубчатых зубьев акул с полумесяцей, подобной микроструктуре, были протестированы в резервуаре для воды с использованием таблицы тяги в качестве слайда. Эксперимент показал, что поверхность с зубцами испытывала 10% снижение сопротивления по сравнению с гладким образцом. Причиной этого сокращения сопротивления было то, что турбулентные вихри оказались в ловушке между зубцами, создавая «подушку, подобную» барьеры против ламинарного потока. [ 40 ] Этот же тип эксперимента был выполнен другой исследовательской группой, которая внедрила больше различий в их биомиметическом образце. Вторая группа пришла к тому же выводу, что и первый. Однако, поскольку их эксперимент содержал больше различий в образцах, они смогли достичь высокой степени экспериментальной точности. В заключение, они заявили, что более практические формы были более долговечны, чем те, у которых были замысловатые линии гребня. Практические формы были низкими и содержали трапециевидные или полукруглые поперечные сечения, похожие на проводку, и были менее эффективными, но, тем не менее, уменьшенным сопротивлением на 6 или 7%. [ 41 ]
Сокращение сопротивления
[ редактировать ]

Акулы уменьшают сопротивление и общую стоимость транспорта (COT) через несколько разных путей. Перетаскивание давления создается из разности давления между передней и задней стороны акулы из -за количества объема, который протолкнут мимо акулы, чтобы продвинуться вперед. [ 42 ] Этот тип сопротивления также прямо пропорционален ламинарному потоку . Когда ламинарный поток увеличивается вокруг рыбы, сопротивление давления тоже. [ 43 ] Трещное сопротивление является результатом взаимодействия между жидкостью против кожи акулы и может варьироваться в зависимости от того, как пограничный слой изменяется на поверхности рыбы. [ 42 ]
Риблы препятствуют трансляции поперечных потоков потоковых вихрей в вязком субалере. Механизм сложный и еще не понят полностью. По сути, буйки ингибируют формирование вихря вблизи поверхности, потому что вихрь не может вписаться в долины, образованные в бунтах. Это толкает вихрь дальше от поверхности, взаимодействуя только с кончиками бунтика, не вызывая какого-либо высокоскоростного потока в долинах. Поскольку этот высокоскоростный поток в настоящее время взаимодействует только с бугорным, который представляет собой очень небольшую площадь поверхности, перенос импульса, который вызывает сопротивление, теперь намного ниже, чем раньше, тем самым эффективно снижая сопротивление. Кроме того, это уменьшает колебания скорости поперечного потока, что также помогает при переносе импульса. [ 38 ]
Недавнее исследование показало, что в пограничном слое ближнехстен есть режим до и после перерыва, где подстелк сгущается с уменьшающейся скоростью, а затем внезапно подвергается распаду в турбулентные вихри, прежде чем, наконец, разрушится. Эта система полностью саморегулируется и опосредует цикл роста и распада; Вихри накапливаются в течение периода роста и внезапно ликвидируются в массивы вихрей шпильки, поднимающихся со стены. Подъем вихрей - это то, что выталкивает пограничный слой и от поверхности акулы, что приводит к уменьшению общего сопротивления, испытываемого рыбой. [ 44 ]
Техническая применение
[ редактировать ]Грубая наждачная бумага , подобная текстуре акулы и лучей, в сочетании с ее жесткостью, привела к тому, что она оценивается как источник кожи из сырья , называемого Шагрином . Одним из многих исторических применений Акулы Шагрин был вручную для рук для мечей . Грубая текстура кожи также используется в японской кухне, чтобы сделать товарные, называемые Орошики , прикрепляя кусочки кожи акулы к деревянным доскам. Небольшой размер весов очень тонко обогащает пищу.

В морской промышленности существует чрезвычайно большой рынок и необходимость в противоречивых поверхностях . С точки зрения мирян, загрязнение известно как процесс, посредством которого что -то становится инкрустированным материалом из окружающей среды, такой как саралы , водоросли и зеленый ил . ДАНТАЛЬНЫЕ ДАЛИКИ - чрезвычайно многообещающая область исследований для этого типа применения из -за того, что акулы являются одними из единственных рыб без наращивания или роста в их масштабах. Исследования ВМС США показали, что если биомиметический материал может быть спроектирован, он может потенциально привести к экономии затрат на топливо для военных судов до 45%. [ 45 ]
Есть много примеров биомиметических материалов и поверхностей, основанных на структуре водных организмов, включая акул. Такие приложения намереваются обеспечить более эффективное движение через жидкие среды, такие как воздух, вода и нефть.
Поверхности, которые имитируют кожу акул, также использовались, чтобы не допустить, чтобы микроорганизмы и водоросли покрывали корпус подводных лодок и кораблей. Один сорт торгуется как « Шарклет ». [ 46 ] [ 47 ]
Многие новые методы для воспроизведения кожи акулы включают использование полидиметилсилоксана (PDMS) для создания формы. Обычно процесс включает в себя съемку плоского куска кожи акулы, покрывая его PDM, чтобы сформировать плесень и снова залить PDM в эту форму, чтобы получить копию кожи акулы. Этот метод был использован для создания биомиметической поверхности, которая обладает сверхгидрофобными свойствами, демонстрируя эффект лотоса . [ 46 ] Одно исследование показало, что эти биомиметические поверхности снижают сопротивление до 9%, [ 37 ] в то время как с расплывчатым движением, снижение сопротивления достигло 12,3%. [ 48 ]
Данкулирование также обеспечивает снижение сопротивления на объектах, где основная форма сопротивления вызвана турбулентным потоком на поверхности. Большая часть общего сопротивления на длинных объектах с относительно плоскими сторонами обычно происходит от турбулентности у стены, поэтому бунтины будут иметь заметный эффект. Наряду с морскими применениями, аэрокосмическая промышленность может извлечь большую пользу от этих биомиметических дизайнов. Другие приложения включают трубы, где они набирают внутренности до шероховатости, похожей на бунтику и обнаружили 5% снижение сопротивления, а сокращение на несколько процентов претендует на конкурентоспособные купальники. [ 49 ]
Параметрическое моделирование было сделано на зубчатых зубцах акулы с широким спектром вариаций проектирования, таких как низкие и высокопрофильные вихревые генераторы. [ 50 ] С помощью этого метода была завершена наиболее тщательная характеристика для симметричных двухмерных буйтов с пилотушными, зубчатыми и поперечными сечениями лезвия. [ 49 ] Эти биомиметические модели были разработаны и проанализированы, чтобы увидеть эффекты применения зубчатых структур к крыльям различных самолетов. Во время симуляции было отмечено, что образец изменил то, как низкие и высокие углы атаки реагировали . Как геометрия зубья, так и их расположение оказывают глубокое влияние на аэродинамический отклик аэродинамической почты. Из протестированных как низких, так и из протестированных образцов генераторы с низким содержанием вихрей превзошли токовые гладкие структуры крыла на 323%. Это увеличение производительности связано с пузырьком разделения в вихрях зубного следа и потоком, которые пополняют импульс, потерянный в пограничном слое из-за трения кожи. [ 50 ]
Закуски
[ редактировать ]
Scutes аналогичны масштабам и выполняют ту же функцию. В отличие от весов рыбы, которые образуются из эпидермиса, в нижнем сосудистом слое кожи образуются заказки, а эпидермальный элемент - только верхняя поверхность. Формируя в живой дерме, изберы производят возбужденный внешний слой, который поверхностно похож на шкалы.
Scute происходит от латыни для щита и может принять форму:
- внешняя костная пластина, похожая на щит, или
- модифицированная, утолщенная шкала, которая часто является килевой или колючей, или
- Проецирующая, модифицированная (грубая и сильно рискую) шкалу, обычно связанную с латеральной линией, или на хвостовом цветоносе, образующем хвостовые кили или вдоль вентрального профиля.
Некоторые рыбы, такие как рыба -пейно -закуски , полностью или частично покрыты закусками. Речные сельды и нить с брюшной полостью сражаются, которые представляют собой масштаб с поднятыми, острыми точками, которые используются для защиты. У некоторых домкратов есть ряд задуц после боковой линии с обеих сторон.
Разработка масштаба
[ редактировать ]Весы обычно появляются в конце развития рыбы. В случае рыбок данио требуется 30 дней после оплодотворения, прежде чем различные слои, необходимые для начала формирования масштабов, дифференцировались и становятся организованными. Для этого необходимо, чтобы консолидация мезенхимы произошла , затем индуцируется морфогенез , и, наконец, происходит процесс дифференцировки или поздней метаморфозы . [ 51 ] [ 52 ]
- Мезенхима Консолидация: консолидация или структурирование мезенхимы возникают во время развития дермы . Этот процесс зависит от того, является ли рыба хрящевая или костная. Для хрящной рыбы структурирование происходит через образование двух слоев. Первый поверхностный и широкий, а второй тонкий и компактный. Эти два слоя разделены мезенхимальными клетками . Conhish Fish генерирует вицеллельный субстрат, организованный перпендикулярно коллагеновыми волокнами . Впоследствии для обеих рыб фибробласты удлиняются. Они проникают в компактный слой мезенхимы, который консолидируется до образования шкалы, чтобы инициировать дермальную пластину. [ 51 ] [ 52 ] [ 53 ]
- Индукция морфогенеза: морфогенез связан с образованием эпидермального сосования , которое генерируется путем соединения эпидермиса и дермы посредством процесса инвагинации . Морфогенез начинается в то время, когда фибробласты перемещаются в верхнюю часть компактной мезенхимы. На протяжении всего этого процесса базальные клетки эпителия образуют разделительный слой , который расположен в верхней части мезенхимы. Впоследствии эти клетки будут дифференцироваться в той области, где будет возникнуть масштабная зачатка . [ 51 ] [ 52 ] [ 53 ]
- Дифференциация или поздняя метаморфоза: эта дифференциация генерируется двумя разными формами в соответствии с типом формируемой шкалы. Образование эламоидных масштабов (циклоидов и Ctenoids) происходит посредством образования пространства между матрицей эпидермального сосования. Это пространство содержит коллагеновые волокна. Вокруг этого пространства эласмобласты дифференцируются и несут ответственность за создание необходимого материала для формирования шкалы. Впоследствии происходит минерализация матрицы , позволяя масштабе приобретать жесткую характеристику, которая их идентифицирует. [ 51 ] [ 52 ] [ 53 ]
В отличие от эламоидных масштабов, ганоидные масштабы состоят из минерализованного и неминерализованного коллагена в разных регионах. Образование их происходит посредством въезда поверхностных клеток мезенхимы в матрицу, последний состоит из коллагеновых волокон и расположена вокруг сосудистых капилляров, что приводит к возникновению сосудистых полостей. В этот момент элазобласты заменяются остеобластами , образуя тем самым кость. Пласти матрицы шкалы, которые не выявляются, состоят из компактного коллагена, которые позволяют ему поддерживать объединение с мезенхимой. Это известно как волокна Шарпи . [ 51 ] [ 52 ] [ 53 ]
Одним из генов, которые регулируют развитие формирования масштаба в Fish, является ген звукового хеджирования (SHH), который посредством белка (SHH), участвующего в органогенезе и в процессе клеточной связи , позволяет образовать шкалы. [ 54 ] [ 55 ] Аполипопротеин E (APOE), который позволяет транспортировать и метаболизм триглицеридов и холестерина , взаимодействует с SHH, потому что APOE обеспечивает холестерин для сигнального пути Shh . Было показано, что во время процесса клеток дифференцировки и взаимодействия уровень транскрипции APOE высок, что привело к выводу, что этот белок важен для поздних развития масштабов. [ 54 ] [ 55 ]
Модифицированные масштабы
[ редактировать ]



Различные группы рыбы развили ряд модифицированных шкал для выполнения различных функций.
- Почти все рыбы имеют боковую линию , систему механорецепторов , которые обнаруживают движения воды. У костных рыб весы вдоль боковой линии имеют центральные поры, которые позволяют воде контактировать с сенсорными клетками.
- Спинные плавники колючи о собачьих рыбах и чимаерах, жгучих хвостовых шипов скатрей и «пилы» зубов пили -рыб и пилледов сплавлены и модифицированными плакоидными масштабами.
- Хирург-рыба имеет скальпель-лезвие, которое представляет собой модифицированную шкалу по обе стороны от хвостового цветоноса . [ 56 ]
- Некоторые сельди , анчоусы и полубески имеют лиственные масштабы , которые легко проливают и помогают в уходе с хищниками.
- Самцы -пирожники перцины имеют ряд увеличенных кадукных масштабов между тазовыми плавниками и анусом .
- Рыбы дикобраза имеют масштабы, модифицированные на большие внешние шипы .
- Напротив, Pufferfish имеет более тонкие, более скрытые шипины, чем дикобраза, которые становятся видимыми только тогда, когда рыба вспыхнет. В отличие от рыб дикобраза, эти шипы не являются модифицированными масштабами, но развиваются под контролем той же сети генов, которые производят перья и волосы у других позвоночных. [ 57 ] [ 58 ]
-
 Рыба дикобраза имеет масштабы, модифицированные в шипы .
Рыба дикобраза имеет масштабы, модифицированные в шипы . -
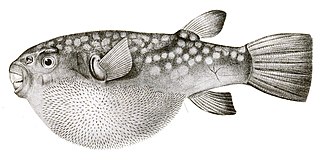 Колючицы Pufferfish не являются модифицированными масштабами, но разработаны независимой генной сетью.
Колючицы Pufferfish не являются модифицированными масштабами, но разработаны независимой генной сетью.


Рыба без чешуи
[ редактировать ]-
 Мандарин -рыба не хватает весов и защищает себя слоем вонючей и горькой слизи.
Мандарин -рыба не хватает весов и защищает себя слоем вонючей и горькой слизи.

Рыба без чешуйков обычно развивает альтернативы защитных чешуйке, например, жесткая кожаная кожа или костные тарелки.
- Без челюстная рыба ( миноги и мусорные рыбы ) имеют гладкую кожу без чешуйков и без кожной кости. [ 59 ] Миногды получают некоторую защиту от жесткой кожистой кожи. Hagfish излучает обильные количества слизи или слизи, если им угрожают. [ 60 ] Они могут завязать себя в узел из -за надруга , соскабливая слизь, когда они уходят и освобождая себя от хищника. [ 61 ]
- Большинство угрей являются без мельчайших, хотя некоторые виды покрыты крошечными гладкими циклоидными масштабами.
- У большинства умов не хватает весов, хотя в нескольких семействах есть бронежина в виде кожных пластин или какого -то раскатывания. [ 62 ]
- Мандариновая рыба не имеет масштабов и имеет слой вонючей и горькой слизи, который блокирует болезнь и, вероятно, препятствует хищникам, подразумевая, что их яркая окраска является апосематичной . [ 63 ]
- Рыба с рыбами имеет свободную тонкую кожу, часто покрытую тонкими раздвоенными кожными кольцами или бугорками , но у них нет обычных весов. Они полагаются на камуфляж, чтобы избежать внимания хищников, в то время как их свободная кожа затрудняет их схватить хищникам.
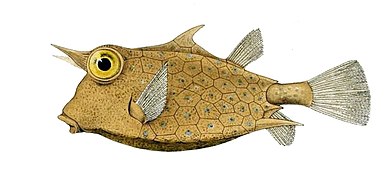
Многие группы костных рыб, в том числе трубные рыбы , морские коньки , коробку , браконьеры и несколько семейств стипку , разработали внешние костиные пластины, структурно напоминающие плакоидные чешуйки, как защитные доспехи против хищников.
- Морские коньки не имеют чешуйки, но у растягивалась тонкая кожа на косниционной броне, расположенной в кольцах по длине их тела.
- В Boxfish пластины объединяются, чтобы сформировать жесткую оболочку или экзоскелет, окружающую все тело. Эти костиные пластины не являются модифицированными масштабами, а кожей, которая была очерчена . Из -за этой тяжелой бронийской рыбы ограничены медленными движениями, но немногие другие рыбы могут есть взрослых.

| |
Угрят кажутся без мельки, но некоторые виды покрыты крошечными гладкими циклоидными масштабами.
| |
-
 В ящиках есть пластины с окостенеенной кожей, слитые вместе, чтобы сформировать жесткую оболочку.
В ящиках есть пластины с окостенеенной кожей, слитые вместе, чтобы сформировать жесткую оболочку. -
 Морские коньки имеют тонкую кожу, растянутую на костяных пластинах, расположенных в кольцах.
Морские коньки имеют тонкую кожу, растянутую на костяных пластинах, расположенных в кольцах.


Некоторая рыба, такая как Хоки и Рыба -Меч, рождаются с веслами, но проливают их по мере их роста.
Filefish имеет грубые непересекающиеся шкалы с небольшими шипами, поэтому они называются Filefish. Некоторые Filefish кажутся безыскальными, потому что их масштабы такие малы.
появляется заметное масштабирование На тунте только вдоль боковой линии и в корслете , защитная полоса утолщенных и увеличенных масштабов в области плеча. На большинстве их тела тунца есть настолько маленькие масштабы, что для повседневного осмотра они кажутся без мельки. [ 64 ]
-
 Некоторые Filefish кажутся безыскальными, потому что их масштабы такие малы.
Некоторые Filefish кажутся безыскальными, потому что их масштабы такие малы. -
 Для случайного обследования тунец кажется в значительной степени свободным от масштабов, но это не так.
Для случайного обследования тунец кажется в значительной степени свободным от масштабов, но это не так.


Левит
[ редактировать ]Отрывок в Левит заявляет, что «обо всех, кто находится в водах ... в морях, а в реках« те, у кого нет ни плавников, так и масштабов », - это мерзости для вас» и не может быть съедено. [ 65 ] Это устраняет всех водных беспозвоночных как мерзости и нечистых , а также любую рыбу, в которой отсутствуют весы (кажется, что рыба не хватает плавников).
Согласно удушью или божественным указам Торы и Талмуда , для того, чтобы рыба была объявлена кошерной , она должна иметь весы и плавники. [ 66 ] Определение «масштаба» отличается от определений, представленных в биологии, тем, что весы кошерной рыбы должны быть видны для глаз и могут быть легко удалены с кожи либо ручной, либо масштабным ножом. [ 66 ] Согласно Кошерному агентству по сертификации Православного союза , рыба является кошерной, если весы можно удалить, не разрывая кожу. [ 67 ] Таким образом, карп и лосось являются кошерными, в то время как акула, масштабы которых являются микроскопическими, осетр, чьишнуты не могут быть легко удалены без вырезания их из тела, не являются кошерными. Другие некошерные рыбы включают сома, угри, пресноводную треску , змеиные скумбрии и рыбу-пуховики. [ 66 ]
Лепидофагия
[ редактировать ]
Лепидофагия (древнегреческий для поедания в масштабе )-это специализированное поведение в кормлении у рыб, которое включает в себя питание чешуек другой рыбы. [ 69 ] Лепидофагия независимо развивалась по меньшей мере в пяти пресноводных семьях и семи морских семьях. [ 70 ]
Шкалы рыбы могут быть питательными, содержащие кожную часть и слой богатой белком слизи, помимо слоев кератина и эмали . Они являются богатым источником фосфата кальция . [ 70 ] Тем не менее, энергия, потраченная на удар, в сравнении с количеством весов, потребляемых за забастовку, ограничивает размер чешуе говорящей рыбы, и они обычно намного меньше их добычи . [ 70 ] Поведение питания в масштабе обычно развивается из -за отсутствия пищи и экстремальных условий окружающей среды. Питание весов и кожа, окружающая весы, обеспечивает богатые белком питательные вещества, которые могут быть недоступны в другом месте ниши. [ 71 ]
Рыбные челюсти обычно показывают двустороннюю симметрию . Исключение происходит с помощью цихлидного Perissodus Microlepis Cichlid Eating Cichlid Microlepis . Челюсти этой рыбы встречаются в двух различных морфологических формах. Одно морф заставляет свою челюсть скрученной влево, позволяя ему легко есть весы на правом фланге своей жертвы. У другого морфа есть челюсть, скрученная вправо, что облегчает употребление весов на левом фланге ее жертвы. Относительное изобилие двух морфов в популяциях регулируется частотно-зависимым отбором . [ 68 ] [ 72 ] [ 73 ]
Смотрите также
[ редактировать ]- Определение возраста в рыбе
- Окраска животных
- Отражатели животных
- Фотонные кристаллы
- Рептилий шкала
- Масштаб (зоология)
- Шкала доспехи
- Змеиные весы
- Urokotori - японский рыб
Ссылки
[ редактировать ]- ^ Шкала Etymonline . Получено 28 апреля 2019 года.
- ^ Mongera, A.; Nüsslein-Volhard, C. (2013). «Шкалы рыбы возникают из мезодермы» . Текущая биология . 23 (9): R338 - R339. doi : 10.1016/j.cub.2013.02.056 . PMID 23660349 .
- ^ Sharpe, PT (2001). "Развитие рыбы: сегодня волосы, зубы и чешуйки вчера?" Полем Текущая биология . 11 (18): R751 - R752. doi : 10.1016/s0960-9822 (01) 00438-9 . PMID 11566120 . S2CID 18868124 .
- ^ Turner, S.; Тарлинг, DH (1982). «Телодонт и другие распределения Агнатана в качестве тестов нижних палеозойских континентальных реконструкций». Палеогеография, палеоклиматология, палеоэкология . 39 (3–4): 295–311. Bibcode : 1982ppp .... 39..295t . doi : 10.1016/0031-0182 (82) 90027-x .
- ^ Märss, T. (2006). «Экзоскелетная ультраскуляция ранних позвоночных». Журнал палеонтологии позвоночных . 26 (2): 235–252. doi : 10.1671/0272-4634 (2006) 26 [235: euoev] 2.0.co; 2 . S2CID 85993241 .
- ^ Подпрыгнуть до: а беременный Janvier, Philippe (1998). «Ранние позвоночные и их существующие родственники». Ранние позвоночные . Издательство Оксфордского университета . С. 123–127. ISBN 978-0-19-854047-2 .
- ^ Подпрыгнуть до: а беременный Тернер С. (1991). «Монофилия и взаимосвязь Theelodonti». В мм Чанг; YH Liu; Гр Чжан (ред.). Ранние позвоночные и связанные с ними проблемы эволюционной биологии . Science Press, Пекин. С. 87–119.
- ^ Märss, T. (1986). «Сквамация TheLodont Agnathan Phlebolepis ». Журнал палеонтологии позвоночных . 6 (1): 1–11. doi : 10.1080/02724634.1986.10011593 .
- ^ Botella, H.; Джи Валенсуэла-Риос; П. Карлс (2006). «Новый ранний Девонский Телодонт из Селтиберии (Испания), с пересмотром испанских Телодонтов» . Палеонтология . 49 (1): 141–154. doi : 10.1111/j.1475-4983.2005.00534.x . S2CID 128939911 .
- ^ Ferrón, Humberto G.; Botella, Héctor (2017). и экология Сквалог Plos один 12 (2): E0 Bibcode : 2017plos . doi : 13.1371/Журнал . PMC 53328365 . PMID 28241029 .
- ^ Майкл Аллаби "Космоидная шкала". Словарь зоологии. Полем Encyclopedia.com. 29 октября 2019 г. < https://www.encyclopedia.com >
- ^ Zylberberg, L., Meunier, FJ, Laurin, M. (2010). Микроанатомическое и гистологическое исследование посткраниального дермального скелета в девонском саркоптеригии эустеноптероне Фоорди , Acta Palaeontologica Polonica 55: 459–470.
- ^ Берген, Дилан Дж. М; Кагу, Эрика; Хаммонд, Крисси Л. (2019). «Рвиграфы как появляющаяся модель остеопороза: первичная платформа тестирования для скрининга новых осте-активных соединений» . Границы в эндокринологии . 10 : 6. doi : 10.3389/fendo.2019.00006 . ISSN 1664-2392 . PMC 6361756 . PMID 30761080 .
- ^ de Vrieze, E.; Ван Кессель, Махдж; Петерс, HM; Охваты, толстые; Flik, G.; Метц, младший (1 февраля 2014 г.). «Преднизолон индуцирует остеопороз-подобный фенотип в регенерирующих масштабах рыбок данио». Остеопороз International . 25 (2): 567–578. doi : 10.1007/s00198-013-2441-3 . ISSN 1433-2965 . PMID 23903952 . S2CID 21829206 .
- ^ Подпрыгнуть до: а беременный Zylberberg, L.; Sire, J. -y.; Нанчи А. (1997). «Иммуноректирование амелогениноподобных белков в гануине экспериментально регенерирующих масштабов каламоихтис Калабарикус, примитивной актиноптерийской рыбы» . Анатомическая запись . 249 (1): 86–95. doi : 10.1002/(SICI) 1097-0185 (199709) 249: 1 <86 :: AID-AR11> 3.0.CO; 2-X . PMID 9294653 .
- ^ Сир, Жан-Ив; Donoghue, Philip CJ; Vickaryous, Matthews K. (2009). «Происхождение и эволюция покровного скелета у неретраподных позвоночных» . Журнал анатомии . 214 (4): 409–440. doi : 10.1111/j.1469-7580.2009.01046.x . ISSN 0021-8782 . PMC 2736117 . PMID 19422423 .
- ^ Подпрыгнуть до: а беременный в Рихтер М. (1995). «Микроструктурное исследование ганойновой ткани выбранных нижних позвоночных». Зоологический журнал Линневого общества . 114 (2): 173–212. doi : 10.1006/zjls.1995.0023 .
- ^ Подпрыгнуть до: а беременный Брю, BJF; Song, J.; Бойс, MC; Ортис, С. (2008). «Принципы дизайна материалов древней рыбной брони». Природные материалы . 7 (9): 748–756. Bibcode : 2008natma ... 7..748b . doi : 10.1038/nmat2231 . PMID 18660814 .
- ^ Подпрыгнуть до: а беременный Шерман, Винсент Р.; Яраги, Николас А.; Кисайлс, Дэвид; Мейерс, Марк А. (1 декабря 2016 г.). «Микроструктурные и геометрические влияния в защитных масштабах Atractosteus spula» . Журнал интерфейса Королевского общества . 13 (125): 20160595. DOI : 10.1098/RSIF.2016.0595 . ISSN 1742-5689 . PMC 5221522 . PMID 27974575 .
- ^ «Миссури Аллигатор Гар Управление и План восстановления» (PDF) . Отдел Миссури Департамента охраны природы. 22 января 2013 года. Архивировано из оригинала (PDF) 6 мая 2016 года . Получено 12 апреля 2019 года .
- ^ Lagler, KF, Je Bardach и RR Miller (1962) Ichthyology . Нью -Йорк: Джон Уайли и сыновья.
- ^ Баллард, Бонни; Чик, Райан (2 июля 2016 г.). Экзотическое лекарство для животных для ветеринарного техника . Джон Уайли и сыновья. ISBN 978-1-118-92421-1 .
- ^ McGrouther, Марк (2 декабря 2019 г.). «Циклоидные и этеноидные масштабы» . Австралийский музей . Получено 29 декабря 2021 года .
- ^ Pouyaud, L.; Sudarto, Guy G. Teugels (2003). «Различные цвета цветовых сортов азиатских склеропаж арованы (Osteoglossidae) являются отдельными видами: морфологические и генетические доказательства». Кибий . 27 (4): 287–305.
- ^ Исмаил М. (1989). Систематика, зоогеография и сохранение пресноводных рыб на полуостровах Малайзии (докторская диссертация изд.). Университет штата Колорадо.
- ^ EJ Brill (1953). Рыбы индо-австралийского архипелага . EJ Brill. С. 306–307.
- ^ Подпрыгнуть до: а беременный в дюймовый Кавасаки, Кента С., «Генетический анализ морфологии шкалы цихлид» (2016). Мастерские тезисы, май 2014 - текущий. 425. http://scholarworks.umass.edu/masters_theses_2/425
- ^ Helfman, Gene (2009). Разнообразие биологии, эволюции и экологии рыб . Wiley-Blackwell.
- ^ Подпрыгнуть до: а беременный в Херринг, Питер (2002). Биология глубокого океана . Оксфорд: издательство Оксфордского университета. С. 193–195. ISBN 9780198549567 .
- ^ «Вероятно, в вашей помаде есть рыбные весы» . Huffpost India . 23 апреля 2015 года . Получено 6 мая 2019 года .
- ^ Мартин, Р. Эйдан. "Кожа зубов" . Получено 28 августа 2007 года .
- ^ Fürstner, Reiner; Бартлот, Вильгельм; Неййнхуис, Кристоф; Вальзель, Питер (1 февраля 2005 г.). «Смачивающиеся и самоочищающиеся свойства искусственных супергидрофобных поверхностей». Langmuir . 21 (3): 956–961. doi : 10.1021/la0401011 . ISSN 0743-7463 . PMID 15667174 .
- ^ Лодер, Джордж В.; Wainwright, Dylan K.; Домель, август Г.; Уивер, Джеймс С.; Вэнь, Ли; Бертольди, Катия (2016). «Структура, биомиметика и жидкая динамика поверхностей кожи рыб» . Физические обзоры жидкости . 1 (6): 060502. BIBCODE : 2016 PHRVF ... 1F0502L . doi : 10.1103/physrevfluids.1.060502 . S2CID 18118663 .
- ^ Подпрыгнуть до: а беременный Фелд, Катрин; Колборг, Энн Ноер; Ниборг, Камилла Мари; Салевски, Мирко; Стеффенсен, Джон Фленг; Берг-Сёренсен, Кирстин (24 мая 2019 г.). «Кожные зубные зубцы трех медленно плавающих видов акул: микроскопия и визуализация потока» . Биомиметика . 4 (2): 38. doi : 10.3390/biomimetics4020038 . ISSN 2313-7673 . PMC 6631580 . PMID 31137624 .
- ^ Флетчер, Томас; Алтрингем, Джон; Пикл, Джеффри; Вигнал, Пол; Доррелл, Роберт (7 августа 2014 г.). «Гидродинамика ископаемых рыб» . Труды Королевского общества B: Биологические науки . 281 (1788): 20140703. DOI : 10.1098/rspb.2014.0703 . ISSN 0962-8452 . PMC 4083790 . PMID 24943377 .
- ^ Мартин, Р. Эйдан. «Важность быть хрящевым» . Центр REEFQUEST для исследований акул . Получено 29 августа 2009 года .
- ^ Подпрыгнуть до: а беременный Hage, W.; Bruse, M.; Бехерт, DW (1 мая 2000 г.). «Эксперименты с трехмерными буйками как идеализированная модель кожи акулы». Эксперименты в жидкостях . 28 (5): 403–412. Bibcode : 2000exfl ... 28..403b . doi : 10.1007/s003480050400 . ISSN 1432-1114 . S2CID 122574419 .
- ^ Подпрыгнуть до: а беременный Мотта, Филипп; Хабеггер, Мария Лаура; Ланг, Эми; Хьютер, Роберт; Дэвис, Джессика (1 октября 2012 г.). «Морфология масштаба и гибкость в короткометражном мако -иурусе oxyrinchus и Blacktip Shark Carcharhinus limbatus». Журнал морфологии . 273 (10): 1096–1110. doi : 10.1002/jmor.20047 . ISSN 1097-4687 . PMID 22730019 . S2CID 23881820 .
- ^ Подпрыгнуть до: а беременный Доу, Чжаолиан; Ван, Цзядао; Чен, Даронг (1 декабря 2012 г.). «Бионические исследования по шкалам рыб для снижения сопротивления» . Журнал Bionic Engineering . 9 (4): 457–464. doi : 10.1016/s1672-6529 (11) 60140-6 . ISSN 1672-6529 . S2CID 137143652 .
- ^ «Экспериментальные исследования по характеристикам снижения сопротивления бионической поверхности с помощью заглощения воды микроструктуры масштабов рыб» (PDF) .
- ^ Палмер, Колин; Янг, Марк Т. (14 января 2015 г.). «Снижение поверхностного сопротивления и контроль разделения потока у пелагических позвоночных, что имеет значение для интерпретации морфологий шкалы в ископаемых таксонах» . Королевское общество открыто наука . 2 (1): 140163. Bibcode : 2015rsos .... 240163p . doi : 10.1098/rsos.140163 . ISSN 2054-5703 . PMC 4448786 . PMID 26064576 .
- ^ Подпрыгнуть до: а беременный Лодер, Джордж В.; Wainwright, Dylan K.; Домель, август Г.; Уивер, Джеймс С.; Вэнь, Ли; Бертольди, Катия (18 октября 2016 года). «Структура, биомиметика и жидкая динамика поверхностей кожи рыб» . Физические обзоры жидкости . 1 (6): 060502. BIBCODE : 2016 PHRVF ... 1F0502L . doi : 10.1103/physrevfluids.1.060502 .
- ^ Мутурамалингам, Мутукумар; Villemin, Leo S.; Брюкер, Кристоф (29 апреля 2019 г.). «Образование полос в потоке по биомиметическим массивам шкалы рыб». Журнал экспериментальной биологии . 222 (Pt 16): Jeb205963. Arxiv : 1904.12752 . BIBCODE : 2019ARXIV190412752M . doi : 10.1242/jeb.205963 . PMID 31375542 . S2CID 139103148 .
- ^ Bandyopadhyay, Promode R.; Хеллум, Арен М. (23 октября 2014 г.). «Моделирование того, как узоры кожи акулы и дельфинов контролируют паттерны завихренности переходной стены с использованием механизмов сброса пространственно-временной фазы» . Научные отчеты . 4 : 6650. Bibcode : 2014natsr ... 4e6650b . doi : 10.1038/srep06650 . ISSN 2045-2322 . PMC 4206846 . PMID 25338940 .
- ^ Магин, Челси М.; Купер, Скотт П.; Бреннан, Энтони Б. (1 апреля 2010 г.). «Нетоксичные стратегии антиобрабатывания» . Материалы сегодня . 13 (4): 36–44. doi : 10.1016/s1369-7021 (10) 70058-4 . ISSN 1369-7021 .
- ^ Подпрыгнуть до: а беременный Лю, Юнхон; Ли, Гуанджи (15 декабря 2012 г.). «Новый метод создания« эффекта лотоса »на коже биомиметической акулы». Журнал коллоидной и интерфейсной науки . 388 (1): 235–242. Bibcode : 2012jcis..388..235L . doi : 10.1016/j.jcis.2012.08.033 . ISSN 0021-9797 . PMID 22995249 .
- ^ «Sharklet Discovery | Sharklet Technologies, Inc» . www.sharklet.com . Получено 26 сентября 2018 года .
- ^ Лодер, Джордж В.; Оффнер, Йоханнес (1 марта 2012 г.). «Гидродинамическая функция кожи акулы и двух биомиметических применений» . Журнал экспериментальной биологии . 215 (5): 785–795. doi : 10.1242/jeb.063040 . ISSN 1477-9145 . PMID 22323201 .
- ^ Подпрыгнуть до: а беременный Дин, Брайан и Бхушан, Бхарат. (2010). Споры акулы для снижения турбулентного потока в турбулентном потоке: обзор. Философские транзакции. Серия A, математические, физические и инженерные науки. 368. 4775-806. 10.1098/rsta.2010.0201.
- ^ Подпрыгнуть до: а беременный Домель, август Г.; Саадат, Мехди; Уивер, Джеймс С.; Хадж-Харири, Хоссейн; Бертольди, Катия; Лодер, Джордж В. (28 февраля 2018 г.). «Проекты, вдохновленные кожей акулы, которые улучшают аэродинамические характеристики» . Журнал интерфейса Королевского общества . 15 (139): 20170828. DOI : 10.1098/rsif.2017.0828 . PMC 5832729 . PMID 29436512 .
- ^ Подпрыгнуть до: а беременный в дюймовый и Сир, JY; Huysseune, Ann (2003). «Образование кожных скелетных и зубных тканей в рыбе: сравнительный и эволюционный подход». Биологические обзоры . 78 (2): 219–249. doi : 10.1017/s1464793102006073 . PMID 12803422 . S2CID 19556201 .
- ^ Подпрыгнуть до: а беременный в дюймовый и Le Guellec, D.; Morvan-Dubois, G.; Sire, JY (2004). «Развитие кожи у конитовой рыбы с особым акцентом на осаждение коллагена в дерме рыбок данио ( Данио Ририо )» . Международный журнал биологии развития . 48 (2–3): 217–231. doi : 10.1387/ijdb.15272388 . PMID 15272388 .
- ^ Подпрыгнуть до: а беременный в дюймовый Sire, JY (2001). «Зубы за ртом у телеострных рыб: как извлечь выгоду из несчастного случая в развитии». Эволюция и развитие . 3 (2): 104–108. doi : 10.1046/j.1525-142x.2001.003002104.x . PMID 11341672 . S2CID 13353402 .
- ^ Подпрыгнуть до: а беременный Сир, JY; Akimenko, MA (2003). «Развитие масштаба в рыбе: обзор с описанием выражения Sonic Hedgehog (SHH) у рыбок данио ( Danio Rerio )» . Международный журнал биологии развития . 48 (2–3): 233–247. doi : 10.1387/ijdb.15272389 . PMID 15272389 .
- ^ Подпрыгнуть до: а беременный Monnot, MJ; Бабин, PJ; Poleo, G.; Андре, М.; LaForest, L.; Ballagny, C.; Akimenko, MA (1999). «Эпидермальная экспрессия гена аполипопротеина Е во время развития плавника и масштаба и регенерации плавников у рыбок данио» . Динамика развития . 214 (3): 207–215. doi : 10.1002/(SICI) 1097-0177 (199903) 214: 3 <207 :: AID-AJA4> 3.0.CO; 2-5 . PMID 10090147 .
- ^ Соренсон, Л.; Сантини, Ф.; Carnevale, G.; Альфаро, я (2013). «Многолокусная срока хирурга (Acanthuridae, Percomorpha), с пересмотренной семейной таксономией». Молекулярная филогенетика и эволюция . 68 (1): 150–160. doi : 10.1016/j.ympev.2013.03.014 . PMID 23542000 .
- ^ Как Pufferfish получил свои дурацкие шипы Phys.org , 25 июля 2019 года.
- ^ Shono, T.; Tiery, ap; Купер, RL; Kurokawa, D.; Бритц, Р.; Okabe, M.; Фрейзер, GJ (2019). «Эволюция и разнообразие развития кожных шипов в рыбах» . ISCAING . 19 : 1248–1259. Bibcode : 2019isci ... 19.1248s . doi : 10.1016/j.isci.2019.06.003 . PMC 6831732 . PMID 31353167 .
- ^ Coolidge E, Hedrick MS и Milsom WK (2011) «Вентиляционные системы» . В: McKenzie DJ, Farrell AP и Brauner CJ (Eds) Физиология рыбы: примитивные рыбы , Elsevier, стр. 182–213. ISBN 9780080549521
- ^ Ротшильд, Анна (1 апреля 2013 г.). "Hagfish Slime: одежда будущего?" Полем BBC News . Получено 2 апреля 2013 года .
- ^ Йонг, изд (23 января 2019 г.). «Никто не готовится к слизилому мусора» . Атлантика . Получено 26 января 2019 года .
- ^ Фрил, JP; Lundberg, JG (1996). « Микромизон Акамай , генерал. Et sp. Nov., Маленький и без глазного сома банджо (Siluriformes: Aspredinidae) из речных каналов нижнего бассейна Амазонки». Копея . 1996 (3): 641–648. doi : 10.2307/1447528 . JSTOR 1447528 .
- ^ Sadovy, Y.; Рэндалл, JE; Расотто, Мария Б. (май 2005 г.). «Структура кожи у шести видов Dragonet (Gobiesociformes; Callionymidae): межвидовые различия в типах железичных клеток и секреции слизи». Журнал рыбной биологии . 66 (5): 1411–1418. doi : 10.1111/j.0022-1112.2005.00692.x .
- ^ Есть ли тунцы весы? Научный центр северо -востока рыболовства, NOAA Fisheries. Доступ 4 августа 2019 года.
- ^ Левит 11: 9–10
- ^ Подпрыгнуть до: а беременный в Ари Цитрон, "Все о кошерной рыбе"
- ^ Проверка кошерной рыбы OU Kosher Certification . Получено 9 августа 2019 года.
- ^ Подпрыгнуть до: а беременный Ли, HJ; Kusche, H.; Мейер, А. (2012). «Рукобавление по поведению кормления в цихлидных рыбах, питающейся масштабам: его потенциальная роль в формировании морфологической асимметрии» . Plos один . 7 (9): E44670. Bibcode : 2012ploso ... 744670L . doi : 10.1371/journal.pone.0044670 . PMC 3435272 . PMID 22970282 .
- ^ Фрозе, Р. и Д. Поли. Редакторы. «Глоссарий: чешуе говорящая» . Рыбная база . Получено 12 апреля 2007 года .
{{cite web}}:|author=имеет общее имя ( справка ) - ^ Подпрыгнуть до: а беременный в Джановец, Джефф (2005). «Функциональная морфология кормления в специализированном катоприоне , специализирующемся на шкале » (PDF) . Журнал экспериментальной биологии . 208 (Pt 24): 4757–4768. doi : 10.1242/jeb.01938 . PMID 16326957 . S2CID 15566769 .
- ^ Мартин, C.; ПК Wainwright (2011). «Трофическая новинка связана с исключительными показателями морфологической диверсификации в двух адаптивных излучениях кипринодонской куколки» . Эволюция 65 (8): 2197–2212. doi : 10.1111/j.1558-5646.2011.01294.x . PMID 21790569 . S2CID 23695342 .
- ^ Хори, М. (1993). «Частотно-зависимый естественный отбор в руке цихлидной рыбы, питающейся шкалой». Наука . 260 (5105): 216–219. Bibcode : 1993sci ... 260..216h . doi : 10.1126/science.260.5105.216 . PMID 17807183 . S2CID 33113282 .
- ^ Стюарт, Та; Albertson, RC (2010). «Эволюция уникального аппарата хищного кормления: функциональная анатомия, развитие и генетический локус для латеральности челюсти в цихлидах, питающихся шкалой, по шкале, питающихся на озере,» . BMC Biology . 8 (1): 8. doi : 10.1186/1741-7007-8-8 . PMC 2828976 . PMID 20102595 .
Дальнейшее чтение
[ редактировать ]- Helfman, GS, BB Collette and De Facey (1997). Разнообразие рыб . Blackwell Science. С. 33–36. ISBN 978-0-86542-256-8 .
{{cite book}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - Schultze, HP (2016). «Весы, эмаль, космин, ганойн и ранние остеихтианцы» . Comptes rendus palevol . 15 (1–2): 83–102. doi : 10.1016/j.crpv.2015.04.001 .

Внешние ссылки
[ редактировать ]| Базы данных управления авторитетом : национальный |
|---|