Эндонуклеаза
В молекулярной биологии эндонуклеазы — это ферменты , расщепляющие фосфодиэфирную связь внутри полинуклеотидной цепи (а именно ДНК или РНК ). Некоторые из них, такие как дезоксирибонуклеаза I , разрезают ДНК относительно неспецифично (независимо от последовательности), в то время как многие, обычно называемые эндонуклеазами рестрикции или ферментами рестрикции , расщепляют только очень специфические нуклеотидные последовательности. Эндонуклеазы отличаются от экзонуклеаз , которые расщепляют концы последовательностей распознавания вместо средней ( эндо ) части. Однако некоторые ферменты, известные как « экзо-эндонуклеазы », не ограничиваются ни одной из нуклеазных функций, проявляя как эндо-, так и экзо-подобные качества. [ 1 ] Данные свидетельствуют о том, что активность эндонуклеазы отстает от активности экзонуклеазы. [ 2 ]
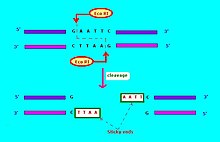
Ферменты рестрикции — это эндонуклеазы эубактерий и архей , которые распознают специфическую последовательность ДНК. [ 3 ] Нуклеотидная последовательность, распознаваемая для расщепления ферментом рестрикции, называется сайтом рестрикции . Обычно сайт рестрикции представляет собой палиндромную последовательность длиной около четырех-шести нуклеотидов. Большинство эндонуклеаз рестрикции расщепляют цепь ДНК неравномерно, оставляя комплементарные одноцепочечные концы. Эти концы могут повторно соединяться посредством гибридизации и называются «липкими концами». После спаривания фосфодиэфирные связи фрагментов могут быть соединены ДНК-лигазой . Известны сотни эндонуклеаз рестрикции, каждая из которых атакует отдельный сайт рестрикции. Фрагменты ДНК, расщепленные одной и той же эндонуклеазой, могут соединяться независимо от происхождения ДНК. Такая ДНК называется рекомбинантной ДНК ; ДНК образуется в результате объединения генов в новые комбинации. [ 4 ] Эндонуклеазы рестрикции ( ферменты рестрикции ) в зависимости от механизма действия делятся на три категории: тип I, тип II и тип III. Эти ферменты часто используются в генной инженерии для создания рекомбинантной ДНК для введения в клетки бактерий, растений или животных, а также в синтетической биологии . [ 5 ] Одна из наиболее известных эндонуклеаз — Cas9 .
Категории
[ редактировать ]В конечном счете, существует три категории эндонуклеаз рестрикции , которые вносят относительный вклад в расщепление специфических последовательностей. Типы I и III представляют собой большие мультисубъединичные комплексы, которые включают в себя как эндонуклеазную , так и метилазную активности. Тип I может расщеплять случайные сайты длиной около 1000 пар оснований или более из последовательности распознавания и требует АТФ в качестве источника энергии. Тип II ведет себя несколько иначе и был впервые выделен Гамильтоном Смитом в 1970 году. Они представляют собой более простые версии эндонуклеаз и не требуют АТФ в процессах деградации. Некоторые примеры эндонуклеаз рестрикции типа II включают Bam HI, Eco RI, Eco RV, HindIII и Hae III. Тип III, однако, расщепляет ДНК примерно по 25 парам оснований от последовательности узнавания, а также требует в этом процессе АТФ. [ 4 ]
Обозначения
[ редактировать ]Обычно используемые обозначения для эндонуклеаз рестрикции [ 6 ] имеет форму « Vwx yZ», где « Vwx » — курсивом первая буква рода и первые две буквы вида, в котором может быть обнаружена эта эндонуклеаза рестрикции, например, Escherichia coli , Eco и Haemophilus. грипп , Хин . За ним следует необязательный символ «y», не выделенный курсивом, который указывает тип или идентификацию штамма, например Eco R для штаммов E. coli , несущих фактор передачи лекарственной устойчивости RTF-1. [ 6 ] Eco B для штамма E. coli B, [ 7 ] и Hind для H. influenzae штамма d . [ 6 ] Наконец, когда конкретный тип или штамм имеет несколько различных эндонуклеаз рестрикции, они обозначаются римскими цифрами, поэтому эндонуклеазы рестрикции из штамма d H. influenzae называются Hin dI, Hin dII, Hind III и т. д. Другой пример: « Hae II» и « Hae III» относятся к бактериям Haemophilus aegyptius (штамм не указан), эндонуклеазы рестрикции номер II и номер III соответственно. [ 4 ] : 64–64 Ферменты рестрикции, используемые в молекулярной биологии, обычно распознают короткие целевые последовательности длиной около 4–8 пар оснований. Например, фермент EcoRI распознает и расщепляет последовательность 5' – GAATTC – 3'. [ 8 ]
Эндонуклеазы рестрикции бывают нескольких типов. Эндонуклеаза рестрикции обычно требует сайта узнавания и схемы расщепления (обычно нуклеотидных оснований: A, C, G, T). Если сайт узнавания находится за пределами области рисунка расщепления, то эндонуклеаза рестрикции относится к типу I. Если последовательность узнавания перекрывается с последовательностью расщепления, то эндонуклеаза рестрикции относится к типу II. [ нужна ссылка ]
Процессы, связанные с эндонуклеазами
[ редактировать ]Эндонуклеазы играют роль во многих аспектах биологической жизни. Ниже приведены несколько примеров процессов, в которых эндонуклеазы играют решающую роль.
восстановление ДНК
[ редактировать ]Эндонуклеазы играют роль в репарации ДНК. AP-эндонуклеаза , в частности, катализирует разрез ДНК исключительно в AP-сайтах и, следовательно, подготавливает ДНК для последующего вырезания, репарационного синтеза и лигирования ДНК. Например, когда происходит депуринация, в результате этого поражения остается сахар дезоксирибоза с отсутствующим основанием. [ 9 ] Эндонуклеаза AP распознает этот сахар и по существу разрезает ДНК в этом сайте, а затем позволяет продолжить восстановление ДНК. [ 10 ] Клетки E. coli содержат две AP-эндонуклеазы: эндонуклеазу IV (endoIV) и экзонуклеазу III (exoIII), тогда как у эукариот имеется только одна AP-эндонуклеаза. [ 11 ]

восстановление перекрестных связей ДНК
[ редактировать ]Восстановление ДНК , в котором две комплементарные цепи соединены межцепочечной ковалентной сшивкой, требует множественных разрезов, чтобы разъединить цепи и устранить повреждение. Надрезы необходимы с обеих сторон сшивки и на обеих цепях дуплексной ДНК. В эмбриональных стволовых клетках мыши промежуточная стадия репарации поперечных связей включает образование двухцепочечных разрывов. [ 12 ] MUS81 / EME1 представляет собой структурно-специфическую эндонуклеазу, участвующую в преобразовании межцепочечных поперечных связей в двухцепочечные разрывы зависимым от репликации способом. [ 12 ] После введения двухцепочечного разрыва необходимы дальнейшие шаги для завершения процесса восстановления. Если сшивка не восстановлена должным образом, она может заблокировать репликацию ДНК . [ нужна ссылка ]
Восстановление димера тимина
[ редактировать ]Воздействие бактериофага (фага) Т4 облучением ультрафиолетовым индуцирует появление димеров тимина в ДНК фага. фага Т4 Ген denV кодирует эндонуклеазу V , которая катализирует начальные этапы восстановления этих димеров тимина, индуцированных УФ-излучением. [ 13 ] Эндонуклеаза V сначала расщепляет гликозиловую связь на 5'-стороне пиримидинового димера, а затем катализирует расщепление фосподиэфирной связи ДНК, которая первоначально связывала два нуклеотида димера. Последующие этапы процесса репарации включают удаление остатков димера и синтез репарации для заполнения образовавшегося одноцепочечного разрыва с использованием неповрежденной цепи в качестве матрицы. [ нужна ссылка ]
Апоптоз
[ редактировать ]Во время апоптоза апоптотическая эндонуклеаза DFF40 активируется, чтобы инициировать контролируемую клеточную разборку. Этот распад характеризуется расщеплением геномной ДНК на специфические фрагменты. Точная роль эндонуклеаз в этом контексте заключается в расщеплении ДНК в определенных сайтах, генерируя фрагменты определенной длины. Эти фрагменты затем упаковываются в апоптотические тельца, обеспечивая аккуратное и эффективное удаление умирающей клетки, не вызывая воспаления или повреждения соседних клеток. [ 14 ]
Репликация ДНК
[ редактировать ]Эндонуклеаза лоскута 1 (FEN1) и эндонуклеаза Dna2 являются неотъемлемой частью репликации ДНК на отстающей цепи, участвуя в таких важных процессах, как удаление праймера и обработка фрагмента Оказаки . Эндонуклеазы активно участвуют в процессинге этих фрагментов, расщепляя фосфодиэфирные связи между ними. Этот процесс является неотъемлемой частью плавного синтеза и соединения фрагментов Окадзаки, способствуя общей непрерывности вновь реплицированной цепи ДНК. [ 15 ] [ 16 ]
Процессинг РНК
[ редактировать ]Эндонуклеазы, точнее эндорибонуклеаза , играют решающую роль в процессинге РНК, фундаментальном этапе экспрессии генов. Этот процесс включает в себя точное расщепление молекул РНК-предшественников под руководством эндонуклеаз с образованием функциональных РНК, необходимых для различных клеточных функций. Эндонуклеазы избирательно расщепляют РНК-предшественники в определенных сайтах, определяя границы функциональных сегментов РНК во время процессинга РНК. Результатом процессинга РНК является образование функциональных молекул РНК, таких как транспортные РНК (тРНК) и рибосомальные РНК (рРНК) . Эндонуклеазы способствуют точности этого процесса, обеспечивая образование зрелых и функциональных видов РНК.
Эндонуклеазы, такие как РНКаза P и тРНКаза Z (ELAC2), превращают тРНК-предшественники в зрелые функциональные тРНК, что имеет решающее значение для точной трансляции во время синтеза белка. [ 17 ] В биогенезе рибосом эндонуклеазы из семейства РНКаз III , такие как DROSHA , играют роль в процессинге рРНК-предшественников, способствуя сборке функциональных рибосом. [ 18 ]
DICER и DROSHA также из семейства РНКаз III играют роль в процессинге пре-миРНК в функциональную микроРНК. [ 19 ]
Созревание ногтей и волос
[ редактировать ]Эндонуклеаза DNase1L2 также вносит значительный вклад в удаление ДНК во время формирования волос и ногтей. Этот процесс важен для созревания структур волос и ногтей и имеет решающее значение для превращения клеток в прочные и ороговевшие структуры, обеспечивающие прочность и целостность волос и ногтей. [ 20 ]
Дальнейшее обсуждение
[ редактировать ]Могут быть обнаружены эндонуклеазы рестрикции, которые расщепляют стандартную дцДНК (двухцепочечную ДНК), или оцДНК (одноцепочечную ДНК), или даже РНК. [ нужна ссылка ] Это обсуждение ограничивается дцДНК; однако обсуждение можно распространить на следующее:
- Стандартная дцДНК
- Нестандартная ДНК
- Праздничные перекрестки
- Трехцепочечная ДНК , четырехцепочечная ДНК ( G-квадруплекс )
- Двухцепочечные гибриды ДНК и РНК (одна цепь — ДНК, другая — РНК). [ 4 ] : 72–73
- Синтетическая или искусственная ДНК (например, содержащая основания, отличные от A, C, G, T, см. работу Эрика Т. Кула ). Исследования с синтетическими кодонами см. исследования С. Беннера, а увеличение набора аминокислот в полипептидах, тем самым увеличивая протеом или протеомику , см. исследования П. Шульца. [ 4 ] : глава 3
Кроме того, сейчас ведутся исследования по созданию синтетических или искусственных эндонуклеаз рестрикции, особенно с сайтами узнавания, уникальными в пределах генома. [ нужна ссылка ]
Эндонуклеазы рестрикции или ферменты рестрикции обычно расщепляют двумя способами: с тупыми концами или с липкими концами. Пример эндонуклеазы рестрикции типа I. [ 4 ] : 64
Кроме того, существуют ДНК/РНК-неспецифические эндонуклеазы , такие как те, которые обнаружены у Serratia marcescens , которые действуют на дцДНК, оцДНК и РНК. [ нужна ссылка ]
Общие эндонуклеазы
[ редактировать ]Ниже приведены таблицы распространенных прокариотических и эукариотических эндонуклеаз. [ 21 ]
| Прокариотический фермент | Источник | Комментарии |
|---|---|---|
| RecBCD Эннуклеаза | кишечная палочка | Частично АТФ-зависимый; также экзонуклеаза; участвует в рекомбинации и репарации |
| Эндонуклеаза Т7 ( P00641 ) | фаг Т7 (ген 3) | Необходим для репликации; предпочтение одноцепочечной ДНК двухцепочечной |
| Эндонуклеаза Т4 II ( P07059 ) | фаг Т4(денА) | Разделяет последовательность -TpC- с получением олигонуклеотидов с концевыми 5'-dCMP; Длина цепочки продукта зависит от условий |
| Эндонуклеаза Bal 31 | П. эспехиана | Также экзонуклеаза; откусывает 3'- и 5'-концы дуплексной ДНК. Смесь как минимум двух нуклеаз, быстрой и медленной. [ 22 ] |
| Эндонуклеаза I (эндо I; P25736 ) | E. coli (endA) | Периплазматическая локализация; средняя длина цепочки продукта – 7; ингибируется тРНК; производит двухцепочечный разрыв ДНК; образует разрыв при образовании комплекса с тРНК; эндо I-мутанты растут нормально |
| Микрококковая нуклеаза ( P00644 ) | стафилококк | Производит 3'-P-концы; требует Ca2+; также действует на РНК; предпочитает одноцепочечную ДНК и богатые АТ участки |
| Эндонуклеаза II (эндо VI, экзо III; P09030 ) | кишечная палочка (xthA) | Расщепление рядом с сайтом AP; также 3'→5'-экзонуклеаза; фосфомоноэстераза на 3'-P-конце |
| Эукариотический фермент | Источник | Комментарии |
| Эндонуклеаза нейроспоры [ 23 ] | Neurospora crassa, митохондрии | Также действует на РНК. |
| Нуклеаза S1 ( P24021 ) | Аспергилл оризае | Также действует на РНК |
| P1-нуклеаза ( P24289 ) | Лимонный карандаш | Также действует на РНК |
| Нуклеаза бобов мунг I | ростки фасоли маш | Также действует на РНК |
| Нуклеаза Устилаго (Дназа I) [ 24 ] | Ustilago maydis | Также действует на РНК |
| ДНКа I ( P00639 ) | Поджелудочная железа крупного рогатого скота | Средняя длина цепочки продукта – 4; производит двойной разрыв цепи в присутствии Mn2+ |
| AP-эндонуклеаза | Ядро, митохондрии | Участвует в пути эксцизионной репарации оснований ДНК. |
| Эндо Р [ 25 ] | клетки HeLa | Специально для сайтов GC |
| ЗАСЛОН1 | Ядро | Отвечает за обработку Оказаки фрагментов во время репликации ДНК. |
Мутации
[ редактировать ]Пигментная ксеродерма — редкое аутосомно-рецессивное заболевание, вызываемое дефектной эндонуклеазой, специфичной к УФ-излучению. Пациенты с мутациями не способны восстанавливать повреждения ДНК, вызванные солнечным светом. [ 26 ]
Серповидноклеточная анемия – заболевание, вызванное точечной мутацией. Последовательность, измененная мутацией, устраняет сайт узнавания эндонуклеазы рестрикции MstII, которая распознает нуклеотидную последовательность. [ 27 ]
Мутации эндонуклеазы сплайсинга тРНК вызывают понтоцеребеллярную гипоплазию. Понтоцеребеллярные гипоплазии (ПГК) представляют собой группу нейродегенеративных аутосомно-рецессивных заболеваний, вызванных мутациями в трех из четырех различных субъединиц эндонуклеазного комплекса сплайсинга тРНК. [ 28 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «Свойства экзонуклеаз и эндонуклеаз» . Биолаборатории Новой Англии . 2017 . Проверено 21 мая 2017 г.
- ^ Слор, Ханох (14 апреля 1975 г.). «Дифференциация между экзонуклеазами и эндонуклеазами, а также между гаплотомными и диплотомными эндонуклеазами с использованием в качестве субстрата лунок пластиковых депрессивных пластин, покрытых 3-h-ДНК» . Исследования нуклеиновых кислот . 2 (6): 897–903. дои : 10.1093/нар/2.6.897 . ПМЦ 343476 . ПМИД 167356 .
- ^ Стивен Т. Килпатрик; Джоселин Э. Кребс; Левин, Бенджамин; Гольдштейн, Эллиотт (2011). Гены Левина X. Бостон: Джонс и Бартлетт. ISBN 978-0-7637-6632-0 .
- ^ Jump up to: а б с д и ж Кокс М., Нельсон Д.Р., Ленинджер А.Л. (2005). Ленингерские принципы биохимии . Сан-Франциско: WH Freeman. стр. 952 . ISBN 978-0-7167-4339-2 .
- ^ Саймон М (2010). Новые вычисления: акцент на биоинформатике . Нью-Йорк: Спрингер. п. 437. ИСБН 978-1441919632 .
- ^ Jump up to: а б с Смит, ХО; Натанс, Д. (15 декабря 1973 г.). «Предлагаемая номенклатура бактериальных систем модификации и рестрикции-хозяина и их ферментов» . Журнал молекулярной биологии . 81 (3): 419–23. дои : 10.1016/0022-2836(73)90152-6 . ПМИД 4588280 .
- ^ Рубин, РА; Модрич, П. (25 октября 1977 г.). «ЭкоРИ-метилаза» . Журнал биологической химии . 252 (20): 7265–72. дои : 10.1016/S0021-9258(19)66964-4 . ПМИД 332688 .
- ^ Лосик Р., Уотсон Дж.Д., Бейкер Т.А., Белл С., Ганн С., Левин М.В. (2008). Молекулярная биология гена . Сан-Франциско: Пирсон/Бенджамин Каммингс. ISBN 978-0-8053-9592-1 .
- ^ Элленбергер Т., Фридберг Э.К., Уокер Г.С., Вольфрам С., Вуд Р.Дж., Шульц Р. (2006). Репарация ДНК и мутагенез . Вашингтон, округ Колумбия: ASM Press. ISBN 978-1-55581-319-2 .
- ^ Альбертс Б (2002). Молекулярная биология клетки . Нью-Йорк: Garland Science. ISBN 978-0-8153-3218-3 .
- ^ Нишино Т., Морикава К. (декабрь 2002 г.). «Структура и функции нуклеаз в репарации ДНК: форма, захват и лезвие ножниц ДНК» . Онкоген . 21 (58): 9022–32. дои : 10.1038/sj.onc.1206135 . ПМИД 12483517 .
- ^ Jump up to: а б Ханада, К.; Будзовска, М.; Модести, М.; Маас, А.; Вайман, К.; Эссерс, Дж.; Канаар, Р. (2006). «Структурно-специфическая эндонуклеаза Mus81-Eme1 способствует превращению межцепочечных связей ДНК в двухцепочечные разрывы» . Журнал ЭМБО . 25 (20): 4921–4932. дои : 10.1038/sj.emboj.7601344 . ПМК 1618088 . ПМИД 17036055 .
- ^ Бернштейн, К. (1981). «Репарация дезоксирибонуклеиновой кислоты в бактериофаге» . Микробиологические обзоры . 45 (1): 72–98. дои : 10.1128/мр.45.1.72-98.1981 . ПМК 281499 . ПМИД 6261109 .
- ^ Ёсида, Акира; Помье, Ив; Уэда, Таканори (01 февраля 2006 г.). «Активация эндонуклеазы и фрагментация хромосомной ДНК во время апоптоза в клетках лейкемии» . Международный журнал гематологии . 84 (1): 31–37. дои : 10.1007/BF03342699 . ISSN 1865-3774 . ПМИД 16867899 . S2CID 25475000 .
- ^ Джин, Ён Хван; Оберт, Робин; Бургерс, Питер М.Дж.; Кункель, Томас А.; Резник, Майкл А.; Горденин, Дмитрий А. (24 апреля 2001 г.). «3'→5'-экзонуклеаза ДНК-полимеразы δ может заменить эндонуклеазу 5'-клапана Rad27/Fen1 при обработке фрагментов Оказаки и предотвращении нестабильности генома» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (9): 5122–5127. дои : 10.1073/pnas.091095198 . ISSN 0027-8424 . ПМК 33174 . ПМИД 11309502 .
- ^ Лю, Юань; Као, Хуэй-И; Бамбара, Роберт А. (июнь 2004 г.). «Лоскутная эндонуклеаза 1: центральный компонент метаболизма ДНК» . Ежегодный обзор биохимии . 73 (1): 589–615. doi : 10.1146/annurev.biochem.73.012803.092453 . ISSN 0066-4154 . ПМИД 15189154 .
- ^ Хартманн, Роланд К.; Гессрингер, Маркус; Шпет, Беттина; Фишер, Сьюзен; Марчфельдер, Анита (2009). «Создание тРНК и многого другого — РНКазы P и тРНКазы Z» . Прогресс молекулярной биологии и трансляционной науки . 85 : 319–368. дои : 10.1016/S0079-6603(08)00808-8 . ISSN 1877-1173 . ПМИД 19215776 .
- ^ Лежарс, Максенс; Кобаяши, Асаки; Хайнсдорф, Элиан (декабрь 2021 г.). «РНКаза III, биогенез рибосом и не только» . Микроорганизмы . 9 (12): 2608. doi : 10.3390/microorganisms9122608 . ПМЦ 8708148 . ПМИД 34946208 .
- ^ Кюбахер, Анжелика; Урбич, Кармен; Зейхер, Андреас М.; Диммелер, Стефани (6 июля 2007 г.). «Роль Дайсера и Дроши в экспрессии эндотелиальных микроРНК и ангиогенезе» . Исследование кровообращения . 101 (1): 59–68. дои : 10.1161/CIRCRESAHA.107.153916 . ISSN 0009-7330 . ПМИД 17540974 .
- ^ Фишер, Хайнц; Сабо, Сандра; Шерц, Дженнифер; Джагер, Карин; Росситер, Хайдемари; Бухбергер, Мария; Ганнадан, Мину; Германн, Марсела; Тойссль, Ганс-Кристиан; Тобин, Десмонд Дж.; Вагнер, Эрвин Ф.; Чахлер, Эрвин; Экхарт, Леопольд (июнь 2011 г.). «Основная роль кератиноцитарной эндонуклеазы DNase1L2 в удалении ядерной ДНК из волос и ногтей» . Журнал исследовательской дерматологии . 131 (6): 1208–1215. дои : 10.1038/jid.2011.13 . ISSN 0022-202X . ПМЦ 3185332 . ПМИД 21307874 .
- ^ Таня А. Бейкер; Корнберг, Артур (2005). Репликация ДНК . Университетская наука. ISBN 978-1-891389-44-3 .
- ^ Вэй, CF; Алианелл, Джорджия; Бенсен, GH; Грей Х.Б.-младший (25 ноября 1983 г.). «Выделение и сравнение двух молекулярных видов нуклеазы BAL 31 из Alteromonas espejiana с различными кинетическими свойствами» . Журнал биологической химии . 258 (22): 13506–12. дои : 10.1016/S0021-9258(17)43942-1 . ПМИД 6643438 .
- ^ Линн, С; Леман, ИК (10 июня 1966 г.). «Эндонуклеаза из митохондрий Neurospora crassa» . Журнал биологической химии . 241 (11): 2694–9. дои : 10.1016/S0021-9258(18)96595-6 . ПМИД 4287861 .
- ^ Холломан, ВК; Холлидей, Р. (10 декабря 1973 г.). «Исследования нуклеазы Ustilago maydis. I. Очистка, свойства и участие в рекомбинации фермента» . Журнал биологической химии . 248 (23): 8107–13. дои : 10.1016/S0021-9258(19)43199-2 . ПМИД 4201782 .
- ^ Готлиб, Дж; Музычка, Н. (5 июля 1990 г.). «Очистка и характеристика эндонуклеазы HeLa R. G-специфической эндонуклеазы млекопитающих» . Журнал биологической химии . 265 (19): 10836–41. дои : 10.1016/S0021-9258(19)38522-9 . ПМИД 2358441 .
- ^ Краткий обзор медицинской биохимии . Нью-Йорк: Уайли. 2012. ISBN 978-0-470-65451-4 .
- ^ Ферье Д.Р., Шамп ПК, Харви Р.П. (2008). Биохимия . Филадельфия: Уолтерс Клювер/Липпинкотт Уильямс и Уилкинс. ISBN 978-0-7817-6960-0 .
- ^ Бадд Б.С., Намавар И., Барт П.Г., Полл-Те БТ, Нюрнберг Г., Беккер С., ван Рейссен Ф., Ветерман М.А., Флюитер К., Те Бик Э.Т., Ароника Э., ван дер Кнаап М.С., Хёне В., Толиат М.Р., Кроу Ю.Дж. , Стейнлинг М, Войт Т, Руленсо Ф, Брюссель В, Брокманн К, Киллерман М, Больтшаузер Е, Хаммерсен Г, Виллемсен М, Базель-Ванагайте Л, Крегело-Манн И, де Врис Л.С., Штриха Л, Мунтони Ф, Ферри КД , Баттини Р., Хеннекам Р.К., Грилло Э., Бимер Ф.А., Стойтс Л.М., Воллник Б., Нюрнберг П., Баас Ф. (сентябрь 2008 г.). «Мутации эндонуклеазы сплайсинга тРНК вызывают понтоцеребеллярную гипоплазию». Нат. Жене. 40 (9): 1113–8. дои : 10.1038/ng.204 . ПМИД 18711368 . S2CID 205345070 .