Фораминифера
| Фораминифера Временный диапазон: Последний эдиакаран - решающий
| |
|---|---|

| |
| Live Amonnia Tepida (Rotaliida) | |
| Научная классификация | |
| Домен: | Эукариота |
| Клада : | Диафоретики |
| Клада : | Сар - |
| Филум: | Рейтарария |
| Субфилум: | Фораминифера D'Orbigny , 1826 |
| Подразделения | |
|
" Моноталамея "
неопределенность | |
Foraminifera ( / f ə ˌ r æ m ə ˈ n ɪ f ə r ə / fə- ram - nih -fə-rə ; латынь для "отверстий"; неофициально называется " forams ")- одноклетные организмы , члены филома характеризующихся или класса ресолярных ; протистов, потоковой гранулированной эктоплазмой для ловли пищи и других видов использования и обычно внешняя оболочка (называемая « тест ») разнообразных форм и материалов. Считается, что тесты хитина (обнаруженные в некоторых простых родах, и в частности, текстолярия ) являются наиболее примитивным типом. Большинство фораминиферов являются морскими, большинство из которых живут на или внутри морского дна (то есть, являются бентическими , с видами разных размеров, играющих роль в макробентосе , мейобентосе и Microbenthos ), [ 2 ] в то время как меньшее число плавает в толще воды на различных глубинах (то есть планктонно ), которые принадлежат подряд Globigerinina . [ 3 ] Меньше известно из пресной воды [ 4 ] или солоноватый [ 5 ] Условия, и некоторые очень немногие (неакватические) виды почвы были идентифицированы с помощью молекулярного анализа малой субъединицы рибосомной ДНК . [ 6 ] [ 7 ]
Foraminifera обычно производит тест или оболочку, которая может иметь одну или несколько камер, некоторые становятся довольно сложными по структуре. [ 8 ] Эти раковины обычно изготавливаются из карбоната кальция ( CACO
3 ) или агглютинированные частицы отложений. Признаны более 50 000 видов, оба живут (6700–10 000) [ 9 ] [ 10 ] и ископаемое (40 000). [ 11 ] [ 12 ] Они обычно имеют размер менее 1 мм, но некоторые гораздо больше, самый большой вид, достигающий до 20 см. [ 13 ]
В современном научном английском термин «Фораминифера» происхождения слова является как единственным, так и множественным числом (независимо от латинского ), и используется для описания одного или нескольких образцов или таксонов: его использование как единственное или множественное число должно определяться из контекста. Foraminifera часто используется неофициально для описания группы, и в этих случаях, как правило, нижний регистр. [ 14 ]
История обучения
[ редактировать ]Самая ранняя известная ссылка на Фораминифера исходит от Геродота , который в 5 веке до н.э. отметил их как создание скалы, которая образует великую пирамиду Гизы . Они сегодня признаны представителями рода Nummulites . Strabo , в 1 веке до нашей эры, отметил одни и те же фораминиферы и предположил, что это были остатки чечевицы, оставленные рабочими, которые построили пирамиды. [ 15 ]
Роберт Хук наблюдал фораминифера под микроскопом, как описано и проиллюстрировано в его книге 1665 года : Micrographia :
Я пробовал несколько маленьких и одиночных увеличительных очков и небрежно просмотрел участок с белым песком, когда я воспринял один из зерен точно формируется и вен способ с лучшим микроскопом и обнаружил его с обеих сторон, так и с краями, чтобы напоминать оболочку небольшой воды с плоской спиральной оболочкой [...] [ 16 ]
Антони Ван Леувенхук описала и проиллюстрировал фораминиферальные тесты в 1700 году, описывая их как мельчайшие кокеты; Его иллюстрация узнаваемо как эльфидий . [ 17 ] Ранние работники классифицировали фораминифер в рамках рода Nautilus , отметив их сходство с определенными головоногими . признал, В 1781 году Лоренц Спенглер что у Фараминифер были дыры в септе, что в конечном итоге предоставит группе свое название. [ 18 ] Спенглер также отметил, что перегородка фораминиферы вышла на противоположную сторону от септов на Наутили и что им не хватало нервной трубки. [ 19 ]

Alcide D'Orbigny , в его работе в 1826 году, считал их группой мельчайших головоногих и отметил свою странную морфологию, интерпретируя псевдоподию как щупальца и отмечая сильно уменьшенную (на самом деле отсутствие) головы. [ 20 ] Он назвал группу фораминифер , или «носители отверстий», поскольку члены группы имели дыры в разделениях между отсеками в их оболочках, в отличие от наутили или аммонитов . [ 14 ]
Протозойская природа Фораминифер была впервые признана Дюджардина в 1835 году. [ 18 ] Вскоре после этого в 1852 году D'Orbigny создал схему классификации, признавая 72 рода фораминифер, которые он классифицировал на основе формы теста - схемы, которая вызвала серьезную критику со стороны коллег. [ 17 ]
Монография HB Brady 1884 года описала фораминиферальные находки Challenger экспедиции . Брэди признал 10 семей с 29 подсемействами, не обращая внимания на стратиграфический диапазон; Его таксономия подчеркнула идею о том, что несколько разных символов должны разделять таксономические группы и, как таковые размещенные агглютинированные и известные роды в тесной связи.
Эта общая схема классификации останется до работы Кушмана в конце 1920 -х годов. Кушман рассматривал композицию стен как единственную наиболее важную черту в классификации фораминифер; Его классификация стала широко принятой, но также вызвала критику со стороны коллег за то, что они «не биологически звучат».
Геолог Ирен Кресс провел обширные исследования в этой области, опубликовав несколько девяносто статей, включая заметную работу над фораминиферой, как единственного автора, а также более двадцати в сотрудничестве с другими учеными. [ 21 ]
Схема Кушмана, тем не менее, оставалась доминирующей схемой классификации до тех пор, пока классификация Таппана и Лобеблиха 1964 года, которая вкладывала фораминиферы в общие группировки, все еще использовалась сегодня, основываясь на микроструктуре испытательной стены. [ 17 ] Эти группы были по-разному перемещены в соответствии с различными схемами классификации более высокого уровня. Использование Pawlowski (2013) молекулярной систематики в целом подтверждало группировки Таппана и Лобеблиха, причем некоторые из них обнаруживаются как полифилетические или парафилетические; Эта работа также помогла определить отношения более высокого уровня между основными фораминиферальными группами. [ 22 ]
Таксономия
[ редактировать ]
| |||||||||||||||||||||||||||||||||||||||||||||
| Филогения фораминиферы после Pawlowski et al. 2013. [ 22 ] Моноталамида упорядочивает астрорхизида и аллогромида, оба парафилетические. |
Таксономическая позиция фораминиферы варьировалась с Schultze в 1854 году, [ 23 ] который назвал приказом, Фораминиферида. Loeblich (1987) и Tappan (1992) переиграли фораминифер как класс [ 24 ] как это сейчас общепринято.
Фораминифера обычно были включены в простейшие , [ 25 ] [ 26 ] [ 27 ] или в аналогичной протоклисте или протеллическом королевстве . [ 28 ] [ 29 ] Убедительные доказательства, основанные в основном на молекулярной филогенетике , существуют для их принадлежности к крупной группе в рамках простейших, известной как Rhizaria . [ 25 ] До признания эволюционных отношений между членами Rhizaria фораминифера обычно сгруппировали с другими амебоидами в качестве филома -ризоподеи (или саркодина) в класс Granuloreticulosa.
Рузария проблематична, так как их часто называют «супергруппой», а не используют устоявшееся таксономическое звание, такое как тип . Кавалер-Смит определяет корневию как инфра-цалье в рамках простейшего царства. [ 25 ]
Некоторые таксономии поместили фораминифера в свой собственный тип, ставя их на одну одну амебоидную саркодину, в которую они были размещены.
Хотя молекулярные данные пока не поддерживаются морфологическими коррелятами, убедительно предполагают, что фораминифера тесно связаны с Cercozoa и Radiolaria , оба из которых также включают амебоиды со сложными оболочками; Эти три группы составляют корневику. [ 26 ] Тем не менее, точные отношения Forams с другими группами и друг с другом все еще не совсем ясны. Фораминифера тесно связаны с амебами тестирования . [ 30 ]
| Таксономия из Mikhalevich 2013 [ 31 ] |
|---|
Анатомия
[ редактировать ]
Самым ярким аспектом большинства фораминиферов являются их твердые оболочки или тесты. Они могут состоять из одной из нескольких камер и могут состоять из белка, частиц отложений, кальцита, арагонита или (в одном случае) кремнезема. [ 24 ] Некоторым фораминифера не хватает тестов полностью. [ 32 ] В отличие от других организмов, секретирующих раковину, таких как моллюски или кораллы , тесты фораминиферы расположены внутри клеточной мембраны , внутри протоплазмы . Оргалелы ячейки расположены в пределах отсека (ы) теста, а отверстие (ы) теста позволяет переносить материал из псевдоподий во внутреннюю ячейку и обратно. [ 33 ]
Фараминиферальная клетка делится на гранулирующую эндоплазму и прозрачную эктоплазму, из которой псевдоподиальная сеть может возникать через одно отверстие или через многие перфорации в тесте. Отдельные псевдоподы характерно имеют небольшие гранулы, потоковые в обоих направлениях. [ 34 ] Foraminifera уникальны в наличии гранулоретулосовой псевдоподий ; То есть их псевдоподия кажутся гранулированными под микроскопом; Эти псевдоподии часто удлинены и могут расколоться и присоединиться друг к другу. Они могут быть расширены и втянуты в соответствии с потребностями клетки. Псевдоподы используются для локомоции, привязки, экскреции, тестовой конструкции и захвата пищи, которое состоит из небольших организмов, таких как диатомовые или бактерии. [ 35 ] [ 33 ]
Помимо тестов, клетки на фораминиферации поддерживаются цитоскелетом микротрубочек , которые свободно расположены без структуры, наблюдаемой в других амебоидах. FORAMS развили специальные клеточные механизмы для быстрого сборки и разборки микротрубочек, что позволило бы быстрому образованию и ретракции удлиненной псевдоподии. [ 24 ]
В Gamont (сексуальная форма) фораминифера обычно имеет только одно ядро, в то время как агамонт (бесполая форма) имеет тенденцию иметь несколько ядер. По крайней мере, у некоторых видов ядра являются диморфными, с соматическими ядрами, содержащими в три раза больше белка и РНК, чем генеративные ядра. Тем не менее, ядерная анатомия кажется очень разнообразной. [ 36 ] Ядра не обязательно ограничены одной камерой у многоканированных видов. Ядра могут быть сферическими или иметь много доли. Ядра обычно имеют диаметр 30-50 мкм. [ 37 ]
Некоторые виды фораминифер имеют большие пустые вакуоли в своих клетках; Точная цель этого неясна, но их предложили функционировать как резервуар нитрата. [ 37 ]
Митохондрии распределяются равномерно по всей клетке, хотя у некоторых видов они сосредоточены под полями и вокруг внешнего края клетки. Это было предположительно, чтобы адаптация к низкоохищенным средам. [ 37 ]
несколько видов ксенофиофора Было обнаружено, что имеют необычайно высокие концентрации радиоактивных изотопов в своих клетках, среди самых высоких из всех эукариоте. Цель этого неизвестна. [ 38 ]
Экология
[ редактировать ]
Современные фораманифера - это в основном морские организмы, но живые люди были найдены в солоноватой пресной воде [ 34 ] и даже наземные места обитания. [ 7 ] Большинство видов являются бентическими , и еще 50 морфоспектов планктонно . [ 35 ] Этот счет может, однако, представлять собой лишь часть фактического разнообразия, поскольку многие генетически различные виды могут быть морфологически неразличимыми. [ 40 ]
Бентические фораминиферы обычно встречаются в мелкозернистых отложениях, где они активно перемещаются между слоями; Тем не менее, многие виды обнаруживаются на подложках твердой породы, прикрепленными к водородам или сидят на поверхности отложения. [ 24 ]
Большинство планктонных фораминиферов встречаются в Globigerinina , происхождении в Роталии . [ 22 ] Тем не менее, по крайней мере еще одна существующая роталийская линия, Neogallitellia , кажется, независимо развивала планктонный образ жизни. [ 41 ] [ 42 ] Кроме того, было высказано предположение, что некоторые ископаемые юрскими ископаемыми фосмами, возможно, также независимо развивались планктонный образ жизни и, возможно, были членами Робертиниды. [ 43 ]
Ряд фурамов, как бентических, так и планктонных, [ 44 ] [ 45 ] Имеют одноклеточные водоросли в качестве эндосимбионтов , из различных линий, таких как зеленые водоросли , красные водоросли , золотые водоросли , диатомовые терапии и динофлагелляты . [ 35 ] Эти миксотрофные фораминифер особенно распространены в океанических водах с бедными питательными веществами. [ 46 ] Некоторые Forams являются kleptoplastic , сохраняя хлоропласты от проглатываемых водорослей для проведения фотосинтеза . [ 47 ]
Большинство фораминиферов являются гетеротрофными, потребляют меньшие организмы и органические вещества; Некоторые меньшие виды представляют собой специализированные кормушки на фитодетрит , а другие специализируются на потреблении диатомов. Некоторые бентические отверстия построены питательные кисты, используя псеуодоподию, чтобы воздействовать на себя внутри отложений и органических частиц. [ 24 ] Определенная добыча на форуме на мелких животных, таких как копеподы или пауманов ; Некоторые отверстия даже предшествуют другим отверстиям, сверляют отверстия в тестах их добычи. [ 48 ] Одна группа, ксенофиофоры, была предложена для фермерских бактерий в рамках их тестов, хотя исследования не смогли найти поддержку этой гипотезы. [ 49 ] Подвесное кормление также распространено в группе, и, по крайней мере, некоторые виды могут воспользоваться растворенным органическим углеродом . [ 24 ]
Несколько видов FORAM являются паразитными , заражающими губками, моллюсками, кораллами или даже другими фораминиферами. Паразитические стратегии различаются; Некоторые действуют как эктопаразиты, используя свою псевдоподию для кражи пищи у хозяина, в то время как другие прорываются через раковину или стенку тела своего хозяина, чтобы питаться его мягкими тканями. [ 24 ]
Фораминиферы сами поедают множеством более крупных организмов, включая беспозвоночных, рыб, береговые птицы и другие фораминиферы. Однако было высказано предположение, что в некоторых случаях хищники могут быть больше заинтересованы в кальцие от оболочек FORAM, чем в самих организмах. Известно, что несколько видов водных улиток селективно питаются фораминифера, часто даже предпочитая отдельные виды. [ 50 ]
Было обнаружено, что некоторые бентические фораминиферы способны выжить в течение более 24 часов, что указывает на то, что они способны к избирательному анаэробному дыханию . Это интерпретируется как адаптация к выживанию изменяющихся кислородных условий вблизи границы раздела отложений. [ 51 ]
Фораминифера встречаются в самых глубоких частях океана, таких как траншея Мариана , в том числе «Челленджер Глуб» , самая глубокая известная часть. На этих глубинах, ниже глубины карбонатной компенсации , карбонат кальция испытаний растворим в воде из -за крайнего давления. Фораминифера, обнаруженные в глубине Challenger, таким образом не имеют тестирования карбоната, но вместо этого имеют один из органических материалов. [ 52 ]
Немариновые фораминиферы традиционно пренебрегали в исследованиях FORAM, но недавние исследования показывают, что они существенно более разнообразны, чем ранее известные. Известно, что они населяют разрозненные экологические ниши, в том числе мхи , реки, озера и пруды, водно -болотные угодья, почвы, торфяные болоты и песчаные дюны. [ 53 ]
Размножение
[ редактировать ]Обобщенный фораминиферальный жизненный цикл включает в себя чередование между гаплоидными и диплоидными поколениями, хотя они в основном похожи по форме. [ 23 ] [ 54 ] Гаплоид или Гамонт изначально имеет одно ядро и делятся на производство многочисленных гамет , которые обычно имеют два жгутика . Диплоид или агамонт является многоядерным , и после того, как мейоз делится на получение новых абонентов. Многочисленные раунды бесполого воспроизведения между сексуальными поколениями не редкость в бентических формах. [ 34 ]

Фораминифера демонстрирует морфологический диморфизм, связанный с их репродуктивным циклом. Гамонт, или сексуально воспроизводящая гаплоидную форму, является мегалосферной , то есть его пролокулусом или первой камерой, пропорционально большой. Гамонт также известен как форма . Гэмы, несмотря на то, что они обычно имеют большие пролокули, также обычно имеют меньший общий диаметр испытаний, чем агамонты.
После достижения зрелости Gamont делится через митоз, чтобы произвести тысячи гаметов, которые также являются гаплоидными. Все эти гаметы имеют полный набор органеллов и изгнаны из теста в окружающую среду, оставляя тест неповрежденным. Гамет не дифференцируется в сперму и яйцо , и любые два гамета от вида, как правило, могут оплодотворять друг друга.

Когда два гамета объединяются, они создают диплоидную многонациональную клетку, известную как Агамонта или B. форма В отличие от Gamont, агамонт является микросферентом , с пропорционально небольшой первой камерой, но, как правило, большим общим диаметром с большим количеством камер. Агамонт - это асексуальная фаза воспроизведения фораминифер; После достижения взрослой жизни протоплазма полностью освобождает тест и делит свою цитоплазму мейотически через множественное деление, образуя ряд гаплоидных потомков. Эти потомки затем начинают формировать свою мегалосферную первую камеру, прежде чем рассеять.
В некоторых случаях гаплоидный молодой может созревать в мегалосферную форму, которая затем воспроизводит асексально для получения другого мегалосферного, гаплоидного потомства. этом случае первая мегалосферная форма называется Schizont или формой 1 В , в то время как вторая называется Gamont или 2 -й формой.

Созревание и размножение встречаются медленнее в более холодной и более глубокой воде; Эти условия также приводят к увеличению фурамов. Формы всегда кажутся гораздо более многочисленными, чем формы B , вероятно, из -за сниженной вероятности двух гамет встретятся друг с другом и успешно объединяются. [ 55 ] [ 33 ]
Вариации в репродуктивном режиме
[ редактировать ]Существует высокая степень разнообразия в репродуктивных стратегиях в разных фораминиферальных группах.
У однородовых видов форма A и B -форма все еще присутствуют. Как и в микросферном морф мультилокулярных Forams, асексально воспроизводимая B -форма больше, чем сексуально воспроизводя форма .
У Forams в семействе Spirillinidae есть амебоидные гаметы, а не жгутиковые. Другие аспекты размножения в этой группе, как правило, похожи на аспекты других групп Forams.
Извеска -спириллинид Patellina Corrugata имеет немного отличную репродуктивную стратегию, чем большинство других фораминифер. Асексально воспроизводящая форма B -форма производит кисту, которая окружает всю клетку; Затем он делится в этой кисте, и ювенильные клетки каннибализируют кальцит теста родителя, чтобы сформировать первую камеру их собственного теста. Эти формы после зрелости собираются в группы до девяти человек; Затем они образуют защитную кисту вокруг всей группы. Гаметогенез происходит в этой кисте, производя очень низкое количество гамет. Личинки формы B продуцируются внутри кисты; Любые ядра, которые не связаны в клетки, потребляются в качестве пищи для развивающихся личинок. Patellina в форме , как сообщается, является диологическим , с полами, называемыми «плюс» и «минус»; Эти полов различаются по количеству ядер, причем форма «плюс» имеет три ядра и форму «минус», имеющая четыре ядра. Форма B снова больше, чем A. форма [ 33 ] [ 55 ] [ 48 ]
Тесты
[ редактировать ]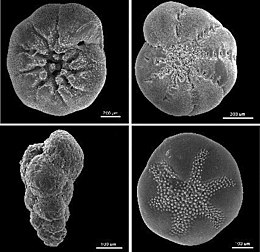
| Часть серии, связанной с |
| Биоминерализация |
|---|
 |
Фораминиферальные испытания служат для защиты организма внутри. Из -за их в целом жесткой и долговечной конструкции (по сравнению с другими протистами) тесты фораминифер являются основным источником научных знаний о группе.
Отверстия в тесте, которые позволяют цитоплазме вытягивать снаружи, называются апертурами. [ 56 ] Первичная апертура, приводящая к внешней части, принимает множество различных форм у разных видов, включая, помимо прочего, округление, в форме полумесяца, щелеобразная, капюшона, излучение (звездообразная), дендритная (разветвление). У некоторых фораминиферов «зубчатые», фланцевые или плитанные первичные отверстия. Может быть только одна первичная диафрагма или множественная; Когда несколько присутствуют, они могут быть кластеризованными или экваториальными. В дополнение к первичной апертуре, многие фораминиферы имеют дополнительные апертуры. Они могут образовываться как реликтные апертуры (прошлые первичные отверстия с более ранней стадии роста) или как уникальные структуры.
Форма теста сильно варьируется среди разных фораминиферов; Они могут быть однокамерными (универсальными) или многокамерными (многоядерными). В многоотлательных формах новые камеры добавляются по мере роста организма. Широкий спектр морфологий испытаний обнаруживается как в универсальных, так и в многоотлательных формах, включая спиралию, серийные и милиолины, среди прочего. [ 33 ]
Многие фораминифера демонстрируют диморфизм в своих тестах, с мегалосферными и микросферными людьми. Эти имена не должны восприниматься как ссылка на размер полного организма; Скорее, они относятся к размеру первой камеры или Proloculus . Тесты, как окаменелости, известны с эдиакарана , [ 57 ] и многие морские отложения составлены в основном из них. Например, известняк, составляющий пирамиды Египта, состоит почти полностью из нумерутских бентических фораминиферов. [ 58 ] По оценкам, риф -фораминифер генерирует около 43 миллионов тонн карбоната кальция в год. [ 59 ]
Генетические исследования идентифицировали обнаженную амебу ретикуломиксу и своеобразные ксенофиофоры в качестве фораминиферов без тестов. Несколько других амебоидов производят псевдопод ретикулозы и ранее были классифицированы с помощью FORAMS как гранулеретикулоза, но это больше не считается естественной группой, и большинство в настоящее время размещаются среди Cercozoaa. [ 60 ]
Эволюционная история
[ редактировать ]Молекулярные часы указывают на то, что группа короны фораминифер, вероятно, развивалась во время неопротерозоя , между 900 и 650 миллионами лет назад; Это время согласуется с неопротерозойскими окаменелостями близкородственных амебы с филозы . Поскольку ископаемые форатаминиферы не были обнаружены до самого конца эдиакарана , вполне вероятно, что большинство из этих протерозойских форм не имели тестов с жестким оболочкой. [ 61 ] [ 62 ]
Из-за их не-минерализированных тестов « аллограммиды » не имеют ископаемых записей. [ 61 ]

таинственные вендозоицы Предполагается, что эдиакарского периода представляют ископаемые ксенофиофоры . [ 63 ] Тем не менее, открытие диагенетически измененных C 27 Стеролов , связанных с остатками Дикинсонии, ставит под сомнение эту идентификацию и предполагает, что это может быть животным. [ 64 ] Другие исследователи предположили, что неуловимый трассировка ископаемого палеодиктиона и его родственников могут представлять собой ископаемое ксенофиофор [ 65 ] и отметил сходство существующего ксенофиофора Occultammina с ископаемым; [ 66 ] Тем не менее, современные примеры палеодиктиона не смогли понять проблему, и след может попеременно представлять нору или стеклянную губку. [ 67 ] Поддержка этого понятия является сходной средой обитания живых ксенофиофоров к предполагаемой среде обитания ископаемых графоглиптидов; Тем не менее, большой размер и регулярность многих графоглиптидов, а также очевидное отсутствие Xenophyae в их окаменелостях ставят под сомнение возможность. [ 66 ] По состоянию на 2017 год не было обнаружено определенных окаменелостей Xenophyophore. [ 68 ]
Тестовая фораминифера имеет отличную ископаемой записи на протяжении всего фанерозойского эона. Самые ранние известные определенные фораминиферы появляются в ископаемом отчете к самому концу Эдиакарана; Все эти формы имеют агглютинированные тесты и являются однонаумилярными. К ним относятся такие формы, как платисолениты и спиросоленты . [ 69 ] [ 57 ] Одноканированные фораминифера продолжали диверсифицироваться по всему кембрийу. Некоторые обычно встречающиеся формы Ammodiscus , Glomospira , Psammosphera и включают Turritellellel ; Эти виды все агглютинированы. Они составляют часть Ammodiscina , линии спириллинидов , которая до сих пор содержит современные формы. [ 70 ] [ 22 ] Более поздние спириллиниды будут развивать многоотборность и кальцитовые тесты, причем первые такие формы появляются во время триаса ; Группа увидела небольшое влияние на разнообразие из-за вымирания K-PG . [ 71 ]
Самыми ранними многокамерными фораминифера являются агглютинированными видами и появляются в ископаемом отчете в течение среднего кембрийского периода. Из -за их плохого сохранения их нельзя положить положительное назначение в какую -либо крупную группу FORAM. [ 70 ]

Самыми ранними известными ракообразными фарраминиферами являются фусулиниды , которые появляются в ископаемом отчете во время Лланковеровской эпохи раннего силурия . Самыми ранними из них были микроскопические, планоспирские спирали и эволютные; Более поздние формы развили разнообразие форм, включая линзоулярные, глобулярные и удлиненные формы в форме риса. [ 72 ] [ 73 ]
Более поздние виды фусулинидов выросли до гораздо большего размера, при этом некоторые формы достигли 5 см в длину; Как сообщается, некоторые образцы достигают длины до 14 см, что делает их одним из самых больших фораминиферов или вымерших. Фусулиниды - самая ранняя линия фораминифер, которые, как считается, развили симбиоз с фотосинтетическими организмами. Окаменелости фусулинидов были обнаружены на всех континентах, кроме Антарктиды ; Они достигли своего наибольшего разнообразия во время висейской эпохи каменного . Затем группа постепенно снизилась в разнообразии, пока, наконец, не вымерла во время пермо-триссового вымирания . [ 33 ] [ 71 ] [ 74 ]
Во время турниазийской эпохи каменноугольной, милиолидные фораминиферы впервые появились в записи окаменелости, расходящиеся от спириллинидов в туботаламеи . Милиолиды понесли около 50% потери во время промистических и вымираний K-PG, но выжили до сегодняшнего дня. Некоторые ископаемые милиолиды достигли 2 см в диаметре. [ 71 ]

Самые ранние известные лагенидные окаменелости появляются во время московской эпохи каменноугольного происхождения. Увидев небольшой эффект из-за пермо-триассического или вымирания K-PG, группа диверсифицировалась во времени. Во втором турнирном таксоне развивалось во время юрского периода и мела.
Самые ранние употребляющие вновь окаменелости появляются во время Пермского; Линия диверсифицировалась на протяжении всего мезозоя Евразии, прежде чем явно исчез из ископаемого рекорда после сеномано-туронического океана . «Devante Group Planispirililinidae» была передана в Awantinida, но это остается предметом дебатов. [ 75 ] [ 71 ]
Робертинида анисийской впервые появляется в ископаемом отчете во время эпохи триаса. Группа оставалась в низком разнообразии на протяжении всей своей ископаемой истории; Все живые представители принадлежат Robertinidae , которые впервые появились во время палеоцена . [ 71 ]
Первые определенные ископаемые роталиид не появляются в ископаемом отчете до тех пор , пока в Плейенсбачской эпохе юрского периода после триасового события . [ 76 ] Разнообразие группы оставалось низким до последствий сеномано-туронского события, после чего группа увидела быструю диверсификацию. Из этой группы планктонная Globigerinina - первая известная группа планктонных фурамов - сначала появляется после того, как Toarcian Turnover ; Группа увидела тяжелые потери как во время вымирания K-PG, так и во время вымирания эоцена-олигоцена , но остается существующим и разнообразным по сей день. [ 71 ] Дополнительная эволюция планктонного образа жизни произошла в миоцене или плиоцене, когда ротальид Neogallitellia независимо развивал планктонный образ жизни. [ 41 ] [ 42 ]
Палеонтологические применения
[ редактировать ]Умирающие планктонные фораминиферы непрерывно дождь на морском дне в огромных количествах, их минерализованные тесты сохраняются как окаменелости в накопленном отложении . Начиная с 1960-х годов и в основном под эгидой глубокого моря бурового бурения , бурения океана и международных буровых программ океана, а также для целей разведки нефти, продвинутые методы бурения глубоководных сил поднимают ядра с отложениями. Полем [ 77 ] доступные для ядер, вызвали исключительно высококачественные планктонные отчеты ископаемого на форы Эффективно неограниченное снабжение этих ископаемых тестов и относительно высокие модели по возрасту , эволюционный процесс. [ 77 ] Исключительное качество ископаемого записи позволило разработать впечатляюще подробную картину видовых взаимосвязи, которые будут разработаны на основе окаменелостей, во многих случаях впоследствии независимо подтверждены путем молекулярных генетических исследований по существующим образцам [ 78 ]
Поскольку определенные типы фораминиферов встречаются только в определенных условиях, их окаменелости могут быть использованы для выяснения вида окружающей среды, в которой были отложены древние морские отложения; Такие условия, как соленость, глубина, кислородные условия и условия освещения, могут быть определены из различных предпочтений среды обитания различных видов FORAM. Это позволяет работникам отслеживать изменение климата и условия окружающей среды с течением времени, агрегируя информацию о присутствующих фораминифере. [ 79 ]
В других случаях относительная доля окаменелостей с планктоническими или бентическими окаменелостями, обнаруженными в скале, может использоваться в качестве прокси для глубины данной местности при откладывании. [ 80 ]


По крайней мере, с 1997 года максимум термического палеоцена -эоцена (PETM) был исследован как аналогия для понимания воздействия глобального потепления и массивного углеродного входа в океан и атмосферу, включая подкисление океана . [ 85 ] Люди сегодня излучают около 10 GT углерода (около 37 GT CO2E) в год, и с этой скоростью выпустит сопоставимую сумму для PETM примерно через тысячу лет. Основное отличие состоит в том, что во время PETM планета была без льда, так как проход Дрейка еще не открылся, а центральная Америка еще не закрылась. [ 86 ] Хотя PETM в настоящее время считается тематическим исследованием для глобального потепления и массового излучения углерода, причина, детали и общее значение события остаются неопределенными. [ 87 ] [ 88 ] [ 89 ] [ 90 ]

Фораминифера имеет значительное применение в области биостратиграфии . Из -за их небольшого размера и твердых раковинов фораминифер может быть сохранена в большом численности и с высоким качеством сохранения; Из -за их сложной морфологии отдельные виды легко узнаваемы. Виды Foraminifera в ископаемом отчете имеют ограниченные диапазоны между первой эволюцией вида и их исчезновением; Стратиграфы разработали последовательные изменения в сборках FORAM на протяжении большей части фанерозой . Таким образом, сборка Foraminifera в данном месте может быть проанализирована и сравнивается с известными датами внешнего вида и исчезновения, чтобы сузить возраст скал. Это позволяет палеонтологам интерпретировать возраст осадочных пород , когда радиометрическое датирование не применимо. [ 91 ] Это применение Foraminifera было обнаружено Alva C. Ellisor в 1920 году. [ 92 ]

Извешенные ископаемые ископаемые фораминифера образуются из элементов, найденных в древних морях, где они жили. Таким образом, они очень полезны в палеоклиматологии и палеокеанографии . их можно использовать В качестве климатического прокси -сервера для реконструкции прошлого климата, изучая стабильные коэффициенты изотопного и содержимое микроэлемента в оболочках (тесты). Глобальная температура и объем льда могут быть обнаружены изотопами кислорода, а также в истории углеродного цикла и продуктивности океана путем изучения стабильных изотопных соотношений углерода; [ 93 ] См. Δ18O и Δ13C . Концентрация микроэлементов, таких как стронций (SR), [ 94 ] магний (мг), [ 95 ] литий (li) [ 96 ] и бор (б), [ 97 ] Также храните множество информации о глобальных температурных циклах, континентальном выветривании и роли океана в глобальном углеродном цикле. Географические закономерности, наблюдаемые в ископаемых записях планктонных фурматов, также используются для восстановления древних океанских течений .
Современное использование
[ редактировать ]Нефтяная промышленность в значительной степени зависит от микрофоссилий, таких как Forams, чтобы найти потенциальные отложения углеводородов. [ 98 ]

По тем же причинам они делают полезные биостратиграфические маркеры, живые фораминиферальные собрания использовались в качестве биоиндикаторов в прибрежной среде, включая показатели здоровья коралловых рифов. Поскольку карбонат кальция подвержен растворению в кислых условиях, на фораминифера может быть особенно влиять на изменение климата и подкисления океана .

Foraminifera обладает много применения в разведке нефти и обычно используется для интерпретации возрастов и палеоража осадочных слоев в нефтяных скважинах. [ 99 ] Агглютинированные ископаемые отверстия, похороненные глубоко в осадочных бассейнах, могут использоваться для оценки тепловой зрелости, что является ключевым фактором для генерации нефти. Фораминиферальный индекс окраски [ 100 ] (FCI) используется для количественной оценки изменений цвета и оценки температуры захоронения. Данные FCI особенно полезны на ранних стадиях генерации нефти (около 100 ° C).
Foraminifera также можно использовать в археологии в проведении некоторых типов каменного сырья. Некоторые типы камней, такие как известняк , обычно содержат окаменелых фомпилированных фораминиферов. Типы и концентрации этих окаменелостей в образце камня могут быть использованы, чтобы соответствовать этому образцу с источником, который, как известно, содержит ту же «ископаемое подпись». [ 101 ]
Галерея
[ редактировать ]-
 Фораминифера острова Паг, Адриатическое море -60 м, ширина поля 5,5 мм
Фораминифера острова Паг, Адриатическое море -60 м, ширина поля 5,5 мм -
 Фораминифера острова Паг, Адриатическое море -60 м, ширина поля 5,5 мм
Фораминифера острова Паг, Адриатическое море -60 м, ширина поля 5,5 мм -
 Фораминифера острова Паг, Адриатическое море -60 м, ширина поля 5,5 мм
Фораминифера острова Паг, Адриатическое море -60 м, ширина поля 5,5 мм -
 Фораминифера острова Паг, Адриатическое море -60 м, ширина поля 5,5 мм
Фораминифера острова Паг, Адриатическое море -60 м, ширина поля 5,5 мм -
 Фораминифера Индийского океана, юго-восточного побережья Бали, ширина поля 5,5 мм
Фораминифера Индийского океана, юго-восточного побережья Бали, ширина поля 5,5 мм -
 Фораминифера Индийского океана, юго-восточного побережья Бали, ширина поля 5,5 мм
Фораминифера Индийского океана, юго-восточного побережья Бали, ширина поля 5,5 мм -
 Фораминифера Индийского океана, юго-восточного побережья Бали, ширина поля 5,5 мм
Фораминифера Индийского океана, юго-восточного побережья Бали, ширина поля 5,5 мм -
 Foraminifera в Нгапали, Мьянма, ширина поля 5,22 мм
Foraminifera в Нгапали, Мьянма, ширина поля 5,22 мм -
 Foraminifera Heterostegina Depressa , ширина поля 4,4 мм
Foraminifera Heterostegina Depressa , ширина поля 4,4 мм









Ссылки
[ редактировать ]- ^ Лаура Вегенер Парфри ; Даниэль Дж.Г. Лара; Эндрю Х Кнолл ; Лаура Кац (16 августа 2011 г.). «Оценка времени ранней эукариотической диверсификации с помощью многогенных молекулярных часов» (PDF) . Труды Национальной академии наук Соединенных Штатов Америки . 108 (33): 13624–9. BIBCODE : 2011PNAS..10813624P . doi : 10.1073/pnas.1110633108 . ISSN 0027-8424 . PMC 3158185 . PMID 21810989 . Wikidata Q24614721 .
- ^ Бинцевска, Анна; Половодова Астенан, Ирина; Farmer, Elizabeth J. (2014), «Фораминиферы (бентические)» , в Harff, Jan; Месхеде, Мартин; Петерсен, Свен; Тиде, Йорн (ред.), Энциклопедия морских героссинсов , Дордрехт: Спрингер Нидерланды, с. 1–8, doi : 10.1007/978-94-6644-0_60-1 , ISBN 978-94-007-6644-0 , Получено 13 мая 2021 года
- ^ Katsunori (2015), Ohtsuka Susumu ; Kimoto , , doi : 10.1007/978-4-431-55130-0_7 , ISBN 978-4-431-55130-0 , Получено 13 мая 2021 года
- ^ Сиеменсма, паром; Apothéloz-Perret-Gentil, Laure; Хольцманн, Мария; Клаусс, Штеффен; Völcker, Eckhard; Pawlowski, январь (1 августа 2017 г.). «Таксономическое пересмотр пресноводных фораминифер с описанием двух новых агглютинированных видов и родов» . Европейский журнал профистологии . 60 : 28–44. doi : 10.1016/j.ejop.2017.05.006 . ISSN 0932-4739 . PMID 28609684 .
- ^ Болтовскоя, Эстебан; Райт, Рамиль (1976), Болтовскоя, Эстебан; Райт, Рамиль (ред.), «Бентонические фораминифер из солоноватых, гиперсолиновых, ультразалиновых и пресных вод» , недавние фораминиферы , Дордрехт: Спрингер Нидерланды, стр. 139–150, doi : 10.1007/978-94-017-2860-7_5. , ISBN 978-94-017-2860-7 , Получено 13 мая 2021 года
- ^ Джир, Олав (2009). Мейобентология: микроскопическая подвижная фауна водных отложений (2 -е изд.). Берлин: Спрингер. ISBN 978-3540686576 .
- ^ Jump up to: а беременный Lejzerwicz, Франк; Павловский, Ян; Fraissinet-Tachet, Лоуренс; Marmeisse, Роланд (1 сентября 2010 года). «Молекулярные доказательства широко распространенного возникновения фораминифер в почвах». Экологическая микробиология . 12 (9): 2518–26. BIBCODE : 2010ENVMI..12.2518L . doi : 10.1111/j.1462-2920.2010.02225.x . PMID 20406290 . S2CID 20940138 .
- ^ Кеннетт, JP; Шринивасан, MS (1983). Неоген -планктонные фораминиферы: филогенетический атлас . Хатчинсон Росс. ISBN 978-0-87933-070-5 .
- ^ Pawlowski, J.; Lejzerwicz, F.; Esling, P. (1 октября 2014 г.). «Обследования разнообразия окружающей среды следующего поколения на фораминифере: подготовка будущего» . Биологический бюллетень . 227 (2): 93–106. doi : 10.1086/bblv227n2p93 . ISSN 0006-3185 . PMID 25411369 . S2CID 24388876 .
- ^ Ald, SM et al . (2007) Разнообразие, номенклатура и таксономия протистов , Syst. Биол. 56 (4), 684–689, doi: 10.1080/10635150701494127.
- ^ Pawlowski, J., Lejzerwicz, F. & Esling, P. (2014). Опросы экологического разнообразия следующего поколения Foraminifera: подготовка будущего . Биологический бюллетень, 227 (2), 93-106.
- ^ «Мировой база данных Foraminifera» .
- ^ Маршалл М (3 февраля 2010 г.). «Зоологигер:« Живой пляжный мяч » - это гигантская одиночная камера» . Новый ученый .
- ^ Jump up to: а беременный Lipps JH, Finger KL, Walker SE (октябрь 2011 г.). «Как мы должны назвать фораминифера» (PDF) . Журнал фораминиферальных исследований . 41 (4): 309–313. Bibcode : 2011jforr..41..309L . doi : 10.2113/gsjfr.41.4.309 . Получено 10 апреля 2018 года .
- ^ «Foraminifera | Fossil Focus | Время | Обнаружение геологии | Британская геологическая служба (BGS)» . bgs.ac.uk. Получено 20 июля 2020 года .
- ^ «Микрография, или, некоторые физиологические описания мельчайших тел, изготовленных увеличительными очками? С наблюдениями и запросами за ней . Интернет -архив . 1665 . Получено 20 июля 2020 года .
- ^ Jump up to: а беременный в Sen Gupta, Barun K. (2003), «Систематика модера Foraminifera», в Sen Gupta, Barun K. (ed.), Modern Foraminifera , Springer Netherlands, с. 7–36, doi : 10.1007/0-306-48104 -9_2 , ISBN 978-0-306-48104-8
- ^ Jump up to: а беременный Boudagher-Fadel, Marcelle K. (2018), «Биология и эволюционная история более крупных бентических фораминиферов», Эволюция и геологическая значимость более крупных бентических фораминиферов (2 изд.), UCL Press, стр. 1–44, doi : 10.2307/j .ctvqhsq3.3 , Isbn 978-1-911576-94-5 , JSTOR J.CTVQHSQ3.3
- ^ Хансен Х. (1 января 1981 г.). «На Лоренце Спенглер и неотип для фораминифер Калларина Спенглери» .
- ^ D'Orbigny, Alcide (1826). «Методичная таблица класса головоногих» . Анналы естественных наук, Париж . Серия 1. 7 : 245–314 - через биологическую библиотеку наследия биоразнообразия.
- ^ «Кеннеди, Маргарет Мур (1896–1967)» , Оксфордский словарь национальной биографии , издательство Оксфордского университета, 6 февраля 2018 года, DOI : 10.1093/ODNB/9780192683120.013.34281 , извлечен 3 ноября 2022 года.
- ^ Jump up to: а беременный в дюймовый Павловский, Ян; Хольцманн, Мария; Тайска, Ярослав (1 апреля 2013 г.). «Новая надвергальная классификация Foraminifera: молекулы соответствуют морфологии» . Морская микропалеонтология . 100 : 1–10. Bibcode : 2013marmp.100 .... 1p . doi : 10.1016/j.marmicro.2013.04.002 . ISSN 0377-8398 .
- ^ Jump up to: а беременный Loeblich, AR Jr.; Таппан Х. (1964). «Фараминиферида». Часть C, Protista 2 . Трактат о палеонтологии беспозвоночных. Геологическое общество Америки. с. C55 - C786. ISBN 978-0-8137-3003-5 .
- ^ Jump up to: а беременный в дюймовый и фон глин Покойный Гупта, Барун К. (2002). Современные фора . Спрингер. П. 978-1-4020-0598-5 .
- ^ Jump up to: а беременный в Cavalier-Smith, T (2004). «Только шесть королевств жизни» (PDF) . Разбирательство. Биологические науки . 271 (1545): 1251–62. doi : 10.1098/rspb.2004.2705 . PMC 1691724 . PMID 15306349 .
- ^ Jump up to: а беременный Cavalier-Smith, T (2003). «Протестная филогения и высокоуровневая классификация простейших». Европейский журнал профистологии . 34 (4): 338–348. doi : 10.1078/0932-4739-00002 .
- ^ "Толвеб Cercozoa" . Архивировано с оригинала 25 декабря 2019 года . Получено 21 июня 2010 года .
- ^ Европейский реестр морских видов
- ^ eforams-taxonomy Archived 3 октября 2011 года на машине Wayback
- ^ Амебы проверены как индикаторы экологических технологий (PDF) , архивировав из оригинала (PDF) 27 ноября 2016 года , извлеченные 27 ноября 2016 года.
- ^ Михалевич, VI (2013). «Новое понимание систематики и эволюции фораминифер» . Микропалеонтология . 59 (6): 493–527. Bibcode : 2013mipal..59..493m . doi : 10.47894/mpal.59.6.01 . S2CID 90243148 .
- ^ Павловский, Ян; Боливар, Игнасио; Фарни, Джозеф Ф.; Варгас, Колумбан де; Баузер, Сэмюэль С. (1999). (Молекулярные доказательства). А 46 (6): 612–617. doi : 10.111/ j.1550-7 ISSN 1550-7408 . PMID 10568034 . S2CID 3649475 .
- ^ Jump up to: а беременный в дюймовый и фон Сарасвати, Пратул Кумар; Сринивасан, MS (2016), Сарасвати, Пратул Кумар; "Микрофоссии с известны , MS ( , Srinivasan . ) Eds 978-3-319-14574-7
- ^ Jump up to: а беременный в Сен Гупта, Барун К. (1982). «Экология бентических фораминиферов» . В Бродхеде, TW (ed.). Foraminifera: Заметки на короткий курс, организованный Ma Buzas и BK Sen Gupta . Исследования по геологии. Тол. 6. Университет Теннесси, кафедра геологических наук. С. 37–50. ISBN 978-0910249058 Полем OCLC 9276403 .
- ^ Jump up to: а беременный в GEMLEBEN, C.; Андерсон, или; Spindler, M. (1989). Современные планктонные фораминиферы . Springer-Verlag. ISBN 978-3-540-96815-3 .
- ^ Грелл, кг (1 января 1979 г.). «Цитогенетические системы и эволюция в фораминифере» . Журнал фораминиферальных исследований . 9 (1): 1–13. Bibcode : 1979jforr ... 9 .... 1g . doi : 10.2113/gsjfr.9.1.1 . ISSN 0096-1191 .
- ^ Jump up to: а беременный в Лекифре, Шарлотта; Бернхард, Джоан М.; Мабилло, Гийом; Филипссон, Хелена Л.; Мейбом, Андерс; Геслин, Эммануэль (1 января 2018 года). «Обзор клеточной ультраструктуры в бентических фораминифере: новые наблюдения роталлидных видов в контексте существующей литературы» . Морская микропалеонтология . 138 : 12–32. Bibcode : 2018marmp.138 ... 12L . doi : 10.1016/j.marmicro.2017.10.005 . HDL : 1912/9530 . ISSN 0377-8398 .
- ^ Domanov, MM (июль 2015 г.). «Естественные 226RA и 232 -й радионуклиды в ксенофиофорах Тихого океана» . Геохимия International . 53 (7): 664–669. Bibcode : 2015geoci..53..664d . doi : 10.1134/s0016702915070034 . ISSN 0016-7029 . S2CID 127121951 .
- ^ Takagi, H., Kimoto, K., Fujiki, T., Saito, H., Schmidt, C., Kucera, M. and Moriya, K. (2019) «Характеризация фотосимбиоза в современных планктонных фораминифера». Biogeosciences , 16 (17). Два : 10.5194/BG-16-3377-2019 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Kucera, M.; Дарлинг, KF (апрель 2002 г.). «Загадочные виды планктонных фораминиферов: их влияние на палеоооооооооооооооооооооооооооооооооооооооооооо для реконструкции». Philos Trans Royal Soc a . 360 (1793): 695–718. Bibcode : 2002rspta.360..695K . doi : 10.1098/rsta.2001.0962 . PMID 12804300 . S2CID 21279683 .
- ^ Jump up to: а беременный Ujiié, Юрика; Кимото, Катсунори; Павловский, Январь (декабрь 2008 г.). «Молекулярные доказательства независимого происхождения современного тризерного планктонного фораминифера от бентических предков». Морская микропалеонтология . 69 (3–4): 334–340. Bibcode : 2008marmp..69..334u . doi : 10.1016/j.marmicro.2008.09.003 .
- ^ Jump up to: а беременный Özdikmen, Hüsein (июнь 2009 г.). ( ) PDF . 4.
- ^ Дубика, Зофия (2019). «Расположение камеры по сравнению с структурой стенки в высокопоставленной филогенетической классификации фораминифер» . Acta Palaeontologica Polonica . 64 doi : 10.4202/app.00564.2018 . ISSN 0567-7920 .
- ^ Симбиоз и гибкость микробиома в кальцификационном бентическом фораминифе большого барьерного рифа
- ^ Характеристика фотосимбиоза в современных планктонных фораминифере - BG
- ^ Достижения в области микробной экологии, Volum 11
- ^ Бернхард, JM; Баузер, С.М. (1999). «Бентические фораминиферы дизоксических отложений: секвестрация хлоропластов и функциональная морфология». Земля-наука обзоров . 46 (1): 149–165. Bibcode : 1999esrv ... 46..149b . doi : 10.1016/s0012-8252 (99) 00017-3 .
- ^ Jump up to: а беременный Goldstein, Susan T. (2003), «Foraminifera: биологический обзор», в Sen Gupta, Barun K. (ed.), Modern Foraminifera , Springer Netherlands, с. 37–55, DOI : 10.1007/0-306-48104 -9_3 , ISBN 978-0-306-48104-8
- ^ Цучия, Масаши; Номаки, Хидетака (1 октября 2021 г.). «Быстрый отклик гигантских протеристических ксенофиофоров (Foraminifera, Rhizaria) на поставку органических веществ на глубине абиссала, выявленное экспериментом in situ с двумя стабильными изотопами» . Deep Sea Research Part I: океанографические исследовательские работы . 176 : 103608. Bibcode : 2021dsri..17603608t . doi : 10.1016/j.dsr.2021.103608 . ISSN 0967-0637 .
- ^ Калвер, Стивен Дж.; Липпс, Джере Х. (2003), Келли, Патриция Х.; Ковалевски, Михал; Хансен, Тор А. (ред.), «Хищничество на фораминифере» , Хищник-Прайские взаимодействия в ископаемом записи , Бостон, Массачусетс: Спрингер США, с. 7–32, doi : 10.1007/978-1-4615- 0161-9_2 , ISBN 978-1-4613-4947-1 , Получено 30 сентября 2020 года
- ^ Moodley, L.; Hess, C. (1 августа 1992 г.). «Толерантность к инфунальным бентическим фораминифера для низких и высоких концентраций кислорода» . Биологический бюллетень . 183 (1): 94–98. doi : 10.2307/1542410 . ISSN 0006-3185 . JSTOR 1542410 . PMID 29304574 .
- ^ Гудэй, AJ; Todo, y.; Uematsu, K.; Китазато, Х. (июль 2008 г.). «Новые органические стены фораминиферы (Protista) из самой глубокой точки океана, Challenger Deep (Западная часть Тихого океана)» . Зоологический журнал Линневого общества . 153 (3): 399–423. doi : 10.1111/j.1096-3642.2008.00393.x .
- ^ Хольцманн, Мария; Гудэй, Эндрю Дж.; Сиеменсма, паром; Павловский, январь (29 октября 2021 г.). «Обзор: пресноводная и почвенная фораминифера-история давно забытых родственников» . Журнал фораминиферальных исследований . 51 (4): 318–331. Bibcode : 2021jforr..51..318h . doi : 10.2113/gsjfr.51.4.318 . ISSN 0096-1191 . S2CID 240240437 .
- ^ Мур, RC; Lalicker, Ag; Фишер, CG (1952). «Ch 2 Foraminifera и Radiolaria». Беспозвоночные окаменелости . МакГроу-Хилл. OCLC 547380 .
- ^ Jump up to: а беременный Хейнс, младший (18 июня 1981 года). Фораминифера . Спрингер. ISBN 978-1-349-05397-1 .
- ^ Лана, С. (2001). «Брицеловая Картерина (фораминифера)». Морская микропалеонтология . 41 (1–2): 97–102. Bibcode : 2001marmp..41 ... 97L . doi : 10.1016/s0377-8398 (00) 00050-5 .
- ^ Jump up to: а беременный Искаженно, аэ; Варлами, ИИ; Спасибо, DV; Карлова, Джорджия; Klets, Ag; Искаженно, va; Saraev, SV; Территориальный, аа; Beliaeav, S. Yu.; Вараксина, IV; Эфимов, как (1 Deceives 2008). Время западной сибирской тарелки Русская геология и геофизика 49 (12): 932–939. Bibcode : 2008king ... 49..932K . doi : 10.1016/j.rgg . ISSN 1068-7971 .
- ^ Foraminifera: История обучения , Университетский колледж Лондон , получен 20 сентября 2007 г.
- ^ Лангер, мистер; Шелк, MTB; Lipps, JH (1997). «Глобальное производство карбоната океана и углекислого газа: роль рифовых фораминиферов» . Журнал фораминиферальных исследований . 27 (4): 271–277. Bibcode : 1997jforr..27..271l . doi : 10.2113/gsjfr.27.4.271 .
- ^ Adl, sm; Симпсон, Агб; Фермер, Массачусетс; Андерсон; и др. (2005). «Новая классификация эукариот более высокого уровня с акцентом на таксономию протистов» . Журнал эукариотической микробиологии . 52 (5): 399–451. doi : 10.1111/j.1550-7408.2005.00053.x . PMID 16248873 . S2CID 8060916 .
- ^ Jump up to: а беременный Павловский, Ян; Хольцманн, Мария; Берни, Седрик; Фарни, Хосе; Гудэй, Эндрю Дж.; Седхаген, Томас; Хабура, Андреа; Баузер, Сэмюэль С. (30 сентября 2003 г.). «Эволюция ранних фораминиферов» . Труды Национальной академии наук . 100 (20): 11494–11498. Bibcode : 2003pnas..10011494p . doi : 10.1073/pnas.2035132100 . ISSN 0027-8424 . PMC 208786 . PMID 14504394 .
- ^ Груссин, Матье; Павловский, Ян; Ян, Зихенг (1 октября 2011 г.). «Байесовская расслабленная оценка часов дивергенции в фораминифере» . Молекулярная филогенетика и эволюция . 61 (1): 157–166. Bibcode : 2011molpe..61..157G . doi : 10.1016/j.ympev.2011.06.008 . ISSN 1055-7903 . PMID 21723398 .
- ^ Сейлахер А. (1 января 2007 г.). «Природа вендобионтов» . Геологическое общество, Лондон, Специальные публикации . 286 (1): 387–397. Bibcode : 2007gslsp.286..387s . doi : 10.1144/sp286.28 . ISSN 0305-8719 . S2CID 128619251 .
- ^ Бобровский, Илья; Надежда, Джанет М.; Авантсов, Андрей; Nettersheim, Benjamin J.; Халманн, Кристиан; Брокс, Джохен Дж. (21 сентября 2018 г.). «Древние стероиды устанавливают эдиакаран ископаемого Дикинсонии как одного из самых ранних животных» . Наука . 361 (6408): 1246–1249. Bibcode : 2018sci ... 361.1246b . doi : 10.1126/science.aat7228 . HDL : 1885/230014 . ISSN 0036-8075 . PMID 30237355 .
- ^ Swinbanks, DD (1 октября 1982 г.). "Piaeodicton: следы инфунальных ксенофиофоров?" Полем Наука . 218 (4567): 47–49. Bibcode : 1982sci ... 218 ... 47 с . doi : 10.1126/science.218.4567.47 . ISSN 0036-8075 . PMID 17776707 . S2CID 28690086 .
- ^ Jump up to: а беременный Левин, Лиза А. (1994). «Палеоэкология и экология ксенофиофоров». Палаис . 9 (1): 32–41. Bibcode : 1994Palai ... 9 ... 32L . doi : 10.2307/3515076 . ISSN 0883-1351 . JSTOR 3515076 .
- ^ Рона, Питер А.; Сейлахер, Адольф; де Варгас, Колумбан; Гудэй, Эндрю Дж.; Бернхард, Джоан М.; Баузер, Сэм; Ветриани, Костантино; Wirsen, Carl O.; Маллино, Лорен; Шеррелл, Роберт; Фредерик Грассл, Дж. (1 сентября 2009 г.). «Палеодиктион Nodosum: живое ископаемое на глубоководном этаже» . Deep Sea Research Часть II: Актуальные исследования в области океанографии . Морская бентическая экология и биоразнообразие: сборник недавних достижений в честь Дж. Фредерика Грассли. 56 (19): 1700–1712. Bibcode : 2009dsrii..56.1700r . doi : 10.1016/j.dsr2.2009.05.015 . ISSN 0967-0645 .
- ^ Гудэй, Эндрю Дж; Хольцманн, Мария; Caulle, Clémence; Гоино, Аурели; Каменская, Ольга; Вебер, Александра А. -t.; Pawlowski, январь (1 марта 2017 г.). «Гигантские протисты (Xenophyophores, Foraminifera) являются исключительно разнообразными в частях абиссала восточной части Тихого океана, лицензированного для исследования полиметаллических узелков» . Биологическое сохранение . 207 : 106–116. Bibcode : 2017bcons.207..106G . doi : 10.1016/j.biocon.2017.01.006 . ISSN 0006-3207 .
- ^ Макилрой, Дункан; Зеленый, или; Brasier, MD (2001). «Палеобиология и эволюция самых ранних агглютинированных фораминиферов: платисольениты, спиросоленты и родственные формы». Летая . 34 (1): 13–29. Bibcode : 2001letha..34 ... 13m . doi : 10.1080/002411601300068170 . ISSN 1502-3931 .
- ^ Jump up to: а беременный Скотт, Дэвид Б.; Медиоли, Франко; Браун, Риган (1 июня 2003 г.). «Foraminifera из кембрия из Новой Шотландии: самый старый многоамбер Foraminifera» . Микропалеонтология . 49 (2): 109–126. doi : 10.2113/49.2.109 . ISSN 1937-2795 .
- ^ Jump up to: а беременный в дюймовый и фон Таппан, Хелен; Loeblich, Alfred R. (1988). «Фараминиферальная эволюция, диверсификация и вымирание». Журнал палеонтологии . 62 (5): 695–714. ISSN 0022-3360 . JSTOR 1305391 .
- ^ Вагнер, Роберт Херман (1983). Канальный мир: Китай, Корея, Япония и Се -Азия . Igme. п. 88. ISBN 978-84-300-9949-8 .
- ^ Голдберг, Уолтер М. (4 октября 2013 г.). Биология рифов и рифовых организмов . Университет Чикагской Прессы. п. 73. ISBN 978-0-226-92537-0 Полем Получено 10 января 2023 года .
- ^ «Фусулиниды | Геоканзас» . Geokansas.ku.edu . Получено 16 мая 2020 года .
- ^ Czaplewski, Джон Дж. "PBDB Navigator" . paleobiodb.org . Получено 16 мая 2020 года .
- ^ Gräfe, Ku (2005). «Бентические фораминифер и палеоэронометры в нижней и средней юрской игре западного баскского кантабрийского бассейна (северная Испания)». Журнал иберийской геологии . 31 (2): 217–233. S2CID 55664447 .
- ^ Jump up to: а беременный Пирсон, Пол (19 ноября 1998 г.). "Природа дебаты" . Природа : 1–3. doi : 10.1038/nature28135 .
- ^ Журнал биоинформатика и биологические идеи, используя множественную подход анализа для реконструкции филогенетических отношений между планктоническими фораминиферами из сильно дивергентных и длина-полиморфных последовательностей SSU RDNA
- ^ Гебхардт, Холгер (1 февраля 1997 г.). «Сеноманский к туронскому фораминифера от Ашаки (NE Нигерия): количественный анализ и палеоэкологическая интерпретация» . Мерашные исследования . 18 (1): 17–36. Bibcode : 1997crres..18 ... 17g . doi : 10.1006/cres.1996.0047 . ISSN 0195-6671 .
- ^ Балди, Каталин; Benkovics, László; Sztanó, Orsolya (1 мая 2002 г.). «Средний миоцен) развитие бассейна в SW Hugingar: история субдианности с Qutantito Peobathytry of Formigera» Международный журнал наук о Земле 91 (3): 490–5 Bibcode : 2002ijeas..91..490b Doi : 10.1007/s053101010026 . ISSN 1437-3 S2CID 129296067
- ^ Робинсон, Марси (2021) Планктические фораминифер из Геологической службы Южного Мэриленда США .
- ^ Зачос, Джеймс С.; Шеклтон, Николас Дж.; Ревено, Джастин С.; Пяковы, Хейко; Цветок, Бенджамин П. (13 апреля 2001 г.). «Климатическая реакция на орбитальное воздействие через олигоценовую границу» (PDF) . Наука . 292 (5515). Американская ассоциация по развитию науки (AAAS): 274–278. Bibcode : 2001sci ... 292..274Z . doi : 10.1126/science.1058288 . ISSN 0036-8075 . PMID 11303100 . S2CID 38231747 .
- ^ Raymo, Me и Lisiecki, Le (2005) «Плиоцено-плейстоценовый стек из 57 глобально распределенных бентических записей Δ18O», палеоооооооооооооооооооооооооооооо , 20 : PA1003.
- ^ Пети, младший; Jouzel, J.; Raynaud, D.; Барков, Ни; Barnola, J.-M.; Базиль, я.; Bender, M.; Chappellaz, J.; Дэвис, М.; Задержка, Г.; Delmotte, M.; Котляков, виртуальная машина; Legrand, M.; Липенков, VY; Лориус, C.; Pépin, L.; Ritz, C.; Saltzman, E.; Stievenard, M. (1999). «История климата и атмосферы за последние 420 000 лет из ледяного ядра Востока, Антарктида» . Природа . 399 (6735). Springer Science and Business Media LLC: 429–436. Bibcode : 1999natur.399..429p . doi : 10.1038/20859 . ISSN 0028-0836 . S2CID 204993577 .
- ^ Диккенс, Гр; Кастильо, мм; Уокер, JCG (1997). «Взрыв газа в последнем палеоцене; имитация эффектов первого порядка массивной диссоциации гидрата океанического метана». Геология . 25 (3): 259–262. Bibcode : 1997geo .... 25..259d . doi : 10.1130/0091-7613 (1997) 025 <0259: abogit> 2.3.co; 2 . PMID 11541226 . S2CID 24020720 .
- ^ «Петм странность» . Realclimate. 2009. Архивировано из оригинала 12 февраля 2016 года . Получено 3 февраля 2016 года .
- ^ Макинхерни, Фа; Wing, S. (2011). «Возмущение углеродного цикла, климата и биосферы с последствиями для будущего» . Ежегодный обзор земли и планетарных наук . 39 : 489–516. Bibcode : 2011Areps..39..489m . doi : 10.1146/annurev-arth-040610-133431 . Архивировано с оригинала 14 сентября 2016 года . Получено 3 февраля 2016 года .
- ^ Zeebe, R.; Зачос, JC; Диккенс, Гр (2009). «Одно только вынуждение углекислого газа, недостаточно для объяснения палеоцен -эоценового теплового максимального потепления». Природа Геонаука . 2 (8): 576–580. Bibcode : 2009natge ... 2..576z . Citeseerx 10.1.1.704.7960 . doi : 10.1038/ngeo578 .
- ^ Марси М Робинсон (2022) Древние микрофоссии являются ключом к будущему восстановлению климата Геологическая служба США .
 Эта статья включает текст из этого источника, который находится в общественном доступе .
Эта статья включает текст из этого источника, который находится в общественном доступе .
- ^ Бабила, Тали Л.; Пенман, Дональд Э.; Standish, Christopher D.; Дубрава, Моника; Bralower, Timothy J.; Робинсон, Марси М.; Самостоятельная работа, Джин М.; Speijer, Robert P.; Стассен, Питер; Фостер, Гэвин Л.; Зачос, Джеймс С. (18 марта 2022 г.). «Поверхностное потепление океана и подкисление, обусловленное быстрым высвобождением углерода, предшествует максимуму палеоцен-эоценового термического» . Наука достижения . 8 (11). Американская ассоциация по развитию науки (AAAS): EABG1025. Бибкод : 2022Scia .... 8G1025B . doi : 10.1126/sciadv.abg1025 . ISSN 2375-2548 . PMC 8926327 . PMID 35294237 . S2CID 247498325 .
- ^ Австралия, C \ = Au \; O \ = Правительство Австралии \; ou \ = Geoscience (15 мая 2014 г.). «Биостратиграфия» . Ga.gov.au. Получено 20 июля 2020 года .
{{cite web}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Кушман, Джозеф А.; Ellisor, Alva C. (1 января 1945 г.). «Фараминиферальная фауна формирования Анахуака». Журнал палеонтологии . 19 (6): 545–572. JSTOR 1299203 .
- ^ Зачос, JC; Pagani, M.; Sloan, L.; Томас, E.; Billups, K. (2001). «Тенденции, ритмы и аберрации в глобальном климате, 65 млн. Лет» (PDF) . Наука . 292 (5517): 686–693. Bibcode : 2001sci ... 292..686Z . doi : 10.1126/science.1059412 . PMID 11326091 . S2CID 2365991 .
- ^ Кюл, Нина; Дольше, г.; Thoms, S.; Невропейский, ЖЖ; Рейкарт, GJ; Приложение, J. (апрель 2017 г.). «Исследование фораминиферала SR/CA в качестве нового прокси -карбонатной системы» (PDF) . Gechemica et cosmomochimica acta . 202 : 374–386. Код BIB : 2017Gecoa.202..374K . doi : 10,1016/j.gc.2016,022 . HDL : 1874/380107 .
- ^ Брэнсон, Оскар; Редферн, Саймон в; Tyliszczak, Tolek; Садеков, Алекси; Лангер, Джеральд; Кимото, Катсунори; Элдерфилд, Генри (декабрь 2013 г.). «Координация Mg в кальците на фораминице» . Земля и планетарные научные письма . 383 : 134–141. BIBCODE : 2013E & PSL.383..134B . doi : 10.1016/j.epsl.2013.09.037 .
- ^ Мисра, с.; Froelich, PN (26 января 2012 г.). «Изотопная история лития кенозойской морской воды: изменения силикатного выветривания и обратного выветривания» . Наука . 335 (6070): 818–823. Bibcode : 2012sci ... 335..818m . doi : 10.1126/science.1214697 . PMID 22282473 . S2CID 42591236 .
- ^ Хемминг, нг; Хансон, GN (январь 1992). «Изотопный состав бора и концентрация в современных морских карбонатах». Geochimica et Cosmochimica Acta . 56 (1): 537–543. Bibcode : 1992gecoa..56..537h . doi : 10.1016/0016-7037 (92) 90151-8 .
- ^ Boardman, Rs; Читам, ах; Rowell, AJ (1987). Ископаемые беспозвоночные . Уайли. ISBN 978-0865423022 .
- ^ Джонс, RW (1996). Микропалеонтология в разведке нефти . Кларендон Пресс. ISBN 978-0-19-854091-5 .
- ^ Макнил, DH; Issler, Dr; Сноудон, Л.Р. (1996). Изменение цвета, тепловое зрелость и диагенез захоронения у ископаемых фораминифер . Геологическая служба канадского бюллетеня. Тол. 499. Геологическая служба Канады. ISBN 978-0-660-16451-9 .
- ^ Уилкинсон, Ян П.; Уильямс, Марк; Янг, Джереми Р.; Кук, Саманта Р.; Фулфорд, Майкл Г.; Лотт, Грэм К. (1 августа 2008 г.). «Применение микрофоссилий при оценке происхождения мела, используемого при изготовлении римской мозаики в Силчестере» . Журнал археологической науки . 35 (8): 2415–2422. Bibcode : 2008jarsc..35.2415w . doi : 10.1016/j.jas.2008.03.010 . ISSN 0305-4403 .
Внешние ссылки
[ редактировать ]- Общая информация
- музея палеонтологии Университета Калифорнийского университета На сайте знакомство с Foraminifera
- Исследователи из Университета Южной Флориды разработали систему, использующую Foraminifera для мониторинга среды коралловых рифов, архивные 15 января 2021 года на машине Wayback
- Университетский колледж Лондонского микропалеонтологии имеет обзор фораминифер, в том числе многие высококачественные SEMS
- Иллюстрированный глоссарий терминов, используемые в фораминиферальных исследованиях, архивировав 21 июня 2012 года в The Wayback Machine -это глоссарий Lukas Hottinger, опубликованный в OA e-Journal "Carnets de Géologie-записные книжки по геологии" Архивировали 29 июня 2017 года на машине Wayback Machine
- Информация о Foraminifera Martin Langer's Micropaleontology страница
- Бентистская информация о Фораминифера из летней школы палеоклиматологии 2005 года 2005 года
- Онлайн-обратные книги
- Иллюстрированный глоссарий терминов, используемых в фораминиферальных исследованиях Лукаса Хоттингера (альтернативная версия той, которая опубликована в «Carnets de Géologie - ноутбуки по геологии», архивировав 29 июня 2017 года на машине Wayback )
- Ресурсы
- pforams@mikrotax - онлайн -база данных, подробно описывающая таксономию планктонных фораминиферов
- Проект Star*Sand (часть Micro*Scope ) - это кооперативная база данных информации о Foraminifera
- 3D-модели FORAMS, сгенерированные рентгеновской томографией
- Chronos имеет несколько ресурсов Foraminifera , в том числе страница поиска таксонов и раздел Micro-Paleo NB Большая часть этого контента теперь включена в веб-сайт pforams@mikrotax
- Eforams - это веб -сайт, ориентированный на фораминифера и моделирование фораминиферальных оболочек
- Галерея Foraminifera проиллюстрировал каталог недавних и ископаемых фораминиферов по роду и местности
- « Фораминифера » . NCBI таксономия браузер . 29178.