Фораминиферы
| Фораминиферы Временной диапазон: Последний эдиакарский язык – недавний
| |
|---|---|

| |
| Живой аммиак тепловатый (Rotaliida) | |
| Научная классификация | |
| Домен: | Эукариоты |
| Клэйд : | потогонные средства |
| Клэйд : | САР |
| Тип: | Ретария |
| Подтип: | Фораминиферы д'Орбиньи , 1826 г. |
| Подразделения | |
|
« Моноталамея »
неопределенное место | |
Фораминиферы ( / f ə ˌ r æ m ə ˈ n ɪ f ə r ə / fə - RAM -ə- NIH -fə-rə ; по латыни «носители дыр»; неофициально называемые « форамы ») — одноклеточные организмы , члены представителя типа или класса ризарианских характеризующихся простейших, струящейся гранулярной эктоплазмой для ловли пищи и другое использование; и обычно внешняя оболочка (называемая « тестом ») различных форм и материалов. раковины Хитиновые (встречающиеся у некоторых простых родов, в частности Textularia ) считаются наиболее примитивным типом. Большинство фораминифер являются морскими, большинство из которых обитают на отложениях морского дна или внутри них (т. е. являются бентосными , причем виды разного размера играют роль в макробентосе , мейобентосе и микробентосе ), [ 2 ] тогда как меньшее их количество плавает в толще воды на различных глубинах (т. е. является планктоном ), которые относятся к подотряду Globigerinina . [ 3 ] Меньше известно из пресноводных [ 4 ] или солоноватый [ 5 ] условиях, а некоторые очень немногие (неводные) виды почв были идентифицированы посредством молекулярного анализа малых субъединиц рибосомальной ДНК . [ 6 ] [ 7 ]
Фораминиферы обычно образуют раковину или раковину, которая может иметь одну или несколько камер, некоторые из которых имеют весьма сложную структуру. [ 8 ] Эти раковины обычно состоят из карбоната кальция ( CaCO
3 ) или агглютинированные частицы осадка. Выявлено более 50 000 видов, оба из которых являются живыми (6 700–10 000). [ 9 ] [ 10 ] и ископаемые (40 000). [ 11 ] [ 12 ] Обычно они имеют размер менее 1 мм, но некоторые из них намного крупнее, самые крупные виды достигают 20 см. [ 13 ]
В современном научном английском языке термин фораминифера происхождения этого слова имеет как единственное, так и множественное число (независимо от латинского ) и используется для описания одного или нескольких экземпляров или таксонов: его использование в единственном или множественном числе должно определяться из контекста. Фораминиферы часто неофициально используются для описания группы, и в этих случаях обычно пишутся строчными буквами. [ 14 ]
История обучения
[ редактировать ]Самое раннее известное упоминание о фораминиферах принадлежит Геродоту , который в V веке до нашей эры отметил, что они составляют скалу, образующую Великую пирамиду в Гизе . Сегодня они признаны представителями рода Nummulites . Страбон в I веке до нашей эры заметил те же фораминиферы и предположил, что это остатки чечевицы, оставленные рабочими, строившими пирамиды. [ 15 ]
Роберт Гук наблюдал фораминиферу под микроскопом, как описано и проиллюстрировано в его книге «Микрография» 1665 года :
Я пробовал несколько маленьких и одиночных увеличительных стекол и случайно рассматривал комочек белого песка, когда увидел одно из зерен точной формы и в виде венка, похожего на ракушку [...]. Я рассматривал его каждый раз. с помощью лучшего микроскопа и обнаружил, что с обеих сторон и по краям он напоминает раковину маленькой водяной улитки с плоской спиральной раковиной [...] [ 16 ]
Антони ван Левенгук описал и проиллюстрировал раковины фораминифер в 1700 году, описав их как крошечные моллюски; в его иллюстрации можно узнать Эльфидиум . [ 17 ] Ранние исследователи отнесли фораминиферы к роду Nautilus , отметив их сходство с некоторыми головоногими моллюсками . признал В 1781 году Лоренц Шпенглер , что у фораминифер есть отверстия в перегородках, что в конечном итоге дало группе такое название. [ 18 ] Шпенглер также отметил, что перегородки фораминифер имеют дугу, противоположную перегородкам наутили, и что у них отсутствует нервная трубка. [ 19 ]

Альсид д'Орбиньи в своей работе 1826 года считал их группой мелких головоногих моллюсков и отмечал их странную морфологию, интерпретируя псевдоподии как щупальца и отмечая сильно уменьшенную (на самом деле отсутствующую) голову. [ 20 ] Он назвал группу фораминиферами , или «носителями дырок», поскольку у членов группы были отверстия в перегородках между отсеками в их раковинах, в отличие от наутили или аммонитов . [ 14 ]
Простейшая природа фораминифер впервые была признана Дюжарденом в 1835 году. [ 18 ] Вскоре после этого, в 1852 году, д'Орбиньи разработал схему классификации, признав 72 рода фораминифер, которые он классифицировал на основе формы раковины - схема, вызвавшая резкую критику со стороны коллег. [ 17 ]
В монографии Х.Б. Брейди 1884 г. описаны находки фораминифер, сделанные Челленджера экспедицией . Брейди выделил 10 семейств с 29 подсемействами, не обращая особого внимания на стратиграфический диапазон; его таксономия подчеркивала идею о том, что несколько разных признаков должны разделять таксономические группы, и, как таковая, помещала агглютинированные и известковые роды в тесное родство.
Эта общая схема классификации сохранялась до работы Кушмана в конце 1920-х годов. Кушман рассматривал состав стенок как самый важный признак в классификации фораминифер; его классификация получила широкое признание, но также вызвала критику со стороны коллег за то, что она «биологически необоснованна».
Геолог Ирен Креспин провела обширные исследования в этой области, опубликовав около девяноста статей, в том числе заметные работы по фораминиферам, в качестве единственного автора, а также более двадцати в сотрудничестве с другими учеными. [ 21 ]
Тем не менее схема Кушмана оставалась доминирующей схемой классификации до тех пор, пока Таппан и Леблих не разработали классификацию 1964 года, которая поместила фораминиферы в общие группы, используемые до сих пор, на основе микроструктуры тестовой стенки. [ 17 ] Эти группы по-разному перемещались в соответствии с разными схемами классификации более высокого уровня. Использование Павловским (2013) молекулярной систематики в целом подтвердило группировки Таппана и Леблиха, причем некоторые из них оказались полифилетическими или парафилетическими; эта работа также помогла выявить взаимоотношения более высокого уровня между основными группами фораминифер. [ 22 ]
Таксономия
[ редактировать ]
| |||||||||||||||||||||||||||||||||||||||||||||
| Филогения фораминифер по данным Pawlowski et al. 2013. [ 22 ] Моноталамидные отряды Astrorhizida и Allogromiida являются парафилетическими. |
Таксономическое положение фораминифер менялось со времен Шульца в 1854 году. [ 23 ] которые назывались отрядом Foraminiferida. Леблих (1987) и Таппан (1992) отнесли фораминиферы к классу. [ 24 ] как это сейчас принято считать.
Фораминиферы обычно относят к простейшим . [ 25 ] [ 26 ] [ 27 ] или в аналогичном протоктисте или протистском королевстве . [ 28 ] [ 29 ] Существуют убедительные доказательства, основанные главным образом на молекулярной филогенетике , их принадлежности к основной группе простейших, известной как ризарии . [ 25 ] До признания эволюционных связей между представителями Rhizaria фораминиферы обычно группировались с другими амебоидами как тип Rhizopodea (или Sarcodina) в классе Granuloreticulosa.
Ризарии являются проблематичными, поскольку их часто называют «супергруппой», вместо того, чтобы использовать установленный таксономический ранг, такой как тип . Кавальер-Смит определяет Ризарию как инфрацарство внутри царства простейших. [ 25 ]
Некоторые систематики относят фораминиферы к отдельному типу, ставя их в один ряд с амебоидными саркодинами, к которым они были отнесены.
Хотя молекулярные данные еще не подтверждены морфологическими коррелятами, молекулярные данные убедительно свидетельствуют о том, что фораминиферы тесно связаны с Cercozoa и Radiolaria , оба из которых также включают амебоидов со сложными панцирями; эти три группы составляют Ризарию. [ 26 ] Однако точные взаимоотношения форумов с другими группами и друг с другом до сих пор не совсем ясны. Фораминиферы тесно связаны с раковинными амебами . [ 30 ]
| Таксономия от Михалевича 2013 г. [ 31 ] |
|---|
Анатомия
[ редактировать ]
Самым поразительным аспектом большинства фораминифер являются их твердые панцири или панцири. Они могут состоять из одной или нескольких камер и могут состоять из белка, частиц осадка, кальцита, арагонита или (в одном случае) кремнезема. [ 24 ] У некоторых фораминифер раковины полностью отсутствуют. [ 32 ] В отличие от других панцирно-секретирующих организмов, например моллюсков или кораллов , раковины фораминифер расположены внутри клеточной мембраны , внутри протоплазмы . Органеллы клетки расположены внутри отсека(ов) теста, а отверстие(я) теста позволяют переносить материал из псевдоподий во внутреннюю клетку и обратно. [ 33 ]
Клетка фораминифер разделена на зернистую эндоплазму и прозрачную эктоплазму, из которой псевдоподиальная сеть может выходить через одно отверстие или через множество перфораций в панцире. Для отдельных ложноножек характерно наличие мелких гранул, струящихся в обоих направлениях. [ 34 ] Фораминиферы уникальны тем, что имеют гранулоретикулезные псевдоподии ; то есть их псевдоподии под микроскопом кажутся зернистыми; эти псевдоподии часто удлинены, могут разделяться и вновь соединяться друг с другом. Их можно выдвигать и убирать в соответствии с потребностями ячейки. Локоподы используются для передвижения, закрепления, выделения, построения тестов и захвата пищи, состоящей из мелких организмов, таких как диатомовые водоросли или бактерии. [ 35 ] [ 33 ]
Помимо раковин, клетки фораминифер поддерживаются цитоскелетом из микротрубочек, которые расположены рыхло и не имеют структуры, наблюдаемой у других амебоидов. Форамы развили специальные клеточные механизмы для быстрой сборки и разборки микротрубочек, что позволяет быстро формировать и сокращать удлиненные псевдоподии. [ 24 ]
Гамонт (половая форма) фораминифер обычно имеет только одно ядро, тогда как агамонт (бесполая форма) имеет тенденцию иметь несколько ядер. По крайней мере, у некоторых видов ядра диморфны: соматические ядра содержат в три раза больше белка и РНК, чем генеративные ядра. Однако ядерная анатомия кажется весьма разнообразной. [ 36 ] Ядра не обязательно ограничены одной камерой у многокамерных видов. Ядра могут иметь сферическую форму или иметь много долей. Ядра обычно имеют диаметр 30-50 мкм. [ 37 ]
Некоторые виды фораминифер имеют внутри клеток большие пустые вакуоли; их точная цель неясна, но предполагается, что они действуют как резервуар нитратов. [ 37 ]
Митохондрии распределены по клетке равномерно, хотя у некоторых видов они концентрируются под порами и вокруг внешнего края клетки. Было высказано предположение, что это адаптация к среде с низким содержанием кислорода. [ 37 ]
несколько видов ксенофиофоров Было обнаружено, что имеют необычно высокие концентрации радиоактивных изотопов внутри своих клеток, одни из самых высоких среди всех эукариот. Цель этого неизвестна. [ 38 ]
Экология
[ редактировать ]
Современные фораминиферы — это преимущественно морские организмы, но живые особи были обнаружены в солоноватой пресной воде. [ 34 ] и даже наземные среды обитания. [ 7 ] Большинство видов донные , еще 50 морфовидов — планктонные . [ 35 ] Однако это количество может отражать лишь часть фактического разнообразия, поскольку многие генетически различные виды могут быть морфологически неотличимы. [ 40 ]
Бентосные фораминиферы обычно встречаются в мелкозернистых отложениях, где они активно перемещаются между слоями; однако многие виды обитают на твердых породах, прикреплены к морским водорослям или сидят на поверхности отложений. [ 24 ]
Большинство планктонных фораминифер встречается в globigerinina , линии внутри rotaliida . [ 22 ] Однако, по крайней мере, еще одна сохранившаяся линия роталиид, Neogallitellia , по-видимому, независимо развила планктонный образ жизни. [ 41 ] [ 42 ] Кроме того, было высказано предположение, что некоторые ископаемые фораминиферы юрского периода, возможно, также независимо развили планктонный образ жизни и могут быть представителями Robertinida. [ 43 ]
Ряд отверстий, как донных, так и планктонных, [ 44 ] [ 45 ] одноклеточные водоросли являются эндосимбионтами из различных линий, таких как зеленые водоросли , красные водоросли , золотые водоросли , диатомовые водоросли и динофлагелляты . [ 35 ] Эти миксотрофные фораминиферы особенно распространены в бедных питательными веществами океанических водах. [ 46 ] Некоторые отверстия являются клептопластическими , сохраняя хлоропласты из проглоченных водорослей для проведения фотосинтеза . [ 47 ]
Большинство фораминифер гетеротрофны, питаются более мелкими организмами и органическим веществом; некоторые более мелкие виды специализируются на питании фитодетритом , тогда как другие специализируются на потреблении диатомовых водорослей. Некоторые донные образования образуют питающиеся кисты, используя псевдоподии для инцистирования внутри отложений и органических частиц. [ 24 ] Некоторые фораминиферы охотятся на мелких животных, таких как копеподы или кумовые ; некоторые отверстия даже охотятся на других, просверливая отверстия в яйцах своей добычи. [ 48 ] Было предложено выращивать бактерии в одной группе, ксенофиофоры, в ходе своих испытаний, хотя исследования не смогли найти подтверждения этой гипотезе. [ 49 ] В этой группе также распространено суспензионное кормление , и, по крайней мере, некоторые виды могут использовать растворенный органический углерод . [ 24 ]
Некоторые виды форамов являются паразитами , заражая губки, моллюски, кораллы и даже другие фораминиферы. Паразитарные стратегии различаются; некоторые действуют как эктопаразиты, используя свои псевдоподии, чтобы украсть пищу у хозяина, в то время как другие прорываются сквозь панцирь или стенки тела хозяина, чтобы питаться его мягкими тканями. [ 24 ]
Сами фораминиферы поедаются множеством более крупных организмов, включая беспозвоночных, рыб, куликов и других фораминифер. Однако было высказано предположение, что в некоторых случаях хищников может больше интересовать кальций из раковин форамов, чем сами организмы. Известно, что несколько видов водных улиток избирательно питаются фораминиферами, часто даже отдавая предпочтение отдельным видам. [ 50 ]
Было обнаружено, что некоторые донные фораминиферы способны выживать в бескислородных условиях более 24 часов, что указывает на их способность к избирательному анаэробному дыханию . Это интерпретируется как адаптация к выживанию в изменяющихся кислородных условиях вблизи границы раздела осадок-вода. [ 51 ]
Фораминиферы встречаются в самых глубоких частях океана, таких как Марианская впадина , включая Бездну Челленджера , самую глубокую из известных частей. На этих глубинах, ниже глубины компенсации карбонатов , карбонат кальция из тестов растворяется в воде из-за экстремального давления. Таким образом, фораминиферы, обнаруженные в Бездне Челленджера, не имеют карбонатного теста, а вместо этого содержат органический материал. [ 52 ]
Неморские фораминиферы традиционно игнорировались в исследованиях форумов, но недавние исследования показывают, что они значительно более разнообразны, чем считалось ранее. Известно, что они населяют разрозненные экологические ниши, включая мхи , реки, озера и пруды, водно-болотные угодья, почвы, торфяные болота и песчаные дюны. [ 53 ]
Воспроизведение
[ редактировать ]Обобщенный жизненный цикл фораминифер включает чередование гаплоидных и диплоидных поколений, хотя по форме они в основном схожи. [ 23 ] [ 54 ] Гаплоид или гамонт изначально имеет одно ядро и делится, образуя многочисленные гаметы , которые обычно имеют два жгутика . Диплоид или агамонт многоядерный и после мейоза делится с образованием новых гамонтов. Множественные циклы бесполого размножения между половыми поколениями нередки для донных форм. [ 34 ]

Фораминиферы демонстрируют морфологический диморфизм, связанный с их репродуктивным циклом. Гамонт, или гаплоидная форма, размножающаяся половым путем, является мегалосферной , то есть ее пролокулюс , или первая камера, пропорционально велика. Гамонт также известен как А. форма Гамонты, несмотря на то, что они обычно имеют более крупные пролокулы, обычно имеют меньший общий диаметр раковины, чем агамонты.
Достигнув зрелости, гамонт делится митозом, образуя тысячи гамет, которые также являются гаплоидными. Все эти гаметы имеют полный набор органелл и выбрасываются из теста в окружающую среду, оставляя тест неповрежденным. Гаметы не дифференцируются на сперму и яйцеклетку , и любые две гаметы одного вида обычно могут оплодотворять друг друга.

Когда две гаметы объединяются, они образуют диплоидную многоядерную клетку, известную как агамонт, или B. форма В отличие от гамонта, агамонт имеет микросферическую форму с пропорционально небольшой первой камерой, но обычно с большим общим диаметром и большим количеством камер. Агамонт - фаза бесполого размножения фораминифер; по достижении взрослой жизни протоплазма полностью покидает тест и делится свою цитоплазму мейотически посредством множественного деления с образованием ряда гаплоидных потомков. Затем это потомство начинает формировать свою первую мегасферную камеру, прежде чем рассеяться.
В некоторых случаях гаплоидное потомство может созреть в мегалосферную форму, которая затем размножается бесполым путем, давая еще одно мегалосферное, гаплоидное потомство. В этом случае первая мегасферная форма называется шизонтом или формой А 1 , а вторая — гамонтом или формой А 2 .

В более прохладной и глубокой воде созревание и размножение происходят медленнее; эти условия также приводят к увеличению размеров отверстий. Формы A всегда кажутся гораздо более многочисленными, чем формы B , вероятно, из-за меньшей вероятности встречи двух гамет друг с другом и успешного объединения. [ 55 ] [ 33 ]
Вариации репродуктивного режима
[ редактировать ]Репродуктивные стратегии разных групп фораминифер весьма разнообразны.
У одногнездных видов форма А и форма В. все еще присутствуют Как и в микросферической морфе многокамерных отверстий, форма B , размножающаяся бесполым путем , крупнее формы A , размножающейся половым путем .
Форамы семейства Spirillinidae имеют амебоидные гаметы, а не жгутиковые. Остальные аспекты размножения этой группы в целом аналогичны таковым у других групп форумов.
Известковая спириллинида Patellina corrugata имеет несколько иную репродуктивную стратегию, чем большинство других фораминифер. , размножающаяся бесполым путем, Форма B образует кисту, окружающую всю клетку; затем он делится внутри этой кисты, и ювенильные клетки поглощают кальцит родительского теста, образуя первую камеру своего собственного теста. Эти формы А по мере взросления собираются в группы до девяти особей; затем они образуют защитную кисту вокруг всей группы. Внутри этой кисты происходит гаметогенез , в результате чего образуется очень небольшое количество гамет. Личинки формы B образуются внутри цисты; любые ядра, не связанные с клетками, потребляются в пищу развивающимся личинкам. Пателлина в форме А Сообщается, что раздельнополая , причем полы называются «плюс» и «минус»; эти полы различаются количеством ядер: форма «плюс» имеет три ядра, а форма «минус» - четыре ядра. Форма B снова больше A. формы [ 33 ] [ 55 ] [ 48 ]
Тесты
[ редактировать ]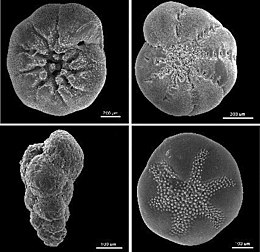
| Часть серии, посвященной |
| Биоминерализация |
|---|
 |
фораминифер Тесты служат для защиты внутреннего организма. Благодаря своей в целом твердой и прочной конструкции (по сравнению с другими протистами) раковины фораминифер являются основным источником научных знаний об этой группе.
Отверстия в тесте, позволяющие цитоплазме выходить наружу, называются апертурами. [ 56 ] Первичное отверстие, ведущее наружу, у разных видов принимает множество различных форм, включая, помимо прочего, округлую , серповидную, щелевидную, покрытую капюшоном, лучистую (звездообразную), дендритную (ветвящуюся). Некоторые фораминиферы имеют «зубчатые», фланцевые или губчатые первичные отверстия. Основная апертура может быть только одна или несколько; если их несколько, они могут быть сгруппированными или экваториальными. Помимо основного отверстия у многих фораминифер имеются дополнительные отверстия. Они могут образовываться как реликтовые отверстия (первичные отверстия более ранней стадии роста) или как уникальные структуры.
Форма раковины сильно различается у разных фораминифер; они могут быть однокамерными (однокамерными) или многокамерными (многокамерными). В многокамерных формах по мере роста организма добавляются новые камеры. Широкое разнообразие морфологии тестовых клеток обнаружено как в однокамерных, так и в многокамерных формах, включая, среди прочего, спиральную, серийную и миолилиновую формы. [ 33 ]
Многие фораминиферы демонстрируют диморфизм в своих раковинах с мегалосферными и микросферическими особями. Эти названия не следует воспринимать как относящиеся к размеру всего организма; скорее, они относятся к размеру первой камеры, или пролокула . Пробы в виде окаменелостей известны еще в эдиакарский период. [ 57 ] и многие морские отложения состоят в основном из них. Например, известняк, из которого состоят пирамиды Египта, почти полностью состоит из нуммулитовых бентосных фораминифер. [ 58 ] Подсчитано, что рифовые фораминиферы производят около 43 миллионов тонн карбоната кальция в год. [ 59 ]
Генетические исследования без тестов идентифицировали голую амебу Reticulomyxa и своеобразные ксенофиофоры как фораминиферы. Несколько других амебоидов производят ретикулезные ложноножки, и раньше их классифицировали вместе с отверстиями как Granuloreticulosa, но они больше не считаются естественной группой, и большинство из них теперь отнесены к Cercozoa. [ 60 ]
Эволюционная история
[ редактировать ]Молекулярные часы показывают, что кронная группа фораминифер, вероятно, возникла в неопротерозое , между 900 и 650 миллионами лет назад; это время согласуется с неопротерозойскими окаменелостями близкородственных нитевидных амеб . Поскольку окаменелости фораминифер не были обнаружены до самого конца эдиакарского периода , вполне вероятно, что у большинства этих протерозойских форм не было панцирей с твердым панцирем. [ 61 ] [ 62 ]
Из-за неминерализованных форм « аллогромииды » не имеют летописи окаменелостей. [ 61 ]

Было высказано предположение, что загадочные вендозои эдиакарского периода представляют собой ископаемые ксенофиофоры . [ 63 ] Однако открытие диагенетически измененных C 27 стеринов , связанных с останками дикинсонии, поставило под сомнение эту идентификацию и позволило предположить, что это может быть животное. [ 64 ] Другие исследователи предположили, что неуловимый след ископаемого Палеодиктиона и его родственников может представлять собой ископаемый ксенофиофор. [ 65 ] и отметил сходство существующего ксенофиофора Occultammina с ископаемым; [ 66 ] однако современные примеры Палеодиктиона не смогли прояснить этот вопрос, и след может попеременно представлять собой нору или стеклянную губку. [ 67 ] Подтверждением этому является сходство среды обитания живых ксенофиофоров с предполагаемой средой обитания ископаемых графоглиптидов; однако большой размер и регулярность многих графоглиптидов, а также очевидное отсутствие ксенофий в их окаменелостях ставят под сомнение такую возможность. [ 66 ] По состоянию на 2017 год никаких определенных окаменелостей ксенофиофоров обнаружено не было. [ 68 ]
Фораминиферы-испытатели имеют отличную летопись окаменелостей на протяжении всего фанерозоя . Самые ранние известные определенные фораминиферы появляются в летописи окаменелостей ближе к самому концу эдиакарского периода; все эти формы имеют агглютинирующие реакции и являются однокамерными. К ним относятся такие формы, как платизолениты и спиросолениты . [ 69 ] [ 57 ] Однокамерные фораминиферы продолжали диверсифицироваться на протяжении всего кембрия. Некоторые часто встречающиеся формы Ammodiscus , Glomospira , Psammosphera и включают Turritellella ; все эти виды агглютинированы. Они составляют часть Ammodiscina , линии спириллинид , которая до сих пор содержит современные формы. [ 70 ] [ 22 ] Позже спириллиниды разовьют многокамерность и кальцитовые раковины, причем первые такие формы появились в триасе ; группа не заметила незначительного влияния на разнообразие из-за вымирания K-Pg . [ 71 ]
Самые ранние многокамерные фораминиферы представляют собой агглютинированные виды и появляются в летописи окаменелостей в среднем кембрийском периоде. Из-за плохой сохранности их нельзя однозначно отнести к какой-либо крупной форумной группе. [ 70 ]

Самые ранние известные фораминиферы с известковыми стенками — это Fusulinids , которые появляются в летописи окаменелостей в лландоверскую эпоху раннего силура . Самые ранние из них были микроскопическими, плоскоспирально закрученными и эволюционирующими; более поздние формы развили разнообразие форм, включая линзовидные, шаровидные и удлиненные формы в форме риса. [ 72 ] [ 73 ]
Более поздние виды фузулинид выросли до гораздо больших размеров, некоторые формы достигали 5 см в длину; как сообщается, некоторые экземпляры достигают 14 см в длину, что делает их одними из крупнейших фораминифер, сохранившихся или вымерших. Фузулиниды - самая ранняя линия фораминифер, которая, как полагают, развила симбиоз с фотосинтезирующими организмами. Окаменелости фузулинид обнаружены на всех континентах, кроме Антарктиды ; своего наибольшего разнообразия они достигли в визейскую эпоху каменноугольного периода . Затем разнообразие группы постепенно уменьшалось, пока окончательно не вымерло во время пермо-триасового вымирания . [ 33 ] [ 71 ] [ 74 ]
В турнейскую эпоху каменноугольного периода Miliolid foraminifera впервые появились в летописи окаменелостей, отделившись от спириллинид в пределах Tubothalamea . Милиолиды понесли около 50% потерь как во время пермо-триасового, так и во время K-Pg вымирания, но дожили до наших дней. Некоторые ископаемые милилиолиды достигали до 2 см в диаметре. [ 71 ]

Самые ранние известные окаменелости лагенидов появляются в московскую эпоху каменноугольного периода. Не видя особого эффекта из-за пермо-триасового или K-Pg вымирания, группа со временем диверсифицировалась. Вторичные однокамерные таксоны возникли в юрском и меловом периодах.
Самые ранние окаменелости инволютинид появляются в перми; эта линия диверсифицировалась на протяжении мезозоя Евразии, прежде чем, по-видимому, исчезла из летописи окаменелостей после сеноманско-туронского океанического бескислородного события . Существующую группу planispirillinidae отнесли к инволютинидам, но это остается предметом дискуссий. [ 75 ] [ 71 ]
Робертиниды эпоху впервые появляются в летописи окаменелостей в анизийскую триаса. Группа оставалась с низким разнообразием на протяжении всей своей истории окаменелостей; все ныне живущие представители принадлежат к отряду Robertinidae , впервые появившемуся в палеоцене . [ 71 ]
Первые определенные окаменелости роталиид не появляются в летописи окаменелостей до плинсбахской эпохи юрского периода, следующей за триасово-юрским событием . [ 76 ] Разнообразие группы оставалось низким до последствий сеноманско-туронского события, после которого в группе произошла быстрая диверсификация. Из этой группы планктонная Globigerinina — первая известная группа планктонных форам — впервые появляется после тоарского оборота ; группа понесла тяжелые потери как во время вымирания K-Pg, так и во время эоцен-олигоценового вымирания , но остается существующей и разнообразной по сей день. [ 71 ] Дополнительная эволюция планктонного образа жизни произошла в миоцене или плиоцене, когда роталииды Neogallitellia самостоятельно развили планктонный образ жизни. [ 41 ] [ 42 ]
Палеонтологические приложения
[ редактировать ]Умирающие планктонные фораминиферы постоянно опадают на морское дно в огромных количествах, их минерализованные частицы сохраняются в виде окаменелостей в накапливающихся осадках . Начиная с 1960-х годов, и в основном под эгидой программ глубоководного бурения , океанского бурения и международных программ океанского бурения, а также в целях разведки нефти, передовые методы глубоководного бурения позволили извлечь керны отложений, содержащие окаменелости фораминифер. . [ 77 ] Практически неограниченное количество этих тестов окаменелостей и относительно высокоточные модели контроля возраста, доступные для кернов, позволили создать исключительно высококачественную летопись окаменелостей планктонных фораминифер, относящихся к середине юрского периода , и представляют собой беспрецедентные данные для ученых, проверяющих и документирующих. эволюционный процесс. [ 77 ] Исключительное качество летописи окаменелостей позволило составить впечатляюще подробную картину взаимоотношений видов на основе окаменелостей, во многих случаях впоследствии подтвержденную независимо посредством молекулярно-генетических исследований на дошедших до нас образцах. [ 78 ]
Поскольку определенные типы фораминифер встречаются только в определенных средах, их окаменелости можно использовать для выяснения типа среды, в которой откладывались древние морские отложения; такие условия, как соленость, глубина, кислородные условия и условия освещенности, можно определить на основе различных предпочтений среды обитания различных видов форм. Это позволяет работникам отслеживать изменение климата и условий окружающей среды с течением времени, собирая информацию о присутствующих фораминиферах. [ 79 ]
В других случаях относительная пропорция окаменелостей планктонных и бентосных фораминифер, обнаруженных в породе, может использоваться в качестве показателя глубины данного места, когда откладывались породы. [ 80 ]


По крайней мере, с 1997 года палеоцен-эоценовый тепловой максимум (PETM) исследуется как аналогия для понимания последствий глобального потепления и массивного поступления углерода в океан и атмосферу, включая закисление океана . [ 85 ] Сегодня человечество выбрасывает в атмосферу около 10 Гт углерода (около 37 Гт CO2-экв.) в год, и примерно через тысячу лет выделит такое же количество, что и ПЭТМ. Основное отличие состоит в том, что во время PETM планета была свободна ото льда, поскольку пролив Дрейка еще не открылся Центральноамериканский морской путь . и не закрылся [ 86 ] Хотя в настоящее время PETM обычно считается примером глобального потепления и массовых выбросов углерода, причина, детали и общее значение этого события остаются неясными. [ 87 ] [ 88 ] [ 89 ] [ 90 ]

Фораминиферы имеют важное применение в области биостратиграфии . Благодаря небольшому размеру и твердой раковине фораминиферы могут сохраняться в большом количестве и с высоким качеством сохранности; благодаря сложной морфологии отдельные виды легко узнаваемы. Виды фораминифер в летописи окаменелостей имеют ограниченный диапазон между первой эволюцией вида и его исчезновением; стратиграфы разработали последовательные изменения в комплексах отверстий на протяжении большей части фанерозоя . Таким образом, комплекс фораминифер в пределах данной местности можно проанализировать и сравнить с известными датами появления и исчезновения, чтобы сузить возраст пород. Это позволяет палеонтологам интерпретировать возраст осадочных пород , когда радиометрическое датирование неприменимо. [ 91 ] Такое применение фораминифер было обнаружено Альвой К. Эллисором в 1920 году. [ 92 ]

Известковые ископаемые фораминиферы образовались из элементов, обнаруженных в древних морях, где они жили. Таким образом, они очень полезны в палеоклиматологии и палеоокеанографии . Их можно использовать в качестве индикатора климата для реконструкции климата прошлого путем изучения соотношений стабильных изотопов и содержания микроэлементов в раковинах (тесты). Глобальную температуру и объем льда можно определить по изотопам кислорода, а также по истории углеродного цикла и продуктивности океана путем изучения соотношений стабильных изотопов углерода; [ 93 ] см. δ18O и δ13C . Концентрация микроэлементов, таких как стронций (Sr), [ 94 ] магний (Mg), [ 95 ] литий (Li) [ 96 ] и бор (В), [ 97 ] также содержат огромное количество информации о глобальных температурных циклах, выветривании континентов и роли океана в глобальном углеродном цикле. Географические закономерности, наблюдаемые в окаменелостях планктонных форм, также используются для реконструкции древних океанских течений .
Современное использование
[ редактировать ]Нефтяная промышленность в значительной степени полагается на микроископаемые, такие как отверстия, для поиска потенциальных месторождений углеводородов. [ 98 ]

По тем же причинам, по которым они являются полезными биостратиграфическими маркерами, сообщества живых фораминифер использовались в качестве биоиндикаторов в прибрежной среде, включая индикаторы здоровья коралловых рифов. Поскольку карбонат кальция подвержен растворению в кислых условиях, фораминиферы могут особенно пострадать от изменения климата и закисления океана .

Фораминиферы широко используются в разведке нефти и регулярно используются для интерпретации возраста и палеосреды осадочных пластов в нефтяных скважинах. [ 99 ] Агглютинированные ископаемые фораминиферы, захороненные глубоко в осадочных бассейнах, можно использовать для оценки термической зрелости, которая является ключевым фактором для добычи нефти. Индекс окраски фораминифер [ 100 ] (FCI) используется для количественной оценки изменений цвета и оценки температуры захоронения. Данные FCI особенно полезны на ранних стадиях производства нефти (около 100 °C).
Фораминиферы также могут использоваться в археологии для поиска источников некоторых видов каменного сырья. Некоторые типы камней, такие как известняк , обычно содержат окаменелые фораминиферы. Типы и концентрации этих окаменелостей в образце камня можно использовать для сопоставления этого образца с источником, который, как известно, содержит ту же «подпись окаменелостей». [ 101 ]
Галерея
[ редактировать ]-
 Фораминиферы острова Паг, Адриатическое море -60 м, ширина поля 5,5 мм.
Фораминиферы острова Паг, Адриатическое море -60 м, ширина поля 5,5 мм. -
 Фораминиферы острова Паг, Адриатическое море -60 м, ширина поля 5,5 мм.
Фораминиферы острова Паг, Адриатическое море -60 м, ширина поля 5,5 мм. -
 Фораминиферы острова Паг, Адриатическое море -60 м, ширина поля 5,5 мм.
Фораминиферы острова Паг, Адриатическое море -60 м, ширина поля 5,5 мм. -
 Фораминиферы острова Паг, Адриатическое море -60 м, ширина поля 5,5 мм.
Фораминиферы острова Паг, Адриатическое море -60 м, ширина поля 5,5 мм. -
 Фораминиферы Индийского океана, юго-восточное побережье Бали, ширина поля 5,5 мм.
Фораминиферы Индийского океана, юго-восточное побережье Бали, ширина поля 5,5 мм. -
 Фораминиферы Индийского океана, юго-восточное побережье Бали, ширина поля 5,5 мм.
Фораминиферы Индийского океана, юго-восточное побережье Бали, ширина поля 5,5 мм. -
 Фораминиферы Индийского океана, юго-восточное побережье Бали, ширина поля 5,5 мм.
Фораминиферы Индийского океана, юго-восточное побережье Бали, ширина поля 5,5 мм. -
 Фораминиферы в Нгапали, Мьянма, ширина поля 5,22 мм.
Фораминиферы в Нгапали, Мьянма, ширина поля 5,22 мм. -
 Foraminifera Heterostegina depressa , ширина поля 4,4 мм.
Foraminifera Heterostegina depressa , ширина поля 4,4 мм.









Ссылки
[ редактировать ]- ^ Лаура Вегенер Парфри ; Дэниел Дж. Г. Лар; Эндрю Х. Нолл ; Лаура А. Кац (16 августа 2011 г.). «Оценка времени ранней диверсификации эукариот с помощью мультигенных молекулярных часов» (PDF) . Труды Национальной академии наук Соединенных Штатов Америки . 108 (33): 13624–9. Бибкод : 2011PNAS..10813624P . дои : 10.1073/PNAS.1110633108 . ISSN 0027-8424 . ПМК 3158185 . ПМИД 21810989 . Викиданные Q24614721 .
- ^ Бинцевская, Анна; Половодова Астеман Ирина; Фармер, Элизабет Дж. (2014), «Фораминиферы (Бентос)» , в Харффе, январь; Мешеде, Мартин; Петерсен, Свен; Тиде, Йорн (ред.), Энциклопедия морских геолого-геофизических наук , Дордрехт: Springer Нидерланды, стр. 1–8, doi : 10.1007/978-94-007-6644-0_60-1 , ISBN 978-94-007-6644-0 , получено 13 мая 2021 г.
- ^ Кимото, Кацунори (2015), Оцука, Сусуму; Сузаки, Тошинобу; Судзуки, Норитоши (ред.), «Планктические фораминиферы» , Морские протисты: разнообразие и динамика , Токио: Springer Japan, стр. 129–178. дои : 10.1007/978-4-431-55130-0_7 , ISBN 978-4-431-55130-0 , получено 13 мая 2021 г.
- ^ Сименсма, Ферри; Апотелоз-Перре-Жантиль, Лор; Хольцманн, Мария; Клаусс, Штеффен; Фёлькер, Экхард; Павловский, Январь (1 августа 2017 г.). «Таксономическая ревизия пресноводных фораминифер с описанием двух новых агглютинирующих видов и родов» . Европейский журнал протистологии . 60 : 28–44. дои : 10.1016/j.ejop.2017.05.006 . ISSN 0932-4739 . ПМИД 28609684 .
- ^ Болтовской, Эстебан; Райт, Рамиль (1976), Болтовской, Эстебан; Райт, Рамиль (ред.), «Бентонные фораминиферы солоноватой, гиперсоленой, ультрасоленой и пресной воды» , Современные фораминиферы , Дордрехт: Springer Нидерланды, стр. 139–150, doi : 10.1007/978-94-017-2860-7_5 , ISBN 978-94-017-2860-7 , получено 13 мая 2021 г.
- ^ Жир, Олав (2009). Мейобентология: микроскопическая подвижная фауна водных отложений (2-е изд.). Берлин: Шпрингер. ISBN 978-3540686576 .
- ^ Jump up to: а б Лейзерович, Франк; Павловский, Ян; Фрессине-Таше, Лоуренс; Мармейс, Роланд (1 сентября 2010 г.). «Молекулярные доказательства широкого распространения фораминифер в почвах». Экологическая микробиология . 12 (9): 2518–26. Бибкод : 2010EnvMi..12.2518L . дои : 10.1111/j.1462-2920.2010.02225.x . ПМИД 20406290 . S2CID 20940138 .
- ^ Кеннетт, JP; Шринивасан, М.С. (1983). Неогеновые планктонные фораминиферы: филогенетический атлас . Хатчинсон Росс. ISBN 978-0-87933-070-5 .
- ^ Павловский Дж.; Лейзерович, Ф.; Эслинг, П. (1 октября 2014 г.). «Исследования экологического разнообразия фораминифер следующего поколения: подготовка будущего» . Биологический вестник . 227 (2): 93–106. дои : 10.1086/BBLv227n2p93 . ISSN 0006-3185 . ПМИД 25411369 . S2CID 24388876 .
- ^ Альд, С.М. и др . (2007) Разнообразие, номенклатура и таксономия протистов , Syst. Биол. 56(4), 684–689, DOI: 10.1080/10635150701494127.
- ^ Павловский Дж., Лейзерович Ф. и Эслинг П. (2014). Исследования экологического разнообразия фораминифер нового поколения: готовим будущее . Биологический бюллетень, 227(2), 93-106.
- ^ «Мировая база данных фораминифер» .
- ^ Маршалл М. (3 февраля 2010 г.). «Зоолог: «Живой пляжный мяч» — это гигантская одноклеточная клетка» . Новый учёный .
- ^ Jump up to: а б Липпс Дж. Х., Фингер К. Л., Уокер С. Е. (октябрь 2011 г.). «Как нам называть фораминиферы» (PDF) . Журнал исследований фораминиферов . 41 (4): 309–313. Бибкод : 2011JForR..41..309L . дои : 10.2113/gsjfr.41.4.309 . Проверено 10 апреля 2018 г.
- ^ «Фораминиферы | Фокус ископаемых | Время | Открытие геологии | Британская геологическая служба (BGS)» . bgs.ac.uk. Проверено 20 июля 2020 г.
- ^ «Микрография, или Некоторые физиологические описания мельчайших тел, сделанные с помощью лупы? с наблюдениями и последующими исследованиями / Р. Гук ...: Гук, Роберт,: Бесплатная загрузка, заимствование и потоковая передача» . Интернет-архив . 1665 . Проверено 20 июля 2020 г.
- ^ Jump up to: а б с Сен Гупта, Барун К. (2003), «Систематика современных фораминифер», в Сен Гупта, Барун К. (ред.), Современные фораминиферы , Springer Нидерланды, стр. 7–36, doi : 10.1007/0-306-48104 -9_2 , ISBN 978-0-306-48104-8
- ^ Jump up to: а б БУДАГЕР-ФАДЕЛЬ, МАРСЕЛЬ К. (2018), «Биология и эволюционная история крупных донных фораминифер», Эволюция и геологическое значение крупных донных фораминифер (2-е изд.), UCL Press, стр. 1–44, doi : 10.2307/j .ctvqhsq3.3 , ISBN 978-1-911576-94-5 , JSTOR j.ctvqhsq3.3
- ^ Хансен, Х. (1 января 1981 г.). «О Лоренце Шпенглере и неотипе фораминиферы Calcarina spengleri» .
- ^ д'Орбиньи, Альсид (1826). «Методическая таблица класса головоногих» . Анналы естественных наук, Париж . Серия 1. 7 : 245–314 – через Библиотеку наследия биоразнообразия.
- ^ «Кеннеди, Маргарет Мур (1896–1967)» , Оксфордский национальный биографический словарь , Oxford University Press, 6 февраля 2018 г., doi : 10.1093/odnb/9780192683120.013.34281 , получено 3 ноября 2022 г.
- ^ Jump up to: а б с д Павловский, Ян; Хольцманн, Мария; Тышка, Ярослав (1 апреля 2013 г.). «Новая надпорядковая классификация фораминифер: молекулы соответствуют морфологии» . Морская микропалеонтология . 100 : 1–10. Бибкод : 2013МарМП.100....1П . дои : 10.1016/j.marmicro.2013.04.002 . ISSN 0377-8398 .
- ^ Jump up to: а б Леблих, А.Р. младший; Таппан, Х. (1964). «Фораминиферида». Часть C, Протиста 2 . Трактат по палеонтологии беспозвоночных. Геологическое общество Америки. стр. C55–C786. ISBN 978-0-8137-3003-5 .
- ^ Jump up to: а б с д и ж г Сен Гупта, Барун К. (2002). Современные фораминиферы . Спрингер. п. 16. ISBN 978-1-4020-0598-5 .
- ^ Jump up to: а б с Кавальер-Смит, Т. (2004). «Только шесть королевств жизни» (PDF) . Слушания. Биологические науки . 271 (1545): 1251–62. дои : 10.1098/rspb.2004.2705 . ПМК 1691724 . ПМИД 15306349 .
- ^ Jump up to: а б Кавальер-Смит, Т. (2003). «Филогения протистов и классификация простейших высокого уровня». Европейский журнал протистологии . 34 (4): 338–348. дои : 10.1078/0932-4739-00002 .
- ^ «Толвеб Церкозоа» . Архивировано из оригинала 25 декабря 2019 года . Проверено 21 июня 2010 г.
- ^ Европейский регистр морских видов
- ^ eForams-taxonomy. Архивировано 3 октября 2011 г. в Wayback Machine.
- ^ Панцирные амебы как индикаторы окружающей среды (PDF) , заархивировано из оригинала (PDF) 27 ноября 2016 г. , получено 27 ноября 2016 г.
- ^ Михалевич, В.И. (2013). «Новый взгляд на систематику и эволюцию фораминифер» . Микропалеонтология . 59 (6): 493–527. Бибкод : 2013MiPal..59..493M . дои : 10.47894/mpal.59.6.01 . S2CID 90243148 .
- ^ Павловский, Ян; Боливар, Игнасио; Фарни, Хосе Ф.; Варгас, Коломбан Де; Баузер, Сэмюэл С. (1999). «Молекулярные доказательства того, что Reticulomyxa Filosa является пресноводным голым фораминифером». Журнал эукариотической микробиологии . 46 (6): 612–617. дои : 10.1111/j.1550-7408.1999.tb05137.x . ISSN 1550-7408 . ПМИД 10568034 . S2CID 36497475 .
- ^ Jump up to: а б с д и ж Сарасвати, Пратул Кумар; Шринивасан, М.С. (2016), Сарасвати, Пратул Кумар; Шринивасан, MS (ред.), «Микрокаменелости с известковой стенкой», Микропалеонтология: принципы и приложения , Springer International Publishing, стр. 81–119, doi : 10.1007/978-3-319-14574-7_6 , ISBN 978-3-319-14574-7
- ^ Jump up to: а б с Сен Гупта, Барун К. (1982). «Экология донных фораминифер» . В Бродхеде, ТВ (ред.). Фораминиферы: конспекты краткого курса, организованного М.А. Бузасом и Б.К. Сеном Гуптой . Исследования по геологии. Том. 6. Университет Теннесси, факультет геологических наук. стр. 37–50. ISBN 978-0910249058 . OCLC 9276403 .
- ^ Jump up to: а б с Хемлебен, К.; Андерсон, Орегон; Шпиндлер, М. (1989). Современные планктонные фораминиферы . Спрингер-Верлаг. ISBN 978-3-540-96815-3 .
- ^ Грелль, КГ (1 января 1979 г.). «Цитогенетические системы и эволюция фораминифер» . Журнал исследований фораминиферов . 9 (1): 1–13. Бибкод : 1979JForR...9....1G . дои : 10.2113/gsjfr.9.1.1 . ISSN 0096-1191 .
- ^ Jump up to: а б с Лекиффр, Шарлотта; Бернхард, Джоан М.; Мабийо, Гийом; Филипссон, Хелена Л.; Мейбом, Андерс; Жеслен, Эммануэль (1 января 2018 г.). «Обзор клеточной ультраструктуры бентосных фораминифер: новые наблюдения за видами роталид в контексте существующей литературы» . Морская микропалеонтология . 138 : 12–32. Бибкод : 2018мартMP.138...12L . дои : 10.1016/j.marmicro.2017.10.005 . hdl : 1912/9530 . ISSN 0377-8398 .
- ^ Доманов, М.М. (июль 2015 г.). «Природные радионуклиды 226Ra и 232Th в ксенофиофорах Тихого океана» . Геохимия Интернэшнл . 53 (7): 664–669. Бибкод : 2015GeocI..53..664D . дои : 10.1134/S0016702915070034 . ISSN 0016-7029 . S2CID 127121951 .
- ^ Такаги Х., Кимото К., Фуджики Т., Сайто Х., Шмидт К., Кучера М. и Мория К. (2019) «Характеристика фотосимбиоза современных планктонных фораминифер». Биогеонауки , 16 (17). два : 10.5194/bg-16-3377-2019 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Кучера, М.; Дарлинг, К.Ф. (апрель 2002 г.). «Загадочные виды планктонных фораминифер: их влияние на палеоокеанографические реконструкции». Филос Транс Роял Сок А. 360 (1793): 695–718. Бибкод : 2002RSPTA.360..695K . дои : 10.1098/rsta.2001.0962 . ПМИД 12804300 . S2CID 21279683 .
- ^ Jump up to: а б Уджие, Юрика; Кимото, Кацунори; Павловский, Ян (декабрь 2008 г.). «Молекулярные доказательства независимого происхождения современных трехсерийных планктонных фораминифер от донных предков». Морская микропалеонтология . 69 (3–4): 334–340. Бибкод : 2008МарМП..69..334У . doi : 10.1016/j.marmicro.2008.09.003 .
- ^ Jump up to: а б Оздикмен, Хусейн (июнь 2009 г.). «Замените названия некоторых таксонов одноклеточных животных (Protozoa» (PDF) . Munis Entomology & Zoology . 4 (1): 233–256.
- ^ Дубицкая, Зофья (2019). «Расположение камер и структура стенок в филогенетической классификации высокого ранга фораминифер» . Acta Palaeontologica Polonica . 64 . дои : 10.4202/app.00564.2018 . ISSN 0567-7920 .
- ^ Симбиоз и гибкость микробиома при кальцинировании бентосных фораминифер Большого Барьерного рифа.
- ^ Характеристика фотосимбиоза современных планктонных фораминифер - BG.
- ^ Достижения в микробной экологии, Том 11.
- ^ Бернхард, Дж. М.; Баузер, С.М. (1999). «Бентосные фораминиферы дизоксичных отложений: секвестрация хлоропластов и функциональная морфология». Обзоры наук о Земле . 46 (1): 149–165. Бибкод : 1999ESRv...46..149B . дои : 10.1016/S0012-8252(99)00017-3 .
- ^ Jump up to: а б Гольдштейн, Сьюзен Т. (2003), «Фораминиферы: биологический обзор», в Сен Гупта, Барун К. (ред.), Современные фораминиферы , Springer Нидерланды, стр. 37–55, doi : 10.1007/0-306-48104 -9_3 , ISBN 978-0-306-48104-8
- ^ Цучия, Масаси; Номаки, Хидетака (1 октября 2021 г.). «Быстрая реакция гигантских протистских ксенофиофоров (Foraminifera, Rhizaria) на поступление органического вещества на абиссальные глубины, выявленная в ходе эксперимента по мечению двойным стабильным изотопом in situ» . Глубоководные исследования. Часть I: Статьи океанографических исследований . 176 : 103608. Бибкод : 2021DSRI..17603608T . дои : 10.1016/j.dsr.2021.103608 . ISSN 0967-0637 .
- ^ Калвер, Стивен Дж.; Липпс, Джер Х. (2003), Келли, Патрисия Х.; Ковалевский, Михал; Хансен, Тор А. (ред.), «Хищничество фораминифер и со стороны фораминифер» , «Взаимодействия хищников и жертв в летописи окаменелостей » , Бостон, Массачусетс: Springer US, стр. 7–32, doi : 10.1007/978-1-4615- 0161-9_2 , ISBN 978-1-4613-4947-1 , получено 30 сентября 2020 г.
- ^ Мудли, Л.; Гесс, К. (1 августа 1992 г.). «Толерантность инфаунальных бентосных фораминифер к низким и высоким концентрациям кислорода» . Биологический вестник . 183 (1): 94–98. дои : 10.2307/1542410 . ISSN 0006-3185 . JSTOR 1542410 . ПМИД 29304574 .
- ^ Добрый день, Эй Джей; Тодо, Ю.; Уэмацу, К.; Китазато, Х. (июль 2008 г.). «Новые фораминиферы (Protista) с органическими стенками из самой глубокой точки океана, бездны Челленджера (западная часть Тихого океана)» . Зоологический журнал Линнеевского общества . 153 (3): 399–423. дои : 10.1111/j.1096-3642.2008.00393.x .
- ^ Хольцманн, Мария; Добрый день, Эндрю Дж.; Сименсма, Ферри; Павловский, Ян (29 октября 2021 г.). «Обзор: Пресноводные и почвенные фораминиферы – история давно забытых родственников» . Журнал исследований фораминиферов . 51 (4): 318–331. Бибкод : 2021JForR..51..318H . дои : 10.2113/gsjfr.51.4.318 . ISSN 0096-1191 . S2CID 240240437 .
- ^ Мур, RC; Лаликер, АГ; Фишер, К.Г. (1952). «Глава 2 Фораминиферы и Радиолярии». Окаменелости беспозвоночных . МакГроу-Хилл. OCLC 547380 .
- ^ Jump up to: а б Хейнс-младший (18 июня 1981 г.). Фораминиферы . Спрингер. ISBN 978-1-349-05397-1 .
- ^ Лана, К. (2001). «Меловая Картерина (Фораминифера)». Морская микропалеонтология . 41 (1–2): 97–102. Бибкод : 2001МарМП..41...97Л . дои : 10.1016/S0377-8398(00)00050-5 .
- ^ Jump up to: а б Конторович А.Е.; Варламов А.И.; Гражданкин Д.В.; Карлова, Г.А.; Клец, АГ; Конторович, В.А.; Сараев С.В.; Терлеев А.А.; Беляев С. Ю.; Вараксина, IV; Ефимов А.С. (1 декабря 2008 г.). "Разрез венда на востоке Западно-Сибирской плиты (по данным скважины Восток 3)" . Российская геология и геофизика . 49 (12): 932–939. Бибкод : 2008РуГГ...49..932К . дои : 10.1016/j.rgg.2008.06.012 . ISSN 1068-7971 .
- ^ Фораминиферы: История изучения , Университетский колледж Лондона , получено 20 сентября 2007 г.
- ^ Лангер, MR; Шелк, МТБ; Липпс, Дж. Х. (1997). «Глобальное производство карбонатов и углекислого газа в океане: роль рифовых фораминифер» . Журнал исследований фораминиферов . 27 (4): 271–277. Бибкод : 1997JForR..27..271L . дои : 10.2113/gsjfr.27.4.271 .
- ^ Адл, С.М.; Симпсон, AGB; Фермер, Массачусетс; Андерсон; и др. (2005). «Новая классификация эукариот более высокого уровня с упором на таксономию протистов» . Журнал эукариотической микробиологии . 52 (5): 399–451. дои : 10.1111/j.1550-7408.2005.00053.x . ПМИД 16248873 . S2CID 8060916 .
- ^ Jump up to: а б Павловский, Ян; Хольцманн, Мария; Берни, Седрик; Фарни, Хосе; Добрый день, Эндрю Дж.; Седхаген, Томас; Хабура, Андреа; Баузер, Сэмюэл С. (30 сентября 2003 г.). «Эволюция ранних фораминифер» . Труды Национальной академии наук . 100 (20): 11494–11498. Бибкод : 2003PNAS..10011494P . дои : 10.1073/pnas.2035132100 . ISSN 0027-8424 . ПМК 208786 . ПМИД 14504394 .
- ^ Груссен, Матье; Павловский, Ян; Ян, Цзыхэн (1 октября 2011 г.). «Оценка времени расхождения фораминифер по байесовским расслабленным часам» . Молекулярная филогенетика и эволюция . 61 (1): 157–166. Бибкод : 2011МОЛПЭ..61..157Г . дои : 10.1016/j.ympev.2011.06.008 . ISSN 1055-7903 . ПМИД 21723398 .
- ^ Сейлахер, А. (1 января 2007 г.). «Природа вендобионтов» . Геологическое общество, Лондон, специальные публикации . 286 (1): 387–397. Бибкод : 2007GSLSP.286..387S . дои : 10.1144/SP286.28 . ISSN 0305-8719 . S2CID 128619251 .
- ^ Бобровский Илья; Хоуп, Джанет М.; Иванцов Андрей; Неттерсхайм, Бенджамин Дж.; Халлманн, Кристиан; Брокс, Йохен Дж. (21 сентября 2018 г.). «Древние стероиды делают эдиакарскую ископаемую дикинсонию одним из самых ранних животных» . Наука . 361 (6408): 1246–1249. Бибкод : 2018Sci...361.1246B . дои : 10.1126/science.aat7228 . hdl : 1885/230014 . ISSN 0036-8075 . ПМИД 30237355 .
- ^ Суинбэнкс, Д.Д. (1 октября 1982 г.). «Пьеодиктон: следы инфаунальных ксенофиофоров?» . Наука . 218 (4567): 47–49. Бибкод : 1982Sci...218...47S . дои : 10.1126/science.218.4567.47 . ISSN 0036-8075 . ПМИД 17776707 . S2CID 28690086 .
- ^ Jump up to: а б Левин, Лиза А. (1994). «Палеоэкология и экология ксенофиофоров». ПАЛЕОС . 9 (1): 32–41. Бибкод : 1994Палай...9...32Л . дои : 10.2307/3515076 . ISSN 0883-1351 . JSTOR 3515076 .
- ^ Рона, Питер А.; Зейлахер, Адольф; де Варгас, Колумбан; Добрый день, Эндрю Дж.; Бернхард, Джоан М.; Баузер, Сэм; Ветриани, Константино; Вирсен, Карл О.; Муллино, Лорен; Шеррелл, Роберт; Фредерик Грассл, Дж. (1 сентября 2009 г.). «Paleodictyon nodosum: живое ископаемое на глубоководном дне» . Глубоководные исследования, часть II: Актуальные исследования в океанографии . Морская бентическая экология и биоразнообразие: сборник последних достижений в честь Дж. Фредерика Грассла. 56 (19): 1700–1712. Бибкод : 2009DSRII..56.1700R . дои : 10.1016/j.dsr2.2009.05.015 . ISSN 0967-0645 .
- ^ Добрый день, Эндрю Дж.; Хольцманн, Мария; Колле, Клеманс; Гуано, Орели; Каменская, Ольга; Вебер, Александра А.-Т.; Павловский, Ян (1 марта 2017 г.). «Гигантские протисты (ксенофиофоры, фораминиферы) исключительно разнообразны в тех частях абиссальной восточной части Тихого океана, где разрешено разведку полиметаллических конкреций» . Биологическая консервация . 207 : 106–116. Бибкод : 2017BCons.207..106G . дои : 10.1016/j.biocon.2017.01.006 . ISSN 0006-3207 .
- ^ Макилрой, Дункан; Грин, Орегон; Брейзер, доктор медицины (2001). «Палеобиология и эволюция самых ранних агглютинированных фораминифер: платизоленитов, спирозоленитов и родственных форм». Летайя . 34 (1): 13–29. Бибкод : 2001Лета..34...13М . дои : 10.1080/002411601300068170 . ISSN 1502-3931 .
- ^ Jump up to: а б Скотт, Дэвид Б.; Медиоли, Франко; Браунд, Риган (1 июня 2003 г.). «Фораминиферы из кембрия Новой Шотландии: древнейшие многокамерные фораминиферы» . Микропалеонтология . 49 (2): 109–126. дои : 10.2113/49.2.109 . ISSN 1937-2795 .
- ^ Jump up to: а б с д и ж Таппан, Хелен; Леблих, Альфред Р. (1988). «Эволюция, разнообразие и вымирание фораминифер». Журнал палеонтологии . 62 (5): 695–714. ISSN 0022-3360 . JSTOR 1305391 .
- ^ Вагнер, Роберт Герман (1983). Карбон мира: Китай, Корея, Япония и Юго-Восточная Азия . ИГМЕ. п. 88. ИСБН 978-84-300-9949-8 .
- ^ Голдберг, Уолтер М. (4 октября 2013 г.). Биология рифов и рифовых организмов . Издательство Чикагского университета. п. 73. ИСБН 978-0-226-92537-0 . Проверено 10 января 2023 г.
- ^ «Фусулиниды | ГеоКанзас» . geokansas.ku.edu . Проверено 16 мая 2020 г.
- ^ Чаплевски, Джон Дж. «Навигатор PBDB» . paleobiodb.org . Проверено 16 мая 2020 г.
- ^ Грефе, КУ (2005). «Бентосные фораминиферы и палеосреда в нижней и средней юре Западного Баскско-Кантабрийского бассейна (Северная Испания)». Журнал иберийской геологии . 31 (2): 217–233. S2CID 55664447 .
- ^ Jump up to: а б Пирсон, Пол (19 ноября 1998 г.). «Споры о природе» . Природа : 1–3. дои : 10.1038/nature28135 .
- ^ Журнал биоинформатики и биологии, Использование подхода множественного анализа для реконструкции филогенетических отношений между планктонными фораминиферами на основе сильно расходящихся и полиморфных по длине последовательностей рДНК SSU.
- ^ Гебхардт, Хольгер (1 февраля 1997 г.). «Фораминиферы сеномана и турона из Ашаки (северо-восток Нигерии): количественный анализ и палеоэкологическая интерпретация» . Меловые исследования . 18 (1): 17–36. Бибкод : 1997CrRes..18...17G . дои : 10.1006/cres.1996.0047 . ISSN 0195-6671 .
- ^ Балди, Кэтрин; Бенкович, Ласло; Штано, Орсоля (1 мая 2002 г.). «Развитие баденского (среднего миоцена) бассейна на юго-западе Венгрии: история опускания на основе количественной палеобатиметрии фораминифер». Международный журнал наук о Земле . 91 (3): 490–504. Бибкод : 2002IJEaS..91..490B . дои : 10.1007/s005310100226 . ISSN 1437-3262 . S2CID 129296067 .
- ^ Робинсон, Марси (2021) Планктические фораминиферы из Геологической службы США Южного Мэриленда .
- ^ Захос, Джеймс С.; Шеклтон, Николас Дж.; Ревена, Джастин С.; Пялике, Хейко; Флауэр, Бенджамин П. (13 апреля 2001 г.). «Реакция климата на орбитальное воздействие через границу олигоцена и миоцена» (PDF) . Наука . 292 (5515). Американская ассоциация содействия развитию науки (AAAS): 274–278. Бибкод : 2001Sci...292..274Z . дои : 10.1126/science.1058288 . ISSN 0036-8075 . ПМИД 11303100 . S2CID 38231747 .
- ^ Раймо, М.Э. и Лисецки, Л.Е. (2005) «Плиоцен-плейстоценовая стопка из 57 глобально распределенных записей бентического δ18O», Палеокеанография , 20 : PA1003.
- ^ Пети, младший; Жузель, Дж.; Рейно, Д.; Барков Н.И.; Барнола, Ж.-М.; Базиль, И.; Бендер, М.; Чапеллаз, Дж.; Дэвис, М.; Делайг, Г.; Дельмотт, М.; Котляков В.М.; Легран, М.; Липенков В.Ю.; Лориус, К.; Пепин, Л.; Ритц, К.; Зальцман, Э.; Стивенард, М. (1999). «Климатическая и атмосферная история последних 420 000 лет ледяного ядра Восток, Антарктида» . Природа . 399 (6735). ООО «Спрингер Сайенс энд Бизнес Медиа»: 429–436. Бибкод : 1999Natur.399..429P . дои : 10.1038/20859 . ISSN 0028-0836 . S2CID 204993577 .
- ^ Диккенс, Г. Р.; Кастильо, ММ; Уокер, JCG (1997). «Взрыв газа в последнем палеоцене; моделирование эффектов первого порядка массивной диссоциации океанического гидрата метана». Геология . 25 (3): 259–262. Бибкод : 1997Geo....25..259D . doi : 10.1130/0091-7613(1997)025<0259:abogit>2.3.co;2 . ПМИД 11541226 . S2CID 24020720 .
- ^ «Странности PETM» . RealClimate. 2009. Архивировано из оригинала 12 февраля 2016 года . Проверено 3 февраля 2016 г.
- ^ Макинерни, ФА; Винг, С. (2011). «Нарушение углеродного цикла, климата и биосферы с последствиями для будущего» . Ежегодный обзор наук о Земле и планетах . 39 : 489–516. Бибкод : 2011AREPS..39..489M . doi : 10.1146/annurev-earth-040610-133431 . Архивировано из оригинала 14 сентября 2016 года . Проверено 3 февраля 2016 г.
- ^ Зибе, Р.; Зачос, Джей Си; Диккенс, GR (2009). «Одного только воздействия углекислого газа недостаточно, чтобы объяснить палеоцен-эоценовое термическое максимальное потепление». Природа Геонауки . 2 (8): 576–580. Бибкод : 2009NatGe...2..576Z . CiteSeerX 10.1.1.704.7960 . дои : 10.1038/ngeo578 .
- ^ Марси М. Робинсон (2022) Древние микроископаемые — ключ к будущему восстановлению климата Геологическая служба США .
 В данной статье использован текст из этого источника, находящегося в свободном доступе .
В данной статье использован текст из этого источника, находящегося в свободном доступе .
- ^ Бабила, Тали Л.; Пенман, Дональд Э.; Стэндиш, Кристофер Д.; Дубрава, Моника; Бралоуэр, Тимоти Дж.; Робинсон, Марси М.; Self-Trail, Джин М.; Спейер, Роберт П.; Стассен, Питер; Фостер, Гэвин Л.; Захос, Джеймс К. (18 марта 2022 г.). «Потепление и закисление поверхности океана, вызванные быстрым выбросом углерода, предшествуют палеоцен-эоценовому термальному максимуму» . Достижения науки . 8 (11). Американская ассоциация содействия развитию науки (AAAS): eabg1025. Бибкод : 2022SciA....8G1025B . дои : 10.1126/sciadv.abg1025 . ISSN 2375-2548 . ПМЦ 8926327 . ПМИД 35294237 . S2CID 247498325 .
- ^ Австралия, c\=AU\;o\=Правительство Австралии\;ou\=Geoscience (15 мая 2014 г.). «Биостратиграфия» . ga.gov.au. Проверено 20 июля 2020 г.
{{cite web}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Кушман, Джозеф А.; Эллисор, Альва К. (1 января 1945 г.). «Фораминиферовая фауна формации Анауак». Журнал палеонтологии . 19 (6): 545–572. JSTOR 1299203 .
- ^ Зачос, Джей Си; Пагани, М.; Слоан, Л.; Томас, Э.; Биллапс, К. (2001). «Тенденции, ритмы и отклонения в глобальном климате от 65 млн лет до наших дней» (PDF) . Наука . 292 (5517): 686–693. Бибкод : 2001Sci...292..686Z . дои : 10.1126/science.1059412 . ПМИД 11326091 . S2CID 2365991 .
- ^ Кеул, Нина; Лангер, Г.; Томс, С.; де Нуйер, LJ; Райхарт, Дж.Дж.; Биджма, Дж. (апрель 2017 г.). «Изучение фораминифер Sr/Ca как представителя новой карбонатной системы» (PDF) . Geochimica et Cosmochimica Acta . 202 : 374–386. Бибкод : 2017GeCoA.202..374K . дои : 10.1016/j.gca.2016.11.022 . hdl : 1874/380107 .
- ^ Брэнсон, Оскар; Редферн, Саймон А.Т.; Тылищак, Толек; Садеков, Алексей; Лангер, Джеральд; Кимото, Кацунори; Элдерфилд, Генри (декабрь 2013 г.). «Координация магния в фораминиферовом кальците» . Письма о Земле и планетологии . 383 : 134–141. Бибкод : 2013E&PSL.383..134B . дои : 10.1016/j.epsl.2013.09.037 .
- ^ Мисра, С.; Фрёлих, ПН (26 января 2012 г.). «История изотопов лития в кайнозойской морской воде: изменения в силикатном и обратном выветривании» . Наука . 335 (6070): 818–823. Бибкод : 2012Sci...335..818M . дои : 10.1126/science.1214697 . ПМИД 22282473 . S2CID 42591236 .
- ^ Хемминг, штат Нью-Йорк; Хэнсон, Дж. Н. (январь 1992 г.). «Изотопный состав и концентрация бора в современных морских карбонатах». Geochimica et Cosmochimica Acta . 56 (1): 537–543. Бибкод : 1992GeCoA..56..537H . дои : 10.1016/0016-7037(92)90151-8 .
- ^ Бордман, RS; Читам, АХ; Роуэлл, Эй Джей (1987). Ископаемые беспозвоночные . Уайли. ISBN 978-0865423022 .
- ^ Джонс, RW (1996). Микропалеонтология в разведке нефти . Кларендон Пресс. ISBN 978-0-19-854091-5 .
- ^ Макнил, Д.Х.; Исслер, доктор медицинских наук; Сноудон, ЛР (1996). Изменение цвета, термическая зрелость и погребальный диагенез ископаемых фораминифер . Бюллетень Геологической службы Канады. Том. 499. Геологическая служба Канады. ISBN 978-0-660-16451-9 .
- ^ Уилкинсон, Ян П.; Уильямс, Марк; Янг, Джереми Р.; Кук, Саманта Р.; Фулфорд, Майкл Г.; Лотт, Грэм К. (1 августа 2008 г.). «Применение микрокаменелостей для оценки происхождения мела, используемого при изготовлении римских мозаик в Силчестере» . Журнал археологической науки . 35 (8): 2415–2422. Бибкод : 2008JArSc..35.2415W . дои : 10.1016/j.jas.2008.03.010 . ISSN 0305-4403 .
Внешние ссылки
[ редактировать ]- Общая информация
- На сайте Музея палеонтологии Калифорнийского университета есть введение в фораминиферы.
- Исследователи из Университета Южной Флориды разработали систему с использованием Foraminifera для мониторинга среды коралловых рифов. Архивировано 15 января 2021 года на Wayback Machine.
- Университетского колледжа Лондона На сайте микропалеонтологии представлен обзор фораминифер, включая множество высококачественных SEM.
- Иллюстрированный глоссарий терминов, используемых в исследованиях фораминифер. Архивировано 21 июня 2012 года в Wayback Machine. Это глоссарий Лукаса Хоттингера, опубликованный в электронном журнале OA «Carnets de Géologie - Notebooks on Geology». Архивировано 29 июня 2017 года в Wayback Machine.
- Информация о странице микропалеонтологии Фораминифер Мартина Лангера
- Информация о бентосных фораминиферах, полученная на Летней школе палеоклиматологии в Урбино 2005 г.
- Онлайн-флип-книги
- Иллюстрированный глоссарий терминов, используемых Лукасом Хоттингером в исследованиях фораминифер ( альтернативная версия словаря, опубликованного в «Carnets de Géologie – Notebooks on Geology». Архивировано 29 июня 2017 г. в Wayback Machine ).
- Ресурсы
- pforams@mikrotax - онлайн-база данных с подробным описанием таксономии планктонных фораминифер.
- Проект «Звездный песок» (часть микроскопа ) представляет собой совместную базу данных информации о фораминиферах.
- 3D-модели отверстий, созданные методом рентгеновской томографии.
- У CHRONOS есть несколько ресурсов по фораминиферам , включая страницу поиска таксонов и раздел микропалео. Примечание. Большая часть этого контента теперь включена в веб-сайт pforams@mikrotax.
- eForams - это веб-сайт, посвященный фораминиферам и моделированию раковин фораминифер.
- Галерея фораминифер Иллюстрированный каталог современных и ископаемых фораминифер по родам и местонахождению.
- « Фораминиферы » . Браузер таксономии NCBI . 29178.