Модель эволюции с плескающимся ковшом
| Часть серии о |
| Эволюционная биология |
|---|
 |
Модель эволюции «плескающего ведра» — это теория эволюционной биологии , которая описывает, как нарушения окружающей среды различной величины будут влиять на существующие виды. [ 1 ] [ 2 ] [ 3 ] Теория подчеркивает причинно-следственную связь между факторами окружающей среды, которые влияют на генеалогические системы, обеспечивая всеобъемлющую точку зрения, определяющую взаимосвязь между разнообразием биологических систем.
Эту теорию разработал Найлс Элдридж , американский биолог и палеонтолог . [ 4 ] и опубликовано в журнале «Эволюционная динамика: изучение взаимодействия отбора, случайности, нейтральности и функции», где Элдридж представляет свою модель плескающегося ведра в статье под названием «Плюшащее ведро: как физическая сфера контролирует эволюцию». [ 5 ]
Краткое содержание
[ редактировать ]Модель плескающегося ведра использует образы воды, представляющие виды, плещущиеся взад и вперед в окружающей среде, представленной ведерком. Нарушения в окружающей среде представлены движением ведра, вызывающим выплескивание. Начнем с того, что небольшие выплески/помехи не приводят к проливанию воды; стазис современных видов являются доминирующими. Однако по мере того, как физические возмущения растут по величине и размеру, выплески приведут к выливанию большого количества воды, что представляет собой вымирание и видообразование присутствующих организмов.
Примером, который использует Элдридж, являются динозавры, которые были распространенной формой жизни на Земле на протяжении 150 миллионов лет. [ 6 ] выживание при небольших выплескиваниях в ведро без особых эволюционных изменений. Только после удара астероида в мезозойскую эпоху динозавры вымерли, и после задержки в пять-семь миллионов лет млекопитающие начали видоизменяться и диверсифицироваться. [ 7 ] [ 8 ]
Три модели жизни
[ редактировать ]Стазис
[ редактировать ]Основанный непосредственно на его предыдущей статье под названием «Периодические равновесия», [ 9 ] застойный образ жизни представляет собой периоды «динамических, нерегулярных колебаний». [ 5 ] внутрипопуляционной изменчивости. Стазис не означает, что коллективный геном вида стабилен, он все еще находится в постоянном движении и вариациях, только в неспецифических направлениях. Это приводит к схожим фенотипам между организмами.
Записи окаменелостей дополнительно подтверждают эту идею, поскольку при незначительных или небольших нарушениях в записях окаменелостей не обнаруживается никакой эволюции или адаптивных изменений. И только после более масштабных и масштабных экологических нарушений летопись окаменелостей радикально изменится. [ 10 ]
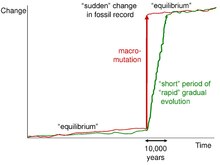
Причины этого застоя кроются в том, что для того, чтобы вызвать генетические сдвиги у всего вида, требуется сила отбора, охватывающая всех членов вида. Факторы окружающей среды, хотя и широко распространены, опережают перемещение видов, которые находят узнаваемые места обитания для расселения и остаются неизменными. [ 11 ] Эта идея, называемая отслеживанием среды обитания, утверждает, что виды способны отслеживать среду обитания лучше, чем естественный отбор может следовать за меняющейся средой.
Даже в исключительных обстоятельствах, таких как резкое изменение климата , вариации многих факторов, таких как исходные геномы, мутационная история и давление отбора внутри вида, означают, что весь вид вряд ли будет двигаться в определенном направлении эволюции.
В целом, застойный образ жизни является доминирующим в жизни и обычно не приводит к каким-либо чистым эволюционным изменениям.
Видообразование и адаптивные изменения
[ редактировать ]В то время как застой был доминирующим паттерном в жизни, адаптивные изменения и, как следствие, создание новых видов возникают в короткие сроки, «подчеркивая» равновесие, установленное застоем. Прерывистость видов возникает не только в результате накопления генетических изменений, но и в сочетании с репродуктивной изоляцией . [ 5 ]
Эта форма аллопатрического видообразования имеет множество правдоподобных моделей, одну из которых описывает Элдридж. Оптимальное место обитания обычно находится в центре ареала вида, причем внешние границы этого места малополезны. Отдельные виды в этих периферийных зонах могут адаптироваться к отличающейся экосистеме, превращая, таким образом, окраинную зону обитания в теперь оптимальную зону для вновь изолированной популяции. [ 12 ]
Синхронное видообразование и вымирание
[ редактировать ]Объединение паттерна 1 (застой) и паттерна 2 (адаптивные изменения) создает окончательный паттерн, о котором свидетельствуют крупномасштабные изменения видов, вызванные достаточно значительными изменениями в окружающей среде в глобальном масштабе. Когда растущая экологическая нагрузка достигает определенного порога, это наряду с миграцией вызывает повсеместное вымирание и видообразование. [ 5 ] Эта закономерность включает в себя целые регионы, охватывающие все таксоны видового уровня и в равной степени затрагивающие их всех. Однако каждый вид реагирует по-разному. Некоторые виды выживают в неизмененном виде, тогда как другие вымирают или видоизменяются.
Существует множество задокументированных явлений, которые очень хорошо согласуются с этой закономерностью. Карлтон Э.Э. Бретт продемонстрировал «скоординированный застой» с помощью ископаемых моделей, продемонстрировав как период застоя, когда 70–85% видов остаются в течение определенного периода, так и после крупномасштабного регионального события только около 20% переживают следующий этап. период стаза. [ 13 ]
Эта модель также была названа Элизабет Врба термином «пульс оборота». [ 14 ] Она задокументировала постепенное понижение температуры на территории нынешней Южной Африки в эпоху плиоцена, которое первоначально не имело большого эффекта. Затем внезапно, через полмиллиона лет, это вызвало резкое изменение окружающей среды: от влажных лесов к саваннам. Здесь произошла та же самая картина застоя, перемежающегося видообразованием и изменениями.
Элдридж предполагает, что эти события массового вымирания «вместо того, чтобы стимулировать видообразование, просто увеличивают вероятность выживания молодых видов». [ 5 ]
Генеалогическая и экологическая двойная иерархия
[ редактировать ]Двойная иерархия генеалогии и экологии необходима в силу двойственной природы организмов. Все организмы делают две основные вещи; они существуют, взаимодействуя с окружающей средой для получения энергии, и размножаются. [ 14 ] Эти два отдельных действия тогда существуют каждое в своей собственной иерархии, но связаны друг с другом на уровне организма посредством естественного отбора и изменчивости.
Генеалогическая иерархия
[ редактировать ]Генеалогическая иерархия существует как следствие пространственного распределения воспроизводства видов. Уровни внутри иерархии повышаются с увеличением размера и географического диапазона, и каждый из них подвергается воздействию соответствующих факторов экологической иерархии. [ 8 ]
Самый низкий уровень генеалогической иерархии — это организм, особенно в его репродуктивном смысле. Эти организмы участвуют в воспроизводстве всего вида. Следующий уровень в иерархии — « деме », представляющие собой скрещивающуюся местную популяцию вида. Их можно рассматривать как специфические региональные вариации скрещивающихся видов. Следующий и второй по высоте уровень в иерархии — виды . Последний и высший уровень генеалогической иерархии — это монофилетические таксоны , которые все имеют общего предка и происходят от него.
Экологическая иерархия
[ редактировать ]Экологическая иерархия аналогичным образом начинается на самом низком уровне организма, хотя в данном случае основное внимание уделяется экономическому стремлению к выживанию. Эти организмы либо конкурируют, сотрудничают или нейтральны по отношению друг к другу в своем выживании. Следующий уровень — «аватары», отличающиеся от дем. Аватары — это местные взаимодействующие сородичи, ориентированные на выживание, а не на размножение. [ 5 ] Комбинация и взаимодействие между аватарами образуют следующий уровень: локальную экосистему . Самый верхний и последний уровень иерархии включает экосистемы региона, которые представляют собой совокупность местных экосистем.
Плескающееся ведро
[ редактировать ]Интегрируя вышеупомянутую систему двойной иерархии вместе с установленными тремя моделями эволюционной жизни, можно полностью реализовать модель эволюции «плескающегося ведра». Пространственно-временной масштаб экологических или физических нарушений можно рассматривать через определенные уровни иерархии, в зависимости от их величины и воздействия. [ 8 ]
Первый уровень: Кратковременные эффекты на уровне демы или аватара. Нет никаких чистых эволюционных изменений, приводящих к застою.
Второй уровень: среднесрочные эффекты, локализованные в конкретных регионах. Стабилизирующий отбор происходит по мере того, как соседние демы или виды заполняют утраченные компоненты, восстанавливая ту же предыдущую иерархию. [ 15 ]
Третий уровень: Крупномасштабные изменения окружающей среды. Эти изменения длятся от десятков лет до тысячи и более лет и достаточно медленны, чтобы позволить видам мигрировать в более оптимальные условия. Следовательно, из-за отслеживания среды обитания происходят некоторые изменения на нижних уровнях иерархии, но в целом виды преобладают и сохраняется стазис.
Четвертый уровень: Региональные беспорядки. Там, где изменения слишком быстры для отслеживания среды обитания, возникает третий образец жизни. На этом пороге в больших масштабах запускаются вымирание и видообразование неродственных видов.
Пятый уровень: Глобальные беспорядки. По сути, это массовое вымирание, которое полностью изменит существующие виды. Вымирание и видообразование широко распространены во всем мире. Примеры включают вымирание в конце перми. [ 16 ]
Прием и критика
[ редактировать ]Модель эволюции с плещущимся ведром была хорошо принята некоторыми биологами-эволюционистами. Коллега-философ и биолог Тельмо Пиевани утверждает, что «теория иерархии способна охватить все уровни, которые делают эволюционную игру такой сложной: от генов до организмов, видов и крупнейших экологических сценариев». [ 17 ] Палеонтологи Брюс Либерман и Элизабет Врба заявили, что модель плескающегося ведра, содержащаяся в теории иерархии, «играет (играет) заметную роль в формировании основных особенностей разнообразия и биологической организации». [ 18 ]
Модель также подверглась критике. Одна из самых больших проблем в модели эволюции с плещущимся ведром – это отсутствие механизма экологического наследования . Поскольку эта форма наследования не предполагает воспроизводства или экономического выживания, она не вписывается ни в одну из двух иерархий, оставляя концептуальный разрыв в теории. [ 19 ] Точно так же различные методы наследования, такие как эпигенетический и символический, которые присутствуют в других теориях эволюционной биологии, нарушают иерархическую структуру модели плещущегося ведра. Кроме того, более экстерналистская точка зрения, принятая в модели плещущегося ведра, хотя и отличается от адаптационизма , представляет собой еще одну концепцию, с которой некоторым биологам трудно согласиться. [ 19 ]
Эволюционный биолог Стивен Джей Гулд далее критикует произвольное и исключительно избирательное определение индивида и последующих групп, которые затем следуют из этих индивидов. [ 20 ] Биолог Дэвид Моррисон также указывает на запутанную природу сетевых взаимодействий со строгой иерархией. [ 21 ] И экологическая, и генеалогическая системы не являются строго иерархическими, и возникает вопрос, «в какой степени иерархии доминируют в сетевых связях». [ 21 ]
Приложения модели
[ редактировать ]Модель эволюции с плещущимся ведром была применена Дэниелом Бруксом для объяснения эволюционной биологии возникающих инфекционных заболеваний (ВИЗ). [ 22 ] Опубликованная в 2007 году, она использует модель плескающегося ведра для объяснения движущих причин этих EID, заявляя, что существуют «эволюционные катастрофы, ожидающие своего часа, требующие только катализатора изменения климата ». . .' [ 22 ] Поскольку региональные экологические нарушения становятся все более частыми, эпизодические вспышки новых мутировавших и потенциально более инфекционных и смертоносных заболеваний также станут более частыми.
Более того, было замечено, что именно генеалогическая иерархия является основой области эволюционной психологии. [ 23 ] Это объясняется тем, что социальные системы являются продуктом биологии, результатом которой является генеалогическая иерархия.
Ссылки
[ редактировать ]- ^ «Ведерное мышление: будущая основа эволюционного объяснения» . Исследовательские ворота . Проверено 11 августа 2019 г.
- ^ Войе, Кьетил Л.; Холен, Эйстейн Х.; Лиоу, Ли Сян; Стенсет, Нильс Хр. (07.06.2015). «Роль биотических сил в макроэволюции: за пределами Красной Королевы» . Труды Королевского общества B: Биологические науки . 282 (1808): 20150186. doi : 10.1098/rspb.2015.0186 . ПМЦ 4455800 . ПМИД 25948685 .
- ^ Майерс, Корин Э.; Саупе, Эрин Э. (2013). «Макроэволюционное расширение современного синтеза и важность внешних абиотических факторов» . Палеонтология . 56 (6). Уайли-Блэквелл : 1179–1198. Бибкод : 2013Palgy..56.1179M . дои : 10.1111/пала.12053 . S2CID 86272884 .
- ^ «Библиография» . Найлс Элдридж -- Палеонтология и эволюция . Проверено 10 апреля 2019 г.
- ^ Jump up to: а б с д и ж Кратчфилд, Джеймс Патрик. Шустер, Питер, 1939- ... (2003). Эволюционная динамика: исследование взаимодействия отбора, случайности, нейтральности и функции . Издательство Оксфордского университета. стр. 3–32. ISBN 0195142640 . OCLC 469426263 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) CS1 maint: числовые имена: список авторов ( ссылка ) - ^ «В какие три периода времени жили динозавры?» . Наука . Проверено 7 июня 2019 г.
- ^ Вейшампель, Дэвид Б.; Додсон, Питер; Осмолска, Гальшка (12 июня 2004 г.), «Введение», The Dinosauria , University of California Press, стр. 1–3, doi : 10.1525/california/9780520242098.003.0001 , ISBN 9780520242098
- ^ Jump up to: а б с Элдридж, Найлз (21 ноября 2007 г.). «Иерархии и плескающееся ведро: к унификации эволюционной биологии» . Эволюция: образование и информационно-пропагандистская деятельность . 1 (1): 10–15. дои : 10.1007/s12052-007-0007-6 . ISSN 1936-6426 .
- ^ «ПРИЛОЖЕНИЕ: Прерывистые равновесия: альтернатива филетическому градуализму», Time Frames , Princeton University Press, 1989-12-31, стр. 193–224, doi : 10.1515/9781400860296.193 , ISBN 9781400860296
- ^ Элдридж, Найлз (2001), «Возвращение к природе и происхождению надвидовых таксонов - с особым акцентом на трилобиту», Окаменелости, филогения и формы , Темы геобиологии, том. 19, Springer US, стр. 341–375, номер документа : 10.1007/978-1-4615-0571-6_10 , ISBN. 9781461351375
- ^ Динесиус, М.; Янссон, Р. (1 августа 2000 г.). «Эволюционные последствия изменений в географическом распространении видов, вызванных климатическими колебаниями Миланковича» . Труды Национальной академии наук . 97 (16): 9115–9120. Бибкод : 2000PNAS...97.9115D . дои : 10.1073/pnas.97.16.9115 . ISSN 0027-8424 . ПМК 16831 . ПМИД 10922067 .
- ^ Хехт, Макс К.; Элдридж, Найлз; Гулд, Стивен Джей (1974), «Морфологическая трансформация, летопись окаменелостей и механизмы эволюции: дебаты», Evolutionary Biology , Springer US, стр. 295–308, doi : 10.1007/978-1-4615-6944- 2_8 , ISBN 9781461569466
- ^ Бретт, Карлтон Э. (2012), «Пересмотр скоординированного стазиса: перспектива через пятнадцать лет», Земля и жизнь , Springer Нидерланды, стр. 23–36, doi : 10.1007/978-90-481-3428-1_2 , ISBN 9789048134274
- ^ Jump up to: а б Врба, Элизабет С.; Элдридж, Найлз (1984). «Люди, иерархии и процессы: к более полной эволюционной теории». Палеобиология . 10 (2): 146–171. Бибкод : 1984Pbio...10..146V . дои : 10.1017/s0094837300008149 . ISSN 0094-8373 . S2CID 86346683 .
- ^ Джонсон, Р.Г. (1972). «Концептуальные модели донных морских сообществ». Модели в палеобиологии : 148–159.
- ^ Рауп, ДМ (28 марта 1986 г.). «Биологическое вымирание в истории Земли» . Наука . 231 (4745): 1528–1533. Бибкод : 1986Sci...231.1528R . дои : 10.1126/science.11542058 . ПМИД 11542058 .
- ^ Элдридж, Найлз, редактор. Пиевани, Тельмо, редактор. Серрелли, Эмануэле, редактор. Темкин Илья, редактор. (23 сентября 2016 г.). Эволюционная теория: иерархическая перспектива . Издательство Чикагского университета. ISBN 9780226426198 . OCLC 1026792474 .
{{cite book}}:|last=имеет общее имя ( справка ) CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Либерман, Брюс С.; Врба, Элизабет С. (июнь 1995 г.). «Теория иерархии, отбор и сортировка». Бионаука . 45 (6): 394–399. дои : 10.2307/1312719 . hdl : 1808/9187 . ISSN 0006-3568 . JSTOR 1312719 . S2CID 54668531 .
- ^ Jump up to: а б Фабрегас-Техеда, Алехандро; Вергара-Сильва, Франциско (01 ноября 2017 г.). «Иерархическая теория эволюции и расширенный эволюционный синтез: некоторые эпистемические мосты, некоторые концептуальные разломы» . Эволюционная биология . 45 (2): 127–139. дои : 10.1007/s11692-017-9438-3 . ISSN 0071-3260 . S2CID 24287468 .
- ^ Даниэли, Джан Антонио (2013). Стивен Дж. Гулд: Научное наследие . Спрингер. ISBN 9788847054240 . OCLC 858012881 .
- ^ Jump up to: а б Моррисон, Дэвид А. (01 января 2018 г.). «Эволюционная теория: иерархическая перспектива. Под редакцией Найлза Элдриджа, Тельмо Пьевани, Эмануэле Серрелли и Ильи Темкина» . Систематическая биология . 67 (1): 175–177. дои : 10.1093/sysbio/syw120 . ISSN 1063-5157 .
- ^ Jump up to: а б Брукс, Дэниел Р.; Хоберг, Эрик П. (21 ноября 2007 г.). «Необходимое несоответствие Дарвина и плескающееся ведро: эволюционная биология новых инфекционных заболеваний» . Эволюция: образование и информационно-пропагандистская деятельность . 1 (1): 2–9. дои : 10.1007/s12052-007-0022-7 . hdl : 1807/87200 . ISSN 1936-6426 .
- ^ Капораэль, Линда Р.; Брюэр, Мэрилин Б. (январь 1995 г.). «Иерархическая эволюционная теория: альтернатива есть, и это не креационизм». Психологический опрос . 6 (1): 31–34. дои : 10.1207/s15327965pli0601_2 . ISSN 1047-840X .