Седореовирусиды
| Седореовирусиды | |
|---|---|
 | |
| Крио-ЭМ реконструкция ротавируса | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область : | Рибовирия |
| Королевство: | Орторнавиры |
| Тип: | Дуплорнавирикота |
| Сорт: | Ресентовирицетес |
| Заказ: | Реовирусы |
| Семья: | Седореовирусиды |
| Подсемейства и роды | |
Sedoreoviridae (ранее Reoviridae ) — семейство вирусов с двухцепочечной РНК . Вирусы-члены имеют широкий круг хозяев , включая позвоночных , беспозвоночных , растения, простейших и грибы. [1] У них нет липидной оболочки , и их сегментированный геном упакован в многослойные капсиды . Отсутствие липидной оболочки позволило трехмерные структуры этих крупных сложных вирусов (диаметром ~60–100 нм получить цистовирусов ), выявив структурное и, вероятно, эволюционное родство с семейством бактериофагов . [2] насчитывается 97 видов В настоящее время в этом семействе , разделенных на 15 родов и разделенных на два подсемейства. [3] Реовирусы могут поражать желудочно-кишечную систему (например, ротавирусы ) и дыхательные пути . [4] Название «рео-» является аббревиатурой от « кишечно респираторных сиротских - » вирусов . [5] Термин « вирус-сирота » относится к тому факту, что некоторые из этих вирусов не связаны с каким-либо известным заболеванием. Несмотря на то, что вирусы семейства Reoviridae совсем недавно были идентифицированы как вызывающие различные заболевания, первоначальное название все еще используется.
Реовирусные инфекции часто встречаются у людей, но в большинстве случаев они протекают в легкой или субклинической форме. ротавирусы Однако могут вызывать тяжелую диарею и кишечные расстройства у детей, а лабораторные исследования на мышах показали, что ортореовирусы участвуют в проявлении целиакии у предрасположенных к этому людей. [6] Вирус можно легко обнаружить в кале , а также выделить из глоточных или носовых выделений , мочи, спинномозговой жидкости и крови. Несмотря на легкость обнаружения реовирусов в клинических образцах, их роль в заболеваниях и лечении человека все еще неясна.
Некоторые вирусы этого семейства, например фитореовирусы и оризавирусы , поражают растения. Большинство реовирусов, инфицирующих растения, передаются между растениями насекомыми-переносчиками . Вирусы реплицируются как в растении, так и в насекомом, обычно вызывая заболевание растения, но не нанося вреда инфицированному насекомому или не нанося ему никакого вреда. [7] : 148
Структура
[ редактировать ]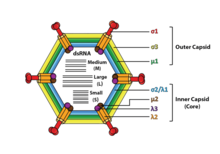
Реовирусы не имеют оболочки и имеют икосаэдрический капсид, состоящий из внешней ( Т =13) и внутренней (Т=2) белковых оболочек. [1] [8] Исследования ультраструктуры показывают, что капсиды вирионов состоят из двух или трех отдельных слоев, что зависит от вида вида. Самый внутренний слой (ядро) имеет икосаэдрическую симметрию T = 1 и состоит из 60 различных типов структурных белков. Ядро содержит сегменты генома, каждый из которых кодирует различную ферментную структуру, необходимую для транскрипции. Ядро покрыто капсидным слоем икосаэдрической симметрии Т=13. Реовирусы имеют уникальную структуру, которая содержит гликолизированный шиповидный белок на поверхности. [9]
Геном
[ редактировать ]Геномы вирусов семейства Reoviridae содержат 9–12 сегментов, которые сгруппированы в три категории, соответствующие их размеру: L (большие), M (средние) и S (маленькие). Размер сегментов варьируется от 0,2 до 3 т.п.н., и каждый сегмент кодирует 1–3 белка (всего 10–14 белков). [1] ). Белки вирусов семейства Reoviridae обозначаются греческим символом, соответствующим сегменту, из которого они были транслированы (сегмент L кодирует белки λ, сегмент M кодирует белки μ и сегмент S кодирует белки σ). [8]
Жизненный цикл
[ редактировать ]
Вирусы семейства Reoviridae имеют геномы, состоящие из сегментированной двухцепочечной РНК (дцРНК). [4] Из-за этого репликация происходит исключительно в цитоплазме, и вирус кодирует несколько белков, которые необходимы для репликации и преобразования генома дцРНК в РНК с положительным смыслом. [10]
Вирус может проникнуть в клетку-хозяина через рецептор на поверхности клетки. Рецептор неизвестен, но считается, что он включает сиаловую кислоту и молекулы соединительной адгезии (JAM). [10] Вирус частично не покрыт протеазами в эндолизосоме, где капсид частично переваривается, обеспечивая дальнейшее проникновение в клетку. Затем центральная частица попадает в цитоплазму посредством пока неизвестного процесса, при котором геном транскрибируется консервативно, вызывая избыток нитей с положительным смыслом, которые используются в качестве шаблонов информационной РНК для синтеза нитей с отрицательным смыслом. [10]
Геном ротавируса разделен на 11 сегментов. Эти сегменты связаны с молекулой VP1, которая отвечает за синтез РНК. На ранних этапах процесс отбора происходит так, что в клетку попадают 11 различных сегментов РНК. Эту процедуру выполняют вновь синтезированные РНК. Это событие гарантирует получение одного из 11 различных сегментов РНК. В поздних событиях процесс транскрипции происходит снова, но на этот раз не ограничен, в отличие от ранних событий. Для вируса требуется разное количество РНК, поэтому на этапе трансляции существует механизм контроля. Существует одинаковое количество сегментов РНК, но разное количество белков. Причина этого в том, что сегменты РНК транслируются с разной скоростью. [7]
Вирусные частицы начинают собираться в цитоплазме через 6–7 часов после заражения. Трансляция происходит путем негерметичного сканирования, подавления терминации и пропуска рибосом . Вирус покидает клетку-хозяина путем моночастного движения вируса, не направляемого канальцами, движения от клетки к клетке, а также существования в окклюзионных тельцах после гибели клетки и сохранения инфекционности до тех пор, пока не будет найден другой хозяин. [1]
| Род | Сведения о хосте | Тканевой тропизм | Детали входа | Подробности выпуска | Сайт репликации | Монтажный участок | Передача инфекции |
|---|
Реактивация множественности
[ редактировать ]Реактивация множественности (MR) — это процесс, посредством которого два или более вирусных генома, каждый из которых содержит инактивирующие повреждения генома, могут взаимодействовать внутри инфицированной клетки с образованием жизнеспособного вирусного генома. Макклейн и Спендлав [11] продемонстрировали МР для трех типов реовируса после воздействия ультрафиолетового облучения. В их экспериментах частицы реовируса подвергались воздействию УФ-излучения в дозах, которые были бы смертельными при единичных инфекциях. Однако когда двум или более инактивированным вирусам было позволено инфицировать отдельные клетки-хозяева, происходила МР и производилось жизнеспособное потомство. Как они заявили, реактивация множественности по определению включает в себя некоторый тип восстановления. Мишо и др. [12] рассмотрели многочисленные примеры MR у различных вирусов и предположили, что MR является распространенной формой полового взаимодействия у вирусов, которая обеспечивает рекомбинационное восстановление повреждений генома. [ нужна ссылка ]
Таксономия
[ редактировать ]Семейство Reoviridae делится на два подсемейства. [13] основан на наличии белка «башенки» на внутреннем капсиде. [14] [15] Из сообщений ICTV: «Название Spinareovirinae будет использоваться для идентификации подсемейства, содержащего вирусы с шипами или турелями, и происходит от слова «reovirus» и латинского слова «spina» в качестве префикса, что означает шип, обозначающий наличие шипов или турелей. на поверхности ядерных частиц. Термин «шипчатый» является альтернативой термину «башенчатый», который использовался в ранних исследованиях для описания структуры частиц, особенно с циповирусами. Название Sedoreovirinae будет использоваться для идентификации подсемейства. содержащий роды вирусов без турелей, и происходит от слова «реовирус» и латинского слова «sedo», что означает «гладкий», обозначая отсутствие шипов или турелей на основных частицах этих вирусов, которые имеют относительно гладкую морфологию». [16]

Семейство Reoviridae делится на следующие подсемейства и роды:
Терапевтическое применение
[ редактировать ]Хотя реовирусы в большинстве своем непатогенны для человека, эти вирусы послужили очень продуктивными экспериментальными моделями для изучения вирусного патогенеза . [17] Новорожденные мыши чрезвычайно чувствительны к реовирусным инфекциям и используются в качестве предпочтительной экспериментальной системы для изучения патогенеза реовирусов. [2]
Было продемонстрировано, что реовирусы обладают онколитическими (уничтожающими рак) свойствами, что поощряет разработку методов лечения рака на основе реовирусов. [18] [19] [20]
Реолизин представляет собой препарат реовируса ( штамм ортореовируса млекопитающих серотипа 3). [21] ), который в настоящее время проходит клинические испытания для лечения различных видов рака, [22] включая исследования, проводимые в настоящее время для изучения роли реолизина в сочетании с другими видами иммунотерапии. [21]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д «Вирусная зона» . ЭксПАСи . Проверено 15 июня 2015 г.
- ^ Перейти обратно: а б Гульельми, К.М.; Джонсон, EM; Штеле, Т; Дермоди, ТС (2006). «Прикрепление и проникновение в клетки ортореовируса млекопитающих». Реовирусы: вход, сборка и морфогенез . Актуальные темы микробиологии и иммунологии. Том. 309. стр. 1–38. дои : 10.1007/3-540-30773-7_1 . ISBN 978-3-540-30772-3 . ПМИД 16909895 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ «Таксономия вирусов: выпуск 2019 г.» . talk.ictvonline.org . Международный комитет по таксономии вирусов . Проверено 11 мая 2020 г.
- ^ Феннер, Ф. (июнь 1976 г.). «Классификация и номенклатура вирусов. Сводка результатов заседаний Международного комитета по таксономии вирусов в Мадриде, сентябрь 1975 г.» . Вирусология . 71 (2): 371–8. дои : 10.1016/0042-6822(76)90364-0 . ПМЦ 7131526 . ПМИД 820065 .
- ^ Бузиа, Р; и др. (7 апреля 2017 г.). «Реовирусная инфекция вызывает воспалительную реакцию на пищевые антигены и развитие целиакии» . Наука . 356 (6333): 44–50. Бибкод : 2017Sci...356...44B . дои : 10.1126/science.aah5298 . ПМК 5506690 . ПМИД 28386004 .
- ^ Перейти обратно: а б Картер, Джон; Сондерс, Венеция (2007). Вирусология: принципы и приложения . Западный Суссекс: Уайли. ISBN 978-0-470-02386-0 .
- ^ Перейти обратно: а б «Реовирусы» . МикробиологияBytes . Архивировано из оригинала 21 мая 2015 г.
- ^ Пейн С. (2017). «Семейство Reoviridae» . Вирусы : 219–226. дои : 10.1016/B978-0-12-803109-4.00026-X . ISBN 9780128031094 .
- ^ Перейти обратно: а б с Бартон, ES; Форрест, Джей Си; Коннолли, Дж.Л.; Чаппелл, доктор медицинских наук; Лю, Ю; Шнелл, Ф.Дж.; Нусрат, А; Паркос, Калифорния; Дермоди, Т.С. (9 февраля 2001 г.). «Молекула соединительной адгезии является рецептором реовируса» . Клетка . 104 (3): 441–51. дои : 10.1016/S0092-8674(01)00231-8 . ПМИД 11239401 .
- ^ Макклейн М.Э., Спендлав Р.С. (ноябрь 1966 г.). «Множественная реактивация реовирусных частиц после воздействия ультрафиолета» . Дж. Бактериол . 92 (5): 1422–9. дои : 10.1128/JB.92.5.1422-1429.1966 . ПМК 276440 . ПМИД 5924273 .
- ^ Мишо, RE; Бернштейн, Х.; Недельку, AM (2008). «Адаптационное значение пола у микробных патогенов». Инфекция, генетика и эволюция . 8 (3): 267–285. дои : 10.1016/j.meegid.2008.01.002 . ПМИД 18295550 .
- ^ Карстенс, Э.Б. (январь 2010 г.). «Ратификационное голосование по таксономическим предложениям Международного комитета по таксономии вирусов (2009 г.)» . Архив вирусологии . 155 (1): 133–146. дои : 10.1007/s00705-009-0547-x . ПМК 7086975 . ПМИД 19960211 .
- ^ Хилл С., Бут Т. и др. (1999). «Строение циповируса и функциональная организация дцРНК-вирусов». Структурная биология природы . 6 (6): 565–9. дои : 10.1038/9347 . ПМИД 10360362 . S2CID 28217302 .
- ^ Найп Д., Хоули П. и др. (2006). Поля вирусологии . Филадельфия, Пенсильвания: Уолтерс Клювер, Липпинкотт Уильямс и Уилкинс. п. 1855. ISBN 978-0-7817-6060-7 .
- ^ Аттуи, Хусам; Мертенс, Питер. «Шаблон таксономического предложения Исполнительному комитету ICTV по созданию нового подсемейства в существующем семействе» . Международный комитет по таксономии вирусов . 2007.127-129V.v2.Spina-Sedoreovirinae. стр. 1–9. Архивировано из оригинала 5 марта 2010 года.
- ^ Ачесон, Николас Х. Основы молекулярной вирусологии . Джон Уайли и сыновья (2011). стр.234
- ^ Канаи, Юта; Кобаяши, Такеши (29 сентября 2021 г.). «Быстрые белки: разработка и использование систем обратной генетики для вирусов реовирусов» . Ежегодный обзор вирусологии . 8 (1): 515–536. doi : 10.1146/annurev-virology-091919-070225 . ISSN 2327-056X . ПМИД 34586868 .
- ^ Лал Р., Харрис Д., Постель-Винай С. , де Боно Дж. (октябрь 2009 г.). «Реовирус: обоснование и обновленная информация о клинических испытаниях». Курс. Мнение. Мол. Там . 11 (5): 532–9. ПМИД 19806501 .
- ^ Келланд, К. (13 июня 2012 г.). «Вирус простуды пытается убить рак: исследование» . Рейтер . Проверено 17 июня 2012 г.
- ^ Перейти обратно: а б Бабикер, HM; Риаз, ИБ; Хуснейн, М.; Борад, MJ (февраль 2017 г.). «Онколитическая виротерапия, включая Ригвир, и стандартные методы лечения злокачественной меланомы» . Онколитическая виротерапия . 6 . Dovepress, Новая Зеландия NLM: 11–18. дои : 10.2147/OV.S100072 . ISSN 2253-1572 . ПМК 5308590 . ПМИД 28224120 . 101629828.
- ^ Тируккумаран С., Моррис Д.Г. (2009). «Онколитическая вирусная терапия с использованием реовируса». Генная терапия рака . Методы молекулярной биологии. Том. 542. стр. 607–34. дои : 10.1007/978-1-59745-561-9_31 . ISBN 978-1-934115-85-5 . ПМИД 19565924 .
Внешние ссылки
[ редактировать ]- ICTV: Reoviridae
- Описание вирусов растений: Reoviridae.
- ВиПР: Реовирусиды
- « Реовирусиды » . Браузер таксономии NCBI . 10880.
