Конкуренция (биология)

Конкуренция — это взаимодействие между организмами или видами , при котором оба требуют ресурса ограниченного еды (например, , воды или территории ) . [1] Конкуренция снижает приспособленность обоих участвующих организмов, поскольку присутствие одного из организмов всегда уменьшает количество ресурсов, доступных другому. [2]
При изучении экологии сообществ конкуренция внутри вида и между членами вида является важным биологическим взаимодействием . Конкуренция — один из многих взаимодействующих биотических и абиотических факторов, которые влияют на структуру сообщества , видовое разнообразие и динамику популяций (сдвиги популяции с течением времени). [3]
Существует три основных механизма конкуренции: вмешательство, эксплуатация и очевидная конкуренция (в порядке от наиболее прямого к наименее прямому). Вмешательство и конкуренция в эксплуатации могут быть классифицированы как «настоящие» формы конкуренции, в то время как очевидная конкуренция таковой не является, поскольку организмы не разделяют ресурс, а вместо этого разделяют хищника. [3] Конкуренция между представителями одного и того же вида называется внутривидовой конкуренцией , а конкуренция между особями разных видов — межвидовой конкуренцией .
Согласно принципу конкурентного исключения , виды, менее приспособленные к конкуренции за ресурсы, должны либо адаптироваться , либо вымереть , хотя конкурентное исключение редко встречается в естественных экосистемах. [3] Согласно эволюционной теории , конкуренция внутри видов и между видами за ресурсы важна для естественного отбора . Однако совсем недавно исследователи предположили, что эволюционное биоразнообразие позвоночных было обусловлено не конкуренцией между организмами, а адаптацией этих животных к колонизации пустого жизненного пространства; это называется гипотезой «комнаты для передвижения». [4]
Конкурс помех
[ редактировать ]
Во время интерференционной конкуренции, также называемой соревновательной конкуренцией, организмы взаимодействуют напрямую, борясь за ограниченные ресурсы. Например, крупные тли защищают места кормления на листьях тополя , выбрасывая более мелких тлей с лучших мест. Конкуренция самцов и самцов у благородных оленей во время гона является примером интерференционной конкуренции, возникающей внутри вида.
Вмешательная конкуренция происходит непосредственно между особями посредством агрессии, когда особи мешают добыванию пищи, выживанию и размножению других, или путем прямого предотвращения их физического закрепления в определенной части среды обитания. Пример этого можно увидеть между муравьем Novomessor Cockerelli и красными муравьями-жнецами , где первый мешает способности вторых добывать пищу, закупоривая входы в их колонии небольшими камнями. [5] Самцы шалашников , которые создают сложные конструкции, называемые беседками, для привлечения потенциальных партнеров, могут напрямую снижать приспособленность своих соседей, крадя украшения из их построек. [6]
У животных интерференционная конкуренция — это стратегия, которую в основном применяют более крупные и сильные организмы в среде обитания. Таким образом, популяции с высокой интерференционной конкуренцией имеют циклы поколений, управляемые взрослыми. [7] Поначалу рост молоди замедляется более крупными взрослыми конкурентами. Однако, как только молодые особи достигают взрослой жизни, у них начинается вторичный цикл роста. [7] Растения, с другой стороны, в первую очередь участвуют в конкуренции со своими соседями посредством аллелопатии или производства биохимических веществ. [8]
Вмешательную конкуренцию можно рассматривать как стратегию, имеющую очевидную цену (травму или смерть) и выгоду (получение ресурсов, которые достались бы другим организмам). [9] Чтобы справиться с сильной мешающей конкуренцией, другие организмы часто либо делают то же самое, либо вступают в конкуренцию за эксплуатацию. Например, в зависимости от сезона более крупные самцы копытных благородных оленей являются конкурентно доминантными из-за мешающей конкуренции. Однако самки и оленята справились с этим посредством временного разделения ресурсов — добывая пищу только в отсутствие взрослых самцов. [10]
Эксплуатационный конкурс
[ редактировать ]Конкуренция за эксплуатацию, или конкуренция за схватку , возникает косвенно, когда организмы используют общий ограничивающий ресурс или общий пищевой продукт. Вместо борьбы или проявления агрессивного поведения с целью завоевания ресурсов возникает эксплуататорская конкуренция, когда использование ресурсов одним организмом истощает общее количество, доступное другим организмам. Эти организмы, возможно, никогда не будут взаимодействовать напрямую, а будут конкурировать, реагируя на изменения уровня ресурсов. Очень очевидные примеры этого явления включают дневные виды и ночные виды, которые, тем не менее, используют одни и те же ресурсы, или растение, которое конкурирует с соседними растениями за свет, питательные вещества и пространство для роста корней. [11] [8]
Эта форма конкуренции обычно вознаграждает те организмы, которые первыми претендуют на ресурс. Таким образом, конкуренция за эксплуатацию часто зависит от размера, и предпочтение отдается более мелким организмам, поскольку более мелкие организмы обычно имеют более высокую скорость поиска пищи. [7] Поскольку более мелкие организмы имеют преимущество, когда в экосистеме важна эксплуататорская конкуренция, этот механизм конкуренции может привести к циклу генерации, управляемому молодью: отдельные молодые особи преуспевают и быстро растут, но когда они созревают, их вытесняют более мелкие организмы. [7]
У растений эксплуататорская конкуренция может возникать как над землей, так и под землей. Надземные растения снижают приспособленность своих соседей, конкурируя за солнечный свет. Растения потребляют азот его , поглощая своими корнями, делая азот недоступным для близлежащих растений. Растения, у которых много корней, обычно снижают содержание азота в почве до очень низкого уровня, что в конечном итоге приводит к гибели соседних растений.
Также было показано, что эксплуататорская конкуренция имеет место как внутри вида (внутривидовая), так и между разными видами (межвидовая). Более того, многие конкурентные взаимодействия между организмами представляют собой комбинацию эксплуататорской и интерференционной конкуренции, а это означает, что эти два механизма далеко не исключают друг друга. Например, недавнее исследование 2019 года показало, что местный вид трипса Frankliniella intonsa конкурентно доминировал над инвазивным видом трипса Frankliniella occidentalis, поскольку он не только уделял больше времени кормлению (эксплуатационная конкуренция), но и больше времени на охрану своих ресурсов (интерференционная конкуренция). [12] Растения также могут проявлять обе формы конкуренции, не только борясь за место для роста корней, но и напрямую подавляя развитие других растений посредством аллелопатии.
Очевидная конкуренция
[ редактировать ]Очевидная конкуренция возникает, когда два не связанных друг с другом вида добычи косвенно конкурируют за выживание через общего хищника . [13] Эта форма конкуренции обычно проявляется в новом равновесном количестве каждого вида добычи. Например, предположим, что существуют два вида (вид A и вид B), на которых охотятся хищники вида C, ограниченные в пище. Ученые наблюдают увеличение численности вида A и снижение численности вида B. очевидная модель конкуренции, установлено, что эти отношения опосредованы хищником C; популяционный взрыв вида A увеличивает численность хищников вида C из-за большего общего источника пищи. Поскольку хищников теперь стало больше, на виды A и B будут охотиться чаще, чем раньше. Таким образом, успех вида А был в ущерб виду Б — не потому, что они конкурировали за ресурсы, а потому, что их увеличение численности опосредованно влияло на популяцию хищников.
Эта модель «один хищник/две жертвы» была исследована экологами еще в 1925 году, но термин «очевидная конкуренция» был впервые введен экологом из Университета Флориды Робертом Д. Холтом в 1977 году. [13] [14] Холт обнаружил, что полевые экологи в то время ошибочно приписывали негативные взаимодействия между видами-жертвами разделению ниш и конкурентному исключению , игнорируя роль хищников с ограниченной пищей. [13]
Очевидная конкуренция и реализованная ниша
[ редактировать ]вида Очевидная конкуренция может помочь сформировать реализованную нишу или территорию или ресурсы, которые вид может фактически сохранить благодаря межвидовым взаимодействиям. Влияние на реализованные ниши может быть невероятно сильным, особенно при отсутствии более традиционного вмешательства или эксплуататорской конкуренции. Реальный пример был изучен в конце 1960-х годов, когда интродукция зайцев-снегоступов ( Lepus americanus ) в Ньюфаундленд сократила ареал обитания местных арктических зайцев ( Lepus arcticus ). В то время как некоторые экологи предположили, что это произошло из-за перекрытия ниш, другие экологи утверждали, что более правдоподобным механизмом было то, что популяции зайцев-беляков привели к взрывному росту популяций рыси , ограниченной в пище , общего хищника обоих видов добычи. Поскольку заяц-беляк обладает относительно более слабой защитной тактикой, чем заяц-беляк, он был исключен из лесных массивов по причине дифференциального нападения хищников. Однако как очевидная конкуренция, так и конкуренция за эксплуатацию могут в некоторой степени объяснить ситуацию. [13] Поддержка влияния конкуренции на широту реализованной ниши в отношении диеты становится все более распространенной в различных системах, основанных на изотопных и пространственных данных, включая как хищных, так и хищников. [15] и мелких млекопитающих. [16]
Асимметричная очевидная конкуренция
[ редактировать ]Явная конкуренция может быть симметричной или асимметричной. [17] Симметричная кажущаяся конкуренция одинаково негативно влияет на оба вида (-,-), из чего можно сделать вывод, что оба вида сохранятся. Однако асимметричная кажущаяся конкуренция возникает, когда один вид страдает меньше, чем другой. Самый крайний сценарий асимметричной очевидной конкуренции – это когда на один вид вообще не влияет увеличение численности хищника, что можно рассматривать как форму аменсализма (0, -). [18] Воздействие человека на виды, находящиеся под угрозой исчезновения, ученые-экологи охарактеризовали как крайнюю форму асимметричной очевидной конкуренции, часто посредством внедрения видов-хищников в экосистемы или субсидий на ресурсы. Примером явно асимметричной конкуренции, которая часто возникает вблизи городских центров, являются субсидии в виде человеческого мусора или отходов. В начале 2000-х годов популяция обыкновенного ворона ( Corvus corax ) в пустыне Мохаве увеличилась из-за притока человеческого мусора, что привело к косвенному негативному воздействию на молодь пустынных черепах ( Gopherus agassizii ). [19] Асимметрия в явной конкуренции может также возникнуть вследствие конкуренции за ресурсы. Эмпирическим примером служат два вида мелких рыб в послеледниковых озерах Западной Канады, где конкуренция за ресурсы между колючим бычком и трехиглой колюшкой приводит к смещению пространственной ниши преимущественно у трехиглой колюшки. [20] В результате этого сдвига хищничество со стороны общего хищника форели увеличивается для колюшки, но уменьшается для бычка в озерах, где эти два вида встречаются одновременно, по сравнению с озерами, в которых каждый вид встречается сам по себе вместе с хищниками форели. Поскольку совместное использование хищников часто сопровождается конкуренцией за общие пищевые ресурсы, очевидная конкуренция и конкуренция за ресурсы часто могут взаимодействовать в природе. [21]
Явная конкуренция в микробиоме человека
[ редактировать ]Очевидная конкуренция также наблюдалась внутри и на человеческом теле. Иммунная система человека может действовать как универсальный хищник, а высокая численность определенных бактерий может вызвать иммунный ответ, повреждающий все патогены в организме. Другой пример — две популяции бактерий, которые могут поддерживать хищный бактериофаг. В большинстве ситуаций тот, кто наиболее устойчив к заражению общим хищником, заменит другого. [17]
Очевидная конкуренция также рассматривается как явление, которое можно использовать при лечении рака. Узкоспециализированные вирусы, разработанные для поражения злокачественных раковых клеток, часто локально вымирают, прежде чем искоренить весь рак. Однако если бы был разработан вирус, который в некоторой степени нацелен как на здоровые, так и на нездоровые клетки-хозяева, большое количество здоровых клеток будет поддерживать хищнический вирус достаточно долго, чтобы уничтожить все злокачественные клетки. [17]
Асимметричная конкуренция по размеру
[ редактировать ]Конкуренция может быть либо полностью симметричной (все особи получают одинаковое количество ресурсов, независимо от их размера), совершенно симметричной по размеру (все особи используют одинаковое количество ресурсов на единицу биомассы), либо абсолютно асимметричной по размеру (самые крупные особи эксплуатируют все доступный ресурс).
Среди растений асимметрия размеров зависит от контекста, и конкуренция может быть как асимметричной, так и симметричной в зависимости от наиболее ограничивающего ресурса. В лесных насаждениях подземная конкуренция за питательные вещества и воду симметрична по размеру, поскольку корневая система дерева обычно пропорциональна биомассе всего дерева. [22] И наоборот, надземная конкуренция за свет асимметрична по размеру — поскольку свет имеет направленность, в пологе леса полностью доминируют самые большие деревья. Эти деревья непропорционально используют большую часть ресурсов своей биомассы, что делает размер взаимодействия асимметричным. [23] Независимо от того, являются ли надземные или подземные ресурсы более ограниченными, это может оказать серьезное влияние на структуру и разнообразие экологических сообществ; например, в смешанных буковых насаждениях асимметричная по размеру конкуренция за свет является более сильным предиктором роста по сравнению с конкуренцией за почвенные ресурсы. [24]
Внутри и между видами
[ редактировать ]
Конкуренция может возникать между особями одного и того же вида, называемая внутривидовой конкуренцией, или между разными видами, называемая межвидовой конкуренцией. Исследования показывают, что внутривидовая конкуренция может регулировать динамику популяции (изменение численности популяции с течением времени). Это происходит потому, что по мере роста населения людям становится скучно. Поскольку индивидуумам в популяции требуются одни и те же ресурсы, скученность приводит к тому, что ресурсы становятся более ограниченными. Некоторые особи (обычно маленькие молодые особи) в конечном итоге не приобретают достаточно ресурсов и умирают или не размножаются. Это уменьшает численность населения и замедляет его рост. [ нужна ссылка ]
Виды также взаимодействуют с другими видами, которым требуются те же ресурсы. Следовательно, межвидовая конкуренция может одновременно изменять размеры популяций многих видов. Эксперименты показывают, что когда виды конкурируют за ограниченный ресурс, один вид в конечном итоге приводит к вымиранию популяций других видов. Эти эксперименты показывают, что конкурирующие виды не могут сосуществовать (они не могут жить вместе на одной территории), поскольку лучший конкурент будет исключать все другие конкурирующие виды. [ нужна ссылка ]
Внутривидовой
[ редактировать ]Внутривидовая конкуренция возникает, когда представители одного и того же вида конкурируют за одни и те же ресурсы в экосистеме. [25] Простой пример — насаждение из растений одного возраста, расположенных на равном расстоянии друг от друга. Чем выше плотность растений , тем больше растений будет присутствовать на единицу площади земли и тем сильнее будет конкуренция за такие ресурсы, как свет, вода или питательные вещества.
Межвидовой
[ редактировать ]Межвидовая конкуренция может возникнуть, когда особи двух отдельных видов используют ограниченный ресурс на одной и той же территории. Если ресурс не может поддерживать обе популяции, то снижение плодовитости , роста или выживаемости может привести как минимум к одному виду. Межвидовая конкуренция может изменить популяции , сообщества и эволюцию взаимодействующих видов. Примером среди животных могут быть гепарды и львы ; поскольку оба вида питаются одинаковой добычей, присутствие другого отрицательно влияет на них, поскольку у них будет меньше еды, однако они по-прежнему сохраняются вместе, несмотря на предсказание, что в условиях конкуренции один вытеснит другой. Фактически, львы иногда крадут добычу, убитую гепардами. Потенциальные конкуренты также могут убивать друг друга, используя так называемое « внутригильдейское хищничество ». Например, в южной Калифорнии койоты часто убивают и поедают серых лисиц и рысей, причем все три хищника делят одну и ту же стабильную добычу (мелких млекопитающих). [26]
Примером простейших являются Paramecium aurelia и Paramecium caudatum . Русский эколог Георгий Гаузе изучал конкуренцию между двумя видами Paramecium , возникшую в результате их сосуществования. В ходе своих исследований Гаузе предложил принцип конкурентного исключения , наблюдая за конкуренцией, которая возникает, когда их разные экологические ниши перекрываются. [27]
Конкуренция наблюдалась между особями, популяциями и видами, но мало свидетельств того, что конкуренция была движущей силой эволюции больших групп. Например, млекопитающие жили рядом с рептилиями в течение многих миллионов лет, но не могли получить конкурентное преимущество, пока динозавры не были опустошены мел-палеогеновым вымиранием . [4]
Эволюционные стратегии
[ редактировать ]В эволюционном контексте конкуренция связана с концепцией теории отбора r/K, которая относится к отбору черт , способствующих успеху в определенных условиях. Теория берет свое начало в работах по биогеографии островов экологов Роберта Макартура и Э.О. Уилсона . [28]
В теории отбора r/K предполагается, что селективное давление ведет эволюцию в одном из двух стереотипных направлений: r- или K -отбор. [29] Эти термины r и K взяты из стандартной экологической алгебры , как показано в простом уравнении Ферхюльста динамики численности населения : [30]

где r — темп роста населения ( — N ), а K несущая способность местной окружающей среды. Обычно виды, отобранные с помощью r, используют пустые ниши и производят много потомков , каждое из которых имеет относительно низкую вероятность дожить до взрослого возраста. Напротив, виды, отобранные K-отбором, являются сильными конкурентами в густонаселенных нишах и вкладывают больше средств в гораздо меньшее количество потомков, каждое из которых имеет относительно высокую вероятность дожить до взрослого возраста. [30]
Принцип конкурентного исключения
[ редактировать ]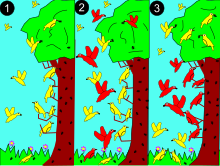
2: более крупный (красный) вид конкурирует за ресурсы.
3: красный цвет доминирует в середине для более обильных ресурсов. Желтый адаптируется к новым нишам, избегая конкуренции.
Чтобы объяснить, как сосуществуют виды, в 1934 году Георгий Гаузе предложил принцип конкурентного исключения , который также называют принципом Гаузе: виды не могут сосуществовать, если они занимают одну и ту же экологическую нишу . Слово «ниша» относится к потребностям вида для выживания и размножения. Эти требования включают как ресурсы (например, пищу), так и надлежащие условия среды обитания (например, температуру или pH). Гаус рассуждал, что если бы у двух видов были одинаковые ниши (требовались одинаковые ресурсы и среда обитания), они попытались бы жить на одной и той же территории и конкурировали бы за одни и те же ресурсы. Если бы это произошло, вид, который был лучшим конкурентом, всегда исключал бы своих конкурентов из этой области. Следовательно, чтобы сосуществовать, виды должны иметь хотя бы несколько разные ниши. [31] [32]
Смещение персонажа
[ редактировать ]
Конкуренция может привести к появлению у видов различий в признаках. Это происходит потому, что особи вида, имеющие признаки, сходные с конкурирующими видами, всегда испытывают сильную межвидовую конкуренцию. У этих особей меньше воспроизводства и выживаемости, чем у особей с признаками, отличающимися от их конкурентов. Следовательно, они не принесут много потомства будущим поколениям. Например, дарвиновы зяблики можно встретить поодиночке или вместе на Галапагосских островах . В популяциях обоих видов на самом деле больше особей с клювами среднего размера, когда они живут на островах без присутствия других видов. Однако, когда оба вида присутствуют на одном острове, конкуренция между особями, имеющими клювы среднего размера у обоих видов, становится острой, поскольку всем им требуются семена среднего размера. Следовательно, особи с маленькими и большими клювами имеют большую выживаемость и размножение на этих островах, чем особи с клювами среднего размера. Разные виды вьюрков могут сосуществовать, если у них есть особенности (например, размер клюва), которые позволяют им специализироваться на определенных ресурсах. Когда Geospiza fortis и Geospiza fuliginosa присутствуют на одном и том же острове, G. fuliginosa имеет тенденцию развивать небольшой клюв, а G. fortis - большой клюв. Наблюдение о том, что черты конкурирующих видов более различаются, когда они живут на одной и той же территории, чем когда конкурирующие виды живут на разных территориях, называется смещением характера. У двух видов вьюрков размер клюва изменился: у одного вида клювы стали меньше, а у другого больше. Исследования смещения характера важны, поскольку они доказывают, что конкуренция важна для определения экологических и эволюционных закономерностей в природе. [33]
См. также
[ редактировать ]- Биологическое взаимодействие
- Смещение персонажа
- Сообщество
- Минимальная жизнеспособная популяция
- Схватка соревнований
- Ресурс (биология)
- Разделение ресурсов
Ссылки
[ редактировать ]- ^ Бегон, М.; Харпер, Дж.Л.; Таунсенд, Ч.Р. (1996) Экология: Индивиды, популяции и сообщества Blackwell Science.
- ^ "Соревнование" . globalchange.umich.edu . Проверено 8 декабря 2021 г.
- ^ Перейти обратно: а б с «Взаимодействие видов и конкуренция | Изучайте науку в Scitable» . www.nature.com . Проверено 8 декабря 2021 г.
- ^ Перейти обратно: а б Сахни, Сарда; Бентон, Майкл Дж.; Ферри, Пол А. (23 августа 2010 г.). «Связь между глобальным таксономическим разнообразием, экологическим разнообразием и распространением позвоночных на суше» . Письма по биологии . 6 (4): 544–547. дои : 10.1098/rsbl.2009.1024 . ПМК 2936204 . ПМИД 20106856 .
- ^ Бартон, Кейси Э.; Сандерс, Натан Дж.; Гордон, Дебора М. (1 октября 2002 г.). «Влияние близости и возраста колонии на межвидовую интерференционную конкуренцию между пустынными муравьями Pogonomyrmex barbatus и Aphaenogaster Cockerelli» . Американский натуралист из Мидленда . 148 (2): 376–382. doi : 10.1674/0003-0031(2002)148[0376:TEOPAC]2.0.CO;2 . ISSN 0003-0031 . S2CID 7668877 .
- ^ Борджиа, Джеральд (1985). «Разрушение беседки и сексуальная конкуренция у атласного шалашника (Ptilonorhynchus violaceus)» . Поведенческая экология и социобиология . 18 (2): 91–100. дои : 10.1007/BF00299037 . ISSN 0340-5443 . JSTOR 4599867 . S2CID 36871646 .
- ^ Перейти обратно: а б с д Ле Бурло, Винсент; Талли, Томас; Классен, Дэвид (01 ноября 2014 г.). «Вмешательство против эксплуататорской конкуренции в регулирование численности населения с разбивкой по размеру» . Американский натуралист . 184 (5): 609–623. дои : 10.1086/678083 . ISSN 0003-0147 . ПМИД 25325745 . S2CID 206002300 .
- ^ Перейти обратно: а б Шенк, Х. Йохен (24 марта 2006 г.). «Коренная конкуренция: за пределами истощения ресурсов: Коренная конкуренция: за пределами истощения ресурсов» . Журнал экологии . 94 (4): 725–739. дои : 10.1111/j.1365-2745.2006.01124.x . S2CID 86320966 .
- ^ Кейс, Тед Дж.; Гилпин, Майкл Э. (август 1974 г.). «Интерференционная конкуренция и теория ниш» . Труды Национальной академии наук Соединенных Штатов Америки . 71 (8): 3073–3077. Бибкод : 1974PNAS...71.3073C . дои : 10.1073/pnas.71.8.3073 . ISSN 0027-8424 . ПМЦ 388623 . ПМИД 4528606 .
- ^ Стоун, Дэвид Б.; Мартин, Джеймс А; Коэн, Брэдли С; Пребил, Томас Дж; Киллмастер, Чарли; Миллер, Карл V (06 июля 2018 г.). «Внутривидовое распределение временных ресурсов в местах кормления белохвостых оленей» . Современная зоология . 65 (2): 139–146. дои : 10.1093/cz/zoy051 . ISSN 2396-9814 . ПМК 6430969 . ПМИД 30936902 .
- ^ Дженсен, Алабама (1 февраля 1987 г.). «Простые модели эксплуататорской и мешающей конкуренции» . Экологическое моделирование . 35 (1): 113–121. Бибкод : 1987EcMod..35..113J . дои : 10.1016/0304-3800(87)90093-7 . hdl : 2027.42/26823 . ISSN 0304-3800 .
- ^ Бхуайн, Мохаммад Мошароф Хоссейн; Лим, Ун Тэк (14 июня 2019 г.). «Конкуренция по вмешательству и эксплуатации между Frankliniella occidentalis и F. intonsa (Thysanoptera: Thripidae) в лабораторных анализах» . Флоридский энтомолог . 102 (2): 322–328. дои : 10.1653/024.102.0206 . ISSN 0015-4040 . S2CID 196662034 .
- ^ Перейти обратно: а б с д Холт, Роберт Д. (1 октября 1977 г.). «Хищничество, очевидная конкуренция и структура сообществ добычи» . Теоретическая популяционная биология . 12 (2): 197–229. дои : 10.1016/0040-5809(77)90042-9 . ISSN 0040-5809 . ПМИД 929457 .
- ^ Шрайбер, Себастьян Дж.; Крживань, Властимил (01.06.2020). «Холт (1977) и очевидная конкуренция» . Теоретическая популяционная биология . Пятьдесят лет теоретической популяционной биологии. 133 : 17–18. дои : 10.1016/j.tpb.2019.09.006 . ISSN 0040-5809 . ПМИД 31711965 . S2CID 207952477 .
- ^ Фаулер, Нидерланды; Петроэлье, ТР; Каутц, ТМ; Свобода, Нью-Джерси; Дюкетт, Дж. Ф.; Келлнер, К.Ф.; Бейер, Делавэр; Белант, Джей Джей (2022). «Переменное влияние волков на ширину ниши и плотность конкурентов внутри гильдии» . Экология и эволюция . 12 (2): e8542. Бибкод : 2022EcoEv..12E8542F . дои : 10.1002/ece3.8542 . ПМЦ 8829107 . ПМИД 35154647 .
- ^ Аллен, AG; Рёрс, З.П.; Севилья, РС; Ланье, ХК (2022). «Конкурентный выпуск во время пожарной сукцессии влияет на экологический оборот в сообществе мелких млекопитающих» . Экология . 103 (8): 1–12. Бибкод : 2022Ecol..103E3733A . дои : 10.1002/ecy.3733 . ПМЦ 9891167 . ПМИД 35430726 .
- ^ Перейти обратно: а б с Холт, Роберт Д.; Бонсолл, Майкл Б. (2 ноября 2017 г.). «Явная конкуренция» . Ежегодный обзор экологии, эволюции и систематики . 48 (1): 447–471. doi : 10.1146/annurev-ecolsys-110316-022628 . ISSN 1543-592X .
- ^ Шанетон, Энрике Дж.; Бонсолл, Майкл Б. (2000). «Очевидная конкуренция, опосредованная противником: эмпирические закономерности и доказательства» . Ойкос . 88 (2): 380–394. Бибкод : 2000Oikos..88..380C . дои : 10.1034/j.1600-0706.2000.880217.x . ISSN 0030-1299 . JSTOR 3547034 .
- ^ Де Чезаре, Нью-Джерси; Хебблуайт, М.; Робинсон, HS; Мусиани, М. (2010). «По всей видимости, находящиеся под угрозой исчезновения: роль очевидной конкуренции в сохранении исчезающих видов» . Охрана животных . 13 (4): 353–362. Бибкод : 2010AnCon..13..353D . дои : 10.1111/j.1469-1795.2009.00328.x . ISSN 1469-1795 . S2CID 83416826 .
- ^ Роести, М., Гро, Дж. С., Блейн, С. А., Хусс, М., Рассиас, П. и Болник, Д. И. и др. (2023) Дивергенция видов в условиях конкуренции и совместного хищничества. Письма об экологии, 26, 111–123. Доступно по адресу: https://doi.org/10.1111/ele.14138.
- ^ Роести, М., Гро, Дж. С., Блейн, С. А., Хусс, М., Рассиас, П. и Болник, Д. И. и др. (2023) Дивергенция видов в условиях конкуренции и совместного хищничества. Письма об экологии, 26, 111–123. Доступно по адресу: https://doi.org/10.1111/ele.14138.
- ^ Уэст, ПВ; Ратковский, Д.А. (04 октября 2021 г.). «Проблемы с моделями, оценивающими влияние размера дерева и конкурентных процессов между деревьями на рост отдельных деревьев: поучительная история» . Журнал лесных исследований . 33 (2): 565–577. дои : 10.1007/s11676-021-01395-9 . ISSN 1993-0607 . S2CID 244202914 .
- ^ Преч, Ганс; Бибер, Питер (15 февраля 2010 г.). «Симметричная по размеру и асимметричная по размеру конкуренция и разделение роста деревьев в лесных насаждениях вдоль экологического градиента в Центральной Европе» . Канадский журнал лесных исследований . 40 (2): 370–384. дои : 10.1139/x09-195 . ISSN 0045-5067 .
- ^ дель Рио, Мирен; Кондес, Соня; Претч, Ганс (1 августа 2014 г.). «Анализ размерно-симметричной и асимметричной, а также внутривидовой и межвидовой конкуренции в смешанных насаждениях бука (Fagus sylvatica L.)» . Лесная экология и управление . 325 : 90–98. дои : 10.1016/j.foreco.2014.03.047 .
- ^ Таунсенд, Колин Р.; Бегон, Майкл (2008). Основы экологии . стр. 103–105. ISBN 978-1-4051-5658-5 .
- ^ .Федриани, Дж. М., Т. К. Фуллер, Р. М. Соважо и ЕС Йорк. 2000. Конкуренция и внутригильдийное хищничество среди трех симпатрических хищников. Экология , 125:258-270.
- ^ Гаузе, Г. Ф. (1934). Борьба за существование . Балтимор, Мэриленд: Уильямс и Уилкинс.
- ^ Макартур, Р. и Уилсон, Э.О. (1967). Теория биогеографии островов , Princeton University Press (переиздание 2001 г.), ISBN 0-691-08836-5 .
- ^ Пианка, ER (1970). О выборе r и K. Американский натуралист «104» , 592–597.
- ^ Перейти обратно: а б Верхюльст, П.Ф. (1838 г.). Обратите внимание на закон, что население продолжает расти. Корр. Математика. Физ. «10» , 113–121.
- ^ Хардин, Гаррет (1960). «Принцип конкурентного исключения» (PDF) . Наука . 131 (3409): 1292–1297. Бибкод : 1960Sci...131.1292H . дои : 10.1126/science.131.3409.1292 . ПМИД 14399717 . Архивировано из оригинала (PDF) 17 ноября 2017 г. Проверено 10 января 2017 г.
- ^ Пошевиль, Арно (2015). «Экологическая ниша: история и недавние противоречия» . В Химсе, Томас; Хьюнеман, Филипп; Лекуантр, Гийом; и др. (ред.). Справочник по эволюционному мышлению в науке . Дордрехт: Спрингер. стр. 547–586. ISBN 978-94-017-9014-7 .
- ^ Браун, В.Л. и Э.О. Уилсон . 1956. «Перемещение характера». Систематическая зоология 5:49–65.
Внешние ссылки
[ редактировать ]- http://www.encyclopedia.com/topic/Competition_%28Biology%29.aspx
- Биологическая основа «ощущения победы»
| Базы данных органов управления : Национальные |
|---|