Камаразавр
| Камаразавр Временной диапазон: поздняя юра ,
| |
|---|---|

| |
| Установленный слепок скелета в Музее юрского периода в Астурии. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | † Зауроподоморфа |
| Клэйд : | † Зауропода |
| Клэйд : | † Макронария |
| Семья: | † Камаразавриды |
| Подсемейство: | † Камаразаврины Коуп, 1878 г. |
| Род: | † Камаразавр Коуп, 1877а. |
| Типовой вид | |
| † Камаразавр верховный Коуп, 1877а.
| |
| Другие виды | |
| |
| Синонимы | |
| |
Камаразавр ( / ˌ k æ m ər ə ˈ s ɔː r ə s / KAM -ər-ə- травоядных динозавров и является наиболее SOR -əs ) — род четвероногих распространенным зауроподом ископаемым в Северной Америке . Его ископаемые останки были найдены в формации Моррисон , относящейся к поздней юрской эпохе ( от кимериджа до титона ), между 155 и 145 миллионами лет назад.
Камаразавр имел характерный черепной профиль с тупой мордой и дугообразным черепом, который был удивительно квадратным, типичным для базальных макронорианцев .
Название означает «камерная ящерица», имея в виду полые камеры, известные как плевроцели , в ее шейных позвонках ( греческое καμαρα ( камара ), означающее «сводчатая камера», или что-либо с арочной крышкой, и σαυρος ( саурос ), означающее «ящерица» ).
Камаразавр включает четыре вида, которые обычно признаны действительными: Camarasaurus grandis , Camarasaurus lentus , Camarasaurus lewisi и Camarasaurus supremus . C. supremus , типовой вид , является самым крупным и геологически самым молодым из четырех. Camarasaurus — типовой род Camarasauridae , в который также входит его европейский близкий родственник Lourinhasaurus .
Камаразавр был назван в 1877 году Эдвардом Дринкером Коупом , в период научного соперничества между ним и Отниэлем Чарльзом Маршем, известного как « Костяные войны» . Вскоре после этого Марш назвал род Morosaurus , но впоследствии было показано, что он является синонимом Camarasaurus .
История
[ редактировать ]Первоначальное открытие
[ редактировать ]Первое упоминание о камаразавре относится к весне 1877 года, когда г-н Орамель Уильям Лукас из Каньон-Сити , штат Колорадо, обнаружил несколько крупных позвонков в Гарден-Парке , которые он отправил Эдварду Дринкеру Коупу , который жил в Филадельфии, штат Пенсильвания . [ 1 ] Исходный присланный материал состоял из частичного шейного позвонка, который впоследствии стал тезкой таксона, трех спинных позвонков и четырех хвостовых позвонков. [ 2 ] [ 1 ] Сейчас считается, что этот экземпляр состоял из нескольких особей. [ 2 ] На основе этих первоначальных фрагментарных останков Коуп сделал свое первоначальное описание Camarasaurus supremus («ящерица с высшими камерами») и основал этот род; эти останки сейчас находятся в Американском музее естественной истории под каталожным номером AMNH 560. [ 2 ] Получив оригинальные кости, Коуп нанял коллекционеров, которые собрали больше материала, описанного в 1921 году Генри Осборном и Чарльзом Муком. [ 2 ]

Количество материала было велико, он состоял из нескольких перемешанных частичных скелетов. [ 2 ] Не все было подготовлено сразу, но значительная часть была очищена Якобом Гейсмаром под руководством Коупа на протяжении 1870-1890-х годов. [ 2 ] В 1877 году реконструкция скелета Камаразавра была нарисована доктором Джоном Райдером на нескольких полотнах под руководством профессора Коупа, который использовал их в лекциях, чтобы произвести впечатление на свою аудиторию. [ 2 ] Эта реконструкция должна была стать первой в истории реконструкцией динозавра-зауропода, иметь естественный размер и представлять собой останки ряда особей. Реконструкция имела длину более пятидесяти футов. Коллекционеры Коупа прислали дополнительные материалы с 1877 по 1878 год. [ 2 ] [ 3 ] и по мере того, как Коуп получал больше материала, он называл таксоны на основе этих недавно присланных останков. Большинство этих дополнительных таксонов сейчас считаются сомнительными или синонимами камаразавра . [ 3 ] К концу сбора в Гарден-Парке было обнаружено как минимум четыре человека и несколько сотен костей почти из каждой части скелета. [ 2 ]
Комо-Блафф находит морозавра
[ редактировать ]Следующее открытие камаразавра произошло позже, в 1877 году, когда фрагментарный задний череп и частичный посткраниальный скелет были обнаружены и собраны в Карьере 1, Комо-Блафф , Вайоминг, бригадами, работавшими на Отниэля Чарльза Марша . Этот скелет был бы наиболее хорошо сохранившейся отдельной особью камаразавра на то время, и в 1877 году он был назван новым видом апатозавра. Образец не был полностью собран до 1879 года, и он содержит большую часть скелета молодого человека (голотип YPM 1901). ) [ 4 ] создал новый вид Amphicoelias Тем временем команды, работавшие на Эдварда Коупа в Гарден-Парке, собрали фрагментарный экземпляр, состоящий из бедренной кости и двух хвостовых позвонков, и Коуп , который он назвал Amphicoelias latus в 1877 году. [ 5 ] Этот вид был предварительно синонимом C. supremus в 1921 году. В 1998 году Кеннет Карпентер утверждал, что стратиграфическое положение находки предполагает, что он, скорее всего, является синонимом C. grandis . [ 6 ] Однако в исследовании биостратиграфического распространения Camarasaurus в 2005 году Такехито Икеджири сохранил его в качестве синонима C. supremus . [ 7 ] В 1878 году крестец зауроподов был обнаружен вместе с несколькими другими беспорядочными посткраниальными элементами зауроподов, снова в Комо-Блафф. Останки также были отправлены в Марш, а в 1878 году крестец был отнесен к новому роду и виду Morosaurus impar («непарная глупая ящерица»). В конце 19 века морозавр получил несколько новых видов, а в 1892 году даже стал частью нового семейства Morosauridae . Большинство видов морозавров сейчас считаются сомнительными. [ 8 ] включая типовой вид, или переклассифицированы. [ 8 ] В 1889 году новый вид морозавра был назван на основе частичного черепа и скелета из Комо-Блафф. Morosaurus lentus — название скелета (голотип YPM 1910). [ 7 ] а скелет был установлен в зале окаменелостей Йельского музея Пибоди в 1930 году.
Находки «Второй гонки динозавров»
[ редактировать ]
В конце 1890-х годов Американский музей естественной истории и Полевой музей обнаружили дополнительные материалы о морозаврах в Комо-Блафф и Фрута соответственно. Новый материал морозавров , в основном состоящий из материала конечностей, привел к новым реконструкциям зауроподов . структуры кисти и стопы [ 9 ] [ 10 ] AMNH сделал важное открытие в 1899 году в своем карьере Bone Cabin в Вайоминге, открыв первый полный череп и нижнюю челюсть камаразавра с соответствующими шейными позвонками. [ 11 ] Основная переоценка морозавров и камаразавров произошла в 1901 году. Переоценка, проведенная Элмером Риггсом, пришла к выводу, что из пяти видов морозавров, названных Маршем, только три действительны. [ 10 ] Morosaurus grandis , Morosaurus lentus и Morosaurus agilis (теперь известный как Smitanosaurus ) были признаны действительными, причем Morosaurus нечетный синоним M. grandis. Возможная синонимия между морозавром и камаразавром была также предложена Риггсом. [ 10 ] В 1905 году первый установленный скелет зауропода был установлен в AMNH бронтозавра . Череп животного, как известно, был основан на материале, который, вероятно, принадлежал камаразавру из Комо-Блафф. [ 12 ]
В 1909 году в Музее Карнеги был камаразавром , теперь под номером экземпляра CM 11338. Примечательно, что экземпляр был найден в позе смерти и выставлен на видном месте в зале Музея Карнеги. обнаружен почти полный скелет подростка, обнаруженный [ 13 ] Эрл Дуглас обнаружил этот образец, и он был собран с 1909 по 1910 год командой Музея Карнеги, работавшей в Национальном памятнике динозавров . Этот скелет не был описан до 1925 года Чарльзом У. Гилмором. Этот экземпляр был отнесен к Camarasaurus lentus . Скелет — один из лучших известных экземпляров зауроподов, в котором почти каждый элемент сохранился в сочленении, включая хрупкие шейные позвонки. [ 13 ]
Еще один скелет камаразавра был найден в 1918 году, снова в Национальном памятнике динозавров , командой Карнеги. Этот экземпляр можно увидеть в Национальном музее естественной истории . Образец, известный как USNM V 13786, был продан USNM в 1935 году, а подготовительные работы над ним начались в 1936 году на выставке столетия Техаса в Далласе , где его могли увидеть посетители мероприятия. [ 14 ] Подготовительные работы продолжались до 1947 года, когда скелет был установлен в мертвой позе в зале окаменелостей. [ 14 ] USNM Камаразавр также упоминается как C. lentus . [ 15 ] [ 7 ]
В 1919 году У. Дж. Холланд назвал Uintasaurus douglassi, основываясь на другом экземпляре зауропода из DNM, который был обнаружен Музеем Карнеги в 1909 году. [ 16 ] Типовой экземпляр был неполным и состоял из 5 передних шейных позвонков. [ 16 ] и является синонимом Camarasaurus lentus. [ 7 ] Дополнительный материал камаразавра был найден недалеко от Черной Мезы в западной Оклахоме в 1940-х годах. [ 17 ] и был отнесен к Camarasaurus supremus. [ 7 ] , материал состоит из множества крупных позвонков и некоторых элементов черепа. [ 7 ]
Возрождающиеся открытия
[ редактировать ]
не произошло Никаких серьезных открытий в отношении камаразавра до тех пор, пока в 1967 году Джеймс Дженсен не собрал хорошо сохранившийся и сочлененный частичный посткраниальный скелет, включая большую часть позвоночного столба, на холме Ункомпагре в западном Колорадо. [ 18 ] [ 19 ] и был передан на хранение в Университет Бригама Янга под номером образца BYU 9740. [ 20 ] [ 18 ] Скелет был полностью готов только спустя годы. [ 18 ] и был описан в 1988 году как новый род и вид камаразавридов , Cathetosaurus lewisi . [ 19 ] Первоначальное описание C. lewisi было кратким, но позже, в 1996 году, Джон Макинтош и его коллеги дали скелету полную остеологию и отнесли его к виду камаразавра . В своей статье они определили, что C. supremus, C. grandis, C. lentus и C. lewisi действительны. [ 18 ] В 2013 году Октавио Матеус и Эмануэль Чопп утверждали, что C. lewisi на самом деле представлял собой отдельный род, основанный на образце, найденном в карьере Хоу в 1992 году. [ 21 ] что они относились к виду. [ 22 ] Дальнейшие исследования Чоппа пришли к выводу, что экземпляр из карьера Хоу, скорее всего, все-таки представлял камаразавра . [ 23 ] По состоянию на 2019 год большинство исследователей считали C. lewisi разновидностью камаразавра . [ 24 ]
В 1992 году Джеффри Паркер и его коллеги собрали еще один солидный и сочлененный скелет камаразавра AMNH возле карьера Bone Cabin Quarry в Комо-Блафф. [ 25 ] Этот скелет был отнесен к Camarasaurus grandis. [ 26 ] и является одним из наиболее полных экземпляров, отнесенных к этому виду. Сейчас он находится в Музее естественной истории Гумма в Токио под номером экземпляра GMNH-PV 101. [ 27 ] [ 7 ] В 1992 году еще один камаразавра скелет был обнаружен дальше на север, в карьере Хоу , штат Вайоминг, командой, работающей в Sauriermuseum Aathal в Швейцарии. Скелет является одним из самых известных, почти каждый его элемент имеет шарнирные соединения и отпечатки кожи черепа и задних конечностей. [ 28 ] [ 21 ] Образец SMA 002 еще не прошел полную идентификацию, но предполагается, что это образец C. lewisi . [ 21 ] В 1996 году несколько фрагментарных останков камаразавра. было описано на западе Южной Дакоты [ 29 ] и Нью-Мексико , [ 30 ] расширяющий северо-восточный и южный ареал рода за счет останков Нью-Мексико из формации Саммервилл . [ 30 ] Самый северный экземпляр камаразавра был обнаружен в 2005 году в районе Снежных гор в центральной Монтане и состоит из почти полного черепа и нескольких посткраниальных элементов. [ 15 ]
Ископаемая запись
[ редактировать ]
камаразавра очень распространены. Окаменелости [ 31 ] Известно более 500 экземпляров, в том числе множество изолированных костей и около 50 частичных скелетов. [ 32 ] Он встречается на обширной территории на западе Соединенных Штатов, от севера до Монтаны до юга до Нью-Мексико, в скалах формации Моррисон . [ 33 ] Благодаря такому обилию камаразавр является очень известным зауроподом. Молодой экземпляр камаразавра , CM 11338, представляет собой наиболее полный скелет зауропода, когда-либо обнаруженный. Известны многочисленные черепа. [ 34 ] Хотя у зауроподов редко встречаются целые шеи, у пяти экземпляров камаразавра сохранились все или почти все шейные позвонки. [ 35 ] Большинство опознаваемых экземпляров Camarasaurus принадлежат к одному из двух видов: C. grandis и C. lentus ; C. lewisi и C. supremus встречаются реже. [ 36 ]
Описание
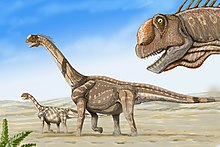
[ редактировать ]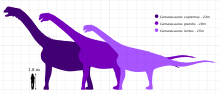
Камаразавр является одним из наиболее распространенных и часто хорошо сохранившихся динозавров-зауроподов, обнаруженных и хорошо описанных в многочисленных публикациях. [ 21 ] [ 13 ] [ 2 ] Подобно другим макронарианцам, он имел типичные большие ноздри, длинные передние конечности и короткий хвост по сравнению с современными диплодоцидами. [ 2 ] [ 37 ] Камаразавр был зауроподом среднего размера по сравнению с современными видами той же формации, но в титоне достиг крупных размеров вместе с C. supremus . [ 7 ] [ 2 ] Максимальный размер наиболее распространенного вида C. lentus составлял около 15 м (49 футов) в длину. Самый крупный вид, C. supremus , достигал максимальной длины 18–23 м (75 футов) и максимального расчетного веса 47 метрических тонн (51,8 тонны ). [ 38 ] В 2016 году Грегори С. Пол оценил его вес в 23 метрических тонны (25,4 тонны ). [ 39 ] тогда как в 2020 году Джон Фостер оценил его вес в 42,3 метрических тонны (46,6 тонны ). [ 31 ]
Изогнутый череп камаразавра множество был удивительно квадратным, а тупая морда имела окон . Прочный череп камаразавра сохранился гораздо лучше, чем у многих других зауроподов, в отличие от изящных черепов диплодоцидов , которые также встречаются в формации Моррисон. [ 37 ] длиной 19 см (7,5 дюйма) Зубы имели форму долота (лопаточки) и располагались равномерно вдоль челюсти. Прочность зубов указывает на то, что камаразавр, вероятно, питался более грубым растительным материалом, чем тонкозубые диплодоциды . [ нужна ссылка ]
Образец камаразавра под названием SMA 0002 (который также был отнесен к катетозавру ) из карьера Хоу-Стивенс в Вайоминге, называемый «ET», демонстрирует признаки мягких тканей. [ 21 ] Вдоль линии челюсти были обнаружены окостеневшие остатки того, что, по-видимому, было деснами животного, что указывает на то, что у него были глубоко посаженные зубы, покрытые деснами, и выступали только кончики коронок. После смерти зубы были выдвинуты дальше из лунок, поскольку десны втягивались, высыхали и сжимались из-за разрушения. Исследования экземпляра также показывают, что зубы были покрыты жесткими внешними чешуйками и, возможно, какой-то разновидностью клюва, хотя наверняка это неизвестно. [ 21 ]
Шея камаразавра была умеренной длины по стандартам зауроподов. Он состоял из 12 позвонков. [ 40 ] Большинство шейных нервных отростков были раздвоенными, и по мере роста животного раздвоенные нервные отростки развивались у большего количества позвонков. [ 41 ] Как и у других зауроподов, позвонки шеи и туловища содержали камеры, которые при жизни были заполнены воздушными мешками, связанными с дыхательной системой . Воздушные мешки могут занимать более половины пространства внутри позвонков, что делает их такими же пневматическими, как кости птиц. [ 42 ] Именно эти камеры дали камаразавру название «камерная ящерица».
Хвост камаразавра состоял из 53 позвонков. [ 40 ] [ 43 ]
Классификация и виды
[ редактировать ]Camarasaurus — типовой род семейства Camarasauridae среднего размера , членами которого являются зауроподы -макронарии , которые в основном датируются поздней юрой . У камаразаврид были более короткие передние конечности, чем задние, большие лопаточно-коракоидные кости и более длинные хвосты, чем шеи. [ 44 ] [ 2 ] Когда Эдвард Коуп описал камаразавра в 1877 году, он полагал, что это динозавр, тесно связанный с цетиозавром , ботриоспондилом , орнитопсисом , анхизавром ( мегадактилусом ) и пневматостеусом . [ 1 ] но не назвал группу этих таксонов до описания Amphicoelias, когда он выделил Camarasauridae. [ 5 ] Камаразавр — единственный таксон, который бесспорно считается действительным родом камаразавридов. Он содержит четыре вида: C. grandis , C. lentus , C. lewisi и C. supremus . C. lewisi может представлять собой отдельный род Cathetosaurus . Lourinhasaurus , типовой вид которого ранее был отнесен к Camarasaurus , в большинстве исследований рассматривается как камаразаврид, хотя он также считается базальным эузавроподом. [ 45 ] [ 46 ]
Упрощенная кладограмма базальной макронарии по Тану и др. (2020) показана ниже:
Камаразавр считается базальным макронарием, более тесно связанным с общим предком всех макронарий, чем с более производными формами, такими как брахиозавр .
Разновидность
[ редактировать ]Camarasaurus Большинство исследователей считают, что содержит четыре действительных вида: C. grandis , C. lentus , C. lewisi и C. supremus . [ 24 ] C. supremus , вид, названный Коупом в 1877 году, является типовым видом . C. grandis был назван в 1877 году, а C. lentus - в 1889 году. Четвертый вид, C. lewisi , имеет неопределенное родство. Первоначально он был описан как отдельный род Cathetosaurus в 1988 году, но реклассифицирован как вид Camarasaurus в 1996 году. [ 47 ] [ 48 ] Некоторые исследователи предположили, что Cathetosaurus следует восстановить в качестве отдельного рода. [ 22 ] [ 49 ] [ 24 ] тогда как другие предположили, что C. lewisi может быть синонимом другого вида Camarasaurus . [ 7 ] [ 50 ]

C. supremus , как следует из названия, является крупнейшим известным видом камаразавра. [ 2 ] и один из самых массивных зауроподов, известных из позднеюрской формации Моррисон . За исключением огромных размеров, он был почти неотличим от C. lentus . C. supremus не был типичен для рода в целом, известен лишь из самых поздних, самых верхних частей свиты и крайне редок. [ 7 ] И C. grandis, и C. lentus, и C. lewisi были меньше по размеру и встречались на более ранних стадиях Моррисона.

Стратиграфические данные свидетельствуют о том, что хронологическая последовательность соответствует физическим различиям между тремя видами и описывает эволюционное развитие внутри формации Моррисон. C. grandis — древнейший вид, обитающий в самых нижних слоях горных пород Моррисона. C. lewisi лишь недолго сосуществовал с C. grandis в самых нижних слоях верхнего Моррисона, пока не вымер. [ 7 ] но возможно, это связано с отсутствием экземпляров C. lewisi . [ 7 ] C. lentus появился позже и сосуществовал с C. grandis в течение нескольких миллионов лет, возможно, из-за разных экологических ниш, о чем свидетельствуют различия в анатомии позвоночника этих двух видов. На более позднем этапе C. grandis исчез из наскальной летописи, остался только C. lentus . [ 7 ] Затем C. lentus исчез и ; в то же время C. supremus появился в самых верхних слоях. Эта непосредственная последовательность видов, а также очень близкое сходство между ними позволяют предположить, что C. supremus , возможно, произошел непосредственно от C. lentus , представляя собой более крупную, выжившую позже популяцию животных. [ 51 ]
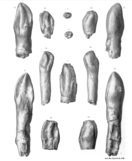
Синонимы и сомнительные виды
[ редактировать ]- Amphicoelias latus был назван Эдвардом Коупом в 1877 году. [ 52 ] основан на правом бедре и четырех хвостовых позвонках, найденных в Гарден-Парке, и является синонимом C. supremus или C. grandis . [ 6 ] [ 7 ]
- Caulodon diversidens был также назван Коупом в 1877 году по теперь сомнительным зубам, которые можно отнести только к макронарианцам или как синонимы Camarasaurus supremus . [ 20 ] [ 7 ] [ 2 ]
- Caulodon leptoganus был назван Коупом в 1878 году по двум частичным зубам и также считается не поддающимся классификации за пределами Macronaria или синонимом Camarasaurus supremus . [ 20 ] [ 7 ] [ 2 ]
- Morosaurus impar был назван Маршем в 1878 году типовым видом Morosaurus , а материал состоял только из крестца и, возможно, дополнительного посткраниального материала, найденного в Комо-Блафф . Сейчас он считается синонимом C. grandis. [ 7 ]
- Morosaurusrobustus был назван на основе подвздошной кости, собранной Маршем в 1878 году в Комо-Блафф. Сейчас он считается синонимом C. grandis . [ 7 ] [ 20 ]
- Camarasaurus leptodirus был еще одним зауроподом из Гарден-Парка Коупа и был назван в 1879 году по трем частичным шейным позвонкам. Было высказано предположение, что это синоним C. supremus . [ 7 ] [ 20 ]
- Diplodocus lacustris был назван Отниелом Маршем в 1884 году на основе нескольких зубов, предчелюстной кости и верхней челюсти из Моррисона , штат Колорадо, которые были собраны Артуром Лейксом и Бенджамином Маджем в 1877 году. [ 53 ] Хотя зубы и зубная кость D. lacustris относятся к Flagellicaudatan , материал черепа, вероятно, принадлежит камаразаврам . [ 53 ]
- Pleurocoelus montanus был также назван Маршем в 1896 году как новый вид Pleurocoelus , материал, состоящий из нескольких центров позвонков и различных посткраний юной особи из Комо-Блафф. Обычно его считают синонимом C. grandis. [ 7 ] [ 20 ]
- Uintasaurus douglassi был назван в 1919 году У. Дж. Холландом в честь пяти передних шейных позвонков из Национального памятника динозавров. [ 16 ] Позже этот вид был расценен как синоним Camarasaurus lentus . [ 7 ]
- Camarasaurus annae был назван Тейджем Эллингером на основании переднего спинного позвонка в 1950 году. Этот вид обычно считается синонимом C.lentus. [ 7 ]
Переназначенные виды
[ редактировать ]- Morosaurus agilis был назван Маршем в 1889 году на основе части черепа и трех позвонков из Гарден-Парка, штат Колорадо. [ 54 ] Вид оставался в таксономической неопределенности до тех пор, пока в 2020 году он не был помещен в новый род Smitanosaurus и реклассифицирован как дикреозаврид . [ 8 ]
- Camarasaurus becklesiii был описан как Pelorosaurus becklesii в 1842 году Гидеоном Мантеллом на основе части передней конечности из Сассекса, Великобритания . Он был помещен в Морозавра Маршем в 1889 году. [ 55 ] и Камаразавр фон Хюне в [ 56 ] пока в 2015 году его не отнесли к отдельному роду Haestasaurus . [ 57 ]
- Morosaurus Marchei был назван Соважем в 1898 году на основании неполного дистального хвостового позвонка и зуба из слоев верхней юры формации Алькобака в Португалии. [ 58 ] Лаппарент и Збышевский упомянули голотип. [ 59 ] позвонка к Megalosaurus insignis , а Madsen et al ., 1995 отнесли его к Megalosauria . [ 60 ] Указанный зуб был идентифицирован как принадлежащий Turiasauria в 2017 году. [ 61 ]
- Camarasaurus alenquerensis был назван как вид апатозавра в 1957 году Альбертом-Феликсом де Лаппарентом и Жоржем Збивески на частичном посткраниальном скелете из Лориньи, Португалия . [ 62 ] Он был помещен в Камаразавр Джоном Макинтошом в 1990 году. [ 63 ] но в 1998 году ему был предоставлен новый род — Lourinhasaurus .
Палеобиология
[ редактировать ]
Кормление
[ редактировать ]Ранее ученые предположили, что камаразавр и другие зауроподы, возможно, глотали гастролиты (камни), чтобы перемалывать пищу в желудке, срыгивая или выдавая ее, когда она становилась слишком гладкой. Однако более поздний анализ данных о камнях в желудке показывает, что это не так. Сильные и крепкие зубы камаразавра были более развиты, чем у большинства зауроподов, и сменялись в среднем каждые 62 дня (М. Д'Эмик и др.), Это указывает на то, что камаразавр мог в некоторой степени пережевывать пищу во рту, прежде чем проглотить. [ 64 ] Другие данные показывают, что Camarasaurus spp. предпочитали растительность, отличную от других зауроподов, что позволяло им жить в одной и той же среде, не конкурируя. [ 65 ]
Рост
[ редактировать ]длинных костей Гистология позволяет исследователям оценить возраст, которого достиг конкретный человек. Исследование Griebeler et al. (2013) изучили гистологические данные длинных костей и пришли к выводу, что Camarasaurus sp. CM 36664 весил 14 247 килограммов (15,7 коротких тонн), достиг половой зрелости в 20 лет и умер в 26 лет. [ 66 ]
Метаболизм
[ редактировать ]Игл и др. выполнил термометрию слипшихся изотопов на эмали, покрывающей зубы различных юрских зауроподов, включая камаразавра . Были получены температуры 32,4–36,9 ° C (90,3–98,4 ° F), что сопоставимо с температурой современных млекопитающих. [ 67 ] Камаразавр быстро рос в размерах, чтобы ограничить время, в течение которого он будет уязвим для хищников. Это означало бы, что в юном возрасте у него был относительно высокий уровень метаболизма. [ 68 ]
Палеопатология
[ редактировать ]На тазе камаразавра , найденном в Национальном памятнике динозавров в штате Юта, видны выбоины, приписываемые аллозавру. [ 69 ] а на подвздошной кости голотипа C. lewisi имеются крупные следы укусов теропод. [ 20 ]
В 1992 году частичный скелет C. grandis был обнаружен в карьере стегозавра Брайана Смолла в формации Моррисон недалеко от Кэнон-Сити, штат Колорадо . [ 70 ] Этот экземпляр сохранил частичную правую плечевую кость, каталогизированную как DMNH 2908, и связанные с ней позвонки сзади и хвоста. [ 70 ] В 2001 году Лори Маквинни, Кеннет Карпентер и Брюс Ротшильд опубликовали описание патологии, наблюдаемой на плечевой кости. [ 71 ] Они отметили юкстакортикальное поражение шириной 25 на 18 см, состоящее из кости, напоминающее переплетенные волокна. [ 72 ] Хотя плетеная кость образуется в добавочной зубной кости, в других местах это признак травмы или болезни. [ 72 ] Наблюдались «волнистые волокнистые пучки» плетеной кости, ориентированные в направлении m. брахиалис . [ 72 ] очага поражения Сращение и отсутствие пористости на ближнем и дальнем концах указывают на то, что периостит был неактивен или зажил. [ 72 ] Маквинни и другие исследователи утверждали, что эта травма стала бы постоянным источником страданий для животного. [ 73 ] Это оказало бы давление на мышцы. [ 71 ] Это давление могло бы сжать кровеносные сосуды и нервы мышц, уменьшив диапазон движений мышц- сгибателей и разгибателей конечности . [ 71 ] Этот эффект помешал бы M. brachialis , m. brachoradialis и в меньшей степени m. biceps brachii к месту поражения на плечевой кости. [ 71 ] Исследователи пришли к выводу, что воспаление мышц и надкостницы также могло вызвать дополнительные осложнения в нижней части передней конечности. [ 74 ] Поражение также могло вызвать длительный фасциит и миозис . [ 71 ] Совокупный эффект этих патологических процессов будет иметь умеренные или серьезные последствия для способности конечности двигаться и «затруднит выполнение повседневных действий, таких как поиск пищи и спасение от хищников». [ 73 ] Чтобы определить причину патологии, Маквинни и другие исследователи провели компьютерную томографию с шагом 3 мм. [ 75 ] КТ показала, что образование имело постоянную радиоплотность и было отделено от коры кости рентгенопрозрачной линией. [ 76 ] Никаких доказательств стрессового перелома или инфекционных процессов, таких как остеомиелит или инфекционный периостит, обнаружено не было. [ 75 ] Они также исключили остеохондрому, поскольку ось шпоры находится под углом 25° к вертикальной оси плечевой кости, тогда как остеохондрома могла образоваться под углом 90° к оси плечевой кости. [ 72 ] Другие кандидаты, идентифицированные учеными как причина образования шпоры, включали:
- Гипертрофическая остеоартропатия – хотя это исключалось наличием шпорообразного отростка [ 77 ]
- Остеоид-остеома – но это не объясняет шпору или остеобластическую реакцию. [ 78 ]
- Расколотая голень или синдром стресса большеберцовой кости - возможное происхождение, поскольку многие симптомы могут быть общими, но расколотая голень не может объяснить появление шпоры. [ 79 ]
- Травматический оссифицирующий миозит (ограниченный) – возможный, но маловероятный источник. [ 80 ]
- Отрывная травма . Маквинни и другие исследователи считали отрывную травму, вызванную «повторяющимся перенапряжением мышц», наиболее вероятным источником поражения плечевой кости. [ 71 ] Исследователи полагали, что поражение возникло в результате отрыва m. brachialis, вызывая образование «наклонной эллиптической массы». [ 73 ] Костная шпора возникла в результате остеобластической реакции после разрыва основания m. brachioradialis, вызванное движением сгибателей. [ 73 ]
Палеоэкология
[ редактировать ]среда обитания
[ редактировать ]
Формация Моррисон, расположенная вдоль восточного склона Скалистых гор , является домом для богатого окаменелостями участка позднеюрских пород. Здесь можно встретить большое количество видов динозавров, в том числе родственников камаразавров, таких как диплодок , апатозавр и брахиозавр , но камаразавры являются наиболее многочисленными из динозавров в формации. [ 81 ] Окаменелости камаразавров были обнаружены почти во всех крупных населенных пунктах и имеют одно из самых известных распространенных мест среди динозавров Моррисона: окаменелости были найдены в местах от Нью-Мексико до Монтаны и от Юты до Оклахомы. [ 7 ] Согласно радиометрическому датированию , осадочные слои Моррисона колеблются от 156,3 миллионов лет назад (млн лет назад) у основания до 146,8 млн лет назад наверху, что помещает их в поздний оксфордский , кимериджский и ранний титонский этапы позднеюрского периода. [ 82 ] [ 83 ] Его окружающая среда интерпретируется как полузасушливая с четко выраженными влажными и засушливыми сезонами .
динозавров и Окаменелости их следы встречаются, в частности, в бассейне Моррисон, который простирается от Нью-Мексико до Альберты и Саскачевана и образовался, когда предшественники Переднего хребта Скалистых гор начали продвигаться на запад. обращенных на восток, Эродированный материал из их водосборных бассейнов, переносился ручьями и реками и откладывался в заболоченных низменностях, озерах, речных руслах и поймах рек . [ 84 ] По возрасту эта формация аналогична формации Лоринья в Португалии и формации Каньядон Калькарео в Аргентине. Окаменелости камаразаврид были обнаружены в двух формациях. [ 62 ] [ 85 ] В 1877 году он стал центром Войн костей , соперничества за сбор окаменелостей между ранними палеонтологами Отниэлем Чарльзом Маршем и Эдвардом Дринкером Коупом , причем сам Камаразавр был обнаружен и назван последним палеонтологом во время конфликта.
Палеофауна
[ редактировать ]В формации Моррисон зафиксирована среда и время, в котором доминировали гигантские динозавры-зауроподы, такие как мараапунизавр , амфикоэлия , барозавр , диплодок , апатозавр , бронтозавр и брахиозавр . Динозавры, живущие рядом с камаразавром, включали травоядных птицетазовых камптозавров , горгульеозавров , дриозавров , стегозавров и нанозавров . Хищниками в этой палеосреде были тероподы Saurophaganax , Torvosaurus , Ceratosaurus , Marshosaurus , Stokesosaurus , Ornitholestes , [ 86 ] и аллозавр , который составлял до 75% экземпляров теропод и находился на верхнем трофическом уровне Моррисона пищевой сети . [ 87 ] [ 88 ] Камаразавр обычно встречается в тех же местах, что и аллозавр , апатозавр , стегозавр и диплодок . [ 89 ]
Другие организмы в этом регионе включали двустворчатых моллюсков , улиток , лучепёрых рыб , лягушек , саламандр , черепах , сфенодонтов , ящериц , наземных и водных крокодиломорфов , а также несколько видов птерозавров, таких как Harpactognathus и Mesadactylus . Ранними млекопитающими были докодонты (такие как докодон ), мультитуберкуляты , симметродонты и триконодонты . Флора этого периода представлена окаменелостями зеленых водорослей , грибов , мхов , хвощей , саговников , гинкго и некоторых семейств хвойных . Растительность варьировалась от прибрежных лесов из древовидных папоротников и папоротников ( галерейных лесов ) до папоротниковых саванн с редкими деревьями, такими как подобное араукарии хвойное дерево Brachyphyllum, . [ 90 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Коуп, Э.Д., 1877a, О гигантском ящере из дакотской эпохи Колорадо: Палеонтологический бюллетень, т. 25, с. 5-10.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п Осборн, Генри Фэрфилд; Мук, Чарльз Крейг (1921). «Камаразавр, амфикоэлии и другие зауроподы Коупа: Мемуары Американского музея естественной истории, т. 3, стр. 247-387» . hdl : 2246/5724 . Архивировано из оригинала 15 января 2016 года.
- ^ Перейти обратно: а б Коуп, Э.Д., 1877c, Об останках рептилий из отложений Дакоты в Колорадо: Палеонтологический бюллетень, т. 26, с. 193-196.
- ^ Марш, О.К. (1877). « Уведомление о новых динозавровых рептилиях из юрской формации ». Американский научный журнал. 3-я серия. 14 (84): 514–516. Бибкод: 1877AmJS...14..514M. doi:10.2475/ajs.s3-14.84.514. S2CID 130488291.
- ^ Перейти обратно: а б Коуп, ED, 1877b, Об амфикоэлиях, новом роде ящеров из эпохи Дакоты в Колорадо: Палеонтологический бюллетень, т. 1878; 27, с. 2-5
- ^ Перейти обратно: а б Карпентер, К. (1998). Биостратиграфия позвоночных формации Моррисон возле Каньон-Сити, штат Колорадо. Современная геология , 23 , 407–426.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В Икеджири, Такехито (2005). Распространение и биохронология камаразавров (Dinosauria, Sauropoda) из юрской формации Моррисон региона Скалистых гор . Геологическое общество Нью-Мексико, январь 2005 г.
- ^ Перейти обратно: а б с Уитлок, Джон А.; Мантилья, Джеффри А. УИЛСОН (10 декабря 2020 г.). «Позднеюрский динозавр-зауропод 'Morosaurus' agilis Marsh, 1889 повторно исследован и переосмыслен как дикреозаврид» . Журнал палеонтологии позвоночных . 40 (6): e1780600. Бибкод : 2020JVPal..40E0600W . дои : 10.1080/02724634.2020.1780600 . ISSN 0272-4634 . S2CID 234424022 .
- ^ Осборн, HF (1901). Передние и задние конечности зауроподов из карьера Bone Cabin (Том 3).
- ^ Перейти обратно: а б с Риггс, Элмер (1901). «Передняя нога и грудной пояс морозавра». Полевой музей естественной истории. Публикация 63, Геологическая серия . 1 (10): 275–281.
- ^ «AMNH 467 — запись Camarasaurus sp.» .
- ↑ Миллер, Б. (30 октября 2014 г.). « Хулиган для Камаразавра ». Динозавры.
- ^ Перейти обратно: а б с Гилмор, К. (1925). «Почти полный шарнирный скелет камаразавра, ящеротазового динозавра из Национального памятника динозавров» . Мемуары музея Карнеги . 10 : 347–384. дои : 10.5962/стр.217807 . S2CID 128077427 .
- ^ Перейти обратно: а б Миллер, Бен (29 декабря 2016 г.). «Диплодок и камаразавр (серия NMNH)» . Архивировано из оригинала 13 августа 2020 года.
- ^ Перейти обратно: а б Вудрафф и Фостер 2017 .
- ^ Перейти обратно: а б с Голландия, WJ (1924). «Описание типа Uintasaurus douglassi HOLLAND». Анналы музея Карнеги. 15 (2–3): 119–138.
- ^ Ведель, Мэтт (2018). «OMNH 1811, дорсальный камаразавр из Чёрной Мезы» . Архивировано из оригинала 3 февраля 2018 года.
- ^ Перейти обратно: а б с д Макинтош Дж.С., Миллер В.Е., Стадтман К.Л. и Джилетт Д.Д., 1996b, Остеология Camarasaurus lewisi (Дженсен, 1988) : Геологические исследования Университета Бригама Янга, т. 41, стр. 73-115.
- ^ Перейти обратно: а б Дженсен, Дж. А. (1988). «Четвертый новый динозавр-зауропод из верхней юры плато Колорадо и прямохождение зауроподов». Натуралист Великого Бассейна . 48 (2): 121–145.
- ^ Перейти обратно: а б с д и ж г Форд, Трейси. «Палеофил Камаразавр» . Архивировано из оригинала 26 января 2020 года.
- ^ Перейти обратно: а б с д и ж Виерсма К. и Сандер П.М. (2017). Зубной ряд хорошо сохранившегося экземпляра Camarasaurus sp.: влияние на функцию, замену зубов, реконструкцию мягких частей и потребление пищи. ПалЗ , 91 (1), 145-161.
- ^ Перейти обратно: а б Матеус О. и Чопп Э. (2013). Катетозавр как действительный род зауроподов и сравнение с камаразавром . Журнал палеонтологии позвоночных, программа и тезисы, 2013. 173.
- ^ Чопп, Эмануэль; Крылья, Оливер; Фрауэнфельдер, Томас; Ротшильд, Брюс (2016). «Патологические фаланги у камаразавридных динозавров-зауроподов и их влияние на поведение». Acta Palaeontologica Polonica . 61 (1): 125–134. дои : 10.4202/app.00119.2014 . HDL : 2318/1526045 . ISSN 0567-7920 . S2CID 53542173 .
- ^ Перейти обратно: а б с Чопп, Эмануэль; Мейдмент, Сюзанна ЧР; Ламанна, Мэтью С.; Норелл, Марк А. (4 ноября 2019 г.). «Переоценка исторической коллекции динозавров зауропод из формации Северный Моррисон в Вайоминге с последствиями для биогеографии зауроподов» . Бюллетень Американского музея естественной истории . 2019 (437): 1. дои : 10.1206/0003-0090.437.1.1 . ISSN 0003-0090 . S2CID 207890316 .
- ^ Макинтош и др. 1996 , стр. 1–5.
- ^ Макинтош и др. 1996 , с. 30
- ^ Макинтош и др. 1996 , с. 6.
- ^ Чопп, Э.Д., Оливер, В., Томас, Ф., и Винанд, Б. (2015). Сочлененные костные наборы рук и ног камаразавра (Sauropoda, Dinosauria).
- ^ Фостер, младший (1996). Динозавры-зауроподы формации Моррисон (верхняя юра), Блэк-Хиллз, Южная Дакота и Вайоминг. Геология Скалистых гор , 31 (1), 1-25.
- ^ Перейти обратно: а б ЛУКАС, С.Г., и ХЕККЕРТ, AB (1993). Юрские динозавры в Нью-Мексико. Динозавры Нью-Мексико: Бюллетень 17 , 17 , 43.
- ^ Перейти обратно: а б Фостер 2020 .
- ^ Вудрафф и Фостер, 2017 , стр. 52–53.
- ^ Вудрафф и Фостер 2017 , с. 54.
- ^ Мэдсен, Макинтош и Берман 1995 , стр. 2.
- ^ Тейлор 2022 , с. 6.
- ^ Вудрафф и Фостер 2017 , с. 48.
- ^ Перейти обратно: а б Макинтош Дж.С., Берман Д.С. 1975. Описание неба и нижней челюсти динозавра-зауропода Diplodocus (Reptilia: Saurischia) с замечаниями о характере черепа апатозавра. Журнал Палеонтологии 49:187–199.
- ^ Бентон, Майкл Дж. (2012). Доисторическая жизнь . Эдинбург, Шотландия: Дорлинг Киндерсли. стр. 270–271. ISBN 978-0-7566-9910-9 .
- ^ Пол, GS (2016). Принстонский полевой справочник по динозаврам (2-е изд.). Принстон и Оксфорд: Издательство Принстонского университета. ISBN 978-0-691-16766-4 .
- ^ Перейти обратно: а б Гилмор 1925 , с. 367.
- ^ Ведель и Тейлор, 2013 , стр. 11–12.
- ^ Тейлор и Ведель, 2013 , стр. 14–15.
- ^ Макинтош и др. 1996 , стр. 15.
- ^ Макинтош и др. 1996 год .
- ^ МОЧО, Педро; Ройо-Торрес, Рафаэль; Ортега, Франциско (19 февраля 2014 г.). «Филогенетическая переоценка Lourinhasaurus alenquerensis , базальной макронарии (Sauropoda) из поздней юры Португалии» . Зоологический журнал Линнеевского общества . 170 (4): 875–916. дои : 10.1111/zoj.12113 . ISSN 0024-4082 .
- ^ Апчерч, Пол; Барретт, Пол М.; Додсон, Питер (2004). «Зауропода». В Вейшампеле, Дэвид Б.; Додсон, Питер; Осмольска, Гальшка (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 259–322 . ISBN 0-520-24209-2 .
- ^ Дженсен, Джеймс А. (30 апреля 1988 г.). «Четвертый новый динозавр-зауропод из верхней юры плато Колорадо и прямохождение зауроподов» . Натуралист Великого Бассейна . 48 (2): 121–145. ISSN 0017-3614 . JSTOR 41712420 .
- ^ Макинтош, Джон С; Миллер, Уэйд Э; Штадтман, Кеннет Л.; Джилетт, Дэвид Д. (1996). «Остеология Camarasaurus lewisi (Дженсен, 1988)». Геологические исследования в Университете Бригама Янга . 41 : 73–115. ISSN 0068-1016 .
- ^ Чопп, Э.; Матеус, О.; Косма, Р.; Сандер, премьер-министр; Джогер, У.; Крылья, ОВМ (2014). «Кладистический анализ камаразавров (Dinosauria, Sauropoda) на уровне экземпляров и пересмотр таксономии камаразавридов». 74-е ежегодное собрание Общества палеонтологии позвоночных, журнал палеонтологии позвоночных, программа и тезисы .
- ^ Woodruff & Foster 2017 , стр. 50–51.
- ^ « Camarasaurus grandis », Фостер (2007). Страница 204.
- ^ Коуп, ЭД (1877c). «Об амфикоэлиях, роде ящеров из эпохи Дакоты в Колорадо». Труды Американского философского общества (17): 242–246.
- ^ Перейти обратно: а б Чопп, Эмануэль; Матеус, Октавиус; Бенсон, Роджер Би Джей (7 апреля 2015 г.). «Филогенетический анализ на уровне конкретного уровня и таксономический обзор Diplodocidae (Dinosauria, Sauropoda)» . ПерДж . 3 : е857. дои : 10.7717/peerj.857 . ISSN 2167-8359 . ПМЦ 4393826 . ПМИД 25870766 .
- ^ ОК Марш. 1889. Уведомление о новой американской динозаврии. Американский журнал науки и искусства, серия 3 38: 331-336.
- ^ Марш, О.К. (1889). Приложение; Сравнение основных форм динозавров Европы и Америки . Американский научный журнал , 3 (220), 323–330.
- ^ Хуэне, Ф.В. (1932). Ископаемые рептилии отряда Saurischia, их развитие и история. Монографии по геологии и палеонтологии (сер. 1).
- ^ Апчерч, Пол; Мэннион, Филип Д.; Тейлор, Майкл П. (3 июня 2015 г.). «Анатомия и филогенетические взаимоотношения Pelorosaurus becklesii (Neosauropoda, Macronaria) из раннего мела Англии» . ПЛОС ОДИН . 10 (6): e0125819. Бибкод : 2015PLoSO..1025819U . дои : 10.1371/journal.pone.0125819 . ISSN 1932-6203 . ПМЦ 4454574 . ПМИД 26039587 .
- ^ Уайлд, HE (1898). Ископаемые позвоночные Португалии: вклад в изучение рыб и рептилий юрского и мелового периода . Королевская академия наук.
- ^ Ольшевский, Георгий (24 октября 1991 г.). «Пересмотр параинфракласса Archosauria Cope, 1869, исключая продвинутые Crocodylia». Мезозойские извилины . 2 : 1–196.
- ^ Мэдсен, Макинтош и Берман, 1995. Череп и комплекс оси атланта верхнеюрского зауропода Camarasaurus Cope (Reptilia: Saurischia). Бюллетень Музея естественной истории Карнеги. 31, 1-115.
- ^ Мочо, П., Ройо-Торрес, Р., Эскасо, Ф., Малафайя, Э., де Мигель Чавес, К., Нарваес, И., ... и Ортега, Ф. (2017). Находка верхнеюрских зауроподов в Лузитанском бассейне (Португалия): географическое и литостратиграфическое распространение. Электронная палеонтология , 20 (2).
- ^ Перейти обратно: а б А. Ф. де Лаппарент и Г. Збышевский, 1951, «Открытие богатой фауны динозавров-рептилий в верхней юре Португалии», Comptes Rendus de l'Académie des Sciences à Paris 233 : 1125-1127.
- ^ Макинтош, Дж. С. 1990. Зауроподы. В Вейшампеле, Д.Б.; Додсон, П.; и Осмольска, Х. (ред.): Динозаврия. Беркли (Калифорнийский университет Press): 345-401.
- ^ Крылья и Сандер (2006).
- ^ «Высокая скорость замены зубов у крупнейших динозавров способствовала их эволюционному успеху» .
- ^ Грибелер, Э.М.; Кляйн, Н; Сандер, премьер-министр (2013). «Старение, созревание и рост динозавров-зауроподоморфов, определенные на основе кривых роста с использованием гистологических данных длинных костей: оценка методологических ограничений и решений» . ПЛОС ОДИН . 8 (6): e67012. Бибкод : 2013PLoSO...867012G . дои : 10.1371/journal.pone.0067012 . ПМЦ 3686781 . ПМИД 23840575 .
- ^ Игл, РА; Тюткен, Т.; Мартин, Т.С.; Трипати, АК; Фрике, ХК; Коннели, М.; Чифелли, РЛ; Эйлер, Дж. М. (22 июля 2011 г.). «Температура тела динозавров, определенная на основе изотопного порядка (13C-18O) в ископаемых биоминералах». Наука . 333 (6041): 443–445. Бибкод : 2011Sci...333..443E . дои : 10.1126/science.1206196 . ПМИД 21700837 . S2CID 206534244 .
- ^ Васков, Катя; Сандер, П. Мартин (7 июня 2014 г.). «Рекорды роста и гистологические вариации спинных ребер Camarasaurus sp. (Sauropoda)» . Журнал палеонтологии позвоночных . 34 (4): 852–869. Бибкод : 2014JVPal..34..852W . дои : 10.1080/02724634.2014.840645 . ISSN 0272-4634 . S2CID 86736713 .
- ^ «Камаразавр», Додсон и др. Страница 56.
- ^ Перейти обратно: а б Маквинни, Карпентер и Ротшильд 2001 , с. 365.
- ^ Перейти обратно: а б с д и ж Маквинни, Карпентер и Ротшильд 2001 , с. 364.
- ^ Перейти обратно: а б с д и Маквинни, Карпентер и Ротшильд 2001 , с. 367.
- ^ Перейти обратно: а б с д Маквинни, Карпентер и Ротшильд 2001 , с. 376.
- ^ Маквинни, Карпентер и Ротшильд 2001 , стр. 369.
- ^ Перейти обратно: а б Маквинни, Карпентер и Ротшильд 2001 , с. 370.
- ^ Маквинни, Карпентер и Ротшильд 2001 , стр. 370–371.
- ^ Маквинни, Карпентер и Ротшильд 2001 , стр. 372–373.
- ^ Маквинни, Карпентер и Ротшильд 2001 , стр. 373–374.
- ^ Маквинни, Карпентер и Ротшильд 2001 , стр. 374.
- ^ Маквинни, Карпентер и Ротшильд 2001 , стр. 374–375.
- ^ « Высший камаразавр », Фостер (2007). Стр. 201. «Изобилие и разнообразие», там же. Страница 248
- ^ Трухильо, КЦ; Чемберлен, КР; Стрикленд, А. (2006). «Оксфордский возраст U/Pb на основе анализа SHRIMP для верхнеюрской формации Моррисон на юго-востоке Вайоминга с последствиями для биостратиграфических корреляций». Геологическое общество Америки Рефераты с программами . 38 (6): 7.
- ^ Билби, ЮАР (1998). «Карьер динозавров Кливленд-Ллойд - возраст, стратиграфия и среда осадконакопления». В Карпентере, К.; Чуре, Д.; Киркланд, Дж.И. (ред.). Формация Моррисона: междисциплинарное исследование . Современная геология. Том. 22. Группа Тейлора и Фрэнсиса. стр. 87–120. ISSN 0026-7775 .
- ^ Рассел, Дейл А. (1989). Одиссея во времени: Динозавры Северной Америки . Миноква, Висконсин: NorthWord Press. стр. 64–70. ISBN 978-1-55971-038-1 .
- ^ КАРБАЛЛИДО, ХОЗЕ Л.; РАУХУТ, ОЛИВЕР ВМ; ПОЛ, ДИЕГО; САЛЬГАДО, ЛЕОНАРДО (24 августа 2011 г.). «Остеология и филогенетические взаимоотношения Tehuelchesaurus benitezii (Dinosauria, Sauropoda) из верхней юры Патагонии» . Зоологический журнал Линнеевского общества . 163 (2): 605–662. дои : 10.1111/j.1096-3642.2011.00723.x . hdl : 11336/71888 . ISSN 0024-4082 .
- ^ Фостер, Дж. (2007). «Приложение». Юрский Запад: динозавры формации Моррисон и их мир. Издательство Университета Индианы. стр. 327-329.
- ^ Фостер, Джон (2007). « Аллозавр ломкий ». Юрский Запад: динозавры формации Моррисон и их мир . Блумингтон, Индиана: Издательство Университета Индианы. стр. 170–176. ISBN 978-0-253-34870-8 . OCLC 77830875 .
- ^ Фостер, Джон Р. (2003). Палеоэкологический анализ фауны позвоночных формации Моррисон (верхняя юра), регион Скалистых гор, США . Бюллетень Музея естественной истории и науки Нью-Мексико, 23 . Альбукерке, Нью-Мексико: Музей естественной истории и науки Нью-Мексико. п. 29.
- ^ Додсон, Питер; Беренсмейер, АК; Баккер, Роберт Т.; Макинтош, Джон С. (1980). «Тафономия и палеоэкология отложений динозавров юрской формации Моррисон». Палеобиология . 6 (2): 208–232. дои : 10.1017/S0094837300025768 .
- ^ Карпентер, Кеннет (2006). «Самый большой из больших: критическая переоценка мегазауропода Amphicoelias fragillimus ». В Фостере, Джон Р.; Лукас, Спенсер Г. (ред.). Палеонтология и геология верхнеюрской формации Моррисон . Бюллетень Музея естественной истории и науки Нью-Мексико. Том. 36. Альбукерке, Нью-Мексико: Музей естественной истории и науки Нью-Мексико. стр. 131–138.
Цитируемые работы
[ редактировать ]- Коуп, ЭД (1878). «О позвоночных эпохи Дакоты в Колорадо». Палеонтологический вестник . 17 (28): 233–247. JSTOR 982303 .
- Коуп, ЭД (1879). «Новая юрская динозаврия». Американский натуралист . 13 : 401–404.
- Додсон, Питер; Бритт, Брукс; Карпентер, Кеннет; Форстер, Кэтрин А.; Джилетт, Дэвид Д.; Норелл, Марк А.; Ольшевский, Георгий; Пэрриш, Дж. Майкл; Вейшампель, Дэвид Б. (1 января 1993 г.). Эпоха динозавров . Публикации Интернешнл, ООО. ISBN 978-0-7853-0443-2 .
- Фостер, Дж. (2007). Юрский Запад: динозавры формации Моррисон и их мир . Издательство Университета Индианы . ISBN 978-0-253-34870-8 .
- Фостер, Джон Рассел (2020). Юрский Запад: динозавры формации Моррисон и их мир . Жизнь прошлого (2-е изд.). Блумингтон, Индиана: Издательство Университета Индианы. ISBN 978-0-253-05158-5 .
- Голландия, WJ (1919). «Раздел V. Палеонтология». Годовой отчет директора за год, закончившийся 31 марта 1919 года (Отчет). Музей Карнеги. п. 38. hdl : 2027/osu.32435027945948 .
- Марш, О.К. (1877 г.). «Уведомление о новых динозавровых рептилиях из юрской формации» . Американский научный журнал . 3-я серия. 14 (84): 514–516. Бибкод : 1877AmJS...14..514M . дои : 10.2475/ajs.s3-14.84.514 . S2CID 130488291 .
- Марш, О.К. (1878a). «Уведомление о новых рептилиях-динозаврах» . Американский научный журнал . 3-я серия. 15 (87): 241–244. Бибкод : 1878AmJS...15..241M . дои : 10.2475/ajs.s3-15.87.241 . S2CID 131371457 .
- Марш, О.К. (1878b). «Главные персонажи американских юрских динозавров, часть I». Американский научный журнал . 3-я серия. 16 (95): 411–416. дои : 10.2475/ajs.s3-16.95.411 . hdl : 2027/hvd.32044107172876 . S2CID 219245525 .
- Марш, О.К. (1889 г.). «Сравнение основных форм динозавров Европы и Америки» . Американский научный журнал . 3-я серия. 37 (220): 323–330. дои : 10.2475/ajs.s3-37.220.323 . S2CID 130946518 .
- Мэдсен, Джеймс Х.; Макинтош, Джон С.; Берман, Дэвид С. (10 октября 1995 г.). «Комплекс черепа и оси атланта верхнеюрского зауропода Camarasaurus Cope (Reptilia: Saurischia)» . Бюллетень Музея естественной истории Карнеги . 31 : 1–115. дои : 10.5962/стр.240778 . ISSN 0145-9058 . S2CID 198252192 .
- Макинтош, Джон С.; Майлз, Клиффорд А.; Кловард, Карен С.; Паркер, Джеффри Р. (24 декабря 1996 г.). «Новый почти полный скелет камаразавра». Вестник Музея естественной истории Гумма . 1 :1–87. ISSN 1342-4092 .
- Маквинни, Лорри; Карпентер, Кеннет ; Ротшильд, Брюс (2001), «25. Плечевой периостит динозавров: случай юкстакортикального поражения в ископаемых записях», Жизнь мезозойских позвоночных , Indiana University Press, стр. 364–377
- Тейлор, Майкл П. (24 января 2022 г.). «Почти все известные шеи зауроподов неполные и искаженные» . ПерДж . 10 : е12810. дои : 10.7717/peerj.12810 . ISSN 2167-8359 . ПМЦ 8793732 . ПМИД 35127288 .
- Тейлор, Майкл П.; Ведель, Мэтью Дж. (12 февраля 2013 г.). «Почему у зауроподов были длинные шеи и почему у жирафов шеи короткие» . ПерДж . 1 : е36. дои : 10.7717/peerj.36 . ISSN 2167-8359 . ПМЦ 3628838 . ПМИД 23638372 .
- Тидвелл, В.; Карпентер, К.; Мейер, С. (2001). Танке, DH; Карпентер, К. (ред.). «Новый титанозаврообразный (Sauropoda) из члена Ядовитой полосы формации Кедровая гора (нижний мел), Юта» . Жизнь мезозойских позвоночных : 139–165.
- Ведель, Мэтью Дж.; Тейлор, Майкл П. (2013). «Бифуркация нервного отдела позвоночника у динозавров-зауроподов формации Моррисон: онтогенетические и филогенетические последствия». Журнал палеонтологии позвоночных PalArch . 10 (1): 1–34. ISSN 1567-2158 .
- Крылья, Оливер; Сандер, П. Мартин (2007). «У динозавров-зауроподов нет желудочной мельницы: новые данные анализа массы и функции гастролита у страусов» (PDF) . Труды Королевского общества Б. 274 (1610): 635–640. дои : 10.1098/rspb.2006.3763 . ПМК 2197205 . ПМИД 17254987 . Архивировано из оригинала (PDF) 13 июля 2011 года . Проверено 24 июля 2009 г.
- Вудрафф, Д. Кэри; Фостер, Джон Р. (31 мая 2017 г.). «Первый экземпляр камаразавра (Dinosauria: Sauropoda) из Монтаны: самое северное возникновение рода» . ПЛОС ОДИН . 12 (5): e0177423. Бибкод : 2017PLoSO..1277423W . дои : 10.1371/journal.pone.0177423 . ISSN 1932-6203 . ПМК 5451207 . ПМИД 28562606 .











