Сауропода
| Извините | ||
|---|---|---|

| ||
| Шесть сауропод (вверху слева внизу справа): патаготитан , барозавр , жирафатитан , омеизавр , Шунозавр и Амаргасавр | ||
| Научная классификация | ||
| Домен: | Эукариота | |
| Королевство: | Животное | |
| Филум: | Chordata | |
| Клада : | Динозаурия | |
| Клада : | Сауриския | |
| Клада : | † Sauropodomorpha | |
| Клада : | † Анчисавр | |
| Клада : | † Сауропода Болото , 1878 | |
| Подгруппы | ||
| ||
| Синонимы | ||
| ||
Вау ( // ɔːˈrɒpədə/), whose members are known as sauropods (/ˈsɔːrəpɒdz/. [ 1 ] [ 2 ] От sauro- + -pod « ящерица -фотеда»), это клада сауришских , («ящеристая») динозавров . У Sauropods были очень длинные шеи, длинные хвосты, маленькие головы (по сравнению с остальной частью их тела) и четыре толстых, похожих на столб ноги. Они известны огромными размерами, достигнутыми некоторыми видами, и в группу входят самые большие животные, которые когда -либо жили на суше. Известные роды включают апатозавра , argentinosaurus , Alamosaurus , Brachiosaurus , Camarasaurus , Diplodocus и Mamenchisaurus . [ 3 ] [ 4 ]
Самые старые известные однозначные динозавры Sauropod известны из ранней юры . [ 5 ] Исанозавр и антетонитрус были первоначально описаны как триасовые сауроподы, [ 6 ] [ 7 ] Но их возраст, и в случае с Antetonitrus также его статус Sauropod были впоследствии допрошены. [ 8 ] [ 5 ] [ 9 ] Сауропод, подобные Sauropodomorph Tracks из формирования Freming Fjord ( Гренландия ), однако, указывать на возникновение группы в позднем триасе . [ 5 ] ( Покойной юрской игрой 150 миллионов лет назад) Sauropods стали широко распространенными (особенно диплодоциды и брахиозавриды ). По позднему мела , одна группа сауропод, титанозавров , заменила все остальные и имела почти глобальное распределение. Однако, как и во всех других неавийских динозаврах в то время, в то время титанозавры вымерли на мели-палеогеновом вымирании . окаменелые На каждом континенте были обнаружены останки сауропод, включая Антарктиду . [ 10 ] [ 11 ] [ 12 ] [ 13 ]
Название Сауропода было придумано Ониэлем Чарльзом Маршем в 1878 году и получено из древнегреческого , что означает «ящерица». [ 14 ] Sauropods являются одной из самых узнаваемых групп динозавров и стали фиксацией в популярной культуре из -за их впечатляющего размера.
Полные ископаемые находки Sauropod чрезвычайно редки. Многие виды, особенно крупнейшие, известны только из изолированных и рассеянных костей. Во многих почти полных образцах не хватает голов, кончиков хвоста и конечностей.
Описание
[ редактировать ]Сауроподы были травоядными (поедание растений), как правило, довольно длинно шея [ 15 ] Четырехпредвиденные (четвероногие), часто с лопаточником (Spamulafice: широкий на кончике, узкие на шее) зубы. У них были крошечные головы, массивные тела, и у большинства были длинные хвосты. Их задние ноги были густыми, прямыми и мощными, заканчивающимися клубными ногами с пятью пальцами, хотя только три (или в некоторых случаях четыре) носили когти. Их передние конечности были более стройными и обычно заканчивались руками, похожими на колонны, построены для поддержки веса; Часто только большой палец носил когти. Многие иллюстрации сауропод во плоти упускают эти факты, неточно изображая сауропод с копытами, покрывающими цифры без когтей ног, или более трех когтей или копыт на руках. Проксимальные являются каудальные позвонки чрезвычайно диагностическими для сауропод. [ 16 ]
Размер
[ редактировать ]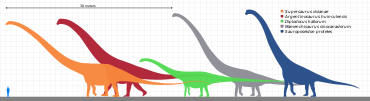
Самой определяющей характеристикой Sauropods была их размер. Даже карликовые сауроподы (возможно, от 5 до 6 метров или 20 футов) были подсчитаны среди самых больших животных в их экосистеме . Их единственные реальные конкуренты с точки зрения размера - это роркулы , такие как синий кит . Но, в отличие от китов, сауроподы были в основном наземными животными .
Их структура тела не изменилась, как и другие динозавры, возможно, из -за ограничений размера, но они продемонстрировали достаточное разнообразие. Некоторые, как диплодоциды , обладали чрезвычайно длинными хвостами, которые, возможно, смогли взломать, как кнут, как сигнал, или сдерживать или повредить хищникам, [ 17 ] или сделать звуковые бумы . [ 18 ] [ 19 ] Supersaurus , длиной от 33 до 34 метров (от 108 до 112 футов), [ 20 ] был самым длинным сауроподом, известным с разумно полных останков, но другие, как старый держатель рекордса, Diplodocus , также были чрезвычайно длинными. Голотип Maraapunisaurus (и теперь потерянный) позвонщик амфикоэлиаса Fragillimus (ныне ) мог быть получен из животного длиной 58 метров (190 футов); [ 21 ] Его позвоночная колонка была бы значительно дольше, чем у синего кита. Тем не менее, исследования, опубликованные в 2015 году, предположили, что размеры оценки A. fragillimus , возможно, были сильно преувеличены. [ 22 ] Самым длинным динозавром, известным из разумных ископаемых материалов, вероятно, является argentinosaurus huinculensis с оценками длины от 35 метров (от 115 футов) до 36 метров (118 футов) в соответствии с самыми последними исследованиями. [ 23 ] [ 24 ] Однако гигантский образец Barosaurus BYU 9024 мог бы быть еще большим достижением длины 45–48 метров (148–157 футов). [ 23 ] [ 24 ] [ 25 ]
Самое длинное наземное животное, живое сегодня, африканский слон , может достигать длины 7,3 метра (24 фута). [ 26 ]
Другие, такие как брахиозавриды , были очень высокими, с высокими плечами и очень длинными шеями. Самым высоким сауроподом был гигантский образец Barosaurus высотой 22 м (72 фута). [ 23 ] Для сравнения, жираф , самый высокий из всех живых земельных животных, составляет всего от 4,8 до 5,6 метра (от 15,74 до 18,3 фута).
Лучшие доказательства указывают на то, что наиболее массивными были аргентинозавр (65–80 метрических тонн [ 27 ] [ 23 ] [ 24 ] ), Mamenchisaurus sinocanadorum (60-80 метрических тонн [ 24 ] ), гигантский образец Barosaurus (60-80+ метрические тонны [ 23 ] [ 24 ] [ 25 ] ) и патаготитан с Пуэртасавром (50-55 метрических тонн [ 23 ] [ 24 ] ) Между тем, «мега-сауроподы», такие как Bruhathkayosaurus, давно тщательно изучались из-за спорных дебатов о ее обоснованности, но недавние фотографии перерывы в 2022 году узаконили его, [ 28 ] позволяя получить более обновленные оценки, которые варьируются от 110 до 170 тонн, что соперничает с синим китом по размеру. [ 29 ] Вес Amphicoelias fragillimus был оценен в 122,4 метрических тоннах с длиной до почти 60 метров [ 21 ] Но исследование 2015 года утверждало, что эти оценки были основаны на диплодоциду, а не на более современном рефбахисавриде, что указывает на гораздо более короткую длину 35–40 метров с массой от 80 до 120 тонн. [ 22 ] Дополнительные находки указывают на то, что ряд видов, вероятно, достигли или превысили веса 40 тонн. [ 30 ] Самое большое земельное животное, живое сегодня, слон Буша , весит не более 10,4 метрических тонн (11,5 коротких тонн). [ 31 ]
Среди самых маленьких сауропод были примитивные омденозавра (4 м или 13 футов длиной), гномочный титанозавр магьярозавр (6 м или 20 футов), а также гномового брахиозавридного европазавра , который длился 6,2 метра в длину до полного взрослого. [ 32 ] Его небольшой рост, вероятно, был результатом островного карлика , возникающего в популяции сауропод, изолированных на острове позднего юрского периода в том, что сейчас является районом Лангенберга в северной Германии . [ 33 ] [ 34 ] Диплодокоидный . Sauropod Brachytrachelopan был самым коротким членом своей группы из -за необычайно короткой шеи В отличие от других сауропод, шеи которых могли расти до четырех раз длиной их спины, шея брахитрачелопана была короче, чем ее основание.
Окаменелости из, пожалуй, самого большого динозавра, когда -либо обнаруженного, были обнаружены в 2012 году в провинции Нойкен на северо -западной Патагонии, Аргентина. Считается, что они из титанозавра, которые были одними из самых больших сауропод. [ 35 ] [ 30 ]
Или незадолго до 29 марта 2017 года в Уоуроподе в регионе Кимберли в регионе Кимберли в регионе Кимберли в регионе Кимберли было обнаружено в регионе Кимберли. [ 36 ] В отчете говорится, что это было самым самым известным. В 2020 году Молина-Перез и Ларраменди оценили размер животного на 31 метрах (102 фута) и 72 тонн (79,4 коротких тонн) на основе длиной 1,75 метра (5,7 фута). [ 23 ]
Конечности и ноги
[ редактировать ]Как массивные четвероногих , Sauropods разработали специализированные «гравипортальные» (весовые) конечности. Задние ноги были широкими и сохранили три когти у большинства видов. [ 37 ] Особенно необычно по сравнению с другими животными были высоко измененные передние ноги ( Manus ). Передние ноги сауропод были очень разными от ног современных крупных четвероногих, таких как слоны . Вместо того, чтобы раскрыть боковые стороны, чтобы создать широкую ногу, как у слонов, кости манус -сауроподов были расположены в полностью вертикальных колоннах, с чрезвычайно уменьшенными костями пальцев (хотя неясно, наиболее примитивные сауроподы, такие как Вулканодон и Барапасавр , имел такую переднюю часть). [ 38 ] Передние ноги были настолько изменены в Eusauropods , что отдельные цифры не были бы видны в жизни.
Расположение колонн кости передней передней части ( метакарпала ) в Eusauropods было полукруглым, так что отпечатки передней части стопы Sauropod являются подковообразными. В отличие от слонов, печатные данные показывают, что у Sauropods не было каких -либо мясистых прокладков, чтобы поднять передние ноги, что делает их вогнутыми. [ 38 ] Единственным когтям, видимым в большинстве сауропод, был отличительный когтя пальца (связанный с цифрой I). Почти у всех зауроподов был такой когти, хотя какая цель он служил, неизвестно. Коготь был самым большим (а также высоким и боковым сплюсением) в диплодоцидах и очень маленьким в брахиозавридах, некоторые из которых, по -видимому, потеряли когти, полностью основанный на доказательствах Trackway. [ 39 ] Титанозавры, возможно, полностью потеряли когти большого пальца (за исключением ранних форм, таких как Джоненшия ).
Титанозавры были наиболее необычными среди сауроподов, так как в своей истории как клада они потеряли не только внешний когти, но и полностью потеряли цифры передней ноги. У продвинутых титанозавров не было цифр или костей цифр, и они ходили только по подковообразным «пнятам», состоящим из столбчатых костей метакарпала. [ 40 ]
Печатные доказательства из Португалии показывают, что, по крайней мере, в некоторых сауроподах (вероятно, брахиозавридах) дно и стороны колонны стопы, вероятно, были покрыты небольшими колючими масштабами, которые оставили оценки в печатных изданиях. [ 41 ] У титанозавров концы костей метакарпала, которые связывались с землей, были необычно широкими и квадратными, а некоторые образцы сохраняют остатки мягких тканей, покрывающих эту область, что позволяет предположить, что передние ноги были заправлены с некоторой прокладкой у этих видов. [ 40 ]
Мэтью Боркс [ 42 ] [ 43 ] показал, что длинные кости динозавров Sauropod выросли изометрически : то есть не было никаких изменений в форме, так как ювенильные сауроподы стали гигантскими взрослыми. Bonnan предположил, что эта нечетная паттерна масштабирования (большинство позвоночных показывают значительные изменения формы в длинных костях, связанных с увеличением поддержки веса), может быть связана с принципом с дыханием (предполагаемый ученый-любитель Джим Шмидт), в котором разрешены длинные ноги взрослых сауропод. Легко покрыть большие расстояния, не изменяя их общую механику.
Воздушные мешки
[ редактировать ]Наряду с другими сауришскими динозаврами (такими как тероподы , включая птиц), у Sauropods была система воздушных мешков , о чем свидетельствуют углубления и полые полости в большинстве их позвонков , которые были захвачены ими. Пневматические, полые кости являются характерной особенностью всех сауропод. [ 44 ] Эти воздушные пространства уменьшили общий вес массивных шеей, которые были у сауроподов, и система воздушного SAC в целом, что позволила получить одно направление воздушного потока через жесткие легкие, позволила сауроподам получить достаточно кислорода. [ 45 ] Эта адаптация имела бы преимущественную силу, особенно в относительно низких условиях кислорода юрского и раннего мела. [ 46 ]
Подобная птицам, впаловая костей Сауропода была признана в начале изучения этих животных, и, по сути, по крайней мере один образец Sauropod, обнаруженный в 19 веке ( Ornithopsis ), был первоначально ошибочно идентифицирован как птерозавр (летающая рептилия) из-за этого Полем [ 47 ]
Доспехи
[ редактировать ]
У некоторых сауропод были броня . Были роды с небольшими клубами на их хвостах, выдающимся примером «Шунозавра» , и у нескольких титанозавров , таких как Saltasaurus и Ampelosaurus , были небольшие костные остеодермы, покрывающие части своего тела.
Зубы
[ редактировать ]Исследование, проведенное Майклом Д'Имиком и его коллегами из Университета Стони Брук, показало, что Sauropods эволюционировали высокие показатели замены зубов, чтобы не отставать от своих больших аппетитов. Исследование показало, что Нигрансирс , например, заменял каждый зуб каждые 14 дней, камаразавр заменял каждый зуб каждые 62 дня, а диплодокус заменял каждый зуб раз каждые 35 дней. [ 48 ] Ученые обнаружили, что качества зуба повлияли на то, сколько времени потребовалось для роста нового зуба. Зубы Camarasaurus заняли больше времени, чем на диплодокус , потому что они были больше. [ 49 ]
Это также было отмечено D'Emic и его командой, что различия между зубами сауропод также указывали на разницу в рационе. Diplodocus съел растения низко до земли, и Camarasaurus просмотрел листья из верхней и средней ветви. По словам ученых, специализация их рационов помогла различным травоядным динозаврам сосуществовать. [ 48 ] [ 49 ]
Шейки
[ редактировать ]Шеи Sauropod были обнаружены на расстоянии более 15 метров (49 футов) в длину, в шесть раз дольше, чем шея жирафа мирового рекорда. [ 45 ] Включение этого было ряд важных физиологических особенностей. Общий большой размер тела и четверопрейк динозавров обеспечил стабильное основание для поддержки шеи, а голова была эволюционирована, чтобы быть очень маленькой и легкой, теряя способность устной обработки пищи. Снив их головы до простых инструментов для сбора урожая, которые привели растения в организм, Sauropods требовалось меньше силы, чтобы поднять головы, и, таким образом, смогли развивать шеи с менее плотными мышцами и соединительной тканью. Это резко уменьшило общую массу шеи, что позволило дальнейшему удлинению.
Sauropods также имел большое количество адаптаций в своей скелетной структуре. У некоторых сауропод было целых 19 шейных позвонков , тогда как почти все млекопитающие ограничены только семь. Кроме того, каждый позвонок был чрезвычайно длинным и имел в них несколько пустых пространств, которые были бы заполнены только воздухом. Система воздуха-SAC, подключенная к пространствам, не только осветляла длинные шеи, но и эффективно увеличивала поток воздуха через трахею, помогая существам дышать достаточно воздухом. Эволюционируя позвонки, состоящие из 60% воздуха, сауроподы смогли минимизировать количество плотной тяжелой кости, не жертвуя способностью делать достаточно большие вдохи, чтобы питать все тело кислородом. [ 45 ] По словам Кента Стивенса, компьютерные реконструкции скелетов, изготовленных из позвонков, указывают на то, что шеи сауропода были способны вытащить большие области кормления без необходимости перемещать свои тела, но не смогли быть втянуты в положение намного выше плеч область или достигая выше. [ 50 ]
Другой предложенной функцией длинных шеей Sauropods была, по сути, радиатором, чтобы справиться с крайним количеством тепла, полученного из их большой массы тела. Учитывая, что метаболизм выполнял бы огромный объем работы, он, безусловно, принесет бы большое количество тепла, и устранение этого избыточного тепла было бы необходимым для выживания. [ 51 ] Также было предложено, чтобы длинные шеи охладили вены и артерии, идущие в мозг, избегая чрезмерно нагретой крови от достижения головы. На самом деле было обнаружено, что увеличение скорости метаболизма в результате шеей сауропод было немного больше, чем компенсируется дополнительной площадью поверхности, с которой тепло может рассеиваться. [ 52 ]
Палеобиология
[ редактировать ]Экология
[ редактировать ]Когда сначала были обнаружены зауропод, их огромный размер заставил многих ученых сравнить их с современными китами . Большинство исследований в 19 -м и начале 20 -го веков пришли к выводу, что сауроподы были слишком большими, чтобы поддерживать свой вес на суше, и поэтому они, должно быть, были в основном водными . Большинство реставраций жизни сауропод в искусстве в течение первых трех четвертей 20 -го века изображали их полностью или частично погружены в воду. [ 53 ] Это раннее представление было вызвано сомнением, начиная с 1950 -х годов, когда исследование Кермака (1951) продемонстрировало, что, если животное было погружено в несколько метров воды, давление было бы достаточным для смертельного разрушения легких и дыхательных путей. [ 54 ] Sauropod Тем не менее, эти и другие ранние исследования экологии были ошибочны в том смысле, что они игнорировали существенное количество доказательств того, что тела сауропод были сильно проницались с воздушными мешками . В 1878 году палеонтолог Эд Коуп даже назвал эти структуры «поплавками».
Начиная с 1970 -х годов, началось влияние воздушных мешков Sauropod на их предполагаемый водный образ жизни. Палеонтологи, такие как Кумбс и Баккер, использовали это, а также доказательства седиментологии и биомеханики , чтобы показать, что сауроподы были в основном наземными животными. В 2004 году Д.М. Хендерсон отметил, что из -за их обширной системы воздушных мешочков Сауроподы были бы оживленными и не смогли бы погрузить свои туловища полностью под поверхностью воды; Другими словами, они будут плавать, и не подвергались бы опасности коллапса легких из -за давления воды при плавании. [ 53 ]
Свидетельство о плавании в сауроподах поступает от ископаемых дорожек, которые иногда были обнаружены, которые сохраняют только впечатления перед передней (MANUS). Хендерсон показал, что такие дорожки могут быть объяснены сауроподами с длинными передними передниками (такими как макронарианцы ), плавающие в относительно мелкой воде, достаточно глубоко, чтобы держать более короткие задних лапок свободными от дна, и использование передних конечностей для удара вперед. [ 53 ] Однако из -за их пропорций тела плавающие сауроподы также были бы очень нестабильными и не работали в течение длительных периодов в воде. Этот режим водного локомоции в сочетании с его нестабильностью привел Хендерсона к обозначению Sauropods в воде как «плавными игроками». [ 53 ]
В то время как Sauropods, следовательно, не могли быть водными, как исторически изображено, есть доказательства того, что они предпочитают влажную и прибрежную среду обитания. Следы Sauropod обычно встречаются после береговых линий или пересечения пойменных залогов, а окаменелости Sauropod часто встречаются во влажной среде или смешаются с окаменелостями морских организмов. [ 53 ] Хорошим примером этого могут быть массивные юрские дорожки Сауропод, найденные в месторождениях лагуны на Шотландии острове Скай . [ 55 ] Исследования, опубликованные в 2021 году, показывают, что Сауроподы не могли обитать полярные регионы. Это исследование предполагает, что они были в значительной степени ограничены тропическими районами и имели метаболизм, которые сильно отличались от метаболистов других динозавров, возможно, промежуточных между млекопитающими и рептилиями. [ 56 ] Новые исследования, опубликованные Taia Wyenberg-Henzler в 2022 году, показывают, что Sauropods в Северной Америке снизились из-за неопределенных причин в отношении их ниш и распространения в конце юрского периода и в последних меловых. Почему это остается неясным, но было предложено некоторые сходства в кормлении ниш между игуанодонтианцами, хадрозавроидами и сауроподами и, возможно, привели к некоторым соревнованиям. Тем не менее, это не может полностью объяснить полное снижение распределения Sauropods, поскольку конкурентное исключение привело бы к гораздо более быстрому снижению, чем то, что показано в ископаемом отчете. Более того, должно быть определено, что Sauropod снижение в Северной Америке является результатом изменения предпочтительной флоры, которую eauropods, климат или другие факторы. В этом же исследовании также предполагается, что игуанодонтианцы и хадрозавроиды воспользовались недавно освобожденными нишами, оставленными снижением разнообразия Sauropod во время поздней юры и меловой в Северной Америке. [ 57 ]
Почист и родительская помощь
[ редактировать ]
Многие линии ископаемых доказательств, как из костных слоев, так и с дорожек, указывают на то, что сауроподы были общительными животными, которые образовали стада . Тем не менее, состав стад варьировался между видами. участок из средней юры Аргентины Некоторые костяные кровати, например , , показывают стада, состоящие из людей разных возрастных групп, смешивая несовершеннолетних и взрослых. Тем не менее, ряд других окаменелостей и дорожных путей указывают на то, что многие виды Sauropod, путешествующие в стаде, отделенные по возрасту, а несовершеннолетние образуют стада, отделенные от взрослых. Такие сегрегированные стратегии пала были обнаружены у таких видов, как Alamosaurus , Bellusaurus и некоторые диплодоциды . [ 58 ]
В обзоре доказательств различных типов стада Майерс и Фиорилло попытались объяснить, почему сауроподы, по -видимому, часто образовали отдельные стада. Исследования микроскопического зубного износа показывают, что у юношеских сауропод были диеты, которые отличались от их взрослых коллег, поэтому уделение вместе не было бы столь продуктивным, как пасти отдельно, где отдельные члены стада могли бы добывать координированным образом. Огромная разница в размерах между несовершеннолетними и взрослыми также может сыграть свою роль в различных стратегиях кормления и пасти. [ 58 ]

Поскольку сегрегация несовершеннолетних и взрослых, должно быть, произошла вскоре после вылупления и в сочетании с тем фактом, что птенцы Sauropod, скорее всего, были предварительными , Майерс и Фиорилло пришли к выводу, что виды с возрастными стадами не будут проявлять большой родительской помощи. [ 58 ] С другой стороны, ученые, которые изучали стада Sauropod, изучали возрастные стада Sauropod, предположили, что эти виды, возможно, заботились о своих молодых в течение длительного периода времени, прежде чем молодые достигли взрослой жизни. [ 59 ] Исследование в 2014 году показало, что время от укладки яйца до времени вылупления, вероятно, было между 65 и 82 днями. [ 60 ] Точно то, как сегрегированное по сравнению с возрастным пастом варьировалось в разных группах сауропод, неизвестно. Дальнейшие примеры общительного поведения должны быть обнаружены от большего количества видов Sauropod, чтобы начать обнаружение возможных моделей распределения. [ 58 ]
Выращивание позиции
[ редактировать ]
С начала истории своего исследования ученые, такие как Осборн , предположили, что Sauropods могут поднять свои задние ножки, используя хвост в качестве третьей «ноги» штатива. [ 61 ] Скелетное крепление, изображающее диплодоцид Barosaurus Lentus, поднимающийся на задних ногах в Американском музее естественной истории, является одной из иллюстраций этой гипотезы. В документе 2005 года Ротшильд и Молнар рассуждали, что если бы Сауроподс время от времени принял двуногих позов, будут свидетельствуют о стрессовых переломах в передней основе «руки». Тем не менее, ни один из них не был найден после того, как они исследовали большое количество скелетов Sauropod. [ 62 ]

Генрих Маллисон (в 2009 году) был первым, кто изучил физический потенциал для различных сауропод, чтобы сразиться в триподальную позицию. Маллисон обнаружил, что некоторые символы, ранее связанные с адаптацией выращивания, были на самом деле не связаны (например, широкоизвестные кости бедра титанозавров ) или было бы препятствовало бы выращивания. Например, у титанозавров была необычайно гибкая магистраль, которая бы снизилась бы устойчивости в тройной осанке и могло бы увеличить нагрузку на мышцы. Аналогично, маловероятно, что брахиозавриды могли подняться на задние ножки, так как их центр тяжести был намного дальше вперед, чем другие сауроподы, что приведет к нестабильной стойке. [ 63 ]
Диплодоциды, с другой стороны, по -видимому, были хорошо адаптированы для подъема в триподальную позицию. Диплодоциды имели центр масс прямо над бедрами, что дало им больший баланс на двух ногах. У диплодоцидов также были самые подвижные шеи сауропод, хорошо мышечный тазовый пояс и хвостовые позвонки со специализированной формой, которая позволила бы хвосту иметь вес в точке, который он касался земли. Маллисон пришел к выводу, что диплодоциды были лучше адаптированы к выращиванию, чем слоны , которые иногда делают это в дикой природе. Он также утверждает, что стрессовые переломы в дикой природе не происходят из повседневного поведения, [ 63 ] такие как активность, связанные с кормлением (Contra Rothschild и Molnar). [ 62 ]
Поза головы и шеи
[ редактировать ]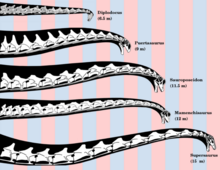
Существует мало соглашения о том, как Sauropods держали головы и шеи, и о позах, которые они могли бы достичь в жизни.
Можно ли использовать длинные шеи Sauropods для просмотра высоких деревьев, на основе расчетов, предполагающих, что просто накапливая кровь до головы в такой позе [ 64 ] Надолго использовал бы немного половины его потребления энергии. [ 65 ] Далее, чтобы перемещать кровь к такой высоте - дискизирование поставлено вспомогательные сердца в шею [ 66 ] -потребовалось бы сердце в 15 раз больше, чем кит одинакового размера. [ 67 ]
Вышеупомянутое использовалось для утверждения, что длинная шея должна быть более или менее горизонтально, предполагается, что позволяет питаться растениями на обширной площади с меньшим количеством необходимости двигаться, что дает большую энергию для такого большого животного. Поэтому реконструкции шеей диплодока и апатозавра часто изображают их в почти горизонтальной, так называемой «нейтральной, нефтьющей позе». [ 68 ]
Тем не менее, исследования на живых животных показывают, что почти все существующие тетрапод держат основание их шеев, резко сгибаясь, когда настороженно, показывая, что любой вывод от костей о привычных «нейтральных позах» [ 68 ] глубоко ненадежно. [ 69 ] [ 70 ] Между тем, компьютерное моделирование шеей страусов вызвало сомнения по поводу гибкости, необходимой для стационарного выпаса. [ 71 ] [ 72 ] [ 73 ]
Треки и передвижение
[ редактировать ]


Sauropod Trackways и другие ископаемые следы (известные как «ichnites») известны из обильных доказательств, присутствующих на большинстве континентов. Ихниты помогли поддерживать другие биологические гипотезы о сауроподах, включая анатомию общего и задних ног (см. Конечности и ноги выше). Как правило, отпечатки от передней части намного меньше задних ног и часто в форме полумесяца. Иногда iChnites сохраняют следы когтей и помогают подтвердить, какие группы Sauropod потеряли когти или даже цифры на их передней части. [ 74 ]
Sauropod Tracks из формации Villar del Arzobispo раннего возраста Берриаса в Испании поддерживает общительное поведение группы. Треки, возможно, больше похожи на Sauropodichnus giganteus , чем на любой другой iChnogenera, хотя они предположительно из базальной титанозавриформ. Треки широко калика, и группировка как можно ближе к Sauropodichnus , также поддерживается расстоянием мануса к PES, морфология мануса была в форме фасоли и морфологией PES субтриангулярной. Нельзя определить, были ли следы стада, вызванные несовершеннолетними или взрослыми из -за отсутствия предыдущей идентификации возрастного возраста. [ 75 ]

Как правило, дорожки Sauropod разделены на три категории на расстоянии между противоположными конечностями: узкий маномотр, средний датчик и широкий калибр. Значение дорожного движения может помочь определить, насколько широкими были конечности различных сауропод и как это могло повлиять на то, как они ходили. [ 74 ] Исследование 2004 года днем, и его коллеги показали, что среди групп продвинутых сауропод можно найти общую модель, причем каждая семейство Sauropod характеризуется определенными датчиками Trackway. Они обнаружили, что у большинства сауропод, кроме титанозавров, были узкие конечности, с сильными впечатлениями от большого когтя большого пальца на передней части. Треки среднего значения с оттисками когтя на передней части, вероятно, принадлежат брахиозавридам и другим примитивным титанозавриформ , которые развивали более широкие конечности, но сохраняли свои когти. Примитивные настоящие титанозавры также сохранили свой когти стопы, но развивали полностью широкие конечности. Усовершенствованные титанозавры сохранили широкие датчики, с которыми проводятся дорожки, из которых показывают широкий диапазон и отсутствие каких -либо когтей или цифр на передней части. [ 76 ]
Иногда обнаруживаются только треки от передней части. Falkingham et al. [ 77 ] Используется компьютерное моделирование, чтобы показать, что это может быть связано со свойствами субстрата. Они должны быть правы, чтобы сохранить следы. [ 78 ] Различия в задней конечности и площади поверхности передней конечности, и, следовательно, контактное давление с подложкой могут иногда привести к сохранению только напрасений перед передней.
Биомеханика и скорость
[ редактировать ]
В исследовании, опубликованном в PLOS One 30 октября 2013 года, Билл Селлерс , Родольфо Кори , Ли Маргеттс и соавт. , Аргентинозавр был реконструирован в цифровом виде, чтобы впервые проверить его локомоцию. Перед исследованием наиболее распространенным способом оценки скорости было изучение гистологии кости и ихнологии . Обычно исследования о гистологии кости и скорости Sauropod и скорости сосредоточены на посткраниальном скелете, который обладает многими уникальными особенностями, такими как увеличенный процесс на локте , широкая доля на подвздошной кости , внутренняя верхняя треть бедра и чрезвычайно Яйцевидный вал бедренной кости. Эти функции полезны при попытке объяснить схемы следов животных гравипортала. При изучении ихнологии для расчета скорости SauroPod существует несколько проблем, таких как предоставление только оценки для определенных походков из -за смещения сохранения и подверженности гораздо большему количеству проблем точности. [ 79 ]
Чтобы оценить походку и скорость Аргентинозавра , исследование провели анализ опорно -двигательного аппарата. Единственный предыдущий анализ опорно -двигательного аппарата проводился на гоминоидах , террористических птицах и других динозаврах . Прежде чем они смогли провести анализ, команде пришлось создать цифровой скелет рассматриваемого животного, показать, где будет наслоение мышц, найти мышцы и суставы и, наконец, найти свойства мышц, прежде чем найти походку и скорость. Результаты исследования биомеханики показали, что аргентинозавр был механически компетентен на максимальной скорости 2 м/с (5 миль в час), учитывая большой вес животного, и штамм, который способен поддерживать его суставы. [ 80 ] Результаты также показали, что гораздо более крупные наземные позвоночные могут быть возможными, но потребуют значительного ремоделирования тела и возможных достаточных изменений в поведении для предотвращения обрушения суставов. [ 79 ]
Размер тела
[ редактировать ]
Сауроподы были гигантскими потомками удивительно маленьких предков. Базальные динозавриформ, такие как Pseudolagosuchus и Marasuchus из среднего триаса Аргентины, весили приблизительно 1 кг (2,2 фунта) или меньше. Они превратились в Сауришию, которая наблюдалась за быстрым увеличением размера баупланов , хотя более примитивные члены, такие как Эораптор , Панфагия , Пантидрако , Сатурналия и Гуабасавр , все еще сохраняли умеренный размер, возможно, менее 10 кг (22 фунта). Даже с этими небольшими, примитивными формами, среди сауроподоморфов наблюдается заметное увеличение размера, хотя скудные остатки этого периода делают интерпретацию явной. Есть один определенный пример небольшого производного сауроподоморф: анкисавр , менее 50 кг (110 фунтов), даже если он ближе к сауроподам, чем платформы и риоджасаурус , которые были более 1 т (0,98 длинные тонны; 1,1 короткие тонны) в) в масса. [ 51 ]
Развивающиеся от Sauropodomorphs, Sauropods были огромными. Их гигантский размер, вероятно, возникла в результате повышенной скорости роста, ставшей возможной благодаря тахиметаболической эндотермии , черту, которая развивалась в сауроподоморфе. Когда -то разветвившись в сауропод, сауроподоморфы постоянно продолжали расти больше, с более мелкими сауроподами, такими как ранняя юрская барапасавр и котасавр , превращающихся в еще более крупные формы, такие как средняя юрская среда мамансизавра и патагозавр . Отвечая на рост сауропод, их хищники также росли, как показано в аллозавром с целофизоиде из Германии . [ 51 ]
Размер в Neosauropoda
[ редактировать ]Neosauropoda вполне правдоподобно клада динозавров с самыми большими размерами тела, когда -либо существовавшими. Предполагается, что немногие исключения меньшего размера вызваны островным карликом или другим экологическим давлением есть тенденция , хотя в некоторых титанозаврии к меньшему размеру. Титанозавры , однако , были одними из крупнейших сауропод за всю историю. Помимо титанозавров, диплодокоиды также достигли действительно гигантских размеров. Между тем, клада диплодокоидов, называемых dicraeosauridae , идентифицируется небольшим и средним [ нужно разъяснения ] размер тела. Однако ни один сауропод не был очень маленьким, поскольку даже «карликовые» сауроподы превышают 500 кг (1100 фунтов), что достигается только около 10% всех видов млекопитающих. [ 51 ]
Независимый гигантизм
[ редактировать ]Несмотря на то, что в целом, сауроподы были большими, гигантский размер (40 т (39 длинных тонн; 44 коротких тонн) или более) был достигнут независимо в их эволюции несколько раз в их эволюции. Многие гигантские формы существовали в поздней юре (в частности, kimmeridgian ), таких как Turiasaur Turiasaurus , Mamenchisaurids Mamenchisaurus и Xinjiangtitan , Diplodocoids Maraapunisaurus , Diplodocus , Apatosaurus , Supersaurus и Barosaurus , Bracharivis , и Gracharusauris, Grachauris и а также анонца , Благодаря раннему и позднему мелу, гиганты Borealosaurus , Sauroposeidon , Paralitan , Argentinosaurus , Puertasaurus , Antarctosaurus , Dreadnawhoud , Notocolossus , Futalognkosaurus , Patagotitan и Alamosaurus , со всеми возможными. Одним из известных возможных гигантов является Huanghetitan ruyangensis , известный только из 3 м (9,8 фута) ребер. Эти гигантские виды жили в конце юры до позднего мела, появляясь независимо в течение 85 миллионов лет. [ 51 ]
Дварфизм в сауроподах
[ редактировать ]Два известных островных карликовых видов сауропод являются меловодным магьярозавром (в какой-то момент его идентичность как карлика была оспорена) и юрского европаурара , оба из Европы. Несмотря на то, что эти сауроподы маленькие, единственный способ доказать, что они являются настоящими карликами, - это исследование их гистологии кости. Исследование, проведенное Мартином Сандером и коллегами в 2006 году, изучило одиннадцать особей Europasaurus Holgeri с использованием гистологии кости и продемонстрировало, что мелкие виды острова эволюционировали благодаря снижению скорости роста длинных костей по сравнению с темпами роста у наследственных видов на материке. [ 81 ] Двумя другими возможными гномами являются Rapetosaurus , который существовал на острове Мадагаскар , изолированный остров на меловом месте, и Ampelosaurus , титанозавр, который жил на иберийском полуострове на юге Испании и Франции. Аманция из Швейцарии также может быть гномом, но это еще не доказано. [ 51 ] Один из наиболее крайних случаев островного карлизма встречается в Европазавре , родственник гораздо более крупного камаразавра и брахиозавра : это было всего около 6,2 м (20 футов), что идентифицирующая черта вида. Что касается всех видов карликов, их снижение скорости роста привело к их небольшим размерам. [ 32 ] [ 51 ] Еще один таксон крошечных сауропод, титанозавр саласавридов Ибирании , длиной 5,7 м (18,7 фута), жил в необщепринятом контексту в верхней креационной Бразилии и является примером нанизма, полученного в результате других экологических давлений. [ 82 ]
Палеопатология и палеопаразитология
[ редактировать ]Сауроподы редко известны сохраненными травмами или признаками болезней, но более недавние открытия показывают, что они могут страдать от таких патологий. Образец диплодоцид из формирования Моррисона, называемый «Долли», был описан в 2022 году с признаками тяжелой респираторной инфекции. [ 83 ] [ 84 ] Ребра Сауропода из округа Юньян , Чунцин , в юго -западном Китае, показывают свидетельства разрушения ребер в результате травматического перелома, кости инфекции и остеосклероза . [ 85 ]
Ибирания , наноидный ископаемый титанозавр из Бразилии , предполагает, что люди различных родов были восприимчивы к таким заболеваниям, как остеомиелит и заражения паразитами. Образец родом из поздней меловой формирования Сан -Хосе -де -Рио -Пот -Пот -Формирование , Бауру , и был описан в журнале Cretaceous Research Aureliano . et al (2021). [ 86 ] Исследование костей титанозавра выявило то, что, по-видимому, представляют собой паразитические кровные черви, похожие на доисторическую палеолеишманию , но в 10-100 раз больше, что, по-видимому, вызвало остеомиелит. Окаменелость является первым известным примером агрессивного случая, когда остеомиелит вызван кровавыми червями у вымершего животного. [ 87 ] [ 88 ] [ 89 ]
История открытия
[ редактировать ]Первые отходы ископаемых остается теперь признаны как Sauropods, все поступили из Англии и первоначально были интерпретированы различными способами. Их отношения с другими динозаврами не были признаны до тех пор, пока их первоначальное открытие.


Первым ископаемым Sauropod, который был научно описан, был единственный зуб, известный под - Линнея не дескриптором . [ 90 ] Это ископаемое было описано Эдвардом Лхуйдом в 1699 году, но в то время не было признано гигантской доисторической рептилей. [ 91 ] Динозавры не будут признаны группой более столетия спустя.
Ричард Оуэн опубликовал первые современные научные описания Sauropods в 1841 году, в книге и в статье «Кардиодон» и «Цетиозавр» . Cardiodon был известен только из двух необычных зубов в форме сердца (от которых он получил свое название), которые не могли быть идентифицированы помимо того, что они пришли из ранее неизвестной большой рептилии . Цетиозавр был известен с немного лучше, но все еще остается ловко. В то время Оуэн подумал, что Цетиозавр был гигантской морской рептилей, связанной с современными крокодилами , отсюда и его название, что означает «китовую ящерицу». Год спустя, когда Оуэн придумал название Dinosauria , он не включил Cetiosaurus и Cardiodon в эту группу. [ 92 ]
В 1850 году Гидеон Мантелл узнал динозаврский характер нескольких костей, назначенных Цетиозавру Оуэном. Мантелл заметил, что кости ноги содержали медуллярную полость , характерную для сухопутных животных. Он назначил эти образцы новому роду Pelorosaurus и сгруппировал их вместе с динозаврами. Тем не менее, Мантелл до сих пор не признал связь с Цетиозавром . [ 47 ]
Следующим Sauropod считается, что его описание и неверно идентифицированное как нечто иное, чем динозавр, представляли собой набор тазобедренных позвонков, описанных Гарри Сили в 1870 году. Сили обнаружил, что позвонки были очень слегка построены для их размера и содержали отверстия для воздушных мешков ( припневматизация ). Такие воздушные мешки были в то время известны только у птиц и птерозавров , и Сили считал, что позвонки поступают от птерозавра. Он назвал новый род Ornithopsis или «птичье лицо» из -за этого. [ 47 ]
Когда в 1871 году Филлипс был описан более полные образцы Цетиоооооооооура , он, наконец, узнал животное как динозавр, связанный с пелорозавром . [ 93 ] Тем не менее, только в том, что описание новых, почти полных скелетов Sauropod из Соединенных Штатов (представляющих апатозавра и камаразавра ) в том же году, появилась полная картина сауропод. Приблизительная реконструкция полного скелета Sauropod была произведена художником Джоном А. Райдером, нанятым палеонтологом Эд Коупом, на основе останков Camarasaurus , хотя многие признаки все еще были неточными или неполными в соответствии с более поздними находками и биомеханическими исследованиями. [ 94 ] Также в 1877 году Ричард Лидеккер назвал еще одного родственника Цетиозавра , Титанозавра , на основе изолированного позвонка. [ 47 ]

В 1878 году был найден самый полный сауропод, который был найден и описал Отниэль Чарльз Марш , который назвал его Diplodocus . С помощью этой находки Марш также создал новую группу для содержания Diplodocus , Cetiosaurus и их растущего списка родственников, чтобы отличить их от других основных групп динозавров. Марш назвал эту группу Sauropoda, или «ноги ящериц». [ 47 ]
Классификация
[ редактировать ]Первое филогенетическое определение Sauropoda было опубликовано в 1997 году Salgado и коллегами. Они определили кладу как таксон на основе узлов , содержащий « самого последнего общего предка Vulcanodon Karibaensis и Eusauropoda и всех его потомков». [ 95 ] Позже было предложено несколько определений на основе STEM , в том числе Yates (2007), который определил Sauropoda как «самую инклюзивную кладу, которая включает Saltasaurus loricatus, но не Melanorosaurus readi ». [ 96 ] [ 97 ]
Сторонники этого определения также используют название клады Gravisauria , определяемое как самый последний предок Tazoudasaurus naimi и Saltasaurus Loricatus и все его потомки [ 98 ] Для клады, эквивалентной Sauropoda, как определено Salgado et al. [ 99 ] Клада марокканским Грейсисаврия была назначена французским палеонтологом Ронаном Алланом и что палеонтологом Наджатом Акесби в 2008 году, когда кладистический анализ динозавра, обнаруженный Аллайном, Тазудасавром , как результат заключался в том, семейство Вулканодонтидов . Группа включает в себя Tazoudasaurus и Vulcanodon , а также сестринский таксон Eusauropoda, а также некоторые виды , такие как Antetonitrus , Gongxianosaurus и Isanosaurus, которые принадлежат не Vulcanodontidae, но и к еще более базовому положению, занятому в Sauropoda. Имело смысл иметь Sauropoda по сравнению с этой, более производной группой, которая включала Vulcanodontidae и Eusauropoda в определение: определяется как группа, сформированная последним общим предком Tazoudasaurus и Saltasaurus (Bonaparte and Powell, 1980) и все его потомки. характеристики гравизной : Акесби упомянул две синапоморфии, общие Полем Ранее они не считались синапоморфиями Eusauropoda, но Allian обнаружил эти свойства также на Tazoudasaurus . [ 100 ]
Gravisauria разделена в ранней юре , вокруг Плайенсбахианского и Торциана , 183 миллиона лет назад, и Акесби подумал, что это было частью гораздо большей революции в фауне , которая включает в себя исчезновение Prosauropoda , Coelophysoidea и Basal Thyreophora . к всемирному массовому вымиранию . [ 100 ]
Филогенетические отношения сауроподов в последние годы в значительной степени стабилизировались, хотя все еще есть некоторые неопределенности, такие как размещение Euhelopus , Haplocanthosaurus , Jobaria и Nemegtosauridae .
Кладограмма после анализа, представленного Сандером и коллегами в 2011 году. [ 51 ]
| † Сауропода |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ "Сауропод" . Merriam-Webster.com Словарь . Мерриам-Уэбстер.
- ^ "Сауропод" . Dictionary.com unabridged (онлайн). н.д.
- ^ Tschopp, E.; Матеус, О.; Бенсон, RBJ (2015). «Филогетический анализ уровня образца и таксономический пересмотр Diplodocidae (Dinosauria, Sauropoda)» . ПЕРЕЙ . 3 : E857. Doi : 10.7717/peerj.857 . PMC 4393826 . PMID 25870766 .
- ^ blogss.sientificamerican.com Tetrapod-zoology 2015-04-24, что Brontosaurus thing
- ^ Jump up to: а беременный в Jens N. Lallensack; Хендрик Кляйн; Джеспер Милан; Оливер Крылья; Octávio Mateus; Ларс Б. Клеменсен (2017). «Следы по сауроподоморфу динозавров из Фьорда Фьорда Фьорда Восточной Гренландии: свидетельство позднего триасового сауропод» . Acta Palaeontologica Polonica . 62 (4): 833–843. doi : 10.4202/app.00374.2017 . HDL : 10362/33146 .
- ^ Эрик Баффетат; Варавуд Сьютетерн; Жиль Куни; Хайян Тонг; Жан Ле Лоуфф; Сасидхорн Хансубха; Sutee Jongautchariyakul (2000). «Самый ранний известный динозавр Сауропода». Природа . 407 (6800): 72–74. Bibcode : 2000natur.407 ... 72b . doi : 10.1038/35024060 . PMID 10993074 . S2CID 4387776 .
- ^ Адам М. Йейтс; Джеймс У. Китчинг (2003). «Самый ранний известный динозавр Sauropod и первые шаги к локомоции Sauropod» . Труды Королевского общества B: Биологические науки . 270 (1525): 1753–1758. doi : 10.1098/rspb.2003.2417 . PMC 1691423 . PMID 12965005 .
- ^ Блэр У. Макфи; Адам М. Йейтс; Джона Н. Чоиньера; Фернандо Абдала (2014). «Полная анатомия и филогенетические отношения антетонитрусических ингенипов (Sauropodiformes, Dinosauria): последствия для происхождения Sauropoda». Зоологический журнал Линневого общества . 171 (1): 151–205. doi : 10.1111/Zoj.12127 . S2CID 82631097 .
- ^ Блэр У. Макфи; Эмес М. Борди; Лара Сцискио; Jonah N. Choiniere (2017). «Биостратиграфия сауроподоморф о формировании эллиота Южной Африки: отслеживание эволюции сауроподоморфы через триасовую границу» . Acta Palaeontologica Polonica . 62 (3): 441–465. doi : 10.4202/app.00377.2017 .
- ^ Фернандо Э. Новас (2009). Эпоха динозавров в Южной Америке Издательство Университета Индианы. П. 181. ISBN 978-0-253-35289-7 .
- ^ Геология Оклахома Примечания . Геологическая служба Оклахомы. 2003. с. 40
- ^ Beau Riffenburgh (2007). Энциклопедия Антарктики . Тейлор и Фрэнсис. п. 415. ISBN 978-0-415-97024-2 .
- ^ JJ Alistair Crame; Геологическое общество Лондона (1989). Происхождение и эволюция антарктической биоты . Геологическое общество. п. 132. ISBN 978-0-903317-44-3 .
- ^ Марш, OC (1878). «Основные персонажи американских динозавров Американского юрского периода. Часть I» Американский журнал науки и искусства . 16 (95): 411–416. DOI : 10.2475/AJS.S3-16.95.411 . HDL : 2027/Hvd.32044107172876 . . 219245525 .
- ^ Майкл П. Тейлор; Мэтью Дж. Ведель (2013). «Почему у Сауроподс были длинные шеи; и почему у жирафов есть короткие шеи» . ПЕРЕЙ . 1 : E36. doi : 10.7717/peerj.36 . PMC 3628838 . PMID 23638372 .
Шеи динозавров Сауропода были самыми длинными из всех животных ...
- ^ Tidwell, V., Carpenter, K. & Meyer, S. 2001. Новая титанозавриформа (Sauropoda) от члена ядовитой полосы формирования Cedar Mountain (нижние меловые), Юта. В: Жизнь мезозойских позвоночных. DH Tanke & K. Carpenter (Eds.). Издательство Университета Индианы, ред. DH Tanke & K. Carpenter. Издательство Университета Индианы. 139-165.
- ^ Баккер, Роберт (1994). «Укус Бронто». Земля . 3 (6): 26–33.
- ^ Петерсон, Иварс (март 2000 г.). «Кнуты и хвосты динозавров» . Science News. Архивировано из оригинала 2007-07-14 . Получено 2007-07-07 .
- ^ Myhrvold, Nathan P.; Керри, Филипп Джон (1997). «Суперзвуковые ? сауропод Палеобиология . 23 (4): 393–409. Bibcode : 1997pbio ... 23..393m . doi : 10.1017/s0094837300019801 . ISSN 0094-8373 . S2CID 83696153 .
- ^ Lovelace, David M.; Хартман, Скотт А.; Wahl, William R. (2007). «Морфология образца суперсероура (Dinosauria, Sauropoda) из образования Моррисона Вайоминга и переоценка диплодоцидной филогения». Arquivos Do Museu Nacional . 65 (4): 527–544.
- ^ Jump up to: а беременный Карпентер, К. (2006). «Самая большая из больших: критическая переоценка мега-сауропод -амфикоэлиас fragillimus ». Музей естественной истории и науки Нью -Мексико . 36 : 131–138. S2CID 56215581 .
- ^ Jump up to: а беременный Кэри Вудрафф и Джон Р. Фостер (15 июля 2015 г.). «Хрупкое наследие Amphicoelias fragillimus (Dinosauria: Sauropoda; Morrison Formation - последняя юрская игра)» (PDF) . Peerj Preprints . 3 : E1037.
- ^ Jump up to: а беременный в дюймовый и фон глин Molina-Perez & Larramendi (2020). Факты и цифры динозавра: сауроподы и другие сауроподоморфы . Нью -Джерси: издательство Принстонского университета. С. 42–267. Bibcode : 2020dffs.book ..... m .
- ^ Jump up to: а беременный в дюймовый и фон Пол, Грегори С. (2019). «Определение крупнейшего известного землевладельца: критическое сравнение различных методов восстановления объема и массы вымерших животных» (PDF) . Анналы музея Карнеги . 85 (4): 335–358. doi : 10.2992/007.085.0403 . S2CID 210840060 .
- ^ Jump up to: а беременный Тейлор, Майк (2019). «Supersaurus, UltraSaurus и Dystylosaurus в 2019 году, часть 2B: размер животного BYU 9024» .
- ^ Природа охраняемость. «Африканский слон Буша» .
{{cite journal}}: CITE Journal требует|journal=( помощь ) - ^ Мацетта, Гв; Christiansen, P.; Фаринья, Р.А. (2004). «Гиганты и странные: размер тела некоторых южноамериканских меловых динозавров» (PDF) . Историческая биология . 16 (2–4): 71–83. Bibcode : 2004hbio ... 16 ... 71m . Citeseerx 10.1.1.694.1650 . doi : 10.1080/08912960410001715132 . S2CID 56028251 . Получено 7 июля 2024 года .
- ^ Приятель, Саурабх; Айясами, Кришнан (27 июня 2022 года). «Потерянный Титан Кавери». Геология сегодня . 38 (3): 112–116. Bibcode : 2022geolt..38..112p . doi : 10.1111/gto.12390 . ISSN 0266-6979 . S2CID 250056201 .
- ^ Пол, Грегори С.; Larramendi, Asier (11 апреля 2023 г.). «Оценка массы тела Bruhathkayosaurus и других фрагментарных остатков Sauropod предполагает, что крупнейшие земельные животные были примерно такими же большими, как величайшие киты». Летая . 56 (2): 1–11. Bibcode : 2023Letha..56..2.5p . doi : 10.18261/let.56.2.5 . ISSN 0024-1164 . S2CID 259782734 .
- ^ Jump up to: а беременный Отеро, Алехандро; Carballdo, José L.; Салази, Леонардо; Canudo, Хосе Игнасио; Гарридо, Альберто С. (12 января 2021 г.). «Отчет о гигантской титосуре является частью Neuquance, Аргентина » креетей Исследование 122 : 104754. Bibcode : 2021crr . doi : 10.1016/j.creator . S2CID 23358290 .
- ^ Larramendi, A. (2015). «Высота плеча, масса тела и форма пробидейцев» (PDF) . Acta Palaeontologica Polonica . 60 doi : 10.4202/app.00136.2014 . S2CID 2092950 .
- ^ Jump up to: а беременный Мартин Сандер, П.; Матеус, Октавио; Лавен, Томас; Knötschke, Nils (2006). «Гистология кости указывает на островный карлизм в новой поздней юрской динозавре Сауропод». Природа . 441 (7094): 739–41. Bibcode : 2006natur.441..739m . doi : 10.1038/nature04633 . PMID 16760975 . S2CID 4361820 .
- ^ Николь Кляйн (2011). Биология динозавров Сауропода: понимание жизни гигантов . Издательство Университета Индианы. п. 73. ISBN 978-0-253-35508-9 .
- ^ Рейтнер, Йоахим; Ян, Кун; Ван, Юндонг; Рейх, Майк (6 сентября 2013 г.). Палеобиология и геобиология ископаемого лагрстента через историю Земли: совместная конференция «палеонтологического общества» и «Палеонтологическое общество Китая», Геттинген, Германия, 23-27 сентября 2013 года . Университетский издатель Геттинген. п. 21. ISBN 978-3-86395-135-1 .
- ^ Бейкер, Гарри (2021). «Массивный новый динозавр может быть крупнейшим существом, когда -либо бродившим землю» . LivesCience.com . Получено 22 января 2021 года .
- ^ Палаццо, Чиара (28 марта 2017 г.). «Самые большие в мире следы динозавров, найденные в« Парке Юрского периода Австралии » . Телеграф .
- ^ Bonnan, MF 2005. Анатомия PES у динозавров Sauropod: последствия для функциональной морфологии, эволюции и филогения; С. 346-380 в К. Карпентере и В. Тидуэлл (ред.), Громовые ящерицы: динозавры Sauropodomorph. Блумингтон, в: Издательство Индианы Университет.
- ^ Jump up to: а беременный Боннан, Мэтью Ф. (2003). «Эволюция формы Manus у динозавров Sauropod: последствия для функциональной морфологии, ориентации передней передней части и филогения» (PDF) . Журнал палеонтологии позвоночных . 23 (3): 595–613. Bibcode : 2003jvpal..23..595b . doi : 10.1671/a1108 . S2CID 85667519 .
- ^ Upchurch, P. (1994). «Функция когтя Manus в динозаврах Sauropod». Гайя . 10 : 161–171.
- ^ Jump up to: а беременный Apeesteguía, S. (2005). «Эволюция титанозавра метакарпуса». Стр. Eds.) Carpenter, K. ( 321-345 в Tidwell, V. и Индианаполис: издательство Университета Индианы.
- ^ Милан, Дж.; Christiansen, P.; Матеус, О. (2005). «Трехмерное сохраненное впечатление Sauropod Manus от верхней юры Португалии: последствия для формы Sauropod Manus и механики локомотора». Каупия . 14 : 47–52.
- ^ Bonnan, MF (2004). «Морфометрический анализ плечевой кости и формы бедра у Моррисона Сауропод: последствия для функциональной морфологии и палеобиологии» (PDF) . Палеобиология . 30 (3): 444–470. Bibcode : 2004pbio ... 30..444b . doi : 10.1666/0094-8373 (2004) 030 <0444: Maohaf> 2,0.co; 2 . JSTOR 4096900 . S2CID 86258781 .
- ^ Боннан, Мэтью Ф. (2007). «Линейный и геометрический морфометрический анализ моделей масштабирования длинных костей у юрских динозавров неозауропода: их функциональные и палеобиологические последствия» . Анатомическая запись . 290 (9): 1089–1111. doi : 10.1002/ar.20578 . PMID 17721981 . S2CID 41222371 .
- ^ Ведель, MJ (2009). « Свидетельство о птицах, похожих на воздушные мешочки у сауришских динозавров ». (PDF) Журнал экспериментальной зоологии , 311a : 18pp.
- ^ Jump up to: а беременный в Тейлор, МП; Ведель, MJ (2013). «Почему у Сауроподс были длинные шеи; и почему у жирафов есть короткие шеи» . ПЕРЕЙ . 1 : E36. doi : 10.7717/peerj.36 . PMC 3628838 . PMID 23638372 .
- ^ Уорд, Питер Дуглас (2006). «Юра: гегемония динозавров в мире низкого оксигена». Из воздуха: динозавры, птицы и древняя атмосфера Земли . Вашингтон, округ Колумбия: Джозеф Генри Пресс. С. 199–222. ISBN 0-309-10061-5 .
- ^ Jump up to: а беременный в дюймовый и Тейлор, MP (2010). «Исследование динозавров Sauropod: исторический обзор». В Ричарде Муди, Эрике Баффетауте, Дэвиде М. Мартилле и Даррене Найш (ред.), Динозаврах (и других вымерших саурианов): историческая перспектива . HTML Abstract .
- ^ Jump up to: а беременный D'Emic, Michael D.; Уитлок, Джон А.; Смит, Кэтлин М.; Фишер, Даниэль С.; Уилсон, Джеффри А. (17 июля 2013 г.). «Эволюция высоких скоростей замены зубов у динозавров Sauropod» . Plos один . 8 (7): E69235. BIBCODE : 2013PLOSO ... 869235D . doi : 10.1371/journal.pone.0069235 . PMC 3714237 . PMID 23874921 .
- ^ Jump up to: а беременный Барбер, Элизабет (2004-06-09). «Зубная щетка не требуется: динозавры заменяли свою улыбку каждый месяц» . Христианский научный монитор . Получено 2013-08-14 .
- ^ Стивенс, Ка (2013). «Артикуляция шеей Sauropod: методология и мифология» . Plos один . 8 (10): E78572. BIBCODE : 2013PLOSO ... 878572S . doi : 10.1371/journal.pone.0078572 . PMC 3812995 . PMID 24205266 .
- ^ Jump up to: а беременный в дюймовый и фон глин час Сандер, П. Мартин; Кристиан, Андреас; Клаусс, Маркус; Фехнер, Регина; Джи, Кэрол Т.; Грибелер, Ева-Мария; Гунга, Ханнс-христиан; Хаммел, Юрген; Маллисон, Генрих; Перри, Стивен Ф.; и др. (2011). «Биология динозавров Сауропода: эволюция гигантизма» . Биологические обзоры . 86 (1): 117–155. doi : 10.1111/j.1469-185x.2010.00137.x . ISSN 1464-7931 . PMC 3045712 . PMID 21251189 .
- ^ Хендерсон, DM (2013). "Sauropod Necks: действительно ли они потери тепла?" Полем Plos один . 8 (10): E77108. BIBCODE : 2013PLOSO ... 877108H . doi : 10.1371/journal.pone.0077108 . PMC 3812985 . PMID 24204747 .
- ^ Jump up to: а беременный в дюймовый и Хендерсон, Д.М. (2004). «Пятнистые игроки: пневматичность динозавра Sauropod, плавучесть и водные привычки» . Труды Королевского общества Лондона б . 271 (Suppl 4): S180 - S183. doi : 10.1098/rsbl.2003.0136 . PMC 1810024 . PMID 15252977 .
- ^ Кермак, Ка (1951). «Примечание о привычках сауропод». Энн. Маг НАТ Хвост 4 (44): 830–832. doi : 10.1080/00222935108654213 .
- ^ «Гигантское оборудование Sauropod Discovery, сделанное на острове Скай (Wired UK)» . Wired UK . 2015-12-02 . Получено 2016-03-22 .
- ^ «Динозавры Sauropod были ограничены более теплыми областями земли» .
- ^ Wyenberg-Henzler T. 2022. Ecomorphospace оккупация крупных травоядных динозавров от позднего юрского периода до позднего мелового времени в Северной Америке. Peerj 10: E13174 https://doi.org/10.7717/peerj.13174
- ^ Jump up to: а беременный в дюймовый Майерс, Т.С.; Fiorillo, AR (2009). «Доказательства общительного поведения и возрастной сегрегации у динозавров Sauropod» (PDF) . Палеогеография, палеоклиматология, палеоэкология . 274 (1–2): 96–104. Bibcode : 2009ppp ... 274 ... 96M . doi : 10.1016/j.palaeo.2009.01.002 .
- ^ Coria, RA (1994). «О моноспецифической сборке динозавров Sauropod из Патагонии: значение для общительного поведения». Гайя . 10 : 209–213.
- ^ Ruxton, Graeme D.; Берчард, Джеффри Ф.; Deeming, D. Charles (2014). «Время инкубации как важное влияние на производство яиц и распределение в сцеплениях для динозавров Sauropod». Палеобиология . 40 (3): 323–330. Bibcode : 2014pbio ... 40..323r . doi : 10.1666/13028 . S2CID 84437615 .
- ^ Осборн, HF (1899). «Скелет Diplodocus, недавно установленный в американском музее» . Наука . 10 (259): 870–4. Bibcode : 1899sci .... 10..870f . doi : 10.1126/science.10.259.870 . PMID 17788971 .
- ^ Jump up to: а беременный Rothschild, BM & Molnar, RE (2005). «Стресс -переломы Sauropod как подсказки для активности». В Carpenter, K. & Tidswell, V. (Eds.). Громовые ящерицы: динозавры Sauropodomorph . Издательство Университета Индианы. С. 381–391. ISBN 978-0-253-34542-4 .
- ^ Jump up to: а беременный Mallison, H. (2009). «Воспитание для пищи? Кинетическое/динамическое моделирование двуночных/тройных позов в динозаврах Sauropod». С. 63 В Godefroit, P. и Lambert, O. (eds), дань уважения Чарльзу Дарвину и Берниссарту Игуанодонам: новые перспективы на эволюцию позвоночных и ранние меловые экосистемы . Брюссель.
- ^ Bujor, Mara (2009-05-29). "Сауроподы ходили с шеей в вертикальном положении?" Полем ZME Science.
- ^ Сеймур, RS (июнь 2009 г.). «Повышение шеи Sauropod: это стоит дороже, чтобы получить меньше» . Биол. Летал 5 (3): 317–9. doi : 10.1098/rsbl.2009.0096 . PMC 2679936 . PMID 19364714 .
- ^ Choy, DS; Альтман, П (1992-08-29). «Сердечно -сосудистая система Barosaurus: образованное предположение» . Лансет . 340 (8818): 534–6. doi : 10.1016/0140-6736 (92) 91722-K . PMID 1354287 . S2CID 7378155 .
- ^ Seymour, Rs; Lillywhite, HB (сентябрь 2000 г.). «Сердца, поза шеи и интенсивность метаболической динозавров Sauropod» . Прокурор Биол. Наука 267 (1455): 1883–7. doi : 10.1098/rspb.2000.1225 . PMC 1690760 . PMID 11052540 .
- ^ Jump up to: а беременный Стивенс, Ка; Parrish, JM (1999). «Привычки позы и кормления двух юрских динозавров юрского периода». Наука . 284 (5415): 798–800. Bibcode : 1999sci ... 284..798s . doi : 10.1126/science.284.5415.798 . PMID 10221910 .
- ^ Taylor, MP, Wedel, MJ и Naish, D. (2009). « Поза головы и шеи в динозаврах Sauropod выводятся из существующих животных ». Acta palaeontologica Polonica 54 (2), 2009: 213-220 Аннотация
- ^ Музеи и телевидение имеют позы динозавров, все неправильно, утверждают ученых . Guardian, 27 мая 2009 г.
- ^ Cobley MJ; Рэйфилд EJ; Барретт премьер -министр (14 августа 2013 г.). «Межвисполенная гибкость шеи страуса: последствия для оценки гибкости шеи Sauropod» . Plos один . 8 (8): E72187. BIBCODE : 2013PLOSO ... 872187C . doi : 10.1371/journal.pone.0072187 . PMC 3743800 . PMID 23967284 .
- ^ ND (2013-08-14). «Шеи страуса показывают движения Sauropod, пищевые привычки» . Наука ежедневно . Получено 2013-08-16 .
- ^ Ghose, Tia (2013-08-15). «Ой! Динозавры с длинной шеей, возможно, на самом деле имели жесткие шеи» . NBC News Live Science . Получено 2013-08-16 .
- ^ Jump up to: а беременный Рига, BJG; Calvo, Jo (2009). «Новый широкий сайт Sauropod Sauropod от покойного мела Мендосы, бассейн Neuquen, Аргентина» (PDF) . Палеонтология . 52 (3): 631–640. Bibcode : 2009Palgy..52..631G . doi : 10.1111/j.1475-4983.2009.00869.x .
- ^ Castanera, D.; Barco, JL; Смерть Мартина, я.; Гаскон, JSH; Lorent Perez, FL; Canon, Ji (2011). Берриасская группа Палеоикатология, палеоэкология . 310 (3–4): 227–237. BIBCODE : 2011PPP.310..227C doi : 10.1016/j.palaeo .
- ^ День, JJ; Норман, дБ; Гейл, как; Upchurch, P.; Пауэлл, HP (2004). «Средний участок динозавров в среднем юрском периоде от Оксфордшира, Великобритания» . Палеонтология . 47 (2): 319–348. Bibcode : 2004Palgy..47..319d . doi : 10.1111/j.0031-0239.2004.00366.x .
- ^ Фалкингем, PL; Бейтс, KT; Margetts, L.; Мэннинг, PL (2011-02-23). «Моделирование формирования трека только Sauropod Manus с использованием анализа конечных элементов» . Биологические письма . 7 (1): 142–145. doi : 10.1098/rsbl.2010.0403 . ISSN 1744-9561 . PMC 3030862 . PMID 20591856 .
- ^ Фалкингем, PL; Бейтс, KT; Margetts, L.; Мэннинг, PL (2011-08-07). «Эффект« Златовласка »: смещение консервации в скобках дорожек позвоночных» . Журнал интерфейса Королевского общества . 8 (61): 1142–1154. doi : 10.1098/rsif.2010.0634 . ISSN 1742-5689 . PMC 3119880 . PMID 21233145 .
- ^ Jump up to: а беременный Продавцы, WI; Margetts, L.; Coria, Rab; Мэннинг, PL (2013). Перевозчик, Дэвид (ред.). «Март Титанов: локомоторные возможности динозавров Sauropod» . Plos один . 8 (10): E78733. BIBCODE : 2013PLOSO ... 878733S . doi : 10.1371/journal.pone.0078733 . PMC 3864407 . PMID 24348896 .
- ^ Сабо, Джон (2011). Динозавры . Университет Акрона: МакГроу Хилл. п. 35. ISBN 978-1-121-09332-4 .
- ^ Брайан К. Холл; Бенедикт Халгримссон (1 июня 2011 г.). Эволюция Стрикбергера . Jones & Bartlett Publishers. п. 446. ISBN 978-1-4496-4722-3 .
- ^ Наварре, Бруно А.; Гилларди, Алин М.; Аурелиано, Тито; День, День Вероники; Группа, Камила Лн; Каттарус, Эндрю Г.С.; Иори, Фабиан В.; Мартин, Ариэль М.; Carvalo, B.; Anelli, Luiz E.; Фернандес, Марсело А.; Захер, Хуссам (2022-09-15). «Верхний создание Бразилии» . Амегини 59 (5). doi : 10 5710/amgh.25.08.2022 . ISSN 0002-7014 . 251875979S2CID
- ^ «Аху! Респираторная болезнь дала молодой« Долли »симптомы динозавров гриппа» . Живая наука . 10 февраля 2022 года.
- ^ «Обнаружение того, что Ийд Долли Динозавр является первым, говорят исследователи» . CNN . 10 февраля 2022 года.
- ^ Тан, Хао ; ::::::::::::::::: ::::::::::::::::: ::::::::::::::::: :::::::::::::: Динозавры из среднего юрского периода Юньяна, Чунцин, юго -западный Китай » . Историческая биология . 35 (4): 475 –482 .
- ^ Аурелиано, Тито; Насименто, Каролина Си; Фернандес, Марсело А.; Рикарди-Бранко, Фресия; Гиларди, Алин М. (2021-02-01). «Паразиты крови и острый остеомиелит у неавийского динозавра (Sauropoda, Титанозаврия) из верхней меловой формирования Адаманты, бассейн Бауру, юго-восточная Бразилия» . Мерашные исследования . 118 : 104672. Bibcode : 2021crres.11804672a . doi : 10.1016/j.cretres.2020.104672 . ISSN 0195-6671 . S2CID 225134198 .
- ^ Аурелиано, Тито; Насименто, Каролина Си; Фернандес, Марсело А.; Рикарди-Бранко, Фресия; Гиларди, Алин М. (февраль 2021 г.). «Паразиты крови и острый остеомиелит у неавийского динозавра (Sauropoda, Титанозаврия) из верхней меловой формирования Адаманты, бассейн Бауру, юго-восточная Бразилия». Мерашные исследования . 118 : 104672. Bibcode : 2021crres.11804672a . doi : 10.1016/j.cretres.2020.104672 . S2CID 225134198 .
- ^ Бараниюк, Крис (январь 2021 г.). «Ужасные» кровавые черви вторглись в кость ноги динозавра, предлагает ископаемое » . Scientific American .
- ^ «Цивяшник титанозавра страдал от паразитов крови и тяжелого воспаления костей | Палеонтология | sci-news.com» . Последние научные новости | Sci-news.com .
- ^ Delair, JB; Сарджант, был (2002). «Самые ранние открытия динозавров: записи повторно проведены». Материалы Ассоциации геологов . 113 (3): 185–197. Bibcode : 2002prga..113..185d . doi : 10.1016/s0016-7878 (02) 80022-0 .
- ^ Lhuyd, E. (1699). Lithophylacii British Ichnographia, или странная мерцание или другие минуты британского ископаемого . Гледц и Вейдманн, Лондон.
- ^ Оуэн Р. (1842). «Отчет о британских ископаемых рептилиях». Часть II. Отчет Британской ассоциации по развитию науки, Плимут, Англия.
- ^ Phillips, J. (1871). Геология Оксфорда и долины Темзы. Оксфорд: Clarendon Press, 523 стр.
- ^ Осборн, HF; Мук, CC (1919-01-01). «Камаразавр, амфикоэлия и другие сауроподы Cope» . Геологическое общество Америки Бюллетень . 30 (1): 379–388. doi : 10.1130/GSAB-30-379 . ISSN 0016-7606 .
- ^ Sagado, L.; Кория, Ра; Calvo, Jo (1997). Полем Амегина 34 (1): 3–3 ISSN 1851-8
- ^ Peyre de Fabrègues, C.; Allain, R.; Барриэль, В. (2015). «Корневые причины филогенетического несоответствия, наблюдаемые в базальных взаимосвязаниях Sauropodomorph: Sauropodomorph взаимосвязи» . Зоологический журнал Линневого общества . 175 (3): 569–586. doi : 10.1111/Zoj.12290 . S2CID 83180197 .
- ^ Йейтс, А.М. (2007). «Решение динозаврской головоломки: личность Аливалии Рекс Гальтон». Историческая биология . 19 (1): 93–123. Bibcode : 2007hbio ... 19 ... 93y . doi : 10.1080/08912960600866953 . S2CID 85202575 .
- ^ Allain, R.; Aquesbi, N. (2008). «Анатомия и филогенетические отношения тазудасурсауруса Найми (Dinosauria, Sauropoda) из раннего юрского периода Марокко » GeoDiversitass 30 (2): 345–4
- ^ Пол, Д.; Otero, A.; Apaldetti, C.; Мартинес, RN (2021). «Триасы Сауроподоморф Динозавров из Южной Америки: происхождение и диверсификация динозавров, доминирующих травоядными фаунами». Журнал южноамериканских наук о Земле . 107 : 103145. Bibcode : 2021jsaes.10703145p . doi : 10.1016/j.jsames.2020.103145 . S2CID 233579282 .
- ^ Jump up to: а беременный Allain, R. and Aquesbi, N. (2008). «Анатомия и филогенетические отношения Тазудасувра Найми (Dinosauria, Sauropoda) из раннего юрского периода Марокко. GeoDiversitass , 30 (2): 345-4
Смотрите также
[ редактировать ]- Штраус, Боб (2008). «Сауроподы: самые большие динозавры, которые когда -либо жили» . ОБЛЮДА . Типы динозавров.
- Роджерс, KC ; Уилсон, JA (2005). Сауроподы: эволюция и палеобиология . Беркли, Калифорния: Университет Калифорнийской прессы . ISBN 0-520-24623-3 .
- Upchurch, P.; Барретт, премьер -министр; Додсон, П. (2004). "Сауропода". В Weishampel, D . ; Додсон, П.; Osmólska, H. (Eds.). Динозаврия (2 -е изд.). Беркли, Калифорния: Университет Калифорнийской прессы . С. 259–322.











