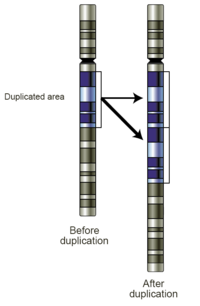
Дупликация генов
Дупликация генов (или хромосомная дупликация , или амплификация генов ) является основным механизмом, посредством которого создается новый генетический материал в ходе молекулярной эволюции . Его можно определить как любое дублирование участка ДНК , содержащего ген . Дупликация генов может возникать в результате нескольких типов ошибок в ДНК механизмах репликации и восстановления , а также в результате случайного захвата эгоистичными генетическими элементами. Общие источники дупликации генов включают эктопическую рекомбинацию , ретротранспозицию , анеуплоидию , полиплоидию и проскальзывание репликации . [1]
Механизмы дублирования
[ редактировать ]Эктопическая рекомбинация
[ редактировать ]Дупликации возникают в результате явления, называемого неравным кроссинговером , которое происходит во время мейоза между смещенными гомологичными хромосомами. Вероятность того, что это произойдет, зависит от степени совместного использования повторяющихся элементов между двумя хромосомами. Продуктами этой рекомбинации являются дупликация в месте обмена и реципрокная делеция. Эктопическая рекомбинация обычно опосредуется сходством последовательностей в дублирующих точках разрыва, которые образуют прямые повторы. Повторяющиеся генетические элементы, такие как мобильные элементы, представляют собой один из источников повторяющейся ДНК, которая может способствовать рекомбинации, и они часто обнаруживаются в точках останова дупликации у растений и млекопитающих. [2]
Проскальзывание репликации
[ редактировать ]Проскальзывание репликации — это ошибка репликации ДНК, которая может привести к дублированию коротких генетических последовательностей. Во время репликации ДНК-полимераза начинает копировать ДНК. В какой-то момент процесса репликации полимераза отделяется от ДНК, и репликация останавливается. Когда полимераза повторно прикрепляется к цепи ДНК, она выравнивает реплицирующуюся цепь в неправильном положении и случайно копирует один и тот же участок более одного раза. Проскальзыванию репликации также часто способствуют повторяющиеся последовательности, но для этого требуется лишь несколько оснований сходства. [ нужна ссылка ]
Ретротранспозиция
[ редактировать ]Ретротранспозоны , главным образом L1 , иногда могут действовать на клеточную мРНК. Транскрипты обратно транскрибируются в ДНК и вставляются в случайное место генома, создавая ретрогены. Полученная последовательность обычно лишена интронов и часто содержит поли(А)-последовательности, которые также интегрированы в геном. Многие ретрогены демонстрируют изменения в регуляции генов по сравнению с последовательностями их родительских генов, что иногда приводит к появлению новых функций. Ретрогены могут перемещаться между разными хромосомами, формируя хромосомную эволюцию. [3]
Анеуплоидия
[ редактировать ]Анеуплоидия возникает, когда нерасхождение одной хромосомы приводит к аномальному количеству хромосом. Анеуплоидия часто бывает вредной и у млекопитающих регулярно приводит к самопроизвольным абортам (выкидышам). Некоторые анеуплоидные особи жизнеспособны, например трисомия 21 у человека, что приводит к синдрому Дауна . Анеуплоидия часто изменяет дозировку генов губительно для организма; поэтому маловероятно, что он распространится среди населения.
Полиплоидия
[ редактировать ]Полиплоидия , или дупликация всего генома, является продуктом нерасхождения во время мейоза, что приводит к образованию дополнительных копий всего генома. Полиплоидия распространена у растений, но она также наблюдалась у животных: два раунда дупликации всего генома ( событие 2R ) в линии позвоночных привели к появлению человека. [4] Это также произошло у дрожжей-гемиаскомицетов ~ 100 млн лет назад. [5] [6]
После дупликации всего генома наступает относительно короткий период нестабильности генома, обширной потери генов, повышенного уровня нуклеотидных замен и перестройки регуляторной сети. [7] [8] Кроме того, значительную роль играют эффекты дозировки генов. [9] Таким образом, большинство дубликатов теряются в течение короткого периода времени, однако значительная часть дубликатов выживает. [10] Интересно, что гены, участвующие в регуляции, преимущественно сохраняются. [11] [12] Более того, сохранение регуляторных генов, особенно Hox-генов , привело к адаптивным инновациям.
Быстрая эволюция и функциональная дивергенция наблюдались на уровне транскрипции дуплицированных генов, обычно за счет точечных мутаций в коротких мотивах связывания транскрипционных факторов. [13] [14] Более того, быстрая эволюция мотивов фосфорилирования белков, обычно встроенных в быстро развивающиеся внутренне неупорядоченные области, является еще одним фактором, способствующим выживанию и быстрой адаптации/неофункционализации дублированных генов. [15] Таким образом, по-видимому, существует связь между регуляцией генов (по крайней мере, на посттрансляционном уровне) и эволюцией генома. [15]
Полиплоидия также является хорошо известным источником видообразования, поскольку потомство, имеющее разное количество хромосом по сравнению с родительскими видами, часто не может скрещиваться с неполиплоидными организмами. Считается, что дупликация всего генома менее вредна, чем анеуплоидия, поскольку относительная дозировка отдельных генов должна быть одинаковой.
Как эволюционное событие
[ редактировать ]
Скорость дупликации генов
[ редактировать ]Сравнение геномов показывает, что дупликация генов распространена у большинства исследованных видов. На это указывают переменные числа копий ( вариации числа копий ) в геноме человека. [16] [17] или плодовые мухи. [18] Однако было трудно измерить скорость, с которой происходит такое дублирование. Недавние исследования дали первую прямую оценку полногеномной скорости дупликации генов у C. elegans , первого многоклеточного эукариота, для которого такая оценка стала доступной. Скорость дупликации генов у C. elegans составляет порядка 10. −7 дупликации/ген/поколение, то есть в популяции из 10 миллионов червей будет дупликация гена на каждое поколение. Эта скорость на два порядка превышает спонтанную скорость точковых мутаций на нуклеотидный сайт у этого вида. [19] Более ранние (косвенные) исследования показали, что частота локус-специфических дупликаций у бактерий, дрозофилы и человека варьируется от 10 −3 до 10 −7 /ген/поколение. [20] [21] [22]
Неофункционализация
[ редактировать ]Дупликация генов является важным источником генетической новизны, которая может привести к эволюционным инновациям. Дупликация создает генетическую избыточность, при которой вторая копия гена часто свободна от селективного давления , то есть мутации ее не оказывают вредного воздействия на организм-хозяин. Если в одной копии гена происходит мутация, которая влияет на его первоначальную функцию, вторая копия может служить «запасной частью» и продолжать функционировать правильно. Таким образом, дублирующиеся гены накапливают мутации быстрее, чем функциональный ген с одной копией, на протяжении поколений организмов, и одна из двух копий может развить новую и другую функцию. Некоторыми примерами такой неофункционализации являются очевидная мутация дублированного пищеварительного гена в семействе ледяных рыб в ген антифриза и дупликация, приводящая к появлению нового гена змеиного яда. [23] и синтез 1-бета-гидрокситестостерона у свиней. [24]
Считается, что дупликация генов играет важную роль в эволюции ; этой позиции придерживаются члены научного сообщества уже более 100 лет. [25] Сусуму Оно был одним из самых известных разработчиков этой теории в своей классической книге «Эволюция путем дупликации генов» (1970). [26] Оно утверждал, что дупликация генов является наиболее важной эволюционной силой с момента появления универсального общего предка . [27] Крупные случаи дупликации генома могут быть довольно частыми. Считается, что весь дрожжей геном подвергся дупликации около 100 миллионов лет назад. [28] Растения являются наиболее плодовитыми дупликаторами генома. Например, пшеница является гексаплоидной (разновидность полиплоида ), то есть имеет шесть копий своего генома.
Субфункциональность
[ редактировать ]Другая возможная судьба дублирующих генов заключается в том, что обе копии в равной степени могут накапливать дегенеративные мутации, пока любые дефекты дополняются другой копией. Это приводит к нейтральной модели « субфункционализации » (процессу конструктивной нейтральной эволюции ) или модели DDC (дупликация-дегенерация-дополнение), [29] [30] при котором функциональность исходного гена распределяется между двумя копиями. Ни один ген не может быть утерян, поскольку оба теперь выполняют важные недублированные функции, но в конечном итоге ни один из них не способен достичь новой функциональности.
Субфункционализация может происходить посредством нейтральных процессов, при которых мутации накапливаются без каких-либо вредных или полезных эффектов. Однако в некоторых случаях субфункционализация может иметь место с явными адаптивными преимуществами. Если предковый ген плейотропен и выполняет две функции, часто ни одну из этих двух функций нельзя изменить, не затрагивая другую функцию. Таким образом, разделение наследственных функций на два отдельных гена может обеспечить адаптивную специализацию подфункций, тем самым обеспечивая адаптивное преимущество. [31]
Потеря
[ редактировать ]Часто возникающие в результате геномные вариации приводят к неврологическим расстройствам, зависящим от дозы гена, таким как синдром Ретта и болезнь Пелицеуса-Мерцбахера . [32] Такие вредные мутации, скорее всего, будут потеряны из популяции, не сохранятся и не разовьют новые функции. Однако многие дупликации на самом деле не являются ни вредными, ни полезными, и эти нейтральные последовательности могут быть потеряны или могут распространиться среди популяции посредством случайных колебаний посредством генетического дрейфа .
Выявление дупликаций в секвенированных геномах
[ редактировать ]Критерии и сканирование одного генома
[ редактировать ]Два гена, которые существуют после события дупликации гена, называются паралогами и обычно кодируют белки со схожей функцией и/или структурой. Напротив, ортологичные гены присутствуют у разных видов, каждый из которых изначально произошел от одной и той же предковой последовательности. (См. Гомология последовательностей в генетике ).
В биологических исследованиях важно (но часто сложно) различать паралоги и ортологи. Эксперименты по функции генов человека часто можно проводить на других видах, если в геноме этого вида можно найти гомолог человеческого гена, но только если гомолог ортологичен. Если они являются паралогами и возникли в результате дупликации генов, их функции, вероятно, будут слишком разными. Одна или несколько копий дуплицированных генов, составляющих семейство генов, могут быть затронуты вставкой мобильных элементов , что вызывает значительные различия между ними в их последовательностях и, в конечном итоге, может стать причиной дивергентной эволюции . Это также может влиять на вероятность и скорость генной конверсии между гомологами дубликатов генов из-за меньшего сходства или отсутствия сходства в их последовательностях.
Паралоги можно идентифицировать в отдельных геномах путем сравнения последовательностей всех аннотированных моделей генов друг с другом. Такое сравнение может быть выполнено на транслированных аминокислотных последовательностях (например, BLASTp, tBLASTx) для идентификации древних дупликаций или на нуклеотидных последовательностях ДНК (например, BLASTn, мегабласт) для идентификации более поздних дупликаций. Большинство исследований по выявлению дупликаций генов требуют взаимных лучших совпадений или нечетких взаимных лучших совпадений, где каждый паралог должен быть единственным лучшим совпадением другого в сравнении последовательностей. [33]
Большинство дупликаций генов существуют в виде малокопийных повторов (LCR), довольно часто повторяющихся последовательностей, таких как мобильные элементы. Чаще всего они обнаруживаются в перицентрономных , субтеломерных и интерстициальных областях хромосомы. Многие LCR из-за своего размера (> 1 КБ), сходства и ориентации очень чувствительны к дупликациям и делециям.
Геномные микрочипы обнаруживают дупликации
[ редактировать ]Такие технологии, как геномные микрочипы , также называемые сравнительной геномной гибридизацией массивов (array CGH), используются для высокопроизводительного обнаружения хромосомных аномалий, таких как микродупликации, из образцов геномной ДНК. В частности, технология ДНК- микрочипов может одновременно отслеживать уровни экспрессии тысяч генов при многих методах лечения или экспериментальных условиях, что значительно облегчает эволюционные исследования регуляции генов после дупликации или видообразования генов . [34] [35]
Секвенирование нового поколения
[ редактировать ]Дупликации генов также можно выявить с помощью платформ секвенирования нового поколения. Самый простой способ выявить дублирования в данных повторного секвенирования генома — это использование считываний парного секвенирования. На тандемные дупликации указывают секвенирование пар прочтений, которые отображаются в аномальной ориентации. Благодаря сочетанию увеличенного охвата последовательностей и аномальной ориентации картирования можно выявить дублирования в данных геномного секвенирования.
Номенклатура
[ редактировать ]
Международная система цитогеномной номенклатуры человека (ISCN) — это международный стандарт хромосом человека номенклатуры , который включает названия полос, символы и сокращенные термины, используемые при описании хромосомных и хромосомных аномалий человека. Сокращения включают dup для дупликаций частей хромосомы. [36] Например, dup(17p12) вызывает болезнь Шарко-Мари-Тута типа 1А. [37]
В качестве усиления
[ редактировать ]Дупликация генов не обязательно представляет собой устойчивое изменение в геноме вида. Фактически, такие изменения часто не сохраняются за пределами первоначального организма-хозяина. С точки зрения молекулярной генетики , амплификация гена одним из многих способов гена сверхэкспрессии является . Генетическая амплификация может происходить искусственно, например, с использованием метода полимеразной цепной реакции для амплификации коротких цепей ДНК in vitro с использованием ферментов , или она может происходить естественным путем, как описано выше. Если это естественная дупликация, она все равно может происходить в соматической клетке , а не в зародышевой клетке (что было бы необходимо для длительных эволюционных изменений).
Роль в раке
[ редактировать ]Дупликации онкогенов являются частой причиной многих видов рака . В таких случаях генетическая дупликация происходит в соматической клетке и затрагивает только геном самой раковой клетки, а не весь организм и тем более любое последующее потомство. Недавняя комплексная классификация на уровне пациентов и количественная оценка движущих событий в когортах TCGA показали, что в среднем на одну опухоль приходится 12 движущих событий, из которых 1,5 являются амплификациями онкогенов. [38]
| Тип рака | Ассоциированный ген усиления | Распространенность усиление по типу рака (в процентах) |
|---|---|---|
| Рак молочной железы | МИК | 20% [39] |
| ЭРББ2 ( HER2 ) | 20% [39] | |
| CCND1 ( Циклин D1 ) | 15–20% [39] | |
| ФГФР1 | 12% [39] | |
| ФГФР2 | 12% [39] | |
| Рак шейки матки | МИК | 25–50% [39] |
| ЭРББ2 | 20% [39] | |
| Колоректальный рак | HRAS | 30% [39] |
| КРАС | 20% [39] | |
| МИБ | 15–20% [39] | |
| Рак пищевода | МИК | 40% [39] |
| CCND1 | 25% [39] | |
| МДМ2 | 13% [39] | |
| Рак желудка | CCNE ( Циклин Е ) | 15% [39] |
| КРАС | 10% [39] | |
| ИЗ | 10% [39] | |
| Глиобластома | ERBB1 ( EGFR ) | 33–50% [39] |
| CDK4 | 15% [39] | |
| Рак головы и шеи | CCND1 | 50% [39] |
| ЭРББ1 | 10% [39] | |
| МИК | 7–10% [39] | |
| Гепатоцеллюлярный рак | CCND1 | 13% [39] |
| Нейробластома | МИКН | 20–25% [39] |
| Рак яичников | МИК | 20–30% [39] |
| ЭРББ2 | 15–30% [39] | |
| АКТ2 | 12% [39] | |
| Саркома | МДМ2 | 10–30% [39] |
| CDK4 | 10% [39] | |
| Мелкоклеточный рак легких | МИК | 15–20% [39] |
Полногеномные дупликации также часто встречаются при раке: они обнаруживаются в 30–36% опухолей наиболее распространенных типов рака. [40] [41] Их точная роль в канцерогенезе неясна, но в некоторых случаях они приводят к потере сегрегации хроматина, что приводит к изменениям конформации хроматина, что, в свою очередь, приводит к онкогенным эпигенетическим и транскрипционным модификациям. [42]
См. также
[ редактировать ]- Сравнительная геномика
- ДбДНВ (2010)
- Опять генное рождение
- Перетасовка экзонов
- Амплификация гена у Paramecium Tetraurelia
- Слияние генов
- Горизонтальный перенос генов
- Геном человека
- Инпараноидальный
- Мобильные генетические элементы
- Молекулярная эволюция
- Псевдоген
- Тандемное дублирование экзонов
- Неравный переход
Ссылки
[ редактировать ]- ^ Чжан Дж (2003). «Эволюция путем дупликации генов: обновленная информация» (PDF) . Тенденции в экологии и эволюции . 18 (6): 292–8. дои : 10.1016/S0169-5347(03)00033-8 .
- ^ «Определение дупликации генов» . Медицинский словарь medterms . МедицинаНет. 19 марта 2012 г. Архивировано из оригинала 6 марта 2014 г. Проверено 1 декабря 2008 г.
- ^ Миллер, Дункан; Чен, Цзяньхай; Лян, Цзянтао; Бетран, Эстер; Лонг, Манюань; Шарахов, Игорь В. (28 мая 2022 г.). «Модели дупликации и экспрессии ретрогенов, сформированные эволюцией половых хромосом у малярийных комаров» . Гены . 13 (6): 968. doi : 10.3390/genes13060968 . ISSN 2073-4425 . ПМЦ 9222922 . ПМИД 35741730 .
- ^ Дехал П., Бур Дж.Л. (октябрь 2005 г.). «Два раунда дупликации всего генома у предков позвоночных» . ПЛОС Биология . 3 (10): е314. дои : 10.1371/journal.pbio.0030314 . ПМК 1197285 . ПМИД 16128622 .
- ^ Вульф, К.Х.; Шилдс, округ Колумбия (12 июня 1997 г.). «Молекулярные доказательства древнего дублирования всего генома дрожжей» . Природа . 387 (6634): 708–713. Бибкод : 1997Natur.387..708W . дои : 10.1038/42711 . ISSN 0028-0836 . ПМИД 9192896 . S2CID 4307263 .
- ^ Келлис, Манолис; Биррен, Брюс В.; Ландер, Эрик С. (08 апреля 2004 г.). «Доказательство и эволюционный анализ древнего дублирования генома дрожжей Saccharomyces cerevisiae» . Природа . 428 (6983): 617–624. Бибкод : 2004Natur.428..617K . дои : 10.1038/nature02424 . ISSN 1476-4687 . ПМИД 15004568 . S2CID 4422074 .
- ^ Отто, Сара П. (2 ноября 2007 г.). «Эволюционные последствия полиплоидии» . Клетка . 131 (3): 452–462. дои : 10.1016/j.cell.2007.10.022 . ISSN 0092-8674 . ПМИД 17981114 . S2CID 10054182 .
- ^ Конант, Гэвин К.; Вулф, Кеннет Х. (апрель 2006 г.). «Функциональное разделение сетей совместной экспрессии дрожжей после дупликации генома» . ПЛОС Биология . 4 (4): е109. дои : 10.1371/journal.pbio.0040109 . ISSN 1545-7885 . ПМЦ 1420641 . ПМИД 16555924 .
- ^ Папп, Балаж; Пал, Чаба; Херст, Лоуренс Д. (10 июля 2003 г.). «Чувствительность к дозировке и эволюция семейств генов дрожжей» . Природа . 424 (6945): 194–197. Бибкод : 2003Natur.424..194P . дои : 10.1038/nature01771 . ISSN 1476-4687 . ПМИД 12853957 . S2CID 4382441 .
- ^ Линч, М.; Конери, Дж. С. (10 ноября 2000 г.). «Эволюционная судьба и последствия дублирования генов» . Наука . 290 (5494): 1151–1155. Бибкод : 2000Sci...290.1151L . дои : 10.1126/science.290.5494.1151 . ISSN 0036-8075 . ПМИД 11073452 .
- ^ Фрилинг, Майкл; Томас, Брайан С. (июль 2006 г.). «Генно-сбалансированные дупликации, такие как тетраплоидия, обеспечивают предсказуемое стремление к увеличению морфологической сложности» . Геномные исследования . 16 (7): 805–814. дои : 10.1101/гр.3681406 . ISSN 1088-9051 . ПМИД 16818725 .
- ^ Дэвис, Джерел К.; Петров, Дмитрий А. (октябрь 2005 г.). «Добавляют ли разные механизмы дупликации в геном схожие гены?» . Тенденции в генетике . 21 (10): 548–551. дои : 10.1016/j.tig.2005.07.008 . ISSN 0168-9525 . ПМИД 16098632 .
- ^ Каснев, Тинеке; Де Бодт, Стефани; Раес, Йерун; Маере, Стивен; Ван де Пер, Ив (2006). «Неслучайное расхождение экспрессии генов после дупликации генов и геномов у цветущего растения Arabidopsis thaliana» . Геномная биология . 7 (2): Р13. дои : 10.1186/gb-2006-7-2-r13 . ISSN 1474-760X . ПМЦ 1431724 . ПМИД 16507168 .
- ^ Ли, Вэнь-Сюн; Ян, Цзин; Гу, Сюнь (ноябрь 2005 г.). «Расхождение экспрессии между дубликатами генов» . Тенденции в генетике . 21 (11): 602–607. дои : 10.1016/j.tig.2005.08.006 . ISSN 0168-9525 . ПМИД 16140417 .
- ^ Перейти обратно: а б Амуциас, Григорис Д.; Он, Ин; Гордон, Джонатан; Моссиалос, Димитрис; Оливер, Стивен Г.; Ван де Пер, Ив (16 февраля 2010 г.). «Посттрансляционная регуляция влияет на судьбу дублированных генов» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (7): 2967–2971. Бибкод : 2010PNAS..107.2967A . дои : 10.1073/pnas.0911603107 . ISSN 1091-6490 . ПМЦ 2840353 . ПМИД 20080574 .
- ^ Себат Дж., Лакшми Б., Троге Дж., Александр Дж., Янг Дж., Лундин П. и др. (июль 2004 г.). «Крупномасштабный полиморфизм числа копий в геноме человека». Наука . 305 (5683): 525–8. Бибкод : 2004Sci...305..525S . дои : 10.1126/science.1098918 . ПМИД 15273396 . S2CID 20357402 .
- ^ Иафрат А.Дж., Феук Л., Ривера М.Н., Листевник М.Л., Донахью П.К., Ци Ю и др. (сентябрь 2004 г.). «Обнаружение крупномасштабных изменений в геноме человека» . Природная генетика . 36 (9): 949–51. дои : 10.1038/ng1416 . PMID 15286789 .
- ^ Эмерсон Дж. Дж., Кардосо-Морейра М., Боревиц Дж. О., Лонг М. (июнь 2008 г.). «Естественный отбор формирует общегеномные закономерности полиморфизма числа копий у Drosophila melanogaster». Наука . 320 (5883): 1629–31. Бибкод : 2008Sci...320.1629E . дои : 10.1126/science.1158078 . ПМИД 18535209 . S2CID 206512885 .
- ^ Липински К.Дж., Фарслоу Дж.К., Фитцпатрик К.А., Линч М., Катью В., Бергторссон Ю. (февраль 2011 г.). «Высокий уровень спонтанной дупликации генов у Caenorhabditis elegans» . Современная биология . 21 (4): 306–10. дои : 10.1016/j.cub.2011.01.026 . ПМК 3056611 . ПМИД 21295484 .
- ^ Андерсон П., Рот Дж. (май 1981 г.). «Спонтанные тандемные генетические дупликации у Salmonella typhimurium возникают в результате неравной рекомбинации между цистронами рРНК (rrn)» . Труды Национальной академии наук Соединенных Штатов Америки . 78 (5): 3113–7. Бибкод : 1981PNAS...78.3113A . дои : 10.1073/pnas.78.5.3113 . ПМК 319510 . ПМИД 6789329 .
- ^ Ватанабэ Ю., Такахаси А., Ито М., Такано-Симидзу Т. (март 2009 г.). «Молекулярный спектр спонтанных мутаций de novo в мужских и женских зародышевых клетках Drosophila melanogaster» . Генетика . 181 (3): 1035–43. дои : 10.1534/genetics.108.093385 . ПМК 2651040 . ПМИД 19114461 .
- ^ Тернер Д.Д., Миретти М., Раджан Д., Фиглер Х., Картер Н.П., Блейни М.Л. и др. (январь 2008 г.). «Уровень зародышевых делеций и дупликаций мейоза de novo, вызывающих ряд геномных нарушений» . Природная генетика . 40 (1): 90–5. дои : 10.1038/ng.2007.40 . ПМЦ 2669897 . ПМИД 18059269 .
- ^ Линч VJ (январь 2007 г.). «Изобретение арсенала: адаптивная эволюция и неофункционализация генов фосфолипазы А2 змеиного яда» . Эволюционная биология BMC . 7 :2. дои : 10.1186/1471-2148-7-2 . ПМЦ 1783844 . ПМИД 17233905 .
- ^ Конант Г.К., Вулф К.Х. (декабрь 2008 г.). «Превращение хобби в работу: как дублированные гены находят новые функции». Обзоры природы. Генетика . 9 (12): 938–50. дои : 10.1038/nrg2482 . ПМИД 19015656 . S2CID 1240225 .
- ^ Тейлор Дж. С., Раес Дж. (2004). «Дупликация и дивергенция: эволюция новых генов и старых идей». Ежегодный обзор генетики . 38 : 615–43. дои : 10.1146/annurev.genet.38.072902.092831 . ПМИД 15568988 .
- ^ Оно, С. (1970). Эволюция путем дупликации генов . Спрингер-Верлаг . ISBN 978-0-04-575015-3 .
- ^ Оно, С. (1967). Половые хромосомы и сцепленные с полом гены . Спрингер-Верлаг. ISBN 978-91-554-5776-1 .
- ^ Келлис М., Биррен Б.В., Ландер Э.С. (апрель 2004 г.). «Доказательство и эволюционный анализ дублирования древнего генома дрожжей Saccharomyces cerevisiae». Природа . 428 (6983): 617–24. Бибкод : 2004Natur.428..617K . дои : 10.1038/nature02424 . ПМИД 15004568 . S2CID 4422074 .
- ^ Форс А, Линч М., Пикетт Ф.Б., Аморес А., Ян Ю.Л., Постлтуэйт Дж. (апрель 1999 г.). «Сохранение дубликатов генов за счет комплементарных дегенеративных мутаций» . Генетика . 151 (4): 1531–45. дои : 10.1093/генетика/151.4.1531 . ПМЦ 1460548 . ПМИД 10101175 .
- ^ Штольцфус А (август 1999 г.). «О возможности конструктивной нейтральной эволюции». Журнал молекулярной эволюции . 49 (2): 169–81. Бибкод : 1999JMolE..49..169S . CiteSeerX 10.1.1.466.5042 . дои : 10.1007/PL00006540 . ПМИД 10441669 . S2CID 1743092 .
- ^ Де Марэ Д.Л., Раушер, доктор медицинских наук (август 2008 г.). «Бегство от адаптивного конфликта после дупликации гена антоцианинового пути». Природа . 454 (7205): 762–5. Бибкод : 2008Natur.454..762D . дои : 10.1038/nature07092 . ПМИД 18594508 . S2CID 418964 .
- ^ Ли Дж.А., Лупски-младший (октябрь 2006 г.). «Геномные перестройки и изменения числа копий генов как причина заболеваний нервной системы» . Нейрон . 52 (1): 103–21. дои : 10.1016/j.neuron.2006.09.027 . ПМИД 17015230 . S2CID 22412305 .
- ^ Хан М.В., Хан М.В., Хан С.Г. (ноябрь 2007 г.). «Эволюция семейства генов в 12 геномах дрозофилы» . ПЛОС Генетика . 3 (11): е197. дои : 10.1371/journal.pgen.0030197 . ПМК 2065885 . ПМИД 17997610 .
- ^ Мао Р., Певснер Дж. (2005). «Использование геномных микрочипов для изучения хромосомных аномалий при умственной отсталости». Обзоры исследований умственной отсталости и нарушений развития . 11 (4): 279–85. дои : 10.1002/mrdd.20082 . ПМИД 16240409 .
- ^ Гу С, Чжан Цз, Хуан В (январь 2005 г.). «Быстрая эволюция экспрессии и регуляторных различий после дупликации генов дрожжей» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (3): 707–12. Бибкод : 2005PNAS..102..707G . дои : 10.1073/pnas.0409186102 . ПМК 545572 . ПМИД 15647348 .
- ^ «Символы ISCN и сокращенные термины» . Институт медицинских исследований Кориелла . Проверено 27 октября 2022 г.
- ^ Кассандра Л. Книффин. «БОЛЕЗНЬ Харко-Мари-Тута, ДЕМИЕЛИНИЗИРУЮЩАЯ, ТИП 1А; ШМТ1А» . ОМИМ . Обновлено: 23 апреля 2014 г.
- ^ Вяткин, Алексей Дмитриевич; Отнюков Данила Владимирович; Леонов Сергей В.; Беликов, Алексей В. (14 января 2022 г.). «Комплексная классификация на уровне пациентов и количественная оценка драйверных событий в когортах TCGA PanCanAtlas» . ПЛОС Генетика . 18 (1): e1009996. дои : 10.1371/journal.pgen.1009996 . ПМЦ 8759692 . ПМИД 35030162 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и Кинцлер К.В., Фогельштейн Б. (2002). Генетическая основа рака человека . МакГроу-Хилл. п. 116. ИСБН 978-0-07-137050-9 .
- ^ Бельски, Крейг М.; Зехир, Ахмет; Пенсон, Александр В.; Донохью, Марк Т.А.; Чатила, Валид; Армения, Джошуа; Чанг, Мэтью Т.; Шрам, Элисон М.; Йонссон, Филип; Бандламуди, Чайтанья; Разави, Педрам; Айер, Гопа; Робсон, Марк Э.; Стадлер, Зофия К.; Шульц, Николаус (2018). «Удвоение генома влияет на эволюцию и прогноз поздних стадий рака» . Природная генетика . 50 (8): 1189–1195. дои : 10.1038/s41588-018-0165-1 . ISSN 1546-1718 . ПМК 6072608 . ПМИД 30013179 .
- ^ Куинтон, Райан Дж.; ДиДомицио, Аманда; Виттория, Марк А.; Котынкова Кристина; Тикас, Карлос Дж.; Патель, Шина; Кога, Юсуке; Вахшурзаде, Жасмин; Эрманс, Николь; Курода, Тарухо С.; Парулекар, Неха; Тейлор, Элисон М.; Мэннинг, Эмити Л.; Кэмпбелл, Джошуа Д.; Ганем, Нил Дж. (2021). «Удвоение всего генома придает опухолевым клеткам уникальную генетическую уязвимость» . Природа . 590 (7846): 492–497. Бибкод : 2021Natur.590..492Q . дои : 10.1038/s41586-020-03133-3 . ISSN 1476-4687 . ПМЦ 7889737 . ПМИД 33505027 .
- ^ Ламбута, Руксандра А.; Нанни, Лука; Лю, Юаньлун; Диас-Мияр, Хуан; Айер, Арвинд; Тавернари, Даниэле; Катанаева Наталья; Чириелло, Джованни; Ориккио, Элиза (15 марта 2023 г.). «Удвоение всего генома приводит к онкогенной потере сегрегации хроматина» . Природа . 615 (7954): 925–933. Бибкод : 2023Natur.615..925L . дои : 10.1038/s41586-023-05794-2 . ISSN 1476-4687 . ПМЦ 10060163 . ПМИД 36922594 .