Прокариоты

Прокариот ə ( / pr oʊ ˈ ) k ær i oʊ t , - , t / реже прокариот пишется [ 1 ] Это одноклеточный организм которого клетка лишена ядра и других мембраносвязанных , органелл . [ 2 ] Слово прокариот происходит от древнегреческого πρό ( pró ) «до» и κάρυον ( káruon ) «орех, ядро». [ 3 ] [ 4 ] В системе двух империй, возникшей на основе работ Эдуарда Шаттона , прокариоты были отнесены к империи Прокариот . [ 5 ] Однако в трехдоменной системе , основанной на молекулярном анализе , прокариоты делятся на два домена : бактерии (ранее эубактерии) и археи (ранее архебактерии). Организмы с ядрами относятся к третьему домену — эукариотам . [ 6 ]
Прокариоты развились раньше эукариотов, и у них отсутствуют ядра, митохондрии и большинство других отдельных органелл , характеризующих эукариотические клетки. Когда-то считалось, что прокариотические клеточные компоненты не заключены в цитоплазме , за исключением внешней клеточной мембраны , но бактериальные микрокомпартменты , которые, как полагают, представляют собой квазиорганеллы, заключенные в белковые оболочки (такие как белковые клетки-инкапсулиновые клетки ). были обнаружены [ 7 ] [ 8 ] наряду с другими прокариотическими органеллами . [ 9 ] Будучи одноклеточными, некоторые прокариоты, такие как цианобактерии , могут образовывать колонии, скрепленные биопленками , а большие колонии могут создавать многослойные микробные маты . Другие, такие как миксобактерии многоклеточные стадии , имеют в своем жизненном цикле . [ 10 ] Прокариоты бесполы и размножаются путем бинарного деления без слияния гамет , хотя горизонтальный перенос генов может иметь место .
Молекулярные исследования позволили понять эволюцию и взаимосвязь трех областей жизни. [ 11 ] Разделение на прокариотов и эукариотов отражает существование двух совершенно разных уровней клеточной организации; только эукариотические клетки имеют покрытое оболочкой ядро, содержащее хромосомную ДНК и другие характерные мембраносвязанные органеллы, включая митохондрии. Отличительные типы прокариот включают экстремофилы и метаногены ; они распространены в некоторых экстремальных условиях. [ 2 ]
История
[ редактировать ]Различие между прокариотами и эукариотами было твердо установлено микробиологами Роджером Станье и К.Б. ван Нилем в их статье 1962 года «Концепция бактерии». [ 12 ] (хотя там пишется прокариот и эукариот). В этой статье цитируется книга Эдуарда Шаттона 1937 года «Названия и научные работы». [ 13 ] за использование этих терминов и признание различия. Одной из причин этой классификации было то, что то, что тогда часто называли сине-зелеными водорослями (теперь называемыми цианобактериями ), не относилось к растениям, а группировалось с бактериями.
Структура
[ редактировать ]Прокариоты имеют прокариотический цитоскелет , более примитивный, чем у эукариот. Помимо гомологов актина и тубулина ( MreB и FtsZ ), спирально расположенный строительный блок жгутика , флагеллин , является одним из наиболее важных белков цитоскелета бактерий, поскольку он обеспечивает структурную основу хемотаксиса , основной физиологической реакции клеток бактерий. . По крайней мере, некоторые прокариоты также содержат внутриклеточные структуры, которые можно рассматривать как примитивные органеллы.
Мембранные органеллы (или внутриклеточные мембраны) известны в некоторых группах прокариот, например вакуоли или мембранные системы, обладающие особыми метаболическими свойствами, такими как фотосинтез или хемолитотрофия . Кроме того, некоторые виды также содержат микрокомпартменты, заключенные в углеводы, которые имеют различную физиологическую роль (например, карбоксисомы или газовые вакуоли).
Размер большинства прокариот составляет от 1 до 10 мкм, но их размер может варьироваться от 0,2 мкм ( Mycoplasmagentium ) до 750 мкм ( Thiomargarita namibiensis ).
| Структура прокариотической клетки | Описание |
|---|---|
| Жгутик (не всегда присутствует) | Длинный, похожий на хлыст выступ, который способствует передвижению клеток, используемому как грамположительными, так и грамотрицательными бактериями . |
| Клеточная мембрана | Окружает цитоплазму клетки и регулирует поток веществ в клетку и из нее. |
| Клеточная стенка (кроме родов Mycoplasma и Thermoplasma ) | Наружное покрытие большинства клеток, защищающее бактериальную клетку и придающее ей форму. |
| Цитоплазма | Гелеобразное вещество, состоящее в основном из воды, которое также содержит ферменты, соли, клеточные компоненты и различные органические молекулы. |
| Рибосома | Клеточные структуры, ответственные за выработку белка. |
| нуклеоид | Область цитоплазмы, содержащая одну молекулу ДНК прокариота. |
| Гликокаликс (только у некоторых типов прокариот) | Гликопротеин оболочка , – полисахаридная окружающая клеточные мембраны. |
| Цитоплазматические включения | Включения , такие как рибосомы и более крупные массы, разбросаны по цитоплазматическому матриксу. |
Морфология
[ редактировать ]Прокариотические клетки имеют различную форму; Четыре основные формы бактерий: [ 14 ]
- Кокки . Бактерия сферической или яйцевидной формы называется кокком (множественное число, кокки). например, стрептококк, стафилококк.
- Бациллы – бактерия цилиндрической формы, называемая палочкой или бациллой (множественное число, бациллы).
- Спиральные бактерии . Некоторые палочки скручиваются в спиральную форму и называются спириллами (единственное число, spirillum).
- Вибрион – в форме запятой
Архей Haloquadratum имеет плоские клетки квадратной формы. [ 15 ]
Воспроизведение
[ редактировать ]Бактерии и археи размножаются путем бесполого размножения, обычно путем бинарного деления . Генетический обмен и рекомбинация все еще происходят, но это форма горизонтального переноса генов , а не репликативный процесс, включающий просто перенос ДНК между двумя клетками, как при бактериальной конъюгации .
перенос ДНК
[ редактировать ]Перенос ДНК между прокариотическими клетками происходит у бактерий и архей, хотя в основном он изучается у бактерий. У бактерий перенос генов происходит посредством трех процессов. Это (1) и опосредованная бактериальным вирусом (бактериофагом) трансдукция, ( 3 ) , (2) плазмидная конъюгация естественная трансформация . Трансдукция бактериальных генов бактериофагом, по-видимому, отражает случайную ошибку во время внутриклеточной сборки вирусных частиц, а не адаптацию бактерий-хозяев. Перенос бактериальной ДНК находится под контролем генов бактериофага, а не бактериальных генов. Конъюгация в хорошо изученной системе E. coli контролируется плазмидными генами и представляет собой адаптацию для распространения копий плазмиды от одного бактериального хозяина к другому. Нечасто во время этого процесса плазмида может интегрироваться в бактериальную хромосому хозяина и впоследствии передавать часть бактериальной ДНК хозяина другой бактерии. Плазмидный перенос бактериальной ДНК хозяина (конъюгация) также является случайным процессом, а не бактериальной адаптацией.
Естественная бактериальная трансформация включает перенос ДНК от одной бактерии к другой через промежуточную среду. В отличие от трансдукции и конъюгации, трансформация, очевидно, является бактериальной адаптацией для переноса ДНК, поскольку она зависит от многочисленных продуктов бактериальных генов, которые специфически взаимодействуют для выполнения этого сложного процесса. [ 16 ] Чтобы бактерия могла связать, забрать и рекомбинировать донорскую ДНК в свою собственную хромосому, она должна сначала войти в особое физиологическое состояние, называемое компетентностью . требуется около 40 генов . у Bacillus subtilis Для развития компетентности [ 17 ] Длина ДНК, перенесенной при трансформации B. subtilis, может достигать трети всей хромосомы. [ 18 ] [ 19 ] Трансформация является распространенным способом переноса ДНК, и на данный момент известно, что 67 видов прокариот естественным образом способны к трансформации. [ 20 ]
Среди архей Halobacterium volcanii образует цитоплазматические мостики между клетками, которые, по-видимому, используются для переноса ДНК из одной клетки в другую. [ 21 ] Другой архей, Sulfolobus solfataricus , переносит ДНК между клетками путем прямого контакта. Фролс и др. (2008) найдено [ 22 ] что воздействие на S. solfataricus агентов, повреждающих ДНК, вызывает агрегацию клеток, и предположили, что клеточная агрегация может усиливать перенос ДНК между клетками, обеспечивая усиленное восстановление поврежденной ДНК посредством гомологичной рекомбинации.
Социальность
[ редактировать ]Хотя прокариоты считаются строго одноклеточными, большинство из них могут образовывать стабильные агрегатные сообщества. [ 23 ] Когда такие сообщества заключены в стабилизирующую полимерную матрицу («слизь»), их можно назвать « биопленками ». [ 24 ] Клетки в биопленках часто демонстрируют различные закономерности экспрессии генов (фенотипическую дифференциацию) во времени и пространстве. Кроме того, как и в случае с многоклеточными эукариотами, эти изменения в экспрессии часто являются результатом передачи сигналов от клетки к клетке — явления, известного как ощущение кворума .
Биопленки могут быть очень гетерогенными и структурно сложными и могут прикрепляться к твердым поверхностям или существовать на границах раздела жидкость-воздух или, возможно, даже на границах раздела жидкость-жидкость. Бактериальные биопленки часто состоят из микроколоний (примерно куполообразных масс бактерий и матрикса), разделенных «пустотами», через которые среда (например, вода) может легко течь. Микроколонии могут объединяться над субстратом, образуя сплошной слой, замыкая сеть каналов, разделяющих микроколонии. Эта структурная сложность в сочетании с наблюдениями о том, что ограничение кислорода (повсеместная проблема для всего, что растет в размерах за пределами масштаба диффузии) по крайней мере частично облегчается движением среды по всей биопленке, заставила некоторых предположить, что это может представлять собой систему кровообращения. [ 25 ] и многие исследователи стали называть прокариотические сообщества многоклеточными (например [ 26 ] ). Дифференциальная клеточная экспрессия, коллективное поведение, передача сигналов, запрограммированная гибель клеток и (в некоторых случаях) дискретное биологическое распространение. [ 27 ] все события, кажется, указывают в этом направлении. Однако эти колонии редко, если вообще когда-либо, основываются одним основателем (так же, как животные и растения создаются отдельными клетками), что представляет собой ряд теоретических проблем. Большинство объяснений сотрудничества и эволюции многоклеточности сосредоточены на высоком родстве между членами группы (или колонии, или целого организма). Если копия гена присутствует у всех членов группы, поведение, способствующее сотрудничеству между членами, может позволить этим членам иметь (в среднем) большую приспособленность, чем аналогичная группа эгоистичных людей. [ 28 ] (см. инклюзивную приспособленность и правило Гамильтона ).
Если эти случаи социальности прокариот окажутся скорее правилом, чем исключением, это будет иметь серьезные последствия для того, как мы рассматриваем прокариотов в целом и как мы обращаемся с ними в медицине. [ 29 ] Бактериальные биопленки могут быть в 100 раз более устойчивы к антибиотикам, чем свободноживущие одноклеточные клетки, и их практически невозможно удалить с поверхностей после их колонизации. [ 30 ] Другие аспекты бактериального сотрудничества, такие как бактериальная конъюгация , опосредованная ощущением кворума и патогенность , представляют дополнительные проблемы для исследователей и медицинских работников, стремящихся лечить сопутствующие заболевания.
Среда
[ редактировать ]
За время своего долгого существования прокариоты сильно диверсифицировались. Метаболизм прокариот гораздо более разнообразен, чем у эукариот, что приводит к появлению множества весьма различных типов прокариот. Например, помимо использования фотосинтеза или органических соединений для получения энергии, как это делают эукариоты, прокариоты могут получать энергию из неорганических соединений, таких как сероводород . Это позволяет прокариотам процветать в суровых условиях, таких холодных, как снежная поверхность Антарктиды , изучаемых в криобиологии , или таких жарких, как подводные гидротермальные источники и наземные горячие источники .
Прокариоты обитают практически во всех средах на Земле. Некоторые археи и бактерии являются экстремофилами , процветающими в суровых условиях, таких как высокие температуры ( термофилы ) или высокая соленость ( галофилы ). [ 31 ] Многие археи растут в виде планктона в океанах. Симбиотические прокариоты живут в телах других организмов, включая человека, или на них. Прокариоты имеют высокие популяции в почве , включая ризосферу и ризооболочку . Почвенные прокариоты все еще недостаточно изучены, несмотря на их близость к человеку и огромное экономическое значение для сельского хозяйства . [ 32 ]
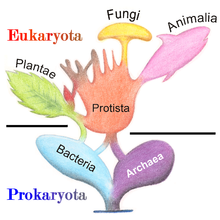
Классификация
[ редактировать ]В 1977 году Карл Везе предложил разделить прокариоты на бактерии и археи (первоначально Eubacteria и Archaebacteria) из-за серьезных различий в структуре и генетике между двумя группами организмов. Первоначально считалось, что археи являются экстремофилами, живущими только в негостеприимных условиях, таких как экстремальные температуры , pH и радиация , но с тех пор они были обнаружены во всех типах среды обитания . Полученное в результате расположение эукариот (также называемых «эукариями»), бактерий и архей называется трехдоменной системой , заменяющей традиционную систему двух империй . [ 33 ] [ 34 ]
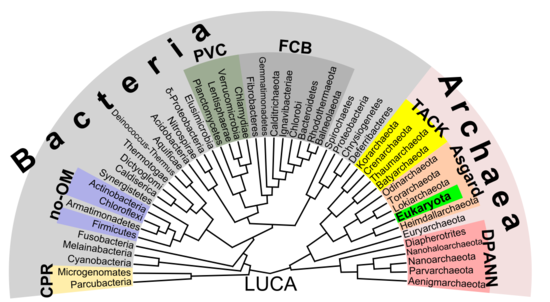
Филогенетическое дерево
[ редактировать ]Согласно филогенетическому анализу Хуга (2016), связи могут быть следующими: [ 35 ]

Эволюция
[ редактировать ]

Широко распространенная современная модель эволюции первых живых организмов состоит в том, что это были некие формы прокариотов, которые, возможно, развились из протоклеток , тогда как эукариоты развились позже в истории жизни. [ 37 ] Некоторые авторы поставили под сомнение этот вывод, утверждая, что нынешний набор видов прокариот, возможно, произошел от более сложных предков эукариот в процессе упрощения. [ 38 ] [ 39 ] [ 40 ]
Другие утверждали, что три области жизни возникли одновременно из набора разнообразных клеток, сформировавших единый генофонд. [ 41 ] Этот спор был подытожен в 2005 году: [ 42 ]
Среди биологов нет единого мнения относительно места эукариотов в общей схеме клеточной эволюции. Современные мнения о происхождении и положении эукариот охватывают широкий спектр, включая взгляды о том, что эукариоты возникли первыми в эволюции и что прокариоты произошли от них, что эукариоты возникли одновременно с эубактериями и архебактериями и, следовательно, представляют собой первичную линию происхождения одинакового возраста и ранга. как и прокариоты, эукариоты возникли в результате симбиотического события, влекущего за собой эндосимбиотическое происхождение ядра, что эукариоты возникли без эндосимбиоза, и что эукариоты возникли в результате симбиотического события, влекущего за собой одновременное эндосимбиотическое происхождение жгутика и ядра, в дополнение ко многим другим моделям, которые были рассмотрены и обобщены в других источниках.
Самые старые известные окаменелые прокариоты возникли примерно 3,5 миллиарда лет назад, всего примерно через 1 миллиард лет после образования земной коры. Эукариоты появляются в летописи окаменелостей только позже и, возможно, образовались в результате эндосимбиоза нескольких предков прокариот. Возраст самых старых известных ископаемых эукариот составляет около 1,7 миллиарда лет. Однако некоторые генетические данные свидетельствуют о том, что эукариоты появились еще 3 миллиарда лет назад. [ 43 ]
Хотя Земля — единственное место во Вселенной, где, как известно, существует жизнь, некоторые предполагают, что на Марсе есть свидетельства существования ископаемых или живых прокариот. [ 44 ] [ 45 ] Однако эта возможность остается предметом серьезных дискуссий и скептицизма. [ 46 ] [ 47 ]
Отношения с эукариотами
[ редактировать ]
Разделение на прокариотов и эукариотов обычно считается наиболее важным различием или различием между организмами. Разница в том, что эукариотические клетки имеют «настоящее» ядро , содержащее их ДНК , тогда как прокариотические клетки не имеют ядра.
И эукариоты, и прокариоты содержат крупные РНК / белка структуры , называемые рибосомами , которые производят белок , но рибосомы прокариот меньше, чем у эукариот. Митохондрии и хлоропласты , две органеллы, обнаруженные во многих эукариотических клетках, содержат рибосомы, аналогичные по размеру и составу рибосомам, обнаруженным у прокариот. [ 48 ] Это одно из многих доказательств того, что митохондрии и хлоропласты произошли от свободноживущих бактерий. Эндосимбиотическая теория утверждает, что ранние эукариотические клетки поглощали примитивные прокариотические клетки путем фагоцитоза и адаптировались, чтобы включить их структуры, что привело к образованию митохондрий и хлоропластов.
Геном называемом прокариот содержится в комплексе ДНК/белок в цитозоле, нуклеоидом , у которого отсутствует ядерная оболочка . [ 49 ] Комплекс содержит одну циклическую двухцепочечную молекулу стабильной хромосомной ДНК, в отличие от множества линейных, компактных, высокоорганизованных хромосом , обнаруженных в эукариотических клетках. Кроме того, многие важные гены прокариот хранятся в отдельных кольцевых структурах ДНК, называемых плазмидами . [ 3 ] Как и эукариоты, прокариоты могут частично дублировать генетический материал и могут иметь гаплоидный хромосомный состав, который частично реплицируется — состояние, известное как меродиплоидия . [ 50 ]
У прокариот отсутствуют митохондрии и хлоропласты . Вместо этого такие процессы, как окислительное фосфорилирование и фотосинтез, происходят через мембрану прокариотической клетки . [ 51 ] Однако прокариоты обладают некоторыми внутренними структурами, такими как прокариотический цитоскелет . [ 52 ] [ 53 ] Было высказано предположение, что бактериальный тип Planctomycetota имеет мембрану вокруг нуклеоида и содержит другие мембраносвязанные клеточные структуры. [ 54 ] Однако дальнейшее исследование показало, что клетки Planctomycetota не разделены и не содержат ядер и, как и другие бактериальные мембранные системы, взаимосвязаны. [ 55 ]
Прокариотические клетки обычно намного меньше эукариотических клеток. [ 3 ] Следовательно, прокариоты имеют большее соотношение площади поверхности к объему , что обеспечивает им более высокую скорость метаболизма , более высокую скорость роста и, как следствие, более короткое время генерации, чем у эукариот. [ 3 ]
Появляется все больше свидетельств того, что корни эукариот можно найти в архейской группе асгардов (или, по крайней мере, рядом с ними) , возможно, Heimdallarchaeota (идея, которая представляет собой современную версию эоцитарной гипотезы 1984 года , где эоциты являются старым синонимом Thermoproteota). , таксон , который можно было найти недалеко от тогда еще неизвестной группы Асгарда). [ 56 ] Например, гистоны , которые обычно упаковывают ДНК в ядрах эукариот, также были обнаружены у нескольких групп архей, что свидетельствует о гомологии . прояснить загадочного предшественника эукариотических клеток ( эуцитов который поглотил альфа-протеобактерию, первый эуцит ( LECA , последний общий эукариот образующую предок Эта идея могла бы ) , ) согласно эндосимбиотической теории . Возможно, существовала некоторая дополнительная поддержка со стороны вирусов, называемая вирусным эукариогенезом . Небактериальную группу, состоящую из архей и эукариот, Neomura в 2002 году назвал Томас Кавальер-Смит . [ 57 ] Однако с кладистической точки зрения эукариоты являются архей в том же смысле, в каком птицы являются динозаврами , поскольку они произошли от группы динозавров манираптора . Напротив, археи без эукариот кажутся парафилетической группой, точно так же, как динозавры без птиц.
Прокариоты можно разделить на две группы.
[ редактировать ]В отличие от вышеизложенного предположения о фундаментальном расколе между прокариотами и эукариотами, важнейшим отличием биоты может быть разделение между бактериями и остальными (археями и эукариотами). [ 56 ] Например, репликация ДНК у бактерий и архей фундаментально различается (в том числе в ядрах эукариот) и может не быть гомологичной между этими двумя группами. [ 58 ] Более того, АТФ-синтаза , хотя и является общей (гомологичной) во всех организмах, сильно различается между бактериями (включая эукариотические органеллы, такие как митохондрии и хлоропласты ) и группой ядер архей/эукариот. Последний общий предок всей жизни (называемый общий предок ) должен был последний универсальный обладать , LUCA ранней версией этого белкового комплекса. Поскольку АТФ-синтаза обязательно связана с мембраной, это подтверждает предположение о том, что LUCA представляет собой клеточный организм. Гипотеза мира РНК могла бы прояснить этот сценарий, поскольку LUCA мог быть рибоцитом (также называемым рибоклеткой) без ДНК, но с геномом РНК, построенным рибосомами как первичными самовоспроизводящимися объектами . [ 59 ] Гипотеза мира пептидов-РНК (также называемая миром RNP ) была предложена на основе идеи о том, что олигопептиды могли быть построены вместе с первичными нуклеиновыми кислотами одновременно, что также подтверждает концепцию рибоцита как LUCA. Особенность ДНК как материальной основы генома затем могла быть принята отдельно у бактерий и архей (а позже и у ядер эукариот), предположительно, с помощью некоторых вирусов (возможно, ретровирусов, поскольку они могли осуществлять обратную транскрипцию РНК в ДНК). [ 60 ] В результате прокариоты, включающие бактерии и археи, также могут быть полифилетическими .
См. также
[ редактировать ]- Актинонин
- Структура бактериальной клетки
- Комбрекс
- Эволюция клеток
- Эволюция полового размножения
- Список секвенированных геномов архей
- Список секвенированных бактериальных геномов
- Морские прокариоты
- Монера — устаревшее царство , включающее архей и бактерий.
- Нанобактерия
- Взрослый
- Паракарион миогенный
- ProGlycProt
Ссылки
[ редактировать ]- ^ «Определение ПРОКАРИОТА» . www.merriam-webster.com . Проверено 30 декабря 2023 г.
- ^ Jump up to: а б Государственный университет Северной Каролины . «Прокариоты: одноклеточные организмы» .
- ^ Jump up to: а б с д Кэмпбелл, Н. «Биология: концепции и связи». Пирсон Образование. Сан-Франциско: 2003.
- ^ «прокариот» . Интернет-словарь этимологии .
- ^ Сапп, Дж . (2005). «Дихотомия прокариот-эукариот: значения и мифология» . Обзоры микробиологии и молекулярной биологии . 69 (2): 292–305. дои : 10.1128/MMBR.69.2.292-305.2005 . ПМЦ 1197417 . ПМИД 15944457 .
- ^ Коте Дж., Де Туллио М. (2010). «Помимо прокариотов и эукариотов: планктомицеты и клеточная организация» . Природа .
- ^ Керфельд К.А., Савая М.Р., Танака С., Нгуен К.В., Филлипс М., Биби М., Йейтс Т.О. (август 2005 г.). «Белковые структуры, образующие оболочку примитивных бактериальных органелл». Наука . 309 (5736): 936–8. Бибкод : 2005Sci...309..936K . CiteSeerX 10.1.1.1026.896 . дои : 10.1126/science.1113397 . ПМИД 16081736 . S2CID 24561197 .
- ^ Мурат Д., Бирн М., Комейли А. (октябрь 2010 г.). «Клеточная биология прокариотических органелл» . Перспективы Колд-Спринг-Харбор в биологии . 2 (10): а000422. doi : 10.1101/cshperspect.a000422 . ПМЦ 2944366 . ПМИД 20739411 .
- ^ Мюрат, Доротея; Бирн, Меган; Комейли, Араш (01 октября 2010 г.). «Клеточная биология прокариотических органелл» . Перспективы Колд-Спринг-Харбор в биологии . 2 (10): а000422. doi : 10.1101/cshperspect.a000422 . ПМЦ 2944366 . ПМИД 20739411 .
- ^ Кайзер Д. (октябрь 2003 г.). «Связь движения клеток с многоклеточным развитием миксобактерий». Обзоры природы. Микробиология . 1 (1): 45–54. дои : 10.1038/nrmicro733 . ПМИД 15040179 . S2CID 9486133 .
- ^ Сун К.Х., Сон Х.К. (22 июля 2014 г.). «Понимание молекулярной эволюции АТФазы HslU посредством биохимического и мутационного анализа» . ПЛОС ОДИН . 9 (7): e103027. Бибкод : 2014PLoSO...9j3027S . дои : 10.1371/journal.pone.0103027 . ПМК 4106860 . ПМИД 25050622 .
- ^ Станье Р.Ю. , Ван Нил CB (1962). «Понятие о бактерии». Архив микробиологии . 42 (1): 17–35. дои : 10.1007/BF00425185 . ПМИД 13916221 . S2CID 29859498 .
- ^ Чаттон Э (1937). Научные звания и работы (1906-1937) Эдуарда Шаттона . Сет: Печать. Э. Соттано.
- ^ Бауман Р.В., Тизард И.Р., Мачунис-Масука Э. (2006). Микробиология . Сан-Франциско: Пирсон Бенджамин Каммингс. ISBN 978-0-8053-7693-7 .
- ^ Стоекениус В. (октябрь 1981 г.). «Квадратная бактерия Уолсби: тонкая структура ортогонального прокариота» . Журнал бактериологии . 148 (1): 352–60. дои : 10.1128/JB.148.1.352-360.1981 . ПМК 216199 . ПМИД 7287626 .
- ^ Чен И, Дубнау Д (март 2004 г.). «Поглощение ДНК во время бактериальной трансформации». Обзоры природы. Микробиология . 2 (3): 241–9. дои : 10.1038/nrmicro844 . ПМИД 15083159 . S2CID 205499369 .
- ^ Соломон Дж. М., Гроссман А. Д. (апрель 1996 г.). «Кто компетентен и когда: регулирование естественной генетической компетентности бактерий». Тенденции в генетике . 12 (4): 150–5. дои : 10.1016/0168-9525(96)10014-7 . ПМИД 8901420 .
- ^ Акамацу Т., Тагучи Х. (апрель 2001 г.). «Включение всей хромосомной ДНК в лизаты протопластов в компетентные клетки Bacillus subtilis» . Бионауки, биотехнологии и биохимия . 65 (4): 823–9. дои : 10.1271/bbb.65.823 . ПМИД 11388459 . S2CID 30118947 .
- ^ Сайто Ю., Тагучи Х., Акамацу Т. (март 2006 г.). «Судьба трансформации бактериального генома после включения в компетентные клетки Bacillus subtilis: непрерывная длина включенной ДНК». Журнал бионауки и биоинженерии . 101 (3): 257–62. дои : 10.1263/jbb.101.257 . ПМИД 16716928 .
- ^ Джонсборг О., Эльдхольм В., Ховарштайн Л.С. (декабрь 2007 г.). «Естественная генетическая трансформация: распространенность, механизмы и функции» . Исследования в области микробиологии . 158 (10): 767–78. дои : 10.1016/j.resmic.2007.09.004 . ПМИД 17997281 .
- ^ Розеншин И., Челет Р., Мевареч М. (сентябрь 1989 г.). «Механизм переноса ДНК в системе спаривания архебактерий». Наука . 245 (4924): 1387–9. Бибкод : 1989Sci...245.1387R . дои : 10.1126/science.2818746 . ПМИД 2818746 .
- ^ Фролс С., Айон М., Вагнер М., Тейхманн Д., Золгадр Б., Фолеа М., Букема Э.Дж., Дриссен А.Ю., Шлепер С., Альберс С.В. (ноябрь 2008 г.). «Индуцируемая УФ-излучением клеточная агрегация гипертермофильных архей Sulfolobus solfataricus опосредована образованием пилей» (PDF) . Молекулярная микробиология . 70 (4): 938–52. дои : 10.1111/j.1365-2958.2008.06459.x . ПМИД 18990182 . S2CID 12797510 .
- ^ Мэдиган Т (2012). Брока биология микроорганизмов (13-е изд.). Сан-Франциско: Бенджамин Каммингс. ISBN 9780321649638 .
- ^ Костертон Дж.В. (2007). «Прямые наблюдения». Праймер для биопленки . Серия Springer о биопленках. Том. 1. Берлин, Гейдельберг: Шпрингер. стр. 3–4. дои : 10.1007/978-3-540-68022-2_2 . ISBN 978-3-540-68021-5 .
- ^ Костертон Дж.В., Левандовски З., Колдуэлл Д.Е., Корбер Д.Р., Лаппин-Скотт Х.М. (октябрь 1995 г.). «Микробные биопленки». Ежегодный обзор микробиологии . 49 (1): 711–45. дои : 10.1146/annurev.mi.49.100195.003431 . ПМИД 8561477 .
- ^ Шапиро Дж. А. (1998). «Думая о бактериальных популяциях как о многоклеточных организмах» (PDF) . Ежегодный обзор микробиологии . 52 (1): 81–104. дои : 10.1146/аннурев.микро.52.1.81 . ПМИД 9891794 . Архивировано из оригинала (PDF) 17 июля 2011 г.
- ^ Чуа С.Л., Лю Ю., Ям Дж.К., Чен Ю., Вейборг Р.М., Тан Б.Г., Кьеллеберг С., Толкер-Нильсен Т., Гивсков М., Ян Л. (июль 2014 г.). «Дисперсные клетки представляют собой отдельную стадию перехода от бактериальной биопленки к планктонному образу жизни» . Природные коммуникации . 5 (1): 4462. Бибкод : 2014NatCo...5.4462C . дои : 10.1038/ncomms5462 . ПМИД 25042103 .
- ^ Гамильтон У. Д. (июль 1964 г.). «Генетическая эволюция социального поведения. II». Журнал теоретической биологии . 7 (1): 17–52. Бибкод : 1964JThBi...7...17H . дои : 10.1016/0022-5193(64)90039-6 . ПМИД 5875340 .
- ^ Балабан Н., Рен Д., Гивсков М., Расмуссен Т.Б. (2008). "Введение". Контроль инфекций биопленок путем манипулирования сигналами . Серия Springer о биопленках. Том. 2. Берлин, Гейдельберг: Шпрингер. стр. 1–11. дои : 10.1007/7142_2007_006 . ISBN 978-3-540-73852-7 .
- ^ Костертон Дж.В., Стюарт П.С., Гринберг Е.П. (май 1999 г.). «Бактериальные биопленки: частая причина персистирующих инфекций» . Наука . 284 (5418): 1318–22. Бибкод : 1999Sci...284.1318C . дои : 10.1126/science.284.5418.1318 . ПМИД 10334980 . S2CID 27364291 .
- ^ Хоган CM (2010). «Экстремофил» . В Моноссон Э, Кливленд С (ред.). Энциклопедия Земли . Национальный совет науки и окружающей среды.
- ^ Кобиан Гуэмес, Ана Джорджина; Юл, Мерри; Канту, Вито Адриан; Фелтс, Бен; Нултон, Джеймс; Ровер, Форест (29 сентября 2016 г.). «Вирусы как победители в игре жизни». Ежегодный обзор вирусологии . 3 (1). Годовые обзоры : 197–214. doi : 10.1146/annurev-virology-100114-054952 . ISSN 2327-056X . ПМИД 27741409 . S2CID 36517589 .
- ^ Вёзе CR (март 1994 г.). «Где-то должен быть прокариот: микробиология ищет себя» . Микробиологические обзоры . 58 (1): 1–9. дои : 10.1128/MMBR.58.1.1-9.1994 . ПМЦ 372949 . ПМИД 8177167 .
- ^ Сапп Дж. (июнь 2005 г.). «Дихотомия прокариот-эукариот: смыслы и мифология» . Обзоры микробиологии и молекулярной биологии . 69 (2): 292–305. дои : 10.1128/MMBR.69.2.292-305.2005 . ПМЦ 1197417 . ПМИД 15944457 .
- ^ Jump up to: а б Обнимаю, Лаура А.; Бейкер, Бретт Дж.; Анантараман, Картик; Браун, Кристофер Т.; Пробст, Александр Дж.; Кастель, Синди Дж.; Баттерфилд, Кристина Н.; Хернсдорф, Алекс В.; Амано, Юки; Исе, Котаро; Сузуки, Йохей; Дудек, Наташа; Релман, Дэвид А.; Финстад, Кари М.; Амундсон, Рональд (11 апреля 2016 г.). «Новый взгляд на древо жизни» . Природная микробиология . 1 (5): 1–6. doi : 10.1038/nmicrobiol.2016.48 . ISSN 2058-5276 .
- ^ Эгель Р. (январь 2012 г.). «Первичный эукариогенез: об коллективной природе доклеточных состояний, предков современной жизни» . Жизнь . 2 (1): 170–212. Бибкод : 2012Life....2..170E . дои : 10.3390/life2010170 . ПМЦ 4187143 . ПМИД 25382122 .
- ^ Циммер С. (август 2009 г.). «Происхождение. О происхождении эукариот». Наука . 325 (5941): 666–8. дои : 10.1126/science.325_666 . ПМИД 19661396 .
- ^ Браун-младший (февраль 2003 г.). «Древний горизонтальный перенос генов». Обзоры природы. Генетика . 4 (2): 121–32. дои : 10.1038/nrg1000 . ПМИД 12560809 . S2CID 22294114 .
- ^ Фортер П. , Филипп Х. (октябрь 1999 г.). «Где корень вселенского древа жизни?». Биоэссе . 21 (10): 871–9. doi : 10.1002/(SICI)1521-1878(199910)21:10<871::AID-BIES10>3.0.CO;2-Q . ПМИД 10497338 .
- ^ Пул А., Джеффарес Д., Пенни Д. (октябрь 1999 г.). «Ранняя эволюция: прокариоты, новые дети на районе». Биоэссе . 21 (10): 880–9. doi : 10.1002/(SICI)1521-1878(199910)21:10<880::AID-BIES11>3.0.CO;2-P . ПМИД 10497339 . S2CID 45607498 .
- ^ Вёзе С. (июнь 1998 г.). «Всемирный предок» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (12): 6854–9. Бибкод : 1998PNAS...95.6854W . дои : 10.1073/pnas.95.12.6854 . ПМК 22660 . ПМИД 9618502 .
- ^ Мартин, Уильям (2005). «Горе древу жизни». В Саппе, Ян (ред.). Микробная филогения и эволюция: концепции и противоречия . Оксфорд: Издательство Оксфордского университета . п. 139.
- ^ Карл Вёзе , Дж. Питер Гогартен , « Когда эукариотические клетки (клетки с ядрами и другими внутренними органеллами) впервые развились? Что мы знаем о том, как они произошли от более ранних форм жизни? » Scientific American , 21 октября 1999 г.
- ^ Максуин HY (июль 1997 г.). «Доказательства жизни в марсианском метеорите?». ГСА сегодня . 7 (7): 1–7. ПМИД 11541665 .
- ^ Маккей Д.С., Гибсон Э.К., Томас-Кепрта К.Л., Вали Х., Романек К.С., Клеметт С.Дж., Чиллиер К.Д., Мэхлинг Ч.Р., Заре Р.Н. (август 1996 г.). «Поиски прошлой жизни на Марсе: возможная реликтовая биогенная активность в марсианском метеорите ALH84001». Наука . 273 (5277): 924–30. Бибкод : 1996Sci...273..924M . дои : 10.1126/science.273.5277.924 . ПМИД 8688069 . S2CID 40690489 .
- ^ Кренсон М. (6 августа 2006 г.). «Спустя 10 лет мало кто верит в жизнь на Марсе» . Associated Press (на сайте space.com). Архивировано из оригинала 9 августа 2006 г. Проверено 6 августа 2006 г.
- ^ Скотт Э.Р. (февраль 1999 г.). «Происхождение карбонат-магнетит-сульфидных комплексов в марсианском метеорите ALH84001» . Журнал геофизических исследований . 104 (Е2): 3803–13. Бибкод : 1999JGR...104.3803S . дои : 10.1029/1998JE900034 . ПМИД 11542931 .
- ^ Брюс Альбертс; и др. (2002). Молекулярная биология клетки (четвертое изд.). Гирляндная наука. п. 808. ИСБН 0-8153-3218-1 .
- ^ Танбихлер М., Ван С.К., Шапиро Л. (октябрь 2005 г.). «Бактериальный нуклеоид: высокоорганизованная и динамичная структура» . Журнал клеточной биохимии . 96 (3): 506–21. дои : 10.1002/jcb.20519 . ПМИД 15988757 . S2CID 25355087 .
- ^ Джонстон С., Каймарис С., Зомер А., Бутсма Х.Дж., Прюдомм М., Гранадель С., Херманс П.В., Полард П., Мартин Б., Клаверис Дж.П. (2013). «Естественная генетическая трансформация создает популяцию меродиплоидов Streptococcus pneumoniae» . ПЛОС Генетика . 9 (9): e1003819. дои : 10.1371/journal.pgen.1003819 . ПМК 3784515 . ПМИД 24086154 .
- ^ Гарольд Ф.М. (июнь 1972 г.). «Сохранение и преобразование энергии бактериальными мембранами» . Бактериологические обзоры . 36 (2): 172–230. дои : 10.1128/MMBR.36.2.172-230.1972 . ПМК 408323 . ПМИД 4261111 .
- ^ Ши Ю.Л., Ротфилд Л. (сентябрь 2006 г.). «Бактериальный цитоскелет» . Обзоры микробиологии и молекулярной биологии . 70 (3): 729–54. дои : 10.1128/MMBR.00017-06 . ПМЦ 1594594 . ПМИД 16959967 .
- ^ Мичи К.А., Лёве Дж (2006). «Динамические нити бактериального цитоскелета» (PDF) . Ежегодный обзор биохимии . 75 (1): 467–92. doi : 10.1146/annurev.biochem.75.103004.142452 . ПМИД 16756499 . Архивировано из оригинала (PDF) 17 ноября 2006 г.
- ^ Фюрст Дж.А. (2005). «Внутриклеточная компартментация планктомицетов». Ежегодный обзор микробиологии . 59 (1): 299–328. дои : 10.1146/annurev.micro.59.030804.121258 . ПМИД 15910279 .
- ^ Сантарелла-Меллвиг Р., Пругналлер С., Роос Н., Маттай И.В., Девос Д.П. (2013). «Трехмерная реконструкция бактерий со сложной эндомембранной системой» . ПЛОС Биология . 11 (5): e1001565. дои : 10.1371/journal.pbio.1001565 . ПМК 3660258 . ПМИД 23700385 .
- ^ Jump up to: а б с Кастель Си Джей, Банфилд Дж. Ф. (март 2018 г.). «Основные новые группы микробов расширяют разнообразие и меняют наше понимание Древа Жизни» . Клетка . 172 (6): 1181–1197. дои : 10.1016/j.cell.2018.02.016 . ПМИД 29522741 .
- ^ Кавалер-Смит Т. (март 2002 г.). «Фаготрофное происхождение эукариот и филогенетическая классификация простейших» . Межд. Дж. Сист. Эвол. Микробиол . 52 (Часть 2): 297–354. дои : 10.1099/00207713-52-2-297 . ПМИД 11931142 .
- ^ Барри Э.Р., Белл С.Д. (декабрь 2006 г.). «Репликация ДНК у архей» . Обзоры микробиологии и молекулярной биологии . 70 (4): 876–87. дои : 10.1128/MMBR.00029-06 . ПМК 1698513 . ПМИД 17158702 .
- ^ Лейн Н (2015). Жизненно важный вопрос: энергия, эволюция и происхождение сложной жизни . WW Нортон . п. 77 . ISBN 978-0-393-08881-6 .
- ^ Фортерре П (2006). «Три клетки РНК для рибосомальных линий и три ДНК-вируса для репликации своих геномов: гипотеза происхождения клеточного домена» . ПНАС . 103 (10): 3669–3674. Бибкод : 2006PNAS..103.3669F . дои : 10.1073/pnas.0510333103 . ПМК 1450140 . ПМИД 16505372 .
Внешние ссылки
[ редактировать ]- Прокариоты против эукариотов, BioMineWiki. Архивировано 25 октября 2012 г. в Wayback Machine.
- Таксономическая схема бактерий и архей
- Дихотомия прокариот-эукариот: значения и мифология
- Викторина по анатомии прокариот
- Страница TOLWEB, посвященная филогении эукариот-прокариот
![]() В этой статье использованы общедоступные материалы из Научный букварь . НКБИ . Архивировано из оригинала 8 декабря 2009 г.
В этой статье использованы общедоступные материалы из Научный букварь . НКБИ . Архивировано из оригинала 8 декабря 2009 г.