Лягушка
| Лягушки Временный диапазон: ранняя юрская болезнь - присутствует ,
| |
|---|---|

| |
| Различные виды лягушки | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Сорт: | Амфибия |
| Клада : | Salientia |
| Заказ: | Приносит Duméril , 1806 (как Anoures) |
| Подгруппы | |
|
Смотрите текст | |
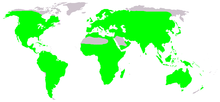
| |
| Нативное распределение лягушек (в зеленом) | |

Лягушка , -любой член разнообразной и в значительной степени плотоядную группу с коротким телом, бесполезными амфибиями сочиняющими порядка ануру [ 1 ] (исходя из древнегреческого ἀνούρα , буквально «без хвоста»). Самая старая ископаемая «прото-фрог» триадобатрах известен из раннего триаса Мадагаскара миллионов (250 лет назад ), но молекулярные часовые знакомства предполагают, что их раскол от других амфибий может распространиться на перми , 265 миллионов лет назад. Лягушки широко распространены, начиная от тропиков до субарктических областей, но наибольшая концентрация разнообразия видов находится в тропических тропических лесах . Лягушки составляют около 88% существующих видов амфибий. Они также являются одним из пяти самых разнообразных позвоночных заказов . Виды Warty лягушек, как правило, называются жабами , но различие между лягушками и жабами неформально, а не таксономии или эволюционной истории.
Взрослая лягушка имеет крепкое тело, выступающие глаза , спереди, прикрепленный языком , конечности, сложенные внизу, и без хвоста (хвост хвостовых лягушек -это расширение мужской клоаки). Лягушки имеют железистую кожу с выделениями от неприятных до токсичных. Их кожа варьируется по цвету от хорошо замаскированного пятнистого, коричневого, серого и зеленого до ярких узоров ярко-красного или желтого и черного, чтобы показать токсичность и отгонять хищников . Взрослые лягушки живут в пресной воде и на сухой земле; Некоторые виды адаптированы для жизни под землей или на деревьях.
Лягушки обычно откладывают яйца в воду. Яйца вылупляются в водные личинки , называемые головастиками с хвостами и внутренними жабрами . У них есть высокоспециализированные части рта, подходящие для травоядных , всеядных или плаковых диет. Жизненный цикл завершается, когда они метаморфизируются на взрослых. Несколько видов откладывают яйца на суше или обходят стадию головастика. У взрослых лягушек обычно есть хищная диета, состоящая из мелких беспозвоночных , но существуют всеядные виды, и некоторые питаются растительными веществами. Кожа лягушка имеет богатый микробиом , который важен для их здоровья. Лягушки чрезвычайно эффективны в превращении того, что они едят в массу тела. Они являются важным источником пищи для хищников и частью динамики пищевой сети мира многих экосистем . Кожа полупроницаемая , что делает их восприимчивыми к обезвоживанию, поэтому они либо живут в влажных местах, либо имеют специальные адаптации для борьбы с сухими средами обитания. Лягушки производят широкий спектр вокализаций , особенно в их сезон размножения и демонстрируют много разных видов сложных поведений , чтобы привлечь товарищей, чтобы отвалить хищников и вообще выжить.
Лягушки ценятся как пищу людьми, а также имеют много культурных ролей в литературе, символике и религии. Они также рассматриваются как экологические колокол , с снижением популяций лягушек, часто рассматриваются как ранние предупреждающие признаки ущерба для окружающей среды. Популяции лягушек значительно снизились с 1950 -х годов. Считается, что более трети видов находятся под угрозой исчезновения , и более 120, как полагают, вымерли с 1980 -х годов. Количество пороков развития среди лягушек растет, и возникающая грибковая болезнь, хитридиомикоз , распространилась по всему миру. Биологи сохранения работают над тем, чтобы понять причины этих проблем и решить их.
Этимология и таксономия
Использование общих имен лягушек и жабы не имеет таксономического оправдания. С точки зрения классификации все члены Anura Order Anura - это лягушки, но только члены семейства Bufonidae считаются «настоящими жабами». Использование термина лягушки в общих названиях обычно относится к видам, которые являются водными или полуакотичными и имеют гладкую влажную кожуру; Термин «жаба» обычно относится к видам, которые являются наземными с сухими, бородавленными шкурами. [ 2 ] [ 3 ] Есть многочисленные исключения из этого правила. Европейская пожарная жаба ( Бомбина Бомбана ) имеет слегка бородачную кожу и предпочитает водянистую среду обитания [ 4 ] Принимая во внимание, что панамская золотая лягушка ( Atelopus Zeteki ) находится в семействе Toad Bufonidae и имеет гладкую кожу. [ 5 ]
Этимология
Происхождение имени Ордена Анура- и его первоначальное правописание- это древнегреческий альфа привативный префикс ἀν- ( an- ἀ- - до гласного) 'без', [ 6 ] и οὐρά ( ourá ) 'Животный хвост'. [ 7 ] Значение «хрустка». Это относится к хрупковому характеру этих амфибий. [ 8 ] [ 9 ] [ 10 ]
Происхождение слова лягушка неясно и обсуждается. [ 11 ] Слово сначала засвидетельствовано на старом английском как Frogga , но обычное старое английское слово для лягушки было Frosc (с такими вариантами, как Frox и ForSC ), и согласно, что слово лягушка каким -то образом связано с этим. Старый английский Frosc оставался в диалекте на английском языке как Frosh и Frosk в девятнадцатом веке, [ 12 ] и широко совпадается с другими германскими языками с примерами на современных языках, включая немецкий фрош , норвежский фроск , исландский фроскир и голландский (Kik) Vors . [ 11 ] Эти слова позволяют реконструкции общего германского предка * Фроскиза . [ 13 ] Третье издание Оксфордского словаря английского языка обнаруживает, что этимология * Фросказа неопределенна, но соглашается с аргументами, что она может правдоподобно происходить от протоиндоевропейской базы в соответствии с * preu , что означает «прыжок». [ 11 ]
Однако, как и английское FROSC дало Frogga , однако, неясно, так как развитие не включает в себя регулярную смену . Вместо этого кажется, что в старом английском языке была тенденция к прозвищам для монеты для животных, заканчивающихся - G , с примерами - они все неопределенные этимологии, включая собаку , свинью , свинью, оленя и (ухо) парик . Лягушка , по -видимому, была адаптирована из FRSC как часть этой тенденции. [ 11 ]
Между тем, слово Toad , впервые засвидетельствованное как старого английского Tādige , является уникальным для английского и также имеет неопределенную этимологию. [ 14 ] Это основа для слова головастика , сначала засвидетельствованное как среднеанглийский Taddepol , очевидно, что означает «голова Toad». [ 15 ]
Таксономия
Около 88% амфибий видов классифицируются в Anura . [ 16 ] К ним относятся более 7600 видов [ 1 ] В 55 семействах , из которых Hylidae (1049 Spp.), Strabomantidae (797 Spp.), Microhylidae (744 Spp.) и Bufonidae (646 Spp.) являются самыми богатыми видами . [ 17 ]

Анура включает в себя все современные лягушки и любые ископаемые виды, которые вписываются в определение Анурана. : 9 или менее предварительные позвонки, наличие уростила, образованного из слитых позвонков, без хвоста, длинного и перспективного подвздошного костюма, более короткие передние конечности, чем радиус конечности локте , себя и задние Характеристики взрослых анурана включают в удлиненные кости лодыжки , отсутствие префронтальной кости, наличие подкованной пластины , нижняя челюсть без зубов (с За исключением Gastrotheca Guentheri ), состоящего из трех пар костей (ангулосплениальная, зубчатая и ментамекелеанская, причем последняя пара отсутствует в Pipoidea ),), [ 18 ] Неподдерживаемый язык, лимфатические пространства под кожей и мышца, протраторная лентис, прикрепленная к линзе глаза . [ 19 ] У личинки или головастика анурана есть один центральный респираторный дух и рта, состоящий из кератиновых клюв и зубных зубьев . [ 19 ]

Лягушки и жабы широко классифицируются на три подчинения: археобатрахия , которая включает в себя четыре семейства примитивных лягушек; Месобатрахия , которая включает в себя пять семей с более эволюционными промежуточными лягушками; и Neobatrachia , безусловно, самая большая группа, которая содержит оставшиеся семейства современных лягушек, включая наиболее распространенные виды по всему миру. Подряд Небатрахия дополнительно разделена на две суперсемейства Hyloidea и Ranoidea . [ 20 ] Эта классификация основана на таких морфологических особенностях, как количество позвонков, структуры грудного пояса и морфологии головастиков. Хотя эта классификация в значительной степени принята, отношения между семьями лягушек все еще обсуждаются. [ 21 ]
Некоторые виды Anurans гибридизуются легко . Например, съедобная лягушка ( Pelophylax Esculentus ) представляет собой гибрид между лягушкой для бассейна ( P. ressowae ) и болотной лягушкой ( P. ridibundus ). [ 22 ] Пожарные жабы Бомба Бомбана и Б. Варигата схожи по формирующим гибридам. Они менее плодородны, чем их родители, что приводит к гибридной зоне , где распространены гибриды. [ 23 ]
Эволюция
Происхождение и эволюционные отношения между тремя основными группами амфибий горячо обсуждаются. Молекулярная филогения , основанная на анализе рДНК , датируемой 2005 году, предполагает, что саламандры и цецилианцы более тесно связаны друг с другом, чем с лягушками, и дивергенция трех групп произошла в палеозое или раннем мезозое до разрушения суперконтинента и палеозоя и раннего мезозоя. Вскоре после их дивергенции от рыб с лепкой . Это поможет объяснить относительный дефицит окаменелостей амфибий из периода, прежде чем группы расстались. [ 24 ] Другой молекулярный филогенетический анализ, проведенный примерно в то же время, пришел к выводу, что Lissamphipians впервые появились около 330 миллионов лет назад и что гипотеза темноспондил-аоригина более заслуживает доверия, чем другие теории. Необатрахи , по -видимому, возникли в Африке/Индии, саламандре в Восточной Азии и противниках в тропической Пангее. [ 25 ] Другие исследователи, соглашаясь с основным направлением этого исследования, подвергли сомнению выбор калибровочных точек, используемых для синхронизации данных. Они предположили, что дата диверсификации Лиссамбиана была помещена в Перми , более 300 миллионов лет назад, дату в лучшем согласии с палеонтологическими данными. [ 26 ] Дальнейшее исследование в 2011 году с использованием как вымерших, так и живых таксонов, выбранных для морфологических данных, а также молекулярных данных, пришло к выводу, что Lissamphibia является монофилетической и что он должен быть введен в Lepospondyli , а не в Temnospondyli . Исследование поступило, что Lissamphibia возникла не раньше, чем позднее каменноугольное , примерно 290-305 миллионов лет назад. Распределение между Anura и Caudata оценивалось как состоявшаяся 292 миллиона лет назад, скорее, чем предполагают большинство молекулярных исследований, при этом Caecilians разделились 239 миллионов лет назад. [ 27 ]

В 2008 году был обнаружен геробатрах Хоттони , термосондил со многими характеристиками, похожими на лягушками и саламандре в Техасе . Он датируется 290 миллионами лет и был провозглашен как недостающее звено , стволовой батрахия, недалеко от общего предка лягушек и саламандров, что согласуется с общепринятой гипотезой о том, что лягушки и саламандры более тесно связаны друг с другом (формируя клад . ), чем они есть, для противя. [ 28 ] [ 29 ] Тем не менее, другие предположили, что геробатрах Хоттони был лишь дискорофоидным темноспондилом, не связанным с существующими амфибиями. [ 30 ]
Salientia (Latin Salire ( Salio ), «прыгать»)-это название общей группы, которая включает в себя современные лягушки в Anura Order, а также их близкие родственники окаменелости, «прото-фроги» или «стволовые фроги». Общие черты, которыми обладают эти прото-фроги, включают 14 предварительных позвонков (современные лягушки имеют восемь или 9), длинный и вперед-перспективный подвздошной кости в тазах , наличие фронтопариетальной кости и нижнюю челюсть без зубов. Самые ранние известные амфибии, которые были более тесно связаны с лягушками, чем с саламандрами, - это триадобатрах Массиноти , с раннего триасового периода Мадагаскара (около 250 миллионов лет назад) и Чекобатрах Полоник , с раннего триаса ( около того же возраста, что и триадобатрах ) [ 31 ] Череп триадобатраха похож на лягушку, широкий с большими глазами, но ископаемое имеет особенности, расходящиеся от современных лягушек. К ним относятся более длинное тело с большим количеством позвонков . Хвост имеет отдельные позвонки, в отличие от слитого уростила или копчика в современных лягушках. Кости большеберцовой кости также разделены, что делает вероятным, что триадобатрахус не был эффективным. [ 31 ] В исследовании 2019 года было отмечено присутствие Salientia от формации Чинл и предположило, что анураны могли бы впервые появиться во время позднего триаса . [ 32 ]
На основании ископаемых доказательств, самые ранние известные «истинные лягушки», которые попадают в должность ануранской линии, все жили в начале юрского периода. [ 2 ] [ 33 ] Один из таких ранних видов лягушек, Prosalirus Bitis , был обнаружен в 1995 году в формировании Кайента в Аризоне и восходит к эпохе ранней юры (с 199,6 до 175 миллионов лет назад), что делает просалирус несколько более недавним, чем Triadobatrachus . [ 34 ] Как и последнее, прозалирус не имел значительно увеличенных ног, но имел типичную трехстороннюю тазовую структуру современных лягушек. В отличие от Triadobatrachus , Prosalirus уже потерял почти весь свой хвост [ 35 ] и был хорошо адаптирован для прыжков. [ 36 ] Еще одна ранняя юрская лягушка - Vieraella herbsti , которая известна только по дорсальным и вентральным впечатлениям одного животного и оценивалась в 33 мм ( 1 + 1 ~ 4 дюйма) от морды до вентиляции. Notobatrachus Degiustoi из средней юры немного моложе, около 155–170 миллионов лет. Основные эволюционные изменения в этом виде включали сокращение тела и потерю хвоста. Эволюция современной Ануры, вероятно, была завершена в юрском периоде. С тех пор эволюционные изменения в количестве хромосом происходили примерно в 20 раз быстрее у млекопитающих, чем у лягушек, что означает, что видообразование происходит быстрее у млекопитающих. [ 37 ]
Согласно генетическим исследованиям, семейства Hyloidea , Microhylidae и Clade Natatanura (составляющие около 88% живых лягушек) диверсифицировались одновременно около 66 миллионов лет назад, вскоре после того, как событие вымирания мела -палеогена, связанное с ударом Chicxulub . Все происхождение древесности (например, в Hyloidea и Natatanura) следуют с того времени и возрождения леса, которое произошло впоследствии. [ 38 ] [ 39 ]
Окаменелости лягушки были найдены на всех континентах Земли. [ 40 ] [ 41 ] на острове Сеймур была обнаружена 40 миллионов лет. на В 2020 году команда позвоночных на острове Сеймур на острове Сеймур острове Сеймур на острове Сеймур на острове Сеймур на острове Сеймур на Сеймур острове Полем [ 42 ]
Филогения
Кладограмма , показывающая отношения различных семейств лягушек в кладской ануре, можно увидеть в таблице ниже. Эта диаграмма в виде дерева показывает, как каждая семья лягушек связана с другими семьями, причем каждый узел представляет точку общего происхождения. Он основан на Frost et al. (2006), [ 43 ] Heinicke et al. (2009) [ 44 ] и Pyron and Wiens (2011). [ 45 ]
| Приносит | |
Морфология и физиология

Лягушки не имеют хвоста, за исключением того, что личинки, и большинство имеют длинные задние лапы, удлиненные кости лодыжки, переносные пальцы, когти, большие глаза и гладкая или бородавная кожа. У них короткие позвоночные колонны, с не более 10 свободными позвонками и плавлеными хвостовыми костями (уростил или копчик). [ 46 ] Размер лягушек варьируется от Paedophryne Amauensis из Папуа -Новой Гвинеи , который составляет 7,7 мм (0,30 дюйма) в длине [ 47 ] до примерно 35 см (14 дюймов) и 3,3 кг (7,3 фунта) лягушки Голиафа ( Конрауа Голиаф ) Центральной Африки. [ 48 ] Есть доисторические, вымершие виды, которые достигли еще больших размеров. [ 49 ]
Ноги и ноги
Структура ног и ног сильно различается среди видов лягушек, частично в зависимости от того, живут ли они в основном на земле, в воде, на деревьях или в норах. У взрослых ануранов четыре пальца на руках и пять пальцев ног на ногах, [ 50 ] Но самые маленькие виды часто имеют руки и ноги, где некоторые цифры являются рудиментарными. [ 51 ] Лягушки должны быть в состоянии быстро перемещаться по окружающей среде, чтобы поймать добычу и избежать хищников, и многочисленные адаптации помогают им сделать это. Большинство лягушек либо опытны в прыжках, либо происходят от предков, которые были, причем большая часть мышечной морфологии, модифицированной для этой цели. Цвета, малоберцовая кость и плавники были слиты в одну сильную кость , как и радиус и локтевая кость в передних конечностях (которые должны поглощать воздействие на посадку). Сночевистые плюсневые костюмы стали удлиненными, чтобы добавить к длине ноги и позволить лягушкам толкаться к земле в течение более длительного периода при взлете. удлиненный Подвздошной подвздошной кости и сформировал мобильный соединение с крестцом , который, в таких специализированных прыгунах, как раниды и гилиды, функционирует как дополнительное соединение конечностей для дальнейшей питания скачков. Хвостовые позвонки слились в уростиле, который втягивается внутри таза. Это позволяет перенести силы от ног в тело во время прыжка. [ 46 ]

( Временная лягушка )

Мышечная система была аналогично изменена. Задние конечности наследственных лягушек, по -видимому, содержали пары мышц, которые действуют в оппозиции (одна мышца для сгибания колена, другая мышца, чтобы расширить его), как это видно у большинства других животных с нитьми. Однако у современных лягушек почти все мышцы были изменены, чтобы способствовать действию прыжков, причем осталось всего несколько небольших мышц, чтобы вернуть конечность в начальную позицию и поддерживать осанку. Мышцы также были значительно увеличены, причем мышцы основных ног составляют более 17% от общей массы лягушек. [ 52 ]
Многие лягушки имеют перепончатые ноги, а степень лямки прямо пропорциональна количеству времени, которое виды тратят в воде. [ 53 ] Полностью водная африканская лягушка ( Hymenochirus sp.) Имеет полностью переносные пальцы ног, тогда как у белой лягушки деревьев ( Litoria Caerulea ), древесной виды, составляют всего четверть или половину перепонки. [ 54 ] Исключения включают летающие лягушки в Hylidae и Rhacophoridae , которые также имеют полностью переносные носки, используемые в скольжении.
Лягушки -леса имеют прокладки, расположенные на концах пальцев ног, чтобы помочь захватить вертикальные поверхности. Это не всасывающие прокладки, поверхность, состоящая вместо столбчатых ячеек с плоскими верхушками с небольшими зазорами между ними, смазанными слизистыми железами. Когда лягушка применяет давление, клетки прилипают к нарушениям на поверхности, а рукоятка поддерживается через поверхностное натяжение . Это позволяет лягушке подниматься на гладкие поверхности, но система не работает эффективно, когда прокладки чрезмерно влажные. [ 55 ]
Во многих древесных лягушках небольшая «интеркалярная структура» на каждом носке увеличивает площадь поверхности, касающуюся подложки . Кроме того, многие древесные лягушки имеют тазобедренные суставы, которые позволяют прыгать и ходить. Некоторые лягушки, которые живут высоко на деревьях, даже обладают сложной степенью лямки между ногами. Это позволяет лягушкам «парашют» или создавать контролируемый скольжение из одной позиции в навесе к другому. [ 56 ]
Лягушки из земли, как правило, не имеют адаптации водных и древесных лягушек. У большинства есть меньшие накладки на пальцы ноги, если таковые имеются, и небольшая лямка. У некоторых ограбления лягушек, таких как Spadefoot Couch ( Scapphiopus couchii ), имеют разгибание ноги, похожее на лоскут на задних ногах, кератинизированный бугорок , который часто называют лопатой, которая помогает им зарываться. [ 57 ]
Иногда во время стадии головастика одна из развивающихся задних ног съедается хищником, такой как нимфа дракона . В некоторых случаях полная нога все еще растет, но в других это не так, хотя лягушка может все равно прожить свою нормальную продолжительность жизни только с тремя конечностями. Время от времени паразитный плоский червя ( рибейроя ondatrae ) выкапывает в задней части головастика, вызывая перестройку клеток зачатка конечностей, а лягушка развивает одну или несколько дополнительных ног. [ 58 ]

Кожа
Кожа лягушки защищает, обладает дыхательной функцией, может поглощать воду и помогает контролировать температуру тела. У него много желез, особенно на голове и спине, которые часто источают неприятные и токсичные вещества ( гранулированные железы ). Секреция часто бывает липкой и помогает сохранять кожу влажной, защищает от въезда плесени и бактерий, делает животное скользким и способным убежать от хищников. [ 59 ] Кожа проливается каждые несколько недель. Обычно он снимается в середине спины и через живот, а лягушка тянет руки и ноги свободными. Затем кожа с разбитой обрабатывается к голове, где ее быстро едят. [ 60 ]
Будучи холодным, лягушки должны принять подходящие модели поведения, чтобы регулировать их температуру. Чтобы согреться, они могут двигаться на солнце или на теплую поверхность; Если они перегреваются, они могут перейти в тень или принять позицию, которая обнажает минимальную область кожи в воздух. Эта поза также используется для предотвращения потери воды и включает в себя приседание лягушки вблизи подложки с руками и ногами, спрятанными под подбородком и телом. [ 61 ] Цвет кожи лягушки используется для терморегуляции. В прохладных влажных условиях цвет будет темнее, чем в жаркий сухой день. Серый пенойкий дерево лягуша ( Chiromantis xerampelina ) даже способна превратиться в белый, чтобы минимизировать вероятность перегрева. [ 62 ]
Многие лягушки способны поглощать воду и кислород непосредственно через кожу, особенно вокруг области таза, но проницаемость кожи лягушки также может привести к потере воды. Железы, расположенные по всему телу, источают слизь, которая помогает сохранить кожу влажной и уменьшает испарение. Некоторые железы на руках и сундуках мужчин специализируются на создании липких выделителей, чтобы помочь в Amplexus . Подобные железы в лягушках деревьев дают клейкоподобное вещество на клейких дисках ног. Некоторые древесные лягушки уменьшают потерю воды, имея водонепроницаемый слой кожи, и несколько южноамериканских видов покрывают их кожу восковой секрецией. Другие лягушки приняли поведение для сохранения воды, в том числе становясь ночными и покоящимися в положении, занимающейся водой. Некоторые лягушки также могут отдыхать в больших группах, причем каждая лягушка прижимает его соседей. Это уменьшает количество кожи, подверженной воздействию воздуха или сухой поверхности, и, таким образом, уменьшает потерю воды. [ 61 ] Жаба Вудхауса ( Bufo Woodhousii ), если получить доступ к воде после удержания в сухом месте, находится на мелководье для регидратации. [ 63 ] У мужской волосатой лягушки ( Trichobatrachus robustus ) есть кожные соски, выступающие из ее нижней части спины и бедер, что придает ей плотно внешний вид. Они содержат кровеносные сосуды и, как полагают, увеличивают площадь кожи, доступную для дыхания. [ 64 ]


У некоторых видов есть костиные пластины, встроенные в кожу, черта, которая, по -видимому, развивалась независимо несколько раз. [ 65 ] У некоторых других видов кожа в верхней части головы уплотняется, а соединительная ткань дермы коофимируется с костями черепа ( экзостоз ). [ 66 ] [ 67 ]
Камуфляж является общим защитным механизмом у лягушек. Такие особенности, как бородавки и кожные складки, обычно находятся на земляных лягушках, для которых гладкая кожа не обеспечит такой эффективный камуфляж. Некоторые лягушки меняют цвет между ночью и ночью, так как свет и влажность стимулируют пигментные клетки и заставляют их расширять или сокращаться. [ 68 ] Некоторые даже способны контролировать текстуру своей кожи. [ 69 ] Тихоокеанская лягушка ( Pseudacris Regilla ) имеет зеленые и коричневые морфы, простые или пятнистые, и меняет цвет в зависимости от времени года и общего цвета фона. [ 70 ] Деревянная лягушка ( Lithobates sylvaticus ) использует разрушительную окраску, включая маркировку черных глаз, похожие на пустоты между листьями, полосы дорсальной кожи (дорсолатеральная дермальная плика), похожие на среднюю заклинания листьев , а также пятна, пятна и полосы ног, сходные с особенностями листьев.
Дыхание и кровообращение
Как и другие амфибии, кислород может проходить через их очень проницаемые шкуры. Эта уникальная функция позволяет им оставаться в местах без доступа к воздуху, дыхал через шкуры. Ребра, как правило, отсутствуют, поэтому легкие заполняются прокачкой из щека , а лягушка, лишенная легких, может поддерживать функции его организма без них. [ 68 ] Полностью лягушка с плоской головой водного борнина ( Barbourula kalimantanensis ) является первой лягушкой, которая, как известно, полностью не хватает легких. [ 71 ]
Лягушки имеют трехчастотные сердца , функция, которую они делятся с ящерицами . Кислотная кровь из легких и деоксигенированная кровь из дыхательных тканей попадает в сердце через отдельную предсердие . Когда эти камеры сокращаются, два крови проходят в общий желудочек , прежде чем перекачивать спиральный клапан в соответствующий сосуд, аорту для оксигенированной крови и легочной артерии для дезоксигенированной крови. [ 72 ]
Некоторые виды лягушки имеют адаптацию, которые позволяют им выжить в воде с дефицитом кислорода. ( Водяная лягушка Titicaca Telmatobius Culeus ) является одним из таких видов и имеет морщинистую кожу, которая увеличивает площадь поверхности для усиления газообмена. Обычно он не использует свои рудиментарные легкие, но иногда ритмично поднимает и понижает свое тело, находясь на ложе озера, чтобы увеличить поток воды вокруг него. [ 73 ]

Пищеварение и экскреция
Лягушки имеют верхнечелюстные зубы вдоль верхней челюсти, которые используются для удержания пищи до ее проглатывания. Эти зубы очень слабые и не могут использоваться для жевания или улавливания и нанесения вреда гибкой добыче. Вместо этого лягушка использует свой липкий, расщелину, чтобы поймать насекомых и другую маленькую добычу. Язык обычно лежит на спину во рту, свободный сзади и прикреплен к нижней челюсти спереди. Его можно выстрелить и отозвать с большой скоростью. [ 53 ] У амфибий на языке есть спасательные железы, которые у лягушек производят так называемую двухфазную вязкоупругую жидкость. При воздействии давления, например, когда язык обертывается вокруг добычи, он становится жидким и покрывает тело добычи. Когда давление падает, оно возвращается в толстое и упругое состояние, что дает языку дополнительную сцепление. [ 74 ] У некоторых лягушек нет языка, и они просто заполняют пищу в рот руками. [ 53 ] Африканская бытка ( Pyxicephalus ), которая охотится на относительно крупных животных, таких как мыши и другие лягушки, имеет костяные проекции в форме конуса, называемые одонтоидными процессами в передней части нижней челюсти, которые функционируют как зубы. [ 16 ] Глаза помогают в глотании пищи, так как их можно втянуть через отверстия в черепе и помочь протолкнуть пищу в горло. [ 53 ] [ 75 ]
Затем пища перемещается через пищевод в желудок, где добавляются пищеварительные ферменты, и оно взлетано. Затем он переходит к тонкой кишке (двенадцатиперстная кишка и подвздошной кишки), где происходит большая часть пищеварения. Сок поджелудочной железы от поджелудочной железы и желчь, производимый печенью и хранящий в желчном пузыре, секретируются в тонкую кишку, где жидкости переваривают пищу, а питательные вещества поглощаются. Остаток пищи проходит в большую кишку, где удаляется избыточная вода, и отходы раздаются через клоаку . [ 76 ]
Несмотря на то, что лягушки адаптированы к наземной жизни, лягушки напоминают пресноводную рыбу в своей неспособности эффективно сохранять воду для тела. Когда они находятся на суше, много воды теряется из -за испарения с кожи. Экскреторная система аналогична системе млекопитающих, и есть две почки , которые удаляют азотные продукты из крови. Лягушки производят большое количество разбавленной мочи, чтобы вымыть токсичные продукты из почечных канальцев. [ 77 ] Азот выделяется как аммиак головастиками и водными лягушками, но в основном как мочевина , менее токсичный продукт, большинством земных взрослых. Несколько видов лягушки деревьев с небольшим доступом к воде выделяют еще менее токсичную мочевую кислоту . [ 77 ] Моча проходит вдоль парных мочеточников в мочевой пузырь, из которого он периодически выпускается в клоаку. Все телесные отходы выходят из тела через клоаку, которая заканчивается в клоакальном вентиляционном отверстии. [ 78 ]
Репродуктивная система
В мужской лягушке два яичка прикрепляются к почкам, а сперма проходит в почки через тонкие трубки, называемые эфферентными протоками . Затем он путешествует через мочеточники, которые, следовательно, известны как мориногенитальные протоки. Там нет пениса, и сперма выталкивается из клоаки прямо на яйца, когда самка их укладывает. Яичники женской лягушки находятся рядом с почками, а яйца пропускают пару яйцеклеток и через клоаку на внешнюю часть. [ 78 ]
Когда лягушки спариваются, мужчина поднимается на заднюю часть самки и обнимает передние конечности вокруг ее тела, либо за передними ногами, либо прямо перед задними ногами. Эта позиция называется Amplexus и может проводиться в течение нескольких дней. [ 79 ] Мужская лягушка имеет определенные гормонозависимые вторичные сексуальные характеристики . К ним относятся разработка специальных прокладков на его больших пальцах в сезон размножения, чтобы дать ему твердую тщательную ткань. [ 80 ] Захват мужской лягушки во время Amplexus стимулирует самку выбросить яйца, обычно завернутые в желе, как нерести. У многих видов мужчина меньше и стройнее, чем самка. У мужчин есть голосовые связки и делают ряд рассыпений, особенно в сезон размножения, а у некоторых видов у них также есть голосовые мешки для усиления звука. [ 78 ]
Нервная система
Лягушки имеют высокоэффективную нервную систему, которая состоит из мозга, спинного мозга и нервов. Многие части мозга лягушки соответствуют частям людей. Он состоит из двух обонятельных долей, двух полушарий головного мозга, корпуса шишковидной мышцы, двух зрительных лепестков, мозжечка и продолговатого мозга. Мышечная координация и поза контролируются мозжечком , а медовая продолговатая регулировала дыхание, пищеварение и другие автоматические функции. Относительный размер головного мозга у лягушек намного меньше, чем у людей. Лягушки имеют десять пар черепных нервов , которые передают информацию снаружи непосредственно к мозгу, и десять пар спинных нервов , которые передают информацию из конечностей к мозгу через спинной мозг. [ 78 ] В отличие от этого, все амниоты (млекопитающие, птицы и рептилии) имеют двенадцать паров черепных нервов. [ 81 ]

Взгляд
Глаза большинства лягушек расположены по обе стороны головы рядом с вершиной и проецируются наружу в виде полусферических выпуклостей. Они обеспечивают бинокулярное зрение на поле 100 ° к передней части и общему визуальному полю почти на 360 °. [ 82 ] Они могут быть единственной частью погруженной лягушки, которая выступает из воды. Каждый глаз имеет приблизительную верхнюю и нижнюю крышку и никитирующую мембрану , которая обеспечивает дальнейшую защиту, особенно когда лягушка плавает. [ 83 ] Члены Aquatic Family Pipidae имеют глаза, расположенные в верхней части головы, положение, лучше подходящее для обнаружения добычи в воде выше. [ 82 ] Ирисы . бывают разных цветов и учеников в различных формах Общая жаба ( Bufo Bufo ) имеет золотые ирисы и горизонтальные зрачки, похожие на щели, у лягушки с красными глазами ( Agalychnis callidryas ) есть вертикальные зрачки, у ядовитой лягушки с темной ирисами, огнеупорная жаба ( Bombina spp. ) Имеет треугольные зрачки, а томатная лягушка ( Dyscophus spp. ) Имеет круглые. Ирисы южной жабы ( Anaxyrus terrestris ) с рисунком, чтобы смешаться с окружающей замаскированной кожей. [ 83 ]
Отдаленное зрение лягушки лучше, чем его близкое зрение. Призыв к лягушкам быстро замолчают, когда они увидят злоумышленника или даже движущуюся тень, но чем ближе объект, тем менее хорошо он видит. [ 83 ] Когда лягушка стреляет из своего языка, чтобы поймать насекомое, она реагирует на небольшой движущийся объект, который не может видеть хорошо и должен точно выровнять его, потому что он закрывает глаза, когда язык вытянут. [ 53 ] Хотя это раньше обсуждалось, [ 84 ] Более поздние исследования показали, что лягушки могут видеть по цвету даже при очень низком свете. [ 85 ]
Слушание
Лягушки могут слышать как в воздухе, так и под водой. У них нет внешних ушей ; Полосы ( барабанные перепонки ) непосредственно обнажаются или могут быть покрыты слоем кожи и видны как круглая область сразу за глаз. Размер и расстояние друг от друга связаны с частотой и длиной волны, на которой вызывает лягушка. У некоторых видов, таких как uklefrog, размер бакалавра указывает на пол лягушки; У мужчин есть барабанные бани, которые больше, чем у их глаз, в то время как у женщин глаза и тампани имеют почти одинаковый размер. [ 86 ] Шум приводит к вибрированию баймпана, а звук передается на среднее и внутреннее ухо. Среднее ухо содержит полукруглые каналы, которые помогают контролировать баланс и ориентацию. Во внутреннем ухе слуховые волосковые клетки расположены в двух областях улитки, базилярной папиллы и амфибийной папилла. Первые обнаруживают высокие частоты и последние низкие частоты. [ 87 ] Поскольку улитка короткая, лягушки используют электрическую настройку , чтобы расширить свой диапазон слышимых частот и помогать различать различные звуки. [ 88 ] Такое расположение позволяет обнаружить территориальные и размножающие звонки их конспецифических . У некоторых видов, которые населяют засушливые регионы, звук грома или сильного дождя может пробудить их из бездействующего государства. [ 87 ] Лягушка может быть поражена неожиданным шумом, но обычно она не предпринимает никаких действий, пока не найдет источник звука. [ 86 ]
Вызов

Вызов или грип лягушка уникальны для его видов. Лягушки создают этот звук, проходя воздух через гортани в горле. В большинстве призванных лягушек звук усиливается одним или несколькими вокальными мешками, мембранами кожи под горлом или на углу рта, которые растягиваются во время усиления вызова. Некоторые звонки с лягушками настолько громкие, что их можно услышать вплоть до мили (1,6 км). [ 89 ] Кроме того, было обнаружено, что некоторые виды используют искусственные структуры, такие как дренажные трубы для искусственного усиления их вызовов. [ 90 ] Прибрежный хвост лягушка ( Ascaphus truei ) живет в горных ручьях в Северной Америке и не озвучивает. [ 91 ]
Основная функция вызова - для мужских лягушек, чтобы привлечь товарищей. Мужчины могут вызывать индивидуально, или может быть хор звука, когда многие мужчины сходились на местах размножения. У многих видов лягушек, таких как общая лягушка дерева ( полипедат Leucomystax ), женщины отвечают на вызовы мужчин, которые действуют для усиления репродуктивной активности в колонии размножения. [ 92 ] Женские лягушки предпочитают мужчин, которые вызывают звуки большей интенсивности и более низкой частоты, атрибуты, которые выделяются в толпе. Считается, что обоснование этого заключается в том, что, демонстрируя его мастерство, мужчина показывает свою пригодность для создания превосходного потомства. [ 93 ]
Другой вызов излучается мужской лягушкой или невосприимчивой женщиной, когда он устанавливается другим мужчиной. Это отчетливый звук, который сопровождается вибрацией тела. [ 94 ] Лягушки деревьев и некоторые неакватические виды имеют дождь, который они делают на основе сигналов влажности перед душем. [ 94 ] Многие виды также имеют территориальный призыв, который используется для отъезда других мужчин. Все эти вызовы излучаются с закрытым ртом лягушки. [ 94 ] Призыв к бедствию, излучаемый некоторыми лягушками, когда они находятся в опасности, производится с открытым ртом, что приводит к более высокому вызову. Обычно он используется, когда лягушка была захвачена хищником и может служить для отвлечения или дезориентирования злоумышленника, чтобы она высвобождала лягушку. [ 94 ]
Многие виды лягушки имеют глубокие звонки. Кроак американского быка ( Рана Кейтсбиана ) иногда пишутся как «кувшин». [ 95 ] Тихоокеанская лягушка ( Pseudacris Regilla ) производит ономатопоэки «ribbit», часто слышащий в фильмах. [ 96 ] Другие визуализации лягушки в речь включают «Brekekekex Koax Koax», зов болотной лягушки ( пелофилакс -Ridibundus ) в лягушках , древнегреческой комической драме Аристофана . [ 97 ] Призывы к вогнутой торрент-лягушке ( Amolops Tormotus ) необычны во многих аспектах. Мужчины известны своими разновидностями звонков, где происходят модуляции вверх и нисходящего частоты. Когда они общаются, они производят звонки, которые падают в диапазоне ультразвуковых частот. Последний аспект, который делает этот вид звонков лягушки необычным, заключается в том, что нелинейные акустические явления являются важными компонентами в их акустических сигналах. [ 98 ]
Торпор
В экстремальных условиях некоторые лягушки попадают в состояние ощущения и остаются неактивными в течение нескольких месяцев. В более холодных регионах многие виды лягушки зимой зимой. Те, кто живет на земле, такой как американская жаба ( Bufo Americanus ), выкапывают нору и делают гибернакулум , в котором можно лежать бездействующим . Другие, менее опытные в копании, находят расщелину или похоронить себя в мертвых листьях. Водные виды, такие как American Bullfrog ( Rana Catesbeiana ), обычно погружаются на дно пруда, где они лежат, полуммируют в грязи, но все еще способны получить доступ к кислороду, растворенному в воде. Их метаболизм замедляется, и они живут в своих энергетических резервах. Некоторые лягушки, такие как деревянная лягушка , майр -лягушка или весенний пипер, могут даже выжить, будучи замороженными. Кристаллы льда образуются под кожей и в полости тела, но основные органы защищены от замораживания при высокой концентрации глюкозы. По -видимому, безжизненная, замороженная лягушка может возобновить дыхание, и ее сердцебиение может перезагружаться, когда условия нагреваются. [ 99 ]
С другой стороны, полосатая лягушка из Беррурующей ( циклорана alboguttata ) регулярно эститирует в жаркий, сухой сезон в Австралии, выживая в состоянии покоя без доступа к еде и воде в течение девяти или десяти месяцев в году. Он зарывается под землей и скручивается в защитном коконе , образованном его сарайкой. Исследователи из Университета Квинсленда обнаружили, что во время эстиции метаболизм лягушки изменяется, а эксплуатационная эффективность митохондрий увеличивается . Это означает, что ограниченное количество энергии, доступной для коматозной лягушки, используется более эффективным образом. Этот механизм выживания полезен только для животных, которые остаются совершенно бессознательными в течение длительного периода времени, и чьи потребности в энергии низкие, потому что они хладнокровны и не нуждаются в генерации тепла. [ 100 ] Другие исследования показали, что, чтобы обеспечить эти энергетические требования, атрофию мышц, но мышцы задней конечности преимущественно не затронуты. [ 101 ] Было обнаружено, что лягушки имеют верхнюю критическую температуру около 41 градуса по Цельсию. [ 102 ]
Локомоция
Различные виды лягушки используют ряд методов передвижения, в том числе прыжки , бег , ходьбу , плавание , смягчение , скалолазание и скольжение .

- Прыжок
Лягушки, как правило, признаются исключительными прыгунами и относительно их размера лучшими прыгунами всех позвоночных. [ 103 ] Полосатая ракетная лягушка , Litoria Nasuta , может перепрыгнуть two metres (6+1⁄2 feet), a distance that is more than fifty times its body length of 55 мм ( 2 + 1 ~ 4 дюйма). [ 104 ] Существуют огромные различия между видами в способности прыгать. Внутри вида расстояние прыжков увеличивается с увеличением размера, но относительное расстояние прыжков (длины тела прыгают) уменьшается. Индийская лягушка шкипера ( euphlyctis cyanophlyctis ) обладает способностью выходить из воды из положения, плавающего на поверхности. [ 105 ] Крошечная северная лягушка для крикета ( крипитаны Acris ) может «перепрыгивать» через поверхность пруда с помощью ряда коротких быстрых прыжков. [ 106 ]
Медленная фотография показывает, что мышцы обладают пассивной гибкостью. Сначала они растянуты, пока лягушка еще находится в приседании, затем они сжимаются, прежде чем снова растягиваются, чтобы запустить лягушку в воздух. Передние ножки сложены на груди, а задние ножки остаются в расширенном, обтекаемом положении на протяжении всего прыжка. [ 52 ] В некоторых чрезвычайно способных прыгунах, таких как кубинская лягушка ( Osteopilus septentrionalis ) и северная лягушка леопарда ( Rana Pipiens ), пиковая мощность, оказываемая во время прыжка, может превышать то, что мышца теоретически способна. Когда мышцы сокращаются, энергия сначала переносится в растянутое сухожилие, которое обернута вокруг кости лодыжки. Затем мышцы снова растягиваются в то же время, когда сухожилие высвобождают свою энергию, как катапульту, чтобы вызвать мощное ускорение за пределами ускорения мышц. [ 107 ] Аналогичный механизм был задокументирован в саранчи и кузнечиках . [ 108 ]
Раннее вылупление лягушек может оказывать негативное влияние на производительность прыжка с лягушки и общую локомоцию. [ 109 ] Задние конечности не могут полностью формироваться, что приводит к тому, что они будут короче и намного слабее по сравнению с нормальным фроглетом. [ 109 ] Ранние выводы лягушек могут чаще зависеть от других форм передвижения, таких как плавание и ходьба. [ 109 ]
- Ходить и бегать

Лягушки в семьях Bufonidae, Rhinophrynidae и Microhylidae имеют короткие задние ноги и имеют тенденцию ходить, а не прыгать. [ 110 ] Когда они пытаются быстро двигаться, они ускоряют скорость движения своих конечностей или прибегают к неуклюже прыгающей походке. Великие равнины с узким ртом ( Gastrophryne Olivacea ) описывалась как имеющая походку, которая является «комбинацией бега и коротких хмелей, которые обычно имеют длину всего лишь дюйм или два». [ 111 ] В эксперименте жаба Фаулера ( Bufo Fowleri ) была помещена на беговую дорожку, которая была повернута на различных скоростях. Измеряя поглощение кислорода жабы, было обнаружено, что прыжки представляют собой неэффективное использование ресурсов во время устойчивого локомоции, но было полезной стратегией во время коротких всплесков высокой интенсивности. [ 112 ]
Ружанная лягушка ( Kassina maculata ) имеет короткие тонкие задних конечностей, не подходящих для прыжков. Он может двигаться быстро, используя бегущую походку, в которой две задние лапы используются попеременно. Медленная фотография показывает, что, в отличие от лошади, которая может вырваться или галоп, походка лягушки оставалась одинаковой на медленных, средних и быстрых скоростях. [ 113 ] Этот вид также может подниматься на деревья и кустарники и делает это ночью, чтобы поймать насекомых. [ 114 ] Индийская лягушка шкипера ( euphlyctis cyanophlyctis ) имеет широкие ноги и может проходить через поверхность воды в течение нескольких метров (ярды). [ 106 ]
- Плавание

Лягушки, которые живут или посещают воду, имеют адаптацию, которые улучшают их способности плавания. Задние конечности сильно мускулистые и сильные. Посимость между пальцами задних ног увеличивает площадь ноги и помогает мощно продвигать лягушку через воду. Члены семьи Pipidae полностью водные и показывают наиболее заметную специализацию. У них есть негибкие позвоночные колонны, сплющенные, обтекаемые тела, системы боковых линий и мощные задние конечности с большими перепончатыми ногами. [ 115 ] Головастики в основном имеют большие хвостовые плавники, которые обеспечивают тягу, когда хвост перемещается из стороны в сторону. [ 116 ]
- Ограбление
Некоторые лягушки стали адаптированными для обрывания и жизни под землей. У них, как правило, есть округлые тела, короткие конечности, маленькие головы с выпуклыми глазами и задние ноги, адаптированные для раскопок. Чрезвычайным примером этого является фиолетовая лягушка ( Nasikabatrachus sahyadrensis ) из южной Индии, которая питается термитами и проводит почти всю свою жизнь под землей. Это происходит кратко во время муссона , чтобы спариваться и размножаться во временных бассейнах. Он имеет крошечную голову с заостренной мордой и пухлым, округлым телом. Из -за этого искоренного существования оно было впервые описано в 2003 году, будучи новым в научном сообществе в то время, хотя ранее известно местным жителям. [ 117 ]

Жалы из северной Америки также адаптированы к подземной жизни. Жажа Spadefoot Plains ( Spea Bombifrons ) типична и имеет лоскут кератинизированной кости, прикрепленной к одному из плюсневых ног, которые он использует, чтобы выкопать себя назад в землю. Когда он копает, жаба падает бедра из стороны в сторону, чтобы опуститься в свободную почву. Летом у него неглубокая нора, из которой он выходит ночью на корм. Зимой он копает гораздо глубже и был записан на глубине 4,5 м (14 футов 9 дюймов). [ 118 ] Туннель заполнен почвой, а жаба спят в небольшой камере в конце. В течение этого времени мочевина накапливается в его тканях, а вода тянутся из окружающей влажной почвы путем осмоса для удовлетворения потребностей жабы. [ 118 ] Жалы Spadefoot - это «взрывные заводчики», все они выходят из их норков одновременно и сходятся на временных бассейнах, привлеченных к одному из них при призвании первого мужчины, чтобы найти подходящее место для размножения. [ 119 ]
The burrowing frogs of Australia have a rather different lifestyle. The western spotted frog (Heleioporus albopunctatus) digs a burrow beside a river or in the bed of an ephemeral stream and regularly emerges to forage. Mating takes place and eggs are laid in a foam nest inside the burrow. The eggs partially develop there, but do not hatch until they are submerged following heavy rainfall. The tadpoles then swim out into the open water and rapidly complete their development.[120] Madagascan burrowing frogs are less fossorial and mostly bury themselves in leaf litter. One of these, the green burrowing frog (Scaphiophryne marmorata), has a flattened head with a short snout and well-developed metatarsal tubercles on its hind feet to help with excavation. It also has greatly enlarged terminal discs on its fore feet that help it to clamber around in bushes.[121] It breeds in temporary pools that form after rains.[122]
- Climbing


Tree frogs live high in the canopy, where they scramble around on the branches, twigs, and leaves, sometimes never coming down to earth. The "true" tree frogs belong to the family Hylidae, but members of other frog families have independently adopted an arboreal habit, a case of convergent evolution. These include the glass frogs (Centrolenidae), the bush frogs (Hyperoliidae), some of the narrow-mouthed frogs (Microhylidae), and the shrub frogs (Rhacophoridae).[110] Most tree frogs are under 10 cm (4 in) in length, with long legs and long toes with adhesive pads on the tips. The surface of the toe pads is formed from a closely packed layer of flat-topped, hexagonal epidermal cells separated by grooves into which glands secrete mucus. These toe pads, moistened by the mucus, provide the grip on any wet or dry surface, including glass. The forces involved include boundary friction of the toe pad epidermis on the surface and also surface tension and viscosity.[123] Tree frogs are very acrobatic and can catch insects while hanging by one toe from a twig or clutching onto the blade of a windswept reed.[124] Some members of the subfamily Phyllomedusinae have opposable toes on their feet. The reticulated leaf frog (Phyllomedusa ayeaye) has a single opposed digit on each fore foot and two opposed digits on its hind feet. This allows it to grasp the stems of bushes as it clambers around in its riverside habitat.[125]
- Gliding
During the evolutionary history of frogs, several different groups have independently taken to the air.[126] Some frogs in the tropical rainforest are specially adapted for gliding from tree to tree or parachuting to the forest floor. Typical of them is Wallace's flying frog (Rhacophorus nigropalmatus) from Malaysia and Borneo. It has large feet with the fingertips expanded into flat adhesive discs and the digits fully webbed. Flaps of skin occur on the lateral margins of the limbs and across the tail region. With the digits splayed, the limbs outstretched, and these flaps spread, it can glide considerable distances, but is unable to undertake powered flight.[127] It can alter its direction of travel and navigate distances of up to 15 m (50 ft) between trees.[128]
Life history

(Rana clamitans)
Reproduction
Two main types of reproduction occur in frogs, prolonged breeding and explosive breeding. In the former, adopted by the majority of species, adult frogs at certain times of year assemble at a pond, lake or stream to breed. Many frogs return to the bodies of water in which they developed as larvae. This often results in annual migrations involving thousands of individuals. In explosive breeders, mature adult frogs arrive at breeding sites in response to certain trigger factors such as rainfall occurring in an arid area. In these frogs, mating and spawning take place promptly and the speed of larval growth is rapid in order to make use of the ephemeral pools before they dry up.[129]
Among prolonged breeders, males usually arrive at the breeding site first and remain there for some time whereas females tend to arrive later and depart soon after they have spawned. This means that males outnumber females at the water's edge and defend territories from which they expel other males. They advertise their presence by calling, often alternating their croaks with neighbouring frogs. Larger, stronger males tend to have deeper calls and maintain higher quality territories. Females select their mates at least partly on the basis of the depth of their voice.[130] In some species there are satellite males who have no territory and do not call. They may intercept females that are approaching a calling male or take over a vacated territory. Calling is an energy-sapping activity. Sometimes the two roles are reversed and a calling male gives up its territory and becomes a satellite.[129]

In explosive breeders, the first male that finds a suitable breeding location, such as a temporary pool, calls loudly and other frogs of both sexes converge on the pool. Explosive breeders tend to call in unison creating a chorus that can be heard from far away. The spadefoot toads (Scaphiopus spp.) of North America fall into this category. Mate selection and courtship is not as important as speed in reproduction. In some years, suitable conditions may not occur and the frogs may go for two or more years without breeding.[129] Some female New Mexico spadefoot toads (Spea multiplicata) only spawn half of the available eggs at a time, perhaps retaining some in case a better reproductive opportunity arises later.[131]
At the breeding site, the male mounts the female and grips her tightly round the body. Typically, amplexus takes place in the water, the female releases her eggs and the male covers them with sperm; fertilization is external. In many species such as the Great Plains toad (Bufo cognatus), the male restrains the eggs with his back feet, holding them in place for about three minutes.[129] Members of the West African genus Nimbaphrynoides are unique among frogs in that they are viviparous; Limnonectes larvaepartus, Eleutherodactylus jasperi and members of the Tanzanian genus Nectophrynoides are the only frogs known to be ovoviviparous. In these species, fertilization is internal and females give birth to fully developed juvenile frogs, except L. larvaepartus, which give birth to tadpoles.[132][133][134]
Life cycle
Eggs / frogspawn

Frogs may lay their in eggs as clumps, surface films, strings, or individually. Around half of species deposit eggs in water, others lay eggs in vegetation, on the ground or in excavations.[135][136][137] The tiny yellow-striped pygmy eleuth (Eleutherodactylus limbatus) lays eggs singly, burying them in moist soil.[138] The smoky jungle frog (Leptodactylus pentadactylus) makes a nest of foam in a hollow. The eggs hatch when the nest is flooded, or the tadpoles may complete their development in the foam if flooding does not occur.[139] The red-eyed treefrog (Agalychnis callidryas) deposits its eggs on a leaf above a pool and when they hatch, the larvae fall into the water below.[140]
In certain species, such as the wood frog (Rana sylvatica), symbiotic unicellular green algae are present in the gelatinous material. It is thought that these may benefit the developing larvae by providing them with extra oxygen through photosynthesis.[141] The interior of globular egg clusters of the wood frog has also been found to be up to 6 °C (11 °F) warmer than the surrounding water and this speeds up the development of the larvae.[142] The larvae developing in the eggs can detect vibrations caused by nearby predatory wasps or snakes, and will hatch early to avoid being eaten.[143] In general, the length of the egg stage depends on the species and the environmental conditions. Aquatic eggs normally hatch within one week when the capsule splits as a result of enzymes released by the developing larvae.[144]
Direct development, where eggs hatch into juveniles like small adults, is also known in many frogs, for example, Ischnocnema henselii,[145] Eleutherodactylus coqui,[146] and Raorchestes ochlandrae and Raorchestes chalazodes.[147]
Tadpoles

The larvae that emerge from the eggs are known as tadpoles (or occasionally polliwogs). Tadpoles lack eyelids and limbs, and have cartilaginous skeletons, gills for respiration (external gills at first, internal gills later), and tails they use for swimming.[116] As a general rule, free-living larvae are fully aquatic, but at least one species (Nannophrys ceylonensis) has semiterrestrial tadpoles which live among wet rocks.[148][149]
From early in its development, a gill pouch covers the tadpole's gills and front legs. The lungs soon start to develop and are used as an accessory breathing organ. Some species go through metamorphosis while still inside the egg and hatch directly into small frogs. Tadpoles lack true teeth, but the jaws in most species have two elongated, parallel rows of small, keratinized structures called keradonts in their upper jaws. Their lower jaws usually have three rows of keradonts surrounded by a horny beak, but the number of rows can vary and the exact arrangements of mouth parts provide a means for species identification.[144] In the Pipidae, with the exception of Hymenochirus, the tadpoles have paired anterior barbels, which make them resemble small catfish.[115] Their tails are stiffened by a notochord, but does not contain any bony or cartilaginous elements except for a few vertebrae at the base which forms the urostyle during metamorphosis. This has been suggested as an adaptation to their lifestyles; because the transformation into frogs happens very fast, the tail is made of soft tissue only, as bone and cartilage take a much longer time to be broken down and absorbed. The tail fin and tip is fragile and will easily tear, which is seen as an adaptation to escape from predators which try to grasp them by the tail.[150]
Tadpoles are typically herbivorous, feeding mostly on algae, including diatoms filtered from the water through the gills. Some species are carnivorous at the tadpole stage, eating insects, smaller tadpoles, and fish. The Cuban tree frog (Osteopilus septentrionalis) is one of a number of species in which the tadpoles can be cannibalistic. Tadpoles that develop legs early may be eaten by the others, so late developers may have better long-term survival prospects.[151]
Tadpoles are highly vulnerable to being eaten by fish, newts, predatory diving beetles, and birds, particularly water birds, such as storks and herons and domestic ducks. Some tadpoles, including those of the cane toad (Rhinella marina), are poisonous. The tadpole stage may be as short as a week in explosive breeders or it may last through one or more winters followed by metamorphosis in the spring.[152]
Metamorphosis
At the end of the tadpole stage, a frog undergoes metamorphosis in which its body makes a sudden transition into the adult form. This metamorphosis typically lasts only 24 hours, and is initiated by production of the hormone thyroxine. This causes different tissues to develop in different ways. The principal changes that take place include the development of the lungs and the disappearance of the gills and gill pouch, making the front legs visible. The lower jaw transforms into the big mandible of the carnivorous adult, and the long, spiral gut of the herbivorous tadpole is replaced by the typical short gut of a predator.[144] Homeostatic feedback control of food intake is largely absent, making tadpoles eat constantly when food is present. But shortly before and during metamorphosis the sensation of hunger is suppressed, and they stop eating while their gut and internal organs are reorganized and prepared for a different diet.[153][154] Also the gut microbiota changes, from being similar to that of fish to resembling that of amniotes.[155] Exceptions are carnivorous tadpoles like Lepidobatrachus laevis, which has a gut already adapted to a diet similar to that of adults. These continue to eat during metamorphosis.[156] The nervous system becomes adapted for hearing and stereoscopic vision, and for new methods of locomotion and feeding.[144] The eyes are repositioned higher up on the head and the eyelids and associated glands are formed. The eardrum, middle ear, and inner ear are developed. The skin becomes thicker and tougher, the lateral line system is lost, and skin glands are developed.[144] The final stage is the disappearance of the tail, but this takes place rather later, the tissue being used to produce a spurt of growth in the limbs.[157] Frogs are at their most vulnerable to predators when they are undergoing metamorphosis. At this time, the tail is being lost and locomotion by means of limbs is only just becoming established.[110]
-
 Larva of the common frog Rana temporaria a day before metamorphosis
Larva of the common frog Rana temporaria a day before metamorphosis -
 Metamorphosis stage with deforming jaws, large eyes, and remains of gill pouch
Metamorphosis stage with deforming jaws, large eyes, and remains of gill pouch -
 Young frog with a stumpy tail, metamorphosis nearly complete
Young frog with a stumpy tail, metamorphosis nearly complete



Adults
Adult frogs may live in or near water, but few are fully aquatic.[158] Almost all frog species are carnivorous as adults, preying on invertebrates, including insects, crabs, spiders, mites, worms, snails, and slugs. A few of the larger ones may eat other frogs, small mammals and reptiles, and fish.[159][160] A few species also eat plant matter; the tree frog Xenohyla truncata is partly herbivorous, its diet including a large proportion of fruit, floral structures and nectar.[161][162] Leptodactylus mystaceus has been found to eat plants,[163][164] and folivory occurs in Euphlyctis hexadactylus, with plants constituting 79.5% of its diet by volume.[165] Many frogs use their sticky tongues to catch prey, while others simply grab them with their mouths.[166] Adult frogs are themselves attacked by many predators. The northern leopard frog (Rana pipiens) is eaten by herons, hawks, fish, large salamanders, snakes, raccoons, skunks, mink, bullfrogs, and other animals.[167]
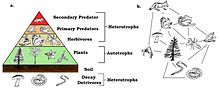
Frogs are primary predators and an important part of the food web. Being cold-blooded, they make efficient use of the food they eat with little energy being used for metabolic processes, while the rest is transformed into biomass. They are themselves eaten by secondary predators and are the primary terrestrial consumers of invertebrates, most of which feed on plants. By reducing herbivory, they play a part in increasing the growth of plants and are thus part of a delicately balanced ecosystem.[168]
Little is known about the longevity of frogs and toads in the wild, but some can live for many years. Skeletochronology is a method of examining bones to determine age. Using this method, the ages of mountain yellow-legged frogs (Rana muscosa) were studied, the phalanges of the toes showing seasonal lines where growth slows in winter. The oldest frogs had ten bands, so their age was believed to be 14 years, including the four-year tadpole stage.[169] Captive frogs and toads have been recorded as living for up to 40 years, an age achieved by a European common toad (Bufo bufo). The cane toad (Rhinella marina) has been known to survive 24 years in captivity, and the American bullfrog (Rana catesbeiana) 14 years.[170] Frogs from temperate climates hibernate during the winter, and four species are known to be able to withstand freezing during this time, including the wood frog (Rana sylvatica).[171]
Parental care


Although care of offspring is poorly understood in frogs, up to an estimated 20% of amphibian species may care for their young in some way.[172] The evolution of parental care in frogs is driven primarily by the size of the water body in which they breed. Those that breed in smaller water bodies tend to have greater and more complex parental care behaviour.[173] Because predation of eggs and larvae is high in large water bodies, some frog species started to lay their eggs on land. Once this happened, the desiccating terrestrial environment demands that one or both parents keep them moist to ensure their survival.[174] The subsequent need to transport hatched tadpoles to a water body required an even more intense form of parental care.[173]
In small pools, predators are mostly absent and competition between tadpoles becomes the variable that constrains their survival. Certain frog species avoid this competition by making use of smaller phytotelmata (water-filled leaf axils or small woody cavities) as sites for depositing a few tadpoles.[175] While these smaller rearing sites are free from competition, they also lack sufficient nutrients to support a tadpole without parental assistance. Frog species that changed from the use of larger to smaller phytotelmata have evolved a strategy of providing their offspring with nutritive but unfertilized eggs.[173] The female strawberry poison-dart frog (Oophaga pumilio) lays her eggs on the forest floor. The male frog guards them from predation and carries water in his cloaca to keep them moist. When they hatch, the female moves the tadpoles on her back to a water-holding bromeliad or other similar water body, depositing just one in each location. She visits them regularly and feeds them by laying one or two unfertilized eggs in the phytotelma, continuing to do this until the young are large enough to undergo metamorphosis.[176] The granular poison frog (Oophaga granulifera) looks after its tadpoles in a similar way.[177]
Many other diverse forms of parental care are seen in frogs. The tiny male Colostethus subpunctatus stands guard over his egg cluster, laid under a stone or log. When the eggs hatch, he transports the tadpoles on his back to a temporary pool, where he partially immerses himself in the water and one or more tadpoles drop off. He then moves on to another pool.[178] The male common midwife toad (Alytes obstetricans) carries the eggs around with him attached to his hind legs. He keeps them damp in dry weather by immersing himself in a pond, and prevents them from getting too wet in soggy vegetation by raising his hindquarters. After three to six weeks, he travels to a pond and the eggs hatch into tadpoles.[179] The tungara frog (Physalaemus pustulosus) builds a floating nest from foam to protect its eggs from predation. The foam is made from proteins and lectins, and seems to have antimicrobial properties.[180] Several pairs of frogs may form a colonial nest on a previously built raft. The eggs are laid in the centre, followed by alternate layers of foam and eggs, finishing with a foam capping.[181]
Some frogs protect their offspring inside their own bodies. Both male and female pouched frogs (Assa darlingtoni) guard their eggs, which are laid on the ground. When the eggs hatch, the male lubricates his body with the jelly surrounding them and immerses himself in the egg mass. The tadpoles wriggle into skin pouches on his side, where they develop until they metamorphose into juvenile frogs.[182] The female gastric-brooding frog (Rheobatrachus sp.) from Australia, now probably extinct, swallows her fertilized eggs, which then develop inside her stomach. She ceases to feed and stops secreting stomach acid. The tadpoles rely on the yolks of the eggs for nourishment. After six or seven weeks, they are ready for metamorphosis. The mother regurgitates the tiny frogs, which hop away from her mouth.[183] The female Darwin's frog (Rhinoderma darwinii) from Chile lays up to 40 eggs on the ground, where they are guarded by the male. When the tadpoles are about to hatch, they are engulfed by the male, which carries them around inside his much-enlarged vocal sac. Here they are immersed in a frothy, viscous liquid that contains some nourishment to supplement what they obtain from the yolks of the eggs. They remain in the sac for seven to ten weeks before undergoing metamorphosis, after which they move into the male's mouth and emerge.[184]
Defence


At first sight, frogs seem rather defenceless because of their small size, slow movement, thin skin, and lack of defensive structures, such as spines, claws or teeth. Many use camouflage to avoid detection, the skin often being spotted or streaked in neutral colours that allow a stationary frog to merge into its surroundings. Some can make prodigious leaps, often into water, that help them to evade potential attackers, while many have other defensive adaptations and strategies.[129]
The skin of many frogs contains mild toxic substances called bufotoxins to make them unpalatable to potential predators. Most toads and some frogs have large poison glands, the parotoid glands, located on the sides of their heads behind the eyes and other glands elsewhere on their bodies. These glands secrete mucus and a range of toxins that make frogs slippery to hold and distasteful or poisonous. If the noxious effect is immediate, the predator may cease its action and the frog may escape. If the effect develops more slowly, the predator may learn to avoid that species in future.[185] Poisonous frogs tend to advertise their toxicity with bright colours, an adaptive strategy known as aposematism. The poison dart frogs in the family Dendrobatidae do this. They are typically red, orange, or yellow, often with contrasting black markings on their bodies. Allobates zaparo is not poisonous, but mimics the appearance of two different toxic species with which it shares a common range in an effort to deceive predators.[186] Other species, such as the European fire-bellied toad (Bombina bombina), have their warning colour underneath. They "flash" this when attacked, adopting a pose that exposes the vivid colouring on their bellies.[4]

Some frogs, such as the poison dart frogs, are especially toxic. The native peoples of South America extract poison from these frogs to apply to their weapons for hunting,[187] although few species are toxic enough to be used for this purpose. At least two non-poisonous frog species in tropical America (Eleutherodactylus gaigei and Lithodytes lineatus) mimic the colouration of dart poison frogs for self-protection.[188][189] Some frogs obtain poisons from the ants and other arthropods they eat.[190] Others, such as the Australian corroboree frogs (Pseudophryne corroboree and Pseudophryne pengilleyi), can synthesize the alkaloids themselves.[191] The chemicals involved may be irritants, hallucinogens, convulsants, nerve poisons or vasoconstrictors. Many predators of frogs have become adapted to tolerate high levels of these poisons, but other creatures, including humans who handle the frogs, may be severely affected.[192]
Some frogs use bluff or deception. The European common toad (Bufo bufo) adopts a characteristic stance when attacked, inflating its body and standing with its hindquarters raised and its head lowered.[193] The bullfrog (Rana catesbeiana) crouches down with eyes closed and head tipped forward when threatened. This places the parotoid glands in the most effective position, the other glands on its back begin to ooze noxious secretions and the most vulnerable parts of its body are protected.[129] Another tactic used by some frogs is to "scream", the sudden loud noise tending to startle the predator. The gray tree frog (Hyla versicolor) makes an explosive sound that sometimes repels the shrew Blarina brevicauda.[129] Although toads are avoided by many predators, the common garter snake (Thamnophis sirtalis) regularly feeds on them. The strategy employed by juvenile American toads (Bufo americanus) on being approached by a snake is to crouch down and remain immobile. This is usually successful, with the snake passing by and the toad remaining undetected. If it is encountered by the snake's head, however, the toad hops away before crouching defensively.[194]
Distribution

Frogs live on every continent except Antarctica, but they are not present on certain islands, especially those far away from continental land masses.[195][196] Many species are isolated in restricted ranges by changes of climate or inhospitable territory, such as stretches of sea, mountain ridges, deserts, forest clearance, road construction, or other human-made barriers.[197] Usually, a greater diversity of frogs occurs in tropical areas than in temperate regions, such as Europe.[198] Some frogs inhabit arid areas, such as deserts, and rely on specific adaptations to survive. Members of the Australian genus Cyclorana bury themselves underground where they create a water-impervious cocoon in which to aestivate during dry periods. Once it rains, they emerge, find a temporary pool, and breed. Egg and tadpole development is very fast compared with those of most other frogs, so breeding can be completed before the pond dries up.[199] Some frog species are adapted to a cold environment. The wood frog (Rana sylvatica), whose habitat extends into the Arctic Circle, buries itself in the ground during winter. Although much of its body freezes during this time, it maintains a high concentration of glucose in its vital organs, which protects them from damage.[53]
Conservation

In 2006, of 4,035 species of amphibians that depend on water during some lifecycle stage, 1,356 (33.6%) were considered to be threatened. This is likely to be an underestimate because it excludes 1,427 species for which evidence was insufficient to assess their status.[200] Frog populations have declined dramatically since the 1950s. More than one-third of frog species are considered to be threatened with extinction, and more than 120 species are believed to have become extinct since the 1980s.[201] Among these species are the gastric-brooding frogs of Australia and the golden toad of Costa Rica. The latter is of particular concern to scientists because it inhabited the pristine Monteverde Cloud Forest Reserve and its population crashed in 1987, along with about 20 other frog species in the area. This could not be linked directly to human activities, such as deforestation, and was outside the range of normal fluctuations in population size.[202] Elsewhere, habitat loss is a significant cause of frog population decline, as are pollutants, climate change, increased UVB radiation, and the introduction of non-native predators and competitors.[203] A Canadian study conducted in 2006 suggested heavy traffic in their environment was a larger threat to frog populations than was habitat loss.[204] Emerging infectious diseases, including chytridiomycosis and ranavirus, are also devastating populations.[205][206]
Many environmental scientists believe amphibians, including frogs, are good biological indicators of broader ecosystem health because of their intermediate positions in food chains, their permeable skins, and typically biphasic lives (aquatic larvae and terrestrial adults).[207] It appears that species with both aquatic eggs and larvae are most affected by the decline, while those with direct development are the most resistant.[208]

Мутации лягушки и генетические дефекты увеличились с 1990 -х годов. Они часто включают отсутствующие ноги или дополнительные ноги. Различные причины были идентифицированы или предполагают, в том числе увеличение ультрафиолетового излучения, влияющего на порождение на поверхности прудов, химическое загрязнение пестицидов и удобрения, а также паразиты, такие как трематод рибейроя . Вероятно, все это сложное в качестве стрессоров , факторы окружающей среды, способствующие показателям заболевания, и уязвимости для атаки паразитами. Раздвижение ухудшает мобильность, и люди могут не выжить во взрослую жизнь. Увеличение количества лягушек, съеденных птицами, может фактически увеличить вероятность паразитизма других лягушек, потому что сложный жизненный цикл трематода включает в себя Ramshorn Snail и несколько промежуточных хозяев, таких как птицы. [ 209 ] [ 210 ]
В некоторых случаях были созданы программы размножения в неволе и в значительной степени были успешными. [ 211 ] [ 212 ] Всемирная ассоциация зоопарков и аквариумов названа 2008 год как «Год лягушки», чтобы привлечь внимание к проблемам сохранения, с которыми они сталкиваются. [ 213 ]
Тростниковая жаба ( Rhinella Marina ) - очень адаптируемый вид, родом из Южной и Центральной Америки. В 1930 -х годах он был введен в Пуэрто -Рико, а затем различные другие острова в Тихоокеанском и Карибском регионе, как биологический агент по борьбе с вредителями . [ 214 ] В 1935 году 3000 жаб были освобождены на полях сахарного тростника в Квинсленде, Австралия, в попытке контролировать жуков тростников, такие как Дермолепида Альбохиртум , личинки которых повреждают и убивают трости. Первоначальные результаты во многих из этих стран были положительными, но впоследствии стало очевидно, что жабы расстраивают экологический баланс в своей новой среде. Они разводились свободно, конкурировали с местными видами лягушек, ели пчел и другие безвредные местные беспозвоночные, имели мало хищников в их приемных местах обитания, а также отравленных домашних животных, плотоядных птиц и млекопитающих. Во многих из этих стран они в настоящее время считаются вредителями и инвазивными видами , и ученые ищут биологический метод для их контроля. [ 215 ]
Человеческое использование
Кулинарный

Ноги лягушки едят люди во многих частях света. Индонезия является крупнейшим в мире экспортером мяса лягушки, экспортируя более 5000 тонн мяса лягушки каждый год, в основном в Францию, Бельгию и Люксембург. [ 216 ] Первоначально они были предоставлены из местных диких популяций, но чрезмерная эксплуатация привела к уменьшению поставки. Это привело к развитию фермерского хозяйства лягушек и глобальной торговле в лягушках. Основными странами импорта являются Франция, Бельгия, Люксембург и Соединенные Штаты, в то время как главными экспортирующими странами являются Индонезия и Китай. [ 217 ] Ежегодная глобальная торговля американским бычьим проведением ( Rana Catesbeiana ), в основном занимающаяся в Китае, варьируется от 1200 до 2400 тонн. [ 218 ]
Горная куриная лягушка , так называемая, как она на вкус с курицей, теперь находится под угрозой исчезновения, отчасти из-за потребления человеком и был основным выбором пищи доминиканцев . [ 219 ] Енот , опоссум , куропатки , курица -прерия и лягушки были среди проезда, записанных в рамках американской кухни. [ 220 ]
Научные исследования
В ноябре 1970 года НАСА отправило двух быков в космос в течение шести дней во время миссии орбитальной лягушки Отолит для проверки невесомости.
Лягушки используются для рассечений в классах анатомии средней школы и университетов, часто сначала вводили цветные вещества для улучшения контрастов между биологическими системами . Эта практика снижается из -за проблем с защитой животных , и теперь доступны «цифровые лягушки» для виртуального рассечения. [ 221 ]
Лягушки служили экспериментальными животными на протяжении всей истории науки. Биолог восемнадцатого века Луиджи Гальвани обнаружил связь между электричеством и нервной системой, изучая лягушек. Он создал один из первых инструментов для измерения электрического тока из лягушки. [ 222 ] В 1852 году HF Stannius использовал сердце лягушки в процедуре, называемой лигатурой Stannius, чтобы продемонстрировать желудочек и предсердий, бит независимо друг от друга и с разными скоростями. [ 223 ] Африканская когтяная лягушка или платана ( Xenopus laevis ) впервые широко использовалась в лабораториях в тестах на беременность в первой половине 20 -го века. Образец мочи от беременной женщины, введенной в женскую лягушку, вызывает ее откладывать яйца , открытие, сделанное английским зоологом Ланселотом Хогбен . Это связано с тем, что гормон, хорионный гонадотропин человека , присутствует в значительных количествах в моче женщин во время беременности. [ 224 ] В 1952 году Роберт Бриггс и Томас Дж. Кинг кловировали лягушку соматической клеточной ядерной переносом . Позже эта же методика была использована для создания Dolly The Sheep , и их эксперимент был первым, когда у более высоких животных была достигнута успешная ядерная трансплантация. [ 225 ]
Лягушки используются в исследованиях клонирования и других ветвей эмбриологии . Хотя были разработаны альтернативные тесты на беременность, биологи продолжают использовать Xenopus в качестве модели организма в биологии развития, поскольку их эмбрионы большие и легко манипулировать, они легко доступны и могут легко храниться в лаборатории. [ 226 ] Xenopus laevis все чаще перемещается из -за его меньшего относительного, Xenopus tropicalis , который достигает своего репродуктивного возраста через пять месяцев, а не от от одного до двух лет для X. laevis , [ 227 ] Таким образом, облегчая более быстрые исследования в разных поколениях.
Геномы Xenopus laevis , X. Tropicalis , Rana Catesbeiana , Rhinella Marina и Nanorana Parkeri были секвенированы и депонированы в NCBI . базе данных генома [ 228 ]
Фармацевтический

Поскольку токсины лягушек чрезвычайно разнообразны, они подняли интерес биохимиков как «естественную аптеку». Алкалоидный эпибатидин , обезболивающий, в 200 раз более мощный, чем морфин , производится некоторыми видами ядовитых лягушек . Другие химические вещества, выделенные из шкуры лягушек, могут оказать устойчивость к ВИЧ -инфекции. [ 229 ] Дарт -яда подвергаются активному расследованию их потенциала в качестве терапевтических препаратов. [ 230 ]
Давно подозревается, что доколумбовые мезоамериканцы использовали токсическую секрецию, производимую тростниковой жабой в качестве галлюциногена , но, скорее всего, они использовали вещества, выделяемые жабой реки Колорадо ( Bufo Alvarius ). Они содержат буфотенин (5-MEO-DMT), психоактивное соединение , которое использовалось в наше время в качестве препарата для отдыха . Как правило, выделения кожи сушат, а затем копчены. [ 231 ] Зарегистрированное употребление наркотиков, облизывая кожу жабы, сообщалось в средствах массовой информации, но это может быть городской миф . [ 232 ]
Эксудации из кожи золотой ядовитой лягушки ( Phyllobates terribilis ) традиционно используются местными колумбийцами для отравления дротиков, которые они используют для охоты. Кончик снаряда протирается над задней частью лягушки, а дротик запускается из выдувного питания . Комбинация двух алкалоидных токсинов батрахотоксина и гомобатрахотоксина настолько мощный, что одна лягушка содержит достаточно яда, чтобы убить около 22 000 мышей. [ 233 ] два других вида, ядовитая лягушка с ядом кокоу ( Phyllobates aurotaenia ) и лягушка с черными ногами ( биколор Phyllobates Для этой цели также используются ). Они менее токсичны и менее распространены, чем золотой ядовитый лягушка. Они пронзаны заостренными палочками и могут быть нагреты над огнем, чтобы максимизировать количество яда, которое может быть передано в дротик. [ 233 ]

Культурное значение
Лягушки были представлены в мифологии, сказках и популярной культуре. В традиционных китайских мифах мир опирается на гигантскую лягушку, которая пыталась проглотить луну, вызывая лунное затмение . Лягушки были представлены в религии, фольклоре и популярной культуре. Древние египтяне изобразили богосекта , защитника новорожденных, с главой лягушки. Для майя лягушки представляли воду, культуру, фертильность и рождение и были связаны с Богом Чааком . В Библии . Моисей раскрывает чуму лягушек на египтяне Средневековые европейцы связаны лягушки и жабы со злом и колдовством . [ 234 ] Братья Гримм Фея Сказка. Лягушка Принц имеет принцессу, принимающую лягушку, и она превращается в красивого принца. [ 235 ] В современной культуре лягушки могут взять комедийную или несчастную роль, такую как г -н Жаба романа 1908 года The Wind in the Willows , Michigan J. Frog of Warner Bros. Cartoons , Muppet Kermit the Frog и в игре Frogger . [ 236 ]
Ссылки
- ^ Jump up to: а беременный Фрост, Даррел Р. (2021) [1998]. "Анура" . Амфибийские виды мира . Американский музей естественной истории . Получено 29 ноября 2022 года .
- ^ Jump up to: а беременный Cannatella, David C. (1997). "Salientia" . Веб -проект Tree of Life . Получено 7 августа 2012 года .
- ^ Барсук, Д.; Netherton, J. (1995). Лягушки . Airlife Publishing. п. 19. ISBN 978-1-85310-740-5 .
- ^ Jump up to: а беременный Kuzmin, Sergius L. (29 сентября 1999 г.). « Бомбара Бомбана » . Amphibiaweb . Калифорнийский университет, Беркли . Получено 15 июня 2012 года .
- ^ Губы, k; Solís, F.; Ибаньес, Р.; Jaramillo, C.; Fuenmayor, Q. (2010). " Atelopus " Zeteki список Красный IUCN 2010 год . 2 августа ,
- ^ Лидделл, Генри Джордж; Скотт, Роберт (1940). "ἀ" . Грек-английский лексикон . Цифровая библиотека Персея.
- ^ Лидделл, Генри Джордж; Скотт, Роберт (1940). "οὐρά" . Грек-английский лексикон . Цифровая библиотека Персея.
- ^ Bailly, Anatole (1981). Сокращено французским греческим словарем . Париж: Хахетт. ISBN 978-2010035289 Полем OCLC 461974285 .
- ^ Бейли, Анатоль. «Греческий франшл словарь онлайн» . www.tabular.be . Получено 9 декабря 2018 года .
- ^ "Ануран, н. и прил." Полем Оксфордский английский словарь (онлайн изд.). Издательство Оксфордского университета . (Требуется членство в учреждении или участвующее учреждение .)
- ^ Jump up to: а беременный в дюймовый "лягушка, № 1 и прил." Полем Оксфордский английский словарь (онлайн изд.). Издательство Оксфордского университета . (Требуется членство в учреждении или участвующее учреждение .)
- ^ "Фрош | Фроск, № 1." Полем Оксфордский английский словарь (онлайн изд.). Издательство Оксфордского университета . (Требуется членство в учреждении или участвующее учреждение .)
- ^ Jerzy Wełna, « Метатетический и неметатический выбор форм в среднеанглийском языке », Studia Anglica Posnaniensia , 30 (2002), 501–18 (стр. 504).
- ^ "Жаба, н". Полем Оксфордский английский словарь (онлайн изд.). Издательство Оксфордского университета . (Требуется членство в учреждении или участвующее учреждение .)
- ^ «Головастик, № 1». Полем Оксфордский английский словарь (онлайн изд.). Издательство Оксфордского университета . (Требуется членство в учреждении или участвующее учреждение .)
- ^ Jump up to: а беременный Паф, FH; Эндрюс, RM; Cadle, Je; Крамп, ML; Савицкий, ах; Уэллс, К.Д. (2003). Герпетология (третья.). Бенджамин Каммингс. ISBN 978-0-13-100849-6 .
- ^ Фрост, Даррел Р. (2023). «Анура Фишер фон Вальдхайм, 1813» . Амфибийские виды мира: онлайн -ссылка. Версия 6.2 . Американский музей естественной истории . Получено 19 сентября 2023 года .
- ^ Дуэльман, Уильям Э. Биология амфибий , с. 319, в Google Books
- ^ Jump up to: а беременный Каннателла, Дэвид (11 января 2008 г.). "Анура" . Веб -проект Tree of Life . Получено 8 августа 2012 года .
- ^ Ford, LS; Cannatella, DC (1993). «Основные клады лягушек». Герпетологические монографии . 7 : 94–117. doi : 10.2307/1466954 . JSTOR 1466954 .
- ^ Faivovich, J.; Хаддад, CFB; Гарсия, PCA; Мороз, доктор; Кэмпбелл, JA; Wheeler, WC (2005). «Систематический обзор семейства лягушек Hylidae, с особой ссылкой на Hylinae: филогенетический анализ и пересмотр». Бюллетень Американского музея естественной истории . 294 : 1–240. Citeseerx 10.1.1.470.2967 . doi : 10.1206/0003-0090 (2005) 294 [0001: srotff] 2.0.co; 2 . S2CID 83925199 .
- ^ Kuzmin, SL (Nover 10, 1999). " Пелофилакс Esculentus Поврежден 12 октября 2012 года .
- ^ Köhler, Sonja (2003). «Механизмы частичной репродуктивной изоляции в гибридной зоне Бомбины в Румынии» (PDF) . Диссертация для диссертации . Архивировано (PDF) из оригинала 9 октября 2022 года . Получено 5 июня 2012 года .
- ^ Сан -Мауро, Диего; Венс, Майкл; Alcobendas, Марина; Зардоя, Рафаэль; Мейер, Аксель (2005). «Первоначальная диверсификация живых амфибий предшествовала разрыву Пангеи » Американский натуралист 165 (5): 590–5 Doi : 10.1086/ 4 JSTOR 429523 PMID 15795855 S2CID 17021360 (Требуется подписка)
- ^ Чжан, Пэн; Чжоу, Хуи; Чен, Юэ-Кин; Лю, Yi-Fei; Q, Liang-Hu (2005). «Митогеномные перспективы происхождения и филогения живых амфибий» . Систематическая биология . 54 (3): 391–400. doi : 10.1080/10635150590945278 . PMID 16012106 .
- ^ Марджанович, Давид; Лаурин, Мишель (2007). «Окаменелости, молекулы, время дивергенции и происхождение лиссамбийцев» . Систематическая биология . 56 (3): 369–388. doi : 10.1080/10635150701397635 . PMID 17520502 .
- ^ Pyron, R. Alexander (2011). «Оценка времени дивергенции с использованием окаменелостей в качестве терминальных таксонов и происхождения Lissamphibia» . Систематическая биология . 60 (4): 466–481. doi : 10.1093/sysbio/syr047 . PMID 21540408 .
- ^ Кассельман, Энн (21 мая 2008 г.). « Лягушка-арандер» окаменелости может быть амфибий-отсутствующим звеном » . National Geographic News . Архивировано из оригинала 25 мая 2008 года . Получено 5 июля 2012 года .
- ^ Андерсон, Джейсон С.; Рейс, Роберт Р.; Скотт, Дайан; Fröbisch, Nadia B.; Сумида, Стюарт С. (2008). «Стебель батрахиян из раннего пермца Техаса и происхождение лягушек и саламандров». Природа . 453 (7194): 515–518. Bibcode : 2008natur.453..515a . doi : 10.1038/nature06865 . PMID 18497824 . S2CID 205212809 .
- ^ Marjanović, D.; Лаурин М. (2009). «Происхождение современных амфибий: комментарий» (PDF) . Эволюционная биология . 36 (3): 336–338. Bibcode : 2009evbio..36..336m . doi : 10.1007/s11692-009-9065-8 . S2CID 12023942 .
- ^ Jump up to: а беременный Cannatella, David (1995). « Триадобатрах Массиноти » . Дерево жизни . Получено 26 июня 2008 года .
- ^ Мишель Р. Стокер; Стерлинг Дж. Несбитт; Бен Т. Клигман; Даниэль Дж. Парух; Адам Д. Марш; Дэвид С. Блэкберн; Уильям Г. Паркер (2019). «Самая ранняя экваториальная запись лягушек из позднего триаса Аризоны» . Биологические письма . 15 (2): ID статьи 20180922. DOI : 10.1098/rsbl.2018.0922 . HDL : 10919/87931 . PMC 6405462 . PMID 30958136 .
- ^ Roček, Z. (2000). «14. Мезозойские амфибии» (PDF) . В Heatwole, H.; Кэрролл, Р.Л. (ред.). Амфибийская биология: палеонтология: эволюционная история амфибий . Тол. 4. Суррей Битти и сыновья. С. 1295–1331. ISBN 978-0-949324-87-0 Полем Архивировано (PDF) из оригинала 9 октября 2022 года.
- ^ Weishampel, DB; Додсон, П.; Osmólska, H., eds. (2004). Распространение динозавров (Ранняя Юра, Северная Америка): Динозаурия (2 -е изд.). Калифорнийский университет. С. 530–532. ISBN 978-0-520-24209-8 .
- ^ Шубин, NH; Дженкинс, FA JR (1995). «Ранняя юрская лягушка». Природа . 377 (6544): 49–52. Bibcode : 1995natur.377 ... 49 с . doi : 10.1038/377049a0 . S2CID 4308225 .
- ^ Foster, J. (2007). «Анура (лягушки)». Юрский Запад: динозавры формирования Моррисона и их мира . Издательство Университета Индианы. С. 135–136. ISBN 978-0-253-34870-8 .
- ^ Уилсон, AC; Сарич, виртуальная машина; Maxson, LR (1974). «Важность перестройки гена в эволюции: данные исследований о показателях хромосомной, белковой и анатомической эволюции» . Труды Национальной академии наук . 71 (8): 3028–3030. Bibcode : 1974pnas ... 71.3028W . doi : 10.1073/pnas.71.8.3028 . PMC 388613 . PMID 4528784 .
- ^ «Эволюция лягушки, связанная с ударом астероидов динозавров» . BBC News . 3 июля 2017 года . Получено 3 июля 2017 года .
- ^ Фенг, Ян-Джи; Блэкберн, Дэвид С.; Лян, Дэн; Хиллис, Дэвид М.; Wake, David B.; Cannatella, David C.; Чжан, Пэн (2017). «Филогеномика выявляет быструю, одновременную диверсификацию трех основных клад гондванан лягушек на границе мелового ипалеогена» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (29): E5864 - E5870. BIBCODE : 2017PNAS..114E5864F . doi : 10.1073/pnas.1704632114 . PMC 5530686 . PMID 28673970 .
- ^ Эванс, SE; Джонс, Мех; Краузе, DW (2008). «Гигантская лягушка с южноамериканским сродством от покойного мела Мадагаскара» . Труды Национальной академии наук . 105 (8): 2951–2956. Bibcode : 2008pnas..105.2951e . doi : 10.1073/pnas.0707599105 . PMC 2268566 . PMID 18287076 .
- ^ Морс, Томас; Регуаро, Марсело; Васильан, Дэвид (2020). «Первая ископаемая лягушка из Антарктиды: последствия для эоценовых высоких климатических условий и гондвананского космополитизма Австралобатрахии» . Научные отчеты . 10 (1): 5051. Bibcode : 2020natsr..10.5051M . doi : 10.1038/s41598-020-61973-5 . PMC 7181706 . PMID 32327670 .
- ^ Джоэл, Лукас (23 апреля 2020 года). «Ископаемые показывают хладнокровные лягушки, жившие на теплой Антарктиде» . New York Times . ISSN 0362-4331 . Архивировано из оригинала 23 апреля 2020 года . Получено 13 мая 2020 года .
- ^ Мороз, доктор; Грант, Т.; Файвович, JN; Бейн, RH; Haas, A.; Хаддад, CLFB; De Sá, ro; Ченнинг, А.; Уилкинсон, М.; Доннеллан, Южная Каролина; Raxworthy, CJ; Кэмпбелл, JA; Блотто, BL; Moler, P.; Дрюс, RC; Nussbaum, ra; Линч, JD; Зеленый, DM; Wheeler, WC (2006). «Амфибийское дерево жизни» . Бюллетень Американского музея естественной истории . 297 : 1–291. doi : 10.1206/0003-0090 (2006) 297 [0001: Tatol] 2.0.co; 2 . HDL : 2246/5781 . S2CID 86140137 .
- ^ Heinicke MP; Дуэльман, мы; TrueB, L.; Означает, дБ; Macculloch, Rd; Хеджес, SB (2009). «Новая семья лягушек (Anura: Terranana) из Южной Америки и расширенная клада прямого развития, выявленная молекулярной филогении» (PDF) . Zootaxa . 2211 : 1–35. doi : 10.11646/Zootaxa.2211.1.1 .
- ^ Р. Александр Пирон; Джон Дж. Винс (2011). «Большая филогения амфибий, в том числе более 2800 видов, и пересмотренная классификация существующих лягушек, саламандров и кесэцилия» . Молекулярная филогенетика и эволюция . 61 (2): 543–583. Bibcode : 2011molpe..61..543a . doi : 10.1016/j.ympev.2011.06.012 . PMID 21723399 .
- ^ Jump up to: а беременный Flam, F. (1995). «Поиск самой ранней настоящей лягушки поможет палеонтологам понять, как лягушка развила свою способность прыгать» . Найт Риддер/Служба новостей Tribune. Архивировано из оригинала 13 мая 2013 года . Получено 10 июня 2012 года .
- ^ «Крошечная лягушка претендовала как самый маленький в мире позвоночный» . Хранитель . 12 января 2012 года . Получено 28 сентября 2012 года .
- ^ Colgan, PV (1982). Книга Гиннеса о фактах и подвигах (3 изд.). Превосходная степень Гиннесса. С. 118–119. ISBN 978-0851122359 .
- ^ Отеро, Ра; П. Хименес-Хумобро; S. Soto-Acuña; Рейри-Яньес (2014). Магаллан, южный Чили, является гинаном земля Американская 55 : 133–140. Bibcode : 2014jsies . doi : 10.1016/j.jsames .
- ^ Морфологические различия в конечностях анурана: ограничения и новинки
- ^ Морфологическая и экологическая конвергенция на пределе более низкого размера для позвоночных, выделенных пятью новыми миниатюрными видами микрогилидных лягушек из трех различных мадагасканских родов
- ^ Jump up to: а беременный Минотт, Кевин (15 мая 2010 г.). «Как лягушки прыгают» . National Geographic . Архивировано с оригинала 4 ноября 2013 года . Получено 10 июня 2012 года .
- ^ Jump up to: а беременный в дюймовый и фон Теслер П. (1999). «Удивительная адаптируемая лягушка» . Exploratorium :: Музей науки, искусства и человеческого восприятия . Получено 4 июня 2012 года .
- ^ Винсент Л. (2001). " Litoria Caerulea " (PDF ) Университет Джеймса Кука. Архивировано из оригинала (PDF) 22 апреля , Получено августа 3 ,
- ^ Эмерсон, SB; Дил Д. (1980). «Морфология падений и механизмы прилипания в лягушках». Биологический журнал Линневого общества . 13 (3): 199–216. doi : 10.1111/j.1095-8312.1980.tb00082.x .
- ^ Харви, MB; Пембертон, AJ; Смит, EN (2002). «Новые и плохо известные парашютные лягушки (Rhacophoridae: Rhacophorus ) от Sumatra и Java». Герпетологические монографии . 16 : 46–92. doi : 10.1655/0733-1347 (2002) 016 [0046: NAPKPF] 2.0.co; 2 . S2CID 86616385 .
- ^ «Spadefoot Couch's ( Scapphiopus couchi )» . Музей пустыни Аризоны-Сонора . Получено 3 августа 2012 года .
- ^ Уокер, М. (25 июня 2009 г.). «Загадка без легтей решены» . BBC News .
- ^ Стеббинс, Роберт С .; Коэн, Натан В. (1995). Естественная история амфибий . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. С. 10–14. ISBN 978-0-691-03281-8 .
- ^ Мороз, SW (1932). «Примечания о кормлении и лищении в лягушках». Американский натуралист . 66 (707): 530–540. doi : 10.1086/280458 . JSTOR 2456779 . S2CID 84796411 .
- ^ Jump up to: а беременный Барсук, Д.; Netherton, J. (1995). Лягушки . Airlife Publishing. п. 27. ISBN 978-1-85310-740-5 .
- ^ Смит, HR (1962). Амфибии и их пути . Макмиллан. ISBN 978-0-02-612190-3 .
- ^ Дикерсон, MC (1969). Книга лягушек: североамериканские лягушки и жабы . Dover Publications. ISBN 978-0-486-21973-8 .
- ^ Блэкберн, округ Колумбия (14 ноября 2002 г.). " Трихобатрахус Робуст " . Amphibiaweb . Получено 18 августа 2012 года .
- ^ Рубилал, Родольфо; Shemaker, Vughan (1985). "Остермии в Аньяне" Журнал герпеологии 18 (3): 313–3 doi : 10.2307/154085 . JSTOR 1564085 .
- ^ Vitt, Laure J.; Caldwell, Janle P. (2013). До этого: вводная биология амфибий и рептов Академическая пресса. П. 50. ISBN 9780123869203 .
- ^ Джаред, C.; Antoniazzi, MM; Навас, Калифорния; Katchburian, E.; Freymüller, E.; Тамбурги, DV; Rodrigues, MT (2005). «Космирование головы, фрагмоз и защита в лягушкой лягушке из каритомантиса Greeningi ». Журнал зоологии . 265 (1): 1–8. doi : 10.1017/s0952836904005953 . S2CID 59449901 .
- ^ Jump up to: а беременный Бертон, Морис (1972). Книга наблюдателя британских диких животных . Frederick Warne & Co. с. 204–209. ISBN 978-0-7232-1503-5 .
- ^ Гуаясамин, Хуан М.; Кринак, Тим; Кринак, Кэтрин; Кулебрас, Хайме; Хаттер, Карл Р. (2015). «Фенотипическая пластичность поднимает вопросы для таксономически важных черт: замечательная новая Андская дояль (Pristimantis) с способностью менять текстуру кожи: фенотипическая пластичность у Андского Районк -края» . Зоологический журнал Линневого общества . 173 (4): 913–928. doi : 10.1111/Zoj.12222 .
- ^ Wede, WH; Филлипс, JB (2003). «Фиксированный зеленый и коричневый цвет морфы и новое изменяющее цвет морф тихоокеанской лягушки Hyla Regilla ». Американский натуралист . 162 (4): 461–473. doi : 10.1086/378253 . JSTOR 10.1086/378253 . PMID 14582008 . S2CID 25692966 .
- ^ Boisvert, Адам (23 октября 2007 г.). " Barbourula kalimantanensis " . Amphibiaweb . Калифорнийский университет, Беркли . Получено 9 июля 2012 года .
- ^ Кимбалл, Джон (2010). «Системы кровообращения животных: три камеры: лягушка и ящерица» . Страницы биологии Кимбалла . Архивировано из оригинала 11 мая 2016 года . Получено 9 июля 2012 года .
- ^ Ли, Дебора (23 апреля 2010 г.). « Телматобий Кулеус » . Amphibiaweb . Калифорнийский университет, Беркли . Получено 9 июля 2012 года .
- ^ Секрет липкости лягушкой плюки | Стао
- ^ Левин, RP; Монрой, JA; Брейнерд, Эл (15 марта 2004 г.). «Вклад в борьбу с глазами в глотающую производительность в северной лягушке леопарда Rana Pipiens » . Журнал экспериментальной биологии . 207 (Pt 8): 1361–1368. doi : 10.1242/jeb.00885 . PMID 15010487 .
- ^ "Лягушательная пищеварительная система" . Tutorvista.com. 2010. Архивировано из оригинала 3 июня 2010 года . Получено 4 августа 2012 года .
- ^ Jump up to: а беременный Дорит, RL; Уокер, WF; Barnes, RD (1991). Зоология . Saunders College Publishing. п. 849 . ISBN 978-0-03-030504-7 .
- ^ Jump up to: а беременный в дюймовый «Внутренние системы лягушки» . Tutorvista.com. 2010. Архивировано из оригинала 21 января 2008 года . Получено 4 июня 2012 года .
- ^ Duellman, WE и L. Trueb (1986). Биология амфибий . Нью-Йорк: издательская компания McGraw-Hill.
- ^ Север, Дэвид М.; Стауб, Нэнси Л. «Гормоны, секс -аксессуары и вторичные сексуальные характеристики у амфибий» (PDF) . Гормоны и размножение позвоночных - том 2: амфибии . С. 83–98. Архивировано (PDF) из оригинала 9 октября 2022 года . Получено 4 августа 2012 года .
- ^ Лаурин, Мишель; Готье, Жак А. (2012). "Амниота" . Веб -проект Tree of Life . Получено 4 августа 2012 года .
- ^ Jump up to: а беременный Говард, Ян П.; Роджерс, Брайан Дж. (1995). Бинокулярное зрение и стереопсис . Издательство Оксфордского университета. п. 651. ISBN 978-0195084764 .
- ^ Jump up to: а беременный в Барсук, Дэвид; Нетертон, Джон (1995). Лягушки . Airlife Publishing Ltd. с. 31–35. ISBN 978-1-85310-740-5 .
- ^ Мунц, WRA; Scientific American Offprints (1964). «Видение у лягушек». Scientific American . 210 (3). WH Freeman: 110–9. ASIN B0006RENBO . Bibcode : 1964sciam.210C.110M . doi : 10.1038/Scientificamerican0364-110 . OCLC 15304238 . PMID 14133069 .
- ^ Келбер, Алмут; Йованович, Карола; Олссон, Питер (5 апреля 2017 г.). «Пороговые значения и ограничения шума цветового зрения в тусклом свете» . Философские транзакции Королевского общества B: биологические науки . 372 (1717): 20160065. DOI : 10.1098/rstb.2016.0065 . ISSN 0962-8436 . PMC 5312015 . PMID 28193810 .
- ^ Jump up to: а беременный Барсук, Дэвид; Нетертон, Джон (1995). Лягушки . Airlife Publishing. п. 38. ISBN 978-1-85310-740-5 .
- ^ Jump up to: а беременный Стеббинс, Роберт С .; Коэн, Натан В. (1995). Естественная история амфибий . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. С. 67–69. ISBN 978-0-691-03281-8 .
- ^ Армстронг, Сесилия Э.; Робертс, Уильям М. (1998). «Электрические свойства саккулярных волосковых клеток лягушки: искажение ферментативной диссоциацией» . Журнал нейробиологии . 18 (8): 2962–2973. doi : 10.1523/jneurosci.18-08-02962.1998 . PMC 6792591 . PMID 9526013 .
- ^ "Бык -экраг" . Департамент природных ресурсов Огайо. Архивировано из оригинала 18 августа 2012 года . Получено 19 июня 2012 года .
- ^ Tan, W.-H.; Цай, С.Г.; Лин, C.; Лин, YK (5 июня 2014 г.). «Эффект городского каньона: штормовые стоки улучшают характеристики вызовов лягушки Mientien Tree» . Журнал зоологии . 294 (2): 77–84. doi : 10.1111/jzo.12154 . ISSN 0952-8369 .
- ^ Нафис, Гэри (2012). " Ascaphus truei : лягушка с прибрежным хвостом" . Калифорнийский герп . Получено 19 июня 2012 года .
- ^ Рой, Дебьяни (1997). «Коммуникационные сигналы и сексуальный отбор у амфибий» (PDF) . Текущая наука . 72 : 923–927. Архивировано из оригинала (PDF) 23 сентября 2012 года.
- ^ Gerhardt, HC (1994). «Эволюция вокализации у лягушек и жаб». Ежегодный обзор экологии и систематики . 25 : 293–324. doi : 10.1146/annurev.es.25.110194.001453 .
- ^ Jump up to: а беременный в дюймовый Барсук, Дэвид; Нетертон, Джон (1995). Лягушки . Airlife Publishing Ltd. с. 39–44. ISBN 978-1-85310-740-5 .
- ^ Хилтон, Билл младший (1986). "9. 'Jug-o-rum': звонок любовного быка». Натуралист Piedmont, том 1 . Йорк, Южная Каролина: Хилтонский пруд Центр по естественной истории Пьемонта. ISBN 978-0-9832151-0-3 .
- ^ Нэш, Пэт (февраль 2005 г.). «Жизнь дерева лягушек» . Архивировано из оригинала 9 марта 2012 года . Получено 4 августа 2012 года .
- ^ Аристофан. "Лягушки" . Архивировано из оригинала 13 мая 2012 года . Получено 19 июня 2012 года .
- ^ Сутерс, Ра; Наринс, премьер -министр; Лин, W; Schnitzler, H; Дензингер, а; Xu, c; Feng, AS (2006). «Голоса мертвых: сложные нелинейные вокальные сигналы из гортани ультразвуковой лягушки» . Журнал экспериментальной биологии . 209 (24): 4984–4993. doi : 10.1242/jeb.02594 . PMID 17142687 .
- ^ Эммер, Рик (24 ноября 1997 г.). «Как лягушки переживают зиму? Почему они не замерзают до смерти?» Полем Scientific American . Получено 15 июня 2012 года .
- ^ Kayes, Sara M.; Судороги, Ребекка Л.; Франклин, Крейг Э. (2009). «Метаболическая депрессия во время эстиции в циклоранах alboguttata ». Сравнительная биохимия и физиология - Часть A: Молекулярная и интегративная физиология . 154 (4): 557–563. doi : 10.1016/j.cbpa.2009.09.001 . PMID 19737622 .
- ^ Хадсон, Нью -Джерси; Lehnert, SA; Ингам, А.Б.; Symonds, B.; Франклин, CE; Harper, GS (2005). «Уроки от обогащающей лягушки: щадящий мышечный белок, несмотря на голод и не используют». AJP: нормативно -правовая, интегративная и сравнительная физиология . 290 (3): R836 - R843. doi : 10.1152/ajpregu.00380.2005 . PMID 16239372 . S2CID 8395980 .
- ^ Уилмер, Пэт (2009). Физиология окружающей среды животных . Уайли. с. 188 . ISBN 9781405107242 .
- ^ «10 лучших животных -джемпер» . Scienceray. Архивировано с оригинала 10 августа 2012 года . Получено 11 июня 2012 года .
- ^ Джеймс, RS; Уилсон, Р.С. (2008). «Взрывные прыжки: экстремальные морфологические и физиологические специализации австралийских ракетных лягушек ( Litoria nasuta )» (PDF) . Физиологическая и биохимическая зоология . 81 (2): 176–185. doi : 10.1086/525290 . PMID 18190283 . S2CID 12643425 . Архивировано (PDF) из оригинала 9 октября 2022 года.
- ^ Nauwelaerts, S.; Schollier, J.; Aerts, P. (2004). «Функциональный анализ того, как лягушки выпрыгивают из воды» . Биологический журнал Линневого общества . 83 (3): 413–420. doi : 10.1111/j.1095-8312.2004.00403.x .
- ^ Jump up to: а беременный Барсук, Дэвид; Нетертон, Джон (1995). Лягушки . Airlife Publishing. п. 51. ISBN 978-1-85310-740-5 .
- ^ Astley, HC; Робертс, TJ (2011). «Свидетельство о катапульте позвоночных: хранение энергии упругого сухожилия Plantaris во время прыжков лягушки» . Биологические письма . 8 (3): 386–389. doi : 10.1098/rsbl.2011.0982 . PMC 3367733 . PMID 22090204 .
- ^ Скотт, Дж. (2005). «Прыжок Саранчи: интегрированное лабораторное исследование». Достижения в области физиологического образования . 29 (1): 21–26. doi : 10.1152/advan.00037.2004 . PMID 15718379 . S2CID 13423666 .
- ^ Jump up to: а беременный в Бакли, Кр; Майкл, SF; Irschick, DJ (2005). «Раннее вылупление снижает производительность прыжков в прямом развитии лягушки, Eleutherodactylus coqui» . Функциональная экология . 19 (1): 67–72. Bibcode : 2005fueco..19 ... 67b . doi : 10.1111/j.0269-8463.2005.00931.x . ISSN 0269-8463 . JSTOR 3599272 .
- ^ Jump up to: а беременный в ; Дуэльман, Уильям Э. (14 мая 2014 г.). "Анура " Britannica Encyclopædia Получено апреля 26 ,
- ^ Fitch, HS (1956). «Экологическое исследование ящерицы с воротником ( Crotaphytus collaris )». Университет Канзаса публикации . 8 : 213–274.
- ^ Уолтон, М.; Андерсон, BD (1988). «Аэробная стоимость соленой локомоции в жабе Фаулера ( Bufo Woodhousei Fowleri )» . Журнал экспериментальной биологии . 136 (1): 273–288. doi : 10.1242/jeb.136.1.273 . PMID 3404074 .

- ^ Ан, Ан; Борозд, E.; Biewener, AA (2004). «Ходьба и бегая в бегущей лягушке с красной ногой, Kassina Maculata » . Журнал экспериментальной биологии . 207 (Pt 3): 399–410. doi : 10.1242/jeb.00761 . PMID 14691087 .
- ^ Pickersgill, M.; Schiøtz, A.; Хауэлл, К.; Минтер Л. (2004). " Кассина Макулата " . МСОП красный список угрожаемых видов . 2004 . Получено 11 июня 2012 года .
- ^ Jump up to: а беременный "Pipidae" . Amphibiaweb . Калифорнийский университет, Беркли . Получено 14 июня 2012 года .
- ^ Jump up to: а беременный Дуэльман, США; Zug, gr "Enra: от головастика до анульта " Британская июля Получено 13 ,
- ^ Radhakrishnan, C.; Gopi, KC (2007). «Расширение диапазона распределения Nasikabatrachus sahyadrensis Biju & Bossuyt (Amphibia: Anura: Nasikabatrachidae) вдоль Западных Гат, с некоторым пониманием его биономики» (PDF) . Текущая наука . 92 (2): 213–216. ISSN 0011-3891 . Архивировано (PDF) из оригинала 9 октября 2022 года.
- ^ Jump up to: а беременный Фаррар, Евгения; Эй, Джейн. " Spea Bombifrons " . Amphibiaweb . Калифорнийский университет, Беркли . Получено 16 июня 2012 года .
- ^ «Scaphiopodidae» . Amphibiaweb . Калифорнийский университет, Беркли . Получено 16 июня 2012 года .
- ^ Робертс, Дейл; Герой, Жан-Марк (2011). « Heleioporus albopunctatus » . МСОП красный список угрожаемых видов . 2011 год Получено 16 июня 2012 года .
- ^ Standzewski, Марк (30 сентября 1998 г.). «Мадагасканские лягушки: Род: Scaphiophyryne (Boulenger, 1882) » Получено июня 16 ,
- ^ Венески, м; Raxworthy, CJ; Nussbaum, ra; Глоу Ф. (2003). «Пересмотр компонента Scaphiophryn Marmorata из мраморных жабей из Мадагаскара, включая описание нового вида» (PDF) . Герпетологический журнал . 13 : 69–79. Архивировано (PDF) из оригинала 9 октября 2022 года.
- ^ Federle, W.; Барнс, WJP; Baumgartner, W.; Drechsler, P.; Смит, JM (2006). «Мокрый, но не скользкий: граничное трение в клейких накладках лягушах» . Журнал интерфейса Королевского общества . 3 (10): 689–697. doi : 10.1098/rsif.2006.0135 . PMC 1664653 . PMID 16971337 .
- ^ Кокран, Дорис Мейбл (1961). Живые амфибии мира . Doubleday. п. 112. ISBN 978-0-241-90338-4 .
- ^ " Phyllomedusa ayaye " . Amphibiaweb . Получено 14 июня 2012 года .
- ^ Эмерсон, Шарон Б.; Koehl, Mar (1990). «Взаимодействие поведенческих и морфологических изменений в эволюции нового локомоторного типа:« летающие лягушки ». . 44 ( 8): 1931–1946. Дои : 10.2307/2409604 . JStor 2409604. 28564439 PMID . Эволюция
- ^ Шах, солнечный; Тивари, Рахна (29 ноября 2001 г.). " Rhacophorus nigropalmatus " . Amphibiaweb . Калифорнийский университет, Беркли . Получено 11 июня 2012 года .
- ^ «Летающая лягушка Уоллеса Rhacophorus nigropalmatus » . National Geographic: животные. 10 сентября 2010 г. Архивировано с оригинала 18 января 2010 года . Получено 5 июня 2012 года .
- ^ Jump up to: а беременный в дюймовый и фон глин Стеббинс, Роберт С .; Коэн, Натан В. (1995). Естественная история амфибий . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. С. 154–162. ISBN 978-0-691-03281-8 .
- ^ Дэвис, NB; Хэллидей, Т.Р. (1978). «Глубокие рассылки и борьба с оценкой в жабах Bufo Bufo ». Природа . 274 (5672): 683–685. Bibcode : 1978natur.274..683d . doi : 10.1038/274683a0 . S2CID 4222519 .
- ^ Лонг, Дэвид Р. (1989). «Энергетика и воспроизводство у женского Scaphiopus multiplicatus из западного Техаса». Журнал герпетологии . 23 (2): 176–179. doi : 10.2307/1564026 . JSTOR 1564026 .
- ^ Искандар, DT; Эванс, BJ; McGuire, JA (2014). «Новый репродуктивный режим у лягушек: новый вид клыка с внутренним оплодотворением и рождением головастиков» . Plos один . 9 (12): E115884. Bibcode : 2014ploso ... 9k5884i . doi : 10.1371/journal.pone.0115884 . PMC 4281041 . PMID 25551466 .
- ^ Ченнинг, Алан; Хауэлл, Ким М. (2006). Амфибии Восточной Африки . Comstock Publishing. С. 104–117. ISBN 978-0-8014-4374-9 .
- ^ Sandberger, L.; Hillers, A.; Doumbia, J.; Луа, NS.; Brede C.; Rödel, Mo. (2010). «Повторное открытие либерийской нимбы Жаба, Nimbaphrynoides liberiensis (Xavier, 1978) (Amphibia: Anura: Bufonidae), и переиздание его таксономического статуса» (PDF) . Zootaxa . 4355 : 56–68. doi : 10.11646/Zootaxa.2355.1.3 . ISSN 1175-5334 . Архивировано (PDF) из оригинала 9 октября 2022 года.
- ^ Маттисон, Крис (2011). Лягушки и жабы миров . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. п. 91. ISBN 978-0-691-14968-4 .
- ^ Дуэльман, США; Zug, gr "Арара: Харлы и вылупление " Британская апреля Получено 9 ,
- ^ Duellman, William E.; TrueB, Линда; Арак, Энтони (2002). «Лягушки и жабы». В Хэллидее Тим; Адлер, Крайг (ред.). Энциклопедия огненных лиц рептилий и амфибий . Книги огня. п. 67. ISBN 978-1-55297-613-5 .
- ^ Эстрада, Альберто Р.; Хеджес, С. Блэр (1996). «При пределе нижнего размера в тетраподах: новая миниатюрная лягушка от Кубы (Leptodactylidae: Eleutherodactylus)». Копея . 1996 (4): 852–859. doi : 10.2307/1447647 . JSTOR 1447647 .
- ^ Уиттакер, Келли; Chantasirivisal, Peera (2 декабря 2005 г.). " Leptodactylus pentadactylus " . Amphibiaweb . Калифорнийский университет, Беркли . Получено 19 июля 2012 года .
- ^ Уиттакер, Келли (27 июня 2007 г.). « Агаличнис каллидрис » . Amphibiaweb . Калифорнийский университет, Беркли . Получено 19 июля 2012 года .
- ^ Гилберт, Перри В. (1942). «Наблюдения на яйцах Ambystoma maculatum с особой ссылкой на зеленые водоросли, обнаруженные в яичных конвертах». Экология . 23 (2): 215–227. Bibcode : 1942ecol ... 23..215G . doi : 10.2307/1931088 . JSTOR 1931088 .
- ^ Вальдман, Брюс; Райан, Майкл Дж. (1983). «Тепловые преимущества общего осаждения массы яиц у деревянных лягушек ( Rana sylvatica )». Журнал герпетологии . 17 (1): 70–72. doi : 10.2307/1563783 . JSTOR 1563783 .
- ^ ВАКЕНТИН, КМ (1995). «Адаптивная пластичность в возрасте вылупления: реакция на компромиссы риска хищничества» . Труды Национальной академии наук . 92 (8): 3507–3510. Bibcode : 1995pnas ... 92.3507W . doi : 10.1073/pnas.92.8.3507 . PMC 42196 . PMID 11607529 .
- ^ Jump up to: а беременный в дюймовый и Стеббинс, Роберт С .; Коэн, Натан В. (1995). Естественная история амфибий . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. С. 179–194. ISBN 978-0-691-03281-8 .
- ^ Голдберг, Хавьер; Taucce, Pedro Pg; Куинцио, Сильвия Инес; Хаддад, Селио Ф.Б; Вера Кандиоти, Флоренсия (1 января 2020 г.). «Увеличение наших знаний о лягушках с прямым развитием: онтогенью Ischnocnema henselii (Anura: Brachycephalidae)» . Зоологиссер Анзегер . 284 : 78–87. Bibcode : 2020zooan.284 ... 78G . Doi : 10.1016/j.jcz.2019.11.001 . ISSN 0044-5231 . S2CID 209576535 .
- ^ Элинсон, Ричард П. (2001). «Прямое развитие: альтернативный способ сделать лягушку» . Бытие . 29 (2): 91–95. doi : 10.1002/1526-968X (200102) 29: 2 <91 :: AID-Gene1009> 3.0.co; 2-6 . ISSN 1526-968X . PMID 11170349 . S2CID 32550621 .
- ^ Seshadri, KS (2015). «Рякофоридные лягушки размножаются в бамбуке: открытие нового репродуктивного режима из западных Гат» . Лягушка . 23 (4) (116): 46–49.
- ^ Wickramasinghe, DeepThi D.; Oseen, Kerri L.; Wassersug, Richard J. (2007). «Онтогенетические изменения в рационе и морфологии кишечника в полутерриальных головастиках нанофрис ceylonensis (dicroglossidae)». Копея . 2007 (4): 1012–1018. doi : 10.1643/0045-8511 (2007) 7 [1012: ocidai] 2.0.co; 2 . JSTOR 25140719 . S2CID 86244218 .
- ^ Янзен, Питер (10 мая 2005 г.). « Наннофрис ceylonensis » . Amphibiaweb . Калифорнийский университет, Беркли . Получено 20 июля 2012 года .
- ^ Хофф, К. против.; Wassersug, RJ (1 февраля 2000 г.). «Локомоция головастика: осевое движение и хвост функционируют в основном позвоночных позвоночных» . Интегративная и сравнительная биология . 40 (1): 62–76. doi : 10.1093/icb/40.1.62 .
- ^ Крамп, Марта Л. (1986). «Каннибализм младших головастиков: еще одна опасность метаморфозы». Копея . 1986 (4): 1007–1009. doi : 10.2307/1445301 . JSTOR 1445301 .
- ^ Стеббинс, Роберт С .; Коэн, Натан В. (1995). Естественная история амфибий . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. С. 173–175. ISBN 978-0-691-03281-8 .
- ^ Бендер, MC; Ху, С.; Pelletier, C.; Денвер, RJ (2018). «Еда или не есть: онтогенство контрольных кормления гипоталамуса и роль лептина в модулировании перехода жизни в амфибии головастиков» . Разбирательство. Биологические науки . 285 (1875). doi : 10.1098/rspb.2017.2784 . PMC 5897637 . PMID 29593109 .
- ^ Головки питаются как сумасшедшие, прежде чем реконструировать их кишки - будущее
- ^ Кол, К.Д.; Кэри, TL; Карасов, WH; Dearing, MD (2013). «Реструктуризация микробиоты кишечника амфибий посредством метаморфозы» . Отчеты о микробиологии окружающей среды . 5 (6): 899–903. Bibcode : 2013envmr ... 5..899k . doi : 10.1111/1758-2229.12092 . PMID 24249298 .
- ^ Фабрези, Марисса; Круз, Хулио Сезар (2020). «Эволюционные и соображения развития диеты и морфологии кишечника в цератофриидных головастиках (Anura)» . BMC Биология развития . 20 (1): 16. doi : 10.1186/s12861-020-00221-5 . PMC 7388516 . PMID 32723314 . S2CID 255787646 .
- ^ Балинский, Борис Иван. «Развитие животных: метаморфоза» . Encyclopædia Britannica . Получено 10 августа 2012 года .
- ^ Маттисон, Крис (2011). Лягушки и жабы миров . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. п. 107. ISBN 978-0-691-14968-4 .
- ^ Маттисон, Крис (2011). Лягушки и жабы миров . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. С. 65–68. ISBN 978-0-691-14968-4 .
- ^ Дуэльман, США; Zug, gr "Enra: кормление рассрочки" . Британская апреля Получено 9 ,
- ^ De-Oliveira-Nogueira, Carlos Henrique; Суза, Убират Феррейра; Мачадо, Тейнара Мендес; Figueiredo-de-Andarde, Caio Antônio; Моника, Александр Таманини; Сазима, Иван; Сазима, Марлис; Толедо, Луис Фелипе (июнь 2023 г.). «Между фруктами, цветами и нектаром: необычайная диета лягушки Xenohyla truncata» . Пищевые сети . 35 : E00281. Bibcode : 2023fwebs..3500281d . Doi : 10.1016/j.fooweb.2023.e00281 .
- ^ Да Силва, HR; De Britto-Pereira, XC (2006). Сколько фруктов едят фрукты, которые едят « ? Журнал зоологии . 270 (4): 692–698. doi : 10.1111/j.1469-7998.2006.00192.x .
- ^ «Диета неотропической лягушки Leptodactylus mystaceus» (PDF) . Архивировано из оригинала (PDF) 7 апреля 2014 года . Получено 2 апреля 2014 года .
- ^ Камеры, Бруно Ф.; Крински, Дайоны; Calvo, Изабелла А. (4 февраля 2014 г.). «Диета неотропической лягушки Leptodactylus mystaceus (Anur: Leptodactylidae)» (PDF ) Герпетология заметок 7 : 31–3 Архивировано из оригинала (PDF) 24 сентября , Получено апреля 26 ,
- ^ Дас И. (апрель 1996 г.). ( ) Panura: Ranidae) (PDF . Журнал зоологии 238 (4): 785–7 doi : 10.111/j.1469-7998.1996 .
- ^ Маттисон, Крис (2011). Лягушки и жабы миров . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. п. 69. ISBN 978-0-691-14968-4 .
- ^ Грэм, Донна. «Северный леопардовый лягушка ( Рана Пипинс )» . Руководство педагога по природным ресурсам Южной Дакоты . Архивировано из оригинала 23 июня 2012 года . Получено 4 августа 2012 года .
- ^ Пимм, Стюарт Л. (1979). «Структура пищевых сетей» (PDF) . Теоретическая биология населения . 16 (2): 144–158. Bibcode : 1979tpbio..16..144p . doi : 10.1016/0040-5809 (79) 90010-8 . PMID 538731 . Архивировано из оригинала (PDF) 27 сентября 2011 года.
- ^ Мэтьюз, Кр; Miaud, C. (2007). «Скелелеронологическое исследование возрастной структуры, роста и долговечности горной лягушки с желтой ножкой, Рана Мускоза , в Сьерра-Неваде, Калифорния». Копея . 2007 (4): 986–993. doi : 10.1643/0045-8511 (2007) 7 [986: assota] 2.0.co; 2 . ISSN 0045-8511 . S2CID 86237494 . (требуется подписка)
- ^ Славяны, Фрэнк; Славянс, Кейт. «Индекс лягушки и жабы» . Долговечность . Архивировано из оригинала 18 августа 2012 года . Получено 4 июля 2012 года .
- ^ Storey, KB (1990). «Жизнь в замороженном состоянии: адаптивные стратегии для естественной толерантности к замораживанию у амфибий и рептилий». Американский журнал физиологии . 258 (3 PT 2): 559–568. doi : 10.1152/ajpregu.1990.258.3.r559 . PMID 2180324 .
- ^ Crump, ML (1996). «Родительская помощь среди амфибий». Родительская помощь: эволюция, механизмы и адаптивное значение . Достижения в изучении поведения. Тол. 25. С. 109–144. doi : 10.1016/s0065-3454 (08) 60331-9 . ISBN 978-0-12-004525-9 .
- ^ Jump up to: а беременный в Браун, JL; Моралес, V.; Саммерс, К. (2010). «Ключевая экологическая черта привела к эволюции двупарной помощи и моногамии в амфибии». Американский натуралист . 175 (4): 436–446. doi : 10.1086/650727 . PMID 20180700 . S2CID 20270737 .
- ^ Шеридан, Дженнифер А.; ОКК, Джоан Ф. (2008). «Родительская помощь в хиромантисе Хансене (Anura: Rhacophoridae)». Копея . 2008 (4): 733–736. doi : 10.1643/CH-07-225 . S2CID 85122799 .
- ^ Браун, JL; Моралес, V.; Саммерс, К. (2008a). «Расхождение в родительской помощи, отборе среды обитания и жизненной истории личинок между двумя видами перуанских ядовитых лягушек: экспериментальный анализ». Журнал эволюционной биологии . 21 (6): 1534–1543. doi : 10.1111/j.1420-9101.2008.01609.x . PMID 18811668 . S2CID 29546555 .
- ^ Грант, Т.; Мороз, доктор; Caldwell, JP; Gagliardo, R.; Хаддад, CFB; Кок, PJR; Означает, дБ; Нунан, BP; Шаргель, мы; Wheeler, W. (2006). "Филогенетическая систематика туманных туманов и их родственников (Amphibia, Athesphanaura, Dendrobatidae) (PDF ) Бюллетень Музея музея Американского музея . 299 : 1–2 Citeseerx 10.1.1.693.8 Doi : 10.1206/0003-0090 (2006) 299 [1: psodfa] 2.0.co ; HDL : 2246/5 ISSN 0003-0 S2CID 82263880 Архивировано из оригинала (PDF) 4 апреля , Получено ноября 13 ,
- ^ Ван Вийгаарден, Рене; Боланьос, Федерико (1992). "Родительская помощь в Дендрове Гранульфера . Журнал герпеологии 26 (1): 102–1 doi : 10.2307/1565037 . JSTOR 1565037 .
- ^ Фандиньо, Мария Клаудия; Lüddecke, Horst; Амезкита, Адольфо (1997). «Вокализация и личиночная транспортировка мужского Colostethus subpunctus (Anura: Denrobatidae)». Амфибийская рептилия . 18 (1): 39–48. Doi : 10.1163/156853897x00297 .
- ^ «Обычная акушерка, Алита Абустетрис » . Repties et Amphibiens de France . Получено 30 июля 2012 года .
- ^ «Белки гнезд пены лягушки» . Университет Глазго. Архивировано из оригинала 3 июня 2013 года . Получено 24 августа 2012 года .
- ^ Далгетти, Лора; Кеннеди, Малкольм В. (2010). «Построение дома от пены-архитектура пены из пены Тунгара и трехфазный процесс строительства» . Биологические письма . 6 (3): 293–296. doi : 10.1098/rsbl.2009.0934 . PMC 2880057 . PMID 20106853 .
- ^ " Асса, Дарлингтони " . Фроги Австралийская сеть . 2005 . Получено 5 августа 2012 года .
- ^ Semeyn, E. (2002). " Rheobatrachus silus " . Интернет -разнообразие животных . Музей зоологии Мичиганского университета . Получено 5 августа 2012 года .
- ^ Sandmeier, Fran (12 марта 2001 г.). " Rhinoderma darwinii " . Amphibiaweb . Получено 5 августа 2012 года .
- ^ Barthalmus, George T.; Zielinski, William J. (1988). « Слизь кожи Xenopus индуцирует пероральные дискинезии, которые способствуют побегу от змей». Фармакология Биохимия и поведение . 30 (4): 957–959. doi : 10.1016/0091-3057 (88) 90126-8 . PMID 3227042 . S2CID 25434883 .
- ^ Дарст, Кэтрин Р.; Каммингс, Молли Э. (2006). «Учебное обучение хищников благоприятствует имитации менее токсичной модели в ядовитых лягушках» . Природа . 440 (7081): 208–211. Bibcode : 2006natur.440..208d . doi : 10.1038/nature04297 . PMID 16525472 .
- ^ Майерс, CW; Дейли, JW (1983). "Дарт-поп-лягушки". Scientific American . 248 (2): 120–133. Bibcode : 1983sciam.248b.120M . doi : 10.1038/Scientificamerican0283-120 . PMID 6836257 .
- ^ Savage, JM (2002). Амфибии и рептилии Коста -Рики . Университет Чикагской Прессы. ISBN 978-0-916984-16-8 .
- ^ Duellman, мы (1978). «Биология экваториальной герпетофауны в Амазонском Эквадоре» (PDF) . Университет Канзасского музея естественной истории Разное публикация . 65 Архивировано из оригинала (PDF) 4 июля 2011 года.
- ^ Сапорито, РА; Гарраффо, HM; Доннелли, Массачусетс; Эдвардс, Ал; Longino, JT; Дейли, JW (2004). «Муравьи -формирование: источник членистоногих для алкалоидов пумилиотоксина дендробатидных ядовитых лягушек» . Труды Национальной академии наук . 101 (21): 8045–8050. Bibcode : 2004pnas..101.8045S . doi : 10.1073/pnas.0402365101 . PMC 419554 . PMID 15128938 .
- ^ Смит, Бп; Тайлер, MJ; Kaneko, T.; Гарраффо, HM; Spande, TF; Дейли, JW (2002). «Свидетельство о биосинтезе псевдофринаминовых алкалоидов австралийской миобатрахидной лягушкой ( псевдофрин ) и для секвестрации пищевых пумилитоксинов». Журнал натуральных продуктов . 65 (4): 439–47. doi : 10.1021/np010506a . PMID 11975476 .
- ^ Грант, Т. "Ядовитая лягушка Vivarium" . Американский музей естественной истории . Получено 7 июля 2012 года .
- ^ Арнольд, Николас; Ovenden, Denys (2002). Рептилии и амфибии Британии и Европы . Харпер Коллинз издатели. С. 73–74. ISBN 978-0-00-219964-3 .
- ^ Хейс, Флойд Э. (1989). «Поведение антипредатора недавно метаморфизированных жаб ( Bufo A. Americanus ) во время встреч со змеями подвязки ( Thamnophis s. Sirtalis )». Копея . 1989 (4): 1011–1015. doi : 10.2307/1445987 . JSTOR 1445987 .
- ^ "Причудливые лягушки" . National Geographic Explorer. Архивировано из оригинала 6 марта 2004 года . Получено 13 июля 2012 года .
- ^ Феррелл, Вэнс (4 марта 2012 г.). «Географическое распределение» . Энциклопедия эволюции . Тол. 3. Факты эволюции. Архивировано из оригинала 5 мая 2012 года . Получено 13 июля 2012 года .
- ^ Райан, Пэдди (25 сентября 2011 г.). «История: лягушки» . Энциклопедия Новой Зеландии . Получено 20 августа 2012 года .
- ^ Дал, Крис; Novotny, vojtech; Моравек, Джири; Ричардс, Стивен Дж. (2009). «Бета -разнообразие лягушек в лесах Новой Гвинеи, Амазония и Европы: контрастные тропические и умеренные сообщества». Журнал биогеографии . 36 (5): 896–904. Bibcode : 2009jbiog..36..896d . doi : 10.1111/j.1365-2699.2008.02042.x . S2CID 13067241 .
- ^ « Циклорана платицефала » . Фроги Австралийская сеть. 23 февраля 2005 г. Получено 20 июля 2012 года .
- ^ Hoekstra, JM; Молнар, JL; Дженнингс, М.; Revenga, C.; Spalding, MD; Boucher, TM; Робертсон, JC; Heibel, TJ; Эллисон, К. (2010). «Количество видов амфибий, находящихся под угрозой глобального уровня с помощью пресноводного экорегиона» . Атлас глобального сохранения: изменения, проблемы и возможности изменить ситуацию . Природа охраняемость . Получено 5 сентября 2012 года .
- ^ Стюарт, Sn; Чансон, JS; Кокс, NA; Молодой, будь; Родригес, ASL; Фишман, DL; Уоллер, RW (2004). «Статус и тенденции амфибийного снижения и вымирания по всему миру» (PDF) . Наука . 306 (5702): 1783–1786. Bibcode : 2004sci ... 306.1783s . Citeseerx 10.1.1.225.9620 . doi : 10.1126/science.1103538 . PMID 15486254 . S2CID 86238651 . Архивировано из оригинала (PDF) 27 октября 2017 года . Получено 27 октября 2017 года .
- ^ Фунт, Дж. Алан; Фогден, Майкл П.Л.; Savage, Jay M.; Горман, Джордж С. (1997). «Испытания нулевых моделей на снижение амфибий на тропической горе». Биология сохранения . 11 (6): 1307–1322. Bibcode : 1997conbi..11.1307p . doi : 10.1046/j.1523-1739.1997.95485.x . S2CID 85382659 .
- ^ «Всемирный амфибия снижается: насколько большая проблема, каковы причины и что можно сделать?» Полем Amphibiaweb. 22 января 2009 г. Получено 15 октября 2012 года .
- ^ «Популяция лягушек уменьшается в основном из -за трафика» . Новый ученый . 7 июля 2006 г. Получено 13 июля 2012 года .
- ^ Voordouw, MJ; Адама, Д.; Хьюстон, Б.; Govindarajulu, P. (2010). «Распространенность патогенного гриба хитридов, батрахохитрия дендробатидис , в исчезающем популяции северных лягушек леопардов, Рана Пипинов » . Экология BMC . 10 (6): 6. Bibcode : 2010bmce ... 10 .... 6V . doi : 10.1186/1472-6785-10-6 . PMC 2846871 . PMID 20202208 .
- ^ Арфа, Элизабет М.; Петранка, Джеймс В. (2006). «Ранавирус в деревянных лягушках ( Rana Sylvatica ): потенциальные источники передачи внутри и между прудами» . Журнал болезней дикой природы . 42 (2): 307–318. doi : 10.7589/0090-3558-42.2.307 . PMID 16870853 .
- ^ Филлипс, Кэтрин (1994). Отслеживание исчезающих лягушек . Книги пингвинов. ISBN 978-0-14-024646-9 .
- ^ Lips, Карен Р. (2008). «Упадок тропической монтанской амфибийской фауны». Биология сохранения . 12 (1): 106–117. doi : 10.1111/j.1523-1739.1998.96359.x . JSTOR 2387466 . S2CID 55870526 .
- ^ Blaustein, Andrew R.; Джонсон, Питер TJ (2003). «Сложность деформированных амфибий» (PDF) . Границы в экологии и окружающей среде . 1 (2): 87–94. doi : 10.1890/1540-9295 (2003) 001 [0087: tcoda] 2.0.co; 2 . Архивировано из оригинала (PDF) 29 октября 2013 года.
- ^ Буркхарт, JG; Анкли, Г.; Bell, H.; Carpenter, H.; Форт, Д.; Gardiner, D.; Gardner, H.; Хейл, Р.; Хельген, JC; Jepson, P.; Джонсон, Д.; Lannoo, M.; Ли, Д.; Lary, J.; Levey, R.; Magner, J.; Meteyer, C.; Шелби, доктор медицины; Люсие, Г. (2000). «Стратегии для оценки последствий уродливых лягушек для здоровья окружающей среды» . Перспективы здоровья окружающей среды . 108 (1): 83–90. doi : 10.2307/3454299 . JSTOR 3454299 . PMC 1637865 . PMID 10620528 .
- ^ Блэк, Ричард (2 октября 2005 г.). «Новый лягушащий центр для лондонского зоопарка» . BBC News . Получено 3 ноября 2008 года .
- ^ «Национальный план восстановления для южной лягушки -корбори (Pseudophryne Corroboree): 5. Предыдущие действия по восстановлению» . Environment.gov.au. Архивировано из оригинала 12 мая 2008 года . Получено 3 ноября 2008 года .
- ^ «2008: Год лягушки» . 15 января 2008 года. Архивировано с оригинала 21 апреля 2012 года . Получено 13 июля 2012 года .
- ^ Тайлер, Майкл Дж. (1989). Австралийские лягушки . Книги пингвинов. п. 111. ISBN 978-0-670-90123-4 .
- ^ Кэмерон, Элизабет (6 сентября 2012 г.). "Тростник жабы" . Австралийский музей . Получено 12 сентября 2012 года .
- ^ ̺ Брих, Екатерина (20 января 2009 г.). «Аппетит к ногам лягушек, наносящих вред дикому населению» . ABC News .
- ^ Warkentin, Ig; Bickford, D.; Содхи, NS; Кори, JA (2009). «Еда лягушек до исчезновения». Биология сохранения . 23 (4): 1056–1059. Bibcode : 2009conbi..23.1056w . doi : 10.1111/j.1523-1739.2008.01165.x . PMID 19210303 . S2CID 1837255 .
- ^ «Информационная программа культивированных видов: Рана Кейтсбейана » . ФАО: Департамент рыболовства и аквакультуры . Получено 5 июля 2012 года .
- ^ Райан Шусслер (28 января 2016 г.). «Первая проблема с горной куриной лягушкой: на вкус как…» National Geographic News . Архивировано с оригинала 29 января 2016 года.
- ^ Марк Твен; Чарльз Дадли Уорнер (1904). Писания Марка Твена [псевдоходов.].: Бродяга за границу . Harper & Bros. p. 263.
- ^ «Калифорнийские школы, ведущие гонку, чтобы остановить рассечения» . Институт защиты животных . 25 апреля 2011 г. Получено 17 июня 2012 года .
- ^ Уэллс, Дэвид Эймс (1859). Наука общих вещей: знакомое объяснение первых принципов физической науки. Для школ, семей и молодых учеников . Ивисон, Финни, Блейкман. п. 290.
- ^ «Станниус лигатура» . Биология онлайн . 3 октября 2005 г. Получено 5 августа 2012 года .
- ^ Саркар С. (1996). «Ланселот Хогбен, 1895–1975» . Генетика . 142 (3): 655–660. doi : 10.1093/Genetics/142.3.655 . PMC 1207007 . PMID 8849876 .
- ^ Браунли, Кристен. «Ядерная передача: привлечение клонов» . Труды Национальной академии наук Соединенных Штатов Америки . Архивировано из оригинала 14 октября 2012 года . Получено 21 октября 2012 года .
- ^ Browder, L.; Iten, L., eds. (1998). « Xenopus как модельная система в биологии развития» . Динамическое развитие . Университет Калгари . Получено 17 июня 2012 года .
- ^ Klein, S. «Развитие потенциала Xenopus tropicalis как генетической модели» . Trans-Nih Xenopus Рабочая группа . Получено 9 марта 2006 г.
- ^ «Список генома - геном» . NCBI . Архивировано из оригинала 13 июля 2019 года . Получено 7 апреля 2019 года .
- ^ Vancompernolle, SE; Тейлор, RJ; Освальд-Рихтер, К.; Цзян, Дж.; Твой, будь; Боуи, JH; Тайлер, MJ; Конлон, М.; Уэйд, Д.; и др. (2005). «Антимикробные пептиды из амфибийной кожи мощно ингибируют инфекцию вируса иммунодефицита человека и перенос вируса из дендритных клеток в Т -клетки» . Журнал вирусологии . 79 (18): 11598–11606. doi : 10.1128/JVI.79.18.11598-11606.2005 . PMC 1212620 . PMID 16140737 .
- ^ Phillipe, G.; Ангенот Л. (2005). «Недавние события в области стрелы и ядов Дарт» . Журнал этнофармакологии . 100 (1–2): 85–91. doi : 10.1016/j.jep.2005.05.022 . PMID 15993556 .
- ^ Lyttle, T.; Гольдштейн, Д.; Гарц Дж. (1996). « Bufo Toads и Bufotenine: факт и художественная литература, окружающие предполагаемую психоделическую». Журнал психоактивных лекарств . 28 (3): 267–290. Citeseerx 10.1.1.688.5926 . doi : 10.1080/02791072.1996.10472488 . PMID 8895112 .
- ^ Lyttle, T. (1993). «Неправомерное использование и легенда в феномене" лизания жабы ". Международный журнал зависимости . 28 (6): 521–538. doi : 10.3109/10826089309039645 . PMID 8486435 .
- ^ Jump up to: а беременный Майерс, Чарльз У.; Дейли, Джон У.; Малкин, Борис (1978). «Опасно токсичная новая лягушка ( Phyllobates ), используемая индейцами Emberá of Western Colombia, с обсуждением изготовления выдувки и отравления дротиками». Бюллетень Американского музея естественной истории . 161 : 307–366. HDL : 2246/1286 .
- ^ Маттисон, Крис (2011). Лягушки и жабы миров . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. С. 91–92. ISBN 978-0-691-14968-4 .
- ^ Sleigh, Charlotee (2012). Лягушка Книги Reaktion. С. 40–42. ISBN 978-1-86189-920-0 .
- ^ Sleigh, Charlotee (2012). Лягушка Книги Reaktion. С. 167–175. ISBN 978-1-86189-920-0 .
Дальнейшее чтение
- Белц, Эллин (2005). Лягушки: В их замечательном мире . Книги огня. ISBN 978-1-55297-869-6 .
- Cogger, Hg; Zweifel, RG; Кристиан, Д. (2004). Энциклопедия рептов и амфибийцев (2 -е изд.). Fog City Press. ISBN 978-1-877019-69-2 .
- Estes, R. и OA Reig. (1973). «Ранняя ископаемая запись о лягушках: обзор доказательств». С. 11–63 в JL Vial (ред.), Эволюционная биология ануранов: современные исследования основных проблем . Университет Миссури Пресс, Колумбия.
- Гисси, Кармела; Сан -Мауро, Диего; Песоле, Грациано; Зардоя, Рафаэль (февраль 2006 г.). «Митохондриальная филогения Anura (амфибия): тематическое исследование конгруэнтной филогенетической реконструкции с использованием аминокислотных и нуклеотидных признаков». Ген . 366 (2): 228–237. doi : 10.1016/j.gene.2005.07.034 . PMID 16307849 .
- Холман, JA (2004). Ископаемые лягушки и жабы Северной Америки . Издательство Университета Индианы. ISBN 978-0-253-34280-5 .
- Сан -Мауро, Диего; Венс, Майкл; Alcobendas, Марина; Зардоя, Рафаэль; Мейер, Аксель (май 2005 г.). «Первоначальная диверсификация живых амфибий предшествовала разрыву Пангеи » Американский натуралист 165 (5): 590–5 Doi : 10.1086/ 4 PMID 15795855 S2CID 17021360
- Тайлер, MJ (1994). Австралийские лягушки естественная история . Рид Книги. ISBN 978-0-7301-0468-1 .
Внешние ссылки
- Amphibiaweb
- Галерея лягушек - фотография и изображения различных видов лягушек
- Весь проект лягушки - виртуальная лягушательная рассечение и анатомия
- «Исчезновение жаб, лягушки обеспокоены некоторыми учеными» Сан -Франциско Хроника , 20 апреля 1992 г.
- Амфибийская фотогалерея от научного названия - много необычных лягушек
- Scientific American : исследователи определяют источник ядовитых лягушек, смертельную защиту
СМИ
- Время за плит видео, показывающее развитие яйца до вылупления
- Лягушка вокализации со всего мира - из британской библиотеки
- Лягушатые звонки - из Манитобы , Канада




Существующие семьи Ануран по подройку |
|---|
| Базы данных управления авторитетом : национальный |
|---|