Лягушка
| Лягушки Временной диапазон: ранняя юра – настоящее время ,
| |
|---|---|

| |
| Различные виды лягушек | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Амфибия |
| Клэйд : | заметность |
| Заказ: | Анура Дюмериль , 1806 (как Анурес) |
| Подгруппы | |
|
Посмотреть текст | |
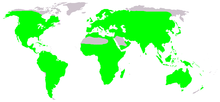
| |
| Местное распространение лягушек (зеленым цветом) | |

Лягушка — это любой представитель разнообразной и в значительной степени плотоядной группы короткотелых бесхвостых земноводных , составляющих отряд Anura. [ 1 ] (происходит от древнегреческого ἀνούρα , буквально «без хвоста»). Самая старая ископаемая «протолягушка» Triadobatrachus известна из раннего триаса Мадагаскара предполагает, что (250 миллионов лет назад ), но датировка по молекулярным часам их отделение от других амфибий может простираться еще дальше, в пермский период , 265 миллионов лет назад. Лягушки широко распространены: от тропиков до субарктических регионов, но наибольшая концентрация видового разнообразия приходится на влажные тропические леса . Лягушки составляют около 88% современных видов земноводных. Они также входят в пятерку самых разнообразных отрядов позвоночных . Виды бородавчатых лягушек обычно называют жабами , но различие между лягушками и жабами носит неформальный характер, а не связано с таксономией или историей эволюции.
У взрослой лягушки крепкое тело, выпученные глаза , прикрепленный вперед язык , загнутые вниз конечности и отсутствие хвоста (хвост у хвостатых лягушек является продолжением клоаки самца). У лягушек железистая кожа, выделения которой варьируются от неприятных до токсичных. Цвет их кожи варьируется от хорошо замаскированных пятнистых коричневых, серых и зеленых до ярких узоров ярко-красного или желтого и черного цветов, демонстрирующих токсичность и отпугивающих хищников . Взрослые лягушки живут в пресной воде и на суше; некоторые виды приспособлены к жизни под землей или на деревьях.
Лягушки обычно откладывают яйца в воду. Из яиц вылупляются водные личинки , называемые головастиками , у которых есть хвосты и внутренние жабры . У них есть узкоспециализированные грубые части рта, подходящие для растительноядных , всеядных или планктоядных диет. Жизненный цикл завершается, когда они превращаются во взрослых особей. Некоторые виды откладывают икру на суше или минуют стадию головастика. Взрослые лягушки обычно имеют плотоядную диету, состоящую из мелких беспозвоночных , но существуют всеядные виды, и некоторые из них питаются растительной пищей. Кожа лягушек имеет богатый микробиом , который важен для их здоровья. Лягушки чрезвычайно эффективно преобразуют то, что они едят, в массу тела. Они являются важным источником пищи для хищников и частью динамики пищевой сети многих мировых экосистем . Кожа полупроницаема , что делает их восприимчивыми к обезвоживанию, поэтому они либо живут во влажных местах, либо имеют специальные приспособления для жизни в сухих местах обитания. Лягушки издают широкий спектр вокализаций , особенно в период размножения. и демонстрируют множество различных видов сложного поведения для привлечения партнеров, защиты от хищников и в целом для выживания.
Люди ценят лягушек как пищу, а также играют важную культурную роль в литературе, символике и религии. Их также считают экологическими лидерами , а сокращение популяции лягушек часто рассматривается как ранний признак экологического ущерба. Популяция лягушек значительно сократилась с 1950-х годов. Считается, что более трети видов находятся под угрозой исчезновения , и более 120 видов вымерли с 1980-х годов. Число пороков развития среди лягушек растет, а новое грибковое заболевание хитридиомикоз распространилось по всему миру. Биологи-природоохранники работают над пониманием причин этих проблем и их решением.
Этимология и таксономия
Использование общих названий лягушка и жаба не имеет таксономического обоснования. С точки зрения классификации все представители отряда Anura являются лягушками, но только представители семейства Bufonidae считаются «настоящими жабами». Использование термина «лягушка» в общих названиях обычно относится к видам, которые являются водными или полуводными и имеют гладкую влажную кожу; Термин «жаба» обычно относится к наземным видам с сухой бородавчатой кожей. [ 2 ] [ 3 ] Из этого правила имеется множество исключений. Европейская огнебрюхая жаба ( Bombina Bombina ) имеет слегка бородавчатую кожу и предпочитает водную среду обитания. [ 4 ] тогда как панамская золотая лягушка ( Atelopus zeteki ) принадлежит к семейству жаб Bufonidae и имеет гладкую кожу. [ 5 ]
Этимология
Происхождение названия отряда Anura - и его первоначального написания Anoures - является древнегреческим приватным альфа- префиксом ἀν- ( an- от ἀ- перед гласной) «без», [ 6 ] и οὐρά ( ourá ) «хвост животного». [ 7 ] означает «бесхвостый». Это относится к бесхвостому характеру этих амфибий. [ 8 ] [ 9 ] [ 10 ]
Происхождение слова « лягушка» неясно и спорно. [ 11 ] Это слово впервые встречается в древнеанглийском языке как frogga , но обычное древнеанглийское слово для обозначения лягушки было frosc (с такими вариантами, как frox и forsc ), и принято считать, что слово frog каким-то образом связано с этим словом. Древнеанглийский frosc оставался в диалектном использовании в английском языке как frosh и frosk до девятнадцатого века. [ 12 ] и широко распространен в других германских языках , с примерами в современных языках, включая немецкий Frosch , норвежский frosk , исландский froskur и голландский (kik)vors . [ 11 ] Эти слова позволяют реконструировать общегерманского предка * froskaz . [ 13 ] В третьем издании Оксфордского словаря английского языка обнаруживается, что этимология * froskaz неясна, но соглашаются с аргументами о том, что оно вполне может происходить от протоиндоевропейской основы, подобной * preu , что означает «прыжок». [ 11 ]
Однако неясно, как древнеанглийское frosc привело к появлению frogga , поскольку это развитие не предполагает регулярного изменения звука . Вместо этого, похоже, в древнеанглийском языке существовала тенденция создавать прозвища для животных, оканчивающиеся на -g , с примерами (которые сами по себе имеют неопределенную этимологию), включая собаку , свинью , свинью, оленя и (ухо)парик . Frog , похоже, был адаптирован из Frosc как часть этой тенденции. [ 11 ]
Между тем, слово жаба , впервые засвидетельствованное как древнеанглийское tādige , уникально для английского языка и также имеет неопределенную этимологию. [ 14 ] Это основа слова головастик , впервые засвидетельствованного как среднеанглийское taddepol , что, очевидно, означает «голова жабы». [ 15 ]
Таксономия
Около 88% земноводных видов относятся к отряду Anura. [ 16 ] Они включают более 7600 видов. [ 1 ] в 55 семействах , из которых видами Hylidae (1049 видов), Strabomantidae (797 видов), Microhylidae (744 вида) и Bufonidae (646 видов) наиболее богаты . [ 17 ]

К ануре относятся все современные лягушки и любые ископаемые виды, подпадающие под определение бесхвостых. Характеристики взрослых бесхвостых животных включают: 9 или меньше пресакральных позвонков, наличие уростиля, образованного из сросшихся позвонков, отсутствие хвоста, длинную и наклоненную вперед подвздошную кость, более короткие передние конечности, чем задние, сросшиеся лучевую и локтевую кости , голени и малоберцовые кости сросшиеся . , удлиненные кости лодыжки , отсутствие предлобной кости, наличие подъязычной пластинки , нижняя челюсть без зубов (за исключением Gastrotheca guentheri отсутствует ), состоящая из трёх пар костей (ангуло-сплениальной, зубной и ментомеккелевой, причем последняя пара у Pipoidea ), [ 18 ] язык без опоры, лимфатические пространства под кожей и мышца, транспортир хрусталика, прикрепленная к хрусталику глаза . [ 19 ] Личинка бесхвостого животного, или головастик, имеет одно центральное дыхательное дыхалец и ротовой аппарат, состоящий из ороговевших клювов и зубчиков . [ 19 ]

Лягушки и жабы в целом подразделяются на три подотряда: Archaeobatrachia , который включает четыре семейства примитивных лягушек; Mesobatrachia , включающая пять семейств более эволюционно промежуточных лягушек; и Neobatrachia , безусловно, самая большая группа, которая содержит оставшиеся семейства современных лягушек, включая наиболее распространенные виды во всем мире. Подотряд Neobatrachia делится на два надсемейства Hyloidea и Ranoidea . [ 20 ] Эта классификация основана на таких морфологических признаках, как количество позвонков, строение грудного пояса , морфология головастиков. Хотя эта классификация широко принята, отношения между семействами лягушек все еще обсуждаются. [ 21 ]
Некоторые виды бесхвостых легко гибридизуются . Например, съедобная лягушка ( Pelophylax esculentus ) представляет собой гибрид прудовой лягушки ( P. Lessae ) и болотной лягушки ( P.ridibundus ). [ 22 ] Огнебрюхие жабы Bombina Bombina и B. variegata сходны в образовании гибридов. Они менее плодовиты, чем их родители, что приводит к образованию гибридной зоны , где гибриды преобладают. [ 23 ]
Эволюция
Происхождение и эволюционные взаимоотношения между тремя основными группами земноводных горячо обсуждаются. Молекулярная филогения, основанная на анализе рДНК , датированном 2005 годом, предполагает, что саламандры и червячие более тесно связаны друг с другом, чем с лягушками, и расхождение этих трех групп произошло в палеозое или раннем мезозое , до распада суперконтинента Пангея и вскоре после их отделения от лопастеперых рыб . Это могло бы помочь объяснить относительную нехватку окаменелостей земноводных периода до разделения групп. [ 24 ] Другой молекулярно-филогенетический анализ, проведенный примерно в то же время, пришел к выводу, что лиссамфибии впервые появились около 330 миллионов лет назад и что гипотеза темноспондильного происхождения более достоверна, чем другие теории. Необатрахи , по-видимому, произошли из Африки/Индии, саламандры – из Восточной Азии, а червяги – из тропической Пангеи. [ 25 ] Другие исследователи, хотя и согласились с основной целью этого исследования, поставили под сомнение выбор точек калибровки, используемых для синхронизации данных. Они предложили относить дату диверсификации лисамфибий к перми , то есть менее 300 миллионов лет назад, — дату, которая лучше согласуется с палеонтологическими данными. [ 26 ] Дальнейшее исследование, проведенное в 2011 году с использованием как вымерших, так и ныне живущих таксонов, отобранных для получения морфологических, а также молекулярных данных, пришло к выводу, что Lissamphibia является монофилетическим и что она должна быть вложена в Lepospondyli, а не в Temnospondyli . Исследование постулировало, что лиссамфибии возникли не ранее позднего каменноугольного периода , примерно 290–305 миллионов лет назад. По оценкам, раскол между Anura и Caudata произошел 292 миллиона лет назад, что гораздо позже, чем предполагают большинство молекулярных исследований: червячие отделились 239 миллионов лет назад. [ 27 ]

В 2008 году был обнаружен Gerobatrachus hottoni , темноспондил со многими характеристиками, напоминающими лягушку и саламандру в Техасе . Он датируется 290 миллионами лет назад и был провозглашен недостающим звеном , стволовым батрахием, близким к общему предку лягушек и саламандр, что согласуется с широко распространенной гипотезой о том, что лягушки и саламандры более тесно связаны друг с другом (образуя кладу под названием Batrachia). ), чем для червяг. [ 28 ] [ 29 ] Однако другие предположили, что Gerobatrachus hottoni был всего лишь диссорофоидным темноспондилом, не связанным с современными амфибиями. [ 30 ]
Salientia (лат. salire ( salio ), «прыгать») — название всей группы, в которую входят современные лягушки отряда Anura, а также их близкие ископаемые родственники, «протолягушки» или «стволовые лягушки». К общим чертам, которыми обладают эти протолягушки, относятся 14 прекрестцовых позвонков (у современных лягушек их восемь или 9), длинная и наклоненная вперед подвздошная кость в области таза , наличие лобно-теменной кости и нижняя челюсть без зубов. Самыми ранними известными земноводными, которые были более тесно связаны с лягушками, чем с саламандрами, являются Triadobatrachus Massinoti из раннего триасового периода Мадагаскара (около 250 миллионов лет назад) и Czatkobatrachus polonicus из раннего триаса в Польше (примерно того же возраста, что и Triadobatrachus). ). [ 31 ] Череп Triadobatrachus похож на лягушку: он широкий, с большими глазницами, но ископаемое имеет черты, отличные от черепа современных лягушек. К ним относятся более длинное тело с большим количеством позвонков . В хвосте имеются отдельные позвонки, в отличие от сросшихся уростиля или копчика у современных лягушек. Кости большеберцовой и малоберцовой костей также разделены, что делает вероятным, что Triadobatrachus не был хорошим прыгуном. [ 31 ] Исследование 2019 года отметило присутствие Salientia из формации Чинле и предположило, что бесхвостые животные могли впервые появиться в позднем триасе . [ 32 ]
Судя по ископаемым свидетельствам, самые ранние известные «настоящие лягушки», относящиеся к собственно линии бесхвостых, жили в раннем юрском периоде. [ 2 ] [ 33 ] Один из таких ранних видов лягушек, Prosalirus bitis , был обнаружен в 1995 году в формации Кайента в Аризоне и относится к раннему юрскому периоду (от 199,6 до 175 миллионов лет назад), что делает Prosalirus несколько более поздним, чем Triadobatrachus . [ 34 ] Как и последний, Prosalirus не имел сильно увеличенных ног, но имел типичное трехлапчатое строение таза современных лягушек. В отличие от Triadobatrachus , Просалирус уже потерял почти весь свой хвост. [ 35 ] и был хорошо приспособлен к прыжкам. [ 36 ] Другая раннеюрская лягушка — Vieraella Hersti , известная только по дорсальным и вентральным отпечаткам одного животного и, по оценкам, 33 мм ( 1 + 1 ⁄ дюйма ) от носика до вентиляционного отверстия. Notobatrachus degiustoi из средней юры немного моложе, около 155–170 миллионов лет. Основные эволюционные изменения этого вида заключались в укорочении тела и потере хвоста. Эволюция современной Ануры, вероятно, завершилась к юрскому периоду. С тех пор эволюционные изменения числа хромосом у млекопитающих происходили примерно в 20 раз быстрее, чем у лягушек, а это означает, что видообразование у млекопитающих происходит быстрее. [ 37 ]
Согласно генетическим исследованиям, семейства Hyloidea , Microhylidae и клада Natatanura (составляющая около 88% ныне живущих лягушек) диверсифицировались одновременно около 66 миллионов лет назад, вскоре после мел -палеогенового вымирания, связанного с ударным камнем Чиксулуб . Все истоки древесной растительности (например, в Hyloidea и Natatanura) проистекают из этого времени и произошедшего впоследствии возрождения леса. [ 38 ] [ 39 ]
Окаменелости лягушек были найдены на всех континентах Земли. [ 40 ] [ 41 ] В 2020 году было объявлено, что шлемовидных лягушек обнаружила окаменелости группа палеонтологов-позвоночных палеонтологов на острове Сеймур на Антарктическом полуострове возрастом 40 миллионов лет , что указывает на то, что этот регион когда-то был домом для лягушек, родственных тем, что сейчас живут в южноамериканских Nothofagus лесах . . [ 42 ]
Филогения
Кладограмму , показывающую родство различных семейств лягушек клады Anura, можно увидеть в таблице ниже. Эта диаграмма в виде дерева показывает, как каждое семейство лягушек связано с другими семействами, причем каждый узел представляет собой точку общего происхождения. Он основан на Frost et al. (2006), [ 43 ] Хейнике и др. (2009) [ 44 ] и Пайрон и Винс (2011). [ 45 ]
| Анура | |
Морфология и физиология

У лягушек нет хвоста, за исключением личинок, и у большинства из них длинные задние ноги, удлиненные лодыжки, перепончатые пальцы, нет когтей, большие глаза и гладкая или бородавчатая кожа. У них короткие позвоночные столбы, не более 10 свободных позвонков и сросшиеся копчики (уростиль или копчик). [ 46 ] Размер лягушек варьируется от Paedophryne amauensis из Папуа-Новой Гвинеи составляет 7,7 мм (0,30 дюйма). , длина морды до отверстия [ 47 ] размером примерно до 35 см (14 дюймов) и весом 3,3 кг (7,3 фунта) . до лягушки-голиафа ( Conraua goliath ) из Центральной Африки [ 48 ] Существуют доисторические, вымершие виды, достигавшие еще больших размеров. [ 49 ]
Ноги и ноги
Строение ступней и ног сильно различается у разных видов лягушек, отчасти в зависимости от того, живут ли они преимущественно на земле, в воде, на деревьях или в норах. У взрослых бесхвостых животных четыре пальца на руках и пять пальцев на ногах. [ 50 ] но у самых маленьких видов часто есть руки и ноги, на которых некоторые пальцы рудиментарны. [ 51 ] Лягушки должны уметь быстро перемещаться по окружающей среде, чтобы ловить добычу и убегать от хищников, и в этом им помогают многочисленные приспособления. Большинство лягушек либо умеют прыгать, либо произошли от их предков, при этом большая часть опорно-двигательного аппарата морфологии была модифицирована для этой цели. Большеберцовая, малоберцовая и предплюсны срослись в одну прочную кость , как и лучевая и локтевая кости передних конечностей (которые должны поглощать удар при приземлении). удлинились Плюсневые кости , чтобы увеличить длину ног и позволить лягушкам дольше прижиматься к земле при взлете. удлинилась Подвздошная кость и образовала подвижный сустав с крестцом , который у прыгунов-специалистов, таких как раниды и гилиды, действует как дополнительный сустав конечностей, обеспечивающий дополнительную мощность прыжков. Хвостовые позвонки срослись в уростиль, который втягивается внутрь таза. Это позволяет передавать силу от ног к телу во время прыжка. [ 46 ]

( Рана временная )

Мышечная система подверглась аналогичным изменениям. Задние конечности предков лягушек, по-видимому, содержали пары мышц, которые действовали противоположно (одна мышца сгибала колено, другая — разгибала его), как это наблюдается у большинства других конечностей животных. Однако у современных лягушек почти все мышцы были модифицированы, чтобы способствовать прыжку, и осталось лишь несколько мелких мышц, которые возвращают конечность в исходное положение и поддерживают позу. Мышцы также значительно увеличены: на основные мышцы ног приходится более 17% общей массы лягушек. [ 52 ]
У многих лягушек лапы перепончатые, и степень перепонок прямо пропорциональна количеству времени, которое вид проводит в воде. [ 53 ] У полностью водной африканской карликовой лягушки ( Hymenochirus sp.) пальцы ног полностью перепончатые, тогда как у древесной лягушки Уайта ( Litoria caerulea ), древесного вида, перепончатые пальцы лишь на четверть или наполовину. [ 54 ] Исключение составляют летающие лягушки Hylidae , у которых также полностью перепончатые пальцы ног , и Rhacophoridae используемые при планировании.
У древесных лягушек на концах пальцев ног есть подушечки, помогающие захватывать вертикальные поверхности. Это не присоски, поверхность которых состоит из столбчатых ячеек с плоскими вершинами и небольшими промежутками между ними, смазанных слизистыми железами. Когда лягушка оказывает давление, клетки прилипают к неровностям на поверхности, и сцепление сохраняется за счет поверхностного натяжения . Это позволяет стрелке карабкаться по гладким поверхностям, но система не работает эффективно, когда подушечки чрезмерно влажные. [ 55 ]
У многих древесных лягушек небольшая «вставочная структура» на каждом пальце увеличивает площадь поверхности, соприкасающейся с субстратом . Кроме того, у многих древесных лягушек есть тазобедренные суставы, позволяющие как прыгать, так и ходить. У некоторых лягушек, живущих высоко на деревьях, между пальцами ног даже имеются сложные перепонки. Это позволяет лягушкам «прыгать с парашютом» или контролируемо планировать из одного положения купола в другое. [ 56 ]
Наземным лягушкам обычно не хватает приспособлений, присущих водным и древесным лягушкам. У большинства из них подушечки пальцев меньше, если таковые имеются, и небольшая перепонка. Некоторые роющие лягушки, такие как чесночница Коуча ( Scaphiopus Couchii ), имеют на задних лапах отросток в виде лоскута, ороговевший бугорок , часто называемый лопатой, который помогает им рыть норы. [ 57 ]
Иногда на стадии головастика одну из развивающихся задних ног съедает такой хищник, как нимфа стрекозы . В некоторых случаях полная нога все же растет, а в других – нет, хотя лягушка все еще может прожить свою нормальную жизнь, имея всего три конечности. Иногда паразитический плоский червь ( Ribeiroia ondatrae ) впивается в заднюю часть головастика, вызывая перестановку клеток зачатков конечностей, и у лягушки развиваются одна или несколько дополнительных ног. [ 58 ]

Кожа
Кожа лягушки является защитной, выполняет дыхательную функцию, поглощает воду и помогает контролировать температуру тела. Имеет множество желез, особенно на голове и спине, которые часто выделяют неприятные и ядовитые вещества ( зернистые железы ). Секрет часто липкий и помогает сохранять кожу влажной, защищает от проникновения плесени и бактерий, делает животное скользким и более способным убежать от хищников. [ 59 ] Кожа сбрасывается каждые несколько недель. Обычно он разделяется посередине спины и живота, и лягушка освобождает руки и ноги. Затем очищенную кожу подносят к голове, где ее быстро съедают. [ 60 ]
Будучи хладнокровными, лягушки должны адаптироваться к соответствующим моделям поведения, чтобы регулировать свою температуру. Чтобы согреться, они могут переместиться на солнце или на теплую поверхность; если они перегреются, они могут уйти в тень или занять позицию, при которой воздуху подвергается воздействию минимальной площади кожи. Эта поза также используется для предотвращения потери воды и предполагает, что лягушка сидит на корточках близко к субстрату, поджимая руки и ноги под подбородок и тело. [ 61 ] Цвет кожи лягушки используется для терморегуляции. В прохладную влажную погоду цвет будет темнее, чем в жаркий сухой день. Серая пенногнездовая древесная лягушка ( Chiromantis xerampelina ) способна даже побелеть, чтобы свести к минимуму вероятность перегрева. [ 62 ]
Многие лягушки способны поглощать воду и кислород непосредственно через кожу, особенно в области таза, но проницаемость кожи лягушки также может привести к потере воды. Железы, расположенные по всему телу, выделяют слизь, которая помогает сохранять кожу влажной и уменьшает испарение. Некоторые железы на руках и груди мужчин специализируются на выработке липкого секрета, помогающего при амплексусе . Подобные железы у квакш вырабатывают клейкое вещество на клейких дисках лапок. Некоторые древесные лягушки сокращают потерю воды благодаря водонепроницаемому слою кожи, а некоторые южноамериканские виды покрывают свою кожу восковым секретом. Другие лягушки научились экономить воду, в том числе стали вести ночной образ жизни и отдыхать в положении, позволяющем экономить воду. Некоторые лягушки также могут отдыхать большими группами, прижимая каждую лягушку к соседям. Это уменьшает количество кожи, подвергающейся воздействию воздуха или сухой поверхности, и, таким образом, уменьшает потерю воды. [ 61 ] Жаба Вудхауса ( Bufo woodhousii ), если после содержания в сухом месте получить доступ к воде, сидит на мелководье для регидратации. [ 63 ] У самца волосатой лягушки ( Trichobatrachus Robustus ) на нижней части спины и бедрах выступают кожные сосочки , придающие ему щетинистый вид. Они содержат кровеносные сосуды и, как полагают, увеличивают площадь кожи, доступную для дыхания. [ 64 ]


У некоторых видов есть костные пластины, встроенные в кожу, и эта черта, по-видимому, несколько раз развивалась независимо. [ 65 ] У некоторых других видов кожа на макушке уплотнена и соединительная ткань дермы ко-окостеневает с костями черепа ( экзостоз ). [ 66 ] [ 67 ]
Камуфляж — распространенный защитный механизм у лягушек. Такие особенности, как бородавки и кожные складки, обычно присущи наземным лягушкам, для которых гладкая кожа не обеспечивает столь эффективный камуфляж. Некоторые лягушки меняют цвет днем и ночью, поскольку свет и влага стимулируют пигментные клетки и заставляют их расширяться или сжиматься. [ 68 ] Некоторые даже способны контролировать текстуру своей кожи. [ 69 ] Тихоокеанская квакша ( Pseudacris regilla ) имеет зеленую и коричневую окраску, однотонную или пятнистую, меняет цвет в зависимости от времени года и общего цвета фона. [ 70 ] Лесная лягушка ( Lithobates sylvaticus ) имеет разрушительную окраску , включая черные отметины под глазами, похожие на пустоты между листьями, полосы спинной кожи (дорсолатеральная кожная складка), похожие на среднюю жилку листа , а также пятна, пятна и полосы на ногах, похожие на особенности опавших листьев.
Дыхание и кровообращение
Как и у других земноводных, кислород может проходить через их высокопроницаемую кожу. Эта уникальная особенность позволяет им оставаться в местах без доступа воздуха, дыша через кожу. Ребер обычно нет, поэтому легкие наполняются за счет щечной перекачки , и лягушка, лишенная легких, может поддерживать функции своего тела без них. [ 68 ] Полностью водная борнейская плоскоголовая лягушка ( Barborula kalimantanensis ) — первая лягушка, у которой полностью отсутствуют легкие. [ 71 ]
У лягушек трехкамерное сердце , что у них общее с ящерицами . Кислородная кровь из легких и деоксигенированная кровь из дышащих тканей поступает в сердце через отдельные предсердия . Когда эти камеры сокращаются, два потока крови проходят в общий желудочек , а затем перекачиваются через спиральный клапан в соответствующий сосуд: аорту для насыщенной кислородом крови и легочную артерию для дезоксигенированной крови. [ 72 ]
У некоторых видов лягушек есть приспособления, которые позволяют им выживать в воде с дефицитом кислорода. ( Водяная лягушка Титикака Telmatobius culeus ) является одним из таких видов и имеет морщинистую кожу, которая увеличивает площадь ее поверхности для улучшения газообмена. Обычно он не использует свои рудиментарные легкие, но иногда, находясь на дне озера, ритмично поднимает и опускает свое тело, чтобы увеличить поток воды вокруг него. [ 73 ]

Пищеварение и выведение
У лягушек есть верхнечелюстные зубы вдоль верхней челюсти, которые используются для удержания пищи перед ее проглатыванием. Эти зубы очень слабы, и их нельзя использовать для жевания, ловли и нанесения вреда подвижной добыче. Вместо этого лягушка использует свой липкий раздвоенный язык, чтобы ловить насекомых и другую мелкую движущуюся добычу. Язык обычно лежит свернутым во рту, свободным сзади и прикрепленным к нижней челюсти спереди. Его можно выдвигать и втягивать на огромной скорости. [ 53 ] У амфибий на языке имеются слюнные железы, которые у лягушек вырабатывают так называемую двухфазную вязкоупругую жидкость. Под воздействием давления, например, когда язык обхватывает добычу, он становится жидким и покрывает тело жертвы. Когда давление падает, он возвращается в густое и эластичное состояние, что обеспечивает дополнительный захват языка. [ 74 ] У некоторых лягушек нет языка, и они просто запихивают пищу в рот руками. [ 53 ] Африканская лягушка-бык ( Pyxicephalus ), которая охотится на относительно крупных животных, таких как мыши и другие лягушки, имеет конусообразные костные выступы, называемые зубчатыми отростками, в передней части нижней челюсти, которые функционируют как зубы. [ 16 ] Глаза помогают глотать пищу, поскольку их можно втянуть через отверстия в черепе и помочь протолкнуть пищу в горло. [ 53 ] [ 75 ]
Затем пища попадает через пищевод в желудок, где добавляются пищеварительные ферменты и перемешиваются. Затем он поступает в тонкую кишку (двенадцатиперстную и подвздошную кишку), где происходит основная часть пищеварения. Панкреатический сок поджелудочной железы и желчь, вырабатываемая печенью и хранящаяся в желчном пузыре, секретируются в тонкую кишку, где жидкости переваривают пищу и всасываются питательные вещества. Остатки пищи попадают в толстый кишечник, где удаляется лишняя вода, а отходы выводятся через клоаку . [ 76 ]
Хотя лягушки адаптированы к наземной жизни, они напоминают пресноводных рыб своей неспособностью эффективно сохранять воду в организме. Когда они находятся на суше, много воды теряется за счет испарения с кожи. Выделительная система аналогична системе млекопитающих, имеются две почки , удаляющие азотистые продукты из крови. Лягушки производят большое количество разбавленной мочи, чтобы вымывать токсичные продукты из почечных канальцев. [ 77 ] Азот выделяется в виде аммиака головастиками и водными лягушками, но в основном в виде мочевины , менее токсичного продукта, большинством наземных взрослых особей. Несколько видов древесных лягушек, имеющих ограниченный доступ к воде, выделяют еще менее токсичную мочевую кислоту . [ 77 ] Моча по парным мочеточникам попадает в мочевой пузырь , из которого периодически выбрасывается в клоаку. Все отходы организма выходят из организма через клоаку, которая заканчивается клоакальным отверстием. [ 78 ]
Репродуктивная система
У самца лягушки два семенника прикреплены к почкам, и сперма поступает в почки через тонкие трубочки, называемые выносящими протоками . Затем он проходит через мочеточники, которые, следовательно, известны как мочеполовые протоки. Пениса нет, и сперма выбрасывается из клоаки прямо на яйцеклетки, когда самка их откладывает. Яичники самки лягушки находятся рядом с почками, и яйца проходят через пару яйцеводов и через клоаку наружу. [ 78 ]
Когда лягушки спариваются, самец забирается на спину самки и обхватывает ее передними конечностями либо позади передних ног, либо прямо перед задними. Эта позиция называется амплексусом и может удерживаться несколько дней. [ 79 ] Самец лягушки имеет определенные гормонозависимые вторичные половые признаки . К ним относятся появление у него на больших пальцах в период размножения специальных подушечек, обеспечивающих надежную фиксацию. [ 80 ] Захват самца лягушки во время амплексуса стимулирует самку выпускать икру, обычно завернутую в желе, в качестве икры. У многих видов самец меньше и стройнее самки. Самцы имеют голосовые связки и издают различные кваканья, особенно в период размножения, а у некоторых видов у них также есть голосовые мешочки для усиления звука. [ 78 ]
Нервная система
У лягушек высокоразвитая нервная система, состоящая из головного, спинного мозга и нервов. Многие части мозга лягушки соответствуют мозгу человека. Он состоит из двух обонятельных долей, двух полушарий головного мозга, шишковидного тела, двух зрительных долей, мозжечка и продолговатого мозга. Мышечная координация и осанка контролируются мозжечком , а продолговатый мозг регулирует дыхание, пищеварение и другие автоматические функции. Относительный размер головного мозга лягушек намного меньше, чем у человека. У лягушек есть десять пар черепно-мозговых нервов , которые передают информацию извне непосредственно в мозг, и десять пар спинномозговых нервов , которые передают информацию от конечностей в мозг через спинной мозг. [ 78 ] Напротив, все амниоты (млекопитающие, птицы и рептилии) имеют двенадцать пар черепных нервов. [ 81 ]

Взгляд
Глаза у большинства лягушек расположены по обе стороны головы ближе к макушке и выступают наружу в виде полусферических выпуклостей. Они обеспечивают бинокулярное зрение в поле зрения 100° вперед и общее поле зрения почти 360°. [ 82 ] Они могут быть единственной частью погруженной в воду лягушки, которая выступает из воды. Каждый глаз имеет закрывающиеся верхние и нижние веки и мигательную перепонку , обеспечивающую дополнительную защиту, особенно когда лягушка плавает. [ 83 ] У представителей водного семейства Pipidae глаза расположены на макушке головы, и это положение лучше подходит для обнаружения добычи в воде над головой. [ 82 ] Радужки . бывают разных цветов, а зрачки — разной формы У обыкновенной жабы ( Bufo bufo ) радужка золотистая и горизонтальные щелевидные зрачки, у красноглазой квакши ( Agalychnis callidryas ) зрачки вертикальные, у древолаза темные ирисы, у огнебрюхой лягушки ( Bombina spp. ) имеет треугольные зрачки, а у томатной лягушки ( Dyscophus spp. ) — круглые. Радужная оболочка южной жабы ( Anaxyrus terrestris ) имеет рисунок, гармонирующий с окружающей замаскированной кожей. [ 83 ]
Видение вдаль у лягушки лучше, чем вблизи. Зовущие лягушки быстро замолкают, когда видят злоумышленника или даже движущуюся тень, но чем ближе объект, тем хуже его видно. [ 83 ] Когда лягушка высовывает язык, чтобы поймать насекомое, она реагирует на небольшой движущийся объект, который она плохо видит и должна заранее точно выровнять его, потому что она закрывает глаза, когда язык вытянут. [ 53 ] Хотя раньше это обсуждалось, [ 84 ] более поздние исследования показали, что лягушки могут видеть цвета даже при очень слабом освещении. [ 85 ]
Слух
Лягушки слышат как в воздухе, так и под водой. У них нет внешних ушей ; барабанные перепонки ( барабанная перепонка ) обнажены непосредственно или могут быть покрыты слоем кожи и видны как круглая область сразу за глазом. Размер и расстояние между барабанными перепонками связаны с частотой и длиной волны, на которой кричит лягушка. У некоторых видов, таких как лягушка-бык, размер барабанной перепонки указывает на пол лягушки; у мужчин барабанные перепонки больше, чем их глаза, а у женщин глаза и барабанные перепонки примерно одинакового размера. [ 86 ] Шум вызывает вибрацию барабанной перепонки, и звук передается в среднее и внутреннее ухо. Среднее ухо содержит полукружные каналы, которые помогают контролировать баланс и ориентацию. Во внутреннем ухе слуховые волосковые клетки расположены в двух областях улитки: базилярном сосочке и сосочке амфибии. Первый обнаруживает высокие частоты, а второй – низкие. [ 87 ] Поскольку улитка короткая, лягушки используют электрическую настройку , чтобы расширить диапазон слышимых частот и помочь различать различные звуки. [ 88 ] Такое расположение позволяет обнаруживать территориальные и племенные сигналы своих сородичей . У некоторых видов, обитающих в засушливых регионах, звук грома или сильного дождя может вывести их из состояния покоя. [ 87 ] Лягушка может испугаться неожиданного шума, но обычно не предпринимает никаких действий, пока не обнаружит источник звука визуально. [ 86 ]
Вызов

Крик или кваканье лягушки уникален для ее вида. Лягушки создают этот звук, пропуская воздух через гортань в горло. У большинства кричащих лягушек звук усиливается одним или несколькими голосовыми мешками, кожными перепонками под горлом или в углах рта, которые раздуваются во время усиления крика. Крики некоторых лягушек настолько громкие, что их можно услышать на расстоянии до 1,6 км. [ 89 ] Кроме того, было обнаружено, что некоторые виды используют искусственные конструкции, такие как водосточные трубы, для искусственного усиления своего крика. [ 90 ] Прибрежная хвостатая лягушка ( Ascaphus truei ) обитает в горных ручьях Северной Америки и не издает вокала. [ 91 ]
Основная функция призыва самцов лягушек – привлечение самок. Самцы могут кричать индивидуально или может раздаваться хор звуков, когда многочисленные самцы собираются в местах размножения. У многих видов лягушек, таких как обыкновенная квакша ( Polypedates leucomystax ), самки отвечают на призывы самцов, что усиливает репродуктивную активность в гнездовой колонии. [ 92 ] Самки лягушек предпочитают самцов, издающих звуки большей интенсивности и более низкой частоты, что выделяет их из толпы. Считается, что объяснением этого является то, что, демонстрируя свое мастерство, самец показывает свою способность производить превосходное потомство. [ 93 ]
Другой крик издает самец лягушки или невосприимчивая самка, когда на нее садится другой самец. Это отчетливый стрекочущий звук, сопровождающийся вибрацией тела. [ 94 ] У древесных лягушек и некоторых неводных видов есть призыв к дождю, который они издают на основе сигналов влажности перед ливнем. [ 94 ] У многих видов также есть территориальный призыв, который используется для отпугивания других самцов. Все эти призывы издаются при закрытой пасти лягушки. [ 94 ] Сигнал бедствия, издаваемый некоторыми лягушками, когда они в опасности, издается с открытым ртом, что приводит к более высокому тону сигнала. Обычно он используется, когда лягушку схватил хищник, и может отвлечь или дезориентировать нападавшего, чтобы тот выпустил лягушку. [ 94 ]
У многих видов лягушек глубокие крики. Кваканье американской лягушки-быка ( Rana catesbiana ) иногда пишется как «кувшин с ромом». [ 95 ] Тихоокеанская древесная лягушка ( Pseudacris regilla ) производит звукоподражательное «ребро», которое часто можно услышать в фильмах. [ 96 ] Другие варианты перевода лягушачьих призывов в речь включают «brekekekex koax koax», зов болотной лягушки ( Pelophylax ridebundus ) в «Лягушках» , древнегреческой комической драме Аристофана . [ 97 ] Крики вогнутоухой речной лягушки ( Amolops tormotus ) во многом необычны. Самцы отличаются разнообразием криков, в которых имеют место модуляции частоты вверх и вниз. Когда они общаются, они издают звуки, попадающие в диапазон ультразвуковых частот. Последний аспект, который делает необычными крики этого вида лягушек, заключается в том, что нелинейные акустические явления являются важными компонентами их акустических сигналов. [ 98 ]
оцепенение
В экстремальных условиях некоторые лягушки впадают в состояние оцепенения и остаются неактивными в течение нескольких месяцев. В более холодных регионах многие виды лягушек впадают в спячку зимой . Те, кто живет на суше, например, американская жаба ( Bufo americanus ), роют нору и устраивают зимнюю спячку , в которой могут лежать в спячке . Другие, менее искусные в копании, находят расщелину или зарываются в опавшие листья. Водные виды, такие как американская лягушка-бык ( Rana catesbeiana ), обычно опускаются на дно пруда, где они лежат, полупогруженные в ил, но все еще способные получить доступ к растворенному в воде кислороду. Их метаболизм замедляется, и они живут за счет своих энергетических запасов. Некоторые лягушки, такие как лесная лягушка , болотная лягушка или весенний глядец , могут даже выжить в замороженном состоянии. Кристаллы льда образуются под кожей и в полостях тела, но важные органы защищены от замерзания высокой концентрацией глюкозы. Очевидно, безжизненная, замороженная лягушка может возобновить дыхание, а ее сердцебиение возобновится, когда условия потеплеют. [ 99 ]
С другой стороны, полосатая роющая лягушка ( Cyclorana alboguttata ) регулярно проводит спячку в жаркий и засушливый сезон в Австралии, выживая в состоянии покоя без доступа к пище и воде в течение девяти или десяти месяцев в году. Он зарывается под землю и сворачивается внутри защитного кокона, образованного сброшенной кожей. Исследователи из Университета Квинсленда обнаружили, что во время зимнего периода метаболизм лягушки изменяется и эффективность работы митохондрий повышается . Это означает, что ограниченное количество энергии, доступное коматозной лягушке, используется более эффективно. Этот механизм выживания полезен только для животных, которые остаются в полном бессознательном состоянии в течение длительного периода времени и чьи энергетические потребности невелики, поскольку они хладнокровны и не нуждаются в выработке тепла. [ 100 ] Другие исследования показали, что для удовлетворения этих энергетических потребностей мышцы атрофируются, но мышцы задних конечностей преимущественно не затрагиваются. [ 101 ] Было обнаружено, что у лягушек верхняя критическая температура составляет около 41 градуса по Цельсию. [ 102 ]
Передвижение
Различные виды лягушек используют ряд методов передвижения, включая прыжки , бег , ходьбу , плавание , рытье норы , лазание и скольжение .

- Прыжки
Лягушки общепризнаны как исключительные прыгуны и, по сравнению с их размером, лучшие прыгуны среди всех позвоночных. [ 103 ] Полосатая ракетная лягушка Литория Насута умеет перепрыгивать через two metres (6+1⁄2 feet), a distance that is more than fifty times its body length of мм ( 2 + 1/4 дюйма 55 ). [ 104 ] Между видами существуют огромные различия в прыгучести. Внутри вида расстояние прыжка увеличивается с увеличением размера, но относительное расстояние прыжка (длина прыжка) уменьшается. Индийская лягушка-шкипер ( Euflyctis cyanophlyctis ) обладает способностью выпрыгивать из воды из положения, плавающего на поверхности. [ 105 ] Крошечная лягушка северного сверчка ( Acris crepitans ) может «скользить» по поверхности пруда серией коротких быстрых прыжков. [ 106 ]
Замедленная фотография показывает, что мышцы обладают пассивной гибкостью. Сначала они растягиваются, пока лягушка все еще находится в приседе, затем сжимаются, а затем снова растягиваются, чтобы подбросить лягушку в воздух. Передние ноги прижаты к груди, а задние остаются в вытянутом обтекаемом положении на протяжении всего прыжка. [ 52 ] У некоторых чрезвычайно способных прыгунов, таких как кубинская квакша ( Osteopilus septentrionalis ) и северная леопардовая лягушка ( Rana pipiens ), пиковая мощность, проявляемая во время прыжка, может превышать ту, которую теоретически способны создать мышцы. Когда мышцы сокращаются, энергия сначала передается в растянутое сухожилие, которое обертывает лодыжку. Затем мышцы снова растягиваются, в то время как сухожилие высвобождает свою энергию, как катапульта , создавая мощное ускорение, выходящее за пределы ускорения, приводимого в движение мышцами. [ 107 ] Подобный механизм был зарегистрирован у саранчи и кузнечиков . [ 108 ]
Раннее вылупление лягушек может отрицательно повлиять на прыгучесть и общее передвижение лягушек. [ 109 ] Задние конечности не могут полностью сформироваться, в результате чего они короче и намного слабее по сравнению с нормальной вылупившейся лягушкой. [ 109 ] Ранние вылупляющиеся лягушки могут чаще зависеть от других форм передвижения, таких как плавание и ходьба. [ 109 ]
- Ходьба и бег

Лягушки семейств Bufonidae, Rhinophrynidae и Microhylidae имеют короткие задние ноги и склонны скорее ходить, чем прыгать. [ 110 ] Когда они пытаются двигаться быстро, они ускоряют скорость движения конечностей или прибегают к неуклюжей подпрыгивающей походке. ( Узкоротая жаба Великих равнин Gastroryne olivacea ) была описана как имеющая походку, которая представляет собой «комбинацию бега и коротких прыжков, длина которых обычно составляет всего один-два дюйма». [ 111 ] В ходе эксперимента жабу Фаулера ( Bufo fowleri ) поместили на беговую дорожку, которая вращалась с различной скоростью. Измерив потребление кислорода жабой, было обнаружено, что прыжки являются неэффективным использованием ресурсов во время длительного передвижения, но являются полезной стратегией во время коротких всплесков высокоинтенсивной активности. [ 112 ]
Красноногая бегущая лягушка ( Kassina maculata ) имеет короткие, тонкие задние конечности, непригодные для прыжков. Он может двигаться быстро, используя беговую походку, при которой поочередно задействуются две задние ноги. Замедленная фотография показывает, что в отличие от лошади, которая может рысью или галопом, походка лягушки оставалась одинаковой на медленной, средней и высокой скорости. [ 113 ] Этот вид также может лазить по деревьям и кустарникам и делает это ночью, чтобы ловить насекомых. [ 114 ] Индийская лягушка-шкипер ( Euflyctis cyanophlyctis ) имеет широкие лапы и может бегать по поверхности воды на несколько метров (ярдов). [ 106 ]
- Плавание

Лягушки, живущие в воде или посещающие ее, обладают приспособлениями, улучшающими их плавательные способности. Задние конечности мускулистые и сильные. Перепонки между пальцами задних лап увеличивают площадь стопы и помогают лягушке мощно продвигаться по воде. Представители семейства Pipidae полностью водны и демонстрируют наиболее выраженную специализацию. У них негибкий позвоночник, уплощенное, обтекаемое тело, системы боковых линий и мощные задние конечности с большими перепончатыми ступнями. [ 115 ] Головастики обычно имеют большие хвостовые плавники, которые обеспечивают толчок при движении хвоста из стороны в сторону. [ 116 ]
- Закапывание
Некоторые лягушки приспособились к рытью нор и жизни под землей. У них, как правило, округлые тела, короткие конечности, маленькие головы с выпученными глазами и задние лапы, приспособленные для раскопок. Крайним примером этого является пурпурная лягушка ( Nasikabatrachus sahyadrensis ) из южной Индии, которая питается термитами и проводит почти всю свою жизнь под землей. Он ненадолго появляется во время сезона дождей, чтобы спариваться и размножаться во временных водоемах. У него крошечная голова с заостренной мордой и пухлое округлое тело. Из-за такого окаменелого существования он был впервые описан в 2003 году и был в то время новым для научного сообщества, хотя ранее был известен местному населению. [ 117 ]

Чесночники Северной Америки также приспособлены к подземной жизни. плюсневых костей задних лап Типична равнинная чесночница ( Spea Bombifrons ) и имеет лоскут ороговевшей кости, прикрепленный к одной из , который она использует, чтобы закапываться назад в землю. Роя, жаба покачивает бедрами из стороны в сторону, погружаясь в рыхлую почву. Летом у него есть неглубокая нора, из которой он выходит ночью в поисках корма. Зимой он роет намного глубже и был зарегистрирован на глубине 4,5 м (14 футов 9 дюймов). [ 118 ] Туннель заполнен землей, и жаба зимует в небольшой камере в конце. За это время в ее тканях накапливается мочевина, а вода всасывается из окружающей влажной почвы путем осмоса для удовлетворения потребностей жабы. [ 118 ] Чесночники - это «взрывные заводчики», все они одновременно выходят из своих нор и собираются во временных лужах, привлеченные к одному из них призывом первого самца, который нашел подходящее место для размножения. [ 119 ]
Роющие лягушки Австралии ведут совсем другой образ жизни. Западная пятнистая лягушка ( Heleioporus albopunctatus ) роет нору на берегу реки или в русле временного ручья и регулярно выходит на корм. Происходит спаривание, и яйца откладываются в пенопластовое гнездо внутри норы. Яйца частично развиваются там, но не вылупляются, пока не попадут в воду после сильного дождя. Затем головастики выплывают в открытую воду и быстро завершают свое развитие. [ 120 ] Мадагаскарские роющие лягушки менее роют и в основном зарываются в опавшие листья. Одна из них, зеленая роющая лягушка ( Scaphiophryne marmorata ), имеет уплощенную голову с короткой мордой и хорошо развитыми плюсневыми бугорками на задних лапах, которые помогают копать землю. У него также сильно увеличенные концевые диски на передних лапах, которые помогают ему карабкаться по кустам. [ 121 ] Размножается во временных лужах, образующихся после дождей. [ 122 ]
- Восхождение


Древесные лягушки живут высоко в кронах деревьев , где они карабкаются по ветвям, веткам и листьям, иногда никогда не спускаясь на землю. «Настоящие» древесные лягушки принадлежат к семейству Hylidae, но представители других семейств лягушек независимо переняли древесный образ жизни, что является примером конвергентной эволюции . К ним относятся стеклянные лягушки (Centrolenidae), кустарниковые лягушки (Hyperoliidae), некоторые узкоротые лягушки (Microhylidae) и кустарниковые лягушки (Rhacophoridae). [ 110 ] Большинство древесных лягушек имеют длину менее 10 см (4 дюйма), с длинными ногами и длинными пальцами с липкими подушечками на кончиках. Поверхность подушечек пальцев ног образована плотно упакованным слоем плоских шестиугольных эпидермальных клеток, разделенных бороздками, в которые железы выделяют слизь . Эти подушечки пальцев, смоченные слизью, обеспечивают сцепление с любой влажной или сухой поверхностью, включая стекло. При этом задействованы силы, включающие граничное трение эпидермиса подушечек пальцев ног о поверхность, а также поверхностное натяжение и вязкость . [ 123 ] Древесные лягушки очень акробатичны и могут ловить насекомых, вися на одном пальце ноги на ветке или цепляясь за лопасть продуваемого ветром тростника. [ 124 ] некоторых представителей подсемейства Phyllomedusinae У пальцы на ногах противоположны. Сетчатая листовая лягушка ( Phyllomedusa ayeaye ) имеет один противоположный палец на каждой передней лапе и два противоположных пальца на задних лапах. Это позволяет ему хвататься за стебли кустов, когда он карабкается по прибрежной среде обитания. [ 125 ]
- Скольжение
За всю историю эволюции лягушек несколько разных групп независимо поднимались в воздух. [ 126 ] Некоторые лягушки тропических лесов специально приспособлены к планированию с дерева на дерево или прыжкам с парашютом на лесную подстилку. Типичным из них является летающая лягушка Уоллеса ( Rhacophorus nigropalmatus ) из Малайзии и Борнео. У него большие ступни, кончики пальцев расширены в плоские липкие диски, а пальцы полностью перепончатые. Лоскуты кожи возникают на боковых краях конечностей и в области хвоста. С растопыренными пальцами, вытянутыми конечностями и расправленными крыльями он может планировать на значительные расстояния, но не может совершать полет с приводом. [ 127 ] Он может менять направление движения и преодолевать расстояния до 15 м (50 футов) между деревьями. [ 128 ]
История жизни

( кваканье лягушки )
Воспроизведение
У лягушек наблюдаются два основных типа размножения: длительное размножение и взрывное размножение. В первом случае, принятом большинством видов, взрослые лягушки в определенное время года собираются у пруда, озера или ручья для размножения. Многие лягушки возвращаются в водоемы, в которых они развивались в виде личинок. Это часто приводит к ежегодным миграциям, в которых участвуют тысячи особей. У взрывных производителей взрослые взрослые лягушки прибывают к местам размножения в ответ на определенные пусковые факторы, такие как осадки, происходящие в засушливой зоне. У этих лягушек спаривание и нерест происходят быстро, а скорость роста личинок высока, чтобы использовать эфемерные лужи до того, как они высохнут. [ 129 ]
У длительно размножающихся самцы обычно первыми прибывают к месту размножения и остаются там некоторое время, тогда как самки, как правило, прибывают позже и уходят вскоре после нереста. Это означает, что самцы численно превосходят самок у кромки воды и защищают территории, с которых изгоняют других самцов. Они сигнализируют о своем присутствии криками, часто чередуя свое кваканье с соседними лягушками. Более крупные и сильные самцы, как правило, имеют более глубокие крики и сохраняют территории более высокого качества. Самки выбирают себе пару, по крайней мере частично, на основе глубины голоса. [ 130 ] У некоторых видов есть самцы-спутники, которые не имеют территории и не звонят. Они могут перехватить приближающихся к зовущему самцу самок или занять освободившуюся территорию. Звонки — это энергозатратное занятие. Иногда две роли меняются местами, и призывающий самец отказывается от своей территории и становится спутником. [ 129 ]

У взрывных заводчиков первый самец, который находит подходящее место для размножения, например, временный водоем, громко кричит, и к водоему собираются другие лягушки обоих полов. Заводчики взрывчатых веществ склонны кричать в унисон, создавая хор, который можно услышать издалека. В эту категорию попадают чесночноногие жабы ( Scaphiopus spp. ) Северной Америки. Выбор партнера и ухаживание не так важны, как скорость воспроизводства. В некоторые годы подходящие условия могут не возникнуть, и лягушки могут не размножаться в течение двух и более лет. [ 129 ] Некоторые самки чесночниц из Нью-Мексико ( Spea multiplicata ) откладывают за раз только половину имеющихся яиц, возможно, сохраняя часть на случай, если позже возникнет лучшая репродуктивная возможность. [ 131 ]
В месте размножения самец садится на самку и крепко обхватывает ее тело. Обычно амплексус происходит в воде: самка выпускает икринки, а самец покрывает их спермой; оплодотворение наружное . У многих видов, таких как жаба Великих равнин ( Bufo cognatus ), самец удерживает яйца задними лапами, удерживая их на месте около трех минут. [ 129 ] Представители западноафриканского рода Nimbaphrynoides уникальны среди лягушек тем, что они живородящие ; Limnonectes larvaepartus , Eleutherodactylus jasperi и представители танзанийского рода Nectophrynoides — единственные лягушки, которые, как известно, являются яйцеживородящими . У этих видов оплодотворение внутреннее , и самки рождают полностью развитых молодых лягушек, за исключением L. larvaepartus , которые рождают головастиков. [ 132 ] [ 133 ] [ 134 ]
Жизненный цикл
Яйца / лягушачья икра

Лягушки могут откладывать яйца в виде комков, поверхностных пленок, нитей или по отдельности. Около половины видов откладывают икру в воду, остальные откладывают яйца в растительности, на земле или в раскопках. [ 135 ] [ 136 ] [ 137 ] Крошечный желтополосый карликовый элеут ( Eleutherodactyluslimbatus ) откладывает яйца поодиночке, закапывая их во влажную почву. [ 138 ] Дымчатая джунглевая лягушка ( Leptodactylus pentadactylus ) строит гнездо из пены в дупле. Яйца вылупляются, когда гнездо затоплено, или головастики могут завершить свое развитие в пене, если не происходит затопления. [ 139 ] Красноглазая древесная лягушка ( Agalychnis callidryas ) откладывает яйца на лист над лужей, а когда они вылупляются, личинки падают в воду внизу. [ 140 ]
У некоторых видов, таких как древесная лягушка ( Rana sylvatica ), симбиотические в студенистом материале присутствуют одноклеточные зеленые водоросли. Считается, что они могут принести пользу развивающимся личинкам, снабжая их дополнительным кислородом посредством фотосинтеза . [ 141 ] Также было обнаружено, что внутренняя часть шаровидных скоплений яиц лесной лягушки на 6 ° C (11 ° F) теплее, чем окружающая вода, и это ускоряет развитие личинок. [ 142 ] Личинки, развивающиеся в яйцах, могут улавливать вибрации, вызванные находящимися поблизости хищными осами или змеями, и вылупляются рано, чтобы их не съели. [ 143 ] В целом продолжительность стадии яйца зависит от вида и условий окружающей среды. Водные яйца обычно вылупляются в течение одной недели, когда капсула расщепляется под действием ферментов, выделяемых развивающимися личинками. [ 144 ]
Прямое развитие , при котором из яиц вылупляются молодые особи, подобные маленьким взрослым особям, известно также у многих лягушек, например, Ischnocnema henselii , [ 145 ] Элеутеродактиль коки , [ 146 ] и Raorchestes ochlandrae и Raorchestes chalazodes . [ 147 ]
Головастики

Личинки, выходящие из яиц, известны как головастики (или иногда головастики). Головастики лишены век и конечностей, имеют хрящевой скелет, жабры для дыхания (сначала наружные, позже внутренние) и хвост, который они используют для плавания. [ 116 ] Как правило, свободноживущие личинки полностью водны, но по крайней мере у одного вида ( Nannophrys ceylonensis ) есть полуназемные головастики, обитающие среди влажных камней. [ 148 ] [ 149 ]
С самого начала развития жаберный мешок покрывает жабры и передние ноги головастика. Легкие вскоре начинают развиваться и используются как вспомогательный орган дыхания. Некоторые виды проходят метаморфозу еще внутри яйца и вылупляются прямо в маленьких лягушек. У головастиков нет настоящих зубов, но у большинства видов челюсти имеют два удлиненных параллельных ряда небольших ороговевших структур, называемых керадонтами, в верхних челюстях. Их нижние челюсти обычно имеют три ряда керадонтов, окруженных роговым клювом, но количество рядов может варьироваться, а точное расположение частей рта позволяет идентифицировать вид. [ 144 ] У Pipidae, за исключением Hymenochirus , головастики имеют парные передние усики, что придает им сходство с небольшими сомами . [ 115 ] Их хвосты укреплены хордой , но не содержат каких-либо костных или хрящевых элементов, за исключением нескольких позвонков у основания, которые во время метаморфоза образуют уростиль. Это было предложено как адаптация к их образу жизни; поскольку превращение в лягушку происходит очень быстро, хвост состоит только из мягких тканей, поскольку кости и хрящи расщепляются и усваиваются гораздо дольше. Хвостовой плавник и кончик хрупкие и легко рвутся, что рассматривается как приспособление к спасению от хищников, которые пытаются схватить их за хвост. [ 150 ]
Головастики, как правило , травоядны и питаются в основном водорослями , в том числе диатомовыми водорослями, которые фильтруются из воды через жабры . Некоторые виды на стадии головастика плотоядны и поедают насекомых, более мелких головастиков и рыбу. Кубинская древесная лягушка ( Osteopilus septentrionalis ) — один из многих видов, у которых головастики могут быть каннибалистами . Головастики, у которых рано развиваются ноги, могут быть съедены другими, поэтому у поздних разработчиков могут быть лучшие долгосрочные перспективы выживания. [ 151 ]
Головастики очень уязвимы для поедания рыбами, тритонами , хищными жуками-плавунами и птицами, особенно водоплавающими , такими как аисты , цапли и домашние утки . Некоторые головастики, в том числе тростниковой жабы ( Rhinella marina ), ядовиты. Стадия головастика может длиться всего неделю у взрывных производителей или может длиться одну или несколько зим с последующим метаморфозом весной. [ 152 ]
Метаморфоза
В конце стадии головастика лягушка претерпевает метаморфозу, при которой ее тело внезапно переходит во взрослую форму. Эта метаморфоза обычно длится всего 24 часа и инициируется выработкой гормона тироксина . Это приводит к тому, что разные ткани развиваются по-разному. Основные изменения, которые происходят, включают развитие легких и исчезновение жабр и жаберного мешка, в результате чего передние ноги становятся видимыми. Нижняя челюсть трансформируется в большую челюсть хищной взрослой особи, а длинная спиральная кишка травоядного головастика заменяется типичной короткой кишкой хищника. [ 144 ] Гомеостатический контроль потребления пищи по обратной связи практически отсутствует, из-за чего головастики едят постоянно, когда пища присутствует. Но незадолго до и во время метаморфоза чувство голода подавляется, и они перестают есть, а их кишечник и внутренние органы реорганизуются и подготавливаются к другому питанию. [ 153 ] [ 154 ] Также меняется микробиота кишечника : от сходства с рыбой до амниот. [ 155 ] Исключением являются плотоядные головастики, такие как Lepidobatrachus laevis , кишечник которых уже адаптирован к диете, аналогичной диете взрослых особей. Они продолжают есть во время метаморфоза. [ 156 ] Нервная система адаптируется к слуху и стереоскопическому зрению, а также к новым способам передвижения и питания. [ 144 ] Глаза перемещаются выше на голове, формируются веки и связанные с ними железы. Развиты барабанная перепонка, среднее и внутреннее ухо. Кожа становится толще и жестче, система боковой линии утрачивается, развиваются кожные железы. [ 144 ] Завершающая стадия — исчезновение хвоста, но происходит это гораздо позже, поскольку ткань используется для резкого роста конечностей. [ 157 ] Лягушки наиболее уязвимы для хищников в период метаморфоза. В это время теряется хвост и только налаживается передвижение с помощью конечностей. [ 110 ]
-
 Личинка обыкновенной лягушки Rana temporaria за сутки до метаморфоза
Личинка обыкновенной лягушки Rana temporaria за сутки до метаморфоза -
 Стадия метаморфоза с деформирующими челюстями, большими глазами и остатками жаберного мешка.
Стадия метаморфоза с деформирующими челюстями, большими глазами и остатками жаберного мешка. -
 Молодая лягушка с коротким хвостом, метаморфоза почти завершена
Молодая лягушка с коротким хвостом, метаморфоза почти завершена



Взрослые
Взрослые лягушки могут жить в воде или рядом с ней, но немногие из них ведут полностью водный образ жизни. [ 158 ] Почти все виды лягушек во взрослом состоянии плотоядны и питаются беспозвоночными, включая насекомых , крабов , пауков , клещей , червей , улиток и слизней . Некоторые из более крупных могут поедать других лягушек, мелких млекопитающих и рептилий, а также рыбу . [ 159 ] [ 160 ] Некоторые виды также едят растительную пищу; Древесная лягушка Xenohyla truncata частично травоядна, ее рацион включает большую долю фруктов, цветочных структур и нектара. [ 161 ] [ 162 ] Leptodactylus mystaceus поедает растения. Было обнаружено, что [ 163 ] [ 164 ] а фоливория встречается у Euphlyctis hexadactylus , причем растения составляют 79,5% его рациона по объему. [ 165 ] Многие лягушки используют свой липкий язык для ловли добычи, а другие просто хватают ее ртом. [ 166 ] Взрослые лягушки сами подвергаются нападению многих хищников. Северную леопардовую лягушку ( Rana pipiens ) поедают цапли , ястребы , рыбы, крупные саламандры , змеи , еноты , скунсы , норки , лягушки-быки и другие животные. [ 167 ]
Лягушки являются основными хищниками и важной частью пищевой сети . Будучи хладнокровными , они эффективно используют потребляемую пищу, расходуя мало энергии на метаболические процессы, а остальная часть преобразуется в биомассу . Сами они поедаются вторичными хищниками и являются основными наземными потребителями беспозвоночных, большинство из которых питаются растениями. Сокращая количество травоядных, они играют роль в ускорении роста растений и, таким образом, являются частью тонко сбалансированной экосистемы. [ 168 ]
Мало что известно о продолжительности жизни лягушек и жаб в дикой природе, но некоторые из них могут жить много лет. Скелетохронология – это метод исследования костей для определения возраста. С помощью этого метода был изучен возраст горных желтоногих лягушек ( Rana muscosa ), при этом на фалангах пальцев ног наблюдаются сезонные линии, где рост зимой замедляется. У самых старых лягушек было десять полос, поэтому считалось, что их возраст составляет 14 лет, включая четырехлетнюю стадию головастика. [ 169 ] Зарегистрировано, что содержащиеся в неволе лягушки и жабы живут до 40 лет — возраста, достигнутого европейской обыкновенной жабой ( Bufo bufo ). тростниковая жаба ( Rhinella marina Известно, что ) выживает в неволе 24 года, а американская лягушка-бык ( Rana catesbeiana ) — 14 лет. [ 170 ] Лягушки из умеренного климата впадают в зимнюю спячку, и известно, что четыре вида способны выдерживать заморозки в это время, в том числе древесная лягушка ( Rana sylvatica ). [ 171 ]
Родительская забота


Хотя забота о потомстве у лягушек плохо изучена, примерно 20% видов земноводных могут тем или иным образом заботиться о своем потомстве. [ 172 ] Эволюция родительской заботы у лягушек обусловлена прежде всего размером водоема, в котором они размножаются. Те, которые размножаются в меньших водоемах, как правило, имеют более серьезное и сложное родительское поведение. [ 173 ] Поскольку в крупных водоемах широко распространено хищничество яиц и личинок, некоторые виды лягушек начали откладывать яйца на суше. Как только это произошло, высыхающая земная среда требует, чтобы один или оба родителя поддерживали влажность детенышей, чтобы обеспечить их выживание. [ 174 ] Последующая необходимость транспортировки вылупившихся головастиков в водоем потребовала еще более интенсивной родительской заботы. [ 173 ]
В небольших водоемах хищники в основном отсутствуют, и конкуренция между головастиками становится переменной, ограничивающей их выживание. Некоторые виды лягушек избегают этой конкуренции, используя более мелкие фитотелмы (заполненные водой пазухи листьев или небольшие древесные полости) в качестве мест для откладывания нескольких головастиков. [ 175 ] Хотя эти небольшие места выращивания свободны от конкуренции, им также не хватает питательных веществ, чтобы прокормить головастика без помощи родителей. Виды лягушек, которые перешли от использования более крупных фитотелм к более мелким, выработали стратегию обеспечения своего потомства питательными, но неоплодотворенными яйцами. [ 173 ] Самка земляничной ядовитой лягушки ( Oophaga pumilio ) откладывает яйца на лесную подстилку. Лягушка-самец охраняет их от хищников и носит в своей клоаке воду, чтобы они оставались влажными. Когда они вылупляются, самка перемещает головастиков на своей спине к бромелиям , содержащим воду , или к другому подобному водоему, откладывая по одному в каждом месте. Она регулярно их посещает и кормит, откладывая в фитотельму одно-два неоплодотворенных яйца, продолжая делать это до тех пор, пока молодь не станет достаточно большой для метаморфоза. [ 176 ] ( зернистая ядовитая лягушка Oophaga granulifera ). Аналогичным образом заботится о своих головастиках [ 177 ]
У лягушек наблюдается множество других разнообразных форм родительской заботы. Крошечный самец Colostethus subpunctatus стоит на страже своей скопления яиц, отложенной под камнем или бревном. Когда икра вылупляется, он переносит головастиков на своей спине во временный водоем, где частично погружается в воду и один или несколько головастиков выпадают. Затем он переходит в другой бассейн. [ 178 ] Самец обыкновенной жабы-акушерки ( Alytes obstetricans ) носит икру, прикрепляясь к задним лапам. Он сохраняет их влажными в сухую погоду, погружаясь в пруд, и не дает им слишком намокнуть в сырой растительности, поднимая задние конечности. Через три-шесть недель он отправляется к пруду, и из яиц вылупляются головастики. [ 179 ] Тунгара лягушка ( Physalaemus pustulosus ) строит плавучее гнездо из пены, чтобы защитить свои яйца от хищников. Пена состоит из белков и лектинов и, по-видимому, обладает противомикробными свойствами. [ 180 ] Несколько пар лягушек могут образовать колониальное гнездо на ранее построенном плоту. Яйца откладываются в центр, затем следуют чередующиеся слои пены и яиц, завершающиеся пенопластовой крышкой. [ 181 ]
Некоторые лягушки защищают свое потомство внутри собственного тела. И самцы, и самки сумчатых лягушек ( Assa darlingtoni ) охраняют свои яйца, откладывающиеся на землю. Когда икра вылупляется, самец смазывает свое тело окружающим их желе и погружается в яичную массу. Головастики извиваются в кожные мешочки на его боку, где развиваются, пока не превращаются в молодых лягушек. [ 182 ] Самка желудочно-высиживающей лягушки ( Reobatrachus sp.) из Австралии , ныне, вероятно, вымершая, заглатывает оплодотворенные яйца, которые затем развиваются внутри ее желудка. Она перестает питаться и перестает выделять желудочную кислоту . Головастики питаются желтками яиц. Через шесть или семь недель они готовы к метаморфозе. Мать срыгивает крошечных лягушек, которые выпрыгивают изо рта. [ 183 ] Самка лягушки Дарвина ( Rinoderma darwinii ) из Чили откладывает на земле до 40 яиц, где их охраняет самец. Когда головастики вот-вот вылупятся, их поглощает самец, который носит их внутри своего значительно увеличенного голосового мешка. Здесь они погружаются в пенистую вязкую жидкость, содержащую питательные вещества, дополняющие то, что они получают из яичных желтков. Они остаются в мешке от семи до десяти недель, прежде чем подвергаются метаморфозу, после чего перемещаются в рот самца и выходят наружу. [ 184 ]
Оборона


На первый взгляд лягушки кажутся довольно беззащитными из-за их небольшого размера, медленного движения, тонкой кожи и отсутствия защитных структур, таких как шипы, когти или зубы. Многие используют камуфляж, чтобы избежать обнаружения: кожа часто имеет пятна или полосы нейтральных цветов, которые позволяют неподвижной лягушке сливаться с окружающей средой. Некоторые могут совершать потрясающие прыжки, часто в воду, что помогает им уклоняться от потенциальных нападающих, в то время как у многих есть другие защитные приспособления и стратегии. [ 129 ]
Кожа многих лягушек содержит мягкие токсичные вещества, называемые буфотоксинами, которые делают их неприятными для потенциальных хищников. У большинства жаб и некоторых лягушек есть большие ядовитые железы, околоушные железы , расположенные по бокам головы за глазами, и другие железы в других частях тела. Эти железы выделяют слизь и ряд токсинов, которые делают лягушек скользкими, неприятными или ядовитыми. Если вредный эффект наступит немедленно, хищник может прекратить свои действия, а лягушка убежит. Если эффект будет развиваться медленнее, хищник может научиться избегать этого вида в будущем. [ 185 ] Ядовитые лягушки склонны демонстрировать свою токсичность яркими цветами — такая адаптивная стратегия известна как апосематизм . Это делают ядовитые лягушки-древоласы семейства Dendrobatidae. Обычно они красного, оранжевого или желтого цвета, часто с контрастными черными отметинами на теле. Allobates zaparo не ядовит, но имитирует внешний вид двух разных токсичных видов, с которыми имеет общий ареал, пытаясь обмануть хищников. [ 186 ] Другие виды, такие как европейская огнебрюхая жаба ( Bombina Bombina ), имеют снизу предупреждающий цвет. Они «вспыхивают» этим, когда на них нападают, принимая позу, обнажающую яркую окраску их животов. [ 4 ]

Некоторые лягушки, например ядовитые древолазы , особенно токсичны. Коренные народы Южной Америки добывают из этих лягушек яд, чтобы применять его к своему оружию . охотничьему [ 187 ] хотя немногие виды достаточно токсичны, чтобы их можно было использовать для этой цели. По крайней мере, два неядовитых вида лягушек тропической Америки ( Eleutherodactylus gaigei и Lithodytes lineatus ) имитируют окраску ядовитых лягушек-дротиков в целях самозащиты. [ 188 ] [ 189 ] Некоторые лягушки получают яд от муравьев и других членистоногих, которых они едят. [ 190 ] Другие, такие как австралийские лягушки-корробори ( Pseudophryne corroboree и Pseudophryne pengilleyi ), могут сами синтезировать алкалоиды . [ 191 ] Используемые химические вещества могут быть раздражителями, галлюциногенами , конвульсантами , нервно-паралитическим ядом или сосудосуживающими средствами . Многие хищники лягушек приспособились переносить высокие уровни этих ядов, но другие существа, в том числе люди, которые обращаются с лягушками, могут серьезно пострадать. [ 192 ]
Некоторые лягушки используют блеф или обман. Европейская обыкновенная жаба ( Bufo bufo ) при нападении принимает характерную стойку: раздувает тело и стоит с поднятой задней частью и опущенной головой. [ 193 ] Лягушка-бык ( Rana catesbeiana ) при угрозе приседает с закрытыми глазами и наклоняет голову вперед. Это помещает околоушные железы в наиболее эффективное положение, другие железы на спине начинают выделять ядовитые выделения, а наиболее уязвимые части тела оказываются защищенными. [ 129 ] Другая тактика, используемая некоторыми лягушками, - это «кричать», внезапный громкий шум, который пугает хищника. Серая квакша ( Hyla versicolor ) издает взрывной звук, который иногда отпугивает землеройку Blarina brevicauda . [ 129 ] Хотя многие хищники избегают жаб, обыкновенная подвязочная змея ( Thamnophis sirtalis ими регулярно питается ). Стратегия, используемая молодыми американскими жабами ( Bufo americanus ) при приближении змеи, заключается в том, чтобы присесть и оставаться неподвижными. Обычно это удается: змея проходит мимо, а жаба остается незамеченной. Однако, если она сталкивается с головой змеи, жаба отпрыгивает, прежде чем присесть и защититься. [ 194 ]
Распределение

Лягушки живут на всех континентах, кроме Антарктиды, но их нет на некоторых островах, особенно на тех, которые находятся вдали от континентальной суши. [ 195 ] [ 196 ] Многие виды изолированы в ограниченных ареалах из-за изменений климата или негостеприимных территорий, таких как морские участки, горные хребты, пустыни, вырубка лесов, строительство дорог или другие искусственные барьеры. [ 197 ] Обычно большее разнообразие лягушек встречается в тропических регионах, чем в регионах с умеренным климатом, таких как Европа. [ 198 ] Некоторые лягушки обитают в засушливых районах, таких как пустыни, и для выживания полагаются на особые приспособления. Представители австралийского рода Cyclorana зарываются под землю, где создают водонепроницаемый кокон, в котором они могут отдыхать в засушливые периоды. Когда идет дождь, они выходят на поверхность, находят временный водоем и размножаются. Развитие яиц и головастиков происходит очень быстро по сравнению с таковыми у большинства других лягушек, поэтому размножение можно завершить до того, как пруд высохнет. [ 199 ] Некоторые виды лягушек приспособлены к холодной среде. Лесная лягушка ( Rana sylvatica ), ареал которой простирается за Полярный круг , на зиму зарывается в землю. Хотя большая часть его тела в это время замерзает, в его жизненно важных органах сохраняется высокая концентрация глюкозы, которая защищает их от повреждений. [ 53 ]
Сохранение

В 2006 году из 4035 видов амфибий, которые на той или иной стадии жизненного цикла зависят от воды, 1356 (33,6%) считались находящимися под угрозой исчезновения. Вероятно, это заниженная оценка, поскольку она исключает 1427 видов, данных по которым было недостаточно для оценки их статуса. [ 200 ] Популяция лягушек резко сократилась с 1950-х годов. Считается, что более трети видов лягушек находятся под угрозой исчезновения , а с 1980-х годов считается, что более 120 видов вымерли. [ 201 ] Среди этих видов — желудочные лягушки Австралии и золотая жаба Коста-Рики. Последний вызывает особую озабоченность у ученых, поскольку он обитал в нетронутом заповеднике облачных лесов Монтеверде , и его популяция резко сократилась в 1987 году вместе с примерно 20 другими видами лягушек в этом районе. Это не могло быть напрямую связано с деятельностью человека, такой как вырубка лесов, и выходило за пределы нормальных колебаний численности населения. [ 202 ] В других странах утрата среды обитания является серьезной причиной сокращения популяции лягушек, равно как и загрязняющие вещества, изменение климата, увеличение УФ-В -излучения, а также появление чужеродных хищников и конкурентов. [ 203 ] Канадское исследование, проведенное в 2006 году, показало, что интенсивное движение транспорта в их среде обитания представляет собой большую угрозу для популяций лягушек, чем потеря среды обитания. [ 204 ] Новые инфекционные заболевания, в том числе хитридиомикоз и ранавирус , также наносят разрушительный ущерб населению. [ 205 ] [ 206 ]
Многие ученые-экологи полагают, что земноводные, в том числе лягушки, являются хорошими биологическими индикаторами в целом здоровья экосистемы из-за их промежуточного положения в пищевых цепях, проницаемой кожи и, как правило, двухфазного образа жизни (водные личинки и наземные взрослые особи). [ 207 ] Похоже, что виды, имеющие как водные яйца, так и личинки, больше всего страдают от сокращения численности, тогда как виды с прямым развитием являются наиболее устойчивыми. [ 208 ]

количество мутаций и генетических дефектов лягушек С 1990-х годов увеличилось . К ним часто относятся отсутствие ног или дополнительные ноги. Были идентифицированы или выдвинуты различные предположения, включая увеличение ультрафиолетового излучения , влияющего на икру на поверхности прудов, химическое загрязнение пестицидами и удобрениями, а также паразитов, таких как трематода Ribeiroia ondatrae . Вероятно, все они задействованы сложным образом в качестве стрессоров , факторов окружающей среды, способствующих распространению заболеваний, а также уязвимости к атакам паразитов. Пороки развития ухудшают подвижность, и люди могут не дожить до взрослого возраста. Увеличение количества поедаемых птицами лягушек может фактически увеличить вероятность паразитизма на других лягушках, поскольку сложный жизненный цикл трематоды включает улитку-баранорога и нескольких промежуточных хозяев, таких как птицы. [ 209 ] [ 210 ]
В некоторых случаях были созданы программы разведения в неволе, которые в основном оказались успешными. [ 211 ] [ 212 ] Всемирная ассоциация зоопарков и аквариумов назвала 2008 год «Годом лягушки», чтобы привлечь внимание к проблемам охраны природы, с которыми они столкнулись. [ 213 ]
Тростниковая жаба ( Rhinella marina ) — очень адаптируемый вид, обитающий в Южной и Центральной Америке. В 1930-х годах он был завезен в Пуэрто-Рико, а затем и на другие острова Тихоокеанского и Карибского региона в качестве биологического средства борьбы с вредителями . [ 214 ] В 1935 году 3000 жаб были освобождены на полях сахарного тростника в Квинсленде, Австралия, в попытке контролировать тростниковых жуков, таких как Dermolepida albohirtum , личинки которых повреждают и убивают тростник. Первоначальные результаты во многих из этих стран были положительными, но позже стало очевидно, что жабы нарушают экологический баланс в новой среде обитания. Они свободно размножались, конкурировали с местными видами лягушек, ели пчел и других безвредных местных беспозвоночных, имели мало хищников в принятых ими местах обитания и травили домашних животных, хищных птиц и млекопитающих. Во многих из этих стран их сейчас считают одновременно вредителями и инвазивными видами , и ученые ищут биологические методы борьбы с ними. [ 215 ]
Человеческое использование
Кулинарный

Лягушачьи лапки едят люди во многих частях мира. Индонезия является крупнейшим в мире экспортером лягушачьего мяса, ежегодно экспортируя более 5000 тонн лягушачьего мяса, в основном во Францию, Бельгию и Люксембург. [ 216 ] Первоначально их поставляли из местных диких популяций, но чрезмерная эксплуатация привела к сокращению поставок. Это привело к развитию лягушачьего хозяйства и мировой торговли лягушками. Основными странами-импортерами являются Франция, Бельгия, Люксембург и США, а основными странами-экспортерами являются Индонезия и Китай. [ 217 ] Ежегодный мировой объем торговли американской лягушкой-быком ( Rana catesbeiana ), выращиваемой в основном в Китае, колеблется от 1200 до 2400 тонн. [ 218 ]
Горная куриная лягушка , названная так по вкусу курицы, в настоящее время находится под угрозой исчезновения, отчасти из-за употребления в пищу человеком, и была основным продуктом питания доминиканцев . [ 219 ] Енот , опоссум , куропатки , прерийные куры и лягушки были среди блюд, которые Марк Твен записал как часть американской кухни. [ 220 ]
Научные исследования
В ноябре 1970 года НАСА отправило в космос двух лягушек-быков на шесть дней во время миссии Orbiting Frog Otolith для проверки невесомости.
Лягушек используют для препарирования на уроках анатомии в средних школах и университетах, часто сначала им вводят цветные вещества, чтобы усилить контраст между биологическими системами . Эта практика приходит в упадок из-за проблем с благополучием животных , и теперь «цифровые лягушки» доступны для виртуального вскрытия. [ 221 ]
лягушки служили экспериментальными животными На протяжении всей истории науки . Биолог восемнадцатого века Луиджи Гальвани обнаружил связь между электричеством и нервной системой , изучая лягушек. Он создал один из первых инструментов для измерения электрического тока из лягушачьей лапки. [ 222 ] В 1852 году Х. Ф. Станниус использовал сердце лягушки в процедуре, называемой лигатурой Станниуса , чтобы продемонстрировать, что желудочек и предсердия сокращаются независимо друг от друга и с разной частотой. [ 223 ] Африканская шпорцевая лягушка или платана ( Xenopus laevis ) впервые широко использовалась в лабораториях для тестов на беременность в первой половине 20 века. Образец мочи беременной женщины, введенный самке лягушки, побуждает ее откладывать яйца - открытие, сделанное английским зоологом Ланселотом Хогбеном . Это связано с тем, что гормон хорионический гонадотропин человека присутствует в значительных количествах в моче женщин во время беременности. [ 224 ] В 1952 году Роберт Бриггс и Томас Дж. Кинг клонировали лягушку путем переноса ядра соматической клетки . Эта же техника позже была использована для создания овцы Долли , и их эксперимент стал первым случаем успешной трансплантации ядра высшим животным. [ 225 ]
Лягушки используются в исследованиях клонирования и других отраслях эмбриологии . Хотя были разработаны альтернативные тесты на беременность, биологи продолжают использовать Xenopus в качестве модельного организма в биологии развития, поскольку их эмбрионы большие, ими легко манипулировать, их легко получить и легко хранить в лаборатории. [ 226 ] Xenopus laevis все чаще вытесняется своим меньшим родственником Xenopus тропическим , который достигает репродуктивного возраста через пять месяцев, а не через один-два года, как у X. laevis . [ 227 ] тем самым способствуя более быстрому обучению между поколениями.
Геномы Xenopus laevis , X.tropicis , Rana catesbeiana , Rhinella marina и Nanorana parkeri были секвенированы и внесены в базу данных геномов NCBI . [ 228 ]
Фармацевтический

Поскольку токсины лягушек чрезвычайно разнообразны, они вызвали интерес биохимиков как «природная аптека». Алкалоид эпибатидин , болеутоляющее средство, в 200 раз более сильное, чем морфин , вырабатывается некоторыми видами ядовитых лягушек-древолагов . Другие химические вещества, выделенные из кожи лягушек, могут обеспечить устойчивость к ВИЧ- инфекции. [ 229 ] Яды дротиков активно исследуются на предмет их потенциала в качестве терапевтических препаратов. [ 230 ]
Давно подозревали, что доколумбовые мезоамериканцы использовали токсичный секрет, вырабатываемый тростниковой жабой, в качестве галлюциногена , но более вероятно, что они использовали вещества, выделяемые колорадской речной жабой ( Bufo alvarius ). Они содержат буфотенин (5-MeO-DMT), психоактивное соединение , которое в наше время используется в качестве рекреационного наркотика . Обычно кожные выделения сушат, а затем коптят. [ 231 ] В средствах массовой информации сообщалось о незаконном употреблении наркотиков путем облизывания кожи жабы, но это может быть городским мифом . [ 232 ]
Выделения из кожи золотой ядовитой лягушки ( Phyllobates terribilis ) традиционно используются коренными колумбийцами для отравления дротиков, которые они используют для охоты. Кончик снаряда трётся о спину лягушки и дротик запускается из духового пистолета . Комбинация двух алкалоидных токсинов, батрахотоксина и гомобатрахотоксина , настолько мощна, что одна лягушка содержит достаточно яда, чтобы убить примерно 22 000 мышей. [ 233 ] Два других вида, ядовитый древолаз Кокое ( Phyllobates aurotaenia ) и черноногий древолаз ( Phyllobates bicolor ), также используются для этой цели. Они менее токсичны и менее распространены, чем золотая ядовитая лягушка. Их нанизывают на заостренные палки и можно нагревать на огне, чтобы максимально увеличить количество яда, которое можно передать в дротик. [ 233 ]

Культурное значение
Лягушки фигурируют в мифологии, сказках и популярной культуре. В традиционных китайских мифах мир держится на гигантской лягушке, которая попытается проглотить Луну, вызвав лунное затмение . Лягушки фигурируют в религии, фольклоре и популярной культуре. Древние египтяне изображали бога Хекета , покровителя новорожденных, с головой лягушки. Для майя лягушки олицетворяли воду, урожай, плодородие и рождение и ассоциировались с богом Чааком . В Библии на египтян Моисей насылает нашествие лягушек . Средневековые европейцы ассоциировали лягушек и жаб со злом и колдовством . [ 234 ] братьев Гримм В сказке «Принц-лягушка» рассказывается, как принцесса принимает лягушку, и она превращается в прекрасного принца. [ 235 ] В современной культуре лягушки могут играть комедийные или незадачливые роли, как, например, мистер Жаба из романа 1908 года «Ветер в ивах» , Мичиган Дж. Фрог из мультфильмов Warner Bros. , кукла-лягушонок Кермит и в игре Frogger . [ 236 ]
Ссылки
- ^ Перейти обратно: а б Фрост, Даррел Р. (2021) [1998]. «Анура» . Виды амфибий мира . Американский музей естественной истории . Проверено 29 ноября 2022 г.
- ^ Перейти обратно: а б Каннателла, Дэвид К. (1997). «Салиентия» . Веб-проект «Древо жизни» . Проверено 7 августа 2012 г.
- ^ Бэджер, Д.; Нетертон, Дж. (1995). Лягушки . Издательство Эйрлайф. п. 19. ISBN 978-1-85310-740-5 .
- ^ Перейти обратно: а б Кузьмин Сергий Львович (29 сентября 1999 г.). « Бомбина бомбина » . АмфибияВеб . Калифорнийский университет в Беркли . Проверено 15 июня 2012 г.
- ^ Липс, К; Солис, Ф.; Ибаньес, Р.; Харамильо, К.; Фуэнмайор, К. (2010). « Ателопус зетеки » . Красный список исчезающих видов МСОП . 2010 . Проверено 2 августа 2012 г.
- ^ Лидделл, Генри Джордж; Скотт, Роберт (1940). «ἀ» . Греко-английский лексикон . Цифровая библиотека Персея.
- ^ Лидделл, Генри Джордж; Скотт, Роберт (1940). "οὐρά" . Греко-английский лексикон . Цифровая библиотека Персея.
- ^ Байи, Анатоль (1981). Сокращено из Французско-греческого словаря . Париж: Хашетт. ISBN 978-2010035289 . OCLC 461974285 .
- ^ Байи, Анатоль. «Греко-французский словарь онлайн» . www.tabularium.be . Проверено 9 декабря 2018 г.
- ^ «ануран, существительное и прил.» . Оксфордский словарь английского языка (онлайн-изд.). Издательство Оксфордского университета . (Требуется подписка или членство участвующей организации .)
- ^ Перейти обратно: а б с д «лягушка, № 1 и прил.» . Оксфордский словарь английского языка (онлайн-изд.). Издательство Оксфордского университета . (Требуется подписка или членство участвующей организации .)
- ^ "фрош | фроск, н.1." . Оксфордский словарь английского языка (онлайн-изд.). Издательство Оксфордского университета . (Требуется подписка или членство участвующей организации .)
- ^ Ежи Велна, « Выбор метатетических и неметатетических форм в среднеанглийском языке », Studia Anglica Posnaniensia , 30 (2002), 501–18 (стр. 504).
- ^ "жаба, н." . Оксфордский словарь английского языка (онлайн-изд.). Издательство Оксфордского университета . (Требуется подписка или членство участвующей организации .)
- ^ "головастик, №1." . Оксфордский словарь английского языка (онлайн-изд.). Издательство Оксфордского университета . (Требуется подписка или членство участвующей организации .)
- ^ Перейти обратно: а б Пау, Ф.Х.; Эндрюс, РМ; Кэдл, Дж. Э.; Крамп, МЛ; Савицкий, А.Х.; Уэллс, К.Д. (2003). Герпетология (третье изд.). Бенджамин Каммингс. ISBN 978-0-13-100849-6 .
- ^ Фрост, Даррел Р. (2023). «Анура Фишер фон Вальдхайм, 1813 год» . Виды амфибий мира: онлайн-справочник. Версия 6.2 . Американский музей естественной истории . Проверено 19 сентября 2023 г.
- ^ Дуэллман, Уильям Э. Биология амфибий , стр. 319, в Google Книгах.
- ^ Перейти обратно: а б Каннателла, Дэвид (11 января 2008 г.). «Анура» . Веб-проект «Древо жизни» . Проверено 8 августа 2012 г.
- ^ Форд, Лос-Анджелес; Каннателла, округ Колумбия (1993). «Основные клады лягушек». Герпетологические монографии . 7 : 94–117. дои : 10.2307/1466954 . JSTOR 1466954 .
- ^ Фаивович Дж.; Хаддад, ЦФБ; Гарсия, PCA; Фрост, ДР; Кэмпбелл, Дж.А.; Уиллер, WC (2005). «Систематический обзор семейства лягушек Hylidae с особым упором на Hylinae: филогенетический анализ и пересмотр». Бюллетень Американского музея естественной истории . 294 : 1–240. CiteSeerX 10.1.1.470.2967 . doi : 10.1206/0003-0090(2005)294[0001:SROTFF]2.0.CO;2 . S2CID 83925199 .
- ^ Кузьмин С.Л. (10 ноября 1999 г.). « Пелофилакс эскулентус » . Проверено 12 октября 2012 г.
- ^ Кёлер, Соня (2003). «Механизмы частичной репродуктивной изоляции в гибридной зоне Бомбина в Румынии» (PDF) . Диссертация на диссертацию . Архивировано (PDF) оригинала 9 октября 2022 г. Проверено 5 июня 2012 г.
- ^ Сан-Мауро, Диего; Винс, Майкл; Альковендас, ВМС; Зардоя, Рафаэль; Мейер, Аксель (2005). «Первоначальное разнообразие живых амфибий предшествовало распаду Пангеи» . Американский натуралист . 165 (5): 590–599. дои : 10.1086/429523 . JSTOR 429523 . ПМИД 15795855 . S2CID 17021360 . (требуется подписка)
- ^ Чжан, Пэн; Чжоу, Хуэй; Чен, Юэ-Цинь; Лю, И-Фэй; Цюй, Лян-Ху (2005). «Митогеномные взгляды на происхождение и филогению современных амфибий» . Систематическая биология . 54 (3): 391–400. дои : 10.1080/10635150590945278 . ПМИД 16012106 .
- ^ Марьянович, Давид; Лорен, Мишель (2007). «Ископаемые, молекулы, время расхождения и происхождение лисамфибий» . Систематическая биология . 56 (3): 369–388. дои : 10.1080/10635150701397635 . ПМИД 17520502 .
- ^ Пайрон, Р. Александр (2011). «Оценка времени дивергенции с использованием окаменелостей в качестве конечных таксонов и происхождение лиссамфибий» . Систематическая биология . 60 (4): 466–481. дои : 10.1093/sysbio/syr047 . ПМИД 21540408 .
- ^ Кассельман, Энн (21 мая 2008 г.). « Ископаемое «лягушка-амандра» может быть недостающим звеном амфибии» . Национальные географические новости . Архивировано из оригинала 25 мая 2008 года . Проверено 5 июля 2012 г.
- ^ Андерсон, Джейсон С.; Рейс, Роберт Р.; Скотт, Дайан; Фрёбиш, Надя Б.; Сумида, Стюарт С. (2008). «Стеблевой батрахий из ранней перми Техаса и происхождение лягушек и саламандр». Природа . 453 (7194): 515–518. Бибкод : 2008Natur.453..515A . дои : 10.1038/nature06865 . ПМИД 18497824 . S2CID 205212809 .
- ^ Марьянович, Д.; Лаурин, М. (2009). «Происхождение современных земноводных: комментарий» (PDF) . Эволюционная биология . 36 (3): 336–338. Бибкод : 2009EvBio..36..336M . дои : 10.1007/s11692-009-9065-8 . S2CID 12023942 .
- ^ Перейти обратно: а б Каннателла, Дэвид (1995). « Триадобатракус массиноти » . Древо Жизни . Проверено 26 июня 2008 г.
- ^ Мишель Р. Стокер; Стерлинг Дж. Несбитт; Бен Т. Клигман; Дэниел Дж. Палух; Адам Д. Марш; Дэвид С. Блэкберн; Уильям Дж. Паркер (2019). «Самая ранняя экваториальная запись лягушек из позднего триаса Аризоны» . Письма по биологии . 15 (2): Идентификатор статьи 20180922. doi : 10.1098/rsbl.2018.0922 . hdl : 10919/87931 . ПМК 6405462 . ПМИД 30958136 .
- ^ Рочек, З. (2000). «14. Мезозойские амфибии» (PDF) . В Хитволе, Х.; Кэрролл, Р.Л. (ред.). Биология амфибий: Палеонтология: эволюционная история земноводных . Том. 4. Суррей Битти и сыновья. стр. 1295–1331. ISBN 978-0-949324-87-0 . Архивировано (PDF) из оригинала 9 октября 2022 г.
- ^ Вейшампель, Д.Б.; Додсон, П.; Осмольска, Х., ред. (2004). Распространение динозавров (ранняя юра, Северная Америка): Динозаврия (2-е изд.). Издательство Калифорнийского университета. стр. 530–532. ISBN 978-0-520-24209-8 .
- ^ Шубин, Нью-Хэмпшир; Дженкинс, Ф.А. младший (1995). «Раннеюрская прыгающая лягушка». Природа . 377 (6544): 49–52. Бибкод : 1995Natur.377...49S . дои : 10.1038/377049a0 . S2CID 4308225 .
- ^ Фостер, Дж. (2007). «Анура (Лягушки)». Юрский запад: динозавры формации Моррисон и их мир . Издательство Университета Индианы. стр. 135–136. ISBN 978-0-253-34870-8 .
- ^ Уилсон, AC; Сарич, В.М.; Мэксон, Л.Р. (1974). «Важность перестройки генов в эволюции: данные исследований скорости хромосомной, белковой и анатомической эволюции» . Труды Национальной академии наук . 71 (8): 3028–3030. Бибкод : 1974PNAS...71.3028W . дои : 10.1073/pnas.71.8.3028 . ПМЦ 388613 . ПМИД 4528784 .
- ^ «Эволюция лягушки связана с ударом динозавра по астероиду» . Новости Би-би-си . 3 июля 2017 г. Проверено 3 июля 2017 г.
- ^ Фэн, Янь-Цзе; Блэкберн, Дэвид К.; Лян, Дэн; Хиллис, Дэвид М.; Уэйк, Дэвид Б.; Каннателла, Дэвид К.; Чжан, Пэн (2017). «Филогеномика обнаруживает быструю и одновременную диверсификацию трех основных клад гондванских лягушек на границе мела и палеогена» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (29): E5864–E5870. Бибкод : 2017PNAS..114E5864F . дои : 10.1073/pnas.1704632114 . ПМК 5530686 . ПМИД 28673970 .
- ^ Эванс, SE; Джонс, MEH; Краузе, Д.В. (2008). «Гигантская лягушка южноамериканского происхождения из позднего мела Мадагаскара» . Труды Национальной академии наук . 105 (8): 2951–2956. Бибкод : 2008PNAS..105.2951E . дои : 10.1073/pnas.0707599105 . ПМК 2268566 . ПМИД 18287076 .
- ^ Мёрс, Томас; Регеро, Марсело; Василян, Давид (2020). «Первая ископаемая лягушка из Антарктиды: последствия для климатических условий эоцена в высоких широтах и гондванского космополитизма Австралобатрахии» . Научные отчеты . 10 (1): 5051. Бибкод : 2020НатСР..10.5051М . дои : 10.1038/s41598-020-61973-5 . ПМК 7181706 . ПМИД 32327670 .
- ^ Джоэл, Лукас (23 апреля 2020 г.). «Окаменелости показывают, что хладнокровные лягушки жили в теплой Антарктиде» . Нью-Йорк Таймс . ISSN 0362-4331 . Архивировано из оригинала 23 апреля 2020 года . Проверено 13 мая 2020 г.
- ^ Фрост, ДР; Грант, Т.; Фаивович Ю.Н.; Бэйн, Р.Х.; Хаас, А.; Хаддад, CLFB; Де Са, Род-Айленд; Ченнинг, А.; Уилкинсон, М.; Доннеллан, Южная Каролина; Раксуорси, CJ; Кэмпбелл, Дж.А.; Блотто, БЛ; Молер, П.; Древес, Колорадо; Нуссбаум, РА; Линч, доктор медицинских наук; Грин, DM; Уиллер, WC (2006). «Древо жизни земноводных» . Бюллетень Американского музея естественной истории . 297 : 1–291. doi : 10.1206/0003-0090(2006)297[0001:TATOL]2.0.CO;2 . hdl : 2246/5781 . S2CID 86140137 .
- ^ Хейнике член парламента; Дуэллман, МЫ; Труб, Л.; Значит, БД; МакКаллох, РД; Хеджес, С.Б. (2009). «Новое семейство лягушек (Anura: Terrarana) из Южной Америки и расширенная клада прямого развития, выявленная с помощью молекулярной филогении» (PDF) . Зоотакса . 2211 : 1–35. дои : 10.11646/zootaxa.2211.1.1 .
- ^ Р. Александр Пайрон; Джон Дж. Винс (2011). «Крупномасштабная филогения земноводных, включающая более 2800 видов, и пересмотренная классификация современных лягушек, саламандр и червяг» . Молекулярная филогенетика и эволюция . 61 (2): 543–583. Бибкод : 2011МОЛПЭ..61..543А . дои : 10.1016/j.ympev.2011.06.012 . ПМИД 21723399 .
- ^ Перейти обратно: а б Флам, Ф. (1995). «Обнаружение самой ранней настоящей лягушки поможет палеонтологам понять, как лягушка развила свою прыгучесть» . Найт Риддер / Служба новостей Tribune. Архивировано из оригинала 13 мая 2013 года . Проверено 10 июня 2012 г.
- ^ «Крошечная лягушка признана самым маленьким позвоночным в мире» . Хранитель . 12 января 2012 года . Проверено 28 сентября 2012 г.
- ^ Колган, П.В. (1982). Книга рекордов Гиннеса по фактам и подвигам животных (3-е изд.). Превосходная степень Гиннесса. стр. 118–119. ISBN 978-0851122359 .
- ^ Отеро, РА; П. Хименес-Уидобро; С. Сото-Акунья; РЭЮри-Яньес (2014). «Свидетельства существования гигантской лягушки в шлеме (Australobatrachia, Calyptocephalellidae) с эоценовых уровней бассейна Магальянес, на самом юге Чили». Журнал южноамериканских наук о Земле . 55 : 133–140. Бибкод : 2014JSAES..55..133O . дои : 10.1016/j.jsames.2014.06.010 .
- ^ Морфологические вариации конечностей бесхвостых животных: ограничения и новинки
- ^ Морфологическая и экологическая конвергенция на нижнем пределе размера позвоночных, подчеркнутая пятью новыми миниатюрными видами микрогилидных лягушек из трех разных мадагаскарских родов.
- ^ Перейти обратно: а б Минотт, Кевин (15 мая 2010 г.). «Как прыгают лягушки» . Нэшнл Географик . Архивировано из оригинала 4 ноября 2013 года . Проверено 10 июня 2012 г.
- ^ Перейти обратно: а б с д и ж Теслер, П. (1999). «Удивительная адаптируемая лягушка» . Эксплораториум:: Музей науки, искусства и человеческого восприятия . Проверено 4 июня 2012 г.
- ^ Винсент, Л. (2001). « Литория голубая » (PDF ) Университет Джеймса Кука. Архивировано из оригинала (PDF) 22 апреля . Получено 3 августа ,
- ^ Эмерсон, С.Б.; Диль, Д. (1980). «Морфология подушечек пальцев и механизмы прилипания у лягушек». Биологический журнал Линнеевского общества . 13 (3): 199–216. дои : 10.1111/j.1095-8312.1980.tb00082.x .
- ^ Харви, МБ; Пембертон, Эй Джей; Смит, Э.Н. (2002). «Новые и малоизвестные лягушки-парашютисты (Rhacophoridae: Rhacophorus ) с Суматры и Явы». Герпетологические монографии . 16 : 46–92. doi : 10.1655/0733-1347(2002)016[0046:NAPKPF]2.0.CO;2 . S2CID 86616385 .
- ^ «Лопастика диванная ( Scaphiopus Couchi )» . Музей пустыни Аризона-Сонора . Проверено 3 августа 2012 г.
- ^ Уокер, М. (25 июня 2009 г.). «Тайна безногих лягушек раскрыта» . Новости Би-би-си .
- ^ Стеббинс, Роберт С .; Коэн, Натан В. (1995). Естественная история амфибий . Издательство Принстонского университета. стр. 10–14. ISBN 978-0-691-03281-8 .
- ^ Фрост, Юго-Запад (1932). «Заметки о питании и линьке лягушек». Американский натуралист . 66 (707): 530–540. дои : 10.1086/280458 . JSTOR 2456779 . S2CID 84796411 .
- ^ Перейти обратно: а б Бэджер, Д.; Нетертон, Дж. (1995). Лягушки . Издательство Эйрлайф. п. 27. ISBN 978-1-85310-740-5 .
- ^ Смит, HR (1962). Земноводные и их пути . Макмиллан. ISBN 978-0-02-612190-3 .
- ^ Дикерсон, MC (1969). Книга о лягушках: Североамериканские лягушки и жабы . Дуврские публикации. ISBN 978-0-486-21973-8 .
- ^ Блэкберн, округ Колумбия (14 ноября 2002 г.). « Трихобатрахус робустус » . АмфибияВеб . Проверено 18 августа 2012 г.
- ^ Руибаль, Родольфо; Шумейкер, Воган (1985). «Остеодермы у бесхвостых». Журнал герпетологии . 18 (3): 313–328. дои : 10.2307/1564085 . JSTOR 1564085 .
- ^ Витт, Лори Дж.; Колдуэлл, Джанали П. (2013). Герпетология: вводная биология амфибий и рептилий . Академическая пресса. п. 50. ISBN 9780123869203 .
- ^ Джаред, К.; Антониацци, ММ; Навас, Калифорния; Качбурян, Э.; Фреймюллер, Э.; Тамбурги, Д.В.; Родригес, Монтана (2005). «Окостенение головы, фрагмоз и защита у каскоголовой древесной лягушки Corythomantis greeningi ». Журнал зоологии . 265 (1): 1–8. дои : 10.1017/S0952836904005953 . S2CID 59449901 .
- ^ Перейти обратно: а б Бертон, Морис (1972). Книга наблюдателя о британских диких животных . Фредерик Уорн и компания, стр. 204–209. ISBN 978-0-7232-1503-5 .
- ^ Гуаясамин, Хуан М.; Крынак, Тим; Крынак, Кэтрин; Кулебрас, Хайме; Хаттер, Карл Р. (2015). «Фенотипическая пластичность поднимает вопросы по таксономически важным признакам: замечательная новая андская дождевая лягушка (Pristimantis) со способностью изменять текстуру кожи: Фенотипическая пластичность андской дождевой лягушки» . Зоологический журнал Линнеевского общества . 173 (4): 913–928. дои : 10.1111/zoj.12222 .
- ^ Венте, штат Вашингтон; Филлипс, Дж. Б. (2003). «Фиксированные зеленые и коричневые цветовые морфы и новая меняющая цвет морфа тихоокеанской древесной лягушки Hyla regilla ». Американский натуралист . 162 (4): 461–473. дои : 10.1086/378253 . JSTOR 10.1086/378253 . ПМИД 14582008 . S2CID 25692966 .
- ^ Буасвер, Адам (23 октября 2007 г.). « Барбурула калимантанская » . АмфибияВеб . Калифорнийский университет в Беркли . Проверено 9 июля 2012 г.
- ^ Кимбалл, Джон (2010). «Системы кровообращения животных: три камеры: лягушка и ящерица» . Страницы биологии Кимбалла . Архивировано из оригинала 11 мая 2016 года . Проверено 9 июля 2012 г.
- ^ Ли, Дебора (23 апреля 2010 г.). « Тельматобиус кулеус » . АмфибияВеб . Калифорнийский университет в Беркли . Проверено 9 июля 2012 г.
- ^ Секрет липкости лягушачьей слюны | СТАО
- ^ Левин, Р.П.; Монрой, Дж.А.; Брейнерд, Эл. (15 марта 2004 г.). «Вклад ретракции глаз в глотательную способность северной леопардовой лягушки Rana pipiens » . Журнал экспериментальной биологии . 207 (Часть 8): 1361–1368. дои : 10.1242/jeb.00885 . ПМИД 15010487 .
- ^ «Пищеварительная система лягушки» . TutorVista.com. 2010. Архивировано из оригинала 3 июня 2010 года . Проверено 4 августа 2012 г.
- ^ Перейти обратно: а б Дорит, РЛ; Уокер, ВФ; Барнс, Р.Д. (1991). Зоология . Издательство Колледжа Сондерса. п. 849 . ISBN 978-0-03-030504-7 .
- ^ Перейти обратно: а б с д «Внутренние системы лягушки» . TutorVista.com. 2010. Архивировано из оригинала 21 января 2008 года . Проверено 4 июня 2012 г.
- ^ Дуэллман, МЫ и Л. Труб (1986). Биология амфибий . Нью-Йорк: Издательская компания McGraw-Hill.
- ^ Север, Дэвид М.; Стауб, Нэнси Л. «Гормоны, половые принадлежности и вторичные половые признаки у амфибий» (PDF) . Гормоны и размножение позвоночных – Том 2: Земноводные . стр. 83–98. Архивировано (PDF) оригинала 9 октября 2022 г. Проверено 4 августа 2012 г.
- ^ Лорен, Мишель; Готье, Жак А. (2012). «Амниота» . Веб-проект «Древо жизни» . Проверено 4 августа 2012 г.
- ^ Перейти обратно: а б Ховард, Ян П.; Роджерс, Брайан Дж. (1995). Бинокулярное зрение и стереопсис . Издательство Оксфордского университета. п. 651. ИСБН 978-0195084764 .
- ^ Перейти обратно: а б с Бэджер, Дэвид; Нетертон, Джон (1995). Лягушки . Airlife Publishing Ltd., стр. 31–35. ISBN 978-1-85310-740-5 .
- ^ Мунц, WRA; Отпечатки Scientific American (1964). «Зрение у лягушек». Научный американец . 210 (3). WH Фриман: 110–9. АСИН B0006RENBO . Бибкод : 1964SciAm.210c.110M . doi : 10.1038/scientificamerican0364-110 . ОСЛК 15304238 . ПМИД 14133069 .
- ^ Кельбер, Альмут; Йованович, Карола; Олссон, Питер (5 апреля 2017 г.). «Пороги и шумовые ограничения цветового зрения при тусклом свете» . Философские труды Королевского общества B: Биологические науки . 372 : 20160065.doi : (1717 ) 10.1098/rstb.2016.0065 . ISSN 0962-8436 . ПМК 5312015 . ПМИД 28193810 .
- ^ Перейти обратно: а б Бэджер, Дэвид; Нетертон, Джон (1995). Лягушки . Издательство Эйрлайф. п. 38. ISBN 978-1-85310-740-5 .
- ^ Перейти обратно: а б Стеббинс, Роберт С .; Коэн, Натан В. (1995). Естественная история амфибий . Издательство Принстонского университета. стр. 67–69. ISBN 978-0-691-03281-8 .
- ^ Армстронг, Сесилия Э.; Робертс, Уильям М. (1998). «Электрические свойства волосковых клеток мешочка лягушки: искажение в результате ферментативной диссоциации» . Журнал неврологии . 18 (8): 2962–2973. doi : 10.1523/JNEUROSCI.18-08-02962.1998 . ПМК 6792591 . ПМИД 9526013 .
- ^ «Лягушка-бык» . Департамент природных ресурсов штата Огайо. Архивировано из оригинала 18 августа 2012 года . Проверено 19 июня 2012 г.
- ^ Тан, В.-Х.; Цай, К.-Г.; Лин, К.; Лин, Ю.К. (5 июня 2014 г.). «Эффект городского каньона: ливневые стоки улучшают звуковые характеристики древесной лягушки Мьентьен» . Журнал зоологии . 294 (2): 77–84. дои : 10.1111/jzo.12154 . ISSN 0952-8369 .
- ^ Нафис, Гэри (2012). « Ascaphus truei : прибрежная хвостатая лягушка» . Калифорнийский герпс . Проверено 19 июня 2012 г.
- ^ Рой, Дебжани (1997). «Коммуникационные сигналы и половой отбор у амфибий» (PDF) . Современная наука . 72 : 923–927. Архивировано из оригинала (PDF) 23 сентября 2012 года.
- ^ Герхардт, ХК (1994). «Эволюция вокализации у лягушек и жаб». Ежегодный обзор экологии и систематики . 25 : 293–324. doi : 10.1146/annurev.es.25.110194.001453 .
- ^ Перейти обратно: а б с д Бэджер, Дэвид; Нетертон, Джон (1995). Лягушки . Airlife Publishing Ltd., стр. 39–44. ISBN 978-1-85310-740-5 .
- ^ Хилтон, Билл младший (1986). «9. «Кувшин-о-ром»: Зов влюбчивой лягушки». Пьемонтский натуралист, Том 1 . Йорк, Южная Каролина: Центр естественной истории Пьемонта Хилтон Понд. ISBN 978-0-9832151-0-3 .
- ^ Нэш, Пэт (февраль 2005 г.). «РРРРРРРЗахватывающая жизнь древесных лягушек» . Архивировано из оригинала 9 марта 2012 года . Проверено 4 августа 2012 г.
- ^ Аристофан. «Лягушки» . Архивировано из оригинала 13 мая 2012 года . Проверено 19 июня 2012 г.
- ^ Сазерс, РА; Наринс, премьер-министр; Лин, В; Шницлер, Х; Денцингер, А; Сюй, С; Фэн, А.С. (2006). «Голоса мертвых: сложные нелинейные голосовые сигналы из гортани ультразвуковой лягушки» . Журнал экспериментальной биологии . 209 (24): 4984–4993. дои : 10.1242/jeb.02594 . ПМИД 17142687 .
- ^ Эммер, Рик (24 ноября 1997 г.). «Как лягушки переживают зиму? Почему они не замерзают насмерть?» . Научный американец . Проверено 15 июня 2012 г.
- ^ Кейс, Сара М.; Судорога, Ребекка Л.; Франклин, Крейг Э. (2009). «Метаболическая депрессия во время летнего периода у Cyclorana alboguttata ». Сравнительная биохимия и физиология – Часть A: Молекулярная и интегративная физиология . 154 (4): 557–563. дои : 10.1016/j.cbpa.2009.09.001 . ПМИД 19737622 .
- ^ Хадсон, Нью-Джерси; Ленерт, SA; Ингхэм, AB; Саймондс, Б.; Франклин, CE; Харпер, Дж.С. (2005). «Уроки спящей лягушки: экономия мышечного белка, несмотря на голодание и неиспользование». AJP: Регуляторная, интегративная и сравнительная физиология . 290 (3): R836–R843. дои : 10.1152/ajpregu.00380.2005 . ПМИД 16239372 . S2CID 8395980 .
- ^ Уилмер, Пэт (2009). Экологическая физиология животных . Уайли. стр. 188 . ISBN 9781405107242 .
- ^ «Топ-10 лучших животных-прыгунов» . Научный луч. Архивировано из оригинала 10 августа 2012 года . Проверено 11 июня 2012 г.
- ^ Джеймс, РС; Уилсон, Р.С. (2008). «Взрывные прыжки: крайние морфологические и физиологические специализации австралийских ракетных лягушек ( Litoria nasuta )» (PDF) . Физиологическая и биохимическая зоология . 81 (2): 176–185. дои : 10.1086/525290 . ПМИД 18190283 . S2CID 12643425 . Архивировано (PDF) из оригинала 9 октября 2022 г.
- ^ Наувелартс, С.; Шольер, Дж.; Аэртс, П. (2004). «Функциональный анализ того, как лягушки выпрыгивают из воды» . Биологический журнал Линнеевского общества . 83 (3): 413–420. дои : 10.1111/j.1095-8312.2004.00403.x .
- ^ Перейти обратно: а б Бэджер, Дэвид; Нетертон, Джон (1995). Лягушки . Издательство Эйрлайф. п. 51. ИСБН 978-1-85310-740-5 .
- ^ Эстли, ХК; Робертс, Ти Джей (2011). «Доказательства существования катапульты позвоночных: накопление упругой энергии в сухожилии подошвенной мышцы во время прыжка лягушки» . Письма по биологии . 8 (3): 386–389. дои : 10.1098/rsbl.2011.0982 . ПМЦ 3367733 . ПМИД 22090204 .
- ^ Скотт, Дж. (2005). «Прыжок саранчи: комплексное лабораторное исследование». Достижения в области физиологического образования . 29 (1): 21–26. дои : 10.1152/advan.00037.2004 . ПМИД 15718379 . S2CID 13423666 .
- ^ Перейти обратно: а б с Бакли, ЧР; Майкл, Сан-Франциско; Иршик, диджей (2005). «Раннее вылупление снижает прыгучесть у лягушки прямого развития Eleutherodactylus coqui» . Функциональная экология . 19 (1): 67–72. Бибкод : 2005FuEco..19...67B . дои : 10.1111/j.0269-8463.2005.00931.x . ISSN 0269-8463 . JSTOR 3599272 .
- ^ Перейти обратно: а б с Цуг, Джордж Р.; Дуэллман, Уильям Э. (14 мая 2014 г.). «Анура» . Британская онлайн-энциклопедия . Проверено 26 апреля 2015 г.
- ^ Фитч, HS (1956). «Экологическое исследование ящерицы с воротником ( Crotaphytus воротникис )». Публикации Канзасского университета . 8 : 213–274.
- ^ Уолтон, М.; Андерсон, Б.Д. (1988). «Аэробные затраты на скачкообразное передвижение у жабы птицелова ( Bufo woodhousei fowleri )» . Журнал экспериментальной биологии . 136 (1): 273–288. дои : 10.1242/jeb.136.1.273 . ПМИД 3404074 .

- ^ Ан, АН; Борозда, Э.; Бивенер, А.А. (2004). «Ходит и бегает красноногая бегущая лягушка Kassina maculata » . Журнал экспериментальной биологии . 207 (Часть 3): 399–410. дои : 10.1242/jeb.00761 . ПМИД 14691087 .
- ^ Пикерсгилл, М.; Шиотц, А.; Хауэлл, К.; Минтер, Л. (2004). « Кассина пятнистая » . Красный список исчезающих видов МСОП . 2004 . Проверено 11 июня 2012 г.
- ^ Перейти обратно: а б «Пипиды» . АмфибияВеб . Калифорнийский университет в Беркли . Проверено 14 июня 2012 г.
- ^ Перейти обратно: а б Дуэллман, МЫ; Цуг Г.Р. «Анура: от головастика до взрослой особи» . Британская энциклопедия . Проверено 13 июля 2012 г.
- ^ Радхакришнан, К.; Гопи, КК (2007). «Расширение ареала распространения Nasikabatrachus sahyadrensis Biju & Bossuyt (Amphibia: Anura: Nasikabatrachidae) вдоль Западных Гат, с некоторым пониманием его биономики» (PDF) . Современная наука . 92 (2): 213–216. ISSN 0011-3891 . Архивировано (PDF) из оригинала 9 октября 2022 г.
- ^ Перейти обратно: а б Фаррар, Евгения; Привет, Джейн. « Спеа бомбифронс » . АмфибияВеб . Калифорнийский университет в Беркли . Проверено 16 июня 2012 г.
- ^ «Скафиоподиды» . АмфибияВеб . Калифорнийский университет в Беркли . Проверено 16 июня 2012 г.
- ^ Робертс, Дейл; Герой, Жан-Марк (2011). « Гелеиопорус альбопунктатус » . Красный список исчезающих видов МСОП . 2011 . Проверено 16 июня 2012 г.
- ^ Станишевский, Марк (30 сентября 1998 г.). «Мадагаскарские роющие лягушки: Род: Scaphiophryne (Boulenger, 1882) » Получено 16 , июня
- ^ Венески, М; Раксуорси, CJ; Нуссбаум, РА; Гло, Ф. (2003). «Пересмотр комплекса мраморных жаб Scaphiophryne marmorata с Мадагаскара, включая описание нового вида» (PDF) . Герпетологический журнал . 13 : 69–79. Архивировано (PDF) из оригинала 9 октября 2022 г.
- ^ Федерле, В.; Барнс, WJP; Баумгартнер, В.; Дрекслер, П.; Смит, Дж. М. (2006). «Мокро, но не скользко: граничное трение в клейких подушечках пальцев древесных лягушек» . Журнал интерфейса Королевского общества . 3 (10): 689–697. дои : 10.1098/rsif.2006.0135 . ПМЦ 1664653 . ПМИД 16971337 .
- ^ Кокран, Дорис Мейбл (1961). Живые амфибии мира . Даблдэй. п. 112. ИСБН 978-0-241-90338-4 .
- ^ « Филломедуза, да! » . АмфибияВеб . Проверено 14 июня 2012 г.
- ^ Эмерсон, Шэрон Б.; Кёль, Маргарита (1990). «Взаимодействие поведенческих и морфологических изменений в эволюции нового локомоторного типа: «летающие лягушки». Эволюция . 44 (8): 1931–1946. : 10.2307 /2409604 . JSTOR 2409604. . PMID 28564439 doi
- ^ Шах, Санни; Тивари, Рахна (29 ноября 2001 г.). « Ракофорус нигропальматус » . АмфибияВеб . Калифорнийский университет в Беркли . Проверено 11 июня 2012 г.
- ^ «Летающая лягушка Уоллеса Rhacophorus nigropalmatus » . National Geographic: Животные. 10 сентября 2010 года. Архивировано из оригинала 18 января 2010 года . Проверено 5 июня 2012 г.
- ^ Перейти обратно: а б с д и ж г Стеббинс, Роберт С .; Коэн, Натан В. (1995). Естественная история амфибий . Издательство Принстонского университета. стр. 154–162. ISBN 978-0-691-03281-8 .
- ^ Дэвис, Северная Каролина; Холлидей, TR (1978). «Глубокое карканье и оценка боя у жаб Bufo bufo ». Природа . 274 (5672): 683–685. Бибкод : 1978Natur.274..683D . дои : 10.1038/274683a0 . S2CID 4222519 .
- ^ Лонг, Дэвид Р. (1989). «Энергетика и размножение самок Scaphiopus multiplicatus из Западного Техаса». Журнал герпетологии . 23 (2): 176–179. дои : 10.2307/1564026 . JSTOR 1564026 .
- ^ Искандар, DT; Эванс, Би Джей; МакГуайр, Дж.А. (2014). «Новый репродуктивный способ лягушек: новый вид клыкастой лягушки с внутренним оплодотворением и рождением головастиков» . ПЛОС ОДИН . 9 (12): e115884. Бибкод : 2014PLoSO...9k5884I . дои : 10.1371/journal.pone.0115884 . ПМК 4281041 . ПМИД 25551466 .
- ^ Ченнинг, Алан; Хауэлл, Ким М. (2006). Земноводные Восточной Африки . Издательство Комсток. стр. 104–117. ISBN 978-0-8014-4374-9 .
- ^ Сандбергер, Л.; Хиллерс, А.; Думбия, Дж.; Луа, Н.С.; Бреде С.; Рёдель, Миссури. (2010). «Повторное открытие либерийской жабы нимба, Nimbaphrynoides liberiensis (Xavier, 1978) (Amphibia: Anura: Bufonidae) и повторная оценка ее таксономического статуса» (PDF) . Зоотакса . 4355 : 56–68. дои : 10.11646/zootaxa.2355.1.3 . ISSN 1175-5334 . Архивировано (PDF) из оригинала 9 октября 2022 г.
- ^ Мэттисон, Крис (2011). Лягушки и жабы миров . Издательство Принстонского университета. п. 91. ИСБН 978-0-691-14968-4 .
- ^ Дуэллман, МЫ; Цуг Г.Р. «Анура: кладка и высиживание яиц» . Британская энциклопедия . Проверено 9 апреля 2022 г.
- ^ Дуэллман, Уильям Э.; Труб, Линда; Арак, Энтони (2002). «Лягушки и жабы». В Холлидее, Тим; Адлер, Крейг (ред.). Энциклопедия рептилий и амфибий Firefly . Книги Светлячка. п. 67. ИСБН 978-1-55297-613-5 .
- ^ Эстрада, Альберто Р.; Хеджес, С. Блэр (1996). «На нижнем пределе размеров четвероногих: новая миниатюрная лягушка с Кубы (Leptodactylidae: Eleutherodactylus)». Копейя . 1996 (4): 852–859. дои : 10.2307/1447647 . JSTOR 1447647 .
- ^ Уиттакер, Келли; Чантасиривисал, Пира (2 декабря 2005 г.). « Лептодактилус пятипалый » . АмфибияВеб . Калифорнийский университет в Беркли . Проверено 19 июля 2012 г.
- ^ Уиттакер, Келли (27 июня 2007 г.). « Агаличнис каллидриас » . АмфибияВеб . Калифорнийский университет в Беркли . Проверено 19 июля 2012 г.
- ^ Гилберт, Перри В. (1942). «Наблюдения за яйцами Ambystoma maculatum с особым упором на зеленые водоросли, обнаруженные в оболочках яиц». Экология . 23 (2): 215–227. Бибкод : 1942Экол...23..215Г . дои : 10.2307/1931088 . JSTOR 1931088 .
- ^ Уолдман, Брюс; Райан, Майкл Дж. (1983). «Термические преимущества совместного отложения яичной массы у лесных лягушек ( Rana sylvatica )». Журнал герпетологии . 17 (1): 70–72. дои : 10.2307/1563783 . JSTOR 1563783 .
- ^ Варкентин, К.М. (1995). «Адаптивная пластичность в возрасте вылупления: ответ на компромиссы с риском нападения хищников» . Труды Национальной академии наук . 92 (8): 3507–3510. Бибкод : 1995PNAS...92.3507W . дои : 10.1073/pnas.92.8.3507 . ПМК 42196 . ПМИД 11607529 .
- ^ Перейти обратно: а б с д и Стеббинс, Роберт С .; Коэн, Натан В. (1995). Естественная история амфибий . Издательство Принстонского университета. стр. 179–194. ISBN 978-0-691-03281-8 .
- ^ Гольдберг, Хавьер; Таучче, Педро П.Г.; КИНИЦИО, Сильвия Инес; Хаддад, Селио ФБ; Вера Кандиоти, Флоренция (1 января 2020 г.). «Улучшение наших знаний о развитии прямых лягушек: онтогенез Ischnocnema henselii (Anura: Brachycephalidae)» . Зоологический обзор . 284 :78–87. Бибкод : 2020ЗооАн.284...78Г . дои : 10.1016/j.jcz.2019.11.001 . ISSN 0044-5231 . S2CID 209576535 .
- ^ Элинсон, Ричард П. (2001). «Прямая разработка: альтернативный способ сделать лягушку» . Бытие . 29 (2): 91–95. doi : 10.1002/1526-968X(200102)29:2<91::AID-GENE1009>3.0.CO;2-6 . ISSN 1526-968X . ПМИД 11170349 . S2CID 32550621 .
- ^ Сешадри, КС (2015). «Лягушки-ракофориды, размножающиеся в бамбуке: открытие нового репродуктивного способа в Западных Гатах» . ЛягушкаЖурнал . 23 (4) (116): 46–49.
- ^ Викрамасингхе, Дипти Д.; Осеин, Керри Л.; Вассерсуг, Ричард Дж. (2007). «Онтогенетические изменения в диете и морфологии кишечника у полуземных головастиков Nannophrys ceylonensis (Dicroglossidae)». Копейя . 2007 (4): 1012–1018. doi : 10.1643/0045-8511(2007)7[1012:ocidai]2.0.co;2 . JSTOR 25140719 . S2CID 86244218 .
- ^ Янзен, Питер (10 мая 2005 г.). « Наннофрис цейлоненсис » . АмфибияВеб . Калифорнийский университет в Беркли . Проверено 20 июля 2012 г.
- ^ Хофф, К. против С.; Вассерсуг, Р.Дж. (1 февраля 2000 г.). «Локомоция головастика: осевое движение и функции хвоста у позвоночных, практически лишенных позвонков» . Интегративная и сравнительная биология . 40 (1): 62–76. дои : 10.1093/icb/40.1.62 .
- ^ Крамп, Марта Л. (1986). «Каннибализм молодых головастиков: еще одна опасность метаморфоза». Копейя . 1986 (4): 1007–1009. дои : 10.2307/1445301 . JSTOR 1445301 .
- ^ Стеббинс, Роберт С .; Коэн, Натан В. (1995). Естественная история амфибий . Издательство Принстонского университета. стр. 173–175. ISBN 978-0-691-03281-8 .
- ^ Бендер, МК; Ху, К.; Пеллетье, К.; Денвер, Р.Дж. (2018). «Есть или не есть: онтогенез гипоталамического контроля питания и роль лептина в модуляции перехода жизненного цикла у головастиков амфибий» . Слушания. Биологические науки . 285 (1875). дои : 10.1098/rspb.2017.2784 . ПМЦ 5897637 . ПМИД 29593109 .
- ^ Головастики едят как сумасшедшие, прежде чем перестроить кишечник - Будущее
- ^ Коль, К.Д.; Кэри, ТЛ; Карасов, WH; Диринг, доктор медицины (2013). «Реструктуризация микробиоты кишечника амфибий посредством метаморфоза» . Отчеты по экологической микробиологии . 5 (6): 899–903. Бибкод : 2013EnvMR...5..899K . дои : 10.1111/1758-2229.12092 . ПМИД 24249298 .
- ^ Фабрези, Марисса; Крус, Хулио Сезар (2020). «Аспекты эволюции и развития диеты и морфологии кишечника головастиков цератофрид (Anura)» . Биология развития BMC . 20 (1): 16. дои : 10.1186/s12861-020-00221-5 . ПМЦ 7388516 . ПМИД 32723314 . S2CID 255787646 .
- ^ Балинский Борис Иван. «Развитие животных: Метаморфозы» . Британская энциклопедия . Проверено 10 августа 2012 г.
- ^ Мэттисон, Крис (2011). Лягушки и жабы миров . Издательство Принстонского университета. п. 107. ИСБН 978-0-691-14968-4 .
- ^ Мэттисон, Крис (2011). Лягушки и жабы миров . Издательство Принстонского университета. стр. 65–68. ISBN 978-0-691-14968-4 .
- ^ Дуэллман, МЫ; Цуг Г.Р. «Анура: Особенности питания» . Британская энциклопедия . Проверено 9 апреля 2022 г.
- ^ де-Оливейра-Ногейра, Карлос Энрике; СОУЗА, Убирата Феррейра; МАЧАДО, Тайнара Мендес; Фигейреду-де-Андраде, Кайо Антониу; Моника, Александр Таманини; САЗИМА Иван; САЗИМА, Марлис; Толедо, Луис Фелипе (июнь 2023 г.). «Среди фруктов, цветов и нектара: Необыкновенный рацион лягушки Xenohyla truncata» . Пищевые сети . 35 : e00281. Бибкод : 2023FWebs..3500281D . doi : 10.1016/j.fooweb.2023.e00281 .
- ^ Да Силва, HR; Де Бритто-Перейра, XC (2006). «Сколько фруктов едят фруктовоядные лягушки? Исследование рациона Xenohyla truncata (Lissamphibia: Anura: Hylidae)». Журнал зоологии . 270 (4): 692–698. дои : 10.1111/j.1469-7998.2006.00192.x .
- ^ «Диета неотропической лягушки Leptodactylus mystaceus» (PDF) . Архивировано из оригинала (PDF) 7 апреля 2014 года . Проверено 2 апреля 2014 г.
- ^ Камера, Бруно Ф.; Кринский, Дионес; Болд, Изабелла А. (4 февраля 2014 г.). «Диета неотропической лягушки Leptodactylus mystaceus (Anura: Leptodactylidae)» (PDF ) Герпетологические заметки . 7 : 31–36. Архивировано из оригинала (PDF) 24 сентября . Получено 26 . апреля
- ^ Дас, И. (апрель 1996 г.). «Фоливорность и сезонные изменения в рационе Rana hexadactyla (Anura: Ranidae)» (PDF) . Журнал зоологии . 238 (4): 785–794. дои : 10.1111/j.1469-7998.1996.tb05430.x .
- ^ Мэттисон, Крис (2011). Лягушки и жабы миров . Издательство Принстонского университета. п. 69. ИСБН 978-0-691-14968-4 .
- ^ Грэм, Донна. «Северная леопардовая лягушка ( Rana pipiens )» . Путеводитель для преподавателей по природным ресурсам Южной Дакоты . Архивировано из оригинала 23 июня 2012 года . Проверено 4 августа 2012 г.
- ^ Пимм, Стюарт Л. (1979). «Структура пищевых сетей» (PDF) . Теоретическая популяционная биология . 16 (2): 144–158. Бибкод : 1979TPBio..16..144P . дои : 10.1016/0040-5809(79)90010-8 . ПМИД 538731 . Архивировано из оригинала (PDF) 27 сентября 2011 года.
- ^ Мэтьюз, КР; Мио, К. (2007). «Скелетохронологическое исследование возрастной структуры, роста и продолжительности жизни горной желтоногой лягушки Rana muscosa в Сьерра-Неваде, Калифорния». Копейя . 2007 (4): 986–993. doi : 10.1643/0045-8511(2007)7[986:ASSOTA]2.0.CO;2 . ISSN 0045-8511 . S2CID 86237494 . (требуется подписка)
- ^ Славенс, Фрэнк; Славенс, Кейт. «Индекс лягушки и жабы» . Долголетие . Архивировано из оригинала 18 августа 2012 года . Проверено 4 июля 2012 г.
- ^ Стори, КБ (1990). «Жизнь в замороженном состоянии: адаптивные стратегии естественной устойчивости к заморозкам у амфибий и рептилий». Американский журнал физиологии . 258 (3 ч. 2): 559–568. дои : 10.1152/ajpregu.1990.258.3.R559 . ПМИД 2180324 .
- ^ Крамп, ML (1996). «Родительская забота среди амфибий». Родительская забота: эволюция, механизмы и адаптивное значение . Достижения в изучении поведения. Том. 25. С. 109–144. дои : 10.1016/S0065-3454(08)60331-9 . ISBN 978-0-12-004525-9 .
- ^ Перейти обратно: а б с Браун, Дж.Л.; Моралес, В.; Саммерс, К. (2010). «Ключевая экологическая черта привела к развитию заботы о двух родителях и моногамии у амфибий». Американский натуралист . 175 (4): 436–446. дои : 10.1086/650727 . ПМИД 20180700 . S2CID 20270737 .
- ^ Шеридан, Дженнифер А.; Окок, Джоан Ф. (2008). «Родительская забота о Chiromantis hansenae (Anura: Rhacophoridae)». Копейя . 2008 (4): 733–736. дои : 10.1643/CH-07-225 . S2CID 85122799 .
- ^ Браун, Дж.Л.; Моралес, В.; Саммерс, К. (2008a). «Расхождения в родительской заботе, выборе среды обитания и истории жизни личинок двух видов перуанских ядовитых лягушек: экспериментальный анализ». Журнал эволюционной биологии . 21 (6): 1534–1543. дои : 10.1111/j.1420-9101.2008.01609.x . ПМИД 18811668 . S2CID 29546555 .
- ^ Грант, Т.; Фрост, ДР; Колдуэлл, JP; Гальярдо, Р.; Хаддад, ЦФБ; Кокс, PJR; Значит, БД; Нунан, БП; Шаргел, МЫ; Уилер, В. (2006). «Филогенетическая систематика древолазов и их родственников (Amphibia, Atesphatanura, Dendrobatidae)» (PDF) . Бюллетень Американского музея естественной истории . 299 : 1–262. CiteSeerX 10.1.1.693.8392 . doi : 10.1206/0003-0090(2006)299[1:PSODFA]2.0.CO;2 . hdl : 2246/5803 . ISSN 0003-0090 . S2CID 82263880 . Архивировано из оригинала (PDF) 4 апреля . Получено 13 , ноября
- ^ «Виноградники», Рене; Боланьос, Федерико (1992). «Родительская забота Dendrobates granuliferus (Anura: Dendrobatidae) с описанием головастика». Журнал герпетологии . 26 (1): 102–105. дои : 10.2307/1565037 . JSTOR 1565037 .
- ^ Фандиньо, Мария Клаудия; Люддеке, Хорст; Амескита, Адольфо (1997). «Вокализация и транспортировка личинок самца Colostethus subpunctatus (Anura: Dendrobatidae)». Земноводные-рептилии . 18 (1): 39–48. дои : 10.1163/156853897X00297 .
- ^ «Обыкновенная жаба-повитуха, Alytes obstetricans » . Repties et Amphibiens de France . Проверено 30 июля 2012 г.
- ^ «Белки пенных гнезд лягушек» . Университет Глазго. Архивировано из оригинала 3 июня 2013 года . Проверено 24 августа 2012 г.
- ^ Далгетти, Лаура; Кеннеди, Малкольм В. (2010). «Строительство дома из пенопласта — архитектура гнезда из пенопласта тунгарской лягушки и трехэтапный процесс строительства» . Письма по биологии . 6 (3): 293–296. дои : 10.1098/rsbl.2009.0934 . ПМЦ 2880057 . ПМИД 20106853 .
- ^ « Асса Дарлингтони » . Сеть лягушек Австралии . 2005 . Проверено 5 августа 2012 г.
- ^ Семейн, Э. (2002). « Реобатрах силус » . Сеть разнообразия животных . Зоологический музей Мичиганского университета . Проверено 5 августа 2012 г.
- ^ Сандмейер, Фрэн (12 марта 2001 г.). « Ринодерма Дарвинии » . АмфибияВеб . Проверено 5 августа 2012 г.
- ^ Бартальмус, Джордж Т.; Зелински, Уильям Дж. (1988). « Слизь кожи Xenopus вызывает дискинезию полости рта, которая способствует бегству от змей». Фармакология Биохимия и поведение . 30 (4): 957–959. дои : 10.1016/0091-3057(88)90126-8 . ПМИД 3227042 . S2CID 25434883 .
- ^ Дарст, Кэтрин Р.; Каммингс, Молли Э. (2006). «Обучение хищников способствует имитации менее токсичной модели ядовитых лягушек» . Природа . 440 (7081): 208–211. Бибкод : 2006Natur.440..208D . дои : 10.1038/nature04297 . ПМИД 16525472 .
- ^ Майерс, CW; Дейли, JW (1983). «Лягушки-дротики». Научный американец . 248 (2): 120–133. Бибкод : 1983SciAm.248b.120M . doi : 10.1038/scientificamerican0283-120 . ПМИД 6836257 .
- ^ Сэвидж, Дж. М. (2002). Земноводные и рептилии Коста-Рики . Издательство Чикагского университета. ISBN 978-0-916984-16-8 .
- ^ Дуэллман, МЫ (1978). «Биология экваториальной герпетофауны в Амазонском Эквадоре» (PDF) . Разные публикации Музея естественной истории Канзасского университета . 65 . Архивировано из оригинала (PDF) 4 июля 2011 года.
- ^ Сапорито, РА; Гарраффо, HM; Доннелли, Массачусетс; Эдвардс, Алабама; Лонгино, Джей Ти; Дейли, JW (2004). «Муравьиные муравьи: источник членистоногих алкалоидов пумилиотоксина дендробатидных ядовитых лягушек» . Труды Национальной академии наук . 101 (21): 8045–8050. Бибкод : 2004PNAS..101.8045S . дои : 10.1073/pnas.0402365101 . ПМК 419554 . ПМИД 15128938 .
- ^ Смит, БП; Тайлер, MJ; Канеко, Т.; Гарраффо, HM; Спанде, ТФ; Дейли, JW (2002). «Доказательства биосинтеза алкалоидов псевдофринамина австралийской миобатрахидной лягушкой ( Pseudophryne ) и секвестрации пищевых пумилиотоксинов». Журнал натуральных продуктов . 65 (4): 439–47. дои : 10.1021/np010506a . ПМИД 11975476 .
- ^ Грант, Т. «Виварий ядовитых лягушек-дротозов» . Американский музей естественной истории . Проверено 7 июля 2012 г.
- ^ Арнольд, Николас; Овенден, Денис (2002). Рептилии и амфибии Британии и Европы . Издательство Харпер Коллинз. стр. 73–74. ISBN 978-0-00-219964-3 .
- ^ Хейс, Флойд Э. (1989). «Антихищное поведение недавно метаморфизованных жаб ( Bufo a. americanus ) во время встреч с подвязочными змеями ( Thamnophis s. sirtalis )». Копейя . 1989 (4): 1011–1015. дои : 10.2307/1445987 . JSTOR 1445987 .
- ^ «Чумовые лягушки» . Национальный географический исследователь. Архивировано из оригинала 6 марта 2004 года . Проверено 13 июля 2012 г.
- ^ Феррелл, Вэнс (4 марта 2012 г.). «Географическое распространение» . Энциклопедия эволюции . Том. 3. Факты эволюции. Архивировано из оригинала 5 мая 2012 года . Проверено 13 июля 2012 г.
- ^ Райан, Пэдди (25 сентября 2011 г.). «Рассказ: Лягушки» . Энциклопедия Новой Зеландии . Проверено 20 августа 2012 г.
- ^ Даль, Крис; Новотный, Войтех; Моравец, Иржи; Ричардс, Стивен Дж. (2009). «Бета-разнообразие лягушек в лесах Новой Гвинеи, Амазонии и Европы: контрастные тропические и умеренные сообщества». Журнал биогеографии . 36 (5): 896–904. Бибкод : 2009JBiog..36..896D . дои : 10.1111/j.1365-2699.2008.02042.x . S2CID 13067241 .
- ^ « Циклорана плоскоголовая » . Сеть лягушек Австралии. 23 февраля 2005 года . Проверено 20 июля 2012 г.
- ^ Хукстра, Дж. М.; Мольнар, Дж.Л.; Дженнингс, М.; Ревенга, К.; Сполдинг, доктор медицины; Баучер, ТМ; Робертсон, Дж. К.; Хейбель, Ти Джей; Эллисон, К. (2010). «Число видов амфибий, находящихся под угрозой исчезновения, по пресноводным экорегионам» . Атлас глобальной охраны природы: изменения, проблемы и возможности изменить ситуацию . Охрана природы . Проверено 5 сентября 2012 г.
- ^ Стюарт, С.Н.; Шансон, Дж.С.; Кокс, Северная Каролина; Янг, Б.Э.; Родригес, ASL; Фишман, Д.Л.; Уоллер, RW (2004). «Состояние и тенденции сокращения и исчезновения земноводных во всем мире» (PDF) . Наука . 306 (5702): 1783–1786. Бибкод : 2004Sci...306.1783S . CiteSeerX 10.1.1.225.9620 . дои : 10.1126/science.1103538 . ПМИД 15486254 . S2CID 86238651 . Архивировано из оригинала (PDF) 27 октября 2017 года . Проверено 27 октября 2017 г.
- ^ Паундс, Дж. Алан; Фогден, Майкл П.Л.; Сэвидж, Джей М.; Горман, Джордж К. (1997). «Испытания нулевых моделей на снижение количества амфибий на тропической горе». Биология сохранения . 11 (6): 1307–1322. Бибкод : 1997ConBi..11.1307P . дои : 10.1046/j.1523-1739.1997.95485.x . S2CID 85382659 .
- ^ «Всемирное сокращение численности амфибий: насколько велика проблема, каковы причины и что можно сделать?» . АмфибияВеб. 22 января 2009 года . Проверено 15 октября 2012 г.
- ^ «Популяция лягушек уменьшается в основном из-за дорожного движения» . Новый учёный . 7 июля 2006 года . Проверено 13 июля 2012 г.
- ^ Вордоу, MJ; Адама, Д.; Хьюстон, Б.; Говиндараджулу, П. (2010). «Распространенность патогенного хитридиевого гриба Batrachochytrium dendrobatidis в находящейся под угрозой исчезновения популяции северных леопардовых лягушек Rana Pipiens » . БМК Экология . 10 (6): 6. Бибкод : 2010BMCE...10....6V . дои : 10.1186/1472-6785-10-6 . ПМЦ 2846871 . ПМИД 20202208 .
- ^ Харп, Элизабет М.; Петранка, Джеймс В. (2006). «Ранавирус лесных лягушек ( Rana sylvatica ): потенциальные источники передачи внутри прудов и между ними» . Журнал болезней дикой природы . 42 (2): 307–318. дои : 10.7589/0090-3558-42.2.307 . ПМИД 16870853 .
- ^ Филлипс, Кэтрин (1994). По следам исчезающих лягушек . Книги о пингвинах. ISBN 978-0-14-024646-9 .
- ^ Липс, Карен Р. (2008). «Упадок тропической горной фауны земноводных». Биология сохранения . 12 (1): 106–117. дои : 10.1111/j.1523-1739.1998.96359.x . JSTOR 2387466 . S2CID 55870526 .
- ^ Блауштайн, Эндрю Р.; Джонсон, Питер Т.Дж. (2003). «Сложность деформированных амфибий» (PDF) . Границы в экологии и окружающей среде . 1 (2): 87–94. doi : 10.1890/1540-9295(2003)001[0087:TCODA]2.0.CO;2 . Архивировано из оригинала (PDF) 29 октября 2013 г.
- ^ Беркхарт, Дж.Г.; Анкли, Г.; Белл, Х.; Карпентер, Х.; Форт, Д.; Гардинер, Д.; Гарднер, Х.; Хейл, Р.; Хелген, JC; Джепсон, П.; Джонсон, Д.; Ланну, М.; Ли, Д.; Лари, Дж.; Леви, Р.; Магнер, Дж.; Метейер, К.; Шелби, доктор медицины; Люсье, Г. (2000). «Стратегии оценки последствий уродливых лягушек для здоровья окружающей среды» . Перспективы гигиены окружающей среды . 108 (1): 83–90. дои : 10.2307/3454299 . JSTOR 3454299 . ПМЦ 1637865 . ПМИД 10620528 .
- ^ Блэк, Ричард (2 октября 2005 г.). «Новый лягушачий центр Лондонского зоопарка» . Новости Би-би-си . Проверено 3 ноября 2008 г.
- ^ «Национальный план восстановления южной лягушки корробори (Pseudophryne corroboree): 5. Предыдущие действия по восстановлению» . Environment.gov.au. Архивировано из оригинала 12 мая 2008 года . Проверено 3 ноября 2008 г.
- ^ «2008: Год Лягушки» . 15 января 2008. Архивировано из оригинала 21 апреля 2012 года . Проверено 13 июля 2012 г.
- ^ Тайлер, Майкл Дж. (1989). Австралийские лягушки . Книги о пингвинах. п. 111. ИСБН 978-0-670-90123-4 .
- ^ Кэмерон, Элизабет (6 сентября 2012 г.). «Тростниковая жаба» . Австралийский музей . Проверено 12 сентября 2012 г.
- ^ ̺ Брагич, Екатерина (20 января 2009 г.). «Аппетит к лягушачьим лапкам наносит вред диким популяциям» . новости АБК .
- ^ Варкентин, И.Г.; Бикфорд, Д.; Содхи, Н.С.; Кори, Дж. А. (2009). «Поедание лягушек до полного вымирания». Биология сохранения . 23 (4): 1056–1059. Бибкод : 2009ConBi..23.1056W . дои : 10.1111/j.1523-1739.2008.01165.x . ПМИД 19210303 . S2CID 1837255 .
- ^ «Информационная программа по культивируемым водным видам: Rana catesbeiana » . ФАО: Департамент рыболовства и аквакультуры . Проверено 5 июля 2012 г.
- ^ Райан Шюсслер (28 января 2016 г.). «Первая проблема горной куриной лягушки: на вкус она как…» National Geographic News . Архивировано из оригинала 29 января 2016 года.
- ^ Марк Твен; Чарльз Дадли Уорнер (1904). Сочинения Марка Твена [псевд.].: Бродяга за границу . Харпер и братья, с. 263.
- ^ «Калифорнийские школы лидируют в гонке за прекращение вскрытий» . Институт защиты животных . 25 апреля 2011 года . Проверено 17 июня 2012 г.
- ^ Уэллс, Дэвид Эймс (1859). Наука об обычных вещах: знакомое объяснение основных принципов физической науки. Для школ, семей и молодых студентов . Айвисон, Финни, Блейкман. п. 290.
- ^ «Лигатура Станниуса» . Биология онлайн . 3 октября 2005 года . Проверено 5 августа 2012 г.
- ^ Саркар, С. (1996). «Ланселот Хогбен, 1895–1975» . Генетика . 142 (3): 655–660. дои : 10.1093/генетика/142.3.655 . ПМК 1207007 . ПМИД 8849876 .
- ^ Браунли, Кристен. «Перенос ядерного оружия: привлечение клонов» . Труды Национальной академии наук Соединенных Штатов Америки . Архивировано из оригинала 14 октября 2012 года . Проверено 21 октября 2012 г.
- ^ Браудер, Л.; Итен Л., ред. (1998). « Xenopus как модельная система в биологии развития» . Динамическое развитие . Университет Калгари . Проверено 17 июня 2012 г.
- ^ Кляйн, С. «Развитие потенциала Xenopus тропического как генетической модели» . Рабочая группа Trans-NIH Xenopus . Проверено 9 марта 2006 г.
- ^ «Список геномов – Геном» . НКБИ . Архивировано из оригинала 13 июля 2019 года . Проверено 7 апреля 2019 г.
- ^ ВанКомпернолле, SE; Тейлор, Р.Дж.; Освальд-Рихтер, К.; Цзян, Дж.; Юри, БЭ; Боуи, Дж. Х.; Тайлер, MJ; Конлон, М.; Уэйд, Д.; и др. (2005). «Антимикробные пептиды из кожи амфибий эффективно подавляют заражение вирусом иммунодефицита человека и передачу вируса от дендритных клеток к Т-клеткам» . Журнал вирусологии . 79 (18): 11598–11606. doi : 10.1128/JVI.79.18.11598-11606.2005 . ПМК 1212620 . ПМИД 16140737 .
- ^ Филипп, Г.; Анжено, Л. (2005). «Последние разработки в области ядов для стрел и дротиков» . Журнал этнофармакологии . 100 (1–2): 85–91. дои : 10.1016/j.jep.2005.05.022 . ПМИД 15993556 .
- ^ Литтл, Т.; Гольдштейн, Д.; Гарц, Дж. (1996). « Жабы Буфо и буфотенин: факты и вымыслы о предполагаемом психоделике». Журнал психоактивных препаратов . 28 (3): 267–290. CiteSeerX 10.1.1.688.5926 . дои : 10.1080/02791072.1996.10472488 . ПМИД 8895112 .
- ^ Литтл, Т. (1993). «Неправильное использование и легенда о феномене «лизания жабы»». Международный журнал зависимостей . 28 (6): 521–538. дои : 10.3109/10826089309039645 . PMID 8486435 .
- ^ Перейти обратно: а б Майерс, Чарльз В.; Дейли, Джон В.; Малкин, Борис (1978). «Опасно токсичная новая лягушка ( Phyllobates ), используемая индейцами Эмбера в западной Колумбии, с обсуждением изготовления духового оружия и отравления дротиками». Бюллетень Американского музея естественной истории . 161 : 307–366. hdl : 2246/1286 .
- ^ Мэттисон, Крис (2011). Лягушки и жабы миров . Издательство Принстонского университета. стр. 91–92. ISBN 978-0-691-14968-4 .
- ^ Сани, Шарлотта (2012). Лягушка . Книги реакции. стр. 40–42. ISBN 978-1-86189-920-0 .
- ^ Сани, Шарлотта (2012). Лягушка . Книги реакции. стр. 167–175. ISBN 978-1-86189-920-0 .
Дальнейшее чтение
- Бельц, Эллин (2005). Лягушки: внутри их удивительного мира . Книги Светлячка. ISBN 978-1-55297-869-6 .
- Коггер, Х.Г.; Цвайфель, Р.Г.; Киршнер, Д. (2004). Энциклопедия рептилий и земноводных (2-е изд.). Пресса Туманного города. ISBN 978-1-877019-69-2 .
- Эстес Р. и О.А. Рейг. (1973). «Ранние окаменелости лягушек: обзор доказательств». стр. 11–63. В книге Дж. Л. Виала (ред.), « Эволюционная биология бесхвостых животных: современные исследования основных проблем» . Университет Миссури Пресс, Колумбия.
- Гисси, Кармела; Сан-Мауро, Диего; Пезоле, Грациано; Зардоя, Рафаэль (февраль 2006 г.). «Митохондриальная филогения Anura (амфибия): пример конгруэнтной филогенетической реконструкции с использованием аминокислотных и нуклеотидных признаков». Джин . 366 (2): 228–237. дои : 10.1016/j.gene.2005.07.034 . ПМИД 16307849 .
- Холман, Дж. А. (2004). Ископаемые лягушки и жабы Северной Америки . Издательство Университета Индианы. ISBN 978-0-253-34280-5 .
- Сан-Мауро, Диего; Винс, Майкл; Альковендас, ВМС; Зардоя, Рафаэль; Мейер, Аксель (май 2005 г.). «Первоначальное разнообразие современных земноводных предшествовало распаду Пангеи» . Американский натуралист . 165 (5): 590–599. дои : 10.1086/429523 . ПМИД 15795855 . S2CID 17021360 .
- Тайлер, MJ (1994). Австралийские лягушки: естественная история . Рид Книги. ISBN 978-0-7301-0468-1 .
Внешние ссылки
- АмфибияВеб
- Галерея лягушек - фотографии и изображения различных видов лягушек.
- Проект Whole Frog - Виртуальное вскрытие и анатомия лягушки
- «Исчезновение жаб и лягушек беспокоит некоторых ученых» San Francisco Chronicle , 20 апреля 1992 г.
- Фотогалерея амфибий по научному названию : много необычных лягушек.
- Scientific American : Исследователи выявили источник смертельной защиты ядовитых лягушек
СМИ
- Покадровое видео, показывающее развитие яйца до вылупления.
- Вокализации лягушек со всего мира – Из звукового архива Британской библиотеки.
- Звонки лягушки – из Манитобы , Канада.




Сохранившиеся семейства бесхвостых по подотряду |
|---|
| Базы данных органов управления : Национальные |
|---|