Губка
| Губки | |
|---|---|

| |
| Губка плиты | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Порифера Грант , 1836 |
| Классы | |
| Синонимы | |
Губки или морские губки являются членами Metazoan Phylum porifera [ 3 ] ( / p IF ɪ f для p мужа Ученый ˌ ɔː ; - / por- - значение « носитель пор»), [ 4 ] базальная животных клада и таксон диплобластов . сестринский [ 5 ] Они представляют собой в сидячие подачи фильтры , которые связаны с морским дном , и являются одним из самых древних членов Macrobenthos , причем многие исторические виды являются важными рифовыми организмами.
Губки представляют собой многоклеточные организмы, состоящие из желе, похожих на желе, зажатые между двумя тонкими слоями клеток , и обычно имеют трубчатые тела, полные пор и каналов, которые позволяют воде циркулировать через них. Они имеют непециализированные клетки, которые могут трансформироваться в другие типы , и которые часто мигрируют между основными клеточными слоями и мезогилом в процессе. У них нет сложных нервных , [ 6 ] пищеварительные или кровообратные системы . Вместо этого больше всего полагается на поддержание постоянного потока воды через их тела для получения пищи и кислорода и удаления отходов, обычно с помощью движений жгутиков так называемых « воротничков ».
Считается, что сегодня некоторые из самых базальных животных живы, губки, возможно, были первой внешней группой , которая разветвляется эволюционным деревом от последнего общего предка всех животных , [ 5 ] с ископаемыми доказательствами примитивных губок, таких как Отавия, в течение всего тонианского периода (около 800 млн. Домов ). Ветвь зоологии , которая изучает губки, известна как спонгиология . [ 7 ]
Этимология
Термин Губка происходит от древнегреческого слова σπόγγος Spóngos . [ 8 ] Научное название Porifera -это среднее множественное число современного латинского термина Porifer , который поступает из корней , означающих «пор, открытие» и -предложить «подшипник или перенос».
Обзор

Губки похожи на других животных в том смысле, что они являются многоклеточными , гетеротрофными , не имеют клеточных стенок и продуцируют сперматозоиды . В отличие от других животных, им не хватает настоящих тканей [ 9 ] и органы . [ 10 ] Некоторые из них радиально симметричны, но большинство из них асимметричны. Формы их тел адаптированы для максимальной эффективности потока воды через центральную полость, где вода откладывает питательные вещества, а затем уходит через отверстие, называемое оскальмом . Одноклеточные губчатых , хоанофлагелляты напоминают клетки хоаноцитов которые используются для управления их системами потока воды и захвата большей части их пищи. Это наряду с филогенетическими исследованиями рибосомных молекул использовались в качестве морфологических данных, чтобы предположить, что губки являются родственной группой для остальных животных. [ 11 ] Подавляющим большинством являются морские (соленую воду) виды, начиная с среды обитания от приливных зон до глубины, превышающих 8 800 м (5,5 миль), хотя есть пресноводные виды. Все взрослые губки являются сидячими , что означает, что они прикрепляются к подводной поверхности и остаются фиксированными на месте (то есть не путешествуют). В то время как на их личиночной стадии жизни они подвижные .
Многие губки имеют внутренние скелеты спикул (скелетные фрагменты карбоната кальция или диоксида кремния ) и/или Spongin (модифицированный тип белка коллагена). [ 9 ] Внутренняя желатиновая матрица, называемая мезохильной функцией в качестве эндоскелета , и это единственный скелет в мягких губках, который инкрустирует такие твердые поверхности, как камни. Чаще всего мезохил укрепляется минеральными спикулами , с помощью волокон или обоих. 90% всех известных видов губки, которые имеют самый широкий ассортимент среды обитания, в том числе все пресноводные, - это демоспоны , которые используют Spongin; Многие виды имеют спикулы кремнезема , тогда как у некоторых видов есть экзоскелеты карбоната кальция . Кальциальные губки имеют спикулы карбоната кальция, и, в некоторых видах экзоскелеты карбоната кальция, ограничены относительно мелкими морскими водами, где производство карбоната кальция является самым простым. [ 12 ] : 179 Хрупкие стеклянные губки , с « лесами » спикул кремнезема, ограничены полярными областями и глубиной океана, где хищники редки. Окаменелости всех этих типов были обнаружены в скалах от 580 миллионов лет назад . Кроме того, археоциатиды , ископаемые которых распространены в скалах с 530 до 490 миллионов лет назад , теперь рассматриваются как тип губки.
Хотя большинство из примерно 5000–10 000 известных видов губок питаются бактериями и другими микроскопическими пищами в воде, некоторые из них принимают фотосинтезирующие микроорганизмы в качестве эндосимбионтов , и эти союзы часто производят больше пищи и кислорода, чем они потребляют. Несколько видов губок, которые живут в бедных пищевых условиях, развивались как плотоядные , которые охотятся в основном на мелких ракообразных . [ 13 ]
Большинство губок размножаются сексуально , но они также могут воспроизводить бесполезно. Сексуально воспроизводя виды высвобождают сперматозоиды в воду, чтобы оплодотворить OVA, высвобождаемый или сохраняемый его партнером или «матери»; Уплотненные яйца превращаются в личинки , которые уплывают в поисках мест для поселения. [ 12 ] : 183–185 Губки известны для восстановления от разбитых фрагментов, хотя это работает только в том случае, если фрагменты включают правильные типы клеток. Некоторые виды размножаются по почте. Когда условия окружающей среды становятся менее гостеприимными для губок, например, по мере того, как температура падает, многие пресноводные виды и несколько морских, производят геммулы , «стручки выживания» неспециализированных клеток, которые остаются бездействующими до тех пор, пока условия не улучшатся; Затем они либо образуют совершенно новые губки, либо реколонизируют скелеты своих родителей. [ 12 ] : 120–127

Несколько видов Demosponge, которые имеют совершенно мягкие волокнистые скелеты без жестких элементов, были использованы людьми в течение тысячелетий в течение нескольких лет для нескольких целей, в том числе в качестве прокладки и в качестве инструментов для очистки. Однако к 1950-м годам они были настолько значительно переполнены , что отрасль почти рухнула, и большинство материалов, похожих на губку, теперь синтетические. Губки и их микроскопические эндосимбионты в настоящее время исследуются в качестве возможных источников лекарств для лечения широкого спектра заболеваний. Дельфины наблюдались с использованием губок в качестве инструментов во время питания . [ 15 ]
Отличительные черты
Губки представляют собой филовую Porifera и были определены как сидячие метазои (многоклеточные неподвижные животные), у которых есть потребление воды и выходные отверстия, соединенные камерами, выстлаемыми хоаноцитами , клетками с хриплыми жгутиками. [ 12 ] : 29 Тем не менее, несколько плотоядных губок потеряли эти системы потока воды и хоаноциты. [ 12 ] : 39 [ 16 ] Все известные живые губки могут переделать свои тела, так как большинство типов их клеток могут перемещаться в их тела, а некоторые могут перейти от одного типа к другому. [ 16 ] [ 17 ]
Даже если несколько губок способны продуцировать слизь, которая действует как микробный барьер у всех других животных, - не было зарегистрировано губки с способностью секретировать функциональный слой слизи. Без такого слоя слизи их живая ткань покрывается слоем микробных симбионтов, который может содержать до 40–50% от губкой влажной массы. Эта неспособность предотвратить проникновение микробов их пористой ткани может быть основной причиной, по которой они никогда не развивали более сложную анатомию. [ 18 ]
Подобно Cnidarians (медуза и т. Д.) И Ctenophores (Check Jellies), и, в отличие от всех других известных метазой, тела губков состоят из неживущей желе, похожей на желе ( мезохил ), зажатых между двумя основными слоями клеток. [ 19 ] [ 20 ] Cnidarians и Ctenophores имеют простые нервные системы, а их клеточные слои связаны внутренними связями и установлены на базальной мембране (тонкий волокнистый коврик, также известный как « базальная пластинка »). [ 20 ] Губки не имеют нервной системы, похожей на систему позвоночных, но могут иметь такую, которая сильно отличается. [ 6 ] Их средние желе-подобные слои имеют большие и разнообразные популяции клеток, и некоторые типы клеток во внешних слоях могут перемещаться в средний слой и изменить их функции. [ 17 ]
| Губки [ 17 ] [ 19 ] | Cnidarians и Ctenophores [ 20 ] | |
|---|---|---|
| Нервная система | Нет/да | Да, просто |
| Ячейки в каждом слое связаны вместе | Нет, за исключением того, что гомосклероморфы имеют подвальные мембраны. [ 21 ] | Да: межклеточные соединения; подвальные мембраны |
| Количество клеток в среднем слое "желе" | Много | Немного |
| Ячейки в внешних слоях могут перемещаться внутрь и изменять функции | Да | Нет |
Базовая структура
Типы клеток
Тело Губки имеет пустое и в форме удерживается мезохилом , вещество, похожее на желе, изготовленное в основном из коллагена и подкрепленное плотной сетью волокон, также сделано из коллагена. 18 различных типов клеток были идентифицированы. [ 23 ] Внутренняя поверхность покрыта хоаноцитами , клетками с цилиндрическими или коническими воротниками, окружающими один жгутичок на хоаноцит. Волноподобное движение блаженого жгутиков ведет воду через тело губки. Все губки имеют остию , каналы, ведущие к внутренней части через мезохил, и в большинстве губок они контролируются с трубчатыми пороцитами , которые образуют близкие впускные клапаны. Пинакоциты , пластинчатые клетки, образуют однослойную внешнюю кожу по всем остальным частям мезогила, которые не покрыты хоаноцитами, а пинкакоциты также переваривают частицы пищи, которые слишком велики, чтобы войти в остию, [ 17 ] [ 19 ] в то время как те, кто у основания животного несет ответственность за его привязку. [ 19 ]
Другие типы клеток живут и перемещаются в мезогиле: [ 17 ] [ 19 ]
- Лофоциты представляют собой амебы , похожие на клетки, которые медленно перемещаются через мезохил и секретные коллагеновые волокна.
- CollenceTes -это еще один тип коллагеновой ячейки.
- Рабдисские клетки секретируют полисахариды , которые также образуют часть мезохила.
- Ооциты и сперматоциты являются репродуктивными клетками.
- Склероциты секретируют минерализованные спикулы («маленькие шипы»), которые образуют скелеты многих губков и в некоторых видах обеспечивают некоторую защиту от хищников.
- В дополнение к или вместо склероцитов, у демоспон, есть спонгоциты , которые выделяют форму коллагена, которая полимеризуется в Spongin , толстый волокнистый материал, который жестко жесткости мезогил.
- Миоциты («мышечные клетки») проводят сигналы и заставляют сокращаться части животного.
- «Серые клетки» действуют как эквивалент губков иммунной системы .
- Археоциты (или амебоциты ) представляют собой амебы, похожие на клетки, которые являются тотипентными , другими словами, каждый способен преобразовать в любой другой тип клетки. Они также играют важную роль в кормлении и в очистке мусора, которые блокируют остию.
без нейронов Многие личиночные губки обладают глазами , которые основаны на криптохромах . Они опосредуют фототаксическое поведение. [ 24 ]
Стеклянные губки представляют собой отличительную вариацию этого основного плана. Их спикулы, изготовленные из кремнезема , образуют каркас, похожие на каркас , между стержнями, живая ткань подвешена как паутина , которая содержит большинство типов клеток. [ 17 ] Эта ткань представляет собой синцитик , который в некотором роде ведет себя как многие клетки, в которых есть одна внешняя мембрана , а в других, таких как одна клетка с несколькими ядрами .
Поток воды и структуры тела

Большинство губки работают довольно как дымоходы : они берут воду внизу и выбросят ее из оскальма наверху. Поскольку окружающие течения быстрее наверху, эффект всасывания, который они производят по принципу Бернулли, выполняет некоторые работы бесплатно. Губки могут контролировать поток воды различными комбинациями, полностью или частично закрывающими оскальм и остию (поры впускного завещания) и изменяя удар жгутиков, и могут закрыть его, если в воде много песка или ила. [ 17 ]
Хотя слои пинакоцитов и хоаноцитов напоминают эпителию более сложных животных, они не связаны тесно связаны с клеточными соединениями или базальной пластинкой (тонкий волокнистый лист внизу). Гибкость этих слоев и повторное моделирование мезогила лофоцитами позволяют животным регулировать свои формы на протяжении всей своей жизни, чтобы максимально использовать местные водные течения. [ 17 ] : 83
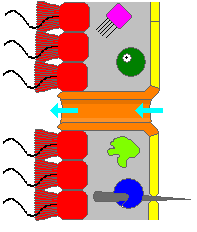
Самая простая структура тела в губках - это форма трубки или вазы, известная как «Asconoid», но это сильно ограничивает размер животного. Структура тела характеризуется стеблем, похожим на губокосолю, окруженное одним слоем хоаноцитов. Если он просто масштабируется, соотношение его объема к площади поверхности увеличивается, поскольку поверхность увеличивается как квадрат длины или ширины, в то время как объем увеличивается пропорционально кубу. Количество ткани, которая нуждается в пище и кислороде, определяется объемом, но насосная способность, которая поставляет пищу и кислород, зависит от области, покрытой хоаноцитами. Асконоидные губки редко превышают 1 мм (0,039 дюймов) в диаметре. [ 17 ]
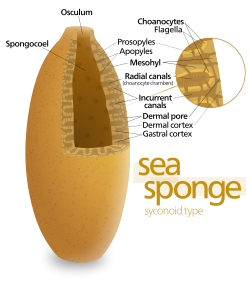
Некоторые губки преодолевают это ограничение, приняв структуру «сионоида», в которой стенка тела плиты . Внутренние карманы складки выстланы хоаноцитами, которые соединяются с внешними карманами складки Ostia. Это увеличение количества хоаноцитов и, следовательно, в насосной способности позволяет синоидным губкам расти до нескольких сантиметров в диаметре.
Паттерн «лейконоида» повышает пропускную способность, заполняя внутреннюю часть почти полностью мезохилом, который содержит сеть камер, выстланную хоаноцитами и подключенные друг к другу, а также в потреблении воды и выходом труб. Лейконидные губки растут до 1 м (3,3 фута) в диаметре, и тот факт, что рост в любом направлении увеличивает число камер хоаноцитов, позволяет им принимать более широкий диапазон форм, «инкрустирующих» губ поверхности, к которым они прикрепляют. Все пресноводные и большинство мелководных морских губок имеют лейконидные тела. Сети водных проходов в стеклянных губках аналогичны структуре лейконида. [ 17 ]
Во всех трех типах структуры площадь поперечного сечения областей, покрытых хоаноцитами, намного больше, чем у впускных и выходных каналов. Это делает поток медленнее вблизи хоаноцитов и, таким образом, облегчает их улавливать частицы пищи. [ 17 ] Например, у лейконии небольшая лейконоидная губка около 10 сантиметров (3,9 дюйма) высотой и 1 сантиметр (0,39 дюйма) в диаметре, вода входит в каждый из более 80 000 впускных каналов со скоростью 6 см в минуту . Однако, поскольку Leuconia имеет более 2 миллионов жгутиковых камер, комбинированный диаметр, намного больше, чем у каналов, вода по течению через камеры замедляется до 3,6 см в час , что облегчает ухаживание хоаноцитов. Вся вода исключена через один ослабление примерно на 8,5 см в секунду , достаточно быстрая, чтобы перевозить отходы на расстоянии. [ 26 ]
- Мезохил
- Спикулы
- Морское дно / рок
- Поток воды
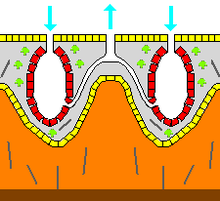
Скелет
В зоологии скелет является какой -либо довольно жесткой структурой животного, независимо от того, есть ли у него суставы и независимо от того, является ли оно биоминерализованным . Мезохильный функционирует как эндоскелет в большинстве губок и является единственным скелетом в мягких губках, который инкрустает твердые поверхности, такие как камни. Чаще мезохил укрепляется минеральными спикулами , с помощью сконсинских волокон или обоих. Спикулы, которые присутствуют в большинстве, но не во всех видах, [ 27 ] может быть изготовлен из кремнезема или карбоната кальция и варьироваться по форме от простых стержней до трехмерных «звезд» с шестью лучей. Спикулы продуцируются клетками склероцитов , [ 17 ] и может быть отдельно, подключен к суставам или слитым. [ 16 ]
Некоторые губки также секретируют экзоскелеты , которые полностью лежат вне их органических компонентов. Например, Sclerosponges («твердые губки») имеют массивные экзоскелеты карбоната кальция, которыми органическое вещество образует тонкий слой с камерами хоаноцита в ямах в минерале. Эти экзоскелеты секретируются пинкацитами , которые образуют шкуры животных. [ 17 ]
Жизненно важные функции

Движение
Хотя взрослые губки являются принципиально сидячими животными, некоторые морские и пресноводные виды могут перемещаться по морскому груду со скоростью 1–4 мм (0,039–0,157 дюйма в день) в результате амебы, подобных движениям пинакоцитов и других клеток. Несколько видов могут сократить все свои тела, и многие могут закрыть свой оска и остию . Младшие дрифты или свободно плавают, а взрослые стационарны. [ 17 ]
Дыхание, кормление и экскреция

Губки не имеют различных систем кровообращения , респираторных , пищеварительных и экскреторных систем - вместо этого система потока воды поддерживает все эти функции. Они фильтруют частицы пищи из воды, протекающей через них. Частицы, более 50 микрометров, не могут войти в остию , а пинакоциты потребляют их путем фагоцитоза (поглощение и внутриклеточное пищеварение). Частицы от 0,5 мкм до 50 мкм попадают в остию, которая сужается от внешних до внутренних концов. Эти частицы потребляются пинкацитами или археоцитами , которые частично выдают себя через стены остии. Частицы размером с бактерии, ниже 0,5 микрометров, проходят через остию и пойманы и потребляются хоаноцитами . [ 17 ] Поскольку самые маленькие частицы являются наиболее распространенными, хоаноциты обычно захватывают 80% поставки продовольствия губки. [ 28 ] Археоциты транспортируют пищу, упакованные в пузырьки из клеток, которые непосредственно переваривают пищу в отношении тех, которые нет. По крайней мере, один вид губки имеет внутренние волокна, которые функционируют как следы для использования с помощью археоцитов, несущих питательные вещества, археоциты, [ 17 ] И эти треки также перемещают инертные объекты. [ 19 ]
Раньше утверждалось, что стеклянные губки могут жить на питательных веществах, растворенных в морской воде, и были очень склонны к илу. [ 29 ] Тем не менее, исследование в 2007 году не обнаружило никаких доказательств этого и пришло к выводу, что они очень эффективно извлекают бактерии и другие микроорганизмы из воды (около 79%) и обрабатывают сусплентные зерна осадка для извлечения такой добычи. [ 30 ] Воротнички переваривают пищу и распределяют ее обернуты в везикулах, которые транспортируются молекулами Dynein «двигатель» вдоль пучков микротрубочек , которые работают по всему синциция . [ 17 ]
Клетки губков поглощают кислород путем диффузии из воды в клетки, когда вода течет через организм, в которые также диффундирует углекислый газ и другие растворимые отходы, такие как аммиак . Археоциты удаляют минеральные частицы, которые угрожают блокировать остию, переносить их через мезохил и, как правило, бросают их в исходящий водный ток, хотя некоторые виды включают их в свои скелеты. [ 17 ]
Плотоядные губки

В водах, где поставка частиц пищи очень плохой, некоторые виды добывают ракообразных и других мелких животных. До сих пор было обнаружено только 137 видов. [ 32 ] Speriopsidae это масло - Esperiopsidae. [ 33 ] В большинстве случаев мало что известно о том, как они на самом деле захватывают добычу, хотя, как полагают, некоторые виды используют либо липкие нити, так и зацепленные спикулы . [ 33 ] [ 34 ] Большинство плотоядных губки живут в глубоких водах, до 8 840 м (5,49 миль), [ 35 ] И разработка методов глубокоокеанского разведки, как ожидается, приведет к открытию еще нескольких. [ 17 ] [ 33 ] Тем не менее, один вид был обнаружен в средиземноморских пещерах на глубине 17–23 м (56–75 футов), наряду с более обычными губками для кормления фильтров . Пещерные хищники захватывают ракообразных под 1 мм (0,039 дюйма) длиной, запутывая их тонкими нитями, переваривают их, окутывая их дальнейшими нитями в течение нескольких дней, а затем вернится к их нормальной форме; Нет никаких доказательств того, что они используют яд . [ 35 ]
Наиболее известные плотоядные губки полностью потеряли систему потока воды и хоаноциты . Тем не менее, род Chondrocladia использует высоко измененную систему потока воды для надувания баллонных структур, которые используются для захвата добычи. [ 33 ] [ 36 ]
Эндосимбионты
Пресноводные губки часто содержат зеленые водоросли в качестве эндосимбионтов в археоцитах и других клетках и получают пользу от питательных веществ, продуцируемых водорослями. Многие морские виды принимают другие фотосинтезирующие организмы, чаще всего цианобактерии , но в некоторых случаях динофлагелляции . Симбиотические цианобактерии могут образовывать треть общей массы живой ткани у некоторых губок, а некоторые губки получают от 48 до 80% от этих микроорганизмов. [ 17 ] В 2008 году команда Университета Штутгарта сообщила, что спикулы, сделанные из силикаса, проводят свет в мезохиле , где живут фотосинтезирующие эндосимбионты. [ 37 ] Губки, которые принимают фотосинтезирующие организмы, наиболее распространены в водах с относительно плохими запасами частиц пищи и часто имеют листовые формы, которые максимизируют количество солнечного света, которое они собирают. [ 19 ]
Недавно обнаруженная плотоядная губка, которая живет вблизи гидротермальных вентиляционных отверстий, хозяина , питающихся бактериями, питающимися и переваривает некоторые из них. [ 19 ]
"Иммунная система
Губки не имеют сложных иммунных систем большинства других животных. Тем не менее, они отвергают трансплантаты от других видов, но принимают их от других представителей их собственных видов. У нескольких морских видов серые клетки играют ведущую роль в отказе от иностранного материала. При вторжении они производят химическое вещество, которое останавливает движение других клеток в пораженной области, что предотвращает использование внутренних транспортных систем губки. Если вторжение сохраняется, серые клетки концентрируются в области и высвобождают токсины, которые убивают все клетки в этой области. «Иммунная» система может оставаться в этом активированном состоянии до трех недель. [ 19 ]
Размножение
Асексуал

Губки имеют три бесполых метода воспроизведения: после фрагментации, почтением и путем производства геммулов . Фрагменты губки могут быть отделены токами или волнами. Они используют подвижность своих пинакоцитов и хоаноцитов и изменяют мезохил, чтобы повторно приспособиться к подходящей поверхности, а затем восстановить себя как небольшие, но функциональные губки в течение нескольких дней. Те же самые возможности позволяют губки, которые были прожаты через тонкую ткань для восстановления. [ 17 ] : 239 Фрагмент губки может регенерировать только в том случае, если он содержит оба CollenceTes для получения мезохил и археоцитов для получения всех других типов клеток. [ 28 ] Очень немногие виды размножаются по почте. [ 17 ] : 90–94
Геммулы - это «стручки для выживания», которые несколько морских губок и многие пресноводные виды производят тысячами при смерти, а некоторые, в основном пресноводные виды, регулярно производят осенью. Спонгоциты делают драгоценные камни, обернув раковины скруна, часто усиленные спикулами, круглые кластеры археоцитов , которые полны питательных веществ. [ 17 ] : 87–88 Пресноводные геммулы также могут включать в себя фотосинтезирование симбионтов. [ 38 ] Затем драгоценные камни становятся бездействующими, и в этом состоянии могут выжить в холоде, высыхая, отсутствие кислорода и экстремальные изменения в солености . [ 17 ] Пресноводные геммулы часто не оживляют, пока температура не падает, не останется холодной в течение нескольких месяцев, а затем достигает почти «нормального» уровня. [ 38 ] Когда геммуле прорастает, археоциты вокруг внешней внешней части кластера превращаются в пинакоциты , мембрану над пор в разрывах оболочки, медленно появляется кластер ячеек, а большинство оставших губка. Геммулы из одного и того же вида, но разные люди могут объединить усилия, чтобы сформировать одну губку. [ 17 ] : 89–90 Некоторые геммулы сохраняются в родительской губке, и весной может быть трудно сказать, возродила ли старая губка или «реколонизирована» своими собственными геммулами. [ 38 ]
Сексуальный
Большинство губок - это гермафродиты (функционируют как оба пола одновременно), хотя губки не имеют гонад (репродуктивные органы). Сперматозоиды продуцируются хоаноцитами или целыми камерами хоаноцитов, которые погружаются в мезохил и образуют сперматические кисты , в то время как яйца образуются путем трансформации археоцитов или хоаноцитов у некоторых видов. Каждое яйцо обычно приобретает желток , потребляя «клетки медсестры». Во время нереста сперма вырвалась из их кист и вытесняется через Osculum . Если они связываются с другой губкой одного и того же вида, поток воды несет их в хоаноциты, которые охватывают их, но вместо того, чтобы переваривать их, метаморфоза в амебоидную форму и переносит сперму через мезогил в яйца, что в большинстве случаев охватывает носитель и его груз. [ 17 ] : 77
Несколько видов выпускают оплодотворенные яйца в воду, но большинство сохраняют яйца, пока они не вылупляются. Сохранение яиц, родители могут переносить симбиотические микроорганизмы непосредственно на свое потомство посредством вертикальной передачи , в то время как виды, которые высвобождают свои яйца в воду, должны приобретать симбионты горизонтально (комбинация обоих, вероятно, является наиболее распространенным, где личинки с вертикально передаваемыми симбионтами (комбинация обоих является наиболее распространенным также приобретать другие горизонтально). [ 39 ] [ 40 ] Существует четыре типа личинок, но все это лецитотрофные (нетокобные) шарики клеток с внешним слоем клеток, чьи жгутики или реснички позволяют личинкам двигаться. После плавания в течение нескольких дней личинки опускаются и ползут, пока они не найдут место для поселения. Большинство клеток превращается в археоциты, а затем в типы, подходящие для их местоположений в миниатюрной губке для взрослых. [ 17 ] : 77 [ 41 ]
Эмбрионы стеклянной губки начинаются с деления на отдельные клетки, но как только 32 клетки сформировались, они быстро превращаются в личинки, которые внешне яйцевидны с полосой ресничек вокруг середины, которую они используют для движения, но внутренне имеют типичную стеклянную губку Основная паутинка, похожая на основной синхтимиум, забита вокруг и между ними, и choanosyncytia с несколькими воротничками в центре. Затем личинки оставляют тела своих родителей. [ 42 ]
Мейоз
Цитологическое прогрессирование оогенеза и сперматогенеза Porifera ( гаметогенез ) очень похоже на развитие других метазоа. [ 43 ] Большинство генов из классического набора мейотических генов, в том числе генов для рекомбинации ДНК и репарации разрыва с двумя целями, которые сохраняются у эукариот , экспрессируются в губках (например, Geodia Hentscheli и Geodia Phlegraei ). [ 43 ] Поскольку Porifera считаются самыми ранними расходящимися животными, эти результаты показывают, что основной инструментарий мейоза, включая возможности для рекомбинации и восстановления ДНК, присутствовали на ранних стадиях эволюции эукариота. [ 43 ]
Жизненный цикл
Губки в умеренных регионах живут не более нескольких лет, но некоторые тропические виды и, возможно, некоторые глубокоокеанские могут жить в течение 200 или более лет. Некоторые кальцифицированные демоспонги растут всего на 0,2 мм (0,0079 дюйма) в год, и, если эта скорость постоянна, образцы шириной 1 м (3,3 фута) должны быть около 5000 лет. Некоторые губки начинают сексуальное размножение, когда ему было всего несколько недель, в то время как другие ждут, пока им не исполнится несколько лет. [ 17 ]
Координация деятельности
У взрослых губок не хватает нейронов или любого другого вида нервной ткани . Тем не менее, большинство видов обладают способностью выполнять движения, которые координируются по всему их телу, в основном сокращения пинакоцитов , сжимая водяные каналы и, таким образом, изгнать избыточные осадка и другие вещества, которые могут вызывать блокировки. Некоторые виды могут сократить осмеревание независимо от остальной части тела. Губки могут также сокращаться, чтобы уменьшить область, которая уязвима для атаки хищников. В тех случаях, когда две губки сливаются, например, если есть большой, но все еще неразвитый почек, эти волны сокращения постепенно координируются в обоих « сиамских близнецах ». Координирующий механизм неизвестен, но может включать химические вещества, похожие на нейротрансмиттеры . [ 44 ] Тем не менее, стеклянные губки быстро передают электрические импульсы через все части синцитика и используют их, чтобы остановить движение их жгутиков, если входящая вода содержит токсины или чрезмерный осадок. [ 17 ] Считается, что миоциты несут ответственность за закрытие оскальма и за передачу сигналов между различными частями тела. [ 19 ]
Губки содержат гены, очень похожие на те, которые содержат «рецепт» для пост -синаптической плотности, важной структуры, связанной с сигналом в нейронах всех других животных. Однако у губок эти гены активируются только в «клетках колбы», которые появляются только у личинок и могут обеспечить некоторую сенсорную способность, в то время как личинки плавают. Это поднимает вопросы о том, представляют ли клетки колбы предшественники истинных нейронов или свидетельствуют о том, что предки губков имели настоящие нейроны, но они потеряли их, когда они адаптировались к сидяче. [ 45 ]
Экология
Среда обитания

Губки во всем мире находятся в своем распределении, живут в широком спектре мест обитания океана, от полярных регионов до тропиков. [ 28 ] Большинство живут в тихих, чистых водах, потому что осадок, разжигаемые волнами или течениями, блокируют свои поры, что затрудняет кормить и дышать. [ 29 ] Наибольшее количество губок обычно встречается на твердых поверхностях, таких как камни, но некоторые губки могут прикрепить себя к мягкому осадкам с помощью корнеподобного основания. [ 46 ]
Губки более распространены, но менее разнообразны в умеренных водах, чем в тропических водах, возможно, потому, что организмы, которые добывают губки, более распространены в тропических водах. [ 47 ] Стеклянные губки являются наиболее распространенными в полярных водах и в глубине умеренного и тропического моря, поскольку их очень пористая конструкция позволяет им извлекать пищу из этих бедных ресурсов с минимальными усилиями. Демоспожи и известковые губки в изобилии и разнообразны в мелких неполярных водах. [ 48 ]
Различные классы губки живут в разных диапазонах среды обитания:
Сорт Тип воды [ 19 ] Глубина [ 19 ] Тип поверхности [ 19 ] Calcarea Морской пехотинец менее 100 м (330 футов) Жесткий Стеклянные губки Морской пехотинец Глубокий Мягкий или твердый осадок Демоспоны Морской пехотинец, солоноватый; и около 150 пресноводных видов [ 17 ] Взаимодействие в абиссаль; [ 19 ] Плотноядающая демонстрация была найдена на уровне 8 840 м (5,49 миль) [ 35 ] Любой
Как основные производители
Губки с фотосинтезирующими эндосимбионтами производят в три раза больше кислорода , чем они потребляют, а также больше органического вещества, чем они потребляют. Такой вклад в ресурсы их среды обитания значительны по сравнению с большим барьерным рифом Австралии , но относительно незначительным в Карибском бассейне. [ 28 ]
Защита


Многие губки проливают спикулы , образуя плотный ковер глубиной на несколько метров, которые удерживают эхинодермы , которые в противном случае охотились бы на губки. [ 28 ] Они также производят токсины, которые предотвращают другие сидячие организмы, такие как мжзильные или морские брызги , от роста или рядом с ним, что делает губ очень эффективными конкурентами для жилой площади. Один из многих примеров включает в себя агелиферин .
Несколько видов, карибская пожарная губка Тедания Игнис , вызывает серьезную сыпь у людей, которые справляются с ними. [ 17 ] Черепахи и некоторые рыбы питаются в основном губками. Часто говорят, что губки производят химическую защиту от таких хищников. [ 17 ] Тем не менее, эксперименты не смогли установить связь между токсичностью химических веществ, производимых губками, и тем, как они на вкус с рыбой, что уменьшит полезность химической защиты в качестве сдерживающих факторов. Хищничество от рыбы может даже помочь распространять губки путем отсоединения фрагментов. [ 19 ] Тем не менее, некоторые исследования показали, что рыба показывает предпочтение не химически защищенным губкам, [ 49 ] И другое исследование показало, что высокий уровень хищничества кораллов действительно предсказывал наличие химически защищенных видов. [ 50 ]
Стеклянные губки не дают токсичных химических веществ и живут в очень глубокой воде, где хищники редки. [ 29 ]
Хищничество
Губцеффи, также известные как Spongillaflies ( Neuroptera , Sisyridae ), являются специализированными хищниками пресноводных губ. Самка откладывает яйца на растительность, нависающую с водой. Личинки вылупляются и падают в воду, где они ищут губки, чтобы питаться. Они используют свои удлиненные рта, чтобы прорвать губку и сосать жидкости внутри. Личинки некоторых видов цепляются за поверхность губки, в то время как другие укрываются во внутренних полостях губки. Полностью выращенные личинки покидают воду и вращают кокон, в котором можно окунуться. [ 51 ]
Биоврозия
Карибская куриная губчатая губка хондриллы гадрилла выделяет токсины, которые убивают коралловые полипы , что позволяет губкам расти над коралловыми скелетами. [ 17 ] Другие, особенно в семействе Clionaidae , используют коррозионные вещества, секретируемые их археоцитами в туннель в скалы, кораллы и раковины мертвых моллюсков . [ 17 ] Губки могут удалять с рифов до 1 м (3,3 фута) в год, создавая видимые выемки чуть ниже уровня низкого прикрытия. [ 28 ]
Болезни
Карибские губки рода Aplysina страдают от синдрома красной полосы Aplysina . Это заставляет Aplysina разрабатывать одну или несколько ржавых полос, иногда с соседними полосами некротической ткани. Эти поражения могут полностью окружать ветви губки. Болезнь, по -видимому, является заразной и воздействует примерно 10 процентов A. cauliformis на багамских рифах. [ 52 ] Ржавованные полосы вызваны цианобактерией , но неизвестно, действительно ли этот организм вызывает заболевание. [ 52 ] [ 53 ]
Сотрудничество с другими организмами
В дополнение к размещению фотосинтезирующих эндосимбионтов, [ 17 ] Губки известны своим широким спектром сотрудничества с другими организмами. Относительно большая инкрустирующая губка lissodendoryx colombiensis наиболее распространена на скалистых поверхностях, но расширяет свой диапазон на луга морских изделий , позволив себе быть окруженным или заросшим губками морских траекторов, которые неприятны для местных звездочек и, следовательно, защищают лиссодендорис от них; В свою очередь, губки морской травы занимают более высокие позиции вдали от отложения морского этажа. [ 54 ]
Шревки рода Synalpheus образуют колонии в губках, и каждый виды креветок обитает в разных видах губки, что делает Synalpheus одним из самых разнообразных родов ракообразных . В частности, Synalpheus Regalis использует губку не только в качестве источника пищи, но и в качестве защиты от других креветок и хищников. [ 55 ] Целых 16 000 человек населяют одну губку Loggerhead , питаясь более крупными частицами, которые собираются на губке, когда он фильтрует океан, чтобы питаться. [ 56 ] Другие ракообразные, такие как крабы отшельника, обычно имеют специфические виды губки, псевдоспонгосоритов , растущие на них, поскольку губчатая и краба занимают гастроподные раковины до тех пор, пока краб и губча оболочка, пока краб не найдет подходящую замену. [ 57 ]

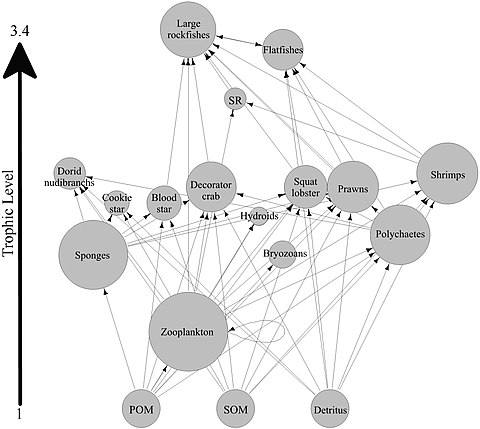
Петля губки
Большинство губков - это детритоядные , которые фильтровали органические частицы мусора и микроскопические формы жизни из океанской воды. В частности, губки занимают важную роль в качестве детритоя в пищевых сетях коралловых рифов путем переработки детрита на более высокие трофические уровни . [ 60 ]
Была выдвинута гипотеза, что губки коралловых рифов облегчают передачу органического вещества, полученного из кораллов, в связанные с ними детритивенные посредством производства губчатого детрита, как показано на диаграмме. Несколько видов губки способны преобразовать коралловые домики в губок детрит, [ 61 ] [ 62 ] и передача органического вещества, производимого кораллами, в дальнейшем в области пищевой сети. Кораллы высвобождают органическое вещество как растворенное и слизи [ 63 ] [ 64 ] [ 65 ] [ 66 ] а также клеточный материал, такой как экстененный симбиодиний . [ 67 ] [ 68 ] [ 60 ]
Органическое вещество может быть перенесено из кораллов в губки всеми этими путями, но DOM, вероятно, составляет наибольшую долю, поскольку большинство (от 56 до 80%) слизь кораллов растворяется в толще воды, [ 64 ] и потеря кораллов фиксированного углерода из -за изгнания симбиодиния обычно незначительна (0,01%) [ 67 ] по сравнению с высвобождением слизи (до ~ 40%). [ 69 ] [ 70 ] Органическое вещество, полученное из кораллов, также может быть косвенно перенесено на губки через бактерии, которые также могут потреблять слизь кораллов. [ 71 ] [ 72 ] [ 73 ] [ 60 ]
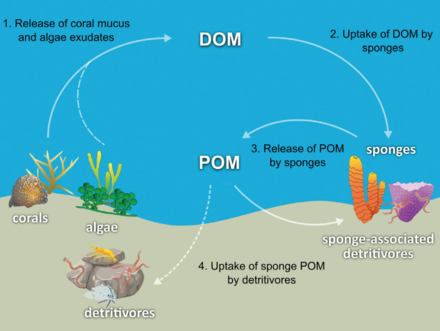

Губка Холобонт
Помимо одно -симбиотического отношения , хозяин может стать симбиотиком с микробным консорциумом , что приведет к разнообразному микробиому губки . Губки способны размещать широкий спектр микробных сообществ , которые также могут быть очень специфичными. Микробные сообщества, которые формируют симбиотическую связь с губкой, могут составлять до 35% биомассы его хозяина. [ 76 ] Термин для этой конкретной симбиотической взаимосвязи, где микробные консорциумы сочетаются с хозяином, называется голобиотическими отношениями . Губка, а также микробное сообщество, связанное с ней, будут производить большой диапазон вторичных метаболитов , которые помогают защитить ее от химинмов, таких как химическая защита . [ 77 ]
Некоторые из этих отношений включают эндосимбионты в клетках бактериоцитов, а также цианобактерии или микроводоросли, обнаруженные под слоем клеток пинакодермы, где они способны получать наибольшее количество света, используемого для фототрофии. Они могут размещать более 50 различных микробных типов и кандидатов, в том числе альфапротоэбактерии, актиномицетота , хлорфлексоты , нитроспироты , « цианобактерий », гамма-таксонов, кандидата Phylum poribacteria и Thaumarchaea . [ 77 ]
Систематика
Таксономия
Карл Линнеус , который классифицировал большинство видов сидячих животных как принадлежащих к Ордену Зофита в классовых вермах , ошибочно идентифицировал род Spongia как растения в Ордене водоросли . [ 78 ] [ необходимо дальнейшее объяснение ] В течение долгого времени после этого губки были назначены для подчинения Parazoa («рядом с животными»), отделенными от эйметазоа который сформировал остальную часть царства , . [ 79 ] Они считались парафилетическим филомом , из которого эволюционировали высшие животные. [ 80 ] Другие исследования показывают, что Porifera является монофилетическим. [ 81 ]
Phylum porifera далее делится на классы в основном в зависимости от состава их скелетов : [ 16 ] [ 28 ]
- Hexactinellida (стеклянные губки) имеют силикатные спикулы, самые большие из которых имеют шесть лучей и могут быть индивидуальными или слитыми. [ 16 ] Основными компонентами их тел являются синцития , в которой большое количество клеток совместно использует одну внешнюю мембрану . [ 28 ]
- У калькалея есть скелеты из кальцита , форма карбоната кальция , которая может образовывать отдельные спикулы или большие массы. Все клетки имеют единое ядро и мембрану. [ 28 ]
- Большинство Demospongiae имеют силикатные спикулы или губки -волокна или обоих в своих мягких тканях. Тем не менее, некоторые также имеют массивные внешние скелеты из арагонита , еще одну форму карбоната кальция. [ 16 ] [ 28 ] Все клетки имеют единое ядро и мембрану. [ 28 ]
- Археоцийта известны только как окаменелости из кембрийского периода. [ 79 ]
В 1970 -х годах губки с массивными карбонатными скелетами кальция были назначены в отдельный класс Sclerospongiae , иначе известный как «кораллические губки». [ 82 ] Однако в 1980 -х годах было обнаружено, что все они были членами каллеи или Demospongiae. [ 83 ]
До сих пор научные публикации определили около 9 000 видов Poriferan, [ 28 ] из которых: около 400 - стеклянные губки; Около 500 являются известными видами; А остальные - демоспон. [ 17 ] Тем не менее, некоторые виды среды обитания, вертикальные камни и стены пещеры и галереи в рок и коралловых валунах были исследованы очень мало, даже в мелких морях. [ 28 ]
Классы
Губки традиционно распространялись в трех классах: известковые губки (калькарея), стеклянные губки (гексактинеллида) и демоспольс (demospongiae). Тем не менее, исследования показали, что гомосклероморфа , группа, которая, как считается, принадлежит Demospongiae , имеет генетическую связь , хорошо отделенную от других классов губки. [ 12 ] : 153–154 Поэтому они недавно были признаны четвертым классом губ. [ 84 ] [ 85 ]
Губки делятся на классы в основном в зависимости от состава их скелетов : [ 19 ] Они расположены в эволюционном порядке, как показано ниже в порядке возрастания их эволюции сверху вниз:
Сорт Тип ячеек [ 19 ] Спикулы [ 19 ] Спунские волокна [ 19 ] Массивный экзоскелет [ 28 ] Форма тела [ 19 ] Hexactinellida В основном синцития у всех видов Кремнезый
Может быть индивидуальным или сплавнымНикогда Никогда Лейконоид Demospongiae Одиночное ядро, единственная внешняя мембрана Кремнезый Во многих видах У некоторых видов.
Сделано из арагонита, если присутствует. [ 16 ] [ 28 ]Лейконоид Calcarea Одиночное ядро, единственная внешняя мембрана Кальцит
Может быть индивидуальные или большие массыНикогда Общий.
Сделано из кальцита, если присутствует.Asconoid, Synoid, Leuconoid или соленоид [ 86 ] Гомосклероморфа Одиночное ядро, единственная внешняя мембрана Кремнезый Во многих видах Никогда Sylleibid или Leuconoid
Эволюционная история
Запись ископаемого



Хотя молекулярные часы и биомаркеры предполагают, что губки существовали задолго до в кембрийском языке взрыва жизни , спикулы кремнезема , такие как у косопонга, отсутствуют в ископаемом записи до кембрия. [ 87 ] Необеспеченный отчет за 2002 год существует в спикулах в скалах, датированных около 750 миллионов лет назад . [ 88 ] Хорошо сохранившиеся ископаемые губки примерно в 580 миллионов лет назад в эдиакаранском периоде были обнаружены в формировании Душантуо . [ 89 ] Эти окаменелости, которые включают в себя: спикулы; пинакоциты ; Пороциты ; археоциты ; склероциты ; и внутренняя полость была классифицирована как демоспожи. Окаменелости стеклянных губков были найдены около 540 миллионов лет назад в скалах в Австралии, Китае и Монголии. [ 90 ] Ранние кембрийские губки из Мексики, принадлежащие к роду , киветинокия показывают признаки слияния нескольких меньших спикул, образуя единую большую спикулу. [ 91 ] Спикулы карбоната кальция из известковых губок были обнаружены в ранних кембрийских породах примерно с 530 до 523 миллионов лет назад в Австралии. Другие вероятные демоспоны были обнаружены в ранней кембрийской фауне Ченгцзян , с 525 до 520 миллионов лет назад . [ 92 ] Окаменелости, найденные на канадских северо -западных территориях, датируемых 890 миллионами лет назад, могут быть губками; Если этот вывод подтвержден, это предполагает, что первые животные появились перед событием неопротерозойской оксигенации. [ 93 ]
Пресноводные губки, по-видимому, намного моложе, так как самые ранние известные окаменелости датируются среднеэоценовым периодом около 48-40 миллионов лет назад . [ 90 ] Хотя около 90% современных губок являются демоспольными , окаменелые остатки этого типа встречаются реже, чем у других типов, потому что их скелеты состоят из относительно мягкой скучки, которая плохо оказывает. [ 94 ] Самые ранние симбионты губки известны из раннего силурия . [ 95 ]
Химический индикатор представляет собой 24-изопропил- холесестан , который представляет собой стабильную производную 24-изопропилового холестерина , который, как говорят, производится демоспонами , но не эйметазоанами («Истинные животные», то есть Cnidarians и Bilaterians ). Поскольку считается, что чонофлагелляты являются самыми близкими одноклельными родственниками животных, группа ученых изучила биохимию и гены одного вида хоанофлагеллята . Они пришли к выводу, что этот вид не может производить 24-изопропилхолестерина, но для того, чтобы доказать, что ископаемое 24-изопропилхолстан могло быть вызвано только исследование более широкого диапазона хоанофлагеллятов. [ 96 ] Хотя в предыдущей публикации сообщалось о следах химического 24-изопропил- холестана в древних породах, начиная с 1800 миллионов лет назад , [ 97 ] Недавнее исследование с использованием гораздо более точно устаревшей серии Rock показало, что эти биомаркеры появляются только до окончания оледенения Мариноан примерно 635 миллионов лет назад , [ 98 ] и что «анализ биомаркеров еще не выявил каких-либо убедительных доказательств древних губке, предварительно датирующих первым глобально обширным неопротерозойским ледниковым эпизодам (Штуртиан, ~ 713 миллионов лет назад в Омане)». Хотя утверждалось, что этот «биомаркер губки» мог возникнуть из морских водорослей, недавние исследования показывают, что способность водорослей продуцировать этот биомаркер развивался только в каменноугольном виде ; Таким образом, биомаркер по -прежнему сильно поддерживает присутствие демоспонов в криогенском. [ 99 ] [ 100 ] [ 101 ]
Археоциатиды , которые некоторые классифицируют как тип кораллической губки, являются очень распространенными окаменелостями в скалах раннего кембрия около 530-520 миллионов лет назад , но, очевидно, умерли к концу кембрийского 490 миллионов лет назад . [ 92 ] Было высказано предположение, что они были произведены: губки; Cnidarians ; водоросли ; Форманиферанс ; Полностью отдельный тип животных, археоциатха; Или даже совершенно отдельное королевство жизни, помеченное архаатей или подводимостью. С 1990 -х годов археоцииды считались отличительной группой губок. [ 79 ]

Трудно вписать канцлорииды в классификации губки или более сложных животных. Анализ в 1996 году пришел к выводу, что они были тесно связаны с губками на том основании, что детальная структура канцлероридных склеритов («Пластины брони») аналогична структуре волокнов Spongin, коллагенового белка , в современных кератозе (роговые) демоспон . Дарвинелла . [ 103 ] Тем не менее, другой анализ в 2002 году пришел к выводу, что канцлориды не являются губками и могут быть промежуточными между губками и более сложными животными, среди прочих причин, потому что их шкуры были толще и более плотно связаны, чем у губ. [ 104 ] В 2008 году подробный анализ склеритов Chancelloriids пришел к выводу, что они были очень похожи на то, что у Halkieriids , мобильных билатеровских животных, которые выглядели как слизняки в цепной почве и чьи ископаемые встречаются в скалах от очень раннего кембрийского до середины кембрийского. Если это правильно, это создало бы дилемму, поскольку крайне маловероятно, что совершенно не связанные организмы могли бы развить такие похожие склериты независимо, но огромная разница в структурах их тела затрудняет, как они могут быть тесно связаны. [ 102 ]
Отношения с другими группами животных

| Упрощенное семейное древо, показывающее известковые губки как самые близкие к более сложным животным [ 105 ] |
| Упрощенное семейное древо, показывающее гомосклероморфу как ближе к более сложным животным [ 106 ] | ||||||||||||||||||
|
В 1990-х годах губки широко рассматривались как монофилетическая группа, и все они происходили от общего предка , который сам был губкой, и как «сестринская группа» для всех других метазоанов (многоклеточные животные), которые сами образуются Монофилетическая группа. С другой стороны, некоторые анализы 1990-х годов также возродили идею о том, что ближайшие эволюционные родственники животных являются хонофлагеллятами , одноклеточные организмы, очень похожие на хоаноциты губ . Не быть монофилетическим, так как те же самые похожие на губчатую предки могли выразить как современные губки, так и для членов Metazoa. [ 105 ]
Анализ с 2001 года пришел к выводу, что эйметазоа (более сложный, чем губки) более тесно связаны с конкретными группами губок, чем с другими губчатыми группами. Такие выводы подразумевают, что губки не являются монофилетическими, потому что последний общий предок всех губок также будет прямым предком Eumetazoa, который не является губками. Исследование в 2001 году, основанное на сравнении рибосомной ДНК, пришло к выводу, что наиболее фундаментальное разделение внутри губки было между стеклянными губками и остальными, а эуметазоа более тесно связано с известковыми губками (с сопикулами карбоната кальция), чем с другими типами губки. [ 105 ] В 2007 году один анализ, основанный на сравнении РНК , а другой, основанный главным образом на сравнении спикул, пришел к выводу, что демоспони и стеклянные губки более тесно связаны друг с другом, чем с любыми, - это известковые губки, которые, в свою очередь, более тесно связаны с эуметазоа. [ 90 ] [ 107 ]
Другие анатомические и биохимические доказательства связывают Eumetazoa с гомосклероморфом , подгруппой Demosponges. Сравнение в 2007 году ядерной ДНК , исключая стеклянные губки и гребневые желе , пришло к выводу, что:
- Гомосклероморфы наиболее тесно связаны с Eumetazoa;
- Известные губки являются ближе всего;
- Другие демоспонги являются эволюционными «тетями» этих групп; и
- Канцллорииды кембрийских , пакетные животные, у которых окаменелости находятся в скалах , могут быть губки. [ 106 ]
Сперма гомосклероморфы разделяет особенности со спермой эйметазоа, чего не хватает сперматозоидов других губ. Как у гомосклероморфы, так и в эйметазоаа-слоях клеток связаны привязанностью к ковровской базальной мембране, состоящей в основном из коллагена «тип IV» , формы коллагена, не обнаруженного в других губках-хотя спонгинские волокна, которые укрепляют мезогил похож на коллаген «Тип IV». [ 21 ]

Анализы, описанные выше, пришли к выводу, что губки наиболее близки к предкам всех метазоа, всех многоклеточных животных, включая как губки, так и более сложные группы. Тем не менее, еще одно сравнение в 2008 году из 150 генов в каждом из 21 родов, от грибов до людей, но в том числе только два вида губки, предположили, что челюры для гребня ( Ctenophora ) являются наиболее базальной линией метазоа, включенной в выборку. [ 108 ] [ 109 ] [ 110 ] [ 111 ] Если это правильно, то либо современные соболезгиватели разработали свои сложные структуры независимо от других метазоа, либо предки губков были более сложными, и все известные губки являются резко упрощенными формами. Исследование рекомендовало дальнейший анализ с использованием более широкого диапазона губок и других простых метазоа, таких как Placozoa . [ 108 ]
Тем не менее, повторный анализ данных показал, что компьютерные алгоритмы, используемые для анализа, были введены в заблуждение из -за наличия определенных генов Ctenophore, которые заметно отличались от других видов, оставляя губки в качестве сестринской группы для всех других животных или исконной паристической степени Полем [ 112 ] [ 113 ] «Семейные деревья», построенные с использованием комбинации всех доступных данных - морфологических, развития и молекулярных, - пришли к выводу, что губки на самом деле являются монофилетической группой, а с Cnidaries образуют родственную группу для билатерианов. [ 114 ] [ 115 ]
Очень большое и внутренне последовательное выравнивание 1719 белков в масштабе метазоу, опубликованное в 2017 году, показало, что (i) губки-представленные гомосклероморфом, калькарея, гексактинеллида и demospongiae-являются монофилетическими, (ii) спонсы являются сестринскими группами ко всем Другие многоклеточные животные, (iii) Ctenophores появляются как вторая самая растопленная линия разветвленных животных, а (iv) плакозозой появляется в качестве третьей линии животных, за которой следует сестринская группа сестрин с билатерианцами . [ 116 ]
В марте 2021 года ученые из Дублина обнаружили дополнительные доказательства того, что губки являются родственной группой для всех других животных, [ 117 ] Находясь в мае 2023 года, Schultz et al. Найденные паттерны необратимых изменений в синтенции генома, которые дают убедительные доказательства того, что Ctenophores являются родственной группой для всех других животных вместо этого. [ 118 ]
Примечательные спонгиологи
Использовать

Дельфинами
В отчете в 1997 году было описано использование губок в качестве инструмента с помощью узких слоев дельфинов в Шак -Бэй в Западной Австралии. Дельфин прикрепит морскую губку к своей роструме , которая, по -видимому, затем используется для защиты ее при поиске пищи на песчаном море . [ 119 ] Поведение, известное как губчатая , наблюдается только в этом заливе и почти исключительно показано женщинами. Исследование в 2005 году пришло к выводу, что матери преподают поведение своим дочерям и что все пользователи губки тесно связаны, что предполагает, что это довольно недавнее новшество. [ 15 ]
Людьми


Скелет
Спикулы кальция или кремнезема карбоната большинства губчатых родов делают их слишком грубыми для большинства применений, но два рода, гиппоспонга и спонгии имеют мягкие, полностью волокнистые скелеты. [ 12 ] : 88 Ранние европейцы использовали мягкие губки для многих целей, в том числе заполнение для шлемов, портативную посуду и муниципальные фильтры воды. До изобретения синтетических губок они использовались в качестве инструментов для чистки, аппликаторов для красок, керамических глазури и осторожных контрацептивов . Тем не менее, к середине 20-го века, перевод, как животных, так и отрасль близко к исчезновению . [ 120 ]
Многие объекты с губчатыми текстурами теперь сделаны из веществ, не полученных из Poriferans. Синтетические губки включают личные и домашние чистящие инструменты , грудные имплантаты , [ 121 ] и противозачаточные губки . [ 122 ] Типичными используемыми материалами являются пена целлюлозы , полиуретановая пена и реже силиконовой пены.
«Губка» Луфы ( , также написанная лаффи , которая обычно продается для использования на кухне или душе, не получена не от животного, а в основном из волокнистого «скелета» губки Luffa aegyptiaca , Cucurbitaceae ) . [ 123 ]
Антибиотические соединения
Губки имеют лекарственный потенциал из -за присутствия самих губок или их микробных симбионтов химических веществ, которые могут использоваться для контроля вирусов , бактерий , опухолей и грибов. [ 124 ] [ 125 ]
Другие биологически активные соединения

Отсутствие какой -либо защитной оболочки или средств побега, губки эволюционировали для синтеза различных необычных соединений. Одним из таких классов являются производные окисленных жирных кислот, называемые оксилипинами . Было обнаружено, что члены этого семейства обладают противораковыми, антибактериальными и противогрибковыми свойствами. Один пример, выделенный из губки Окинавана Плакотиса , Плакоридин А , показал потенциал в качестве цитотоксина к клеткам мышиной лимфомы. [ 126 ] [ 127 ]
Смотрите также
- Списки губ
- Губцовый рифовый проект
- 3-алкилпиридиния , соединения, обнаруженные в морских галлосклерады губчах
Ссылки
- ^ Zumberge, J. Alex; Любовь, Гордон Д.; Кармас, Пако; Сперлинг, Эрик А.; Гунасекера, Сунити; Рорсен, Меган; Грожан, Эммануэль; Grotzinger, John P.; Призыв, Роджер Э. (ноябрь 2018 г.). «Стероидный биомаркер Demosponge 26-метилстигмастан предоставляет доказательства неопротерозойских животных» . Природа экология и эволюция . 2 (11): 1709–1714. Bibcode : 2018natee ... 2.1709Z . doi : 10.1038/s41559-018-0676-2 . PMC 6589438 . PMID 30323207 .
- ^ Pajdzińska, A. (2018). «Животные умирают более мелко: они не умерли, они мертвы. Животные в польском языковом мировоззрении и в современных науках о жизни» . Этнолингвистический . 29 : 147–161. doi : 10.17951/et.2017.29.135 .
- ^ "Порифера" . Мировой реестр морских видов . Фландерс Морский институт. 2024 . Получено 8 мая 2024 года .
- ^ "Порифера" . Merriam-Webster.com Словарь . Мерриам-Уэбстер.
- ^ Jump up to: а беременный Феуда, Роберто; Дорманн, Мартин; Петт, Уокер; Филипп, Эрве; Рота-Стабелли, Омар; Лартиллот, Николас; Wörheide, Gert; Пизани, Давид (декабрь 2017 г.). «Улучшенное моделирование композиционной гетерогенности поддерживает губки как сестры для всех других животных» . Текущая биология . 27 (24): 3864–3870.e4. Bibcode : 2017cbio ... 27e3864f . doi : 10.1016/j.cub.2017.11.008 . PMID 29199080 .
- ^ Jump up to: а беременный Мороз, LL; Ромава, Да (23 декабря 2022 г.). «Альтернативные нейронные системы: что такое нейрон? (Ctenophores, губки и плацеозо)» . Границы в клеточной биологии и развитии . 10 : 1071961. DOI : 10.3389/fcell.2022.1071961 . PMC 9816575 . PMID 36619868 .
- ^ «Спонгиология» . Merriam-Webster.com Словарь . Мерриам-Уэбстер . Получено 27 декабря 2017 года .
- ^ Лидделл, Генри Джордж; Скотт, Роберт (1940). "σπόγγος" . Грек-английский лексикон . Архивировано из оригинала 5 сентября 2021 года . Получено 5 сентября 2021 года - через Персей.
- ^ Jump up to: а беременный Hooper, J. (2018). «Структура губки» . Квинслендский музей . Архивировано с оригинала 26 сентября 2019 года . Получено 27 сентября 2019 года .
- ^ Thacker, Robert W.; Диас, Мария Кристина; Кернер, Аделина; Vignes-lebbe, régine; Сегерделл, Эрик; Хаендель, Мелисса А.; Мунгалл, Кристофер Дж. (8 сентября 2014 г.). «Онтология Porifera (Poro): улучшение систематики губки с помощью онтологии анатомии» . Журнал биомедицинской семантики . 5 (1): 39. doi : 10.1186/2041-1480-5-39 . PMC 4177528 . PMID 25276334 .
- ^ Коллинз, AG (декабрь 1998 г.). «Оценка множества альтернативных гипотез для происхождения Bilateria: анализ молекулярных данных 18S рРНК» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (26): 15458–63. Bibcode : 1998pnas ... 9515458c . doi : 10.1073/pnas.95.26.15458 . PMC 28064 . PMID 9860990 .
- ^ Jump up to: а беременный в дюймовый и фон глин Bergquist, PR (1978). Губки . Лондон: Хатчинсон. ISBN 978-0-520-03658-1 .
- ^ Valet, J.; Duport, E. (2004). «Захват добычи и пищеварение в грилеяльной губчатой гипогеи асбестопума (Porifera: Demospongiae)». Zoomorphology . 123 (4): 179–190. doi : 10.1007/s00435-004-0100-0 . S2CID 24484610 .
- ^ Кларк, Массачусетс; Choi, J.; Дуглас, М. (2018) Биология 2E [ Постоянная мертвая ссылка ] , Стр. 776, OpenStax . ISBN 978-1-947172-52-4 .
- ^ Jump up to: а беременный Крутцен, Майкл; Манн, Джанет; Heithaus, Michael R.; Коннор, Ричард С.; Bejder, Lars; Шервин, Уильям Б. (июнь 2005 г.). «Культурная передача использования инструментов в узкополосных дельфинах» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (25): 8939–43. Bibcode : 2005pnas..102.8939k . doi : 10.1073/pnas.0500232102 . PMC 1157020 . PMID 15947077 .
- ^ Jump up to: а беременный в дюймовый и фон глин Хупер, JNA; Ван Соэт, RWM; Дебренн Ф. (2002). "Phylum Porifera Grant, 1836". В Хупер, JNA; Ван Соэт, RWM (ред.). Система Порифера: Руководство по классификации губок . Нью -Йорк: Kluwer Academic/Plenum. С. 9–14. ISBN 978-0-306-47260-2 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. ведущий с Т в v В х и С аа Аб и объявление Но из в нравиться это к и Рупперт, Эдвард Э.; Fox, Richard S.; Барнс, Роберт Д. (2004). Зоология беспозвоночных (7 -е изд.). Brooks / Cole Publishing . С. 76–97. ISBN 978-0-03-025982-1 .
- ^ Бакшани, Кэсси Р.; Morales-Garcia, Ana L.; Альтаус, Майк; Уилкокс, Мэтью Д.; Пирсон, Джеффри П.; Bythell, John C.; Burgess, J. Grant (2018-07-04). «Эволюционное сохранение антимикробной функции слизи: первая защита от инфекции» . NPJ Биопленки и микробиомы . 4 (1): 14. doi : 10.1038/s41522-018-0057-2 . PMC 6031612 . PMID 30002868 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. ведущий с Т Bergquist, PR (1998). "Порифера". В Андерсоне, DT (ред.). Зоология беспозвоночных . Издательство Оксфордского университета. С. 10–27. ISBN 978-0-19-551368-4 .
- ^ Jump up to: а беременный в Хинде, RT (1998). Полем В Андерсоне, DT (ред.). Инверберат зоология . Пресс стр. 28–57. ISBN 978-0-19-551368-4 .
- ^ Jump up to: а беременный Exposito, Жан-Ив; Клузель, Кэролайн; Гаррон, Роберт; Летиас, Клэр (ноябрь 2002 г.). «Эволюция коллагенов» . Анатомическая запись . 268 (3): 302–16. Doi : 10,1002/ar.10162 . PMID 12382326 .
- ^ Ruppert EE, Fox RS, Barnes Rd (2004). Зоология беспозвоночных (7 -е изд.). Брукс / Коул. п. 82. ISBN 978-0-03-025982-1 .
- ^ Musser, Jacob M.; Schippers, Klaske J.; Никель, Майкл; Мизсон, Джулия; Кон, Андреа Б.; Папе, Константин; и др. (Ноябрь 2021 г.). «Профилирование клеточного разнообразия в губках информирует тип животных клеток и эволюцию нервной системы» . Наука . 374 (6568): 717–723. Bibcode : 2021sci ... 374..717M . doi : 10.1126/science.abj2949 . PMC 9233960 . PMID 34735222 .
- ^ Ривера, как; Ozturk, N.; Фахей, Б.; Plachetzki, DC; ДеГнан, Б.М.; Sancar, A.; Оукли, Т. Т. (апрель 2012 г.). «Криптохромный рецепт с синим светом экспрессируется в губчатом глазу, в которых отсутствуют нейроны и опсин» . Журнал экспериментальной биологии . 215 (Pt 8): 1278–86. doi : 10.1242/jeb.067140 . PMC 3309880 . PMID 22442365 .
- ^ Рупперт, Эдвард Э.; Fox, Richard S.; Барнс, Роберт Д. (2004). Зоология беспозвоночных (7 -е изд.). Брукс / Коул. п. 78. ISBN 978-0-03-025982-1 .
- ^ Хикман, CP; Робертс, LS; Ларсон А. (2001). Интегрированные принципы зоологии (11 -е изд.). Нью-Йорк: МакГроу-Хилл . п. 247. ISBN 978-0-07-290961-6 .
- ^ «Портал идентификации морских видов: Halisarca Dujardini» . виды идентификация.org . Архивировано из оригинала 2020-10-17 . Получено 2019-08-02 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а Bergquist, PR (2001). «Порифера (губки)». Энциклопедия наук о жизни . Джон Уайли и сыновья. doi : 10.1038/npg.els.0001582 . ISBN 978-0-470-01617-6 .
- ^ Jump up to: а беременный в Krautter, M. (1998). «Экология кремнистых губков: применение к окружающей среде интерпретации фаций губки верхней юрского периода (Оксфордский) из Испании» (PDF) . Cuadernos de Geología Ibérica . 24 : 223–239. Архивировано из оригинала (PDF) 19 марта 2009 года . Получено 2008-10-10 .
- ^ Yahel, G.; Уитни, Ф.; Рейсвиг, HM; Eerces-Medrano, di; Лейс, SP (2007). «Кормление in situ и метаболизм стеклянных губок (Hexactinellida, Porifera) изучались в глубоком умеренном фьорде с дистанционно управляемым погружным». Лимнология и океанография . 52 (1): 428–440. Bibcode : 2007limoc..52..428y . Citeseerx 10.1.1.597.9627 . doi : 10.4319/lo.2007.52.1.0428 . S2CID 86297053 .
- ^ Ван Соэт, Роб Вм; Бури-Эзна, Николь; Веклет, Джин; Дорманн, Мартин; Эрпенбек, Дирк; Де Вогд, Николь Дж.; Santodomingo, Nadiezhda; Ванхорн, Барт; Келли, Мишель ; Хупер, Джон На (2012). «Глобальное разнообразие губок (Porifera)» . Plos один . 7 (4): E35105. BIBCODE : 2012PLOSO ... 735105V . doi : 10.1371/journal.pone.0035105 . PMC 3338747 . PMID 22558119 .
- ^ «4 новых вида« убийственных »губок обнаружены у побережья Тихого океана» . CBC News . 19 апреля 2014 года. Архивировано с оригинала 19 апреля 2014 года . Получено 2014-09-04 .
- ^ Jump up to: а беременный в дюймовый Valet, J. (2008). «Новый род хищных губок (Porifera: Poecilosclerida, Cladorhizidae) из Deep Ne Pacific и замечания о роде Neocladia » (PDF) . Zootaxa . 1752 : 57–65. doi : 10.11646/Zootaxa.1752.1.3 . Архивировано (PDF) из оригинала на 2008-09-06 . Получено 2008-10-31 .
- ^ Уотлинг Л. (2007). «Хищничество на копеподах аляскинской губкой кладорхизидов». Журнал Морской биологической ассоциации Соединенного Королевства . 87 (6): 1721–1726. Bibcode : 2007jmbuk..87.1721w . doi : 10.1017/s0025315407058560 . S2CID 86588792 .
- ^ Jump up to: а беременный в Valet, J.; Boury-esnault, N. (1995). «Плотоядные губки» . Природа . 373 (6512): 333–335. Bibcode : 1995natur.373..333v . doi : 10.1038/3733333A0 . S2CID 4320216 .
- ^ Valet, J.; Келли, Мишель (2008). «Новые виды из глубокой части Тихого океана предполагают, что плотоядные губки датируются ранним юрным» . Природа предшествует . doi : 10.1038/npre.2008.2327.1 .
- ^ Брюммер, Франц; Пфаннкучен, Мартин; Балц, Александр; Хаузер, Томас; Тиль, Вера (2008). "Свет внутри губки". Журнал экспериментальной морской биологии и экологии . 367 (2): 61–64. Bibcode : 2008jembe.367 ... 61b . doi : 10.1016/j.jembe.2008.06.036 .
- Уокер, Мэтт (10 ноября 2008 г.). «Эксперты« Волокна -оптики »природы» . BBC News . Архивировано из оригинала 17 декабря 2008 года . Получено 11 ноября 2008 года .
- ^ Jump up to: а беременный в Смит, DG; Пеннак, RW (2001). Пресноводные беспозвоночные Пеннака в Соединенных Штатах: Порифера до ракообразной (4 изд.). Джон Уайли и сыновья . С. 47–50. ISBN 978-0-471-35837-4 .
- ^ Díez-Vives, Кристина; Koutsouveli, Vasiliki; Conejero, Maria; Рисго, Ана (26 октября 2022 г.). «Глобальные закономерности в стратегиях выбора и передачи симбионта в губках» . Границы в экологии и эволюции . 10 doi : 10.3389/fevo.2022.1015592 . ISSN 2296-701X .
- ^ Перевозчик, Тайлер Дж.; Мальдонадо, Мануэль; Шмиттманн, Лара; Пита, Люсия; Бош, Томас К.Г.; Hentschel, Ute (май 2022). «Передача симбионта в морских губках: воспроизведение, развитие и метаморфоза» . BMC Biology . 20 (1): 100. doi : 10.1186/s12915-022-01291-6 . PMC 9077847 . PMID 35524305 .
- ^ Рисго, Ана; Табоада, Серхио; Санчес-Вила, Лора; Сола, Джоан; Бертран, Андреа; Авила, Конксита (18 марта 2015 г.). «Некоторым нравится It Fat: сравнительная ультраструктура эмбриона в двух демонстрациях рода Mycale (порядка Poecilosclerida) из Антарктиды и Карибского бассейна» . Plos один . 10 (3): E0118805. BIBCODE : 2015PLOSO..1018805R . doi : 10.1371/journal.pone.0118805 . ISSN 1932-6203 . PMC 4365022 . PMID 25785444 .
- ^ Лейс, SP (16 февраля 2006 г.). «Эмбриогенез в стеклянной губке oopsacas minuta: образование синциты путем слияния бластомеров». Интегративная и сравнительная биология . 46 (2): 104–117. doi : 10.1093/ICB/ICJ016 . ISSN 1540-7063 . PMID 21672727 .
- ^ Jump up to: а беременный в Koutsouveli, Vasiliki; Карденнас, Пако; Holydomingo, Nadiezda; Марина, Анабель; Морато, Эсперанса; Рапп, Ганс разорвал; Стайго, Ана (16 декабря 2020 г.). «Молекулярное механизм гаметогенеза в геоспособности Geodia (хлеб): эволюционное происхождение консервативного инструментария между животными » Молекулярная биология и эволюция 37 (12): 3485–3 Doi : 10.1093/ molbev/ msea1 ISSN 0737-4 7743902PMC PMID 32929503
- ^ Никель, М. (декабрь 2004 г.). «Кинетика и ритм сокращений тела в губке Тетя Вильгельма (Porifera: Demospongiae)» . Журнал экспериментальной биологии . 207 (Pt 26): 4515–24. doi : 10.1242/jeb.01289 . PMID 15579547 .
- ^ Сакарья, Онур; Армстронг, Кэтрин А.; Адамска, Майя; Адамски, Марцин; Ван, я-фан; Тидор, Брюс; ДеГнан, Бернард М.; Оукли, Тодд Х.; Косик, Кеннет С. (6 июня 2007 г.). «Пост-синаптический каркас на происхождении животного царства» . Plos один . 2 (6): E506. Bibcode : 2007ploso ... 2..506s . doi : 10.1371/journal.pone.0000506 . ISSN 1932-6203 . PMC 1876816 . PMID 17551586 .
- ^ Уивер, Джеймс С.; Айзенберг, Джоанна; Фантнер, Георг Э.; Кисайлс, Дэвид; Woesz, Александр; Аллен, Петр; Филдс, Кирк; Портер, Майкл Дж.; Зок, Фрэнк У.; Хансма, Пол К.; Фратцл, Питер; Морс, Даниэль Э. (2007). «Иерархическая сборка кремнистой скелетной решетки гексактинеллидной губки euplectella aspergillum». Журнал структурной биологии . 158 (1): 93–106. doi : 10.1016/j.jsb.2006.10.027 . PMID 17175169 .
- ^ Рузицка, R; Глисон, DF (январь 2008 г.). «Распространение широтных рыб и эффективность химической защиты губки» (PDF) . Oecologia . 154 (4): 785–94. Bibcode : 2008oecol.154..785r . doi : 10.1007/s00442-007-0874-0 . PMID 17960425 . S2CID 1495896 . Архивировано из оригинала (PDF) на 2008-10-06.
- ^ Гейдж, JD; Тайлер, Пенсильвания (1996). Глубоководная биология: естественная история организмов на глубоководном этаже . Издательство Кембриджского университета . С. 91–93. ISBN 978-0-521-33665-9 .
- ^ Dunlap, M.; Pawlik, Jr (1996). «Видеомонизированное хищничество от карибских рифов рыб на множестве мангровых и рифовых губок». Морская биология . 126 (1): 117–123. Bibcode : 1996marbi.126..117d . doi : 10.1007/bf00571383 . ISSN 0025-3162 . S2CID 84799900 .
- ^ Loh, tl; Pawlik, Jr (март 2014 г.). «Химическая защита и компромиссы ресурсов структурируют губчатые сообщества на карибских коралловых рифах» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (11): 4151–6. Bibcode : 2014pnas..111.4151L . doi : 10.1073/pnas.13216261111 . PMC 3964098 . PMID 24567392 .
- ^ Пайпер, Росс (2007). Чрезвычайные животные: энциклопедия любопытных и необычных животных . Greenwood Publishing Group . п. 148. ISBN 978-0-313-33922-6 .
- ^ Jump up to: а беременный Гочфельд, диджей; и др. (2012). Стеллер, Д.; Лобель, Л. (ред.). «Динамика популяции губки на карибских рифах» . Дайвинг для науки 2012 . Труды Американской академии подводных наук 31 -й симпозиум. Архивировано с оригинала 2015-09-04.
{{cite journal}}: Cs1 maint: непредвзятый URL ( ссылка ) - ^ Олсон, JB; Гочфельд, диджей; Slattery, M. (июль 2006 г.). «Синдром красной полосы Aplysina: новая угроза для карибских губок» . Болезни водных организмов . 71 (2): 163–8. doi : 10.3354/dao071163 . PMID 16956064 . Сводка на: Кларк, М. (2006-10-17). «Новая болезнь угрожает губками» . Практическое рыболовство . Архивировано из оригинала 2007-09-26.
- ^ Wulff, JL (июнь 2008 г.). «Сотрудничество между видами губки увеличивает разнообразие губки и изобилие на лугах по морскому изделиям» . Морская экология . 29 (2): 193–204. Bibcode : 2008marec..29..193w . doi : 10.1111/j.1439-0485.2008.00224.x .
- ^ Даффи, JE (1996). «Границы видов, специализация и радиация губчатых креветков Альфейд» . Биологический журнал Линневого общества . 58 (3): 307–324. doi : 10.1111/j.1095-8312.1996.tb01437.x .
- ^ Мерфи, RC (2002). Коралловые рифы: города под морями . The Darwin Press , Inc. с. 51. ISBN 978-0-87850-138-0 .
- ^ Сэндфорд Ф. (2003). «Ассоциации динамики населения и эпибионтов крабов отшельника (ракообразные: Decapoda: Paguroidea) на острове собаки, Флорида» (PDF) . Мемуары музея Виктория . 60 (1): 45–52. doi : 10.24199/j.mmv.2003.60.6 . ISSN 1447-2554 . S2CID 86167606 . Архивировано (PDF) из оригинала 2018-07-19 . Получено 2022-01-24 .
- ^ Окуковиак, М. (18 декабря 2020 г.). «Использование спикул губки в таксономических, экологических и экологических реконструкциях: обзор» . ПЕРЕЙ . 8 : E10601. doi : 10.7717/peerj.10601 . PMC 7751429 . PMID 33384908 .
- ^ Арчер, Стефани К.; Кан, Аманда С.; Тисс, Мэри; Закон, Лорен; Лейс, Салли П.; Йоханнесен, София С.; Мирян, Крейг А.; Берк, Лили; Данхэм, Аня (24 сентября 2020 г.). «Фонд видов изобилия влияет на топологию пищевой сети на рифах стеклянной губки» . Границы в морской науке . 7 Frontiers Media SA. doi : 10.3389/fmars.2020.549478 . ISSN 2296-7745 .
 Материал был скопирован из этого источника, который доступен в рамках Attribution Creative Commons 4.0 Международная лицензия, архивировавшаяся 2017-10-16 на машине Wayback .
Материал был скопирован из этого источника, который доступен в рамках Attribution Creative Commons 4.0 Международная лицензия, архивировавшаяся 2017-10-16 на машине Wayback .
- ^ Jump up to: а беременный в дюймовый Rix, L.; de Goeij, JM; Van Oevelen, D.; Удар, U.; Аль-Хорани, Фа; Wild, C.; Науманн, MS (23 февраля 2018 г.). «Рифы-губки облегчают передачу органического вещества, полученного из кораллов в связанную фауну через петлю губки». Серия прогресса в морской экологии . 589 : 85–96. Bibcode : 2018meps..589 ... 85r . doi : 10.3354/meps12443 . ISSN 0171-8630 . Материал был скопирован из этого источника, который доступен в рамках Attribution Creative Commons 4.0 Международная лицензия Архивирована 2017-10-16 на машине Wayback
- ^ Rix L, de Goeij JM, Mueller CE, Struck U и другие (2016) «Слизь кораллов питает петлю губки в экосистемах теплых и холодных коралловых рифов». SCI Rep , 6 : 18715.
- ^ Jump up to: а беременный Рикс, Лора; de Goeij, Jasper M.; van Oevelen, Dick; Ударил, Ульрих; Al-Horani, Fuad A.; Дикий, христианин; Науманн, Малик С. (март 2017 г.). «Дифференциальная утилизация кораллов и водорослей растворяет органическое вещество через петлю губки». Функциональная экология . 31 (3): 778–789. Bibcode : 2017fueco..31..778r . doi : 10.1111/1365-2435.12758 .
- ^ Crossland, CJ (июль 1987 г.). «Выпуск слизи и Doc-липида in situ из кораллов acropora variabilis и stylophora pistillata в разных режимах света». Коралловые рифы . 6 (1): 35–42. Bibcode : 1987corre ... 6 ... 35c . doi : 10.1007/bf00302210 .
- ^ Jump up to: а беременный Дикий, христианин; Хуеттель, Маркус; Клютер, Анке; Кремб, Стефан Дж.; Рашид, Мухаммед Им; Jørgensen, Bo B. (2004). «Слизь кораллов функционирует в качестве энергетического носителя и ловушки частиц в рифовой экосистеме». Природа . 428 (6978): 66–70. Bibcode : 2004natur.428 ... 66w . doi : 10.1038/nature02344 . ISSN 0028-0836 . PMID 14999280 .
- ^ Танака, Ясуаки; Мияджима, Тошихиро; Умезава, Ю; Хаясибара, Такеши; Огава, Хироши; Koike, Isao (2009). «Чистый высвобождение растворенного органического вещества склерактинской коралловой акропорой пульчры». Журнал экспериментальной морской биологии и экологии . 377 (2): 101–106. Bibcode : 2009jembe.377..101t . doi : 10.1016/j.jembe.2009.06.023 .
- ^ Науманн, MS; Haas, A.; Удар, U.; Mayr, C.; El-Zibdah, M.; Wild, C. (сентябрь 2010 г.). «Органическое вещество высвобождение доминирующими герматипическими кораллами Северного Красного моря». Коралловые рифы . 29 (3): 649–659. Bibcode : 2010corre..29..649n . doi : 10.1007/s00338-010-0612-7 .
- ^ Jump up to: а беременный Hoegh-Guldberg, O.; McCloskey, LR; Маскатин Л. (апрель 1987). «Изгнание зооксантеллы симбиотическими кубиками из Красного моря». Коралловые рифы . 5 (4): 201-204. Bibcode : 1987corre ... 5..201h . doi : 10.1007/bf00300964 .
- ^ Багдасарян, G; Маскатин, Л. (2000). «Преференциальное изгнание делящихся клеток водорослей в качестве механизма регуляции водоросли-гнидарийского симбиоза» . Биологический бюллетень . 199 (3): 278–286. doi : 10.2307/1543184 . ISSN 0006-3185 . JSTOR 1543184 . PMID 11147708 .
- ^ Crossland, CJ; Барнс, DJ; Borowitzka, MA (1980). «Производство суточных липидов и слизи в Acuminata Acuminata Staghorn Acuminata». Морская биология . 60 (2–3): 81–90. Bibcode : 1980marbi..60 ... 81c . doi : 10.1007/bf00389151 . ISSN 0025-3162 .
- ^ Тремблей, Паскаль; Гровер, Рено; Магуер, Джин Франсуа; Legendre, Луи; Ferier-Pagès, Кристина (15 апреля 2012 года). «Автотрофический углеродный бюджет в коралловой ткани: новая модель фотосинтировки на основе 13C на основе 13C». Журнал экспериментальной биологии . 215 (8): 1384–1393. doi : 10.1242/jeb.065201 . ISSN 1477-9145 . PMID 22442377 .
- ^ Кузнецы-страницы, c; Leclercq, n; Jaubert, J; Pilgrim, S (2000). "Эскудация пикко- Водная микробная экология 21 : 203–209. два 10.3354/ame021203: ISSN 0948-3
- ^ Wild, C.; и др. (Июль 2010). «Органическое вещество высвобождение организмами коралловых рифов красного моря - потенциальное влияние на микробную активность и in situ o 2 доступность ». Серия прогресса в морской экологии . 411 : 61–71. Bibcode : 2010meps..411 ... 61w . doi : 10.3354/meps08653 .
- ^ Танака, Ясуаки; Огава, Хироши; Miyajima, Toshihiro (2011). «Производство и бактериальное разложение растворенного органического вещества в бахроновом коралловом рифе». Журнал океанографии . 67 (4): 427–437. Bibcode : 2011joce ... 67..427t . doi : 10.1007/s10872-011-0046-z . ISSN 0916-8370 .
- ^ Хорошее, Джаспер М.; Oeves, Дик; Избегайте, Марк JA; Осинге, Рональд; Мидлбург, Джек Дж.; Хороший, Антон ФПМ; Адмирал, Вим (4 октября 2013 г.). «Выживание в морской пустыне: петля губки сохраняет ресурсы Wethin угольные рифы». Наука . 342 (6154): 108–110. Код BIB : 2013SCI… 342..108d . doi : 10,1126/science.1241981 . ISSN 0036-8075 .
- ^ Pita, L.; Rix, L.; Slaby, BM; Franke, A.; Hentschel, U. (2018). «Губка Holobiont в изменяющемся океане: от микробов до экосистемы» . Микробиом . 6 (1). doi : 10.1186/s40168-018-0428-1 . ISSN 2049-2618 . PMC 5845141 . PMID 29523192 . Материал был скопирован из этого источника, который доступен в рамках Attribution Creative Commons 4.0 Международная лицензия, архивировавшаяся 2017-10-16 на машине Wayback .
- ^ Иган, Сухелен; Томас, Торстен (16 июня 2015 г.). «Редакция для: микробный симбиоз морских сидячих хозяев- разнообразие и функции» . Границы в микробиологии . 6 doi : 10.3389/fmicb.2015.00585 . ISSN 1664-302X . PMC 4468920 . PMID 26136729 .
- ^ Jump up to: а беременный Webster, NS; Томас, Т. (апрель 2016 г.). «Гугеном губки» . Мбио . 7 (2): E00135-16. doi : 10.1128/mbio.00135-16 . PMC 4850255 . PMID 27103626 .
- ^ "Spongia Linnaeus, 1759" . Мировой реестр морских видов . Архивировано с оригинала 27 марта 2016 года . Получено 18 июля 2012 года .
- ^ Jump up to: а беременный в Роуленд, С.М.; Стивенс Т. (2001). «Археоциата: история филогенетической интерпретации». Журнал палеонтологии . 75 (6): 1065–1078. doi : 10.1666/0022-3360 (2001) 075 <1065: aahopi> 2,0.co; 2 . JSTOR 1307076 .
- ^ Сперлинг, EA; Пизани, д.; Петерсон, KJ (1 января 2007 г.). «Poriferan Paraphyly и его последствия для докембрийской палеобиологии» (PDF) . Геологическое общество, Лондон, Специальные публикации . 286 (1): 355–368. BIBCODE : 2007GSLSP.286..355S . doi : 10.1144/sp286.25 . S2CID 34175521 . Архивировано из оригинала (PDF) 9 мая 2009 года . Получено 2012-08-22 .
- ^ Whelan, Nathan v.; Кокот, Кевин М.; Мороз, Леонид Л.; Галанних, Кеннет М. (май 2015). «Ошибка, сигнал и размещение сестры Ctenophora для всех других животных» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (18): 5773–8. Bibcode : 2015pnas..112.5773w . doi : 10.1073/pnas.1503453112 . PMC 4426464 . PMID 25902535 .
- ^ Хартман, WD; Горо, Т.Ф. (1970). «Ямайские кораллические губки: их морфология, экология и родственники окаменелости». Симпозиум зоологического общества Лондона . 25 : 205–243. (цитируется mgg.rsmas.miami.edu). Архивированный 2018-08-18 на машине Wayback
- ^ Valet, J. (1985). «Кораллические губки и эволюция Porifera». В Conway Morris, S.; Джордж, JD; Гибсон, Р.; Платт, HM (ред.). Происхождение и отношения нижних беспозвоночных . Издательство Оксфордского университета . С. 1–13. ISBN 978-0-19-857181-0 .
- ^ Гэтч, Ева; Лапеби, Паскаль; Ренар, Эммануэль; Веклет, Джин; Рочер, Кэролайн; Эресковский, Александр V.; Лавров, Деннис В.; Borchiellini, Carole (декабрь 2010 г.). «Молекулярная филогения восстанавливает супрагенерическое подразделение гомосклероморфных губок (Porifera, homoscleromorpha)» . Plos один . 5 (12): E14290. BIBCODE : 2010PLOSO ... 514290G . doi : 10.1371/journal.pone.0014290 . PMC 3001884 . PMID 21179486 .
- ^ Гэтч, Ева; Лапеби, Паскаль; Эресковский, Александр V.; Веклет, Джин; Ренар, Эммануэль; Кармас, Пако; Борчиллини, Кэрол (май 2012 г.). «Больше не Demospongiae: Homoscleromorpha формальная номинация как четвертый класс Porifera» (PDF) . Гидробиология . 687 : 3–10. doi : 10.1007/s10750-011-0842-x . S2CID 14468684 . Архивировано (PDF) из оригинала 2019-08-01 . Получено 2019-08-01 .
- ^ Cavalcanti, ff; Клаутау, М. (2011). «Соленоид: новая водная система для Porifera». Zoomorphology . 130 (4): 255–260. doi : 10.1007/s00435-011-0139-7 . S2CID 21745242 .
- ^ Сперлинг, EA; Робинсон, JM; Пизани, д.; Петерсон, KJ (январь 2010 г.). «Где стекло? Биомаркеры, молекулярные часы и микроРНК предполагают 200 -м мильную пропущенную докембрийскую ископаемые записи силисенских губчатых спикул». Геобиология . 8 (1): 24–36. Bibcode : 2010gbio .... 8 ... 24S . doi : 10.1111/j.1472-4669.2009.00225.x . PMID 19929965 . S2CID 41195363 .
- ^ Reitner, J; Wörheide, G. (2002). «Неиспадающие ископаемые Demospongiae-происхождение их палеобиоразнообразии и выделения в истории сохранения». В Hooper, JN; Ван Соэт, RW (ред.). Система Porifera: Руководство по классификации губок (PDF) . Нью -Йорк, Нью -Йорк: Kluwer Academic Plenum. Архивировано (PDF) из оригинала 2008-12-16 . Получено 4 ноября 2008 года .
- ^ Ли, Чиа-Вей; Чен, Джун-Юан; Хуа, Цзы-Эн (февраль 1998 г.). «Докембрийские губки с клеточными структурами». Наука . 279 (5352): 879–882. Bibcode : 1998sci ... 279..879L . doi : 10.1126/science.279.5352.879 . PMID 9452391 .
- ^ Jump up to: а беременный в Мюллер, Вег; Джинхе Ли; Schröder, HC; Ли Цяо; Сяхонг Ван (2007). «Уникальный скелет силисенских губков (Porifera; hexactinellida и demospongiae), которые эволюционировали первым из Urmetazoa во время протерозоя: обзор» . Биогеонов . 4 (2): 219–232. Bibcode : 2007bgeo .... 4..219m . doi : 10.5194/bg-4-219-2007 .
- ^ McMenmin, MA (2008). Полем Геологический акт 6 (4): 363–3
- ^ Jump up to: а беременный Ли, Чиа-Вей; Чен, Джун-Юан; Хуа, Цзы-Эн (февраль 1998 г.). «Докембрийские губки с клеточными структурами». Наука . 279 (5352): 879–82. Bibcode : 1998sci ... 279..879L . doi : 10.1126/science.279.5352.879 . PMID 9452391 . S2CID 38837724 .
- ^ Тернер, ЕС (август 2021 г.). «Возможные окаменелости для тела Poriferan в ранних неопротерозойских микробных рифах» . Природа . 596 (7870): 87–91. Bibcode : 2021natur.596 ... 87t . doi : 10.1038/s41586-021-03773-z . PMC 8338550 . PMID 34321662 .
- ^ «Демоспонгия» . Калифорнийский музей палеонтологии . Беркли, Калифорния: UC Berkeley . Архивировано из оригинала 18 октября 2013 года . Получено 2008-11-27 .
- ^ Винн, Олев; Уилсон, Марк А.; ТУМ, Урсула; Mõtus, Mari-Ann (2015). «Самый ранний известный симбиоз Rugosan-Stromatoporoid из Llandovery of Estonia (Baltica)» . Палеогеография, палеоклиматология, палеоэкология . 31 : 1–5. Bibcode : 2015ppp ... 431 .... 1V . doi : 10.1016/j.palaeo.2015.04.023 . Архивировано из оригинала 2024-04-10 . Получено 2015-06-18 .
- ^ Коднер, Робин Б.; Призыв, Роджер Э.; Пирсон, Энн; Король, Николь; Кнолл, Эндрю Х. (июль 2008 г.). «Стеролы в одноклеточном родственнике метазоя» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (29): 9897–9902. Bibcode : 2008pnas..105.9897k . doi : 10.1073/pnas.0803975105 . PMC 2481317 . PMID 18632573 .
- ^ Николс, с.; Wörheide, G. (апрель 2005 г.). «Губки: новые виды старых животных». Интегративная и сравнительная биология . 45 (2): 333–334. Citeseerx 10.1.1.598.4999 . doi : 10.1093/icb/45.2.333 . PMID 21676777 .
- ^ Любовь, Гордон Д.; Грожан, Эммануэль; Сталви, Шарлотта; Fike, David A.; Grotzinger, John P.; Брэдли, Александр С.; и др. (Февраль 2009 г.). «Окаменечные стероиды регистрируют появление Demospongiae в криогенский период» (PDF) . Природа . 457 (7230): 718–721. Bibcode : 2009natur.457..718L . doi : 10.1038/nature07673 . PMID 19194449 . S2CID 4314662 . Архивировано из оригинала (PDF) 2018-07-24 . Получено 2019-08-01 .
- ^ Antcliffe, JB (2013). Stouge, S. (ed.). «Опрос доказательств органических соединений, называемых биомаркерами губки» . Палеонтология . 56 (5): 917–925. Bibcode : 2013Palgy..56..917a . doi : 10.1111/pala.12030 .
- ^ Золото, да (сентябрь 2018 г.). «Медленный рост сложной жизни, как показано с помощью генетики биомаркера». Новые темы в науках о жизни . 2 (2): 191–199. doi : 10.1042/etls20170150 . PMID 32412622 . S2CID 90887224 .
- ^ Золото, Дэвид А.; Grabenstatter, Джонатан; де Мендоса, Алекс; Рисго, Ана; Руис-Трильо, Иьяки; Призыв, Роджер Э. (март 2016 г.). «Стерол и геномный анализ подтверждают гипотезу биомаркера губки» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (10): 2684–2689. BIBCODE : 2016PNAS..113.2684G . doi : 10.1073/pnas.1512614113 . PMC 4790988 . PMID 26903629 .
- ^ Jump up to: а беременный Портер, С.М. (2008). «Скелетная микроструктура указывает на канцлорииды и Halkieriids тесно связаны» . Палеонтология . 51 (4): 865–879. Bibcode : 2008Palgy..51..865p . doi : 10.1111/j.1475-4983.2008.00792.x .
- ^ Баттерфилд, Нью -Джерси; Николас, CJ (1996). «Берджесс сланцевого типа. Журнал палеонтологии . 70 (6): 893–899. Bibcode : 1996jpal ... 70..893b . doi : 10.1017/s0022336000038579 . JSTOR 1306492 . S2CID 133427906 .
- ^ Jusensen, Dorte; Штайнер, Майкл; Маоян, Чжу (2002). «Новые хорошо сохранившиеся Scletloridae из ранней камбрийской формирования Юансан (Ченгцзян, Китай) и Среднего Камбрийского Уилера-сланца (Юта, США) и палеобиологические последствия» Журнал палеонтологии 76 (4): 596–6 Bibcode : 2002Jpal ... 76..596J Doi : 10.1666/0022-3360 (2002) 076 <10596: NWPSOC> 2,0.co ; S2CID 129127213 Бесплатный текст в Януссен Д. (2002). "(Полный текст без изображений)" . Журнал палеонтологии . Архивировано из оригинала 10 декабря 2008 года . Получено 2008-08-04 .
- ^ Jump up to: а беременный в Borchiellini, C.; Мануэль, М.; Аливон, Е.; Boury-esnault, n.; Valet, J.; Le Parco, Y. (январь 2001 г.). "Губка парафила и оригинал метазоа " Журнал эволюционной биологии 14 (1): 171–1 Doi : 10.1046/j.1420-9101.2001.000244.x . PMID 29280585
- ^ Jump up to: а беременный Сперлинг, EA; Пизани, д.; Петерсон, К.Дж. (2007). «Poriferan Paraphyly и его последствия для докембрийской палеобиологии» (PDF) . Журнал Геологического общества Лондона . 286 (1): 355–368. BIBCODE : 2007GSLSP.286..355S . doi : 10.1144/sp286.25 . S2CID 34175521 . Архивировано из оригинала (PDF) 9 мая 2009 года . Получено 2008-11-04 .
- ^ Медина, Моника; Коллинз, Аллен Дж.; Сильберман, Джеффри Д.; Согин, Митчелл Л. (август 2001 г.). «Оценка гипотез базальной филогения животных с использованием полных последовательностей большой и маленькой субъединицы рРНК» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (17): 9707–12. Bibcode : 2001pnas ... 98,9707M . doi : 10.1073/pnas.171316998 . PMC 55517 . PMID 11504944 .
- ^ Jump up to: а беременный Данн, Кейси В.; Хейнол, Андреас; Матус, Дэвид Q.; Панг, Кевин; Браун, Уильям Э.; Смит, Стивен А.; и др. (Апрель 2008 г.). «Широкая филогеномная выборка улучшает разрешение животного дерева жизни». Природа . 452 (7188): 745–9. Bibcode : 2008natur.452..745d . doi : 10.1038/nature06614 . PMID 18322464 . S2CID 4397099 .
- ^ Хейнол, Андреас; OBST, Матиас; Стаматакис, Александрос; Отт, Майкл; Rouse, Greg W.; Edgecombe, Gregory D.; и др. (Декабрь 2009 г.). «Оценка корня билатерианских животных с помощью масштабируемых филогеномных методов» . Разбирательство. Биологические науки . 276 (1677): 4261–70. doi : 10.1098/rspb.2009.0896 . PMC 2817096 . PMID 19759036 .
- ^ Райан, Джозеф Ф.; Панг, Кевин; Schnitzler, Christine E.; Нгуен, Ан-Дао; Морленд, Р. Трэвис; Симмонс, Дэвид К.; и др. (Декабрь 2013). «Геном Ctenophore mnemiopsis leidyi и его последствия для эволюции клеток» . Наука . 342 (6164): 1242592. DOI : 10.1126/Science.1242592 . PMC 3920664 . PMID 24337300 .
- ^ Мороз, Леонид Л.; Кокот, Кевин М.; Citarella, Mathew R.; Dosung, Sohn; Норекиан, Тигран П.; Povolotkaya, Inna s.; и др. (2014). «Геном Ctenophore и эволюционное происхождение нейронных систем» . Природа . 510 (7503): 109–114. Bibcode : 2014natur.510..109M . doi : 10.1038/nature13400 . ISSN 0028-0836 . PMC 4337882 . PMID 24847885 .
- ^ Пизани, Давид; Петт, Уокер; Дорманн, Мартин; Феуда, Роберто; Рота-Стабелли, Омар; Филипп, Эрве; Лартиллот, Николас; Wörheide, Gert (декабрь 2015 г.). «Геномные данные не поддерживают пробортные желе, как сестринская группа для всех других животных» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (50): 15402–15407. BIBCODE : 2015PNAS..11215402P . doi : 10.1073/pnas.1518127112 . PMC 4687580 . PMID 26621703 .
- ^ Berwald, J. (2017). Беспостол: наука о медузе и искусство выращивания основы . Риверхед книги. [ страница необходима ]
- ^ Ширвотер, Бернд; Эйтель, Майкл; Якоб, Вольфганг; Осигус, Ханс-Юрген; Хэдрис, Хейке; Dellaporta, Stephen L.; Kolokotronis, Sergios-Orestis; Десальле, Роб (январь 2009 г.). «Конкатенированный анализ проливает свет на раннюю эволюцию метазоя и питает современную« гипотезу Urmetazoon » . PLOS Биология . 7 (1): E20. doi : 10.1371/journal.pbio.1000020 . PMC 2631068 . PMID 19175291 .
- ^ Kapli, P.; Телфорд, MJ (декабрь 2020 г.). «Топология-зависимая асимметрия в систематических ошибках влияет на филогенетическое размещение Ctenophora и Xenacoelomorpha» . Наука достижения . 6 (50): EABC5162. Bibcode : 2020scia .... 6.5162K . doi : 10.1126/sciadv.abc5162 . PMC 7732190 . PMID 33310849 .
- ^ Симион, Пол; Филипп, Эрве; Бауэр, Денис; Джагер, Мюриэль; Рихтер, Даниэль Дж.; Ди Франко, Арно; и др. (Апрель 2017). «Большой и последовательный филогеномный набор данных поддерживает губки в качестве родственной группы для всех других животных» (PDF) . Текущая биология . 27 (7): 958–967. Bibcode : 2017cbio ... 27..958s . doi : 10.1016/j.cub.2017.02.031 . PMID 28318975 . Архивировано (PDF) из оригинала 2020-04-25 . Получено 2018-11-04 .
- ^ Редмонд, АК; McLysaght, A. (март 2021 г.). «Свидетельство о губках как сестре для всех других животных от разделенной филогеномики с моделями смесью и перекодированием» . Природная связь . 12 (1): 1783. Bibcode : 2021natco..12.1783r . doi : 10.1038/s41467-021-22074-7 . PMC 7979703 . PMID 33741994 .
- ^ Шульц, Даррин Т.; Хэддок, Стивен HD; Bredeson, Jessen v.; Грин, Ричард Э.; Симаков, Олег; Рохсар, Даниэль С. (июнь 2023 г.). «Древние генные связи поддерживают Ctenophores как сестры для других животных» . Природа . 618 (7963): 110–117. Bibcode : 2023natur.618..110S . doi : 10.1038/s41586-023-05936-6 . PMC 10232365 . PMID 37198475 .
- ^ Смолкер, Рэйчел; Ричардс, Эндрю; Коннор, Ричард; Манн, Джанет; Berggren, per (1997). «Губцовая корни в Индийском океане дельфинов: возможное использование инструмента дельфинида». Этология . 103 (6): 454–465. doi : 10.1111/j.1439-0310.1997.tb00160.x . HDL : 2027.42/71936 .
- ^ McClenachan, L. (2008). «Социальный конфликт, избыточная ловкость и болезнь во Флоридской губке, 1849–1939 гг.» . В Starkey, DJ; Холм, П.; Барнард, М. (ред.). Прошлое океаны: понимание руководства из истории популяций морских животных . Earthscan . С. 25–27. ISBN 978-1-84407-527-0 .
- ^ Джейкобсон, Н. (2000). Расщепление . Рутгерс Университет Пресс. п. 62. ISBN 978-0-8135-2715-4 .
- ^ "Губки" . Общество продвижения шейки матки . 2004. Архивировано из оригинала 14 января 2009 года . Получено 2006-09-17 .
- ^ Портерфилд, Wm (1955). «Лезуа - губка губка». Экономическая ботаника . 9 (3): 211–223. Bibcode : 1955ecbot ... 9..211p . doi : 10.1007/bf02859814 . S2CID 27313678 .
- ^ Имхофф, JF; Stöhr, R. (2003). «Бактерии, связанные с губкой». В Мюллере мы (ред.). Губки (Порифера): Порифера . Спрингер . С. 43–44. ISBN 978-3-540-00968-9 .
- ^ Teyapant, R.; Woerdenbag, HJ; Крейс, П.; Хакер, Дж.; Wray, v.; Witte, L.; Прокш П. (1993). «Антибиотическая и цитотоксическая активность бромированных соединений из морской губки Verongia aerophoba» . Журнал естественных исследований c . 48 (11–12): 939–45. Doi : 10.1515/Znc-1193-11-1218 . PMID 8297426 . S2CID 1593418 .
- ^ Такеучи, Синдзи; Ishibashi, Masami; Kobayashi, Junichi (1994). «Плакоридин А, новый тирамин, содержащий пирролидиновый алкалоид из морской губки Окинавана Плакотис С.П.». Журнал органической химии . 59 (13): 3712–3713. doi : 10.1021/jo00092a039 .
- ^ Etchells, Laura L.; Сардарян, Али; Уайтхед, Роджер С. (18 апреля 2005 г.). «Синтетический подход к плакоридинам, смоделированному на биогенетической теории». Тетраэдр буквы . 46 (16): 2803–2807. doi : 10.1016/j.tetlet.2005.02.124 .
Дальнейшее чтение
- Хикман, С.; Робертс, Л.; Ларсон А. (2003). Разнообразие животных (3 -е изд.). Нью-Йорк: МакГроу-Хилл. ISBN 978-0-07-234903-0 .
- Эресковский, AV (2010). Сравнительная эмбриология губ . Россия: Springer Science+Business Media. ISBN 978-90-481-8575-7 .
- Wörheide, G. (апрель 2008 г.). «Гиперкальцифицированная губка с мягкими родственниками: Valetia - это кератоза -демонгея». Молекулярная филогенетика и эволюция . 47 (1): 433–8. Bibcode : 2008molpe..47..433W . doi : 10.1016/j.ympev.2008.01.021 . PMID 18321733 .
Внешние ссылки
- Поток воды и кормление в филоме Porifera (губки) - флэш -анимация структур корпуса губки, поток воды и кормление
- Губчатка Карстена , информация об экологии и биотехнологический потенциал губок и связанных с ними бактерий.
- История губчатой индустрии Тарпон -Спрингс , Тарпон -Спрингс, Флорида
- Эксперты природы «волокна»
- Проект губки -рифа
- Информация о музее Квинсленда о губках
- Квинслендские музейные коллекции морских беспозвоночных
- Квинслендский музей Сансильские морские беспозвоночные исследования
- Гид Губедж для Британии и Ирландии Архивировал 2008-12-08 в The Wayback Machine , Бернард Пиктон, Кристин Морроу и Роб Ван Соест
- World Porifera Database , Всемирный список существующих губок, включает в себя базу данных для поиска.
- Губки: мировое производство и рынки // Продовольственная и сельскохозяйственная организация
Условные Porifera занятия |
|---|
