КРИСПР
| Каскад (CRISPR-ассоциированный комплекс противовирусной защиты) | |||
|---|---|---|---|
 Белок CRISPR Cascade (голубой), связанный с РНК CRISPR (зеленый) и ДНК фага (красный) [ 1 ] | |||
| Идентификаторы | |||
| Организм | |||
| Символ | КРИСПР | ||
| Входить | 947229 | ||
| ПДБ | 4QYZ | ||
| RefSeq (защита) | НП_417241.1 | ||
| ЮниПрот | P38036 | ||
| |||
| Часть серии о |
| КРИСПР |
|---|
| Редактирование генома : CRISPR-Cas |
|
варианты: Анти-CRISPR - CIRTS - CRISPeYCRISPR-Cas10 - CRISPR-Cas13 - CRISPR-BEST CRISPR-Disp - CRISPR-Gold - CRISPRa - CRISPRi Easi-CRISPR – СДЕЛАЙТЕ ЭТО |
| Фермент |
|
Cas9 - FokI - EcoRI - PstI - SmaI HaeIII – Cas12a (Cpf1) – xCas9 |
| Приложения |
| КАМЕРА - ICE - Направленная генетика |
| другой метод редактирования генома: |
| Прайм-редактирование - Pro-AG - RESCUE - TALEN - ZFN - LEAPER |
| Часть серии о |
| Генная инженерия |
|---|
| Генетически модифицированные организмы |
| История и регулирование |
| Процесс |
| Приложения |
| Споры |
CRISPR ( / ˈ k r ɪ s p ər / ) ( аббревиатура от кластеризованных , коротких регулярно расположенных между собой палиндромных повторов ) таких , — семейство последовательностей ДНК обнаруженных в организмов , геномах прокариотических как бактерии и археи . [ 2 ] Эти последовательности получены из фрагментов ДНК бактериофагов , ранее заразивших прокариот. Их используют для обнаружения и уничтожения ДНК подобных бактериофагов при последующих инфекциях. Следовательно, эти последовательности играют ключевую роль в противовирусной (т.е. антифаговой ) системе защиты прокариот и обеспечивают форму приобретенного иммунитета . [ 2 ] [ 3 ] [ 4 ] [ 5 ] CRISPR обнаружен примерно в 50% секвенированных геномов бактерий и почти в 90% секвенированных архей. [ 6 ]

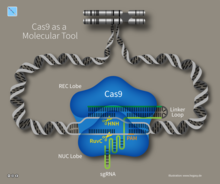
Cas9 (или «CRISPR-ассоциированный белок 9») — это фермент , который использует последовательности CRISPR в качестве руководства для распознавания и открытия определенных цепей ДНК, которые комплементарны последовательности CRISPR. Ферменты Cas9 вместе с последовательностями CRISPR составляют основу технологии, известной как CRISPR-Cas9 , которую можно использовать для редактирования генов в живых организмах. [ 8 ] [ 9 ] Этот процесс редактирования имеет широкий спектр применений, включая фундаментальные биологические исследования, разработку биотехнологических продуктов и лечение заболеваний. [ 10 ] [ 11 ] Разработка метода редактирования генома CRISPR-Cas9 была отмечена Нобелевской премией по химии 2020 года, присужденной Эммануэль Шарпантье и Дженнифер Дудна . [ 12 ] [ 13 ]
История
[ редактировать ]Повторяющиеся последовательности
[ редактировать ]Открытие кластерных повторов ДНК произошло независимо в трех частях света. Первое описание того, что позже будет названо CRISPR, принадлежит из Университета Осаки исследователю Ёсидзуми Исино и его коллегам в 1987 году. Они случайно клонировали часть последовательности CRISPR вместе с геном «iap» (изоферментная конверсия щелочной фосфатазы) из генома эшерихий. палочка [ 14 ] [ 15 ] что было их целью. Организация повторов была необычной. Повторяющиеся последовательности обычно располагаются последовательно, без перемежения разных последовательностей. [ 11 ] [ 15 ] Они не знали функции прерванных кластерных повторов.
В 1993 году исследователи микобактерии туберкулеза в Нидерландах опубликовали две статьи о кластере прерванных прямых повторов (DR) у этой бактерии. Они признали разнообразие последовательностей, которые участвуют в прямых повторах среди разных штаммов M. Tuberculosis. [ 16 ] и использовал это свойство для разработки метода набора текста под названием сполиготипирование , который используется до сих пор. [ 17 ] [ 18 ]
Франсиско Мохика из Университета Аликанте в Испании изучал функцию повторов у архейных видов Haloferax и Haloarcula . Руководитель Мохики предположил, что кластерные повторы играют роль в правильном разделении реплицируемой ДНК на дочерние клетки во время клеточного деления, поскольку плазмиды и хромосомы с идентичными массивами повторов не могли сосуществовать в Haloferax volcanii . Транскрипция прерванных повторов также отмечена впервые; это была первая полная характеристика CRISPR. [ 18 ] [ 19 ] К 2000 году Мохика и его ученики после автоматического поиска опубликованных геномов идентифицировали прерывистые повторы у 20 видов микробов как принадлежащие к одному и тому же семейству. [ 20 ] Поскольку эти последовательности располагались через промежутки, Мохика первоначально назвал эти последовательности «короткими повторами с регулярными интервалами» (SRSR). [ 21 ] В 2001 году Мохика и Рууд Янсен , которые искали дополнительные прерванные повторы, предложили аббревиатуру CRISPR (Clustered Regularly Interspaced Short Palindromic Повторы), чтобы унифицировать многочисленные акронимы, используемые для описания этих последовательностей. [ 19 ] [ 22 ] В 2002 году Тан и др. продемонстрировали доказательства того, что области повторов CRISPR из генома Archaeoglobus fulgidus транскрибируются в длинные молекулы РНК, которые впоследствии обрабатываются в небольшие РНК единичной длины, а также в некоторые более длинные формы из 2, 3 или более единиц спейсерных повторов. [ 23 ] [ 24 ]
В 2005 году йогурта исследователь Родольф Баррангу обнаружил, что Streptococcus thermophilus после повторяющихся фаговых заражений развивает повышенную устойчивость к фагам из-за включения дополнительных спейсерных последовательностей CRISPR. [ 25 ] Работодатель Баррангу, датская пищевая компания Danisco, затем разработала устойчивые к фагам штаммы S. thermophilus для производства йогурта. Позже Danisco была куплена компанией DuPont , которой принадлежит около 50 процентов мирового рынка молочных культур, и технология широко распространилась. [ 26 ]
Системы, связанные с CRISPR
[ редактировать ]Большим прорывом в понимании CRISPR стало наблюдение Янсена о том, что кластер повторов прокариот сопровождается четырьмя гомологичными генами, которые составляют CRISPR-ассоциированные системы, cas 1–4. Белки Cas обнаруживают геликазные и нуклеазные мотивы , что указывает на их роль в динамической структуре локусов CRISPR. [ 27 ] В этой публикации в качестве универсального названия этого паттерна использовалась аббревиатура CRISPR, но его функция осталась загадочной.

В 2005 году три независимые исследовательские группы показали, что некоторые спейсеры CRISPR происходят из фаговой ДНК и внехромосомной ДНК, такой как плазмиды . [ 31 ] [ 32 ] [ 33 ] По сути, спейсеры представляют собой фрагменты ДНК, полученные от вирусов, ранее атаковавших клетку. Источник спейсеров был признаком того, что система CRISPRcas может играть роль в адаптивном иммунитете бактерий . [ 28 ] [ 34 ] Все три исследования, предлагающие эту идею, первоначально были отвергнуты известными журналами, но со временем появились в других журналах. [ 35 ]
Первая публикация [ 32 ] предложив роль CRISPR-Cas в микробном иммунитете, Мохика и его коллеги из Университета Аликанте предсказали роль РНК-транскрипта спейсеров в распознавании мишени в механизме, который может быть аналогичен системе РНК-интерференции, используемой эукариотическими клетками. Кунин и его коллеги расширили эту гипотезу РНК-интерференции, предложив механизмы действия различных подтипов CRISPR-Cas в соответствии с предсказанной функцией их белков. [ 36 ]
Экспериментальная работа нескольких групп выявила основные механизмы иммунитета CRISPR-Cas. В 2007 году были опубликованы первые экспериментальные доказательства того, что CRISPR представляет собой адаптивную иммунную систему. [ 4 ] [ 11 ] Область CRISPR у Streptococcus thermophilus приобрела спейсеры из ДНК инфицирующего бактериофага . Исследователи манипулировали устойчивостью S. thermophilus к различным типам фагов, добавляя и удаляя спейсеры, последовательность которых соответствовала обнаруженной в тестируемых фагах. [ 37 ] [ 38 ] В 2008 году Браунс и Ван дер Ост идентифицировали комплекс белков Cas, названный Cascade, который в E. coli разрезал предшественник РНК CRISPR внутри повторов на зрелые молекулы РНК, содержащие спейсер, называемые CRISPR РНК (crRNA), которые оставались связанными с белком. сложный. [ 39 ] Более того, было обнаружено, что каскад, crRNA и хеликаза/нуклеаза ( Cas3 ) необходимы для обеспечения бактериального хозяина иммунитета против заражения ДНК-вирусом . Разработав антивирусный CRISPR, они продемонстрировали, что две ориентации crRNA (смысловая/антисмысловая) обеспечивают иммунитет, что указывает на то, что направляющие crRNA нацелены на дцДНК . В том же году Марраффини и Сонтхаймер подтвердили, что последовательность CRISPR S. epidermidis нацелена на ДНК, а не на РНК, чтобы предотвратить конъюгацию . Это открытие противоречило предложенному механизму иммунитета CRISPR-Cas, подобному РНК-интерференции, хотя система CRISPR-Cas, нацеленная на чужеродную РНК, была позже обнаружена у Pyrococcus Furiosus . [ 11 ] [ 38 ] Исследование 2010 года показало, что CRISPR-Cas разрезает нити как фаговой, так и плазмидной ДНК у S. thermophilus . [ 40 ]
Красный9
[ редактировать ]Более простая система CRISPR Streptococcus pyogenes основана на белке Cas9 . Cas9 Эндонуклеаза представляет собой четырехкомпонентную систему, включающую две небольшие молекулы: crRNA и трансактивирующую РНК CRISPR (tracrRNA). [ 41 ] [ 42 ] В 2012 году Дженнифер Дудна и Эммануэль Шарпантье модернизировали эндонуклеазу Cas9 в более управляемую двухкомпонентную систему, объединив две молекулы РНК в « РНК с одним проводником », которая в сочетании с Cas9 могла найти и разрезать ДНК-мишень. указанной направляющей РНК. [ 43 ] Этот вклад был настолько значительным, что он был отмечен Нобелевской премией по химии в 2020 году. Манипулируя нуклеотидной последовательностью направляющей РНК, искусственную систему Cas9 можно было запрограммировать для разделения любой последовательности ДНК. [ 43 ] Другое сотрудничество, включающее Вирджиниюса Шикшниса , Гасюнаса, Баррангу и Хорвата, показало, что Cas9 из системы CRISPR S. thermophilus также можно перепрограммировать для нацеливания на выбранный ими сайт путем изменения последовательности его crRNA. Эти достижения стимулировали усилия по редактированию геномов с помощью модифицированной системы CRISPR-Cas9. [ 18 ]
Группы под руководством Фэн Чжана и Джорджа Черча одновременно впервые опубликовали описания редактирования генома в культурах клеток человека с использованием CRISPR-Cas9. [ 11 ] [ 44 ] [ 45 ] С тех пор он использовался для лечения широкого спектра микроорганизмов, включая пекарские дрожжи ( Saccharomyces cerevisiae ), [ 46 ] [ 47 ] [ 48 ] Candida условно-патогенный микроорганизм albicans , [ 49 ] [ 50 ] рыбка данио ( Danio rerio ), [ 51 ] плодовые мушки ( Drosophila melanogaster ), [ 52 ] [ 53 ] муравьи ( Harpegnathos saltator [ 54 ] и Ooceraea biroi [ 55 ] ), комары ( Aedes aegypti [ 56 ] ), нематоды ( Caenorhabditis elegans ), [ 57 ] растения, [ 58 ] мыши ( Mus musculus Domesticus ) , [ 59 ] [ 60 ] обезьяны [ 61 ] и человеческие эмбрионы. [ 62 ]
CRISPR был модифицирован для создания программируемых факторов транскрипции , которые позволяют активировать или подавлять целевые гены. [ 63 ]

Было показано, что система CRISPR-Cas9 эффективно редактирует гены в трехпронуклеарных зиготах человека , как впервые описано в статье 2015 года китайских ученых П. Ляна и Ю. Сюй. Система успешно расщепила мутантный бета-гемоглобин ( HBB ) в 28 из 54 эмбрионов. Четыре из 28 эмбрионов были успешно рекомбинированы с использованием донорской матрицы. Ученые показали, что во время рекомбинации ДНК расщепленной цепи гомологичная эндогенная последовательность HBD конкурирует с экзогенной донорской матрицей. Репарация ДНК в эмбрионах человека гораздо сложнее и специфичнее, чем в производных стволовых клетках. [ 64 ]
Кас12а
[ редактировать ]В 2015 году нуклеаза Cas12a (ранее называвшаяся Cpf1 [ 65 ] ) был охарактеризован в системе CRISPR-Cpf1 бактерии Francesella novicida . [ 66 ] [ 67 ] Его первоначальное название, взятое из TIGRFAMs, определения семейства белков созданного в 2012 году, отражает распространенность его подтипа CRISPR-Cas в линиях Prevotella и Francesella . Cas12a продемонстрировал несколько ключевых отличий от Cas9, в том числе: создание «шахматного» разреза двухцепочечной ДНК в отличие от «тупого» разреза, производимого Cas9, с использованием «T-богатого» PAM (обеспечивающего альтернативные сайты нацеливания для Cas9) и необходимости только РНК CRISPR (crRNA) для успешного нацеливания. Напротив, Cas9 требует как crRNA, так и трансактивирующей crRNA (tracrRNA).
Эти различия могут дать Cas12a некоторые преимущества перед Cas9. Например, небольшие crRNA Cas12a идеально подходят для мультиплексного редактирования генома, поскольку в один вектор можно упаковать больше из них, чем sgRNA Cas9. Липкие 5'-концы, оставленные Cas12a, также можно использовать для сборки ДНК, которая гораздо более специфична для мишени, чем традиционное клонирование ферментов рестрикции. [ 68 ] Наконец, Cas12a расщепляет ДНК на 18–23 пары оснований ниже сайта PAM. Это означает, что после репарации не происходит нарушения последовательности узнавания, и поэтому Cas12a обеспечивает возможность многократного расщепления ДНК. Напротив, поскольку Cas9 разрезает только 3 пары оснований выше сайта PAM, путь NHEJ приводит к индел -мутациям, которые разрушают последовательность узнавания, тем самым предотвращая дальнейшие циклы разрезания. Теоретически, повторные циклы расщепления ДНК должны увеличить вероятность желаемого редактирования генома. [ 69 ] Отличительной особенностью Cas12a по сравнению с Cas9 является то, что после разрезания своей мишени Cas12a остается связанным с мишенью и затем неизбирательно расщепляет другие молекулы оцДНК. [ 70 ] Это свойство называется активностью «побочного расщепления» или «транс-расщепления» и использовалось для разработки различных диагностических технологий. [ 71 ] [ 72 ]
Кас13
[ редактировать ]В 2016 году нуклеаза Cas13a (ранее известный как C2c2 ) из бактерии Leptotrichia shahii . Cas13 представляет собой РНК-ориентированную РНК-эндонуклеазу, что означает, что она не расщепляет ДНК, а только одноцепочечную РНК. Cas13 направляется с помощью crRNA к мишени оцРНК, связывает и расщепляет мишень. Подобно Cas12a, Cas13 остается связанным с мишенью, а затем без разбора расщепляет другие молекулы оцРНК. [ 73 ] Это побочное свойство расщепления было использовано для разработки различных диагностических технологий. [ 74 ] [ 75 ] [ 76 ]
В 2021 году доктор Хуэй Ян охарактеризовал новые миниатюрные варианты белка Cas13 (mCas13): Cas13X и Cas13Y. Использование небольшой части последовательности гена N SARS-CoV-2 в качестве мишени для характеристики mCas13 выявило чувствительность и специфичность mCas13 в сочетании с RT-LAMP для обнаружения SARS-CoV-2 как в синтетических, так и в клинических образцах по сравнению с другими доступные стандартные тесты, такие как RT-qPCR (1 копия/мкл). [ 77 ]
Структура локуса
[ редактировать ]Повторы и проставки
[ редактировать ]Массив CRISPR состоит из богатой АТ лидерной последовательности, за которой следуют короткие повторы, разделенные уникальными спейсерами. [ 78 ] Повторы CRISPR обычно имеют размер от 28 до 37 пар оснований (п.н.), хотя их длина может составлять от 23 до 55 п.н. [ 79 ] Некоторые демонстрируют диадную симметрию , подразумевающую образование вторичной структуры, такой как стебель-петля («шпилька») в РНК, в то время как другие спроектированы так, чтобы быть неструктурированными. Размер спейсеров в разных массивах CRISPR обычно составляет от 32 до 38 п.н. (диапазон от 21 до 72 п.н.). [ 79 ] Новые спейсеры могут быстро появляться как часть иммунного ответа на фаговую инфекцию. [ 80 ] Обычно в массиве CRISPR содержится менее 50 единиц последовательности повтор-спейсер. [ 79 ]
Структуры CRISPR РНК
[ редактировать ]-
 CRISPR-DR2: Вторичная структура взята из базы данных Rfam . Семья RF01315 .
CRISPR-DR2: Вторичная структура взята из базы данных Rfam . Семья RF01315 . -
 CRISPR-DR5: Вторичная структура взята из базы данных Rfam . Семья RF011318 .
CRISPR-DR5: Вторичная структура взята из базы данных Rfam . Семья RF011318 . -
 CRISPR-DR6: Вторичная структура взята из базы данных Rfam . Семья RF01319 .
CRISPR-DR6: Вторичная структура взята из базы данных Rfam . Семья RF01319 . -
 CRISPR-DR8: Вторичная структура взята из базы данных Rfam . Семья RF01321 .
CRISPR-DR8: Вторичная структура взята из базы данных Rfam . Семья RF01321 . -
 CRISPR-DR9: Вторичная структура взята из базы данных Rfam . Семья RF01322 .
CRISPR-DR9: Вторичная структура взята из базы данных Rfam . Семья RF01322 . -
 CRISPR-DR19: Вторичная структура взята из базы данных Rfam . Семья RF01332 .
CRISPR-DR19: Вторичная структура взята из базы данных Rfam . Семья RF01332 . -
 CRISPR-DR41: Вторичная структура взята из базы данных Rfam . Семья RF01350 .
CRISPR-DR41: Вторичная структура взята из базы данных Rfam . Семья RF01350 . -
 CRISPR-DR52: Вторичная структура взята из базы данных Rfam . Семья RF01365 .
CRISPR-DR52: Вторичная структура взята из базы данных Rfam . Семья RF01365 . -
 CRISPR-DR57: Вторичная структура взята из базы данных Rfam . Семья RF01370 .
CRISPR-DR57: Вторичная структура взята из базы данных Rfam . Семья RF01370 . -
 CRISPR-DR65: Вторичная структура взята из базы данных Rfam . Семья RF01378 .
CRISPR-DR65: Вторичная структура взята из базы данных Rfam . Семья RF01378 .










Гены Cas и подтипы CRISPR
[ редактировать ]Небольшие кластеры генов cas часто располагаются рядом с массивами повторов и спейсеров CRISPR. В совокупности 93 гена cas сгруппированы в 35 семейств на основе сходства последовательностей кодируемых белков. 11 из 35 семейств образуют ядро cas , в которое входят семейства белков от Cas1 до Cas9. Полный локус CRISPR-Cas имеет по крайней мере один ген, принадлежащий ядру cas . [ 81 ]
Системы CRISPR-Cas делятся на два класса. Системы класса 1 используют комплекс из нескольких белков Cas для расщепления чужеродных нуклеиновых кислот. Системы класса 2 используют для той же цели один большой белок Cas. Класс 1 делится на типы I, III и IV; 2-й класс делится на типы II, V и VI. [ 82 ] 6 типов систем разделены на 33 подтипа. [ 83 ] Каждый тип и большинство подтипов характеризуются «сигнатурным геном», встречающимся почти исключительно в этой категории. Классификация также основана на составе cas присутствующих генов . Большинство систем CRISPR-Cas содержат белок Cas1. Филогения белков Cas1 в целом соответствует принятой классификационной системе. [ 84 ] но существуют исключения из-за перетасовки модулей. [ 81 ] Многие организмы содержат несколько систем CRISPR-Cas, что позволяет предположить, что они совместимы и могут иметь общие компоненты. [ 85 ] [ 86 ] Спорадическое распространение подтипов CRISPR-Cas позволяет предположить, что система CRISPR-Cas подвержена горизонтальному переносу генов в ходе микробной эволюции .
В этой таблице отсутствует информация о перекрестных ссылках UniProt и InterPro. ( октябрь 2020 г. ) |
| Сорт | Тип Каса | Подтип Cas | Сигнатурный белок | Функция | Ссылка |
|---|---|---|---|---|---|
| 1 | я | — | Кас3 | Одноцепочечная ДНК-нуклеаза (домен HD) и АТФ-зависимая геликаза | [ 87 ] [ 88 ] |
| Я | Кас8а, Кас5 | Cas8 представляет собой субъединицу модуля интерференции, которая важна для нацеливания на вторгающуюся ДНК путем распознавания последовательности PAM . Cas5 необходим для процессинга и стабильности crRNA. | [ 84 ] [ 89 ] | ||
| ИБ | Cas8b | ||||
| IC | Cas8c | ||||
| ИДЕНТИФИКАТОР | Кас10д | содержит домен, гомологичный пальмовому домену полимераз нуклеиновых кислот и нуклеотидциклаз | [ 90 ] [ 91 ] | ||
| ИЕ | Ксе1, Ксе2 | ||||
| ЕСЛИ | Csy1, Csy2, Csy3 | Тип IF-3 участвует в транспозонах, связанных с CRISPR. | [ 84 ] | ||
| ИГ [ Примечание 1 ] | GSU0054 | [ 92 ] | |||
| III | — | Кас10 | Гомолог Cas10d и Cse1. Связывает целевую РНК CRISPR и способствует стабильности интерференционного комплекса. | [ 91 ] [ 93 ] | |
| III-А | Ксм2 | Не определено | [ 84 ] | ||
| III-Б | CMR5 | Не определено | [ 84 ] | ||
| III-С | Кас10 или Csx11 | [ 84 ] [ 93 ] | |||
| III-D | Csx10 | [ 84 ] | |||
| III-E | [ 92 ] | ||||
| III-F | [ 92 ] | ||||
| IV | — | Csf1 | [ 92 ] | ||
| IV-А | [ 92 ] | ||||
| IV-Б | [ 92 ] | ||||
| IV-С | [ 92 ] | ||||
| 2 | II | — | Красный9 | Нуклеазы RuvC и HNH вместе продуцируют DSB , а по отдельности могут производить одноцепочечные разрывы. Обеспечивает приобретение функциональных спейсеров при адаптации. | [ 94 ] [ 95 ] |
| II-А | CSN2 | ДНК-связывающий белок кольцевой формы. Участвует в первичной адаптации в системе CRISPR типа II. | [ 96 ] | ||
| II-Б | Кас4 | Эндонуклеаза, которая работает с cas1 и cas2 для создания спейсерных последовательностей. | [ 97 ] | ||
| II-С | Характеризуется отсутствием Csn2 или Cas4. | [ 98 ] | |||
| V | — | Кас12 | Нуклеаза RuvC. Не хватает ХНХ. | [ 82 ] [ 99 ] | |
| V-A | Кас12а (Cpf1) | Автоматическая обработка активности пре-crRNA для мультиплексной регуляции генов | [ 92 ] [ 100 ] | ||
| V-B | Кас12б (С2с1) | [ 92 ] | |||
| V-C | Кас12с (С2с3) | [ 92 ] | |||
| V-D | Кас12д (CasY) | [ 92 ] | |||
| ВЕ | Кас12е (КасХ) | [ 92 ] | |||
| ВФ | Кас12f (Кас14, C2c10) | [ 92 ] | |||
| V-G | Кас12г | [ 92 ] | |||
| ВХ | Cas12h | [ 92 ] | |||
| V-I | Cas12i | [ 92 ] | |||
| V-K [ Примечание 2 ] | Кас12к (C2c5) | Тип VK вовлечен в CRISPR-ассоциированные транспозоны . | [ 92 ] | ||
| V-U | С2с4, С2с8, С2с9 | [ 92 ] | |||
| МЫ | — | Кас13 | РНК-ориентированная РНКаза | [ 82 ] [ 101 ] | |
| С ПОМОЩЬЮ | Кас13а(C2c2) | [ 92 ] | |||
| VI-Б | Кас13б | [ 92 ] | |||
| ШУТИТЬ | Кас13с | [ 92 ] | |||
| VI-Д | Кас13д | [ 92 ] | |||
| VI-X | Кас13x.1 | РНК-зависимая РНК-полимераза, профилактическое ингибирование РНК-вируса | [ 102 ] | ||
| VI-Y | [ 102 ] |
Механизм
[ редактировать ]
(1) Приобретение начинается с распознавания вторгшейся ДНК с помощью Cas1 и Cas2 и расщепления протоспейсера.
(2) Протоспейсер лигируют с прямым повтором, соседним с лидерной последовательностью, и
(3) удлинение одной цепи восстанавливает CRISPR и дублирует прямой повтор. Стадии обработки и интерференции crRNA происходят по-разному в каждой из трех основных систем CRISPR.
(4) Первичный транскрипт CRISPR расщепляется генами cas с образованием crRNA.
(5) В системах типа I Cas6e/Cas6f расщепляются в месте соединения оцРНК и дцРНК, образованных шпильками в прямом повторе. В системах типа II используется трансактивирующая (tracr) РНК для образования дцРНК, которая расщепляется Cas9 и РНКазой III. В системах типа III используется гомолог Cas6, который не требует наличия шпилек в прямом повторе для расщепления.
(6) В системах типа II и типа III вторичная обрезка выполняется либо на 5'-, либо на 3'-конце для получения зрелых crRNA.
(7) Зрелые crRNA связываются с белками Cas, образуя интерференционные комплексы.
(8) В системах типа I и типа II взаимодействие между белком и последовательностью PAM необходимо для деградации вторгающейся ДНК. Системы типа III не требуют PAM для успешной деградации, а в системах типа III-A спаривание оснований происходит между crRNA и мРНК, а не между ДНК, на которую нацелены системы типа III-B.




Иммунитет CRISPR-Cas — это естественный процесс бактерий и архей. [ 103 ] CRISPR-Cas предотвращает заражение бактериофагом, его конъюгацию и естественную трансформацию путем разрушения чужеродных нуклеиновых кислот, попадающих в клетку. [ 38 ]
Приобретение проставки
[ редактировать ]Когда микроб подвергается вторжению бактериофага , первым этапом иммунного ответа является захват ДНК фага и вставка ее в локус CRISPR в виде спейсера. Cas1 и Cas2 обнаружены в обоих типах иммунных систем CRISPR-Cas, что указывает на их участие в приобретении спейсера. Исследования мутаций подтвердили эту гипотезу, показав, что удаление Cas1 или Cas2 останавливает приобретение спейсера, не влияя на иммунный ответ CRISPR. [ 104 ] [ 105 ] [ 106 ] [ 107 ] [ 108 ]
Охарактеризовано множество белков Cas1 и решены их структуры. [ 109 ] [ 110 ] [ 111 ] Белки Cas1 имеют разнообразные аминокислотные последовательности. Однако их кристаллические структуры схожи, и все очищенные белки Cas1 представляют собой металлозависимые нуклеазы/ интегразы , которые связываются с ДНК независимым от последовательности образом. [ 85 ] Репрезентативные белки Cas2 были охарактеризованы и обладают либо (одноцепочечными) оцРНК- [ 112 ] или (двухцепочечная) дцДНК- [ 113 ] [ 114 ] специфическая эндорибонуклеазная активность.
В системе IE E. coli Cas1 и Cas2 образуют комплекс, в котором димер Cas2 соединяет два димера Cas1. [ 115 ] В этом комплексе Cas2 выполняет роль неферментативного каркаса. [ 115 ] связывание двухцепочечных фрагментов вторгшейся ДНК, тогда как Cas1 связывает одноцепочечные фланги ДНК и катализирует их интеграцию в массивы CRISPR. [ 116 ] [ 117 ] [ 118 ] Новые спейсеры обычно добавляются в начале CRISPR рядом с лидерной последовательностью, создавая хронологическую запись вирусных инфекций. [ 119 ] В E. coli называемый гистонподобный белок, фактором хозяина интеграции ( IHF ), который связывается с лидерной последовательностью. за точность этой интеграции отвечает [ 120 ] IHF также повышает эффективность интеграции в системе типа IF Pectobacterium atrosepticum . [ 121 ] но в других системах могут потребоваться другие факторы хоста [ 122 ]
Смежные мотивы протоспейсера (PAM)
[ редактировать ]Биоинформатический анализ областей фаговых геномов, которые были вырезаны в качестве спейсеров (называемых протоспейсерами), показал, что они не были выбраны случайным образом, а вместо этого были обнаружены рядом с короткими (3–5 п.о.) последовательностями ДНК, называемыми мотивами, прилегающими к протоспейсерам (PAM). Анализ систем CRISPR-Cas показал, что PAM важны для систем типа I и типа II, но не для систем типа III во время сбора данных. [ 33 ] [ 123 ] [ 124 ] [ 125 ] [ 126 ] [ 127 ] В системах типа I и типа II протоспейсеры вырезаются в позициях, прилегающих к последовательности PAM, при этом другой конец спейсера обрезается с помощью механизма линейки, что позволяет поддерживать регулярность размера спейсера в массиве CRISPR. [ 128 ] [ 129 ] Консервативность последовательности PAM различается в разных системах CRISPR-Cas и, по-видимому, эволюционно связана с Cas1 и лидерной последовательностью . [ 127 ] [ 130 ]
Новые спейсеры добавляются в массив CRISPR направленным образом. [ 31 ] происходит преимущественно, [ 80 ] [ 123 ] [ 124 ] [ 131 ] [ 132 ] но не исключительно, прилегающие [ 126 ] [ 129 ] к лидерской последовательности. Анализ системы типа IE из E. coli показал, что первый прямой повтор, соседний с лидерной последовательностью, копируется, при этом вновь приобретенный спейсер вставляется между первым и вторым прямыми повторами. [ 107 ] [ 128 ]
Последовательность PAM, по-видимому, важна при вставке спейсера в системах типа IE. Эта последовательность содержит сильно консервативный конечный нуклеотид (нт), соседний с первым нуклеотидом протоспейсера. Эта нт становится последней основой в первом прямом повторе. [ 108 ] [ 133 ] [ 134 ] Это предполагает, что механизм получения спейсера генерирует одноцепочечные выступы в предпоследнем положении прямого повтора и в PAM во время вставки спейсера. Однако, по-видимому, не все системы CRISPR-Cas разделяют этот механизм, поскольку PAM в других организмах не демонстрируют такой же уровень консервации в конечном положении. [ 130 ] Вполне вероятно, что в этих системах тупой конец образуется на самом конце прямого повтора и протоспейсера во время приобретения.
Варианты вставки
[ редактировать ]Анализ CRISPR Sulfolobus solfataricus выявил дополнительные сложности канонической модели вставки спейсеров, поскольку один из шести локусов CRISPR встраивал новые спейсеры случайным образом по всему массиву CRISPR, а не вставлял их ближе всего к лидерной последовательности. [ 129 ]
Множественные CRISPR содержат множество спейсеров к одному и тому же фагу. Механизм, вызывающий это явление, был открыт в системе типа IE E. coli . Значительное улучшение приобретения спейсеров было обнаружено там, где спейсеры уже нацелены на фаг, даже если они не соответствуют протоспейсеру. Этот «прайминг» требует, чтобы белки Cas, участвующие как в захвате, так и в интерференции, взаимодействовали друг с другом. Вновь приобретенные спейсеры, возникающие в результате механизма прайминга, всегда обнаруживаются на той же цепи, что и прайминг-спейсер. [ 108 ] [ 133 ] [ 134 ] Это наблюдение привело к гипотезе, что механизм сбора скользит по чужеродной ДНК после прайминга, чтобы найти новый протоспейсер. [ 134 ]
Биогенез
[ редактировать ]CRISPR-РНК (crRNA), которая позже направляет нуклеазу Cas к мишени на этапе интерференции, должна быть сгенерирована из последовательности CRISPR. Первоначально crРНК транскрибируется как часть одного длинного транскрипта, охватывающего большую часть массива CRISPR. [ 29 ] Этот транскрипт затем расщепляется белками Cas с образованием crRNA. Механизм производства crRNA различается в разных системах CRISPR-Cas. В системах типа IE и типа IF белки Cas6e и Cas6f соответственно распознают стеблевые петли. [ 135 ] [ 136 ] [ 137 ] создается путем спаривания идентичных повторов, фланкирующих crRNA. [ 138 ] Эти белки Cas расщепляют более длинный транскрипт на краю парной области, оставляя одну crRNA вместе с небольшим остатком парной области повтора.
Системы типа III также используют Cas6, однако их повторы не создают стебель-петли. Вместо этого расщепление происходит за счет обертывания более длинного транскрипта вокруг Cas6, что позволяет расщеплять непосредственно перед повторяющейся последовательностью. [ 139 ] [ 140 ] [ 141 ]
В системах типа II отсутствует ген Cas6, и вместо этого для расщепления используется РНКаза III. Системы функционального типа II кодируют очень малую РНК, которая комплементарна повторяющейся последовательности, известную как транс-активирующая crРНК (tracrRNA). [ 41 ] Транскрипция tracrRNA и первичного транскрипта CRISPR приводит к спариванию оснований и образованию дцРНК в повторяющейся последовательности, которая впоследствии подвергается воздействию РНКазы III для производства crРНК. В отличие от двух других систем, crRNA не содержит полного спейсера, который вместо этого усечен на одном конце. [ 94 ]
CrRNAs связываются с белками Cas, образуя рибонуклеотидные комплексы, которые узнают чужеродные нуклеиновые кислоты. CrRNA не демонстрируют предпочтения между кодирующими и некодирующими цепями, что указывает на систему нацеливания на ДНК, управляемую РНК. [ 5 ] [ 40 ] [ 104 ] [ 108 ] [ 142 ] [ 143 ] [ 144 ] Комплекс типа IE (обычно называемый каскадом) требует пяти белков Cas, связанных с одной crRNA. [ 145 ] [ 146 ]
Помехи
[ редактировать ]На стадии интерференции в системах типа I последовательность PAM распознается на цепи, комплементарной crRNA, и требуется наряду с отжигом crRNA. В системах типа I правильное спаривание оснований между crRNA и протоспейсером сигнализирует о конформационном изменении в Cascade, которое привлекает Cas3 для деградации ДНК.
Системы типа II полагаются на один многофункциональный белок Cas9 на этапе интерференции. [ 94 ] Cas9 требует, чтобы как crRNA, так и tracrRNA функционировали и расщепляли ДНК, используя свои двойные HNH и RuvC/RNaseH-подобные эндонуклеазные домены. Спаривание оснований между PAM и геномом фага необходимо в системах типа II. Однако PAM распознается на той же цепи, что и crRNA (противоположная цепь системам типа I).
Системы типа III, как и типа I, требуют связывания шести или семи белков Cas с crRNA. [ 147 ] [ 148 ] Системы типа III, проанализированные на S. Solfataricus и P. Furiosus, нацелены на мРНК фагов, а не на геном фаговой ДНК. [ 86 ] [ 148 ] что может сделать эти системы уникальными способными воздействовать на геномы фагов на основе РНК. [ 85 ] Также было обнаружено, что системы типа III нацелены на ДНК в дополнение к РНК, используя в комплексе другой белок Cas, Cas10. [ 149 ] Было показано, что расщепление ДНК зависит от транскрипции. [ 150 ]
Механизм различения своей ДНК от чужеродной во время интерференции встроен в crRNA и поэтому, вероятно, является общим для всех трех систем. На протяжении отличительного процесса созревания каждого основного типа все crRNA содержат спейсерную последовательность и некоторую часть повтора на одном или обоих концах. Именно частичная повторяющаяся последовательность не позволяет системе CRISPR-Cas нацеливаться на хромосому, поскольку спаривание оснований помимо спейсерной последовательности сигнализирует о себе, и предотвращает расщепление ДНК. [ 151 ] Ферменты CRISPR, управляемые РНК, классифицируются как ферменты рестрикции типа V.
Эволюция
[ редактировать ]| CRISPR-ассоциированный белок Cas2 (адаптационная РНКаза) | |||
|---|---|---|---|
 Кристаллическая структура гипотетического белка tt1823 Thermus thermophilus. | |||
| Идентификаторы | |||
| Символ | CRISPR_Cas2 | ||
| Пфам | PF09827 | ||
| ИнтерПро | ИПР019199 | ||
| CDD | cd09638 | ||
| |||
| CRISPR-ассоциированный белок CasA/Cse1 (эффекторная ДНКаза типа I) | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | CRISPR_Cse1 | ||
| Пфам | PF09481 | ||
| ИнтерПро | ИПР013381 | ||
| CDD | cd09729 | ||
| |||
| CRISPR-ассоциированный белок CasC/Cse3/Cas6 (эффекторная РНКаза типа I) | |||
|---|---|---|---|
 Кристаллическая структура хруст-ассоциированного белка Thermus thermophilus | |||
| Идентификаторы | |||
| Символ | CRISPR_assoc | ||
| Пфам | PF08798 | ||
| Пфам Клан | CL0362 | ||
| ИнтерПро | ИПР010179 | ||
| CDD | cd09727 | ||
| |||
Считается, что гены cas в адаптерном и эффекторном модулях системы CRISPR-Cas произошли от двух разных предковых модулей. Транспозон -подобный элемент , называемый капозоном, кодирующий Cas1-подобную интегразу и, возможно, другие компоненты адаптационного модуля, был вставлен рядом с наследственным эффекторным модулем, который, вероятно, функционировал как независимая врожденная иммунная система. [ 152 ] Высококонсервативные гены cas1 и cas2 адаптерного модуля произошли от предкового модуля, в то время как различные эффекторные cas гены класса 1 произошли от предкового эффекторного модуля. [ 153 ] Эволюция этих различных генов cas эффекторных модулей класса 1 направлялась различными механизмами, такими как события дупликации. [ 154 ] С другой стороны, каждый тип эффекторного модуля класса 2 возник в результате последующих независимых вставок мобильных генетических элементов. [ 155 ] Эти мобильные генетические элементы заменили множество эффекторных модулей генов для создания эффекторных модулей с одним геном, которые производят большие белки, выполняющие все необходимые задачи эффекторного модуля. [ 155 ] Спейсерные области систем CRISPR-Cas взяты непосредственно из чужеродных мобильных генетических элементов, поэтому их долгосрочную эволюцию трудно проследить. [ 156 ] Было обнаружено, что неслучайная эволюция этих спейсерных областей сильно зависит от окружающей среды и конкретных чужеродных мобильных генетических элементов, которые она содержит. [ 157 ]
CRISPR-Cas может иммунизировать бактерии против определенных фагов и тем самым остановить передачу инфекции. По этой причине Кунин описал CRISPR-Cas как ламарковский механизм наследования. [ 158 ] Однако это было оспорено критиком, который отметил: «Мы должны помнить [Ламарка] за то добро, которое он внес в науку, а не за вещи, которые лишь внешне напоминают его теорию. и элегантный способ, которым действительно работает эволюция». [ 159 ] Но по мере проведения более поздних исследований стало очевидно, что приобретенные спейсерные области систем CRISPR-Cas действительно являются формой ламарковской эволюции, поскольку они представляют собой генетические мутации, которые приобретаются, а затем передаются дальше. [ 160 ] С другой стороны, эволюция генного механизма Cas, обеспечивающего работу системы, происходит посредством классической дарвиновской эволюции. [ 160 ]
Коэволюция
[ редактировать ]Анализ последовательностей CRISPR выявил коэволюцию генома хозяина и вируса. [ 161 ]
Базовая модель эволюции CRISPR — это новые спейсеры, которые заставляют фаги мутировать свои геномы, чтобы избежать бактериального иммунного ответа, создавая разнообразие как в популяциях фагов, так и в популяциях хозяев. Чтобы противостоять фаговой инфекции, последовательность спейсера CRISPR должна идеально соответствовать последовательности гена целевого фага. Фаги могут продолжать заражать своих хозяев заданными точечными мутациями в спейсере. [ 151 ] Аналогичная строгость требуется и при ПАМ, иначе бактериальный штамм останется чувствительным к фагам. [ 124 ] [ 151 ]
Цены
[ редактировать ]Исследование 124 штаммов S. thermophilus показало, что 26% всех спейсеров были уникальными и что разные локусы CRISPR демонстрировали разную скорость приобретения спейсеров. [ 123 ] Некоторые локусы CRISPR развиваются быстрее, чем другие, что позволило определить филогенетические взаимоотношения штаммов. Сравнительный геномный анализ показал, что E. coli и S. enterica эволюционируют гораздо медленнее, чем S. thermophilus . Штаммы последнего, разошедшиеся 250 000 лет назад, все еще содержали тот же спейсерный комплемент. [ 162 ]
Метагеномный анализ двух биопленок с кислотно-минным дренажем показал, что один из проанализированных CRISPR содержал обширные делеции и добавления спейсеров по сравнению с другой биопленкой, что указывает на более высокую активность/распространенность фагов в одном сообществе, чем в другом. [ 80 ] В полости рта временное исследование показало, что 7–22% спейсеров использовались совместно в течение 17 месяцев внутри одного человека, тогда как менее 2% были общими для разных людей. [ 132 ]
В той же среде один штамм был отслежен с использованием праймеров ПЦР, специфичных для его системы CRISPR. Результаты общего уровня присутствия/отсутствия спейсера показали значительное разнообразие. Однако в этот CRISPR за 17 месяцев были добавлены три спейсера. [ 132 ] предполагая, что даже в среде со значительным разнообразием CRISPR некоторые локусы эволюционируют медленно.
CRISPR были проанализированы на основе метагеномов, полученных для проекта «Микробиом человека» . [ 163 ] Хотя большинство из них были специфичны для конкретного участка тела, некоторые из них широко распространены среди людей. Один из этих локусов происходил от видов стрептококков и содержал около 15 000 спейсеров, 50% из которых были уникальными. Подобно целевым исследованиям полости рта, некоторые из них показали небольшую эволюцию с течением времени. [ 163 ]
Эволюцию CRISPR изучали в хемостатах с использованием S. thermophilus для непосредственного изучения скорости приобретения спейсеров. За одну неделю штаммы S. thermophilus приобретали до трех спейсеров при заражении одним фагом. [ 164 ] В течение того же периода времени у фага развились однонуклеотидные полиморфизмы , которые зафиксировались в популяции, что позволяет предположить, что нацеливание предотвратило репликацию фага в отсутствие этих мутаций. [ 164 ]
Другой эксперимент с S. thermophilus показал, что фаги могут инфицировать и реплицироваться в хозяевах, имеющих только один целевой спейсер. Еще один показал, что чувствительные хозяева могут существовать в средах с высокими титрами фагов. [ 165 ] Хемостатические и наблюдательные исследования указывают на множество нюансов CRISPR и (ко)эволюции фагов.
Идентификация
[ редактировать ]CRISPR широко распространены среди бактерий и архей. [ 90 ] и показать некоторые сходства последовательностей. [ 138 ] Их наиболее примечательной характеристикой являются повторяющиеся прокладки и прямые повторы. Эта характеристика позволяет легко идентифицировать CRISPR в длинных последовательностях ДНК, поскольку количество повторов снижает вероятность ложноположительного совпадения. [ 166 ]
Анализ CRISPR в метагеномных данных является более сложным, поскольку локусы CRISPR обычно не собираются из-за их повторяющегося характера или из-за вариаций штаммов, что сбивает с толку алгоритмы сборки. Если доступно много эталонных геномов, полимеразную цепную реакцию (ПЦР). для амплификации массивов CRISPR и анализа содержания спейсеров можно использовать [ 123 ] [ 132 ] [ 167 ] [ 168 ] [ 169 ] [ 170 ] Однако этот подход дает информацию только о специально нацеленных CRISPR и об организмах, достаточно представленных в общедоступных базах данных для разработки надежных полимеразных праймеров для ПЦР. Вырожденные праймеры, специфичные для повторов, можно использовать для амплификации спейсеров CRISPR непосредственно из образцов окружающей среды; ампликоны, содержащие два или три спейсера, можно затем собрать с помощью вычислений для реконструкции длинных массивов CRISPR. [ 170 ]
Альтернативой является извлечение и реконструкция массивов CRISPR на основе метагеномных данных. Это сложнее с вычислительной точки зрения, особенно при использовании технологий секвенирования второго поколения (например, 454, Illumina), поскольку короткая длина считывания предотвращает появление более двух или трех повторяющихся единиц в одном считывании. Идентификация CRISPR в необработанных чтениях была достигнута с использованием чисто de novo. идентификации [ 171 ] или с использованием прямых повторяющихся последовательностей в частично собранных массивах CRISPR из контигов (перекрывающихся сегментов ДНК, которые вместе представляют собой консенсусную область ДНК) [ 163 ] и прямые повторяющиеся последовательности из опубликованных геномов [ 172 ] в качестве крючка для выявления прямых повторов в отдельных прочтениях.
Использование фагами
[ редактировать ]Другой способ защиты бактерий от фаговой инфекции — наличие хромосомных островков . Подтип хромосомных островков, называемый индуцируемым фагом хромосомным островом (PICI), вырезается из бактериальной хромосомы при фаговой инфекции и может ингибировать репликацию фага. [ 173 ] PICI индуцируются, вырезаются, реплицируются и, наконец, упаковываются в небольшие капсиды определенными стафилококковыми умеренными фагами. PICI используют несколько механизмов для блокирования размножения фагов. В первом механизме Ppi, кодируемый PICI, дифференциально блокирует созревание фага путем связывания или специфического взаимодействия с фагом TerS, тем самым блокируя образование комплекса фага TerS/TerL, ответственного за упаковку фаговой ДНК. Во втором механизме PICI CpmAB перенаправляет морфогенетический белок фагового капсида, создавая 95% капсида размером с SaPI, и ДНК фага может упаковывать только 1/3 их генома в эти маленькие капсиды и, следовательно, становиться нежизнеспособным фагом. [ 174 ] Третий механизм включает два белка, PtiA и PtiB, которые нацелены на LtrC, который отвечает за выработку вириона и белков лизиса. Этот механизм интерференции модулируется модулирующим белком PtiM, который связывается с одним из белков, опосредующих интерференцию, PtiA и, следовательно, достигает необходимого уровня интерференции. [ 175 ]
Одно исследование показало, что литический фаг ICP1, специально нацеленный на Vibrio cholerae серогруппы O1, приобрел систему CRISPR-Cas, которая нацелена на PICI-подобный элемент V. cholerae . Система имеет 2 локуса CRISPR и 9 генов Cas. Кажется, она гомологична системе IF, обнаруженной у Yersinia pestis . Более того, как и бактериальная система CRISPR-Cas, ICP1 CRISPR-Cas может приобретать новые последовательности, что позволяет фагу и хозяину развиваться совместно. [ 176 ] [ 177 ]
Было показано, что некоторые архейные вирусы несут массивы мини-CRISPR, содержащие один или два спейсера. Было показано, что спейсеры в вирусных массивах CRISPR нацелены на другие вирусы и плазмиды, что позволяет предположить, что мини-чипы CRISPR представляют собой механизм исключения гетеротипической суперинфекции и участвуют в межвирусных конфликтах. [ 170 ]
Приложения
[ редактировать ]Редактирование генов CRISPR — это революционная технология, которая позволяет точно и целенаправленно модифицировать ДНК живых организмов. CRISPR-Cas9, разработанный на основе естественного защитного механизма, обнаруженного у бактерий, является наиболее часто используемой системой, которая позволяет «разрезать» ДНК в определенных местах и удалять, модифицировать или вставлять генетический материал. Эта технология изменила такие области, как генетика, медицина, [ 178 ] [ 179 ] и сельское хозяйство, [ 180 ] [ 181 ] предлагая потенциальные методы лечения генетических нарушений, достижения в области растениеводства и исследования фундаментальных механизмов жизни. Однако его этические последствия и потенциальные непредвиденные последствия вызвали серьезные споры. [ 182 ] [ 183 ]
См. также
[ редактировать ]- Активация CRISPR
- Анти-CRISPR
- Инструменты CRISPR/Cas
- Редактирование генов CRISPR
- Журнал CRISPR
- « Дизайнерский малыш »
- ДРАКО
- Джин нокаут
- Полногеномный нокаут CRISPR-Cas9
- Глоссарий генетики
- Инженерия зародышевой линии человека
- Человеческая природа (документальный фильм, 2019)
- МАГЕСТИК
- Новая евгеника
- Премьер-редактирование
- РНКи
- миРНК
- Нуклеазный анализ Surveyor
- Синтетическая биология
- Цинковый палец
Примечания
[ редактировать ]Ссылки
[ редактировать ]- ^ ВВП : 4QYZ : Мулепати С., Эру А., Бэйли С. (2014). «Кристаллическая структура комплекса наблюдения, управляемого РНК CRISPR, связанного с мишенью оцДНК» . Наука . 345 (6203): 1479–1484. Бибкод : 2014Sci...345.1479M . дои : 10.1126/science.1256996 . ПМЦ 4427192 . ПМИД 25123481 .
- ^ Jump up to: а б Баррангу Р. (2015). «Роль систем CRISPR-Cas в адаптивном иммунитете и не только». Современное мнение в иммунологии . 32 : 36–41. дои : 10.1016/j.coi.2014.12.008 . ПМИД 25574773 .
- ^ Редман М., Кинг А., Уотсон С., Кинг Д. (август 2016 г.). «Что такое CRISPR/Cas9?» . Архив детских болезней: издание для образования и практики . 101 (4): 213–215. дои : 10.1136/archdischild-2016-310459 . ПМЦ 4975809 . ПМИД 27059283 .
- ^ Jump up to: а б Баррангу Р. , Фремо С., Дево Х., Ричардс М., Бояваль П., Муано С. и др. (март 2007 г.). «CRISPR обеспечивает приобретенную устойчивость прокариот к вирусам». Наука . 315 (5819): 1709–1712. Бибкод : 2007Sci...315.1709B . дои : 10.1126/science.1138140 . hdl : 20.500.11794/38902 . ПМИД 17379808 . S2CID 3888761 . ( требуется регистрация )
- ^ Jump up to: а б Марраффини Л.А., Сонтхаймер Э.Дж. (декабрь 2008 г.). «Интерференция CRISPR ограничивает горизонтальный перенос генов у стафилококков путем воздействия на ДНК» . Наука . 322 (5909): 1843–1845. Бибкод : 2008Sci...322.1843M . дои : 10.1126/science.1165771 . ПМК 2695655 . ПМИД 19095942 .
- ^ Хилле Ф., Рихтер Х., Вонг С.П., Братович М., Рессель С., Шарпантье Э. (март 2018 г.). «Биология CRISPR-Cas: вперед и назад». Клетка . 172 (6): 1239–1259. дои : 10.1016/j.cell.2017.11.032 . hdl : 21.11116/0000-0003-FC0D-4 . ПМИД 29522745 . S2CID 3777503 .
- ^ Хорват П., Баррангу Р. (январь 2010 г.). «CRISPR/Cas, иммунная система бактерий и архей». Наука . 327 (5962): 167–170. Бибкод : 2010Sci...327..167H . дои : 10.1126/science.1179555 . ПМИД 20056882 . S2CID 17960960 .
- ^ Бак Р.О., Гомес-Оспина Н., Портеус М.Х. (август 2018 г.). «Редактирование генов в центре внимания». Тенденции в генетике . 34 (8): 600–611. дои : 10.1016/j.tig.2018.05.004 . ПМИД 29908711 . S2CID 49269023 .
- ^ Чжан Ф, Вэнь Ю, Го Икс (2014). «CRISPR/Cas9 для редактирования генома: прогресс, последствия и проблемы» . Молекулярная генетика человека . 23 (Р1): Р40–6. дои : 10.1093/hmg/ddu125 . ПМИД 24651067 . S2CID 19008095 .
- ^ CRISPR-CAS9, TALENS и ZFNS – битва в редактировании генов https://www.ptglab.com/news/blog/crispr-cas9-talens-and-zfns-the-battle-in-gene-editing/ Архивировано в 2021 г. -05-25 в Wayback Machine
- ^ Jump up to: а б с д и Сюй П.Д., Ландер Э.С., Чжан Ф. (июнь 2014 г.). «Разработка и применение CRISPR-Cas9 для генной инженерии» . Клетка . 157 (6): 1262–1278. дои : 10.1016/j.cell.2014.05.010 . ПМЦ 4343198 . ПМИД 24906146 .
- ^ «Пресс-релиз: Нобелевская премия по химии 2020» . Нобелевский фонд. Архивировано из оригинала 15 января 2021 года . Проверено 7 октября 2020 г.
- ^ Ву К.Дж., Пельтье Э. (7 октября 2020 г.). «Нобелевская премия по химии присуждена двум ученым за работу по редактированию генома — Эммануэль Шарпантье и Дженнифер А. Дудна разработали инструмент Crispr, который может с высокой точностью изменять ДНК животных, растений и микроорганизмов» . Нью-Йорк Таймс . Архивировано из оригинала 8 октября 2020 года . Проверено 7 октября 2020 г.
- ^ Рават А., Рой М., Джьоти А., Кошик С., Верма К., Шривастава В.К. (август 2021 г.). «Цистеиновые протеазы: борьба с патогенными паразитическими простейшими с помощью вездесущих ферментов» . Микробиологические исследования . 249 : 126784. doi : 10.1016/j.micres.2021.126784 . ПМИД 33989978 . S2CID 234597200 .
- ^ Jump up to: а б Исино Ю, Синагава Х, Макино К, Амемура М, Наката А (декабрь 1987 г.). «Нуклеотидная последовательность гена iap, ответственного за превращение изофермента щелочной фосфатазы в Escherichia coli, и идентификация продукта гена» . Журнал бактериологии . 169 (12): 5429–5433. дои : 10.1128/jb.169.12.5429-5433.1987 . ПМК 213968 . ПМИД 3316184 .
- ^ ван Соолинген Д., де Хаас П.Е., Херманс П.В., Гроенен П.М., ван Эмбден Дж.Д. (август 1993 г.). «Сравнение различных повторяющихся элементов ДНК как генетических маркеров штаммовой дифференциации и эпидемиологии микобактерий туберкулеза» . Журнал клинической микробиологии . 31 (8): 1987–1995. doi : 10.1128/JCM.31.8.1987-1995.1993 . ПМК 265684 . ПМИД 7690367 .
- ^ Гроенен П.М., Буншотен А.Е., ван Соолинген Д., ван Эмбден Дж.Д. (декабрь 1993 г.). «Природа полиморфизма ДНК в кластере прямых повторов микобактерии туберкулеза; применение для дифференциации штаммов новым методом типирования». Молекулярная микробиология . 10 (5): 1057–1065. дои : 10.1111/j.1365-2958.1993.tb00976.x . ПМИД 7934856 . S2CID 25304723 .
- ^ Jump up to: а б с Мохика Ф.Дж., Монтолиу Л. (2016). «О происхождении технологии CRISPR-Cas: от прокариот к млекопитающим». Тенденции в микробиологии . 24 (10): 811–820. дои : 10.1016/j.tim.2016.06.005 . ПМИД 27401123 .
- ^ Jump up to: а б Мохика Ф.Дж., Родригес-Валера Ф. (2016). «Открытие CRISPR у архей и бактерий» (PDF) . Журнал ФЭБС . 283 (17): 3162–3169. дои : 10.1111/февраль 13766 . hdl : 10045/57676 . ПМИД 27234458 . S2CID 42827598 . Архивировано (PDF) из оригинала 25 февраля 2021 г. Проверено 29 августа 2019 г.
- ^ Мохика Ф.Дж., Диес-Вильясеньор К., Сориа Э., Хуэс Дж. (апрель 2000 г.). «Биологическое значение семейства регулярно расположенных повторов в геномах архей, бактерий и митохондрий» . Молекулярная микробиология . 36 (1): 244–246. дои : 10.1046/j.1365-2958.2000.01838.x . ПМИД 10760181 .
- ^ Исааксон В. (2021). Взломщик кодов: Дженнифер Дудна, редактирование генов и будущее человечества . Нью-Йорк: Саймон и Шустер. п. 73. ИСБН 978-1-9821-1585-2 . OCLC 1239982737 . Архивировано из оригинала 14 января 2023 г. Проверено 20 октября 2021 г.
- ^ Баррангу Р. , ван дер Ост Дж. (2013). Системы CRISPR-Cas: РНК-опосредованный адаптивный иммунитет у бактерий и архей . Гейдельберг: Спрингер. п. 6. ISBN 978-3-642-34656-9 .
- ^ Тан Т.Х., Бачеллери Дж.П., Рождественский Т., Бортолин М.Л., Хубер Х., Друнговски М. и др. (май 2002 г.). «Идентификация 86 кандидатов на малые непосредственные РНК из археи Archaeoglobus fulgidus» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (11): 7536–7541. Бибкод : 2002PNAS...99.7536T . дои : 10.1073/pnas.112047299 . ПМК 124276 . ПМИД 12032318 .
- ^ Шарпантье Э., Рихтер Х., ван дер Ост Дж., Уайт М.Ф. (май 2015 г.). «Пути биогенеза РНК-направляющих в адаптивном иммунитете CRISPR-Cas архей и бактерий» . Обзоры микробиологии FEMS . 39 (3): 428–441. дои : 10.1093/femsre/fuv023 . ПМЦ 5965381 . ПМИД 25994611 .
- ^ Ромеро Д.А., Мэгилл Д., Миллен А., Хорват П., Фремо К. (ноябрь 2020 г.). «Взаимодействие молочного лактококкового и стрептококкового фага с хозяином: промышленная перспектива в развивающемся фаговом ландшафте». Обзоры микробиологии FEMS . 44 (6): 909–932. дои : 10.1093/femsre/fuaa048 . ПМИД 33016324 .
- ^ Молтени М., Хакинс Дж. (1 августа 2020 г.). «Руководство WIRED по Crispr» . Конде Наст. Проводной журнал. Архивировано из оригинала 23 октября 2021 года . Проверено 23 февраля 2021 г.
- ^ Янсен Р., Эмбден Дж. Д., Гаастра В., Шоулс Л. М. (март 2002 г.). «Идентификация генов, связанных с повторами ДНК у прокариот» . Молекулярная микробиология . 43 (6): 1565–1575. дои : 10.1046/j.1365-2958.2002.02839.x . ПМИД 11952905 . S2CID 23196085 .
- ^ Jump up to: а б Хорват П., Баррангу Р. (январь 2010 г.). «CRISPR/Cas, иммунная система бактерий и архей». Наука . 327 (5962): 167–170. Бибкод : 2010Sci...327..167H . дои : 10.1126/Science.1179555 . ПМИД 20056882 . S2CID 17960960 .
- ^ Jump up to: а б Марраффини Л.А., Сонтхаймер Э.Дж. (март 2010 г.). «CRISPR-интерференция: РНК-направленный адаптивный иммунитет у бактерий и архей» . Обзоры природы Генетика . 11 (3): 181–190. дои : 10.1038/nrg2749 . ПМЦ 2928866 . ПМИД 20125085 .
- ^ Грисса I, Верно Дж., Пурсель К. (май 2007 г.). «База данных CRISPRdb и инструменты для отображения CRISPR и создания словарей спейсеров и повторов» . БМК Биоинформатика . 8 : 172. дои : 10.1186/1471-2105-8-172 . ПМК 1892036 . ПМИД 17521438 .
- ^ Jump up to: а б Пурсель К., Сальвиньоль Дж., Верно Дж. (март 2005 г.). «Элементы CRISPR в Yersinia pestis приобретают новые повторы за счет преимущественного поглощения ДНК бактериофага и предоставляют дополнительные инструменты для эволюционных исследований» . Микробиология . 151 (Часть 3): 653–663. дои : 10.1099/mic.0.27437-0 . ПМИД 15758212 .
- ^ Jump up to: а б Мохика Ф.Х., Диес-Вильясеньор К., Гарсиа-Мартинес Х., Сория Э. (февраль 2005 г.). «Промежуточные последовательности прокариотических повторов с регулярными интервалами происходят от чужеродных генетических элементов». Журнал молекулярной эволюции . 60 (2): 174–182. Бибкод : 2005JMolE..60..174M . дои : 10.1007/s00239-004-0046-3 . ПМИД 15791728 . S2CID 27481111 .
- ^ Jump up to: а б Болотин А, Квинкис Б, Сорокин А, Эрлих С.Д. (август 2005 г.). «Кластерные короткие палиндромные повторы с регулярными интервалами (CRISPR) имеют спейсеры внехромосомного происхождения» . Микробиология . 151 (Часть 8): 2551–2561. дои : 10.1099/mic.0.28048-0 . ПМИД 16079334 .
- ^ Моранж М. (июнь 2015 г.). «Что говорит нам история XXXVII. CRISPR-Cas: открытие иммунной системы у прокариот» (PDF) . Журнал биологических наук . 40 (2): 221–223. дои : 10.1007/s12038-015-9532-6 . ПМИД 25963251 .
- ^ Лендер ES (январь 2016 г.). «Герои CRISPR» . Клетка . 164 (1–2): 18–28. дои : 10.1016/j.cell.2015.12.041 . ПМИД 26771483 .
- ^ Макарова К.С., Гришин Н.В., Шабалина С.А., Вольф Ю.И., Кунин Е.В. (март 2006 г.). «Предполагаемая иммунная система прокариот, основанная на РНК-интерференции: вычислительный анализ предсказанного ферментативного механизма, функциональные аналогии с эукариотическими РНКи и гипотетические механизмы действия» . Биология Директ . 1 :7. дои : 10.1186/1745-6150-1-7 . ПМК 1462988 . ПМИД 16545108 .
- ^ Пенниси Э. (август 2013 г.). «Увлечение CRISPR». Новости Фокус. Наука . 341 (6148): 833–836. Бибкод : 2013Sci...341..833P . дои : 10.1126/science.341.6148.833 . ПМИД 23970676 .
- ^ Jump up to: а б с Марраффини, Лос-Анджелес (октябрь 2015 г.). «CRISPR-Cas иммунитет у прокариот». Природа . 526 (7571): 55–61. Бибкод : 2015Природа.526...55М . дои : 10.1038/nature15386 . ПМИД 26432244 . S2CID 3718361 .
- ^ Браунс С.Дж., Джор М.М., Лундгрен М., Вестра Э.Р., Слиджхуис Р.Дж., Снейдерс А.П. и др. (август 2008 г.). «Маленькие РНК CRISPR обеспечивают противовирусную защиту прокариот» . Наука . 321 (5891): 960–964. Бибкод : 2008Sci...321..960B . дои : 10.1126/science.1159689 . ПМЦ 5898235 . ПМИД 18703739 .
- ^ Jump up to: а б Гарно Дж.Э., Дюпюи М., Виллион М., Ромеро Д.А., Баррангу Р. , Бояваль П. и др. (ноябрь 2010 г.). «Бактериальная иммунная система CRISPR/Cas расщепляет ДНК бактериофага и плазмиды». Природа 468 (7320): 67–71. Бибкод : 2010Природа.468...67Г . CiteSeerX 10.1.1.451.9645 . дои : 10.1038/nature09523 . ПМИД 21048762 . S2CID 205222849 .
- ^ Jump up to: а б Дельчева Е., Чилински К., Шарма С.М., Гонсалес К., Чао Ю., Пирзада З.А. и др. (март 2011 г.). «Созревание РНК CRISPR с помощью транскодируемой малой РНК и фактора хозяина РНКазы III» . Природа . 471 (7340): 602–607. Бибкод : 2011Natur.471..602D . дои : 10.1038/nature09886 . ПМК 3070239 . ПМИД 21455174 .
- ^ Баррангу Р. (ноябрь 2015 г.). «Разнообразие CRISPR-Cas иммунных систем и молекулярных машин» . Геномная биология . 16 : 247. дои : 10.1186/s13059-015-0816-9 . ПМЦ 4638107 . ПМИД 26549499 .
- ^ Jump up to: а б Джинек М., Чилински К., Фонфара И., Хауэр М., Дудна Дж.А. , Шарпантье Э. (август 2012 г.). «Программируемая ДНК-эндонуклеаза, управляемая двойной РНК, в адаптивном бактериальном иммунитете» . Наука . 337 (6096): 816–821. Бибкод : 2012Sci...337..816J . дои : 10.1126/science.1225829 . ПМК 6286148 . ПМИД 22745249 .
- ^ Конг Л., Ран Ф.А., Кокс Д., Лин С., Барретто Р., Хабиб Н. и др. (февраль 2013 г.). «Мультиплексная геномная инженерия с использованием систем CRISPR/Cas» . Наука . 339 (6121): 819–823. Бибкод : 2013Sci...339..819C . дои : 10.1126/science.1231143 . ПМЦ 3795411 . ПМИД 23287718 .
- ^ Мали П., Ян Л., Эсвелт К.М., Аах Дж., Гуэлл М., ДиКарло Дж.Э. и др. (февраль 2013 г.). «Инженерия генома человека под контролем РНК с помощью Cas9» . Наука . 339 (6121): 823–826. Бибкод : 2013Sci...339..823M . дои : 10.1126/science.1232033 . ПМЦ 3712628 . ПМИД 23287722 .
- ^ ДиКарло Дж.Э., Норвилл Дж.Э., Мали П., Риос Х, Аах Дж., Черч ГМ (апрель 2013 г.). «Геномная инженерия Saccharomyces cerevisiae с использованием систем CRISPR-Cas» . Исследования нуклеиновых кислот . 41 (7): 4336–4343. дои : 10.1093/нар/gkt135 . ПМЦ 3627607 . ПМИД 23460208 .
- ^ Чжан Г.К., Конг II, Ким Х, Лю Дж.Дж., Кейт Дж.Х., Джин Ю.С. (декабрь 2014 г.). «Создание четверного ауксотрофного мутанта промышленного полиплоидного штамма saccharomyces cerevisiae с использованием РНК-ориентированной нуклеазы Cas9» . Прикладная и экологическая микробиология . 80 (24): 7694–7701. Бибкод : 2014ApEnM..80.7694Z . дои : 10.1128/АЕМ.02310-14 . ПМЦ 4249234 . ПМИД 25281382 .
- ^ Лю Дж.Дж., Конг II, Чжан Г.К., Джаякоди Л.Н., Ким Х., Ся П.Ф. и др. (апрель 2016 г.). «Метаболическая инженерия пробиотиков Saccharomyces boulardii » . Прикладная и экологическая микробиология . 82 (8): 2280–2287. Бибкод : 2016ApEnM..82.2280L . дои : 10.1128/AEM.00057-16 . ПМЦ 4959471 . ПМИД 26850302 .
- ^ Вяс В.К., Барраса М.И., Финк Г.Р. (2015). « Система CRISPR Candida albicans позволяет осуществлять генную инженерию важнейших генов и семейств генов» . Достижения науки . 1 (3): e1500248. Бибкод : 2015SciA....1E0248V . дои : 10.1126/sciadv.1500248 . ПМЦ 4428347 . ПМИД 25977940 .
- ^ Нг Х, Дин Н (2017). « Candida albicans за счет увеличения экспрессии единой направляющей РНК» . мСфера . 2 (2): e00385–16. дои : 10.1128/mSphere.00385-16 . ПМЦ 5397569 . ПМИД 28435892 .
- ^ Хван В.Ю., Фу Ю., Рейон Д., Мэдер М.Л., Цай С.К., Сандер Дж.Д. и др. (март 2013 г.). «Эффективное редактирование генома рыбок данио с использованием системы CRISPR-Cas» . Природная биотехнология . 31 (3): 227–229. дои : 10.1038/nbt.2501 . ПМЦ 3686313 . ПМИД 23360964 .
- ^ Грац С.Дж., Каммингс А.М., Нгуен Дж.Н., Хэмм Д.К., Донохью Л.К., Харрисон М.М. и др. (август 2013 г.). «Геномная инженерия дрозофилы с помощью нуклеазы Cas9, управляемой РНК CRISPR» . Генетика . 194 (4): 1029–1035. дои : 10.1534/генетика.113.152710 . ПМЦ 3730909 . ПМИД 23709638 .
- ^ Бассетт А.Р., Тиббит С., Понтинг К.П., Лю Дж.Л. (июль 2013 г.). «Высокоэффективный таргетный мутагенез дрозофилы с помощью системы CRISPR/Cas9» . Отчеты по ячейкам . 4 (1): 220–228. дои : 10.1016/j.celrep.2013.06.020 . ПМЦ 3714591 . ПМИД 23827738 .
- ^ Ян Х., Опахалоэмфан С., Манчини Г., Ян Х., Галлитто М., Млейнек Дж. и др. (август 2017 г.). «Инженерная мутация орко приводит к аберрантному социальному поведению и дефектному развитию нервной системы у муравьев» . Клетка . 170 (4): 736–747.e9. дои : 10.1016/j.cell.2017.06.051 . ПМЦ 5587193 . ПМИД 28802043 .
- ^ Трайбл В., Оливос-Сиснерос Л., Маккензи С.К., Сарагости Дж., Чанг Н.К., Мэтьюз Б.Дж. и др. (август 2017 г.). «Мутагенез орко вызывает потерю клубочков антеннальных долей и нарушение социального поведения у муравьев» . Клетка . 170 (4): 727–735.e10. дои : 10.1016/j.cell.2017.07.001 . ПМК 5556950 . ПМИД 28802042 .
- ^ Кистлер К.Е., Восшалл Л.Б., Мэтьюз Б.Дж. (апрель 2015 г.). «Геномная инженерия с использованием CRISPR-Cas9 у комара Aedes aegypti» . Отчеты по ячейкам . 11 (1): 51–60. дои : 10.1016/j.celrep.2015.03.009 . ПМК 4394034 . ПМИД 25818303 .
- ^ Фридланд А.Е., Цур Ю.Б., Эсвелт К.М., депутат Колаяково, генеральный директор Черча, Каларко Х.А. (август 2013 г.). «Наследственное редактирование генома C. elegans с помощью системы CRISPR-Cas9» . Природные методы . 10 (8): 741–743. дои : 10.1038/nmeth.2532 . ПМЦ 3822328 . ПМИД 23817069 .
- ^ Цзян В., Чжоу Х., Би Х., Фромм М., Ян Б., Уикс Д.П. (ноябрь 2013 г.). «Демонстрация целевой модификации генов, опосредованной CRISPR/Cas9/sgRNA, у арабидопсиса, табака, сорго и риса» . Исследования нуклеиновых кислот . 41 (20): e188. дои : 10.1093/nar/gkt780 . ПМЦ 3814374 . ПМИД 23999092 .
- ^ Ван Х., Ян Х., Шивалила К.С., Давлати М.М., Ченг А.В., Чжан Ф. и др. (май 2013 г.). «Одноэтапное создание мышей, несущих мутации в нескольких генах, с помощью геномной инженерии, опосредованной CRISPR/Cas» . Клетка . 153 (4): 910–918. дои : 10.1016/j.cell.2013.04.025 . ПМЦ 3969854 . ПМИД 23643243 .
- ^ Сони Д., Ван Д.М., Регми С.К., Миттал М., Фогель С.М., Шлютер Д. и др. (май 2018 г.). «Дебиквитиназная функция А20 поддерживает и восстанавливает эндотелиальный барьер после повреждения сосудов легких» . Открытие клеточной смерти . 4 (60): 60. дои : 10.1038/s41420-018-0056-3 . ПМЦ 5955943 . ПМИД 29796309 .
- ^ Го X, Ли XJ (июль 2015 г.). «Направленное редактирование генома эмбрионов приматов» . Клеточные исследования . 25 (7): 767–768. дои : 10.1038/cr.2015.64 . ПМЦ 4493275 . ПМИД 26032266 .
- ^ Балтимор Д., Берг П., Ботчан М., Кэрролл Д., Чаро Р.А., Чёрч Дж. и др. (апрель 2015 г.). «Биотехнология. Разумный путь вперед в области геномной инженерии и модификации генов зародышевой линии» . Наука . 348 (6230): 36–38. Бибкод : 2015Sci...348...36B . дои : 10.1126/science.aab1028 . ПМЦ 4394183 . ПМИД 25791083 .
- ^ Ларсон М.Х., Гилберт Л.А., Ван X, Лим В.А., Вайсман Дж.С., Ци Л.С. (ноябрь 2013 г.). «CRISPR-интерференция (CRISPRi) для последовательно-специфического контроля экспрессии генов» . Протоколы природы . 8 (11): 2180–2196. дои : 10.1038/nprot.2013.132 . ПМЦ 3922765 . ПМИД 24136345 .
- ^ Лян П., Сюй Ю, Чжан Х, Дин С., Хуан Р., Чжан З. и др. (май 2015 г.). «CRISPR/Cas9-опосредованное редактирование генов в трехпронуклеарных зиготах человека» . Белок и клетка . 6 (5): 363–372. дои : 10.1007/s13238-015-0153-5 . ПМЦ 4417674 . ПМИД 25894090 .
- ^ Yan MY, Yan HQ, Ren GX, Zhao JP, Guo XP, Sun YC (сентябрь 2017 г.). «Рекомбинация с помощью CRISPR-Cas12a в бактериях» . Прикладная и экологическая микробиология . 83 (17). Бибкод : 2017ApEnM..83E.947Y . дои : 10.1128/AEM.00947-17 . ПМК 5561284 . ПМИД 28646112 .
- ^ Зетше Б., Гутенберг Дж.С., Абудайе О.О., Слеймейкер И.М., Макарова К.С., Эсслетцбихлер П. и др. (октябрь 2015 г.). «Cpf1 представляет собой единственную РНК-ориентированную эндонуклеазу системы CRISPR-Cas класса 2» . Клетка . 163 (3): 759–771. дои : 10.1016/j.cell.2015.09.038 . ПМЦ 4638220 . ПМИД 26422227 .
- ^ Фонфара И, Рихтер Х, Братович М, Ле Рун А, Шарпантье Э (апрель 2016 г.). «Связанный с CRISPR фермент, расщепляющий ДНК Cpf1, также обрабатывает РНК-предшественник CRISPR». Природа . 532 (7600): 517–521. Бибкод : 2016Natur.532..517F . дои : 10.1038/nature17945 . ПМИД 27096362 . S2CID 2271552 .
- ^ Ким Х, Ким С.Т., Рю Дж., Кан Б.С., Ким Дж.С. и Ким С.Г. (февраль 2017 г.). «Редактирование генома растений без ДНК, опосредованное CRISPR / Cpf1» . Природные коммуникации . 8 (14406): 14406. Бибкод : 2017NatCo...814406K . дои : 10.1038/ncomms14406 . ПМК 5316869 . ПМИД 28205546 .
- ^ «Нуклеаза Cpf1» . abmgood.com . Архивировано из оригинала 23 октября 2021 г. Проверено 14 декабря 2017 г.
- ^ Чен Дж.С., Ма Э., Харрингтон Л.Б., Да Коста М., Тянь Х., Палефски Дж.М. и др. (апрель 2018 г.). «Связывание CRISPR-Cas12a с мишенью высвобождает неизбирательную активность одноцепочечной ДНКазы» . Наука . 360 (6387): 436–439. Бибкод : 2018Sci...360..436C . дои : 10.1126/science.aar6245 . ПМК 6628903 . ПМИД 29449511 .
- ^ Бротон Дж.П., Денг Х., Ю.Г., Фашинг К.Л., Сервеллита В., Сингх Дж. и др. (июль 2020 г.). «Обнаружение SARS-CoV-2 на основе CRISPR-Cas12» . Природная биотехнология . 38 (7): 870–874. дои : 10.1038/s41587-020-0513-4 . ПМЦ 9107629 . ПМИД 32300245 .
- ^ Нгуен Л.Т., Смит Б.М., Джайн П.К. (сентябрь 2020 г.). «Усиление активности транс-расщепления Cas12a с помощью сконструированной crRNA позволяет обнаруживать усиленные нуклеиновые кислоты» . Природные коммуникации . 11 (1): 4906. Бибкод : 2020NatCo..11.4906N . дои : 10.1038/s41467-020-18615-1 . ПМЦ 7528031 . ПМИД 32999292 .
- ^ Абудайе О.О., Гутенберг Дж.С., Конерманн С., Йонг Дж., Слеймейкер И.М., Кокс Д.Б. и др. (август 2016 г.). «C2c2 представляет собой однокомпонентный программируемый РНК-направляемый эффектор CRISPR, нацеленный на РНК» . Наука . 353 (6299): aaf5573. doi : 10.1126/science.aaf5573 . ПМК 5127784 . ПМИД 27256883 .
- ^ Гутенберг Дж.С., Абудайе О.О., Ли Дж.В., Эсслетцбихлер П., Ди А.Дж., Йонг Дж. и др. (апрель 2017 г.). «Обнаружение нуклеиновой кислоты с помощью CRISPR-Cas13a/C2c2» . Наука . 356 (6336): 438–442. Бибкод : 2017Sci...356..438G . дои : 10.1126/science.aam9321 . ПМК 5526198 . ПМИД 28408723 .
- ^ Гутенберг Дж.С., Абудайе О.О., Келлнер М.Дж., Йонг Дж., Коллинз Дж.Дж., Чжан Ф. (апрель 2018 г.). «Мультиплексная и портативная платформа для обнаружения нуклеиновых кислот с Cas13, Cas12a и Csm6» . Наука . 360 (6387): 439–444. Бибкод : 2018Sci...360..439G . дои : 10.1126/science.aaq0179 . ПМЦ 5961727 . ПМИД 29449508 .
- ^ Ивасаки Р.С., Бати RT (сентябрь 2020 г.). «СПРИНТ: платформа на основе Cas13a для обнаружения малых молекул» . Исследования нуклеиновых кислот . 48 (17): е101. дои : 10.1093/nar/gkaa673 . ПМЦ 7515716 . ПМИД 32797156 .
- ^ Махас А., Ван Ц, Марсик Т., Махфуз М.М. (октябрь 2021 г.). «Новая миниатюрная система CRISPR-Cas13 для диагностики SARS-CoV-2» . ACS Синтетическая биология . 10 (10): 2541–2551. doi : 10.1021/acsynbio.1c00181 . ПМЦ 8482783 . ПМИД 34546709 .
- ^ Хилле Ф., Шарпантье Э (ноябрь 2016 г.). «CRISPR-Cas: биология, механизмы и актуальность» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 371 (1707): 20150496. doi : 10.1098/rstb.2015.0496 . ПМК 5052741 . ПМИД 27672148 .
- ^ Jump up to: а б с Баррангу Р. , Марраффини Л.А. (апрель 2014 г.). «Системы CRISPR-Cas: переход прокариот к адаптивному иммунитету» . Молекулярная клетка . 54 (2): 234–244. doi : 10.1016/j.molcel.2014.03.011 . ПМК 4025954 . ПМИД 24766887 .
- ^ Jump up to: а б с Тайсон Г.В., Банфилд Дж.Ф. (январь 2008 г.). «Быстро развивающиеся CRISPR, участвующие в приобретенной устойчивости микроорганизмов к вирусам». Экологическая микробиология . 10 (1): 200–207. Бибкод : 2008EnvMi..10..200T . дои : 10.1111/j.1462-2920.2007.01444.x . ПМИД 17894817 .
- ^ Jump up to: а б Кунин Е.В., Макарова К.С. (май 2019). «Происхождение и эволюция систем CRISPR-Cas» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 374 : 20180087.doi : (1772 ) 10.1098/rstb.2018.0087 . ПМК 6452270 . ПМИД 30905284 .
- ^ Jump up to: а б с Райт А.В., Нуньес Х.К., Дудна Х.А. (январь 2016 г.). «Биология и применение систем CRISPR: использование природного инструментария для генной инженерии» . Клетка . 164 (1–2): 29–44. дои : 10.1016/j.cell.2015.12.035 . ПМИД 26771484 .
- ^ Макарова К.С., Вольф Ю.И., Иранзо Дж., Шмаков С.А., Алхнбаши О.С., Браунс С.Дж. и др. (декабрь 2019 г.). «Эволюционная классификация систем CRISPR–Cas: взрыв класса 2 и производные варианты» . Обзоры природы Микробиология . 18 (1): 67–83. дои : 10.1038/s41579-019-0299-x . hdl : 10045/102627 . ПМЦ 8905525 . ПМИД 31857715 .
- ^ Jump up to: а б с д и ж г час Макарова К.С., Вольф Ю.И., Алхнбаши О.С., Коста Ф., Шах С.А., Сондерс С.Дж. и др. (ноябрь 2015 г.). «Обновленная эволюционная классификация систем CRISPR-Cas» . Обзоры природы. Микробиология . 13 (11): 722–736. дои : 10.1038/nrmicro3569 . ПМК 5426118 . ПМИД 26411297 .
- ^ Jump up to: а б с Виденхефт Б., Штернберг Ш., Дудна Дж.А. (февраль 2012 г.). «Системы генетического молчания, управляемые РНК, у бактерий и архей». Природа . 482 (7385): 331–338. Бибкод : 2012Natur.482..331W . дои : 10.1038/nature10886 . ПМИД 22337052 . S2CID 205227944 .
- ^ Jump up to: а б Дэн Л., Гарретт Р.А., Шах С.А., Пэн X, Ше Q (март 2013 г.). «Новый механизм взаимодействия с помощью модуля CRISPR-Cmr типа IIIB в Sulfolobus». Молекулярная микробиология . 87 (5): 1088–1099. дои : 10.1111/mmi.12152 . ПМИД 23320564 .
- ^ Синкунас Т., Гасюнас Г., Фремо С., Баррангу Р. , Хорват П., Сикснис В. (апрель 2011 г.). «Cas3 представляет собой одноцепочечную ДНК-нуклеазу и АТФ-зависимую геликазу в иммунной системе CRISPR/Cas» . Журнал ЭМБО . 30 (7): 1335–1342. дои : 10.1038/emboj.2011.41 . ПМК 3094125 . ПМИД 21343909 .
- ^ Хуо Ю, Нам К.Х., Дин Ф., Ли Х., Ву Л., Сяо Ю. и др. (сентябрь 2014 г.). «Структуры CRISPR Cas3 предлагают механистическое понимание каскадно-активируемого раскручивания и деградации ДНК» . Структурная и молекулярная биология природы . 21 (9): 771–777. дои : 10.1038/nsmb.2875 . ПМК 4156918 . ПМИД 25132177 .
- ^ Брендель Дж., Столл Б., Ланге С.Дж., Шарма К., Ленц С., Стахлер А.Е. и др. (март 2014 г.). «Комплекс белков Cas 5, 6 и 7 необходим для биогенеза и стабильности сгруппированных регулярно расположенных коротких палиндромных повторов (crispr), производных РНК (crrnas) в Haloferax volcanii» . Журнал биологической химии . 289 (10): 7164–77. дои : 10.1074/jbc.M113.508184 . ПМЦ 3945376 . ПМИД 24459147 .
- ^ Jump up to: а б Чилинский К., Макарова К.С., Шарпантье Э., Кунин Е.В. (июнь 2014 г.). «Классификация и эволюция систем CRISPR-Cas типа II» . Исследования нуклеиновых кислот . 42 (10): 6091–6105. дои : 10.1093/nar/gku241 . ПМК 4041416 . ПМИД 24728998 .
- ^ Jump up to: а б Макарова К.С., Аравинд Л., Вольф Ю.И., Кунин Е.В. (июль 2011 г.). «Объединение семейств белков Cas и простой сценарий возникновения и эволюции систем CRISPR-Cas» . Биология Директ . 6:38 . дои : 10.1186/1745-6150-6-38 . ПМК 3150331 . ПМИД 21756346 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В Макарова К.С., Вольф Ю.И., Иранзо Дж., Шмаков С.А., Алхнбаши О.С., Браунс С.Дж. и др. (февраль 2020 г.). «Эволюционная классификация систем CRISPR-Cas: взрыв класса 2 и производные варианты» . Обзоры природы. Микробиология . 18 (2): 67–83. дои : 10.1038/s41579-019-0299-x . hdl : 10045/102627 . ПМЦ 8905525 . ПМИД 31857715 . S2CID 209420490 .
- ^ Jump up to: а б Могила И, Казлаускене М, Валинските С, Тамулатиене Г, Тамулайтис Г, Сикснис В (март 2019 г.). «Генетическое исследование комплекса CSM системы CRISPR-Cas типа III-A выявляет роль отдельных субъединиц» . Отчеты по ячейкам . 26 (10): 2753–2765.e4. дои : 10.1016/j.celrep.2019.02.029 . ПМИД 30840895 .
- ^ Jump up to: а б с Гасюнас Г., Баррангу Р. , Хорват П., Сикснис В. (сентябрь 2012 г.). «Рибонуклеопротеиновый комплекс Cas9-crRNA опосредует специфическое расщепление ДНК для адаптивного иммунитета у бактерий» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (39): E2579–2586. Бибкод : 2012PNAS..109E2579G . дои : 10.1073/pnas.1208507109 . ПМЦ 3465414 . ПМИД 22949671 .
- ^ Хелер Р., Самай П., Моделл Дж.В., Вайнер С., Голдберг Г.В., Бикард Д. и др. (март 2015 г.). «Cas9 определяет функциональные вирусные мишени во время адаптации CRISPR-Cas» . Природа . 519 (7542): 199–202. Бибкод : 2015Natur.519..199H . дои : 10.1038/nature14245 . ПМЦ 4385744 . ПМИД 25707807 .
- ^ Нам К.Х., Куринов И., Ке А. (сентябрь 2011 г.). «Кристаллическая структура кластеризованных коротких палиндромных повторов с регулярными промежутками (CRISPR), ассоциированных с белком Csn2, выявила Ca2+-зависимую активность связывания двухцепочечной ДНК» . Журнал биологической химии . 286 (35): 30759–30768. дои : 10.1074/jbc.M111.256263 . ПМК 3162437 . ПМИД 21697083 .
- ^ Ли Х., Дхингра Ю., Сашитал Д.Г. (апрель 2019 г.). «Комплекс Cas4-Cas1-Cas2 обеспечивает точную обработку преспейсера во время адаптации CRISPR» . электронная жизнь . 8 . doi : 10.7554/eLife.44248 . ПМК 6519985 . ПМИД 31021314 .
- ^ Чилински К., Ле Рун А., Шарпантье Э. (май 2013 г.). «Семейства tracrRNA и Cas9 систем иммунитета CRISPR-Cas типа II» . Биология РНК . 10 (5): 726–737. дои : 10.4161/rna.24321 . ПМЦ 3737331 . ПМИД 23563642 .
- ^ Макарова К.С., Чжан Ф., Кунин Е.В. (январь 2017). «SnapShot: системы CRISPR-Cas класса 2» . Клетка . 168 (1–2): 328–328.e1. дои : 10.1016/j.cell.2016.12.038 . ПМИД 28086097 .
- ^ Пол Б., Монтойя Дж. (февраль 2020 г.). «CRISPR-Cas12a: Функциональный обзор и приложения» . Биомедицинский журнал . 43 (1): 8–17. дои : 10.1016/j.bj.2019.10.005 . ПМК 7090318 . ПМИД 32200959 .
- ^ Кокс Д.Б., Гутенберг Дж.С., Абудайе О.О., Франклин Б., Келлнер М.Дж., Йонг Дж. и др. (ноябрь 2017 г.). «Редактирование РНК с помощью CRISPR-Cas13» . Наука . 358 (6366): 1019–1027. Бибкод : 2017Sci...358.1019C . дои : 10.1126/science.aaq0180 . ПМЦ 5793859 . ПМИД 29070703 .
- ^ Jump up to: а б Сюй С, Чжоу Ю, Сяо Q, Хэ Б, Гэн Г, Ван Цз и др. (май 2021 г.). «Программируемое редактирование РНК с помощью компактных систем CRISPR-Cas13 из некультивируемых микробов» (PDF) . Природные методы . 18 (5): 499–506. дои : 10.1038/s41592-021-01124-4 . ПМИД 33941935 . S2CID 233719501 . Архивировано (PDF) из оригинала 12 мая 2024 г. Проверено 12 мая 2024 г.
- ^ Азангу-Хьяви М., Гасеми М., Ханали Дж., Бороманд-Сабур М., Джамалха М., Сулеймани М. и др. (2020). «CRISPR/Cas: от редактирования опухолевых генов к Т-клеточной иммунотерапии рака» . Границы в иммунологии . 11 : 2062. дои : 10.3389/fimmu.2020.02062 . ПМЦ 7553049 . ПМИД 33117331 .
- ^ Jump up to: а б Алияри Р., Дин С.В. (январь 2009 г.). «Вирусный иммунитет на основе РНК, инициируемый семейством иммунных рецепторов хозяина Dicer» . Иммунологические обзоры . 227 (1): 176–188. дои : 10.1111/j.1600-065X.2008.00722.x . ПМК 2676720 . ПМИД 19120484 .
- ^ Дугар Г., Хербиг А., Фёрстнер К.Ю., Хайдрих Н., Рейнхардт Р., Низельт К. и др. (май 2013 г.). «Карты транскриптома высокого разрешения выявляют штаммоспецифичные регуляторные особенности нескольких изолятов Campylobacter jejuni» . ПЛОС Генетика . 9 (5): e1003495. дои : 10.1371/journal.pgen.1003495 . ПМЦ 3656092 . ПМИД 23696746 .
- ^ Хатум-Аслан А., Манив И., Марраффини Л.А. (декабрь 2011 г.). «Длина зрелых кластеризованных, регулярно расположенных коротких палиндромных повторов РНК (crRNA) измеряется с помощью механизма линейки, закрепленного на сайте процессинга предшественника» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (52): 21218–21222. Бибкод : 2011PNAS..10821218H . дои : 10.1073/pnas.1112832108 . ПМК 3248500 . ПМИД 22160698 .
- ^ Jump up to: а б Йосеф I, Горен М.Г., Кимрон У (июль 2012 г.). «Белки и элементы ДНК, необходимые для процесса адаптации CRISPR у Escherichia coli » . Исследования нуклеиновых кислот . 40 (12): 5569–5576. дои : 10.1093/nar/gks216 . ПМЦ 3384332 . ПМИД 22402487 .
- ^ Jump up to: а б с д Свартс, округ Колумбия, Мостерд С., ван Пассел М.В., Браунс С.Дж. (2012). «Вмешательство CRISPR направляет приобретение спейсера, специфичного для цепи» . ПЛОС ОДИН . 7 (4): e35888. Бибкод : 2012PLoSO...735888S . дои : 10.1371/journal.pone.0035888 . ПМЦ 3338789 . ПМИД 22558257 .
- ^ Бабу М. , Белоглазова Н., Флик Р., Грэм С., Скарина Т., Ночек Б. и др. (январь 2011 г.). «Двойная функция системы CRISPR-Cas: обеспечение бактериального антивирусного иммунитета и восстановление ДНК» . Молекулярная микробиология . 79 (2): 484–502. дои : 10.1111/j.1365-2958.2010.07465.x . ПМК 3071548 . ПМИД 21219465 .
- ^ Хан Д., Леманн К., Краусс Г. (июнь 2009 г.). «SSO1450 - белок CAS1 из Sulfolobus solfataricus P2 с высоким сродством к РНК и ДНК». Письма ФЭБС . 583 (12): 1928–1932. Бибкод : 2009FEBSL.583.1928H . дои : 10.1016/j.febslet.2009.04.047 . ПМИД 19427858 . S2CID 22279972 .
- ^ Виденхефт Б., Чжоу К., Джинек М., Койл С.М., Ма В., Дудна Дж.А. (июнь 2009 г.). «Структурная основа ДНКазной активности консервативного белка, участвующего в CRISPR-опосредованной защите генома» . Структура . 17 (6): 904–912. дои : 10.1016/j.str.2009.03.019 . ПМИД 19523907 .
- ^ Белоглазова Н., Браун Г., Циммерман М.Д., Праудфут М., Макарова К.С., Кудрицка М. и др. (июль 2008 г.). «Новое семейство эндорибонуклеаз, специфичных для последовательностей, связанных с кластеризованными короткими палиндромными повторами с регулярными промежутками» . Журнал биологической химии . 283 (29): 20361–20371. дои : 10.1074/jbc.M803225200 . ПМЦ 2459268 . ПМИД 18482976 .
- ^ Самай П., Смит П., Шуман С. (декабрь 2010 г.). «Структура CRISPR-ассоциированного белка Cas2 из Desulfovibrio vulgaris» . Acta Crystallographica Раздел F. 66 (Часть 12): 1552–1556. дои : 10.1107/S1744309110039801 . ПМК 2998353 . ПМИД 21139194 .
- ^ Нам К.Х., Дин Ф., Хайтджема С., Хуан К., ДеЛиса М.П., Ке А. (октябрь 2012 г.). «Двухцепочечная эндонуклеазная активность в белке Cas2, связанном с регулярно расположенными короткими палиндромными повторами (CRISPR) в Bacillus halodurans» . Журнал биологической химии . 287 (43): 35943–35952. дои : 10.1074/jbc.M112.382598 . ПМЦ 3476262 . ПМИД 22942283 .
- ^ Jump up to: а б Нуньес Дж.К., Кранцуш П.Дж., Ноеске Дж., Райт А.В., Дэвис К.В., Дудна Дж.А. (июнь 2014 г.). «Образование комплекса Cas1-Cas2 опосредует приобретение спейсера во время адаптивного иммунитета CRISPR-Cas» . Структурная и молекулярная биология природы . 21 (6): 528–534. дои : 10.1038/nsmb.2820 . ПМК 4075942 . ПМИД 24793649 .
- ^ Нуньес Дж.К., Ли А.С., Энгельман А., Дудна Х.А. (март 2015 г.). «Приобретение спейсера, опосредованное интегразой, во время адаптивного иммунитета CRISPR-Cas» . Природа . 519 (7542): 193–198. Бибкод : 2015Natur.519..193N . дои : 10.1038/nature14237 . ПМК 4359072 . ПМИД 25707795 .
- ^ Ван Дж., Ли Дж., Чжао Х., Шэн Г., Ван М., Инь М. и др. (ноябрь 2015 г.). «Структурные и механистические основы PAM-зависимого приобретения спейсера в системах CRISPR-Cas» . Клетка . 163 (4): 840–853. дои : 10.1016/j.cell.2015.10.008 . ПМИД 26478180 .
- ^ Нуньес Дж.К., Харрингтон Л.Б., Кранцуш П.Дж., Энгельман А.Н., Дудна Х.А. (ноябрь 2015 г.). «Захват чужеродной ДНК во время адаптивного иммунитета CRISPR-Cas» . Природа . 527 (7579): 535–538. Бибкод : 2015Natur.527..535N . дои : 10.1038/nature15760 . ПМЦ 4662619 . ПМИД 26503043 .
- ^ Сорек Р., Лоуренс К.М., Виденхефт Б. (2013). «CRISPR-опосредованная адаптивная иммунная система у бактерий и архей» . Ежегодный обзор биохимии . 82 (1): 237–266. doi : 10.1146/annurev-biochem-072911-172315 . ПМИД 23495939 .
- ^ Нуньес Дж.К., Бай Л., Харрингтон Л.Б., Хиндер Т.Л., Дудна Дж.А. (июнь 2016 г.). «Иммунологическая память CRISPR требует фактора хозяина для специфичности» . Молекулярная клетка . 62 (6): 824–833. doi : 10.1016/j.molcel.2016.04.027 . ПМИД 27211867 .
- ^ Фагерлунд Р.Д., Уилкинсон М.Е., Клыков О., Барендрегт А., Пирс Ф.Г., Кипер С.Н. и др. (июнь 2017 г.). «Захват и интеграция спейсера адаптационным комплексом CRISPR Cas1-Cas2–3 типа IF» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (26): Е5122–Е5128. Бибкод : 2017PNAS..114E5122F . дои : 10.1073/pnas.1618421114 . ПМЦ 5495228 . ПМИД 28611213 .
- ^ Ролли С., Грэм С., Руйон С., Уайт М.Ф. (февраль 2018 г.). «Обработка преспейсеров и специальная интеграция в систему CRISPR типа IA» . Исследования нуклеиновых кислот . 46 (3): 1007–1020. дои : 10.1093/nar/gkx1232 . ПМК 5815122 . ПМИД 29228332 .
- ^ Jump up to: а б с д Хорват П., Ромеро Д.А., Куте-Монвуазен А.С., Ричардс М., Дево Х., Муано С. и др. (февраль 2008 г.). «Разнообразие, активность и эволюция локусов CRISPR у Streptococcus thermophilus » . Журнал бактериологии . 190 (4): 1401–1412. дои : 10.1128/JB.01415-07 . ПМК 2238196 . ПМИД 18065539 .
- ^ Jump up to: а б с Дево Х., Баррангу Р. , Гарно Дж.Э., Лабонте Дж., Фремо С., Бояваль П. и др. (февраль 2008 г.). «Фаговый ответ на устойчивость, кодируемую CRISPR, у Streptococcus thermophilus » . Журнал бактериологии . 190 (4): 1390–1400. дои : 10.1128/JB.01412-07 . ПМК 2238228 . ПМИД 18065545 .
- ^ Мохика Ф.Д., Диес-Вильясеньор К., Гарсиа-Мартинес Х., Альмендрос К. (март 2009 г.). «Короткие последовательности мотивов определяют цели прокариотической защитной системы CRISPR» . Микробиология . 155 (Часть 3): 733–740. дои : 10.1099/mic.0.023960-0 . ПМИД 19246744 .
- ^ Jump up to: а б Лиллестёль Р.К., Шах С.А., Брюггер К., Реддер П., Фан Х., Кристиансен Дж. и др. (апрель 2009 г.). «Семейства CRISPR рода Sulfolobus кренархей: двунаправленная транскрипция и динамические свойства». Молекулярная микробиология . 72 (1): 259–272. дои : 10.1111/j.1365-2958.2009.06641.x . ПМИД 19239620 . S2CID 36258923 .
- ^ Jump up to: а б Шах С.А., Хансен Н.Р., Гарретт Р.А. (февраль 2009 г.). «Распределение спейсерных совпадений CRISPR в вирусах и плазмидах кренархейных ацидотермофилов и значение их ингибирующего механизма». Труды Биохимического общества . 37 (Часть 1): 23–28. дои : 10.1042/BST0370023 . ПМИД 19143596 . S2CID 19093261 .
- ^ Jump up to: а б Диес-Вильясеньор К., Гусман Н.М., Альмендрос К., Гарсиа-Мартинес Х., Мохика Ф.Дж. (май 2013 г.). «Репортерные плазмиды интеграции CRISPR-спейсера обнаруживают различные подлинные особенности приобретения среди вариантов CRISPR-Cas IE Escherichia coli » . Биология РНК . 10 (5): 792–802. дои : 10.4161/rna.24023 . ПМЦ 3737337 . ПМИД 23445770 .
- ^ Jump up to: а б с Эрдманн С., Гаррет Р.А. (сентябрь 2012 г.). «Селективное и гиперактивное поглощение чужеродной ДНК адаптивной иммунной системой археи посредством двух различных механизмов» . Молекулярная микробиология . 85 (6): 1044–1056. дои : 10.1111/j.1365-2958.2012.08171.x . ПМЦ 3468723 . ПМИД 22834906 .
- ^ Jump up to: а б Шах С.А., Эрдманн С., Мохика Ф.Дж., Гаррет Р.А. (май 2013 г.). «Мотивы узнавания протоспейсеров: смешанные идентичности и функциональное разнообразие» . Биология РНК . 10 (5): 891–899. дои : 10.4161/rna.23764 . ПМЦ 3737346 . ПМИД 23403393 .
- ^ Андерссон А.Ф., Банфилд Дж.Ф. (май 2008 г.). «Динамика популяций вирусов и приобретенная устойчивость к вирусам в природных микробных сообществах». Наука . 320 (5879): 1047–1050. Бибкод : 2008Sci...320.1047A . дои : 10.1126/science.1157358 . ПМИД 18497291 . S2CID 26209623 .
- ^ Jump up to: а б с д Pride DT, Sun CL, Salzman J, Rao N, Loomer P, Armitage GC и др. (январь 2011 г.). «Анализ стрептококковых CRISPR из слюны человека выявляет существенное разнообразие последовательностей внутри и между субъектами с течением времени» . Геномные исследования . 21 (1): 126–136. дои : 10.1101/гр.111732.110 . ПМК 3012920 . ПМИД 21149389 .
- ^ Jump up to: а б Горен М.Г., Йосеф I, Остер О, Кимрон У (октябрь 2012 г.). «Экспериментальное определение сгруппированных коротких палиндромных дупликонов с регулярными промежутками в Escherichia coli ». Журнал молекулярной биологии . 423 (1): 14–16. дои : 10.1016/j.jmb.2012.06.037 . ПМИД 22771574 .
- ^ Jump up to: а б с Даценко К.А., Пугач К., Тихонов А., Ваннер Б.Л., Северинов К., Семенова Е. (июль 2012 г.). «Молекулярная память о предшествующих инфекциях активирует систему адаптивного бактериального иммунитета CRISPR/Cas» . Природные коммуникации . 3 : 945. Бибкод : 2012NatCo...3..945D . дои : 10.1038/ncomms1937 . ПМИД 22781758 .
- ^ Геснер Э.М., Шелленберг М.Дж., Гарсайд Э.Л., Джордж М.М., Макмиллан А.М. (июнь 2011 г.). «Распознавание и созревание эффекторных РНК в пути вмешательства CRISPR». Структурная и молекулярная биология природы . 18 (6): 688–692. дои : 10.1038/nsmb.2042 . ПМИД 21572444 . S2CID 677704 .
- ^ Сашитал Д.Г., Джинек М., Дудна Дж.А. (июнь 2011 г.). «Вызванное РНК конформационное изменение, необходимое для расщепления РНК CRISPR эндорибонуклеазой Cse3». Структурная и молекулярная биология природы . 18 (6): 680–687. дои : 10.1038/nsmb.2043 . ПМИД 21572442 . S2CID 5538195 .
- ^ Хаурвиц Р.Э., Джинек М., Виденхефт Б., Чжоу К., Дудна Дж.А. (сентябрь 2010 г.). «Обработка РНК, специфичная для последовательности и структуры, эндонуклеазой CRISPR» . Наука . 329 (5997): 1355–1358. Бибкод : 2010Sci...329.1355H . дои : 10.1126/science.1192272 . ПМК 3133607 . ПМИД 20829488 .
- ^ Jump up to: а б Кунин В., Сорек Р., Гугенхольц П. (2007). «Эволюционная консервативность последовательности и вторичных структур в повторах CRISPR» . Геномная биология . 8 (4): С61. дои : 10.1186/gb-2007-8-4-r61 . ПМК 1896005 . ПМИД 17442114 .
- ^ Карт Дж., Ван Р., Ли Х., Тернс Р.М., Тернс MP (декабрь 2008 г.). «Cas6 — это эндорибонуклеаза, которая генерирует направляющие РНК для защиты от захватчиков у прокариот» . Гены и развитие . 22 (24): 3489–3496. дои : 10.1101/gad.1742908 . ПМК 2607076 . ПМИД 19141480 .
- ^ Ван Р., Преамплюм Г., Тернс М.П., Тернс Р.М., Ли Х (февраль 2011 г.). «Взаимодействие рибоэндонуклеазы Cas6 с РНК CRISPR: распознавание и расщепление» . Структура . 19 (2): 257–264. дои : 10.1016/j.str.2010.11.014 . ПМК 3154685 . ПМИД 21300293 .
- ^ Нивонер О., Джинек М., Дудна Дж.А. (январь 2014 г.). «Эволюция распознавания и обработки РНК CRISPR эндонуклеазами Cas6» . Исследования нуклеиновых кислот . 42 (2): 1341–1353. дои : 10.1093/нар/gkt922 . ПМК 3902920 . ПМИД 24150936 .
- ^ Семенова Е., Жоре М.М., Даценко К.А., Семенова А., Вестра Э.Р., Ваннер Б. и др. (июнь 2011 г.). «Вмешательство кластеризованных коротких палиндромных повторов (CRISPR) РНК с регулярными промежутками регулируется затравочной последовательностью» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (25): 10098–10103. Бибкод : 2011PNAS..10810098S . дои : 10.1073/pnas.1104144108 . ПМК 3121866 . ПМИД 21646539 .
- ^ Гудбергсдоттир С., Дэн Л., Чен З., Дженсен Дж.В., Дженсен Л.Р., Ше К. и др. (январь 2011 г.). «Динамические свойства систем Sulfolobus CRISPR/Cas и CRISPR/Cmr при воздействии трансмиссивных вирусных и плазмидных генов и протоспейсеров» . Молекулярная микробиология . 79 (1): 35–49. дои : 10.1111/j.1365-2958.2010.07452.x . ПМК 3025118 . ПМИД 21166892 .
- ^ Маника А., Зебек З., Тейхманн Д., Шлепер С. (апрель 2011 г.). «In vivo активность CRISPR-опосредованной вирусной защиты у гипертермофильных архей» . Молекулярная микробиология . 80 (2): 481–491. дои : 10.1111/j.1365-2958.2011.07586.x . ПМИД 21385233 . S2CID 41442419 .
- ^ Джор М.М., Лундгрен М., ван Дуйн Э., Бултема Дж.Б., Вестра Э.Р., Вагмаре С.П. и др. (май 2011 г.). «Структурная основа распознавания ДНК с помощью CRISPR-РНК с помощью Cascade» (PDF) . Структурная и молекулярная биология природы . 18 (5): 529–536. дои : 10.1038/nsmb.2019 . ПМИД 21460843 . S2CID 10987554 . Архивировано (PDF) из оригинала 23 октября 2021 г. Проверено 30 июня 2019 г.
- ^ Виденхефт Б., Ландер Г.К., Чжоу К., Джор М.М., Браунс С.Дж., ван дер Ост Дж. и др. (сентябрь 2011 г.). «Структуры РНК-ориентированного надзорного комплекса бактериальной иммунной системы» . Природа . 477 (7365): 486–489. Бибкод : 2011Natur.477..486W . дои : 10.1038/nature10402 . ПМК 4165517 . ПМИД 21938068 .
- ^ Чжан Дж., Руйон С., Керу М., Рикс Дж., Брюггер К., Грэм С. и др. (февраль 2012 г.). «Структура и механизм комплекса CMR для CRISPR-опосредованного противовирусного иммунитета» . Молекулярная клетка . 45 (3): 303–313. doi : 10.1016/j.molcel.2011.12.013 . ПМЦ 3381847 . ПМИД 22227115 .
- ^ Jump up to: а б Хейл С.Р., Чжао П., Олсон С., Дафф М.О., Грейвли Б.Р., Уэллс Л. и др. (ноябрь 2009 г.). «Расщепление РНК под контролем РНК белковым комплексом CRISPR РНК-Cas» . Клетка . 139 (5): 945–956. дои : 10.1016/j.cell.2009.07.040 . ПМЦ 2951265 . ПМИД 19945378 .
- ^ Эстрелла М.А., Куо Ф.Т., Бэйли С. (2016). «РНК-активируемое расщепление ДНК эффекторным комплексом CRISPR-Cas типа III-B» . Гены и развитие . 30 (4): 460–470. дои : 10.1101/gad.273722.115 . ПМЦ 4762430 . ПМИД 26848046 .
- ^ Самай П., Пьенсон Н. , Цзян В., Голдберг Г.В., Хатум-Аслан А., Марраффини Л.А. (2015). «Котранскрипционное расщепление ДНК и РНК во время иммунитета CRISPR-Cas типа III» . Клетка . 161 (5): 1164–1174. дои : 10.1016/j.cell.2015.04.027 . ПМЦ 4594840 . ПМИД 25959775 .
- ^ Jump up to: а б с Марраффини Л.А., Сонтхаймер Э.Дж. (январь 2010 г.). «Дискриминация себя и чужого во время иммунитета, направленного на РНК CRISPR» . Природа . 463 (7280): 568–571. Бибкод : 2010Natur.463..568M . дои : 10.1038/nature08703 . ПМЦ 2813891 . ПМИД 20072129 .
- ^ Крупович М., Беген П., Кунин Е.В. (август 2017 г.). «Каспозоны: мобильные генетические элементы, породившие механизм адаптации CRISPR-Cas» . Современное мнение в микробиологии . 38 : 36–43. дои : 10.1016/j.mib.2017.04.004 . ПМЦ 5665730 . ПМИД 28472712 .
- ^ Кунин Е.В. , Макарова К.С. (май 2013 г.). «CRISPR-Cas: эволюция системы адаптивного иммунитета на основе РНК у прокариот» . Биология РНК . 10 (5): 679–686. дои : 10.4161/rna.24022 . ПМЦ 3737325 . ПМИД 23439366 .
- ^ Кунин Е.В. , Макарова К.С., Чжан Ф (июнь 2017). «Разнообразие, классификация и эволюция систем CRISPR-Cas» . Современное мнение в микробиологии . 37 : 67–78. дои : 10.1016/j.mib.2017.05.008 . ПМЦ 5776717 . ПМИД 28605718 .
- ^ Jump up to: а б Шмаков С., Смаргон А., Скотт Д., Кокс Д., Пызоча Н., Ян В. и др. (март 2017 г.). «Разнообразие и эволюция систем CRISPR-Cas класса 2» . Обзоры природы. Микробиология . 15 (3): 169–182. дои : 10.1038/nrmicro.2016.184 . ПМЦ 5851899 . ПМИД 28111461 .
- ^ Купчок А., Болбак Дж.П. (февраль 2013 г.). «Вероятностные модели эволюции содержания спейсера CRISPR» . Эволюционная биология BMC . 13 (1): 54. Бибкод : 2013BMCEE..13...54K . дои : 10.1186/1471-2148-13-54 . ПМК 3704272 . ПМИД 23442002 .
- ^ Штернберг С.Х., Рихтер Х., Шарпантье Э., Кимрон У. (март 2016 г.). «Адаптация в системах CRISPR-Cas». Молекулярная клетка . 61 (6): 797–808. doi : 10.1016/j.molcel.2016.01.030 . hdl : 21.11116/0000-0003-E74E-2 . ПМИД 26949040 .
- ^ Кунин Е.В. , Вольф Ю.И. (ноябрь 2009 г.). «Является ли эволюция дарвиновской или/и ламаркистской?» . Биология Директ . 4:42 . дои : 10.1186/1745-6150-4-42 . ПМЦ 2781790 . ПМИД 19906303 .
- ^ Вайс А. (октябрь 2015 г.). «Ламарковские иллюзии» . Тенденции в экологии и эволюции . 30 (10): 566–568. Бибкод : 2015TEcoE..30..566W . дои : 10.1016/j.tree.2015.08.003 . ПМИД 26411613 .
- ^ Jump up to: а б Кунин Е.В. , Вольф Ю.И. (февраль 2016). «Насколько ламаркианский иммунитет CRISPR-Cas: континуум механизмов развития» . Биология Директ . 11 (1): 9. дои : 10.1186/s13062-016-0111-z . ПМК 4765028 . ПМИД 26912144 .
- ^ Гейдельберг Дж. Ф., Нельсон В. К., Шенфельд Т., Бхайя Д. (2009). Ахмед Н. (ред.). «Микробная война в сообществе микробных матов: CRISPR дают представление о совместной эволюции геномов хозяина и вируса» . ПЛОС ОДИН . 4 (1): е4169. Бибкод : 2009PLoSO...4.4169H . дои : 10.1371/journal.pone.0004169 . ПМЦ 2612747 . ПМИД 19132092 .
- ^ Touchon M, Rocha EP (июнь 2010 г.). Рандау Л. (ред.). «Маленькие, медленные и специализированные CRISPR и анти-CRISPR эшерихий и сальмонелл» . ПЛОС ОДИН . 5 (6): е11126. Бибкод : 2010PLoSO...511126T . дои : 10.1371/journal.pone.0011126 . ПМК 2886076 . ПМИД 20559554 .
- ^ Jump up to: а б с Ро М., Ву Ю.В., Тан Х., Доак Т.Г., Йе Ю. (2012). «Разнообразные CRISPR развиваются в микробиомах человека» . ПЛОС Генетика . 8 (6): e1002441. дои : 10.1371/journal.pgen.1002441 . ПМЦ 3374615 . ПМИД 22719260 .
- ^ Jump up to: а б Сан КЛ, Баррангу Р. , Томас БК, Хорват П., Фремо К., Банфилд Дж.Ф. (февраль 2013 г.). «Фаговые мутации в ответ на диверсификацию CRISPR в бактериальной популяции». Экологическая микробиология . 15 (2): 463–470. Бибкод : 2013EnvMi..15..463S . дои : 10.1111/j.1462-2920.2012.02879.x . ПМИД 23057534 .
- ^ Куно С., Сако Ю., Ёсида Т. (май 2014 г.). «Диверсификация CRISPR в пределах сосуществующих генотипов в естественной популяции цветущей цианобактерии Microcystis aeruginosa» . Микробиология . 160 (Часть 5): 903–916. дои : 10.1099/mic.0.073494-0 . ПМИД 24586036 .
- ^ Сорек Р., Кунин В., Гугенхольц П. (март 2008 г.). «CRISPR — широко распространенная система, обеспечивающая приобретенную устойчивость бактерий и архей к фагам» . Обзоры природы. Микробиология . 6 (3): 181–186. дои : 10.1038/nrmicro1793 . ПМИД 18157154 . S2CID 3538077 . Архивировано из оригинала 8 марта 2021 г. Проверено 25 ноября 2019 г.
Таблица 1. Веб-ресурсы для анализа CRISPR
- ^ Прайд Д.Т., Зальцман Дж., Релман Д.А. (сентябрь 2012 г.). «Сравнение кластеризованных коротких палиндромных повторов с регулярными промежутками и виромов в слюне человека выявляет бактериальную адаптацию к слюнным вирусам» . Экологическая микробиология . 14 (9): 2564–2576. Бибкод : 2012EnvMi..14.2564P . дои : 10.1111/j.1462-2920.2012.02775.x . ПМЦ 3424356 . ПМИД 22583485 .
- ^ Состоялось в Нидерландах, Эррера А., Уитакер Р.Дж. (ноябрь 2013 г.). «Реассортация локусов повторов-спейсеров CRISPR у Sulfolobus Islandicus». Экологическая микробиология . 15 (11): 3065–3076. Бибкод : 2013EnvMi..15.3065H . дои : 10.1111/1462-2920.12146 . ПМИД 23701169 .
- ^ Состоялось в Нидерландах, Эррера А., Кадилло-Кирос Х., Уитакер Р.Дж. (сентябрь 2010 г.). «Разнообразие, связанное с CRISPR, в популяции Sulfolobus Islandicus» . ПЛОС ОДИН . 5 (9): e12988. Бибкод : 2010PLoSO...512988H . дои : 10.1371/journal.pone.0012988 . ПМЦ 2946923 . ПМИД 20927396 .
- ^ Jump up to: а б с Медведева С., Лю Ю., Кунин Е.В., Северинов К., Прангишвили Д., Крупович М. (ноябрь 2019 г.). «Массивы мини-CRISPR, переносимые вирусами, участвуют в межвирусных конфликтах» . Природные коммуникации . 10 (1): 5204. Бибкод : 2019NatCo..10.5204M . дои : 10.1038/s41467-019-13205-2 . ПМК 6858448 . ПМИД 31729390 .
- ^ Скеннертон, Коннектикут, Имелфорт М, Тайсон Г.В. (май 2013 г.). «Crass: идентификация и реконструкция CRISPR на основе несобранных метагеномных данных» . Исследования нуклеиновых кислот . 41 (10): е105. дои : 10.1093/нар/gkt183 . ПМЦ 3664793 . ПМИД 23511966 .
- ^ Стерн А., Мик Э., Тирош И., Сэги О., Сорек Р. (октябрь 2012 г.). «Нацеливание на CRISPR выявляет резервуар распространенных фагов, связанных с микробиомом кишечника человека» . Геномные исследования . 22 (10): 1985–1994. дои : 10.1101/гр.138297.112 . ПМК 3460193 . ПМИД 22732228 .
- ^ Новик Р.П., Кристи Дж.Э., Пенадес-младший (август 2010 г.). «Фагосвязанные хромосомные острова грамположительных бактерий» . Обзоры природы Микробиология . 8 (8): 541–551. дои : 10.1038/nrmicro2393 . ПМЦ 3522866 . ПМИД 20634809 .
- ^ Рам Дж., Чен Дж., Кумар К., Росс Х.Ф., Убеда С., Дамле П.К. и др. (октябрь 2012 г.). «Вмешательство острова патогенности стафилококка в репродукцию фагов-помощников является парадигмой молекулярного паразитизма» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (40): 16300–16305. Бибкод : 2012PNAS..10916300R . дои : 10.1073/pnas.1204615109 . ПМЦ 3479557 . ПМИД 22991467 .
- ^ Рам Дж., Чен Дж., Росс Х.Ф., Новик Р.П. (октябрь 2014 г.). «Точно модулированное вмешательство острова патогенности в транскрипцию генов позднего фага» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (40): 14536–14541. Бибкод : 2014PNAS..11114536R . дои : 10.1073/pnas.1406749111 . ПМК 4209980 . ПМИД 25246539 .
- ^ Сид К.Д., Лазински Д.В., Колдервуд С.Б., Камилли А. (февраль 2013 г.). «Бактериофаг кодирует собственный адаптивный ответ CRISPR/Cas, чтобы избежать врожденного иммунитета хозяина» . Природа . 494 (7438): 489–491. Бибкод : 2013Natur.494..489S . дои : 10.1038/nature11927 . ПМЦ 3587790 . ПМИД 23446421 .
- ^ Бойд С.М., Ангермейер А., Хейс С.Г., Барт З.К., Патель К.М., Сид К.Д. (сентябрь 2021 г.). «Бактериофаг ICP1: стойкий хищник холерного вибриона » . Ежегодный обзор вирусологии . 8 (1): 285–304. doi : 10.1146/annurev-virology-091919-072020 . ISSN 2327-056X . ПМК 9040626 . ПМИД 34314595 .
- ^ Фэн Ц, Ли Ц, Чжоу Х, Ван З, Линь С, Цзян Цз и др. (август 2024 г.). «Технология CRISPR в болезнях человека» . Медкомм . 5 (8): е672. дои : 10.1002/mco2.672 . ПМЦ 11286548 . ПМИД 39081515 .
- ^ Ли Т, Ли С, Кан Ю, Чжоу Дж, И М (август 2024 г.). «Использование развивающегося CRISPR/Cas9 для точной онкологии» . Журнал трансляционной медицины . 22 (1): 749. doi : 10.1186/s12967-024-05570-4 . ПМЦ 11312220 . ПМИД 39118151 .
- ^ Мишра С., Наяк С., Тутеджа Н., Пусапати С., Суэйн Д.М., Саху Р.К. (июль 2024 г.). «CRISPR/Cas-опосредованная геномная инженерия растений: применение и перспективы» . Растения . 13 (14). Базель, Швейцария: 1884 г. doi : 10.3390/plants13141884 . ПМЦ 11279650 . ПМИД 39065411 .
- ^ Каур Р., Гупта С., Чаухан А., Мишра В., Шарма М.К., Сингх Дж. (август 2024 г.). «Использование возможностей микрофлюидики на основе кластерных регулярных коротких палиндромных повторов (CRISPR) для молекулярной диагностики следующего поколения». Отчеты по молекулярной биологии . 51 (1): 896. doi : 10.1007/s11033-024-09840-8 . ПМИД 39115550 .
- ^ Шинвари З.К., Танвир Ф., Халил А.Т. (2018). «Этические проблемы редактирования генома с помощью CRISPR» . Актуальные проблемы молекулярной биологии . 26 : 103–110. дои : 10.21775/cimb.026.103 . ПМИД 28879860 .
- ^ Козан Д.В., Фарбер С.А. (февраль 2024 г.). «Разумно ли когда-нибудь редактировать аллели дикого типа? Сконструированные аллели CRISPR против миллионов лет эволюции человека». Атеросклероз, тромбоз и сосудистая биология . 44 (2): 328–333. дои : 10.1161/ATVBAHA.123.318069 . ПМК 10948015. ПМИД 38059350 .
Дальнейшее чтение
[ редактировать ]- Дудна Дж. , Мали П. (23 марта 2016 г.). CRISPR-Cas: Лабораторное руководство . Нью-Йорк: Лабораторное издательство Колд-Спринг-Харбор. ISBN 978-1-62182-131-1 .
- Моханраджу П., Макарова К.С., Зетше Б., Чжан Ф. , Кунин Е.В. , ван дер Ост Дж. (август 2016 г.). «Различные эволюционные корни и механистические вариации систем CRISPR-Cas» (PDF) . Наука . 353 (6299): аад5147. дои : 10.1126/science.aad5147 . hdl : 1721.1/113195 . ПМИД 27493190 . S2CID 11086282 . Архивировано из оригинала 23 октября 2021 г. Проверено 29 августа 2019 г.
- Сандер Дж.Д., Йонг Дж.К. (апрель 2014 г.). «Системы CRISPR-Cas для редактирования, регулирования и нацеливания геномов» . Природная биотехнология . 32 (4): 347–355. дои : 10.1038/nbt.2842 . ПМК 4022601 . ПМИД 24584096 .
- Slaymaker IM, Гао Л., Зетше Б., Скотт Д.А., Ян В.С., Чжан Ф. (январь 2016 г.). «Рационально сконструированные нуклеазы Cas9 с улучшенной специфичностью» . Наука . 351 (6268): 84–88. Бибкод : 2016Sci...351...84S . дои : 10.1126/science.aad5227 . ПМЦ 4714946 . ПМИД 26628643 .
- Тернс Р.М., Тернс МП (март 2014 г.). «Технологии на основе CRISPR: перепрофилирование прокариотического защитного оружия» . Тенденции в генетике . 30 (3): 111–118. дои : 10.1016/j.tig.2014.01.003 . ПМЦ 3981743 . ПМИД 24555991 .
- Вестра Э.Р., Баклинг А., Финеран ПК (май 2014 г.). «Системы CRISPR-Cas: за пределами адаптивного иммунитета». Обзоры природы Микробиология . 12 (5): 317–326. дои : 10.1038/nrmicro3241 . ПМИД 24704746 . S2CID 36575361 .
- Андерссон А.Ф., Банфилд Дж.Ф. (май 2008 г.). «Динамика популяций вирусов и приобретенная устойчивость к вирусам в природных микробных сообществах». Наука . 320 (5879): 1047–1050. Бибкод : 2008Sci...320.1047A . дои : 10.1126/science.1157358 . ПМИД 18497291 . S2CID 26209623 .
- Хейл С., Клеппе К., Тернс Р.М., Тернс член парламента (декабрь 2008 г.). «Прокариотические молчащие (пси)РНК у Pyrococcus Furiosus» . РНК . 14 (12): 2572–2579. дои : 10.1261/rna.1246808 . ПМК 2590957 . ПМИД 18971321 .
- ван дер Плог-младший (июнь 2009 г.). «Анализ CRISPR у Streptococcus mutans предполагает частое возникновение приобретенного иммунитета против заражения M102-подобными бактериофагами» (PDF) . Микробиология . 155 (Часть 6): 1966–1976. дои : 10.1099/mic.0.027508-0 . ПМИД 19383692 .
- ван дер Ост Дж., Браунс С.Дж. (ноябрь 2009 г.). «RNAi: в дело вступают прокариоты» . Клетка . 139 (5): 863–865. дои : 10.1016/j.cell.2009.11.018 . ПМИД 19945373 . S2CID 11863610 .
- Каргинов Ф.В., Хэннон Г.Дж. (январь 2010 г.). «Система CRISPR: защита, управляемая малой РНК, у бактерий и архей» . Молекулярная клетка . 37 (1): 7–19. doi : 10.1016/j.molcel.2009.12.033 . ПМК 2819186 . ПМИД 20129051 .
- Пул У, Вурм Р., Арслан З., Гейссен Р., Хофманн Н., Вагнер Р. (март 2010 г.). «Идентификация и характеристика промоторов CRISPR-cas E. coli и их подавление с помощью H-NS» . Молекулярная микробиология . 75 (6): 1495–1512. дои : 10.1111/j.1365-2958.2010.07073.x . ПМИД 20132443 . S2CID 37529215 .
- Диес-Вильясеньор К., Альмендрос К., Гарсиа-Мартинес Х., Мохика Ф.Дж. (май 2010 г.). «Разнообразие локусов CRISPR в Escherichia coli » . Микробиология . 156 (Часть 5): 1351–1361. дои : 10.1099/mic.0.036046-0 . ПМИД 20133361 .
- Дево Х., Гарно Ж.Э., Муано С. (2010). «Система CRISPR/Cas и ее роль во взаимодействии фагов и бактерий». Ежегодный обзор микробиологии . 64 : 475–493. дои : 10.1146/annurev.micro.112408.134123 . ПМИД 20528693 .
- Кунин Е.В. , Макарова К.С. (декабрь 2009 г.). «CRISPR-Cas: система адаптивного иммунитета у прокариот» . Отчеты о биологии F1000 . 1 : 95. дои : 10.3410/B1-95 . ПМЦ 2884157 . ПМИД 20556198 .
- «Эпоха красного пера» . Экономист . 22 августа 2015. ISSN 0013-0613 . Архивировано из оригинала 24 августа 2015 г. Проверено 25 августа 2015 г.
- Ран А.Ф., Сюй П.Д., Райт Дж., Агарвала В., Скотт Д.А., Чжан Ф. (2013). «Геномная инженерия с использованием системы CRISPR-Cas9» . Протоколы природы . 8 (11): 2281–2308. дои : 10.1038/nprot.2013.143 . ПМЦ 3969860 . ПМИД 24157548 .
Внешние ссылки
[ редактировать ]- «Расширенное редактирование генов: CRISPR-Cas9» (PDF) . Исследовательская служба Конгресса .
- «Доклад Дженнифер Дудна: Геномная инженерия с CRISPR-Cas9: рождение революционной технологии» . 10 сентября 2022 г.
- «Человеческая природа» . НОВА . 47 сезон. 9 серия. 9 сентября 2020. PBS . ВГБХ . Проверено 7 апреля 2023 г.
Банк данных по белкам
[ редактировать ]- Обзор всей структурной информации, доступной в PDB для UniProt : Q46901 (каскадный субблок CasA системы CRISPR) в PDBe-KB .
- Обзор всей структурной информации, доступной в PDB для UniProt : P76632 (каскадный субблок CasB системы CRISPR) в PDBe-KB .
- Обзор всей структурной информации, доступной в PDB для UniProt : Q46899 (каскадный субблок CasC системы CRISPR) в PDBe-KB .
- Обзор всей структурной информации, доступной в PDB для UniProt : Q46898 (каскадный субблок CasD системы CRISPR) в PDBe-KB .
- Обзор всей структурной информации, доступной в PDB для UniProt : Q46897 (каскадный субблок CasE системы CRISPR) в PDBe-KB .