Биология развития
| Часть серии о |
| Биология |
|---|
Биология развития – это изучение процесса роста и развития животных и растений . Биология развития также охватывает биологию регенерации , бесполого размножения , метаморфоза , а также роста и дифференциации стволовых клеток во взрослом организме.
Перспективы
[ редактировать ]Основными процессами, участвующими в эмбриональном развитии животных, являются: формирование паттерна ткани (посредством региональной спецификации и дифференцировки клеток по паттерну ); рост тканей ; и морфогенез тканей .
- Региональная спецификация относится к процессам, которые создают пространственные структуры в шаре или листе изначально похожих клеток. Обычно это связано с действием цитоплазматических детерминант , расположенных внутри частей оплодотворенной яйцеклетки, и индуктивных сигналов, излучаемых сигнальными центрами эмбриона. На ранних стадиях региональной спецификации образуются не функционально дифференцированные клетки, а клеточные популяции, готовые развиваться в определенной области или части организма. Они определяются экспрессией специфических комбинаций факторов транскрипции .
- Дифференциация клеток относится конкретно к образованию функциональных типов клеток, таких как нервные, мышечные, секреторный эпителий и т. д. Дифференцированные клетки содержат большое количество специфических белков, связанных с клеточной функцией.
- Морфогенез связан с образованием трехмерной формы. В основном это включает в себя организованные движения клеточных листов и отдельных клеток. Морфогенез важен для создания трех зародышевых слоев раннего эмбриона ( эктодермы , мезодермы и энтодермы ) и для построения сложных структур во время развития органов.
- Рост ткани включает как общее увеличение размера ткани, так и дифференциальный рост частей ( аллометрия ), который способствует морфогенезу. Рост в основном происходит за счет пролиферации клеток , а также за счет изменения размера клеток или отложения внеклеточного материала.
В развитии растений происходят те же процессы, что и у животных. Однако растительные клетки в основном неподвижны, поэтому морфогенез достигается за счет дифференциального роста без движений клеток. Кроме того, индуктивные сигналы и задействованные гены отличаются от тех, которые контролируют развитие животных.
Генеративная биология
[ редактировать ]Генеративная биология — это генеративная наука , которая исследует динамику, направляющую развитие и эволюцию биологической морфологической формы. [ 1 ] [ 2 ] [ 3 ]
Процессы развития
[ редактировать ]Дифференциация клеток
[ редактировать ]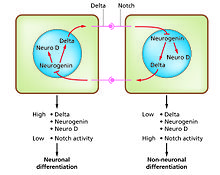
Дифференциация клеток — это процесс, в результате которого в развитии возникают различные функциональные типы клеток. Например, нейроны, мышечные волокна и гепатоциты (клетки печени) являются хорошо известными типами дифференцированных клеток. Дифференцированные клетки обычно производят большое количество нескольких белков, которые необходимы для их специфической функции, и это придает им характерный внешний вид, позволяющий распознавать их под световым микроскопом. Гены, кодирующие эти белки, высокоактивны. Обычно их структура хроматина очень открыта, что обеспечивает доступ ферментам транскрипции, а специфические факторы транскрипции связываются с регуляторными последовательностями ДНК, чтобы активировать экспрессию генов. [ 4 ] [ 5 ] Например, NeuroD является ключевым фактором транскрипции для дифференцировки нейронов, миогенин для дифференцировки мышц и HNF4 для дифференцировки гепатоцитов. Дифференциация клеток обычно является заключительной стадией развития, которой предшествуют несколько состояний детерминации, которые визуально не дифференцируются. Одна ткань, образованная из одного типа клеток-предшественников или стволовых клеток, часто состоит из нескольких дифференцированных типов клеток. Контроль их образования включает процесс латерального торможения, [ 6 ] на основе свойств сигнального пути Notch . [ 7 ] Например, в нервной пластинке эмбриона эта система генерирует популяцию нейрональных клеток-предшественников, в которых высока экспрессия NeuroD.
Регенерация
[ редактировать ]Регенерация указывает на способность восстановить недостающую часть. [ 8 ] Это очень распространено среди растений, которые демонстрируют непрерывный рост, а также среди колониальных животных, таких как гидроиды и асцидии. Но наибольший интерес биологи развития проявляют к регенерации частей свободноживущих животных. В частности, четыре модели стали предметом многочисленных исследований. Два из них обладают способностью регенерировать целые тела: Гидра , способная регенерировать любую часть полипа из небольшого фрагмента, [ 9 ] и планарии , которые обычно могут регенерировать как голову, так и хвост. [ 10 ] В обоих этих примерах наблюдается непрерывный клеточный оборот, питаемый стволовыми клетками , и, по крайней мере, у планарий, по крайней мере, некоторые из стволовых клеток оказались плюрипотентными . [ 11 ] Две другие модели демонстрируют только дистальную регенерацию придатков. Это придатки насекомых, обычно ноги полуметаболических насекомых, таких как сверчок, [ 12 ] и конечности хвостатых амфибий . [ 13 ] В настоящее время имеется значительная информация о регенерации конечностей амфибий, и известно, что каждый тип клеток регенерирует сам, за исключением соединительных тканей, где происходит значительная взаимоконверсия между хрящом, дермой и сухожилиями. Что касается структуры структур, то это контролируется повторной активацией сигналов, активных в эмбрионе. До сих пор ведутся споры по поводу старого вопроса о том, является ли регенерация «первозданным» или «адаптивным» свойством. [ 14 ] Если это так, то с улучшением знаний мы могли бы ожидать, что сможем улучшить регенеративные способности у людей. Если последнее, то предполагается, что каждый случай регенерации возник в результате естественного отбора в обстоятельствах, специфичных для данного вида, поэтому не следует ожидать каких-либо общих правил.
Эмбриональное развитие животных
[ редактировать ]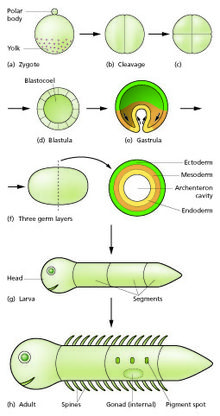

Сперматозоид и яйцеклетка сливаются в процессе оплодотворения, образуя оплодотворенную яйцеклетку или зиготу . [ 15 ] Она подвергается периоду делений с образованием шара или листа одинаковых клеток, называемых бластулой или бластодермой . Эти клеточные деления обычно происходят быстро и без роста, поэтому дочерние клетки составляют половину размера материнской клетки, а весь эмбрион остается примерно того же размера. Их называют делениями расщепления .
мыши эпибласта Первичные зародышевые клетки (см. рисунок: «Начальные стадии эмбриогенеза человека ») подвергаются обширному эпигенетическому перепрограммированию. [ 16 ] Этот процесс включает геному по всему деметилирование ДНК , реорганизацию хроматина и стирание эпигенетического импринта, что приводит к тотипотентности . [ 16 ] Деметилирование ДНК осуществляется с помощью процесса, в котором используется путь эксцизионной репарации оснований ДНК . [ 17 ]
Морфогенетические движения преобразуют клеточную массу в трехслойную структуру, состоящую из многоклеточных листов, называемых эктодермой , мезодермой и энтодермой . Эти листы известны как зародышевые листки . Это процесс гаструляции . Во время дробления и гаструляции происходят первые события региональной спецификации. Помимо формирования самих трех зародышевых листков, они часто образуют внеэмбриональные структуры, такие как плацента млекопитающих , необходимые для поддержки и питания эмбриона. [ 18 ] а также установить различия фиксации по переднезадней оси (голова, туловище и хвост). [ 19 ]
Региональная спецификация инициируется наличием цитоплазматических детерминант в одной части зиготы. Клетки, содержащие детерминанту, становятся сигнальным центром и выделяют индуцирующий фактор. Поскольку индуцирующий фактор вырабатывается в одном месте, диффундирует и распадается, он образует градиент концентрации, высокий вблизи клеток-источников и низкий вдали от них. [ 20 ] [ 21 ] Остальные клетки эмбриона, которые не содержат детерминанту, способны реагировать на различные концентрации путем активации специфических генов контроля развития. В результате создается ряд зон, расположенных на все большем расстоянии от сигнального центра. В каждой зоне активируется различная комбинация генов контроля развития. [ 22 ] Эти гены кодируют факторы транскрипции , которые активируют новые комбинации активности генов в каждой области. Помимо других функций, эти факторы транскрипции контролируют экспрессию генов, придающих специфические адгезивные и подвижные свойства клеткам, в которых они активны. Из-за этих различных морфогенетических свойств клетки каждого зародышевого листка перемещаются, образуя листы, так что эктодерма оказывается снаружи, мезодерма — посередине, а энтодерма — внутри. [ 23 ] [ 24 ]

Морфогенетические движения не только изменяют форму и структуру эмбриона, но, приводя клеточные листы в новые пространственные взаимоотношения, они также делают возможными новые фазы передачи сигналов и реакции между ними. Кроме того, первые морфогенетические движения эмбриогенеза, такие как гаструляция, эпиболия и скручивание , напрямую активируют пути, участвующие в спецификации эндомезодермы посредством процессов механотрансдукции. [ 25 ] [ 26 ] Было высказано предположение, что это свойство эволюционно унаследовано от спецификации эндомезодермы, механически стимулируемой гидродинамическим потоком морской среды у первых животных организмов (первых многоклеточных животных). [ 27 ] Скручивание вдоль оси тела за счет левосторонней хиральности встречается у всех хордовых (включая позвоночных) и рассматривается в теории осевого скручивания . [ 28 ]
Рост эмбрионов в основном автономен. [ 29 ] Для каждой территории клеток скорость роста контролируется комбинацией активных генов. Свободноживущие эмбрионы не растут в массе, поскольку у них нет внешнего источника питания. Но эмбрионы, питающиеся плацентой или внеэмбриональным желтком, могут расти очень быстро, и изменения относительной скорости роста между частями этих организмов помогают создать окончательную общую анатомию.
Весь процесс нужно согласовать во времени и как это контролируется непонятно. Могут существовать главные часы, способные взаимодействовать со всеми частями эмбриона и контролировать ход событий, или время может зависеть просто от локальных причинных последовательностей событий. [ 30 ]
Метаморфоза
[ редактировать ]Процессы развития очень очевидны в процессе метаморфоза . Это происходит у различных видов животных. Хорошо известные примеры можно увидеть у лягушек, которые обычно вылупляются как головастики и превращаются во взрослую лягушку, а также у некоторых насекомых, которые вылупляются как личинки, а затем на стадии куколки реконструируются во взрослую форму.
Все перечисленные выше процессы развития происходят в ходе метаморфоза. Примеры, которые были особенно хорошо изучены, включают потерю хвоста и другие изменения в головастике лягушки Xenopus , [ 31 ] [ 32 ] и биология имагинальных дисков, из которых образуются части тела взрослой особи мухи Drosophila melanogaster . [ 33 ] [ 34 ]
Развитие завода
[ редактировать ]растений Развитие — это процесс, в ходе которого структуры возникают и созревают по мере роста растения. Он изучается в анатомии и физиологии растений , а также в морфологии растений.
Растения на протяжении всей жизни постоянно производят новые ткани и структуры из меристем. [ 35 ] располагаются на кончиках органов или между зрелыми тканями. Таким образом, живое растение всегда имеет эмбриональные ткани. Напротив, эмбрион животного очень рано производит все части тела, которые у него когда-либо будут в жизни. Когда животное рождается (или вылупляется из яйца), у него есть все части тела, и с этого момента оно становится только крупнее и взрослее.
Свойства организации, наблюдаемые в растении, — это эмерджентные свойства , которые представляют собой нечто большее, чем сумма отдельных частей. «Сборка этих тканей и функций в единый многоклеточный организм дает не только характеристики отдельных частей и процессов, но и совершенно новый набор характеристик, который невозможно было бы предсказать на основе изучения отдельных частей». [ 36 ]
Рост
[ редактировать ]начинается Сосудистое растение из одноклеточной зиготы , образующейся в результате оплодотворения яйцеклетки сперматозоидом. С этого момента оно начинает делиться, образуя зародыш растения в процессе эмбриогенеза . При этом образующиеся клетки организуются так, что один конец становится первым корнем, а другой конец образует верхушку побега. У семенных растений у зародыша разовьется один или несколько «семенных листьев» ( семядолей ). К концу эмбриогенеза молодое растение будет иметь все части, необходимые для начала жизни.
Как только эмбрион прорастает из семени или родительского растения, он начинает производить дополнительные органы (листья, стебли и корни) в процессе органогенеза . Новые корни вырастают из корневых меристем, расположенных на кончике корня, а новые стебли и листья вырастают из меристем побега , расположенных на кончике побега. [ 37 ] Ветвление происходит, когда небольшие скопления клеток, оставшиеся от меристемы и еще не подвергшиеся клеточной дифференцировке с образованием специализированной ткани, начинают расти как верхушка нового корня или побега. Рост любой такой меристемы на кончике корня или побега называется первичным ростом и приводит к удлинению этого корня или побега. Вторичный рост приводит к расширению корня или побега за счет деления клеток камбия . [ 38 ]
Помимо роста за счет деления клеток , растение может расти за счет удлинения клеток . [ 39 ] Это происходит, когда отдельные клетки или группы клеток становятся длиннее. Не все растительные клетки вырастают до одинаковой длины. Когда клетки на одной стороне стебля растут длиннее и быстрее, чем клетки на другой стороне, в результате стебель изгибается в сторону более медленно растущих клеток. Этот направленный рост может происходить через реакцию растения на определенный раздражитель, такой как свет ( фототропизм ), гравитация ( гравитропизм ), вода ( гидротропизм ) и физический контакт ( тигмотропизм ).
Рост и развитие растений опосредованы специфическими фитогормонами и регуляторами роста растений (РГР) (Росс и др., 1983). [ 40 ] На уровень эндогенных гормонов влияют возраст растения, холодостойкость, период покоя и другие метаболические условия; фотопериод, засуха, температура и другие внешние условия среды; и экзогенные источники ГРР, например, применяемые извне и ризосферного происхождения.
Морфологическая вариация
[ редактировать ]Растения демонстрируют естественные различия в своей форме и строении. Хотя все организмы варьируются от человека к человеку, растения демонстрируют дополнительный тип вариаций. Внутри одной особи повторяются части, которые по форме и строению могут отличаться от других подобных частей. Эту вариацию легче всего увидеть на листьях растения, хотя и в других органах, таких как стебли и цветы, могут наблюдаться аналогичные вариации. Есть три основные причины этого изменения: позиционные эффекты, воздействие окружающей среды и молодость.
Эволюция морфологии растений
[ редактировать ]Факторы транскрипции и сети регуляции транскрипции играют ключевую роль в морфогенезе растений и их эволюции. Во время посадки растений появилось множество новых семейств транскрипционных факторов, которые преимущественно встраиваются в сети многоклеточного развития, размножения и развития органов, способствуя более сложному морфогенезу наземных растений. [ 41 ]
Большинство наземных растений имеют общего предка — многоклеточные водоросли. Пример эволюции морфологии растений можно увидеть на харофитах. Исследования показали, что харофиты обладают признаками, гомологичными наземным растениям. Существуют две основные теории эволюции морфологии растений: гомологичная теория и антитетическая теория. Общепринятой теорией эволюции морфологии растений является антитетическая теория. Противоположная теория утверждает, что множественные митотические деления, происходящие перед мейозом, вызывают развитие спорофита. Тогда спорофит будет развиваться как самостоятельный организм. [ 42 ]
Модельные организмы развития
[ редактировать ]Большая часть исследований в области биологии развития в последние десятилетия была сосредоточена на использовании небольшого числа модельных организмов . Оказалось, что в животном мире механизмы развития во многом консервативны. На раннем этапе развития все виды позвоночных используют по существу одни и те же индуктивные сигналы и одни и те же гены, кодирующие региональную идентичность. Даже беспозвоночные используют схожий набор сигналов и генов, хотя сформированные части тела существенно различаются. Каждый из модельных организмов обладает определенными экспериментальными преимуществами, которые позволили им стать популярными среди исследователей. В одном смысле они являются «моделями» всего животного мира, а в другом смысле — «моделями» человеческого развития, которое трудно изучать непосредственно как по этическим, так и по практическим причинам. Модельные организмы оказались наиболее полезными для выяснения широкой природы механизмов развития. Чем больше деталей ищется, тем больше они отличаются друг от друга и от человека.
Растения
[ редактировать ]- Кресс-салат ( Arabidopsis thaliana ) [ 43 ]
Позвоночные животные
[ редактировать ]- Лягушка: Ксеноп [ 43 ] ( X. laevis и X. тропический ). [ 44 ] [ 45 ] Хороший запас эмбрионов. Особенно подходит для микрохирургии.
- Рыбка данио : Данио рерио . [ 46 ] Хороший запас эмбрионов. Хорошо развитая генетика.
- Курица: Gallus Gallus . [ 47 ] Ранние стадии аналогичны млекопитающим, но микрохирургия проще. Бюджетный.
- Мышь: Mus musculus . [ 48 ] С мамой [ 43 ] с хорошо развитой генетикой.
Беспозвоночные
[ редактировать ]- Плодовая мушка: Drosophila melanogaster . [ 49 ] Хороший запас эмбрионов. Хорошо развитая генетика.
- Нематода: Caenorhabditis elegans . [ 50 ] Хороший запас эмбрионов. Хорошо развитая генетика. Бюджетный.
Одноклеточный
[ редактировать ]- Водоросли: Хламидомонада [ 43 ]
- Дрожжи: Сахаромицеты [ 43 ]
Другие
[ редактировать ]Также для некоторых целей популярны морские ежи. [ 51 ] [ 43 ] и асцидии . [ 52 ] Для изучения регенерации хвостатых амфибий такие как аксолотль Ambystoma mexicanum , используются [ 53 ] а также планарии, такие как Schmidtea mediterranea . [ 10 ] Органоиды также были продемонстрированы как эффективная модель развития. [ 54 ] Разработка растений была сосредоточена на кресс-салате Arabidopsis thaliana как модельном организме. [ 55 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Вебстер, Джерри; Гудвин, Брайан (13 ноября 1996 г.). «Глава 9 — Генеративная биология» . Форма и трансформация: порождающие и реляционные принципы в биологии . Издательство Кембриджского университета. ISBN 978-0-521-35451-6 .
- ^ «Генеративная биология: разработка биологических лекарств с большей скоростью и успехом» . Амген . 7 июня 2022 г. . Проверено 5 апреля 2024 г.
- ^ «Генеративная биология: учимся программировать клеточные машины» . НАЦИОНАЛЬНЫЕ ИНСТИТУТЫ ЗДРАВООХРАНЕНИЯ США. 15 марта 2024 г. Проверено 5 апреля 2024 г.
- ^ Ли Б., Кэри М., Уоркман Дж.Л. (февраль 2007 г.). «Роль хроматина во время транскрипции» . Клетка . 128 (4): 707–19. дои : 10.1016/j.cell.2007.01.015 . ПМИД 17320508 .
- ^ Хайнцман Н.Д., Стюарт Р.К., Хон Г., Фу Ю., Чинг К.В., Хокинс Р.Д. и др. (март 2007 г.). «Отличные и прогнозируемые хроматиновые сигнатуры промоторов и энхансеров транскрипции в геноме человека». Природная генетика . 39 (3): 311–8. дои : 10.1038/ng1966 . ПМИД 17277777 . S2CID 1595885 .
- ^ Мейнхардт Х, Гирер А (2000). «Формирование паттерна путем локальной самоактивации и латерального торможения» (PDF) . Биоэссе . 22 (8): 753–760. CiteSeerX 10.1.1.477.439 . doi : 10.1002/1521-1878(200008)22:8<753::aid-bies9>3.0.co;2-z . ПМИД 10918306 . Архивировано (PDF) из оригинала 27 октября 2017 г.
- ^ Спринзак Д., Лаханпал А., Лебон Л., Сантат Л.А., Фонтес М.Е., Андерсон Г.А. и др. (май 2010 г.). «Цис-взаимодействия между Notch и Delta генерируют взаимоисключающие состояния сигнализации» . Природа . 465 (7294): 86–90. Бибкод : 2010Natur.465...86S . дои : 10.1038/nature08959 . ПМК 2886601 . ПМИД 20418862 .
- ^ Карлсон Б.М. (2007). Принципы регенеративной биологии . Берлингтон, Массачусетс: Академическая пресса.
- ^ Bosch TC (март 2007 г.). «Почему полипы регенерируют, а мы нет: к клеточной и молекулярной основе регенерации гидры» . Биология развития . 303 (2): 421–33. дои : 10.1016/j.ydbio.2006.12.012 . ПМИД 17234176 .
- ^ Jump up to: а б Реддиен П.В., Санчес Альварадо А (2004). «Основы регенерации планарий». Ежегодный обзор клеточной биологии и биологии развития . 20 : 725–57. doi : 10.1146/annurev.cellbio.20.010403.095114 . ПМИД 15473858 . S2CID 1320382 .
- ^ Вагнер Д.Е., Ван И.Е., Реддиен П.В. (май 2011 г.). «Клоногенные необласты — это плюрипотентные взрослые стволовые клетки, лежащие в основе регенерации планарий» . Наука . 332 (6031): 811–6. Бибкод : 2011Sci...332..811W . дои : 10.1126/science.1203983 . ПМЦ 3338249 . ПМИД 21566185 .
- ^ Накамура Т., Мито Т., Бандо Т., Оучи Х., Нодзи С. (январь 2008 г.). «Рассечение регенерации ног насекомых посредством интерференции РНК» . Клеточные и молекулярные науки о жизни . 65 (1): 64–72. дои : 10.1007/s00018-007-7432-0 . ПМЦ 11131907 . ПМИД 18030418 .
- ^ Саймон А., Танака Э.М. (2013). «Регенерация конечностей». Междисциплинарные обзоры Wiley. Биология развития . 2 (2): 291–300. дои : 10.1002/wdev.73 . ПМИД 24009038 . S2CID 13158705 .
- ^ Слэк Дж. М. (2013). «Глава 20». Основная биология развития . Оксфорд: Уайли-Блэквелл.
- ^ Юнгникель М.К., Саттон К.А., Флорман Х.М. (август 2003 г.). «В начале: уроки оплодотворения у мышей и червей» . Клетка . 114 (4): 401–4. дои : 10.1016/s0092-8674(03)00648-2 . ПМИД 12941269 .
- ^ Jump up to: а б Хакетт Дж.А., Сенгупта Р., Зилич Дж.Дж., Мураками К., Ли С., Даун Т.А., Сурани М.А. (январь 2013 г.). «Динамика деметилирования зародышевой ДНК и стирание отпечатка с помощью 5-гидроксиметилцитозина» . Наука . 339 (6118): 448–52. Бибкод : 2013Sci...339..448H . дои : 10.1126/science.1229277 . ПМЦ 3847602 . ПМИД 23223451 .
- ^ Хайкова П., Джеффрис С.Дж., Ли С., Миллер Н., Джексон С.П., Сурани М.А. (июль 2010 г.). «Полногеномное перепрограммирование в зародышевой линии мыши влечет за собой путь эксцизионной репарации основания» . Наука . 329 (5987): 78–82. Бибкод : 2010Sci...329...78H . дои : 10.1126/science.1187945 . ПМЦ 3863715 . ПМИД 20595612 .
- ^ Стивен Д.Х., изд. (1975). Сравнительная плацентация . Лондон: Академическая пресса.
- ^ Кимельман Д., Мартин Б.Л. (2012). «Передне-задний паттерн в раннем развитии: три стратегии» . Междисциплинарные обзоры Wiley. Биология развития . 1 (2): 253–66. дои : 10.1002/wdev.25 . ПМК 5560123 . ПМИД 23801439 .
- ^ Слэк Дж. М. (1987). «Морфогенетические градиенты - прошлое и настоящее». Тенденции биохимических наук . 12 : 200–204. дои : 10.1016/0968-0004(87)90094-6 .
- ^ Роджерс К.В., Шир А.Ф. (2011). «Градиенты морфогена: от генерации к интерпретации». Ежегодный обзор клеточной биологии и биологии развития . 27 : 377–407. doi : 10.1146/annurev-cellbio-092910-154148 . ПМИД 21801015 . S2CID 21477124 .
- ^ Даманн С., Оутс AC, Бранд М. (январь 2011 г.). «Формирование и поддержание границ в развитии тканей». Обзоры природы. Генетика . 12 (1): 43–55. дои : 10.1038/nrg2902 . ПМИД 21164524 . S2CID 1805261 .
- ^ Хардин Дж., Уолстон Т. (август 2004 г.). «Модели морфогенеза: механизмы и механика клеточной перестройки». Текущее мнение в области генетики и развития . 14 (4): 399–406. дои : 10.1016/j.где.2004.06.008 . ПМИД 15261656 .
- ^ Хаммершмидт М., Ведлих Д. (ноябрь 2008 г.). «Регулируемая адгезия как движущая сила гаструляционных движений» . Разработка . 135 (22): 3625–41. дои : 10.1242/dev.015701 . ПМИД 18952908 .
- ^ Фарж, Эммануэль (2003). «Механическая индукция скручивания передней кишки / зачатка стомодели дрозофилы» . Современная биология . 13 (16): 1365–1377. дои : 10.1016/s0960-9822(03)00576-1 . ПМИД 1293230 .
- ^ Брюне, Тибо; Букле, Адриан; и др. (2013). «Эволюционное сохранение спецификации ранней мезодермы путем механотрансдукции в Bilateria» . Природные коммуникации . 4 : 2821. Бибкод : 2013NatCo...4.2821B . дои : 10.1038/ncomms3821 . ПМК 3868206 . ПМИД 24281726 .
- ^ Нгуен, Нгок-Мин; Мерль, Татьяна; и др. (2022). «Механо-биохимическая морская стимуляция инверсии, гаструляции и спецификации эндомезодермы у многоклеточных эукариот» . Границы клеточной биологии и биологии развития . 10 : 992371. дои : 10.3389/fcell.2022.992371 . ПМЦ 9754125 . ПМИД 36531949 .
- ^ де Люссане, MHE; Оссе, JWM (2012). «Наследственное осевое скручивание объясняет контралатеральную переднюю часть и перекрест зрительных нервов у позвоночных». Биология животных . 62 (2): 193–216. arXiv : 1003.1872 . дои : 10.1163/157075611X617102 . S2CID 7399128 .
- ^ О'Фаррелл PH (2003). «Как многоклеточные животные достигают своего полного размера: естественная история размера». Ин-Холл, Миннесота, Рафф М., Томас Дж. (ред.). Рост клеток: контроль размера клеток . Лабораторный пресс Колд-Спринг-Харбор. стр. 1–21.
- ^ Мосс Э.Г., Ромер-Зайберт Дж. (2014). «Внутреннее клеточное время в развитии животных». Междисциплинарные обзоры Wiley. Биология развития . 3 (5): 365–77. дои : 10.1002/wdev.145 . ПМИД 25124757 . S2CID 29029979 .
- ^ Тата-младший (1996). «Метаморфоз амфибий: изысканная модель гормональной регуляции постэмбрионального развития у позвоночных». Развитие, рост и дифференциация . 38 (3): 223–231. дои : 10.1046/j.1440-169x.1996.t01-2-00001.x . ПМИД 37281700 . S2CID 84081060 .
- ^ Браун Д.Д., Кай Л. (июнь 2007 г.). «Метаморфоза земноводных» . Биология развития . 306 (1): 20–33. дои : 10.1016/j.ydbio.2007.03.021 . ЧВК 1945045 . ПМИД 17449026 .
- ^ Коэн С.М. (1993). «Развитие воображаемого диска». В Бэйт М., Мартинес-Ариас М. (ред.). Развитие дрозофилы melanogaster . Пресса Колд-Спринг-Харбор.
- ^ Мавес Л., Шубигер Г. (октябрь 2003 г.). «Трандетерминация в имагинальных дисках дрозофилы: модель для понимания плюрипотентности и поддержания селекторных генов». Текущее мнение в области генетики и развития . 13 (5): 472–9. дои : 10.1016/j.где.2003.08.006 . ПМИД 14550411 .
- ^ Бёрле I, Ло Т (октябрь 2003 г.). «Апикальные меристемы: источник молодости растения». Обзор. Биоэссе . 25 (10): 961–70. дои : 10.1002/bies.10341 . ПМИД 14505363 .
- ^ Леопольд AC (1964). Рост и развитие растений . Нью-Йорк: МакГроу-Хилл. п. 183 .
- ^ Брэнд Ю, Хобе М, Саймон Р. (февраль 2001 г.). «Функциональные домены в меристемах побегов растений». Обзор. Биоэссе . 23 (2): 134–41. doi : 10.1002/1521-1878(200102)23:2<134::AID-BIES1020>3.0.CO;2-3 . ПМИД 11169586 . S2CID 5833219 .
- ^ Барлоу П. (май 2005 г.). «Определение закономерностей клеток в растительной ткани: вторичная флоэма деревьев». Биоэссе . 27 (5): 533–41. дои : 10.1002/bies.20214 . ПМИД 15832381 .
- ^ Пасифичи Э., Ди Мамбро Р., Делло Иоио Р., Константино П., Сабатини С. (август 2018 г.). «Корень арабидопсиса» . Журнал ЭМБО . 37 (16). дои : 10.15252/embj.201899134 . ПМК 6092616 . ПМИД 30012836 .
- ^ Росс С.Д., Фарис Р.П., Биндер В.Д. (1983). «Регуляторы роста и хвойные деревья: их физиология и потенциальное использование в лесном хозяйстве». В Nickell LG (ред.). Химические вещества, регулирующие рост растений . Том. 2. Бока-Ратон, Флорида: CRC Press. стр. 35–78.
- ^ Цзинь Дж, Хэ К, Тан Икс, Ли З, Лу Л, Чжао Ю и др. (июль 2015 г.). «Карта регуляции транскрипции арабидопсиса раскрывает отличительные функциональные и эволюционные особенности новых факторов транскрипции» . Молекулярная биология и эволюция . 32 (7): 1767–73. дои : 10.1093/molbev/msv058 . ПМЦ 4476157 . ПМИД 25750178 . Архивировано из оригинала 2 июня 2016 г.
- ^ Пирес Н.Д., Долан Л. (февраль 2012 г.). «Морфологическая эволюция наземных растений: новые конструкции со старыми генами» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 367 (1588): 508–518. дои : 10.1098/rstb.2011.0252 . ПМЦ 3248709 . ПМИД 22232763 .
- ^ Jump up to: а б с д и ж Фридман, Уильям Э. (1999). «Экспрессия клеточного цикла в сперме Arabidopsis : значение для понимания закономерностей гаметогенеза и оплодотворения у растений и других эукариот». Разработка . 126 (5). Компания биологов : 1065–75. дои : 10.1242/dev.126.5.1065 . ISSN 0950-1991 . ПМИД 9927606 . S2CID 13397345 .
- ^ Ньюкуп П.Д., Фабер Дж. (1967). Обычный стол Xenopus laevis (Даудин) . Северная Голландия, Амстердам.
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Харланд Р.М., Грейнджер Р.М. (декабрь 2011 г.). «Исследования Xenopus: метаморфизованные генетикой и геномикой» . Тенденции в генетике . 27 (12): 507–15. дои : 10.1016/j.tig.2011.08.003 . ПМК 3601910 . ПМИД 21963197 .
- ^ Лоусон Н.Д., Вулф С.А. (июль 2011 г.). «Прямой и обратный генетические подходы к анализу развития позвоночных рыбок данио» . Развивающая клетка . 21 (1): 48–64. дои : 10.1016/j.devcel.2011.06.007 . ПМИД 21763608 .
- ^ Рашиди Х., Соттиле В. (апрель 2009 г.). «Куриный эмбрион: создание модели для современных биомедицинских исследований». Биоэссе . 31 (4): 459–65. doi : 10.1002/bies.200800168 . ПМИД 19274658 . S2CID 5489431 .
- ^ Берингер Р., Герценштайн М., Винтерстен К., Надь М. (2014). Манипулирование мышиным эмбрионом. Лабораторное руководство (Четвертое изд.). Колд-Спринг-Харбор, Нью-Йорк: Лабораторное издательство Колд-Спринг-Харбор.
- ^ Сент-Джонстон Д. (март 2002 г.). «Искусство и дизайн генетических экранов: Drosophila melanogaster». Обзоры природы. Генетика . 3 (3): 176–88. дои : 10.1038/nrg751 . ПМИД 11972155 . S2CID 195368351 .
- ^ Риддл Д.Л., Блюменталь Т., Мейер Б.Дж., Присс-младший (1997). К.элеганс II . Колд-Спринг-Харбор, Нью-Йорк: Лабораторное издательство Колд-Спринг-Харбор.
- ^ Эттенсон, Калифорния, Sweet HC (2000). «Создание образца раннего эмбриона морского ежа» . Текущие темы биологии развития, том 50 . Том. 50. Академическая пресса. стр. 1–44 . дои : 10.1016/S0070-2153(00)50002-7 . ISBN 9780121531508 . ПМИД 10948448 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Лемэр П. (июнь 2011 г.). «Эволюционный перекресток биологии развития: оболочники» . Разработка . 138 (11): 2143–52. дои : 10.1242/dev.048975 . ПМИД 21558365 .
- ^ Наку Э., Танака Э.М. (2011). «Регенерация конечностей: новая разработка?». Ежегодный обзор клеточной биологии и биологии развития . 27 : 409–40. doi : 10.1146/annurev-cellbio-092910-154115 . ПМИД 21801016 .
- ^ Адер М., Танака Э.М. (декабрь 2014 г.). «Моделирование человеческого развития в 3D-культуре». Современное мнение в области клеточной биологии . 31 : 23–8. дои : 10.1016/j.ceb.2014.06.013 . ПМИД 25033469 .
- ^ Вайгель Д., Глейзбрук Дж. (2002). Арабидопсис. Лабораторное руководство . Колд-Спринг-Харбор, Нью-Йорк: Лабораторное издательство Колд-Спринг-Харбор.
Дальнейшее чтение
[ редактировать ]- Гилберт С.Ф. , Баррези М. (2023). Биология развития (13-е изд.). Нью-Йорк: Издательство Оксфордского университета . ISBN 9780197574591 .
- Слэк Дж. М. (2013). Основная биология развития . Оксфорд: Уайли-Блэквелл.
- Вулперт Л., Тикл С. (2011). Принципы развития . Оксфорд и Нью-Йорк: Издательство Оксфордского университета.