Теория осевого поворота
| Осевое скручивание | |
|---|---|
 Схема развития осевой крутки. Фазы развития (сверху вниз): (1) поворот эмбриона на левый бок; (2) передняя часть головы растет в том же направлении, но остальная часть тела растет в противоположном направлении и закручивается. Так что в конечном итоге (3) внешняя двусторонняя симметрия восстанавливается. Обратите внимание: эволюционное давление на внутреннюю симметрию отсутствует, поэтому сердце (и другие органы) остаются асимметричными. | |
| Подробности | |
| Система | план тела позвоночных |
| Анатомическая терминология | |
Теория осевого скручивания ( также известная как осевого скручивания гипотеза ) — это научная теория, выдвинутая для объяснения ряда необычных аспектов строения тела ( позвоночных включая человека ). [1] Предполагается, что ростральная часть головы «повернута» относительно остального тела. [2] Эта конечная часть состоит из лица (глаза, нос и рот), а также части мозга ( мозг и таламус ). Согласно теории, тело позвоночных имеет левостороннюю хиральность .
Теория осевого скручивания конкурирует с рядом других предложений, которые фокусируются на более ограниченных, конкретных аспектах (см. раздел #Отношение к другим теориям и гипотезам ), большинство из которых объясняют контралатеральную организацию переднего мозга , феномен, которым в основном управляет левое полушарие мозга. правая сторона тела и наоборот. [3] Ни одна из предложенных теорий, объясняющих это явление, включая теорию осевого скручивания, не получила всеобщего признания. [4] Генетическая основа, лежащая в основе такого поворота развития, еще не изучена.
скручивание между концом головы и остальной частью тела Теория осевого скручивания объясняет различные явления и рассматривает, как и когда развивается . Он также обращается к возможной эволюционной истории. Одним из предсказаний теории была аурофациальная асимметрия, которая затем была обнаружена эмпирически . [5] хотя и автором, который также был соавтором оригинальной теории.
Феномены, которые теория может объяснить, включают:
- Контралатеральная организация мозга
- Левосторонняя ориентация сердца.
- Асимметричное положение желудочно-кишечного тракта , печени и поджелудочной железы.
- Оптический перекрест
- перекрест блокового нерва
- Непересеченный обонятельный тракт
- Аурофациальная асимметрия
- Yakovlevian torque
- Асимметрия грудного позвонка
Согласно модели развития осевого поворота , передняя часть головы поворачивается против остального тела, за исключением внутренних органов . Из-за этого поворота передний мозг и лицо поворачиваются так, что лево и право, а также передняя и задняя части позвоночных переворачиваются у взрослых .
История
[ редактировать ]В конце XIX века известный нейробиолог и Нобелевской премии лауреат Сантьяго Рамон-и-Кахаль предложил теорию, объясняющую контралатеральную организацию переднего мозга, которая быстро и широко получила признание. [6] [4] Эта теория визуальных карт предполагает, что перекрест зрительных нервов восстанавливает изображение сетчатки в зрительной коре. [7] Теория Кахаля оставалась практически неоспоримой более века. [8] [9] Хотя были выдвинуты и другие идеи (см. раздел #Отношение к другим теориям и гипотезам ).
Эмбриология
[ редактировать ]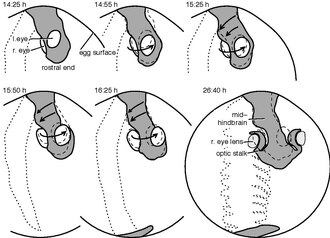
Хотя эмбриологическое развитие осевого поворота подробно не изучалось, имеются четкие указания на рыбок данио и птенцов . [1] Скручивание начинается вскоре после нейруляции и начинается в рострокаудальном (от передней части хвоста) направлении. [10]
Группа Филиппа Келлера отслеживала каждую клетку развивающихся эмбрионов рыбок данио до первого сердцебиения. [11] Прослеживая движения клеток в будущей области глаз и задней части головы, выявили противоположные направления движения в соответствии с осевым поворотом. В то время как клетки области левого глаза имели тенденцию двигаться наружу и вниз ( вентрально ), клетки области правого глаза перемещались наружу и вверх, как показано на замедленном видео . [12] С другой стороны, поверхностные клетки задней стороны головы сместились влево, что соответствует осевому повороту. [1]
Развитие птенцов хорошо изучено. Развитие принято описывать по стадиям Гамбургера-Гамильтона . [13] Скручивание начинается на стадии 6 на ростральной стороне области головы. [14] и начинается до 14 стадии в направлении области сердца. [10] В то время как передняя область головы вращается, причем правая сторона движется вверх, а левая сторона вниз, область сердца движется в противоположном направлении. В конце концов птенец переворачивается на правый бок, а сердце, не принимавшее участия в перекручивании, приземляется на левую сторону тела. [1]
Генетические механизмы
[ редактировать ]Осевой поворот возникает в результате противоположного асимметричного развития. Это можно наблюдать как волну, движущуюся по эмбриону спереди назад. Сейчас хорошо установлено, что Nodal сигнальный каскад и поток справа налево, производимый реснитчатыми клетками в первичной полоске, являются центральными в создании асимметричной организации. Три аспекта этой волны роста: [1]
- асимметричный рост (поворот влево) на переднем конце примитивного узла (он же органайзер Шпемана-Мангольда у амфибий или узел Гензена у птиц), [14] [15]
- лево-правое расположение органов и
- поворот тела на правую сторону.
Nodal, FGF8 и shh , а также моторный белок Kif5c были связаны с асимметричным ростом переднего примитивного узла, хотя только Nodal, по-видимому, экспрессируется до начала асимметрии. [15] [16] Ген Nodal и ген цинкового пальца ( cSnR ) контролируют асимметричное развитие сердца. [17] Поворот тела вправо тесно связан с теми же генетическими механизмами. [17]
Пороки развития
[ редактировать ]При голопрозэнцефалии полушария головного мозга (или его часть) располагаются не на левой и правой сторонах, а только на лобной и затылочной сторонах черепа, а голова обычно остается очень маленькой. Согласно теории осевого скручивания, это представляет собой крайний случай крутящего момента Яковлева : [18] иможет возникнуть, когда головной мозг не поворачивается на ранних стадиях эмбриологии. [1]
Цефалопагические близнецы, или близнецы джаницепсы, — это сиамские близнецы , рождающиеся с двумя лицами, по одному на каждой стороне головы. У этих близнецов два головного и два спинного мозга, но они расположены на левой и правой сторонах тела. [19] Согласно модели осевого скручивания, две нервные системы не могли поворачиваться из-за сложной конфигурации тела и поэтому оставались по обе стороны. [1]
Эволюция
[ редактировать ]Считается, что осевое скручивание развилось у общего предка всех позвоночных, но механизм остается спекулятивным. [1] Анатомия указывает на то, что все позвоночные искривлены, но также большинство сестринских клад позвоночных демонстрируют сходные формы асимметричного, искривленного развития (см. Ниже).
Даже самые отдаленные группы позвоночных — бесчелюстные миноги и миксины — обладают перекрестом зрительных нервов и контралатеральной организацией мозга. [20] а также левостороннее сердце и асимметричный кишечник. [21] Кроме того, у каждого позвоночного имеется контралатеральная организация переднего мозга. [20] Ископаемые отпечатки черепов ранних позвоночных ордовика и более позднего периода показывают наличие перекреста зрительных нервов. [22]
Скручивание и асимметричное развитие хорошо известны у других вторичноротых — таких как Hemichordata , Echinodermata , Cephalochordata и Tunicata . Поворот в сторону или вверх ногами встречается также в этих кладах:
- Морские звезды поворачивают рот вниз после того, как личинка ненадолго прикрепилась к субстрату с помощью рудиментарного стебелька, рот обращен вверх. [23]
- Взрослый ланцетник зарывается спиной вниз и вытянутым ртом (ср. Lancelet#Habitat ).
Некоторые виды рыб склонны разворачиваться при кормлении с поверхности воды.
Морфология
[ редактировать ]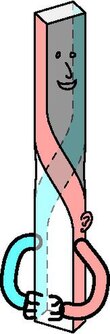
Осевое скручивание происходит у ранних эмбрионов позвоночных. существует эволюционное давление У животных в сторону двусторонней симметрии из-за полового отбора (лучший взгляд на потенциальных партнеров) и функционального отбора (например, лучшее передвижение). Эволюционное давление уменьшается с улучшением симметрии. Соответственно, давление снижается, поскольку часть тела меньше связана с поверхностью тела (меньше полового отбора) и опорно-двигательной системой (меньше функционального отбора). [5] Следовательно, теория осевого скручивания предсказывает, что небольшие систематические асимметрии остаются снаружи тела и что эти асимметрии больше внутри тела. [1] [24] [5]
Крутящий момент мозга и асимметрия позвоночника
[ редактировать ]
Передний мозг ( большой мозг и таламус ) преимущественно представляет противоположную сторону тела (и зрительного мира). Однако для контроля движений обычно требуется информация с обеих сторон тела, поэтому контралатеральное представление ни в коем случае не является абсолютным . Скорее, почти каждая область мозга связана с обеими сторонами тела. [27]
The Yakovlevian torque [25] (он же «крутящий момент мозга против часовой стрелки» [28] относится к анатомической особенности нормального мозга. В среднем лобные доли асимметричны влево (правая доля кажется немного больше левой), тогда как затылочная доля асимметрична вправо; центральная борозда и височная доля правого коркового полушария расположены дальше вперед, чем левые. В целом, эти асимметрии эквивалентны небольшому вращению головного мозга (т.е. крутящему моменту). Такое вращение в точности соответствует теории осевого скручивания, учитывая, что головной мозг не является поверхностной структурой.
Крутящий момент также известен как «затылочный изгиб». [29] если на затылочной стороне он выражен сильнее, чем на лобной.
Позвоночник слегка асимметричен. У здоровых людей грудные позвонки (позвонки Т6-Т12) были в среднем асимметричными, так что средняя линия была направлена вправо (2,5° в Т6). [26] Таким образом, крутящий момент Яковлева и асимметрия позвоночника направлены в противоположные стороны, как и предсказывает теория осевого скручивания.
Дискуссии по центральной нервной системе
[ редактировать ]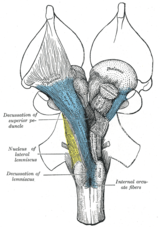

Если правый передний мозг представляет собой преимущественно левое тело, а левый передний мозг — правое тело, то позади переднего мозга должны быть места пересечения основных нервов.
Анатомически контралатеральная организация переднего мозга проявляется в виде крупных перекрестов (на основе латинского обозначения десяти , «дека», как заглавной буквы «Х») и хиазм (от греческой заглавной буквы «Х», хи ). Пересечение означает пересечение пучков аксональных волокон внутри центральной нервной системы . В результате таких перекрестов: эфферентные связи головного мозга с базальными ганглиями , мозжечком и спинным мозгом перекрещиваются ; и афферентные связи от позвоночника, мозжечка и моста к таламусу пересекаются. [20] Таким образом, моторные, соматосенсорные , слуховые и зрительные первичные области переднего мозга преимущественно представляют контрлатеральную сторону тела.
Большинство афферентных и эфферентных связей переднего мозга имеют двусторонние компоненты, особенно за пределами первичных сенсорных и двигательных областей. [27]
Визуальная система
[ редактировать ]В зрительной системе глаза и мышцы располагаются перед поворотом, а сенсорный и двигательный центры — позади поворота. Таким образом, предполагается, что развивающиеся нервы будут искать место прикрепления на противоположной стороне от своего начала.
четыре черепных нерва Непосредственно глаз обслуживают : один сенсорный и три двигательных нерва. Зрительный нерв является чувствительным и пересекает срединную линию перекреста зрительных нервов . Глазодвигательный нерв , блоковый нерв и отводящий нерв — это двигательные нервы, которые контролируют одну или несколько глазных мышц. Глазодвигательный нерв пересекает срединную линию, прежде чем покинуть центральную нервную систему. Блоковый нерв пересекает срединную линию в перекресте на дорсальной стороне, а отводящая мышца иннервирует глазную мышцу на той же стороне. [30] : Рис. 17.8
В свете теории осевого скручивания эту сложную картину можно понять. Глаза, как и рот и нос, берут начало в переднем отделе головы, т. е. перед заворотом. Единственный черепной нерв, берущий начало от переднего мозга, — это обонятельный нерв (см. ниже). Все остальные черепные нервы берут начало из участков центральной нервной системы, лежащих за завитком. [1]
Зрительный нерв прикрепляется к зрительной покрышке среднего мозга . У четвероногих и костистых рыб он также ответвляется от LGN таламуса в переднем мозге, но не у других позвоночных, таких как акулы и скаты . У акул зрительный центр головного мозга получает волокна из зрительной оболочки. По пути эти волокна снова пересекают срединную линию, так что каждое полушарие головного мозга акул представляет собой глаз на одной и той же стороне. [31] [32] Таким образом, зрительный тракт во многом следует предсказаниям теории осевого скручивания. Ветвь к LGN существует только у четвероногих. [30] и поэтому обычно понимается как приобретенное на более позднем этапе эволюции позвоночных и, таким образом, составляет исключение (см. Также Контралатеральный мозг # Контралатеральная организация неполная ).
расположено Отводящее ядро в мосту . Отводящий нерв иннервирует латеральную прямую мышцу глаза у большинства позвоночных, за исключением миног и миксин. [21] Таким образом, кажется, что латеральная прямая мышца развилась позже и независимо от других глазных мышц и представляет собой исключение из модели осевого поворота. [1]
Обонятельная система
[ редактировать ]
Обонятельные пути идут параллельно зрительному тракту, но не образуют перекреста. Соответственно, каждая обонятельная луковица соединяется с односторонними центрами лобного мозга. Это полностью согласуется с теорией осевого скручивания, поскольку нос является частью передней области головы, которая скручивается вместе с передним мозгом. Поскольку первичные обонятельные центры находятся в головном мозге (обоняние — единственное чувство, которое зарождается в головном мозге), предполагается, что каждая обонятельная доля будет представлена мозгом на одной и той же стороне, что действительно так: тогда как обонятельные тракты Соседствуя непосредственно со зрительными путями, они не хиазмируются, а прикрепляются на ипсилатеральной стороне головного мозга (см. рисунок и раздел Обонятельная система # Клиническое значение ).
Аурофациальная асимметрия
[ редактировать ]
Аурофациальная асимметрия выражает положение лица (глаз, носа, рта) относительно плоскости, перпендикулярной оси через уши. Как показано на графике, асимметрия уменьшается до 13 лет. Поскольку осевой поворот расположен между ушами и лицом, прогнозируется, что лицо растет слева к средней линии, что и есть на самом деле. [5]
Ориентация внутренних органов
[ редактировать ]
Внутренние органы туловища — это области тела, которые меньше всего механически связаны с передвижением и внешним телом, и поэтому теория осевого поворота предсказывает, что они являются наиболее асимметричными областями тела. Другие двусторонне-симметричные животные, такие как насекомые и кольчатые черви, также двусторонне симметричны и внутри. Асимметричное развитие сердца хорошо изучено. [17] [33]
Вопрос о том, почему сердце должно иметь левостороннюю ориентацию, был темой научных исследований до того, как была опубликована теория осевого поворота, но ни одна из гипотез не выдержала критической проверки. [34]
Асимметричная ориентация и положение желудочно-кишечного тракта возникает при развитии путем ротации (см. Развитие пищеварительной системы ). Латеральное расположение органов пищеварения у всех позвоночных животных находится на одной стороне: печень и желчный пузырь — справа, а желудок , поджелудочная железа , селезенка и конечная петля толстой кишки — слева. [35]
Связь с другими теориями и гипотезами
[ редактировать ]Других теорий или гипотез, объясняющих весь спектр, охватываемый теорией осевого скручивания, не существует, но ряд теорий и гипотез рассматривают отдельные аспекты на различной глубине.
Гипотеза инверсии
[ редактировать ]В 1822 году французский зоолог Этьен Жоффруа Сен-Илер заметил, что организация спинных и вентральных структур у раков ( членистоногих ) противоположна организации млекопитающих, и предположил, что млекопитающие и другие позвоночные животные перевернуты вверх ногами. [36] [37] Как объяснялось выше, Марсель Кинсборн предположил, что перевернуто тело (сома), но не передняя часть головы (отсюда и гипотеза соматического скручивания). [2]
Молекулярные доказательства гипотезы инверсии имеются почти во всех группах вторичноротых. [38] [39]
Гипотеза соматического поворота
[ редактировать ]Гипотеза соматического скручивания Марселя Кинсборна наиболее тесно связана с теорией осевого скручивания. как улучшение гипотезы дорсовентральной инверсии. начала XIX века Обе теории были представлены натуралистом Этьеном Жоффруа Сен-Илером [1] [2] В отличие от осевого поворота, соматический поворот не затрагивает аспекты развития. Кроме того, это объясняет только контралатеральный мозг, но не ориентацию внутренних органов. [24]
Асимметрия позвоночника
[ редактировать ]Никаких гипотез об асимметрии позвоночника не опубликовано. Другой теории, объясняющей левое положение сердца или асимметричную ориентацию кишечника и связанных с ним органов, не существует. [26] [5]
Контралатеральный передний мозг
[ редактировать ]Несколько гипотез предложили функцию (определенных аспектов) контралатерального мозга (т.е. левый передний мозг представляет в основном правое тело, а правый передний мозг представляет в основном левое тело). Самый известный из них - Кахаля.
Теория визуальных карт Кахаля
[ редактировать ]Теория визуальных карт Сантьяго Рамона-и-Кахаля предполагает, что перекрест зрительных нервов восстанавливает изображение сетчатки в зрительной коре. [7] [8] Несмотря на то, что теория все еще поддерживается, [4] несколько исследований указали на серьезные недостатки теории. [1] [2] [40] Самое главное, что петля оптического излучения отменяет потенциальное восстановление оптической проекции на коре, что является центральной идеей теории. Кроме того, эта теория не справедлива для важных групп позвоночных, таких как акулы, поскольку их головной мозг имеет ипсилатеральное зрительное представление, но контралатеральное соматосенсорное и моторное представительство. [31] [32]
Другие функциональные гипотезы
[ редактировать ]Функциональный [41] [42] или топологический [43] объяснения терпят неудачу, потому что односторонние связи так же важны, как и пересекающиеся. [44] Более того, эти объяснения оставляют открытым вопрос, почему ствол мозга и мозжечок имеют одностороннюю организацию тела. [1]
Латерализация мозга
[ редактировать ]Часто полагают, что крутящий момент Яковлева отражает латерализацию определенных функций мозга. [25]
Теория парцелляции предполагает, что увеличение размера мозга может сохранить случайную контралатеральную организацию. [27] но не объясняет ни перекрест зрительных нервов, ни то, что у большинства позвоночных довольно маленький передний мозг, особенно у ранних форм. [45]
Открытые вопросы
[ редактировать ]Теория осевого скручивания — новая научная дисциплина, и в настоящее время очень немногие научные статьи посвящены ей напрямую. [1] [2] [24] [5] Хотя существует значительный объем исследований генетических и эмбриологических механизмов асимметричного развития, остается открытым вопрос, как инициируется поворот и как устанавливается инверсия осей лево-право и вверх-вниз в передней области головы.
Эмбриология скручивания у птенцов и рыбок данио изучена лишь в зачаточном состоянии. [1] Различия во времени и механизмах между кладами позвоночных совершенно неизвестны.
Эволюция осевого скручивания остается открытым вопросом. Основатели идеи осевого скручивания (де Люссане и Оссе и Кинсборн) согласны с тем, что осевое скручивание универсально для позвоночных и, вероятно, является особенностью всех хордовых. [1] [2] Хотя асимметричное развитие других хордовых, таких как ланцетник, было подробно изучено , ни одно исследование не анализировало это развитие в свете теории осевого скручивания. Более того, даже другие вторичноротые, т. е. иглокожие ( морские звезды , морские лилии и т. д.), демонстрируют заметное асимметричное развитие и даже осевое закручивание. [46] Этот поворот имеет поразительное сходство с таковым у позвоночных, но в настоящее время этому не посвящено ни одно исследование. Наконец, асимметричное и искривленное развитие хорошо известно у брюхоногих моллюсков, и важным вопросом является связь с асимметричным развитием у позвоночных.
Было высказано предположение, что проблемы с развитием осевого скручивания могут играть центральную роль в пороках развития, таких как голопрозэнцефалия. [1] и сколиоз [5] но они не были рассмотрены.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с де Люссане, MHE; Оссе, JWM (2012). «Наследственное осевое скручивание объясняет контралатеральный передний мозг и перекрест зрительных нервов у позвоночных». Биология животных . 62 (2): 193–216. arXiv : 1003.1872 . дои : 10.1163/157075611X617102 . S2CID 7399128 .
- ^ Jump up to: а б с д и ж Кинсборн, М. (2013). «Соматический поворот: модель эволюции перекреста». Нейропсихология . 27 (5): 511–515. дои : 10.1037/a0033662 . ПМИД 24040928 . S2CID 11646580 .
- ^ Виджано, Д.; Пироло, Л. (2002). «Апробация модели формирования перекреста зрительных нервов у человека». Мозговой Рес. Бык . 59 (2): 111–115. дои : 10.1016/S0361-9230(02)00846-8 . ПМИД 12379441 . S2CID 2687785 .
- ^ Jump up to: а б с Мора, Карла; Веласкес, Карлос; Мартино, Хуан (01 сентября 2019 г.). «Теория пересечения срединной линии нервных путей: исторический анализ вклада Сантьяго Рамона-и-Кахаля в церебральную локализацию и контрлатеральную организацию переднего мозга» . Нейрохирургический фокус . 47 (3): Е10. дои : 10.3171/2019.6.FOCUS19341 . ISSN 1092-0684 . ПМИД 31473669 .
- ^ Jump up to: а б с д и ж г час я де Люссане, MHE (2019). «Противоположная асимметрия лица и туловища, а также поцелуев и объятий, как предсказывает гипотеза осевого скручивания» . ПерДж . 7 : е7096. дои : 10.7717/peerj.7096 . ПМК 6557252 . ПМИД 31211022 .
- ^ Флексиг, Пол (1899). «Предисловие к строению перекреста зрительных нервов». Преподобный Трим. Микрографика (на немецком языке). 3 . Издатель Джо А. Барт: III – VI.
- ^ Jump up to: а б Рамон-и-Кахаль, Сантьяго (1898 г.). «Строение Chiasma opticum и общая теория пересечения нервных путей» [Die Structur des Chiasma opticum nebst einer allgemeine Theorie der Kreuzung der Nervenbahnen (нем., 1899, Verlag Joh. A. Barth)]. Преподобный Трим. Микрографика (на испанском языке). 3 : 15–65.
- ^ Jump up to: а б Ллинас, Р.Р. (2003). «Вклад Сантьяго Рамона-и-Кахаля в функциональную нейробиологию». Нат. Преподобный Нейроски . 4 (1): 77–80. дои : 10.1038/nrn1011 . ПМИД 12511864 . S2CID 30442863 .
- ^ Вуллемоз, Серж; Ренето, Оливье; Жабодон, Денис (2005). «Выход за пределы средней линии: почему человеческий мозг переплетен?». Ланцет Нейрол . 4 (2): 87–99. дои : 10.1016/S1474-4422(05)00990-7 .
- ^ Jump up to: а б Беллэрс, Рут; Осмонд, Марк (2005). Атлас развития цыплят (2-е изд.). Нью-Йорк: Академическая пресса. п. 172.
- ^ Келлер, Филипп Дж.; Шмидт, Аннетт Д.; Витбродт, Иоахим; Стельцер, Эрнст Х.К. (2008). «Реконструкция раннего эмбрионального развития рыбок данио с помощью сканирующей световой листовой микроскопии» . Наука . 322 (5904): 1065–1069. Бибкод : 2008Sci...322.1065K . дои : 10.1126/science.1162493 . ПМИД 18845710 . S2CID 7594561 .
- ^ де Люссане, Марк HE (8 сентября 2023 г.). Фильм об эмбриологических изменениях эмбриона рыбы-зебры, выявленных по моделям клеточных движений (Quicktime Movie). Фиговая доля. doi : 10.6084/m9.figshare.24104085 .
- ^ Гамбургер, Виктор; Гамильтон, Ховард Л. (1951). «Ряд нормальных стадий развития куриного эмбриона» . Дев. Дин . 88 (4): 49–92. дои : 10.1002/aja.1001950404 . ПМИД 1304821 . S2CID 32593520 .
- ^ Jump up to: а б Лопес-Санчес, Кармен; Гарсия-Мартинес, Вирджиния; Шенвольф, Гэри К. (2001). «Локализация клеток проспективной нервной пластинки, сердца и сомитов в первичной полоске и эпибласте птичьих эмбрионов на промежуточных стадиях примитивной полоски». Клетки Ткани Органы . 169 (4): 334–346. дои : 10.1159/000047900 . ПМИД 11490112 . S2CID 238761 .
- ^ Jump up to: а б Пипер, Тобиас; Карпай, Мериам; Райнерманн, Йоханна; Сурчев, Лачезар; Вибан, Кристоф; Циколия, Николоз (2020). «Заполненные матрицей микрополости в формирующемся птичьем лево-правом органайзере» . Дев. Дин . 249 (4): 496–508. дои : 10.1002/dvdy.133 . ПМИД 31729123 . S2CID 208037861 .
- ^ Циколия, Н; Шредер, С; Шварц, П; Вибан, К. (2012). «Параксиальная левосторонняя узловая экспрессия и начало формирования лево-правого паттерна у раннего куриного эмбриона». Дифференциация . 84 (5): 380–391. doi : 10.1016/j.diff.2012.09.001 . ПМИД 23142734 .
- ^ Jump up to: а б с Исаак, Элисон; Сарджент, Майкл Г.; Кук, Джонатан (1997). «Контроль лево-правой асимметрии позвоночных с помощью гена цинкового пальца, связанного с улиткой». Наука . 275 (5304): 1301–1304. дои : 10.1126/science.275.5304.1301 . ПМИД 9036854 . S2CID 19753550 .
- ^ Тога, AW; Томпсон, премьер-министр (2003). «Картирование асимметрии мозга». Нат. Преподобный Нейроски . 4 (1): 37–48. дои : 10.1038/nrn1009 . ПМИД 12511860 . S2CID 15867592 .
- ^ Виджано, Д.; Пироло, Л. (2002). «Апробация модели формирования перекреста зрительных нервов у человека». Мозговой Рес. Бык . 59 (2): 111–115. дои : 10.1016/S0361-9230(02)00846-8 . ПМИД 12379441 . S2CID 2687785 .
- ^ Jump up to: а б с Ньювенхейс, Р.; Донкелаар, HJ; Николсон, К.; Смитс, WJAJ; Вихт, Х. (1998). Центральная нервная система позвоночных . Нью-Йорк: Спрингер. ISBN 9783642621277 .
- ^ Jump up to: а б Маринелли, Уильям; Стренгер, Аннелизе (1956). Сравнительная анатомия и морфология позвоночных, 1. Lampetra fluviatilis (L.) 2. Myxine Glutinosa (L.) (на немецком языке). Вена: Франц Дойтике. п. 172.
- ^ Жанвье, П. (1996). Ранние позвоночные . Нью-Йорк: Clarendon Press, Oxford University Press. ISBN 978-0198540472 .
- ^ Геммилл, Джеймс Ф. (1920). «Развитие морской звезды Crossaster papposus , Мюллера и Трошеля». QJ Микроск. Наука . с2-64 (254): 155–187.
- ^ Jump up to: а б с де Люссане, MHE; Оссе, JWM (2015). «Пересечение как осевое скручивание: комментарий к Кинсборну (2013)». Нейропсихология . 29 (5): 713–714. дои : 10.1037/neu0000163 . ПМИД 25528610 .
- ^ Jump up to: а б с Тога, Артур В.; Томпсон, Пол М. (2003). «Картирование асимметрии мозга». Нат. Преподобный Нейроски . 4 (1): 37–48. дои : 10.1038/nrn1009 . ПМИД 12511860 . S2CID 15867592 .
- ^ Jump up to: а б с Кувенховен, Ян-Виллем; Винкен, Коэн Л.; Бартельс, Ламбертус В.; Кастельен, Рене М. (2006). «Анализ существовавшего ранее вращения позвонков в нормальном позвоночнике». Позвоночник . 31 (13): 1467–1472. дои : 10.1097/01.brs.0000219938.14686.b3 . ПМИД 16741456 . S2CID 2401041 .
- ^ Jump up to: а б с Эббесон, Свен О.Э. (1980). «Теория парцелляции и ее связь с межвидовой изменчивостью в организации мозга, эволюционным и онтогенетическим развитием и пластичностью нейронов». Ресурсы клеточных тканей . 213 (2): 179–212. дои : 10.1007/BF00234781 . ПМИД 7459999 .
- ^ Мок, Джеффри; Задина, Джанет; Кори, Дэвид; Коэн, Джереми; Лемен, Лиза; Фундас, Энн (2012). «Атипичное перенапряжение мозга у мальчиков с заиканием в процессе развития» . Развивающая нейропсихология . 37 (5): 434–452. дои : 10.1080/87565641.2012.661816 . ПМЦ 5537737 . ПМИД 22799762 .
- ^ Маллер, Джером Дж.; Андерсон, Родни; Томсон, Ричард Х.; Розенфельд, Джеффри В .; Даскалакис, Зафирис Дж.; Фицджеральд, Пол Б. (2015). «Затылочный изгиб (тормор Яковлева) при биполярной депрессии». Психиатрические исследования: нейровизуализация . 231 (1): 8–14. doi : 10.1016/j.pscychresns.2014.11.008 . ISSN 0925-4927 . ПМИД 25480522 . S2CID 37224862 .
- ^ Jump up to: а б Ньювенхейс, Рудольф; Гардиан, Ян; ван Хейзен, К. (2008). Центральная нервная система человека (4-е изд.). Берлин: Шпрингер.
- ^ Jump up to: а б Люитен, PGM (1981). «Два зрительных пути к конечному мозгу акулы-няньки (Ginglymostoma cirratum). I. Проекции сетчатки». Дж. Комп. Нейрол . 96 (4): 531–538. дои : 10.1002/cne.901960402 . ПМИД 7204669 . S2CID 36572723 .
- ^ Jump up to: а б Люитен, PGM (1981). «Два зрительных пути к телэнцефалону у акулы-няньки (Ginglymostoma cirratum). II. Восходящие таламо-теленцефальные соединения». Дж. Комп. Нейрол . 96 (4): 539–548. дои : 10.1002/cne.901960403 . ПМИД 7204670 . S2CID 13333558 .
- ^ Кидокоро, Хинако; Сайджо, Юкио; Шенвольф, Гэри К. (2022). «Передача узловых сигналов регулирует асимметричное клеточное поведение, приводя в движение сердечную трубку по часовой стрелке у рыбок данио» . Коммуникационная биология . 5 (1): 996. doi : 10.1038/s42003-022-03826-7 . ПМЦ 9492702 . ПМИД 36131094 .
- ^ Воронов Дмитрий А.; Алфорд, Патрик В.; Сюй, Банда; Табер, Ларри А. (2004). «Роль механических сил в правостороннем вращении во время сердечной петли у куриного эмбриона». Биология развития . 272 (2): 339–350. дои : 10.1016/j.ydbio.2004.04.033 .
- ^ Вестхайде, Вильфрид; Ригер, Рейнхард М. (2004). Специальная зоология, Часть 2: Позвоночные животные и черепа (на немецком языке). Гейдельберг: Академическое издательство «Спектр». п. 712. ИСБН 9783827415752 .
- ^ Жоффруа-Сен-Илер, Этьен (1822 г.). «Общие соображения о позвонке». Мемуары Музея естественной истории (на французском языке). 9 :89–119.
- ^ Диксон, А. Фрэнсис (1907). «Почему большие двигательные и сенсорные пути центральной нервной системы перекрещены?» . Дублинский журнал медицинских наук . 124 (1): 1–4. дои : 10.1007/BF02972358 . S2CID 76086581 .
- ^ Нюблер-Юнг, Катарина; Арендт, Детлев (1996). «Энтеропнеусты и эволюция хордовых» . Современная биология . 6 (4): 352–353. Бибкод : 1996CBio....6..352N . дои : 10.1016/S0960-9822(02)00491-8 . ПМИД 8723329 . S2CID 8964529 .
- ^ Арендт, Детлев; Нюблер-Юнг, Катарина (1994). «Инверсия дорсовентральной оси?» . Природа . 371 (6492): 26. Бибкод : 1994Natur.371...26A . дои : 10.1038/371026a0 . ПМИД 8072524 . S2CID 33780610 .
- ^ Виктор, доктор медицинских наук; Апкарян, П. (2000). «Зрительная функция и организация мозга при синдроме непересекающихся ретинально-фугальных волокон». Цереб. Кортекс . 10 (1): 2–22. дои : 10.1093/cercor/10.1.2 . ПМИД 10639391 .
- ^ Бертен, Рене СП (1994). Естественный ум гипотетических животных: о гребцах и светящихся шарах (PDF) (кандидатская диссертация). Утрехт: Утрехтский университет.
- ^ Леб, Жак (1918). Принудительные движения, тропизмы и поведение животных . Филадельфия и Лондон: Липпинкотт. п. 209.
- ^ Шинброт, Т.; Янг, В. (2008). «Зачем перекреститься? Топологические ограничения на 3D-проводку». Анатомическая запись . 291 (10): 1278–1292. CiteSeerX 10.1.1.694.3088 . дои : 10.1002/ar.20731 . ПМИД 18780298 . S2CID 13477741 .
- ^ Брайтенберг, Валентино (1984). Транспортные средства-эксперименты по синтетической психологии . Кембридж, Массачусетс: MIT Press.
- ^ Жанвье, Филипп (1996). Ранние позвоночные . Оксфорд: Издательство Оксфордского университета. п. 409.
- ^ де Люссане, Марк Х.Э. (2011). «Гексамерное происхождение пяти лучей иглокожих [на arXiv.org: с исправлениями]». Эвол. Дев . 13 (2): 228–238. arXiv : 1107.2223v2 . дои : 10.1111/j.1525-142x.2011.00472.x . ПМИД 21410878 . S2CID 1938239 .
Дальнейшее чтение
[ редактировать ]- «Ваша голова может быть повернута набок» . Ютуб . 2020 . Проверено 9 февраля 2020 г.
- Крис Смит (11 августа 2023 г.). «Вопрос недели» . www.thenakedscientists.com (подкаст). Кембриджский университет. Событие происходит в 23:30 . Проверено 14 августа 2023 г.
- «Твои глаза наизнанку» . Ютуб . 29 сентября 2024 г. Проверено 1 августа 2024 г.
{{cite web}}: CS1 maint: дата и год ( ссылка )