CRISPR
| Каскад (комплекс, связанный с CRISPR для противовирусной защиты) | |||
|---|---|---|---|
 CRISPR Каскадный белок (голубой), связанный с РНК CRISPR (зеленый) и фаговой ДНК (красный) [ 1 ] | |||
| Идентификаторы | |||
| Организм | |||
| Символ | CRISPR | ||
| Входить | 947229 | ||
| PDB | 4 квартала | ||
| Refseq (prot) | NP_417241.1 | ||
| Uniprot | P38036 | ||
| |||
| Часть серии на |
| CRISPR |
|---|
| Редактирование генома : crispr-cas |
|
Варианты: Анти -Криспр - Крнис - КриспиCrispr-cas10 - crispr-cas13 - crispr-best Crispr -disp - crispr -gold - crispra - crispri Easi -crispr - сделать |
| Фермент |
|
CAS9 - Основные цели - ECORI - PSTI - SMAI Heaiii - cas12a (cpf1) - xcas9 |
| Приложения |
| Камера - лед - направленная генетика |
| Другой метод редактирования генома: |
| Редактирование Prime - Pro -AG - Rescue - Talen - Zfn - Leaper |
| Часть серии на |
| Генетическая инженерия |
|---|
| Генетически модифицированные организмы |
| История и регулирование |
| Процесс |
| Приложения |
| Противоречия |
CRISPR ( / ˈ k r ɪ s p ər / ) аббревиатура для сияющих эпохи в , , , ( . прокариотических археа бактерии семейства ДНК обнаруженных геномах организмов как таких и в последовательностей [ 2 ] Эти последовательности получены из фрагментов ДНК бактериофагов , которые ранее заразили прокариот. Они используются для обнаружения и уничтожения ДНК из аналогичных бактериофагов во время последующих инфекций. Следовательно, эти последовательности играют ключевую роль в противовирусной (т.е. антифажной ) системе прокариот и обеспечивают форму приобретенного иммунитета . [ 2 ] [ 3 ] [ 4 ] [ 5 ] CRISPR обнаруживается примерно в 50% секвенированных бактериальных геномов и почти 90% секвенированной археи. [ 6 ]

CAS9 (или «CRISPR-ассоциированный белок 9»)-это фермент , который использует последовательности CRISPR в качестве руководства для распознавания и открытия специфических цепей ДНК, которые дополняют последовательность CRISPR. Ферменты CAS9 вместе с последовательностями CRISPR составляют основу технологии, известной как CRISPR-CAS9 , которую можно использовать для редактирования генов в живых организмах. [ 8 ] [ 9 ] Этот процесс редактирования имеет широкий спектр применений, включая основные биологические исследования, разработку биотехнологических продуктов и лечение заболеваний. [ 10 ] [ 11 ] Развитие метода редактирования генома CRISPR-CAS9 была признана Нобелевской премией по химии в 2020 году, присужденной Эммануэль Чарпентье и Дженнифер Дудне . [ 12 ] [ 13 ]
История
[ редактировать ]Повторные последовательности
[ редактировать ]Открытие повторений кластерной ДНК происходило независимо в трех частях света. Первое описание того, что впоследствии будет названо CRISPR от Университета Осаки исследователя Йошизуми Ишино и его коллег в 1987 году. Они случайно клонировали часть последовательности CRISPR вместе с геномом «IAP» (изозим -преобразование щелочной фосфатазы) из генома эшераха (изозим -преобразование щелочной фосфатазы) из генома Escherichia. палочка [ 14 ] [ 15 ] которая была их целью. Организация повторов была необычной. Повторные последовательности обычно расположены последовательно, без впрыскивания разных последовательностей. [ 11 ] [ 15 ] Они не знали функции прерванных кластерных повторов.
В 1993 году исследователи Mycobacterium tuberculosis в Нидерландах опубликовали две статьи о кластере прерванных прямых повторов (DR) в этой бактерии. Они признали разнообразие последовательностей, которые вмешались в прямые повторения между различными штаммами M. tuberculosis [ 16 ] и использовал это свойство для разработки метода печати, называемого Spoligotying , все еще используемым сегодня. [ 17 ] [ 18 ]
Франциско Мохика из Университета Аликанте в Испании изучил функцию повторений в архаальных видах Haloferax и Haloarcula . Супервайзер Мохики предположил, что кластерные повторения играли роль в правильно сегрегированной реплицированной ДНК в дочерние клетки во время деления клеток, потому что плазмиды и хромосомы с идентичными повторными массивами не могли сосуществовать в вулкании Haloferax . Транскрипция прерванных повторов была также отмечена в первый раз; Это была первая полная характеристика CRISPR. [ 18 ] [ 19 ] К 2000 году Мохика и его ученики, после автоматического поиска опубликованных геномов, идентифицировали прерванные повторения у 20 видов микробов, принадлежащих к одному и тому же семейству. [ 20 ] Поскольку эти последовательности были перемещены, Mojica изначально назвал эти последовательности «короткие регулярно распределенные повторения» (SRSR). [ 21 ] В 2001 году Mojica и Ruud Jansen , которые искали дополнительные прерванные повторения, предложили аббревиатуру CRISPR (регулярно складывались в короткие палиндромические повтора), чтобы объединить многочисленные аббревиатуры, используемые для описания этих последовательностей. [ 19 ] [ 22 ] В 2002 году Tang, et al. показали доказательства того, что CRISPR повторения областей из генома археоглобуса фульгидуса были транскрибированы в длинные молекулы РНК, впоследствии обрабатываемые в малые РНК длиной длины, плюс некоторые более длинные формы из 2, 3 или более единиц повторного пропасти. [ 23 ] [ 24 ]
В 2005 году йогурта исследователь Родольф Баррангу обнаружил, что Streptococcus Thermophilus после итерационной инфекции фаговой инфекции развивает повышенную устойчивость к фагам из -за включения дополнительных последовательностей спейсеров CRISPR. [ 25 ] Работодатель Barrangou, датская продовольственная компания Danisco, затем разработала устойчивые к фагам штаммы S. thermophilus для производства йогурта. Позже Даниско был куплен DuPont , который владеет около 50 процентами мирового рынка молочной культуры, и технология широко распространяется. [ 26 ]
CRISPR-ассоциированные системы
[ редактировать ]Основной прогресс в понимании CRISPR пришел с наблюдением Янсена, что кластер для повторов прокариота сопровождался четырьмя гомологичными генами, которые составляют системы, связанные с CRISPR, CAS 1–4. Белки CAS показали геликазы и нуклеазы мотивы , что указывает на роль в динамической структуре локусов CRISPR. [ 27 ] В этой публикации аббревиатура CRISPR использовалась в качестве универсального названия этого шаблона, но его функция оставалась загадочной.

В 2005 году три независимые исследовательские группы показали, что некоторые проставки CRISPR получены из фаговой ДНК и экстрахромосомной ДНК, такой как плазмиды . [ 31 ] [ 32 ] [ 33 ] По сути, проставки представляют собой фрагменты ДНК, собранные из вирусов, которые ранее атаковали клетку. Источником проставки был признак того, что система Crisprs CAS может играть роль в адаптивном иммунитете у бактерий . [ 28 ] [ 34 ] Все три исследования, предлагающие эту идею, были первоначально отвергнуты громкими журналами, но в конечном итоге появились в других журналах. [ 35 ]
Первая публикация [ 32 ] Предлагая роль CRISPR-CAS в микробном иммунитете, MOJICA и соавторов в Университете Аликанте , предсказал роль РНК-транскрипта спейсеров по распознаванию цели в механизме, который может быть аналогичен системе интерференции РНК, используемой эукариотическими клетками. Кунин и его коллеги расширили эту гипотезу интерференции РНК, предлагая механизмы действия для различных подтипов CRISPR-Cas в соответствии с прогнозируемой функцией их белков. [ 36 ]
Экспериментальная работа нескольких групп выявила основные механизмы иммунитета CRISPR-CAS. В 2007 году было опубликовано первые экспериментальные данные о том, что CRISPR была адаптивной иммунной системой. [ 4 ] [ 11 ] Область CRISPR в Streptococcus Thermophilus приобрела проставки от ДНК инфицирующего бактериофага . Исследователи манипулировали сопротивлением S. thermophilus различным типам фагов, добавляя и удаляя проставки, последовательность которых соответствовала тем, которые были обнаружены в тестируемых фагах. [ 37 ] [ 38 ] В 2008 году Brouns и Van Der Oost идентифицировали комплекс белков CAS, называемых каскадом, который в E. coli вырезал предшественник РНК CRISPR в рамках повторений в зрелые спейсер-содержащие молекулы РНК, называемые РНК CRISPR (CRRNA), которые оставались связанными с белком сложный. [ 39 ] Более того, было обнаружено, что каскад, Crrna и геликаза/нуклеаза ( CAS3 ) должны были обеспечить бактериальный хозяин с иммунитетом против инфекции вирусом ДНК . Проектируя антивирусную CRISPR, они продемонстрировали, что две ориентации CRRNA (Sense/Antisende) обеспечивают иммунитет, что указывает на то, что гиды CRRNA нацелены на дцДНК . В этом году Марраффини и Сонтхеймер подтвердили, что последовательность CRISPR S. epidermidis нацеленной на ДНК, а не РНК для предотвращения конъюгации . Это открытие противоречило предложенному РНК-вводимому механизму иммунитета CRISPR-CAS, хотя система CRISPR-CAS, которая нацелена на иностранную РНК впоследствии обнаружена в Furiosus Pyrococcus . [ 11 ] [ 38 ] Исследование 2010 года показало, что CRISPR-CAS сокращает пряди как фаговой, так и плазмидной ДНК в S. thermophilus . [ 40 ]
Cas9
[ редактировать ]Более простая система CRISPR от Streptococcus pyogenes зависит от белка Cas9 . CAS9 Эндонуклеаза представляет собой четырехкомпонентную систему, которая включает в себя две малые молекулы: CRRNA и транса-активирующая РНК CRISPR (TracrRNA). [ 41 ] [ 42 ] В 2012 году Дженнифер Дудна и Эммануэль Чарпентье вновь информировали эндонуклеазу CAS9 в более управляемую двухкомпонентную систему, объединив две молекулы РНК в « РНК с одним парнем », которая в сочетании с CAS9 может найти и сократить мишень ДНК Указано руководством РНК. [ 43 ] Этот вклад был настолько значительным, что он был признан Нобелевской премией по химии в 2020 году. Манипулируя нуклеотидной последовательности руководящей РНК, система искусственной CAS9 может быть запрограммирована для нацеливания на любую последовательность ДНК для разделения. [ 43 ] Другое сотрудничество, состоящее из Вирджини Шикнис , Гасиунаса, Баррангу и Хорвата, показало, что Cas9 из системы S. thermophilus crispr также может быть перепрограммирована, чтобы нацелиться на участок их выбора, изменяя последовательность его Crrna. Эти достижения вызвали усилия по редактированию геномов с помощью модифицированной системы CRISPR-CAS9. [ 18 ]
Группы во главе с Фенгом Чжаном и Джорджем Черч одновременно опубликовали описания редактирования генома в культурах клеток человека, использующих CRISPR-Cas9 впервые. [ 11 ] [ 44 ] [ 45 ] С тех пор он использовался в широком спектре организмов, в том числе пекарских дрожжей ( Saccharomyces cerevisiae ), [ 46 ] [ 47 ] [ 48 ] оппортунистический патоген Candida albicans , [ 49 ] [ 50 ] зебра ( дорожки ), [ 51 ] Фруктовые мухи ( Drosophila melanogaster ), [ 52 ] [ 53 ] муравьи ( Harpegnathos saltator [ 54 ] и Ooceraea biroi [ 55 ] ), комары ( aedes aegypti [ 56 ] ), нематоды ( Caenorhabditis elegans ), [ 57 ] растения, [ 58 ] мыши ( Macaclima Muscle Omestic ) ; [ 59 ] [ 60 ] обезьяны [ 61 ] и человеческие эмбрионы. [ 62 ]
CRISPR был изменен для создания программируемых факторов транскрипции , которые позволяют активировать или молчать целевых генов. [ 63 ]

Было показано, что система CRISPR-CAS9 делает эффективные генные правовой редакции у человеческих трипноядерных зигот , как впервые описано в статье китайских ученых 2015 года П. Лян и Ю. Сюй. Система сделала успешное расщепление мутантного бета-гемоглобина ( HBB ) у 28 из 54 эмбрионов. Четыре из 28 эмбрионов были успешно перечислены с использованием шаблона доноров. Ученые показали, что во время рекомбинации ДНК расщепленной цепи гомологичная эндогенная последовательность HBD конкурирует с экзогенным шаблоном доноров. Репарация ДНК у эмбрионов человека гораздо сложнее и особенно, чем в производных стволовых клетках. [ 64 ]
CAS12A
[ редактировать ]В 2015 году Nuclease CAS12A (ранее назывался CPF1 [ 65 ] ) был охарактеризован в системе CRISPR-CPF1 бактерии Francisella Novicida . [ 66 ] [ 67 ] Его первоначальное название из Tigrfams определения семейства белков , построенного в 2012 году, отражает распространенность его подтипа CRISPR-Cas в линии Prevotella и Francisella . CAS12A показал несколько ключевых отличий от CAS9, в том числе: вызывание «шахматного» разреза в двойной междисфеонечной ДНК, а не «тупого», произведенного CAS9, полагаясь на «богатую» PAM (обеспечивая альтернативные сайты нацеливания CAS9) и требуя. Только РНК CRISPR (CRRNA) для успешного нацеливания. Напротив, CAS9 требует как Crrna, так и транса-активирующей CrRNA (TracrRNA).
Эти различия могут дать CAS12A некоторые преимущества перед CAS9. Например, небольшие CRRNAs CAS12A идеально подходят для мультиплексного редактирования генома, так как больше из них можно упаковать в один вектор, чем SGRNAs CAS9. Липкие 5 'свесы, оставленные CAS12A, также могут использоваться для сборки ДНК, которая гораздо более специфична для цели, чем клонирование традиционного рестрикционного фермента. [ 68 ] Наконец, CAS12A расщепляет ДНК 18–23 пар оснований вниз по течению от сайта PAM. Это означает, что нет нарушения последовательности распознавания после восстановления, и поэтому CAS12A позволяет многочисленным раундам расщепления ДНК. В отличие от этого, поскольку CAS9 сокращает только 3 пары оснований вверх по течению от сайта PAM, путь NHEJ приводит к индильским мутациям, которые разрушают последовательность распознавания, что предотвращает дальнейшие раунды резки. Теоретически, повторные раунды расщепления ДНК должны привести к увеличению возможностей для желаемого геномного редактирования. [ 69 ] Отличительной особенностью CAS12A по сравнению с CAS9 является то, что после сокращения своей цели CAS12A остается связанным с мишенью, а затем не дискриминально расщепляет другие молекулы ssDNA. [ 70 ] Это свойство называется «обеспечение расщепления» или «транс-расщепление» и использовалось для разработки различных диагностических технологий. [ 71 ] [ 72 ]
CAS13
[ редактировать ]В 2016 году нуклеаза Cas13a (ранее известный как C2C2 ) из бактерии C2C2 ) был охарактеризован . CAS13 представляет собой эндонуклеазу РНК с РНК, что означает, что он не расщепляет ДНК, а только одноцепочечную РНК. CAS13 руководствуется его CrRNA к мишени SSRNA и связывает и расщепляет мишень. Подобно CAS12A, CAS13 остается связанным с мишенью, а затем расщепляет другие молекулы ssRNA недискриминтером. [ 73 ] Это собственность по обеспечению расщепления было использовано для разработки различных диагностических технологий. [ 74 ] [ 75 ] [ 76 ]
В 2021 году доктор Хуэй Ян охарактеризовал новые миниатюрные варианты белка Cas13 (MCAS13), CAS13X и CAS13Y. Используя небольшую часть последовательности генов N из SARS-COV-2 в качестве мишени в характеристике MCAS13, выявила чувствительность и специфичность MCAS13 в сочетании с RT-лампом для обнаружения SARS-COV-2 как в синтетических, так и в клинических образцах над другими Доступные стандартные тесты, такие как RT-QPCR (1 копия/мкл). [ 77 ]
Локус Структура
[ редактировать ]Повторяется и проставки
[ редактировать ]Массив CRISPR состоит из последовательности AT-AT-лидеров, за которой следует короткие повторения, которые разделены уникальными проставками. [ 78 ] CRISPR повторяет, как правило, варьируется в размере от 28 до 37 пар оснований (BPS), хотя может быть всего 23 п.н. и до 55 п.н. [ 79 ] Некоторые показывают симметрию диады , подразумевая образование вторичной структуры , такой как петля стебля («шпилька») в РНК, в то время как другие предназначены для неструктурированных. Размер проставки в разных массивах CRISPR обычно составляет от 32 до 38 п.н. (диапазон от 21 до 72 п.н.). [ 79 ] Новые проставки могут быстро появляться в рамках иммунного ответа на фаговую инфекцию. [ 80 ] Обычно в массиве CRISPR в массиве CRISPR составляют менее 50 единиц последовательности повторного пространства. [ 79 ]
CRISPR РНК -структуры
[ редактировать ]-
 CRISPR-DR2: вторичная структура, взятая из базы данных RFAM . Семейство RF01315 .
CRISPR-DR2: вторичная структура, взятая из базы данных RFAM . Семейство RF01315 . -
 CRISPR-DR5: вторичная структура, взятая из базы данных RFAM . Семейство RF011318 .
CRISPR-DR5: вторичная структура, взятая из базы данных RFAM . Семейство RF011318 . -
 CRISPR-DR6: вторичная структура, взятая из базы данных RFAM . Семейство RF01319 .
CRISPR-DR6: вторичная структура, взятая из базы данных RFAM . Семейство RF01319 . -
 CRISPR-DR8: вторичная структура, взятая из базы данных RFAM . Семейство RF01321 .
CRISPR-DR8: вторичная структура, взятая из базы данных RFAM . Семейство RF01321 . -
 CRISPR-DR9: вторичная структура, взятая из базы данных RFAM . Семейство RF01322 .
CRISPR-DR9: вторичная структура, взятая из базы данных RFAM . Семейство RF01322 . -
 CRISPR-DR19: вторичная структура, взятая из базы данных RFAM . Семейство RF01332 .
CRISPR-DR19: вторичная структура, взятая из базы данных RFAM . Семейство RF01332 . -
 CRISPR-DR41: вторичная структура, взятая из базы данных RFAM . Семейство RF01350 .
CRISPR-DR41: вторичная структура, взятая из базы данных RFAM . Семейство RF01350 . -
 CRISPR-DR52: вторичная структура, взятая из базы данных RFAM . Семейство RF01365 .
CRISPR-DR52: вторичная структура, взятая из базы данных RFAM . Семейство RF01365 . -
 CRISPR-DR57: вторичная структура, взятая из базы данных RFAM . Семейство RF01370 .
CRISPR-DR57: вторичная структура, взятая из базы данных RFAM . Семейство RF01370 . -
 CRISPR-DR65: вторичная структура, взятая из базы данных RFAM . Семейство RF01378 .
CRISPR-DR65: вторичная структура, взятая из базы данных RFAM . Семейство RF01378 .










Гены CAS и подтипы CRISPR
[ редактировать ]Небольшие кластеры генов CAS часто расположены рядом с массивами CRISPR-повторного пространства. В совокупности 93 гена CAS сгруппированы в 35 семейств на основе сходства последовательностей кодируемых белков. 11 из 35 семейств образуют ядро CAS , которое включает в себя семейства белков CAS1 через CAS9. В полном локусе CRISPR-CAS есть по крайней мере один ген, принадлежащий ядру CAS . [ 81 ]
Системы CRISPR-CAS делятся на два класса. Системы класса 1 используют комплекс нескольких белков CAS для разложения иностранных нуклеиновых кислот. Системы класса 2 используют один большой белок CAS для той же цели. Класс 1 делится на типы I, III и IV; Класс 2 делится на типы II, V и VI. [ 82 ] 6 типов систем разделены на 33 подтипа. [ 83 ] Каждый тип и большинство подтипов характеризуются «подписью геном», найденным почти исключительно в категории. Классификация также основана на дополнении генов CAS , которые присутствуют. Большинство систем CRISPR-CAS имеют белок CAS1. Филогения , белков CAS1 обычно согласуется с системой классификации [ 84 ] Но исключения существуют из -за перетасовки модуля. [ 81 ] Многие организмы содержат несколько систем CRISPR-CAS, предполагающие, что они совместимы и могут совместно использовать компоненты. [ 85 ] [ 86 ] Спорадическое распределение подтипов CRISPR-CAS предполагает, что система CRISPR-CAS подвержена горизонтальному переносу генов микробов во время эволюции .
В этой таблице отсутствует информация о Uniprot и InterPro перекрестной ссылки. ( Октябрь 2020 г. ) |
| Сорт | Тип CAS | CAS подтип | Фирменный белок | Функция | Ссылка |
|---|---|---|---|---|---|
| 1 | я | — | CAS3 | Одноцепочечная дНК-нуклеаза (HD-домен) и АТФ-зависимая геликаза | [ 87 ] [ 88 ] |
| Я | Cas8a, Cas5 | CAS8 является субъединицей интерференционного модуля, которая важна для нацеливания на вторжение ДНК путем распознавания последовательности PAM . CAS5 необходим для обработки и стабильности CRRNAS. | [ 84 ] [ 89 ] | ||
| IB | Cas8b | ||||
| IC | Cas8c | ||||
| ИДЕНТИФИКАТОР | Cas10d | Содержит домен, гомологичный домен пальмы полимераз нуклеиновой кислоты и нуклеотидных циклаз | [ 90 ] [ 91 ] | ||
| Т.е. | CSE1, CSE2 | ||||
| ЕСЛИ | CSY1, CSY2, CSY3 | Тип IF-3 участвовал в транспозонах, ассоциированных с CRISPR | [ 84 ] | ||
| IG [ Примечание 1 ] | GSU0054 | [ 92 ] | |||
| Iii | — | CAS10 | Гомолог CAS10D и CSE1. Связывает РНК -мишени CRISPR и способствует стабильности интерференционного комплекса | [ 91 ] [ 93 ] | |
| III-A | CSM2 | Не определяется | [ 84 ] | ||
| III-B | CMR5 | Не определяется | [ 84 ] | ||
| III-C | CAS10 или CSX11 | [ 84 ] [ 93 ] | |||
| III-D | CSX10 | [ 84 ] | |||
| Iii-e | [ 92 ] | ||||
| Iii-f | [ 92 ] | ||||
| IV | — | CSF1 | [ 92 ] | ||
| Iv-a | [ 92 ] | ||||
| IV-B | [ 92 ] | ||||
| IV-C. | [ 92 ] | ||||
| 2 | II | — | Cas9 | Нуклеазы RUVC и HNH вместе производят DSB , и отдельно могут производить разрывы в одну цепь. Обеспечивает получение функциональных проставков во время адаптации. | [ 94 ] [ 95 ] |
| II-A | CSN2 | Кольцевой ДНК-связывающий белок. Участвует в заполненной адаптации в системе CRISPR типа II. | [ 96 ] | ||
| II-B | Cas4 | Эндонуклеаза, которая работает с CAS1 и CAS2 для генерации последовательностей спейсеров | [ 97 ] | ||
| II-C. | Характеризуется отсутствием CSN2 или CAS4 | [ 98 ] | |||
| V | — | CAS12 | Nuclease ruvc. Не хватает HNH. | [ 82 ] [ 99 ] | |
| V-A | CAS12A (CPF1) | Автопроцессование пре-межселе | [ 92 ] [ 100 ] | ||
| V-B | CAS12B (C2C1) | [ 92 ] | |||
| V-C | CAS12C (C2C3) | [ 92 ] | |||
| V-D | CAS12D (CASY) | [ 92 ] | |||
| Вежливый | CAS12E (CASX) | [ 92 ] | |||
| Vf. | CAS12F (CAS14, C2C10) | [ 92 ] | |||
| V-G | CAS12G | [ 92 ] | |||
| VH | CAS12H | [ 92 ] | |||
| V-I | Cas12i | [ 92 ] | |||
| V-K [ Примечание 2 ] | CAS12K (C2C5) | Тип VK участвовал в транспозонах, связанных с CRISPR . | [ 92 ] | ||
| V-U | C2C4, C2C8, C2C9 | [ 92 ] | |||
| МЫ | — | CAS13 | РНК-РНКаза | [ 82 ] [ 101 ] | |
| Ты | CAS13A (C2C2) | [ 92 ] | |||
| Vi-b | Cas13b | [ 92 ] | |||
| ШУТИТЬ | Cas13c | [ 92 ] | |||
| Обвенчались | Cas13d | [ 92 ] | |||
| Vi-x | CAS13X.1 | РНК-зависимая РНК-полимераза, ингибирование профилактического РНК-вируса | [ 102 ] | ||
| Vi-y | [ 102 ] |
Механизм
[ редактировать ]
(1) Приобретение начинается с распознавания вторжения ДНК CAS1 и CAS2 и расщепления протоспакера.
(2) Протопсер лигируется с прямым повторением примыкания к последовательности лидеров и
(3) Продление с одной прямой ремонтирует CRISPR и дублирует прямой повтор. Стадии обработки и интерференции CRRNA происходят по -разному в каждой из трех основных систем CRISPR.
(4) Первичный транскрипт CRISPR расщепляется генами CAS для получения CRRNAS.
(5) В системах I типа CAS6E/CAS6F расщепляются на соединении ssRNA и дсРНК, образованной петлями шпильки в прямом повторении. Системы типа II используют транса-активирующую (TRACR) РНК с образованием дцРНК, которая расщепляется CAS9 и RNASEIII. Системы типа III используют гомолог CAS6, который не требует петли шпильки в прямом повторении для расщепления.
(6) В системах типа II и типа III.
(7) Зрелые CRRNAs связываются с белками CAS с образованием интерференционных комплексов.
(8) В системах типа I и типа II взаимодействие между белком и последовательности PAM требуется для ухудшения вторжения ДНК. Системы типа III не требуют PAM для успешной деградации, а в системах систем III типа A происходит между CRRNA и мРНК, а не ДНК, нацеленным на системы типа III-B.



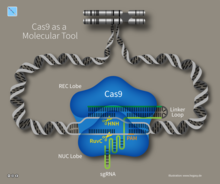

Иммунитет CRISPR-CAS является естественным процессом бактерий и архей. [ 103 ] CRISPR-CAS предотвращает инфекцию бактериофагов, конъюгацию и природную трансформацию путем разложения иностранных нуклеиновых кислот, которые попадают в клетку. [ 38 ]
Приобретение проставки
[ редактировать ]Когда микроб захватывается бактериофагом , первой стадией иммунного ответа является захват фаговой ДНК и вставить ее в локус CRISPR в форме проставки. CAS1 и CAS2 обнаруживаются в обоих типах иммунной системы CRISPR-CAS, что указывает на то, что они участвуют в приобретении проставки. Исследования по мутациям подтвердили эту гипотезу, показав, что удаление CAS1 или CAS2 прекратило приобретение проставки, не влияя на иммунный ответ CRISPR. [ 104 ] [ 105 ] [ 106 ] [ 107 ] [ 108 ]
Было охарактеризовано множество белков CAS1, и их структуры разрешены. [ 109 ] [ 110 ] [ 111 ] Белки Cas1 имеют разнообразные аминокислотные последовательности. Тем не менее, их кристаллические структуры похожи, и все очищенные белки Cas1 являются металлическими нуклеазами/ интегратами , которые связываются с ДНК независимым от последовательности. [ 85 ] Репрезентативные белки Cas2 были охарактеризованы и обладают либо (одиночной нити) ssRNA- [ 112 ] или (двойная прядь) дсдна- [ 113 ] [ 114 ] Конкретная эндорибонуклеазная активность.
В системе IE E. coli CAS1 и CAS2 образуют комплекс, где димер CAS2 соединяет два димера CAS1. [ 115 ] В этом комплексном CAS2 выполняет негриментативную роль каркаса, [ 115 ] Связывание двухцепочечных фрагментов вторжения ДНК, в то время как CAS1 связывает одноцепочечные боки ДНК и катализирует их интеграцию в массивы CRISPR. [ 116 ] [ 117 ] [ 118 ] Новые проставки обычно добавляются в начале CRISPR рядом с последовательности лидеров, создавая хронологическую запись вирусных инфекций. [ 119 ] В E. coli гистон, подобный белке, называемый фактором интеграционного хозяина ( IHF ), который связывается с последовательности лидеров, отвечает за точность этой интеграции. [ 120 ] IHF также повышает эффективность интеграции в типе, если система Pectobacterium atrosepticum . [ 121 ] Но в других системах могут потребоваться различные факторы хоста [ 122 ]
Протоспацер соседних мотивов (PAM)
[ редактировать ]Биоинформатический анализ областей фаговых геномов, которые были вырезаны в виде спейсеров (называемых протоспацеров), показал, что они не были выбраны случайным образом, а вместо этого были обнаружены рядом с короткими (3–5 п.н.) последовательностями ДНК, называемых прилегающими мотивами протоспацера (PAM). Анализ систем CRISPR-CAS показал, что PAMs важны для систем типа I и типа II, но не систем типа III во время приобретения. [ 33 ] [ 123 ] [ 124 ] [ 125 ] [ 126 ] [ 127 ] В системах типа I и типа II, протоспасеры иссекаются в положениях, прилегающих к последовательности PAM, с другим концом проставки с использованием механизма линейки, тем самым поддерживая регулярность размера спейсера в массиве CRISPR. [ 128 ] [ 129 ] Сохранение последовательности PAM отличается между системами CRISPR-CAS и, по-видимому, эволюционно связано с CAS1 и последовательности лидеров . [ 127 ] [ 130 ]
Новые проставки добавляются в массив CRISPR в направлении, [ 31 ] встречается преимущественно, [ 80 ] [ 123 ] [ 124 ] [ 131 ] [ 132 ] но не исключительно, прилегающий [ 126 ] [ 129 ] к последовательности лидера. Анализ системы IE типа от E. coli продемонстрировал, что первое прямое повторение, прилегающее к последовательности лидеров, скопирована, причем недавно приобретенный проставщик вставлен между первым и вторым прямым повторением. [ 107 ] [ 128 ]
Последовательность PAM, по -видимому, важна во время вставки проставки в системах IE типа. Эта последовательность содержит сильно консервативную конечную нуклеотид (NT), прилегающая к первым NT протопэссера. Этот NT становится последней базой в первом прямом повторении. [ 108 ] [ 133 ] [ 134 ] Это говорит о том, что механизм сбора проставки генерирует одноцепочечные свесы в втором положении прямого повторения и в PAM во время вставки проставки. Однако не все системы CRISPR-CAS, по-видимому, разделяют этот механизм, поскольку PAMS в других организмах не показывают такой же уровень сохранения в конечной позиции. [ 130 ] Вполне вероятно, что в этих системах тупой конец генерируется в самом конце прямого повторения и протоспакера во время приобретения.
Варианты вставки
[ редактировать ]Анализ Sulfolobus solfataricus crisprs выявил дополнительные сложности для канонической модели вставки спейсеров, поскольку один из шести локусов CRISPR вставлял новые проставки случайным образом на протяжении всего своего массива CRISPR, в отличие от вставки наиболее близких к последовательности лидеров. [ 129 ]
Многочисленные CRISPR содержат много проставки в одном и том же фаге. Механизм, который вызывает это явление, был обнаружен в системе типа IE E. coli . Было обнаружено значительное улучшение приобретения проставки, когда проставки уже нацелены на фаг, даже несоответствия протоспакеру. Это «праймирование» требует, чтобы белки CAS участвовали как в приобретении, так и в интерференции для взаимодействия друг с другом. Недавно приобретенные проставки, возникающие в результате механизма заполнения, всегда находятся на одной цепи, что и проставка для заполнения. [ 108 ] [ 133 ] [ 134 ] Это наблюдение привело к гипотезе о том, что механизм приобретения скользит вдоль иностранной ДНК после праймирования, чтобы найти новый протосп. [ 134 ]
Биогенез
[ редактировать ]CRISPR-RNA (CRRNA), которая впоследствии направляет нуклеазу CAS к мишени на стадии интерференции, должна генерироваться из последовательности CRISPR. КРРНК первоначально транскрибируется как часть одной длинной транскрипции, охватывающей большую часть массива CRISPR. [ 29 ] Этот транскрипт затем расщепляется белками CAS с образованием CRRNAS. Механизм производства CRRNAS отличается среди систем CRISPR-CAS. В типе IE и типа, если системы, белки CAS6E и CAS6F соответственно, распознают петли стеблей [ 135 ] [ 136 ] [ 137 ] Создан путем сочетания идентичных повторов, которые фланкируют тррен. [ 138 ] Эти белки CAS расщепляют более длинную транскрипцию на краю парной области, оставляя одну сдержанную кровку вместе с небольшим остатком парной области повторения.
Системы типа III также используют CAS6, однако их повтора не производят петли стеблей. Вместо этого расщепление происходит более длинным транскриптом вокруг CAS6, чтобы разрешить расщепление чуть выше последовательности повторения. [ 139 ] [ 140 ] [ 141 ]
Системам типа II не хватает гена CAS6 и вместо этого используют RNASEIII для расщепления. Функциональные системы типа II кодируют дополнительную небольшую РНК, которая комплементарна для последовательности повторений, известной как транса-активирующая крольна (тракрРНК). [ 41 ] Транскрипция тракрРНК и первичной транскрипции CRISPR приводит к спариванию оснований и образованию дцРНК в последовательности повторений, которая впоследствии нацелена на РНКасеиии для получения CRRNAS. В отличие от двух других систем, CRRNA не содержит полного проставки, который вместо этого усечен на одном конце. [ 94 ]
CRRNAs связывают с белками CAS с образованием рибонуклеотидных комплексов, которые распознают иностранные нуклеиновые кислоты. CRRNAs не показывают предпочтения между кодирующими и некодирующими нити, что указывает на систему ДНК-нацеливания РНК. [ 5 ] [ 40 ] [ 104 ] [ 108 ] [ 142 ] [ 143 ] [ 144 ] Комплекс типа IE (обычно называемый каскадом) требует пяти белков CAS, связанных с одной кронной. [ 145 ] [ 146 ]
Вмешательство
[ редактировать ]На этапе помех в системах I типа последовательность PAM распознается на Crrna-In-Land и требуется вместе с отжигом Crrna. В системах I типа Правильное сочетание оснований между Crrna и Protospacer сигнализирует о конформационном изменении в каскаде, которое рекрутирует CAS3 для деградации ДНК.
Системы типа II полагаются на один многофункциональный белок, CAS9 , для этапа помех. [ 94 ] CAS9 требует, чтобы как тррен, так и тракрРНК для функционирования и расщепления ДНК с использованием его двойных HNH и RUVC/RNASEH-подобных доменов эндонуклеазы. В системах типа II требуется базовое выступление между PAM и геномом фага. Тем не менее, PAM распознается на той же цепи, что и с Crrna (противоположная цепочка для систем типа I).
Системы типа III, такие как тип I, требуют шести или семи белков CAS, связывания с CRRNAs. [ 147 ] [ 148 ] Системы типа III, проанализированные из S. solfataricus и P. furiosus, оба нацелены на мРНК фагов, а не генома фага, ДНК, [ 86 ] [ 148 ] что может сделать эти системы однозначно способными нацелить фаг-геномы на основе РНК. [ 85 ] Также было обнаружено, что системы типа III нацелены на ДНК в дополнение к РНК с использованием другого белка CAS в комплексе, CAS10. [ 149 ] Было показано, что расщепление ДНК зависит от транскрипции. [ 150 ]
Механизм для отличия от иностранной ДНК во время помех встроен в CRRNAS и, следовательно, вероятно, является общим для всех трех систем. На протяжении всего отличительного процесса созревания каждого основного типа все CRRNAS содержат проставленную последовательность и некоторую часть повторения на одном или обоих концах. Это частичная последовательность повторений, которая предотвращает нацеливание системы CRISPR-CAS на нацеливание на хромосому в качестве спаривания оснований за пределами сигналов последовательности спейсеров и предотвращает расщепление ДНК. [ 151 ] РНК-управляемые ферменты CRISPR классифицируются как рестрикционные ферменты типа V.
Эволюция
[ редактировать ]| CRISPR, связанный белком CAS2 (адаптация РНКаза) | |||
|---|---|---|---|
 Кристаллическая структура гипотетического белка TT1823 из Thermus Thermophilus | |||
| Идентификаторы | |||
| Символ | Crispr_cas2 | ||
| Pfam | PF09827 | ||
| InterPro | IPR019199 | ||
| CDD | CD09638 | ||
| |||
| CRISPR-ассоциированный белок CASA/CSE1 (эффекторная ДНКаза I типа) | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | Crispr_cse1 | ||
| Pfam | PF09481 | ||
| InterPro | IPR013381 | ||
| CDD | CD09729 | ||
| |||
| CRISPR, связанный с белком CASC/CSE3/CAS6 (эффекторная РНКаза типа) | |||
|---|---|---|---|
 Кристаллическая структура белка, ассоциированного с CRISPR из Thermus Thermophilus | |||
| Идентификаторы | |||
| Символ | Crispr_assoc | ||
| Pfam | PF08798 | ||
| PFAM клан | CL0362 | ||
| InterPro | IPR010179 | ||
| CDD | CD09727 | ||
| |||
Считается, что гены CAS в адаптере и эффекторных модулях системы CRISPR-CAS развивались из двух разных предков. Транспозонподобный элемент , называемый каспозон , кодирующий CAS1 -подобную интегразу и потенциально другие компоненты модуля адаптации, был вставлен рядом с модулем наследственного эффектора, который, вероятно, функционировал как независимая врожденная иммунная система. [ 152 ] Высококонсервативные гены CAS1 и CAS2 модуля адаптера развивались из наследственного модуля, в то время как различные эффекторные гены CAS класса 1 развивались из модуля наследственного эффектора. [ 153 ] Эволюция этих различных генов эффекторных модулей класса 1 руководствовалась различными механизмами, такими как события дублирования. [ 154 ] С другой стороны, каждый тип эффекторного модуля класса 2 возник из -за последующих независимых вставков мобильных генетических элементов. [ 155 ] Эти мобильные генетические элементы заняли место множественных генных эффекторных модулей для создания эффекторных модулей одного гена, которые производят большие белки, которые выполняют все необходимые задачи эффекторного модуля. [ 155 ] Распространные области систем CRISPR-CAS взяты непосредственно из иностранных мобильных генетических элементов, и, следовательно, их долгосрочную эволюцию трудно проследить. [ 156 ] Было обнаружено, что нелупинка этих областей проставки сильно зависит от окружающей среды и конкретных иностранных мобильных генетических элементов, которые он содержит. [ 157 ]
CRISPR-CAS может иммунизировать бактерии против определенных фагов и, таким образом, остановить передачу. По этой причине Кунин описал CRISPR-CAS как механизм наследования Ламаркия . [ 158 ] Тем не менее, это было оспорено критиком, который отметил: «Мы должны помнить [Ламарк] за то, что он внес в науку, а не о вещах, которые напоминают его теорию лишь поверхностно. Действительно, думая о CRISPR и других явлениях, поскольку Ламаркиян только скрывает простое простые. И элегантный способ эволюции действительно работает ». [ 159 ] Но по мере того, как были проведены более поздние исследования, стало очевидно, что приобретенные пространственные области систем CRISPR-CAS действительно являются формой эволюции Ламаркина, поскольку это генетические мутации, которые приобретаются, а затем передаются. [ 160 ] С другой стороны, эволюция механизма гена CAS, которая облегчает систему, развивается благодаря классической дарвинианской эволюции. [ 160 ]
Коэволюция
[ редактировать ]Анализ последовательностей CRISPR выявил коэволюцию хозяина и вирусных геномов. [ 161 ]
Основная модель эволюции CRISPR - это недавно включенные спейсеры, приводящие фаги, чтобы мутировать свои геномы, чтобы избежать бактериального иммунного ответа, создавая разнообразие как в популяциях фага, так и в популяциях фага. Чтобы противостоять фаговой инфекции, последовательность проставки CRISPR должна идеально соответствовать последовательности гена -целевого фага. Фаги могут продолжать заражать заданную точечную мутации своих хозяев в проставке. [ 151 ] Аналогичная строгость требуется в PAM или бактериальной штамме, чувствительной к фагам. [ 124 ] [ 151 ]
Ставки
[ редактировать ]Исследование 124 штаммов S. Thermophilus показало, что 26% всех проставки были уникальными и что разные локусы CRISPR показали различные показатели приобретения проставки. [ 123 ] Некоторые локусы CRISPR развиваются быстрее, чем другие, что позволило определить филогенетические отношения штаммов. Сравнительный геномный анализ показал, что E. coli и S. enterica развиваются гораздо медленнее, чем S. thermophilus . Штаммы последних, которые разошлись 250 000 лет назад, все еще содержали тот же дополнительный комплемент. [ 162 ]
Метагеномный анализ двух биопленок с кислой-мин-дренажом показал, что один из проанализированных CRISPR содержал обширные делеции и добавления спейсеров по сравнению с другой биопленкой, что предполагает более высокую активность/распространенность фага в одном сообществе, чем в другом. [ 80 ] В полости рта временное исследование определило, что 7–22% проставки были распределены в течение 17 месяцев внутри человека, в то время как менее 2% были разделены на отдельных лиц. [ 132 ]
Из той же среды был отслеживал один штамм с использованием праймеров ПЦР , специфичных для его системы CRISPR. Результаты широкого уровня присутствия/отсутствия проставки показали значительное разнообразие. Однако этот CRISPR добавил три проставки за 17 месяцев, [ 132 ] предполагая, что даже в среде со значительным разнообразием CRISPR некоторые локусы развиваются медленно.
CRISPR были проанализированы из метагеномов, продуманных для проекта микробиома человека . [ 163 ] Хотя большинство из них были специфичными для тела, некоторые внутри тела широко распространены среди людей. Один из этих локусов возникла из стрептококковых видов и содержал ≈15000 проставки, 50% из которых были уникальными. Подобно целевым исследованиям полости рта, некоторые показали небольшую эволюцию с течением времени. [ 163 ]
Эволюция CRISPR была изучена в хемостатах с использованием S. Thermophilus для непосредственного изучения скорости сбора проставки. За одну неделю штаммы S. thermophilus приобрели до трех проставки, когда бросали вызов одним фагом. [ 164 ] В том же интервале у фага развивались однонуклеотидные полиморфизмы , которые стали фиксированными в популяции, что позволяет предположить, что нацеливание предотвратило репликацию фага при отсутствии этих мутаций. [ 164 ]
Другой эксперимент S. thermophilus показал, что фаги могут заразить и повторять у хозяев, у которых есть только один нацеленный проставщик. Еще один показал, что чувствительные хозяева могут существовать в средах с высокими фазами титров. [ 165 ] Исследования хемостата и обсерваций предполагают много нюансов для эволюции CRISPR и фага (CO).
Идентификация
[ редактировать ]CRISPR широко распространены среди бактерий и археи [ 90 ] и показывать некоторые сходства последовательностей. [ 138 ] Их наиболее заметной характеристикой являются их повторяющиеся проставки и прямые повторения. Эта характеристика позволяет CRISPR легко идентифицировать в длинных последовательностях ДНК, поскольку количество повторений уменьшает вероятность ложного положительного соответствия. [ 166 ]
Анализ CRISPR в метагеномных данных является более сложным, поскольку локусы CRISPR обычно не собираются из -за их повторяющегося характера или с помощью изменений деформации, что смущает алгоритмы сборки. В тех случаях, когда доступно много эталонных геномов, полимеразная цепная реакция (ПЦР) может использоваться для амплификации массивов CRISPR и анализа содержания спейсеров. [ 123 ] [ 132 ] [ 167 ] [ 168 ] [ 169 ] [ 170 ] Тем не менее, этот подход дает информацию только для специально целенаправленных CRIPPR и для организмов с достаточным представлением в общедоступных базах данных для разработки надежных праймеров ПЦР полимеразы. Вырожденные повтора-специфические праймеры могут быть использованы для усиления проставки CRISPR непосредственно из образцов окружающей среды; Ампликоны, содержащие два или три проставки, могут затем быть собраны вычислительно реконструировать длинные массивы CRISPR. [ 170 ]
Альтернативой является извлечение и восстановление массивов CRISPR из метагеномных данных о дробовике. Это вычислительно сложнее, особенно с технологиями секвенирования второго поколения (например, 454, Illumina), так как короткие длины считывания предотвращают более двух или трех повторяющихся единиц, появляющихся в одном чтении. Идентификация CRISPR в необработанных показаниях была достигнута с использованием чисто de novo идентификации [ 171 ] или с использованием последовательностей прямых повторений в частично собранных массивах CRISPR из контигов (перекрывая сегменты ДНК, которые вместе представляют собой консенсусную область ДНК) [ 163 ] и прямые повторные последовательности из опубликованных геномов [ 172 ] как крючок для определения прямых повторений в отдельных чтениях.
Используйте фаги
[ редактировать ]Еще один способ защиты от бактерий от инфекции фагов - это наличие хромосомных островов . Подтип хромосомных островов, называемый фагом-индуцируемым хромосомным островом (PICI), вырезается из бактериальной хромосомы при инфекции фагов и может ингибировать репликацию фага. [ 173 ] PICE индуцируются, вырезаются, реплицируются и, наконец, упаковываются в небольшие капсиды определенными стафилококковыми умеренными фагами. PICIS использует несколько механизмов для блокировки репродукции фага. В первом механизме PIC-кодируемый PPI дифференциально блокирует созревание фага, связывая или взаимодействуя, специфически с фагами, следовательно, блокируя образование комплекса фагов/Terl, ответственного за упаковку Фаговой ДНК. Во втором механизме PICI CPMAB перенаправляет морфогенетический белок фагового капсида, чтобы сделать 95% капсида размером с SAPI, а ДНК фага может упаковать только 1/3 своего генома в этих небольших капсидах и, следовательно, стать нежизнеспособным фагом. [ 174 ] Третий механизм включает в себя два белка, PTIA и PTIB, которые нацелены на LTRC, который отвечает за производство вириона и лизисных белков. Этот механизм интерференции модулируется модуляторным белком, PTIM, связывается с одним из интерференционных белков, PTIA и, следовательно, достигает необходимого уровня помех. [ 175 ]
Одно исследование показало, что фаг Lytic ICP1, который специально нацелен на Vibrio Cholerae Serogroup O1, приобрел систему CRISPR-Cas, которая нацелена на V. v. jolera pici-подобный элемент. Система имеет 2 локуса CRISPR и 9 генов CAS. Кажется, это гомологично для системы, найденной в Yersinia pestis . Более того, как и бактериальная система CRISPR-CAS, ICP1 CRISPR-CAS может приобрести новые последовательности, которые позволяют PHAGE и HOST-EVELE. [ 176 ] [ 177 ]
Было показано, что определенные архейные вирусы несут массивы мини-кризра, содержащие один или два проставки. Было показано, что расставки в массивах CRISPR, передаваемых вирусом, нацелены на другие вирусы и плазмиды, что позволяет предположить, что мини-массивы CRISPR представляют собой механизм гетеротипического исключения суперинфекции и участвуют в интервенционных конфликтах. [ 170 ]
Приложения
[ редактировать ]Редактирование генов CRISPR - это революционная технология, которая обеспечивает точные, целевые модификации ДНК живых организмов. Разработанный из естественной защиты, обнаруженного у бактерий, CRISPR-CAS9 является наиболее часто используемой системой, которая позволяет «резать» ДНК в определенных местах и удалять, модифицировать или вставлять генетический материал. Эта технология трансформировала такие области, как генетика, медицина, [ 178 ] [ 179 ] и сельское хозяйство, [ 180 ] [ 181 ] Предлагая потенциальные методы лечения генетических расстройств, достижения в области инженерии сельскохозяйственных культур и исследования фундаментальной работы жизни. Однако его этические последствия и потенциальные непреднамеренные последствия вызвали значительные дебаты. [ 182 ] [ 183 ]
Смотрите также
[ редактировать ]- Активация CRISPR
- Антикриспр
- CRISPR/CAS Инструменты
- Редактирование генов CRISPR
- Журнал CRISPR
- " Дизайнерский ребенок "
- Драко
- Ген нокаут
- Обще геном CRISPR-CAS9 нокаут экраны
- Глоссарий генетики
- Человеческая зародышевая инженерия
- Человеческая природа (документальный фильм 2019 года)
- Магический
- Новая евгеника
- Главное редактирование
- РНКИ
- МиРНК
- Анализ нуклеазы геодезиста
- Синтетическая биология
- Цинковый палец
Примечания
[ редактировать ]Ссылки
[ редактировать ]- ^ ВВП : 4QYZ : Mulepati S, Héroux A, Bailey S (2014). «Кристаллическая структура комплекса надзора с помощью CRISPR, связанного с надзором, связана с мишенью ssDNA» . Наука . 345 (6203): 1479–1484. Bibcode : 2014sci ... 345.1479M . doi : 10.1126/science.1256996 . PMC 4427192 . PMID 25123481 .
- ^ Jump up to: а беременный Баррангу Р. (2015). «Роли систем CRISPR-CAS в адаптивном иммунитете и за его пределами». Текущее мнение в иммунологии . 32 : 36–41. doi : 10.1016/j.coi.2014.12.008 . PMID 25574773 .
- ^ Редман М., Кинг А., Уотсон С., Кинг Д. (август 2016 г.). "Что такое crispr/cas9?" Полем Архив болезни в детстве: издание образования и практики . 101 (4): 213–215. doi : 10.1136/archdischild-2016-310459 . PMC 4975809 . PMID 27059283 .
- ^ Jump up to: а беременный Баррунгу Р. , Фримо С., Дево Х., Ричардс М., Бойавал П., Моино С. и др. (Март 2007 г.). «CRISPR обеспечивает приобретенную устойчивость к вирусам у прокариот». Наука . 315 (5819): 1709–1712. Bibcode : 2007sci ... 315.1709b . doi : 10.1126/science.1138140 . HDL : 20.500.11794/38902 . PMID 17379808 . S2CID 3888761 . ( Требуется регистрация )
- ^ Jump up to: а беременный Marraffini LA, Sontheimer EJ (декабрь 2008 г.). «Интерференция CRISPR ограничивает горизонтальный перенос генов у стафилококков путем нацеливания на ДНК» . Наука . 322 (5909): 1843–1845. Bibcode : 2008Sci ... 322.1843M . doi : 10.1126/science.1165771 . PMC 2695655 . PMID 19095942 .
- ^ Hille F, Richter H, Wong SP, Bratovič M, Ressel S, Charpentier E (март 2018 г.). «Биология CRISPR-CAS: назад и вперед». Клетка . 172 (6): 1239–1259. doi : 10.1016/j.cell.2017.11.032 . HDL : 21.11116/0000-0003-FC0D-4 . PMID 29522745 . S2CID 3777503 .
- ^ Horvath P, Barrangou R (январь 2010 г.). «CRISPR/CAS, иммунная система бактерий и археи». Наука . 327 (5962): 167–170. Bibcode : 2010sci ... 327..167H . doi : 10.1126/science.1179555 . PMID 20056882 . S2CID 17960960 .
- ^ Bak Ro, Gomez-Ospina N, Porteus MH (август 2018 г.). «Редактирование генов на центральной сцене». Тенденции в генетике . 34 (8): 600–611. doi : 10.1016/j.tig.2018.05.004 . PMID 29908711 . S2CID 49269023 .
- ^ Zhang F, Wen Y, Guo X (2014). «CRISPR/CAS9 для редактирования генома: прогресс, последствия и проблемы» . Молекулярная генетика человека . 23 (R1): R40–6. doi : 10.1093/hmg/ddu125 . PMID 24651067 . S2CID 19008095 .
- ^ Crispr-cas9, Talens and Zfns-битва в редактировании генов https://www.ptglab.com/news/blog/crispr-cas9-talens-and-zfns-the-battle-in-gene-editing/ архивировано 2021 -05-25 на машине Wayback
- ^ Jump up to: а беременный в дюймовый и HSU PD, Lander ES, Zhang F (июнь 2014 г.). «Разработка и применение CRISPR-CAS9 для инженерии генома» . Клетка . 157 (6): 1262–1278. doi : 10.1016/j.cell.2014.05.010 . PMC 4343198 . PMID 24906146 .
- ^ «Пресс -релиз: Нобелевская премия по химии 2020 года» . Нобелевский фонд. Архивировано из оригинала 15 января 2021 года . Получено 7 октября 2020 года .
- ^ Wu KJ, Peltier E (7 октября 2020 г.). «Нобелевская премия по химии, присужденная 2 ученым за работу по редактированию генома - Эммануэль Чарпентье и Дженнифер А. Дудна разработали инструмент CRISPR, который может изменить ДНК животных, растений и микроорганизмов с высокой точностью» . New York Times . Архивировано из оригинала 8 октября 2020 года . Получено 7 октября 2020 года .
- ^ Рават А., Рой М., Джиоти А., Каушик С., Верма К., Шривастава В.К. (август 2021 г.). «Цистеиновые протеазы: борьба с патогенными паразитными простейшими с вездесущими ферментами» . Микробиологические исследования . 249 : 126784. DOI : 10.1016/j.micres.2021.126784 . PMID 33989978 . S2CID 234597200 .
- ^ Jump up to: а беременный Ишино Й., Шинагава Х., Макино К., Амемура М., Наката А (декабрь 1987). «Нуклеотидная последовательность гена IAP, ответственная за преобразование изозима щелочной фосфатазы в Escherichia coli, и идентификация генного продукта» . Журнал бактериологии . 169 (12): 5429–5433. doi : 10.1128/jb.169.12.5429-5433.1987 . PMC 213968 . PMID 3316184 .
- ^ Van Soolingen D, De Haas PE, Hermans PW, Groenen PM, Van Embden JD (август 1993). «Сравнение различных повторяющихся элементов ДНК в качестве генетических маркеров для дифференцировки штаммов и эпидемиологии микобактерии туберкулеза» . Журнал клинической микробиологии . 31 (8): 1987–1995. doi : 10.1128/jcm.31.8.1987-1995.1993 . PMC 265684 . PMID 7690367 .
- ^ Groenen PM, Bunschoten AE, Van Soolingen D, Van Embden JD (декабрь 1993 г.). «Природа полиморфизма ДНК в прямом повторном кластере микобактерий туберкулеза; применение для дифференцировки штамма с помощью нового метода типирования». Молекулярная микробиология . 10 (5): 1057–1065. doi : 10.1111/j.1365-2958.1993.tb00976.x . PMID 7934856 . S2CID 25304723 .
- ^ Jump up to: а беременный в Mojica FJ, Montoliu L (2016). «О происхождении технологии CRISPR-CAS: от прокариот до млекопитающих». Тенденции в микробиологии . 24 (10): 811–820. doi : 10.1016/j.tim.2016.06.005 . PMID 27401123 .
- ^ Jump up to: а беременный Mojica FJ, Rodriguez-Valera F (2016). «Открытие CRISPR в археи и бактериях» (PDF) . Журнал FEBS . 283 (17): 3162–3169. doi : 10.1111/febs.13766 . HDL : 10045/57676 . PMID 27234458 . S2CID 42827598 . Архивировано (PDF) из оригинала на 2021-02-25 . Получено 2019-08-29 .
- ^ Mojica FJ, Díez-Villaseñor C, Soria E, Juez G (апрель 2000 г.). «Биологическое значение семейства регулярно распределенных повторений в геномах археи, бактерий и митохондрий» . Молекулярная микробиология . 36 (1): 244–246. doi : 10.1046/j.1365-2958.2000.01838.x . PMID 10760181 .
- ^ Исааксон W (2021). Кодекс прерыватель: Дженнифер Дудна, редактирование генов и будущее человеческой расы . Нью -Йорк: Саймон и Шустер. п. 73. ISBN 978-1-9821-1585-2 Полем OCLC 1239982737 . Архивировано из оригинала 2023-01-14 . Получено 2021-10-20 .
- ^ Barrangou R , Van der Oost J (2013). Системы CRISPR-CAS: РНК-опосредованный адаптивный иммунитет у бактерий и археи . Гейдельберг: Спрингер. п. 6. ISBN 978-3-642-34656-9 .
- ^ Тан Т.Х., Бачеллер Дж.П., Рождественский Т., Бортолин М.Л., Хубер Х., Друнгски М. и др. (Май 2002 г.). «Идентификация 86 кандидатов для мелких нецентёрных РНК из археонов археоглобуса Fulgidus» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (11): 7536–7541. Bibcode : 2002pnas ... 99,7536t . doi : 10.1073/pnas.112047299 . PMC 124276 . PMID 12032318 .
- ^ Charpentier E, Richter H, Van der Oost J, White MF (май 2015). «Пути биогенеза РНК-гидов в архейном и бактериальном адаптивном иммунитете CRISPR-Cas» . Обзоры микробиологии FEMS . 39 (3): 428–441. doi : 10.1093/femsre/fuv023 . PMC 5965381 . PMID 25994611 .
- ^ Ромеро Д.А., Магилл Д., Миллен А., Хорват П., Фремо С (ноябрь 2020 г.). «Молочные лактококковые и стрептококковые взаимодействия фага-хоста: промышленная перспектива в развивающемся фаг-ландшафте». Обзоры микробиологии FEMS . 44 (6): 909–932. doi : 10.1093/femsre/fuaa048 . PMID 33016324 .
- ^ Molteni M, Huckins G (1 августа 2020 г.). «Проводное руководство по CRISPR» . Конде Наст. Проводной журнал. Архивировано из оригинала 23 октября 2021 года . Получено 23 февраля 2021 года .
- ^ Янсен Р., Эмбден Д.Д., Гаастра В., Шулс Л.М. (март 2002 г.). «Идентификация генов, которые связаны с повторениями ДНК в прокариотах» . Молекулярная микробиология . 43 (6): 1565–1575. doi : 10.1046/j.1365-2958.2002.02839.x . PMID 11952905 . S2CID 23196085 .
- ^ Jump up to: а беременный Horvath P, Barrangou R (январь 2010 г.). «CRISPR/CAS, иммунная система бактерий и археи». Наука . 327 (5962): 167–170. Bibcode : 2010sci ... 327..167H . doi : 10.1126/science.1179555 . PMID 20056882 . S2CID 17960960 .
- ^ Jump up to: а беременный Marraffini LA, Sontheimer EJ (март 2010 г.). «Вмешательство CRISPR: РНК-направленный адаптивный иммунитет у бактерий и археи» . Nature Reviews Genetics . 11 (3): 181–190. doi : 10.1038/nrg2749 . PMC 2928866 . PMID 20125085 .
- ^ Grissa I, Vergnaud G, Pourcel C (май 2007 г.). «База данных и инструменты CRISPRDB для отображения CRISPR и создания словари проставки и повторений» . BMC Bioinformatics . 8 : 172. DOI : 10.1186/1471-2105-8-172 . PMC 1892036 . PMID 17521438 .
- ^ Jump up to: а беременный Pourcel C, Salvignol G, Vergnaud G (март 2005 г.). «Элементы CRISPR в Yersinia pestis приобретают новые повторения за счет преимущественного поглощения ДНК бактериофагов и предоставляют дополнительные инструменты для эволюционных исследований» . Микробиология . 151 (Pt 3): 653–663. doi : 10.1099/mic.0.27437-0 . PMID 15758212 .
- ^ Jump up to: а беременный Mojica FJ, Díez-Villaseñor C, García-Martínez J, Soria E (февраль 2005 г.). «Промежуточные последовательности регулярно расположенных прокариотических повторов происходят из иностранных генетических элементов». Журнал молекулярной эволюции . 60 (2): 174–182. Bibcode : 2005jmole..60..174M . doi : 10.1007/s00239-004-0046-3 . PMID 15791728 . S2CID 27481111 .
- ^ Jump up to: а беременный Болотин А., Квинквиз Б., Сорокин А., Эрлих С.Д. (август 2005 г.). «Кластерные регулярно межпрессовые короткие палиндромные повтора (CRISPR) имеют проставки экстрахромосомного происхождения» . Микробиология . 151 (Pt 8): 2551–2561. doi : 10.1099/mic.0.28048-0 . PMID 16079334 .
- ^ Morange M (июнь 2015 г.). «То, что история рассказывает нам XXXVII. CRISPR-CAS: открытие иммунной системы у прокариот» (PDF) . Журнал биологических наук . 40 (2): 221–223. doi : 10.1007/s12038-015-9532-6 . PMID 25963251 .
- ^ Lander ES (январь 2016 г.). "Герои Crispr" . Клетка . 164 (1–2): 18–28. doi : 10.1016/j.cell.2015.12.041 . PMID 26771483 .
- ^ Макарова К.С., Гришин Н.В., Шабалина С.А., Вольф Йи, Кунин Э.В. (март 2006 г.). «Предполагаемая иммунная система на основе РНК в прокариотах: вычислительный анализ прогнозируемого ферментативного механизма, функциональные аналогии с эукариотической РНКи и гипотетические механизмы действия» . Биология прямой . 1 : 7. doi : 10.1186/1745-6150-1-7 . PMC 1462988 . PMID 16545108 .
- ^ Pennisi E (август 2013 г.). "Crispr Craze". Новости фокусируется. Наука . 341 (6148): 833–836. Bibcode : 2013sci ... 341..833p . doi : 10.1126/science.341.6148.833 . PMID 23970676 .
- ^ Jump up to: а беременный в Marraffini LA (октябрь 2015). «Иммунитет CRISPR-CAS у прокариот». Природа . 526 (7571): 55–61. Bibcode : 2015natur.526 ... 55m . doi : 10.1038/nature15386 . PMID 26432244 . S2CID 3718361 .
- ^ Brouns SJ, Jore MM, Lundgren M, Westra ER, Section RJ, Spartiary AP, et al. (Август 2008 г.). «Убьем Crisr RNAS Guide Antiviral определяет в прокаритах» . Наука . 321 (5891): 960–964. Код BIB : 2008Sci… 321..960b . doi : 10,1126/science.1159689 . PMC 5898235 . PMID 187039 .
- ^ Jump up to: а беременный Garneau JE, Dupuis Mè, Vollion M, Romero DA, Barangou R , Boyaval P, et al. (Ноябрь 2010). «Бактериальная иммунная система CRISPR/CAS расщепляет бактериофаг и плазмидную ДНК» Природа 468 (7320): 67–7 Bibcode : 2010natur.468 ... 67G Citeseerx 10.1.1.451.9 Doi : 10.1038/ nature0 PMID 21048762 S2CID 205222849
- ^ Jump up to: а беременный Deltcheva E, Chylinski K, Sharma CM, Gonzales K, Chao Y, Pirzada Za, et al. (Март 2011 г.). «Созревание РНК CRISPR с помощью транскодированной небольшой РНК и фактора-хозяина РНКазы III» . Природа . 471 (7340): 602–607. Bibcode : 2011natur.471..602d . doi : 10.1038/nature09886 . PMC 3070239 . PMID 21455174 .
- ^ Баррангу р (ноябрь 2015). «Разнообразие иммунной системы CRISPR-CAS и молекулярных машин» . Биология генома . 16 : 247. DOI : 10.1186/S13059-015-0816-9 . PMC 4638107 . PMID 26549499 .
- ^ Jump up to: а беременный Джинк М., Чилински К., Фонфара И., Хауэр М., Доудна Дж.А. , Чарпентье Е (август 2012 г.). «Программируемая двойная РНК-эндонуклеаза ДНК при адаптивном бактериальном иммунитете» . Наука . 337 (6096): 816–821. Bibcode : 2012sci ... 337..816j . doi : 10.1126/science.1225829 . PMC 6286148 . PMID 22745249 .
- ^ Cong L, Ran FA, Cox D, Lin S, Barretto R, Habib N, et al. (Февраль 2013 г.). «Мультиплексная инженерия генома с использованием систем CRISPR/CAS» . Наука . 339 (6121): 819–823. Bibcode : 2013sci ... 339..819c . doi : 10.1126/science.1231143 . PMC 3795411 . PMID 23287718 .
- ^ Мали П., Ян Л., Эсвельт К.М., Аач Дж., Гуэлл М., Дикарло Дж. И. и др. (Февраль 2013 г.). «РНК-управляемая инженериями генома человека через CAS9» . Наука . 339 (6121): 823–826. Bibcode : 2013sci ... 339..823M . doi : 10.1126/science.1232033 . PMC 3712628 . PMID 23287722 .
- ^ Dicarlo JE, Norville JE, Mali P, Rios X, Aach J, Church GM (апрель 2013 г.). «Инженер генома в Saccharomyces cerevisiae с использованием систем Crispr-Cas» . Исследование нуклеиновых кислот . 41 (7): 4336–4343. doi : 10.1093/nar/gkt135 . PMC 3627607 . PMID 23460208 .
- ^ Zhang GC, Kong II, Kim H, Liu JJ, Cate JH, Jin YS (декабрь 2014 г.). «Строительство четырехкратного ауксотрофного мутанта промышленного полиплоидного штамма Saccharomyces cerevisiae с использованием нуклеазы CAS9 с РНК» . Прикладная и экологическая микробиология . 80 (24): 7694–7701. Bibcode : 2014Apenm..80.7694Z . doi : 10.1128/aem.02310-14 . PMC 4249234 . PMID 25281382 .
- ^ Лю Дж.Дж., Конг II, Чжан Г.С., Джаякоди Л.Н., Ким Х., Ся П.Ф. и др. (Апрель 2016 г.). «Метаболическая инженерия пробиотического Saccharomyces boulardii » . Прикладная и экологическая микробиология . 82 (8): 2280–2287. BIBCODE : 2016Apenm..82.2280L . doi : 10.1128/aem.00057-16 . PMC 4959471 . PMID 26850302 .
- ^ Вьяс В.К., Барраса М.И., Финк Г.Р. (2015). « Система Candida Albicans Crispr позволяет генетической инженерии важных генов и семейств генов» . Наука достижения . 1 (3): E1500248. Bibcode : 2015scia .... 1E0248V . doi : 10.1126/sciadv.1500248 . PMC 4428347 . PMID 25977940 .
- ^ Нг Х, Дин Н. (2017). « Candida albicans по повышению экспрессии РНК с одним направляющим» . MSPHERE . 2 (2): E00385–16. doi : 10.1128/msphere.00385-16 . PMC 5397569 . PMID 28435892 .
- ^ Hwang Wy, Fu Y, Reyon D, Maeder ML, Tsai SQ, Sander JD, et al. (Март 2013). «Эффективное редактирование генома у рыбок данио с использованием системы CRISPR-CAS» . Nature Biotechnology . 31 (3): 227–229. doi : 10.1038/nbt.2501 . PMC 3686313 . PMID 23360964 .
- ^ Гратц С.Дж., Каммингс А.М., Нгуен Дж.Н., Хэмм Д.К., Донохью Л.К., Харрисон М.М. и др. (Август 2013). «Геном инженерия Drosophila с помощью RNA-нуклеазы CAS9, управляемой CRISPR» . Генетика . 194 (4): 1029–1035. doi : 10.1534/Genetics.113.152710 . PMC 3730909 . PMID 23709638 .
- ^ Bassett AR, Tibbit C, Ponting CP, Liu JL (июль 2013). «Высокоэффективный целевой мутагенез дрозофилы с системой CRISPR/CAS9» . Сотовые отчеты . 4 (1): 220–228. doi : 10.1016/j.celrep.2013.06.020 . PMC 3714591 . PMID 23827738 .
- ^ Ян Х., Опалоемфан С., Манчини Г., Ян Х, Галлитто М., Млежнек Дж. И др. (Август 2017). «Инженерная мутация ORCO вызывает аберрантное социальное поведение и дефектное развитие нейронности у муравьев» . Клетка . 170 (4): 736–747.e9. doi : 10.1016/j.cell.2017.06.051 . PMC 5587193 . PMID 28802043 .
- ^ Trible W, Olivos-Cisneros L, McKenzie SK, Saragosti J, Chang NC, Matthews BJ, et al. (Август 2017). «Мутагенез Orco вызывает потерю антеннальной доли клубочков и нарушение социального поведения у муравьев» . Клетка . 170 (4): 727–735.e10. doi : 10.1016/j.cell.2017.07.001 . PMC 5556950 . PMID 28802042 .
- ^ Кистлер К.Е., Восхолл Л.Б., Мэтьюз Б.Дж. (апрель 2015). «Инженерия генома с Crispr-Cas9 в комаре Aedes aegypti» . Сотовые отчеты . 11 (1): 51–60. doi : 10.1016/j.celrep.2015.03.009 . PMC 4394034 . PMID 25818303 .
- ^ Фридленд А.Е., Цур Ю.Б., Эсвельт К.М., Колайаково М.М., Церковь Г.М., Каларко Дж.А. (август 2013 г.). «Редактирование наследственного генома у C. elegans через систему Crispr-Cas9» . Природные методы . 10 (8): 741–743. doi : 10.1038/nmeth.2532 . PMC 3822328 . PMID 23817069 .
- ^ Jiang W, Zhou H, Bi H, Fromm M, Yang B, Weeks DP (ноябрь 2013 г.). «Демонстрация CRISPR/CAS9/SGRNA-опосредованная модификация гена у арабидопсиса, табака, сорго и риса» . Исследование нуклеиновых кислот . 41 (20): E188. doi : 10.1093/nar/gkt780 . PMC 3814374 . PMID 23999092 .
- ^ Wang H, Yang H, Shivalila CS, Dawlaty MM, Cheng AW, Zhang F , et al. (Май 2013). «Одноэтапное поколение мышей, несущих мутации в нескольких генах с помощью CRISPR/CAS-опосредованного генома инженерии» . Клетка . 153 (4): 910–918. doi : 10.1016/j.cell.2013.04.025 . PMC 3969854 . PMID 23643243 .
- ^ Сони Д., Ван Д.М., Регми С.К., Миттал М., Фогель С.М., Шлютер Д. и др. (Май 2018). «Функция Deubiquinase A20 поддерживает и ремонтирует эндотелиальный барьер после травмы сосудов легких» . Открытие гибели клеток . 4 (60): 60. doi : 10.1038/s41420-018-0056-3 . PMC 5955943 . PMID 29796309 .
- ^ Guo X, Li XJ (июль 2015 г.). «Целевое редактирование генома у эмбрионов приматов» . Клеточные исследования . 25 (7): 767–768. doi : 10.1038/cr.2015.64 . PMC 4493275 . PMID 26032266 .
- ^ Балтимор Д., Берг П., Ботчан М., Кэрролл Д., Чаро Р.А., Черч Г. и др. (Апрель 2015). «Биотехнология. Представленный путь вперед для геномной инженерии и модификации генов зародышевой линии» . Наука . 348 (6230): 36–38. Bibcode : 2015sci ... 348 ... 36b . doi : 10.1126/science.aab1028 . PMC 4394183 . PMID 25791083 .
- ^ Larson MH, Gilbert LA, Wang X, Lim WA, Weissman JS, Qi LS (ноябрь 2013 г.). «Интерференция CRISPR (CRISPRI) для последовательно-специфической для контроля экспрессии генов» . Природные протоколы . 8 (11): 2180–2196. doi : 10.1038/nprot.2013.132 . PMC 3922765 . PMID 24136345 .
- ^ Liang P, Xu Y, Zhang X, Ding C, Huang R, Zhang Z, et al. (Май 2015). «Редактирование генов CRISPR/CAS9 у человеческого трипноядерного зигот» . Белок и клетка . 6 (5): 363–372. doi : 10.1007/s13238-015-0153-5 . PMC 4417674 . PMID 25894090 .
- ^ Yan My, Yan HQ, Ren GX, Zhao JP, Guo XP, Sun YC (сентябрь 2017 г.). «Рекомбинирирование с помощью CRISPR-CAS12A у бактерий» . Прикладная и экологическая микробиология . 83 (17). Bibcode : 2017apenm..83e.947y . doi : 10.1128/aem.00947-17 . PMC 5561284 . PMID 28646112 .
- ^ Zetsche B, Goootenberg JS, Abudayyeh OO, Slaymaker IM, Makarova KS, Essletzbichler P, et al. (Октябрь 2015). «CPF1-это единая РНК-эндонуклеаза системы CRISPR-CAS класса 2» . Клетка . 163 (3): 759–771. doi : 10.1016/j.cell.2015.09.038 . PMC 4638220 . PMID 26422227 .
- ^ Фонфара I, Рихтер Х, Братович М., Ле Рун А., Чарпентье Е (апрель 2016 г.). «Ассоциированный CRISPR Фермент ДНК CPF1 также обрабатывает предшественник CRISPR РНК». Природа . 532 (7600): 517–521. Bibcode : 2016natur.532..517f . doi : 10.1038/nature17945 . PMID 27096362 . S2CID 2271552 .
- ^ Ким Х., Ким С., Рю Дж., Кан Б.К., Ким Дж.С. и Ким С.Г. (февраль 2017 г.). «CRISPR/CPF1-опосредованное ДНК-без ДНК Редактирование генома растений» . Природная связь . 8 (14406): 14406. Bibcode : 2017natco ... 814406K . doi : 10.1038/ncomms14406 . PMC 5316869 . PMID 28205546 .
- ^ "CPF1 нуклеаза" . abmgood.com . Архивировано из оригинала 2021-10-23 . Получено 2017-12-14 .
- ^ Chen JS, Ma E, Harrington LB, Da Costa M, Tian X, Palefsky JM, et al. (Апрель 2018). «CRISPR-CAS12A-связывание связывания развязывает неразборчивую одноцепочечную активность ДНКазы» . Наука . 360 (6387): 436–439. Bibcode : 2018sci ... 360..436c . doi : 10.1126/science.aar6245 . PMC 6628903 . PMID 29449511 .
- ^ Broughton JP, Deng X, Yu G, Fasching CL, Servellita V, Singh J, et al. (Июль 2020 г.). «Обнаружение SARS-COV-2 на основе CRISPR-CAS12» . Nature Biotechnology . 38 (7): 870–874. doi : 10.1038/s41587-020-0513-4 . PMC 9107629 . PMID 32300245 .
- ^ Нгуен Л.Т., Смит Б.М., Джайн П.К. (сентябрь 2020 г.). «Улучшение транс-расщепления Cas12a с помощью инженерной крольсы обеспечивает амплифицированное обнаружение нуклеиновых кислот» . Природная связь . 11 (1): 4906. Bibcode : 2020natco..11.4906n . doi : 10.1038/s41467-020-18615-1 . PMC 7528031 . PMID 32999292 .
- ^ Abudayyeh OO, Goootenberg JS, Konermann S, Joung J, Slaymaker IM, Cox DB, et al. (Август 2016 г.). «C2C2 представляет собой однокомпонентный программируемый РНК-контролируемый РНК-целевой эффектор CRISPR» . Наука . 353 (6299): AAF5573. doi : 10.1126/science.aaf5573 . PMC 5127784 . PMID 27256883 .
- ^ Gootenberg JS, Abudayyeh OO, Lee JW, Essletzbichler P, Dy AJ, Joung J, et al. (Апрель 2017). «Обнаружение нуклеиновых кислот с CRISPR-CAS13A/C2C2» . Наука . 356 (6336): 438–442. Bibcode : 2017sci ... 356..438g . doi : 10.1126/science.aam9321 . PMC 5526198 . PMID 28408723 .
- ^ Gootenberg JS, Abudayyeh OO, Kellner MJ, Joung J, Collins JJ, Zhang F (апрель 2018 г.). «Мультиплексированная и портативная платформа обнаружения нуклеиновых кислот с CAS13, CAS12A и CSM6» . Наука . 360 (6387): 439–444. Bibcode : 2018sci ... 360..439G . doi : 10.1126/science.aaq0179 . PMC 5961727 . PMID 29449508 .
- ^ Iwasaki RS, Batey RT (сентябрь 2020 г.). «Спринт: платформа на основе CAS13A для обнаружения мелких молекул» . Исследование нуклеиновых кислот . 48 (17): E101. doi : 10.1093/nar/gkaa673 . PMC 7515716 . PMID 32797156 .
- ^ Махас А., Ван К, Марсич Т., Махфуз М.М. (октябрь 2021 г.). «Новая миниатюрная система CRISPR-CAS13 для диагностики SARS-COV-2» . ACS Синтетическая биология . 10 (10): 2541–2551. doi : 10.1021/acssynbio.1c00181 . PMC 8482783 . PMID 34546709 .
- ^ Hille F, Charpentier E (ноябрь 2016 г.). «CRISPR-CAS: биология, механизмы и актуальность» . Философские транзакции Королевского общества Лондона. Серия B, биологические науки . 371 (1707): 20150496. DOI : 10.1098/rstb.2015.0496 . PMC 5052741 . PMID 27672148 .
- ^ Jump up to: а беременный в Баррангу Р. , Марраффини Л.А. (апрель 2014 г.). «Системы CRISPR-CAS: прокариоты обновляются до адаптивного иммунитета» . Молекулярная клетка . 54 (2): 234–244. doi : 10.1016/j.molcel.2014.03.011 . PMC 4025954 . PMID 24766887 .
- ^ Jump up to: а беременный в Тайсон Г.В., Банфилд Дж.Ф. (январь 2008 г.). «Быстро развивающиеся CRISPR, участвующие в приобретенной устойчивости микроорганизмов к вирусам». Экологическая микробиология . 10 (1): 200–207. Bibcode : 2008envmi..10..200t . doi : 10.1111/j.1462-2920.2007.01444.x . PMID 17894817 .
- ^ Jump up to: а беременный Koonin EV, Makarova KS (май 2019). «Происхождение и эволюция систем CRISPR-CAS» . Философские транзакции Королевского общества Лондона. Серия B, биологические науки . 374 (1772): 20180087. DOI : 10.1098/rstb.2018.0087 . PMC 6452270 . PMID 30905284 .
- ^ Jump up to: а беременный в Райт А.В., Нуньес Дж.К., Доудна Дж.А. (январь 2016 г.). «Биология и применение систем CRISPR: использование инструментов природы для инженерии генома» . Клетка . 164 (1–2): 29–44. doi : 10.1016/j.cell.2015.12.035 . PMID 26771484 .
- ^ Макарова К.С., Вольф Йи, Иранцо Дж., Шмаков С.А., Алхнбаши О.С., Брунс С.Дж. и др. (Декабрь 2019). «Эволюционная классификация систем CRISPR -CAS: взрыв класса 2 и полученных вариантов» . Nature Reviews Microbiology . 18 (1): 67–83. doi : 10.1038/s41579-019-0299-x . HDL : 10045/102627 . PMC 8905525 . PMID 31857715 .
- ^ Jump up to: а беременный в дюймовый и фон глин час Макарова К.С., Вольф Йи, Алхнбаши Ос, Коста Ф., Шах С.А., Сондерс С.Дж. и др. (Ноябрь 2015). «Обновленная эволюционная классификация систем CRISPR-CAS» . Природные обзоры. Микробиология . 13 (11): 722–736. doi : 10.1038/nrmicro3569 . PMC 5426118 . PMID 26411297 .
- ^ Jump up to: а беременный в Wiedenheft B, Sternberg SH, Doudna JA (февраль 2012 г.). «РНК-управляемые генетические системы молчания у бактерий и археи». Природа . 482 (7385): 331–338. Bibcode : 2012natur.482..331W . doi : 10.1038/nature10886 . PMID 22337052 . S2CID 205227944 .
- ^ Jump up to: а беременный Дэн Л., Гаррет Р.А., Шах С.А., Пенг Х, она Q (март 2013 г.). «Новый механизм интерференции с помощью модуля CRISPR-CMR типа IIIB в Sulfolobus». Молекулярная микробиология . 87 (5): 1088–1099. doi : 10.1111/mmi.12152 . PMID 23320564 .
- ^ Sinkunas T, Gasiunas G, Fremaux C, Barrangou R , Horvath P, Siksnys V (апрель 2011 г.). «CAS3 представляет собой одноцепочечную нуклеазу ДНК и АТФ-зависимая геликаза в иммунной системе CRISPR/CAS» . Embo Journal . 30 (7): 1335–1342. doi : 10.1038/emboj.2011.41 . PMC 3094125 . PMID 21343909 .
- ^ Huo Y, Nam KH, Ding F, Lee H, Wu L, Xiao Y, et al. (Сентябрь 2014). «Структуры CRISPR CAS3 дают механистическую информацию о раскручивании и деградации ДНК, активируемой каскадом» . Природа структурная и молекулярная биология . 21 (9): 771–777. doi : 10.1038/nsmb.2875 . PMC 4156918 . PMID 25132177 .
- ^ Brendel J, Stoll B, Lange SJ, Sharma K, Lenz C, Stachler AE, et al. (Март 2014 г.). «Комплекс белков CAS 5, 6 и 7 необходим для биогенеза и стабильности кластерных регулярно межсетенных коротких палиндромических повторений (CRISPR) РНК (CRRNAs) в вулкании Haloferax» . Журнал биологической химии . 289 (10): 7164–77. doi : 10.1074/jbc.m113.508184 . PMC 3945376 . PMID 24459147 .
- ^ Jump up to: а беременный Chylinski K, Makarova KS, Charpentier E, Koonin EV (июнь 2014 г.). «Классификация и эволюция систем CRISPR-CAS типа II» . Исследование нуклеиновых кислот . 42 (10): 6091–6105. doi : 10.1093/nar/gku241 . PMC 4041416 . PMID 24728998 .
- ^ Jump up to: а беременный Макарова К.С., Аравинд Л., Вольф Йи, Кунин Э.В. (июль 2011 г.). «Объединение семейств белков CAS и простой сценарий для происхождения и эволюции систем CRISPR-CAS» . Биология прямой . 6 : 38. doi : 10.1186/1745-6150-6-38 . PMC 3150331 . PMID 21756346 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. ведущий с Т в v В Макарова К.С., Вольф Йи, Иранцо Дж., Шмаков С.А., Алхнбаши О.С., Брунс С.Дж. и др. (Февраль 2020 г.). «Эволюционная классификация систем CRISPR-CAS: взрыв класса 2 и полученных вариантов» . Природные обзоры. Микробиология . 18 (2): 67–83. doi : 10.1038/s41579-019-0299-x . HDL : 10045/102627 . PMC 8905525 . PMID 31857715 . S2CID 209420490 .
- ^ Jump up to: а беременный Mogila I, Kazlauskiene M, Valinskyte S, Tamulaitiene G, Tamulaitis G, Siksnys V (март 2019 г.). «Генетическая рассечение комплекса системы CSM типа III-A CRISPR-CAS выявляет роли отдельных субъединиц» . Сотовые отчеты . 26 (10): 2753–2765.e4. doi : 10.1016/j.celrep.2019.02.029 . PMID 30840895 .
- ^ Jump up to: а беременный в Gasiunas G, Barrangou R , Horvath P, Siksnys V (сентябрь 2012 г.). «Комплекс рибонуклеопротеина Cas9-Crrna опосредует специфическое расщепление ДНК для адаптивного иммунитета у бактерий» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (39): E2579–2586. BIBCODE : 2012PNAS..109E2579G . doi : 10.1073/pnas.1208507109 . PMC 3465414 . PMID 22949671 .
- ^ Heler R, Samai P, Modell JW, Weiner C, Goldberg GW, Bikard D, et al. (Март 2015 г.). «CAS9 определяет функциональные вирусные мишени во время адаптации CRISPR-CAS» . Природа . 519 (7542): 199–202. Bibcode : 2015natur.519..199h . doi : 10.1038/nature14245 . PMC 4385744 . PMID 25707807 .
- ^ Nam KH, Kurinov I, Ke A (сентябрь 2011 г.). «Кристаллическая структура кластеризированных регулярно межсеплеевых коротких палиндромных повторений (CRISPR), ассоциированного с белком CSN2, выявил Ca2+-зависимую активность связывания ДНК» . Журнал биологической химии . 286 (35): 30759–30768. doi : 10.1074/jbc.m111.256263 . PMC 3162437 . PMID 21697083 .
- ^ Lee H, Dhingra Y, Sashital DG (апрель 2019 г.). «Комплекс CAS4-CAS1-CAS2 опосредует точную обработку Prespacer во время адаптации CRISPR» . элиф . 8 doi : 10.7554/elife.44248 . PMC 6519985 . PMID 31021314 .
- ^ Chylinski K, Le Rhun A, Charpentier E (май 2013). «Семейства Tracrrna и CAS9 систем иммунитета II типа II» . РНК -биология . 10 (5): 726–737. doi : 10.4161/rna.24321 . PMC 3737331 . PMID 23563642 .
- ^ Макарова К.С., Чжан Ф., Кунин Э.В. (январь 2017 г.). «Снимок: системы CRISPR-CAS класса 2» . Клетка . 168 (1–2): 328–328.e1. doi : 10.1016/j.cell.2016.12.038 . PMID 28086097 .
- ^ Пол Б., Монтойя Г. (февраль 2020 г.). «CRISPR-CAS12A: функциональный обзор и приложения» . Биомедицинский журнал . 43 (1): 8–17. doi : 10.1016/j.bj.2019.10.005 . PMC 7090318 . PMID 32200959 .
- ^ Cox DB, Gootenberg JS, Abudayyeh OO, Franklin B, Kellner MJ, Joung J, et al. (Ноябрь 2017). «Редактирование РНК с CRISPR-CAS13» . Наука . 358 (6366): 1019–1027. BIBCODE : 2017SCI ... 358.1019C . doi : 10.1126/science.aaq0180 . PMC 5793859 . PMID 29070703 .
- ^ Jump up to: а беременный Xu C, Zhou Y, Xiao Q, He B, Geng G, Wang Z, et al. (Май 2021 г.). «Программируемое редактирование РНК с компактными системами CRISPR-CAS13 из некультивированных микробов» (PDF) . Природные методы . 18 (5): 499–506. doi : 10.1038/s41592-021-01124-4 . PMID 33941935 . S2CID 233719501 . Архивировано (PDF) из оригинала на 2024-05-12 . Получено 2024-05-12 .
- ^ Азангу-Кхьяви М., Гасеми М., Ханали Дж., Борим и-Сабур М., Джамалхах М., Солеймани М. и др. (2020). «CRISPR/CAS: от редактирования генов опухоли до иммунотерапии рака на основе Т-клеток» . Границы в иммунологии . 11 : 2062. DOI : 10.3389/fimmu.2020.02062 . PMC 7553049 . PMID 33117331 .
- ^ Jump up to: а беременный Aliyari R, Ding SW (январь 2009 г.). «Вирусный иммунитет на основе РНК, инициированный семейством иммунных рецепторов хозяина-хозяина» . Иммунологические обзоры . 227 (1): 176–188. doi : 10.1111/j.1600-065x.2008.00722.x . PMC 2676720 . PMID 19120484 .
- ^ Дугар Г., Хербиг А., Форстнер Ку, Хайдрих Н., Рейнхардт Р., Низельт К. и др. (Май 2013). «Карты транскриптома высокого разрешения показывают специфичные для штамма регуляторные особенности множественных изолятов Campylobacter Jejuni» . PLOS Genetics . 9 (5): E1003495. doi : 10.1371/journal.pgen.1003495 . PMC 3656092 . PMID 23696746 .
- ^ Hatoum-Aslan A, Maniv I, Marraffini LA (декабрь 2011 г.). «Зрелая кластеризованная, регулярно межсетенная, короткие палиндромные повторения РНК (CRRNA) измеряется механизмом правителя, закрепленным на месте обработки предшественника» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (52): 21218–21222. Bibcode : 2011pnas..10821218H . doi : 10.1073/pnas.1112832108 . PMC 3248500 . PMID 22160698 .
- ^ Jump up to: а беременный Yosef I, Goren Mg, Qimron U (июль 2012 г.). «Белки и ДНК -элементы, необходимые для процесса адаптации CRISPR в Escherichia coli » . Исследование нуклеиновых кислот . 40 (12): 5569–5576. doi : 10.1093/nar/gks216 . PMC 3384332 . PMID 22402487 .
- ^ Jump up to: а беременный в дюймовый Swarts DC, Mosterd C, Van Passel MW, Brouns SJ (2012). «Вмешательство CRISPR направляет конкретное приобретение проставки с прядью» . Plos один . 7 (4): E35888. BIBCODE : 2012PLOSO ... 735888S . doi : 10.1371/journal.pone.0035888 . PMC 3338789 . PMID 22558257 .
- ^ Бабу М. , Белоглазова Н., Флик Р., Грэм С., Скарина Т., Носек Б. и др. (Январь 2011). «Двойная функция системы CRISPR-CAS в бактериальном антивирусном иммунитете и репарации ДНК» . Молекулярная микробиология . 79 (2): 484–502. doi : 10.1111/j.1365-2958.2010.07465.x . PMC 3071548 . PMID 21219465 .
- ^ Хан Д., Леманн К, Краусс Г (июнь 2009 г.). «SSO1450 - белок CAS1 из Sulfolobus solfataricus p2 с высоким сродством к РНК и ДНК». Письма Febs . 583 (12): 1928–1932. Bibcode : 2009 Febsl.583.1928h . doi : 10.1016/j.febslet.2009.04.047 . PMID 19427858 . S2CID 22279972 .
- ^ Wiedenheft B, Zhou K, Jinek M, Coyle SM, MA W, Doudna JA (июнь 2009 г.). «Структурная основа для ДНКазы консервативного белка, участвующего в CRISPR-опосредованной защите генома» . Структура 17 (6): 904–912. doi : 10.1016/j.str.2009.03.019 . PMID 19523907 .
- ^ Beloglazova N, Brown G, Zimmerman MD, Proudfoot M, Makarova KS, Kudritska M, et al. (Июль 2008 г.). «Новое семейство специфичных для последовательности эндорибонуклеаз, связанных с кластеризированными регулярно межсепесенными короткими палиндромическими повторами» . Журнал биологической химии . 283 (29): 20361–20371. doi : 10.1074/jbc.m803225200 . PMC 2459268 . PMID 18482976 .
- ^ Самай П, Смит П., Шуман С (декабрь 2010 г.). «Структура белка, ассоциированного с CRISPR, из Desulfovibrio vulgaris» . Acta Crystallographica Раздел f . 66 (Pt 12): 1552–1556. doi : 10.1107/s1744309110039801 . PMC 2998353 . PMID 21139194 .
- ^ Nam KH, Ding F, Haitjema C, Huang Q, Delisa MP, KE A (октябрь 2012 г.). «Двухцепочечная эндонуклеационная активность в бациллевых галодуранских складывании регулярно мешает межсеплее коротких палиндромических повторений (CRISPR), связанного с белком CAS2,» . Журнал биологической химии . 287 (43): 35943–35952. doi : 10.1074/jbc.m112.382598 . PMC 3476262 . PMID 22942283 .
- ^ Jump up to: а беременный Nuñez JK, Kranzusch PJ, Noeske J, Wright AV, Davies CW, Doudna JA (июнь 2014 г.). «Формирование комплекса CAS1-CAS2 опосредует приобретение спейсеров во время адаптивного иммунитета CRISPR-CAS» . Природа структурная и молекулярная биология . 21 (6): 528–534. doi : 10.1038/nsmb.2820 . PMC 4075942 . PMID 24793649 .
- ^ Nuñez JK, Lee As, Engelman A, Doudna Ja (март 2015 г.). «Интеграционная опосредованная приобретение проставки во время адаптивного иммунитета CRISPR-CAS» . Природа . 519 (7542): 193–198. Bibcode : 2015natur.519..193n . doi : 10.1038/nature14237 . PMC 4359072 . PMID 25707795 .
- ^ Wang J, Li J, Zhao H, Sheng G, Wang M, Yin M, et al. (Ноябрь 2015). «Структурная и механистическая основа PAM-зависимого приобретения спейсеров в системах CRISPR-CAS» . Клетка . 163 (4): 840–853. doi : 10.1016/j.cell.2015.10.008 . PMID 26478180 .
- ^ Nuñez JK, Harrington LB, Kranzusch PJ, Engelman AN, Doudna JA (ноябрь 2015). «Захват иностранного ДНК во время адаптивного иммунитета CRISPR-Cas» . Природа . 527 (7579): 535–538. Bibcode : 2015natur.527..535n . doi : 10.1038/nature15760 . PMC 4662619 . PMID 26503043 .
- ^ Sorek R, Lawrence CM, Wiedenheft B (2013). «CRISPR-опосредованная адаптивная иммунная система у бактерий и археи» . Ежегодный обзор биохимии . 82 (1): 237–266. doi : 10.1146/annurev-biochem-072911-172315 . PMID 23495939 .
- ^ Nuñez JK, Bai L, Harrington LB, Hinder TL, Doudna JA (июнь 2016 г.). «Иммунологическая память CRISPR требует фактора хозяина для специфичности» . Молекулярная клетка . 62 (6): 824–833. doi : 10.1016/j.molcel.2016.04.027 . PMID 27211867 .
- ^ Fagerlund RD, Wilkinson ME, Klykov O, Barendregt A, Pearce FG, Kieper SN, et al. (Июнь 2017 г.). «Захват и интеграция с помощью типа, если Cas1-Cas2–3 адаптационный комплекс CRISPR» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (26): E5122 - E5128. Bibcode : 2017pnas..114e5122f . doi : 10.1073/pnas.1618421114 . PMC 5495228 . PMID 28611213 .
- ^ Rollie C, Graham S, Rouillon C, White MF (февраль 2018 г.). «Обработка Prespacer и конкретная интеграция в системе CRISPR типа IA» . Исследование нуклеиновых кислот . 46 (3): 1007–1020. doi : 10.1093/nar/gkx1232 . PMC 5815122 . PMID 29228332 .
- ^ Jump up to: а беременный в дюймовый Хорват П., Ромеро Д.А., Коуте-Монвуазин А.С., Ричардс М., Дево Х., Моино С. и др. (Февраль 2008 г.). «Разнообразие, активность и эволюция локусов CRISPR в Streptococcus thermophilus » . Журнал бактериологии . 190 (4): 1401–1412. doi : 10.1128/jb.01415-07 . PMC 2238196 . PMID 18065539 .
- ^ Jump up to: а беременный в Deveau H, Barrangou R , Garneau I, Labonté J, Fremaux C, Boyaval P, et al. (Февраль 2008 г.). «Ответ фага на устойчивость к CRISPR у Streptococcus thermophilus » . Журнал бактериологии . 190 (4): 1390–1400. Doi : 10.1128/jb.01412-07 . PMC 2238228 . PMID 18065545 .
- ^ García-M, Aldeds C (март 2009 г.). "Короткие ходы целей целей Микробиология 155 (Pt 3): 733–7 doi : 10.1099/ mic.023960-0 PMID 19246744 .
- ^ Jump up to: а беременный Lillestøl RK, Shah SA, Brügger K, Redder P, Phan H, Christiansen J, et al. (Апрель 2009 г.). «Семейства CRISPR из рода Crenarchaeal Sulfolobus: двунаправленная транскрипция и динамические свойства». Молекулярная микробиология . 72 (1): 259–272. doi : 10.1111/j.1365-2958.2009.06641.x . PMID 19239620 . S2CID 36258923 .
- ^ Jump up to: а беременный Shah SA, Hansen NR, Garrett RA (февраль 2009 г.). «Распределение спейсеров CRISPR в вирусах и плазмидах ацидоотермофилов Crenarchaeal и последствия для их ингибирующего механизма». Биохимическое общество транзакций . 37 (Pt 1): 23–28. doi : 10.1042/bst0370023 . PMID 19143596 . S2CID 19093261 .
- ^ Jump up to: а беременный Díez-Villaseñor C, Guzmán NM, Almendros C, García-Martínez J, Mojica FJ (май 2013). CRISPR-CAS «Репортерные плазмиды интеграции CRISPR-SPACER выявляют различную подлинную специфику приобретения среди вариантов Escherichia coli » . РНК -биология . 10 (5): 792–802. doi : 10.4161/rna.24023 . PMC 3737337 . PMID 23445770 .
- ^ Jump up to: а беременный в Erdmann S, Garrett RA (сентябрь 2012 г.). «Селективное и гиперактивное поглощение иностранной ДНК адаптивной иммунной системой археона с помощью двух различных механизмов» . Молекулярная микробиология . 85 (6): 1044–1056. doi : 10.1111/j.1365-2958.2012.08171.x . PMC 3468723 . PMID 22834906 .
- ^ Jump up to: а беременный Shah SA, Erdmann S, Mojica FJ, Garrett RA (май 2013). «Мотивы распознавания протоспацера: смешанная идентичность и функциональное разнообразие» . РНК -биология . 10 (5): 891–899. doi : 10.4161/rna.23764 . PMC 3737346 . PMID 23403393 .
- ^ Andersson AF, Banfield JF (май 2008 г.). «Динамика популяции вирусов и приобретенная устойчивость к вирусам в природных микробных сообществах». Наука . 320 (5879): 1047–1050. Bibcode : 2008Sci ... 320.1047a . doi : 10.1126/science.1157358 . PMID 18497291 . S2CID 26209623 .
- ^ Jump up to: а беременный в дюймовый Pride DT, Sun CL, Salzman J, Rao N, Loomer P, Armitage GC, et al. (Январь 2011). «Анализ стрептококковых CRISPR из слюны человека выявляет существенное разнообразие последовательностей у субъектов и между субъектами с течением времени» . Исследование генома . 21 (1): 126–136. doi : 10.1101/gr.111732.110 . PMC 3012920 . PMID 21149389 .
- ^ Jump up to: а беременный Goren MG, Yosef I, Auster O, Qimron U (октябрь 2012 г.). «Экспериментальное определение кластерного регулярного межсепенсации короткого палиндромного дубликона в Escherichia coli ». Журнал молекулярной биологии . 423 (1): 14–16. doi : 10.1016/j.jmb.2012.06.037 . PMID 22771574 .
- ^ Jump up to: а беременный в Datsenko Ka, Pougach K, Tikhonov A, Wanner BL, Severinov K, Semenova E (июль 2012 г.). «Молекулярная память предшествующих инфекций активирует систему адаптивного иммунитета CRISPR/CAS» . Природная связь . 3 : 945. Bibcode : 2012natco ... 3..945d . doi : 10.1038/ncomms1937 . PMID 22781758 .
- ^ Геснер Э.М., Шелленберг М.Дж., Гарсайд Эль, Джордж М.М., Макмиллан А.М. (июнь 2011 г.). «Признание и созревание эффекторных РНК в пути интерференции CRISPR». Природа структурная и молекулярная биология . 18 (6): 688–692. doi : 10.1038/nsmb.2042 . PMID 21572444 . S2CID 677704 .
- ^ Sashital DG, Jinek M, Doudna JA (июнь 2011 г.). «РНК-индуцированное конформационное изменение, необходимое для расщепления РНК CRISPR от эндорибонуклеазы CSE3». Природа структурная и молекулярная биология . 18 (6): 680–687. doi : 10.1038/nsmb.2043 . PMID 21572442 . S2CID 5538195 .
- ^ Haurwitz RE, Jinek M, Wiedenheft B, Zhou K, Doudna JA (сентябрь 2010 г.). «Последовательность и структура-специфическая обработка РНК с помощью эндонуклеазы CRISPR» . Наука . 329 (5997): 1355–1358. Bibcode : 2010sci ... 329.1355H . doi : 10.1126/science.1192272 . PMC 3133607 . PMID 20829488 .
- ^ Jump up to: а беременный Kunin V, Sorek R, Hugenholtz P (2007). «Эволюционное сохранение последовательности и вторичных структур в повторениях CRISPR» . Биология генома . 8 (4): R61. doi : 10.1186/gb-2007-8-4-r61 . PMC 1896005 . PMID 17442114 .
- ^ Carte J, Wang R, Li H, Terns RM, Terns MP (декабрь 2008 г.). «CAS6 - это эндорибонуклеаза, которая генерирует руководство RNA для защиты Invader в прокариотах» . Гены и развитие . 22 (24): 3489–3496. doi : 10.1101/gad.1742908 . PMC 2607076 . PMID 19141480 .
- ^ Wang R, Preamplume G, Terns MP, Terns RM, Li H (февраль 2011 г.). «Взаимодействие рибоннуклеазы CAS6 с CRISPR RNAS: признание и расщепление» . Структура 19 (2): 257–264. doi : 10.1016/j.str.2010.11.014 . PMC 3154685 . PMID 21300293 .
- ^ Niewoehner O, Jinek M, Doudna JA (январь 2014 г.). «Эволюция распознавания и обработки CRISPR РНК с помощью эндонуклеасов CAS6» . Исследование нуклеиновых кислот . 42 (2): 1341–1353. doi : 10.1093/nar/gkt922 . PMC 3902920 . PMID 24150936 .
- ^ Семенова Е., Джор М.М., Датсенко К.А., Семенова А., Вестра Эр, Ваннер Б. и др. (Июнь 2011 г.). «Вмешательство путем кластеризированной регулярной межсетенной РНК с коротким палиндромным повторением (CRISPR) определяется последовательности семян» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (25): 10098–10103. Bibcode : 2011pnas..10810098S . doi : 10.1073/pnas.1104144108 . PMC 3121866 . PMID 21646539 .
- ^ Gudbergsdottir S, Deng L, Chen Z, Jensen JV, Jensen LR, Hes Q, et al. (Январь 2011). «Динамические свойства систем Sulfolobus crispr/CAS и CRISPR/CMR, когда бросают вызов вирусным и плазмидным генам, переносимым вектором» . Молекулярная микробиология . 79 (1): 35–49. doi : 10.1111/j.1365-2958.2010.07452.x . PMC 3025118 . PMID 21166892 .
- ^ Маника А., Зебек З., Тейхманн Д., Шлепер С (апрель 2011 г.). «Активность in vivo опосредованной CRISPR защиты вируса в гипертермофильном археоне» . Молекулярная микробиология . 80 (2): 481–491. doi : 10.1111/j.1365-2958.2011.07586.x . PMID 21385233 . S2CID 41442419 .
- ^ Джор М.М., Лундгрен М., Ван Дуйдж Э., Бултема Дж. Б., Вестра Э.Р., Вагмаре С.П. и др. (Май 2011). «Структурная основа для распознавания ДНК-управляемой CRISPR РНК каскадом» (PDF) . Природа структурная и молекулярная биология . 18 (5): 529–536. doi : 10.1038/nsmb.2019 . PMID 21460843 . S2CID 10987554 . Архивировано (PDF) из оригинала на 2021-10-23 . Получено 2019-06-30 .
- ^ Wiedenheft B, Lander GC, Zhou K, Jore MM, Brouns SJ, Van Der Oost J, et al. (Сентябрь 2011). «Структуры комплекса наблюдения с РНК из бактериальной иммунной системы» . Природа . 477 (7365): 486–489. Bibcode : 2011natur.477..486w . doi : 10.1038/nature10402 . PMC 4165517 . PMID 21938068 .
- ^ Zhang J, Rouillon C, Kerou M, Reeks J, Brugger K, Graham S, et al. (Февраль 2012 г.). «Структура и механизм комплекса CMR для CRISPR-опосредованного противовирусного иммунитета» . Молекулярная клетка . 45 (3): 303–313. doi : 10.1016/j.molcel.2011.12.013 . PMC 3381847 . PMID 22227115 .
- ^ Jump up to: а беременный Hale CR, Zhao P, Olson S, Duff MO, Graveley BR, Wells L, et al. (Ноябрь 2009 г.). «РНК-управляемая РНК-расщепление с помощью белкового комплекса RNA-CAS CRISPR» . Клетка . 139 (5): 945–956. doi : 10.1016/j.cell.2009.07.040 . PMC 2951265 . PMID 19945378 .
- ^ Estrella MA, Kuo FT, Bailey S (2016). «РНК-активированное расщепление ДНК с помощью эффекторного комплекса типа III-B CRISPR-CAS» . Гены и развитие . 30 (4): 460–470. doi : 10.1101/gad.273722.115 . PMC 4762430 . PMID 26848046 .
- ^ Самаи П., Паенсон Н. , Цзян В., Голдберг Г.В., Хатум-Аслан А., Марраффини Л.А. (2015). «Ко-транскрипционная ДНК и РНК-расщепление во время иммунитета Crispr-CAS типа III» . Клетка . 161 (5): 1164–1174. doi : 10.1016/j.cell.2015.04.027 . PMC 4594840 . PMID 25959775 .
- ^ Jump up to: а беременный в Marraffini LA, Sontheimer EJ (январь 2010 г.). «Самостоятельно против не-я дискриминация во время иммунитета к РНК CRISPR» . Природа . 463 (7280): 568–571. Bibcode : 2010natur.463..568m . doi : 10.1038/nature08703 . PMC 2813891 . PMID 20072129 .
- ^ Krupovic M, Béguin P, Koonin EV (август 2017 г.). «Каспозоны: мобильные генетические элементы, которые породили механизм адаптации CRISPR-Cas» . Текущее мнение о микробиологии . 38 : 36–43. doi : 10.1016/j.mib.2017.04.004 . PMC 5665730 . PMID 28472712 .
- ^ Koonin EV , Makarova KS (май 2013 г.). «CRISPR-CAS: эволюция адаптивной системы иммунитета на основе РНК у прокариот» . РНК -биология . 10 (5): 679–686. doi : 10.4161/rna.24022 . PMC 3737325 . PMID 23439366 .
- ^ Koonin EV , Makarova KS, Zhang F (июнь 2017 г.). «Разнообразие, классификация и эволюция систем CRISPR-CAS» . Текущее мнение о микробиологии . 37 : 67–78. doi : 10.1016/j.mib.2017.05.008 . PMC 5776717 . PMID 28605718 .
- ^ Jump up to: а беременный Shmakov S, Smargon A, Scott D, Cox D, Pyzocha N, Yan W, et al. (Март 2017). «Разнообразие и эволюция систем CRISPR-CAS класса 2» . Природные обзоры. Микробиология . 15 (3): 169–182. doi : 10.1038/nrmicro.2016.184 . PMC 5851899 . PMID 28111461 .
- ^ Kupczok A, Bollback JP (февраль 2013 г.). «Вероятностные модели для эволюции контента Spacer CRISPR» . BMC Эволюционная биология . 13 (1): 54. Bibcode : 2013bmcee..13 ... 54K . doi : 10.1186/1471-2148-13-54 . PMC 3704272 . PMID 23442002 .
- ^ Sternberg SH, Richter H, Charpentier E, Qimron U (март 2016 г.). «Адаптация в системах CRISPR-CAS». Молекулярная клетка . 61 (6): 797–808. doi : 10.1016/j.molcel.2016.01.030 . HDL : 21.11116/0000-0003-E74E-2 . PMID 26949040 .
- ^ Koonin EV , Wolf Yi (ноябрь 2009 г.). "Эволюция дарвиниан или/и Ламарккиан?" Полем Биология прямой . 4 : 42. DOI : 10.1186/1745-6150-4-42 . PMC 2781790 . PMID 19906303 .
- ^ Вайс А (октябрь 2015 г.). «Ламаркские иллюзии» . Тенденции в экологии и эволюции . 30 (10): 566–568. Bibcode : 2015tecoe..30..566w . doi : 10.1016/j.tree.2015.08.003 . PMID 26411613 .
- ^ Jump up to: а беременный Koonin EV , Wolf Yi (февраль 2016 г.). «Как Ламарккиан-иммунитет CRISPR-CAS: континуум механизмов эволюции» . Биология прямой . 11 (1): 9. doi : 10.1186/s13062-016-0111-z . PMC 4765028 . PMID 26912144 .
- ^ Heidelberg JF, Nelson WC, Schoenfeld T, Bhaya D (2009). Ахмед Н. (ред.). «Заращивание в сообществе микробных матов: CRISPR дают представление о совместной эволюции хозяина и вирусных геномов» . Plos один . 4 (1): E4169. Bibcode : 2009ploso ... 4.4169h . doi : 10.1371/journal.pone.0004169 . PMC 2612747 . PMID 19132092 .
- ^ Touckon M, Rocha EP (июнь 2010 г.). Рандау Л (ред.). «Маленькая, медленная и специализированная CRISPR и антикриспр Escherichia и Salmonella» . Plos один . 5 (6): E11126. BIBCODE : 2010PLOSO ... 511126T . doi : 10.1371/journal.pone.0011126 . PMC 2886076 . PMID 20559554 .
- ^ Jump up to: а беременный в Rho M, Wu YW, Tang H, Doak TG, Ye Y (2012). «Разнообразные CRISPR, развивающиеся в микробиомах человека» . PLOS Genetics . 8 (6): E1002441. doi : 10.1371/journal.pgen.1002441 . PMC 3374615 . PMID 22719260 .
- ^ Jump up to: а беременный Sun CL, Barrangou R , Thomas BC, Horvath P, Fremaux C, Banfield JF (февраль 2013 г.). «Мутации фага в ответ на диверсификацию CRISPR в бактериальной популяции». Экологическая микробиология . 15 (2): 463–470. Bibcode : 2013envmi..15..463s . doi : 10.1111/j.1462-2920.2012.02879.x . PMID 23057534 .
- ^ Kuno S, Sako Y, Yoshida T (май 2014). «Диверсификация CRISPR в сосуществующих генотипах в естественной популяции цветущей формирующей цветение цианобактерий microcystis aeruginosa» . Микробиология . 160 (Pt 5): 903–916. doi : 10.1099/mic.0.073494-0 . PMID 24586036 .
- ^ Sorek R, Kunin V, Hugenholtz P (март 2008 г.). «CRISPR - широко распространенная система, которая обеспечивает приобретенную устойчивость к фагам в бактериях и археи» . Природные обзоры. Микробиология . 6 (3): 181–186. doi : 10.1038/nrmicro1793 . PMID 18157154 . S2CID 3538077 . Архивировано из оригинала 2021-03-08 . Получено 2019-11-25 .
Таблица 1: Веб -ресурсы для анализа CRISPR
- ^ Pride DT, Salzman J, Relman DA (сентябрь 2012 г.). «Сравнение кластеризированных регулярно межпрессовых коротких палиндромических повторений и вирусов в слюне человека показывают бактериальную адаптацию к вирусам слюны» . Экологическая микробиология . 14 (9): 2564–2576. Bibcode : 2012envmi..14.2564p . doi : 10.1111/j.1462-2920.2012.02775.x . PMC 3424356 . PMID 22583485 .
- ^ Helld NL, Herrera A, Whitaker RJ (ноябрь 2013 г.). «Переворачивание локусов CRISPR повторного пространства в Sulfolobus Islandicus». Экологическая микробиология . 15 (11): 3065–3076. Bibcode : 2013envmi..15.3065H . doi : 10.1111/1462-2920.12146 . PMID 23701169 .
- ^ Hell NL, Herrera A, Cadillo-Quiroz H, Whitaker RJ (сентябрь 2010 г.). «CRISPR связано с разнообразием в популяции Sulfolobus Islandicus» . Plos один . 5 (9): E12988. Bibcode : 2010ploso ... 512988H . doi : 10.1371/journal.pone.0012988 . PMC 2946923 . PMID 20927396 .
- ^ Jump up to: а беременный в Медведева С., Лю Ю., Кунин Э.В., Северинов К., Пранджшвили Д., Крупович М (ноябрь 2019). «Вирусные массивы Mini-Crispr участвуют в интервенционных конфликтах» . Природная связь . 10 (1): 5204. Bibcode : 2019natco..10.5204M . doi : 10.1038/s41467-019-13205-2 . PMC 6858448 . PMID 31729390 .
- ^ Skennerton CT, Imelfort M, Tyson GW (май 2013). «Crass: идентификация и реконструкция CRISPR из незаменимых метагеномных данных» . Исследование нуклеиновых кислот . 41 (10): E105. doi : 10.1093/nar/gkt183 . PMC 3664793 . PMID 23511966 .
- ^ Стерн А., Мик Э., Тирош I, Саги О., Сорек Р (октябрь 2012 г.). «Целью CRISPR выявляет резервуар общих фагов, связанных с микробиомом кишечника человека» . Исследование генома . 22 (10): 1985–1994. doi : 10.1101/gr.138297.112 . PMC 3460193 . PMID 22732228 .
- ^ Novick RP, Christie Ge, Penadés Jr (август 2010 г.). «Связанные с фагом хромосомные острова грамположительные бактерии» . Nature Reviews Microbiology . 8 (8): 541–551. doi : 10.1038/nrmicro2393 . PMC 3522866 . PMID 20634809 .
- ^ Ram G, Chen J, Kumar K, Ross HF, Ubeda C, Damle PK, et al. (Октябрь 2012 г.). «Вмешательство острова стафилококковой патогенности с воспроизводством вспомогательного фага является парадигмой молекулярного паразитизма» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (40): 16300–16305. BIBCODE : 2012PNAS..10916300R . doi : 10.1073/pnas.1204615109 . PMC 3479557 . PMID 22991467 .
- ^ Рам Г., Чен Дж., Росс Х.Ф., Новик Р.П. (октябрь 2014 г.). «Точно модулированные интерференции острова патогенности с транскрипцией позднего фага» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (40): 14536–14541. Bibcode : 2014pnas..11114536R . doi : 10.1073/pnas.1406749111 . PMC 4209980 . PMID 25246539 .
- ^ SEED KD, Lazinski DW, Calderwood SB, Camilli A (февраль 2013 г.). «Бактериофаг кодирует свой собственный адаптивный ответ CRISPR/CAS на уклонение от врожденного иммунитета хозяина» . Природа . 494 (7438): 489–491. Bibcode : 2013natur.494..489s . doi : 10.1038/nature11927 . PMC 3587790 . PMID 23446421 .
- ^ Boyd CM, Angermeyer A, Hays SG, Barth ZK, Patel KM, Seed KD (сентябрь 2021 г.). «Бактериофаг ICP1: постоянный хищник вибрао -холеры » . Ежегодный обзор вирусологии . 8 (1): 285–304. doi : 10.1146/annurev-virology-091919-072020 . ISSN 2327-056X . PMC 9040626 . PMID 34314595 .
- ^ Feng Q, Li Q, Zhou H, Wang Z, Lin C, Jiang Z, et al. (Август 2024). «Технология CRISPR при заболеваниях человека» . Medcomm . 5 (8): E672. doi : 10.1002/mco2.672 . PMC 11286548 . PMID 39081515 .
- ^ Li T, Li S, Kang Y, Zhou J, Yi M (август 2024 г.). «Использование развивающегося CRISPR/CAS9 для точной онкологии» . Журнал трансляционной медицины . 22 (1): 749. DOI : 10.1186/S12967-024-05570-4 . PMC 11312220 . PMID 39118151 .
- ^ Мишра С., Наяк С., Тутея Н., Пусапати С., Суэйн Д.М., Саху Р.К. (июль 2024 г.). «CRISPR/CAS-опосредованная генома инженерия на растениях: применение и перспективы» . Растения 13 (14). Базель, Швейцария: 1884. doi : 10.3390/plants13141884 . PMC 11279650 . PMID 39065411 .
- ^ Каур Р., Гупта С., Чаухан А., Мишра В., Шарма М.К., Сингх Дж (август 2024 г.). «Использование мощности кластеризированных регулярных межсепенсорных микрофлюидиков на основе коротких палиндромических повторений (CRISPR) для молекулярной диагностики следующего поколения». Молекулярная биология отчетов . 51 (1): 896. DOI : 10.1007/S11033-024-09840-8 . PMID 39115550 .
- ^ Shinwari ZK, Tanveer F, Khalil AT (2018). «Этические проблемы, касающиеся редактирования генома CRISPR» . Современные проблемы в молекулярной биологии . 26 : 103–110. doi : 10.21775/cimb.026.103 . PMID 28879860 .
- ^ Kozan DW, Farber SA (февраль 2024 г.). «Неужели разумно редактировать аллели дикого типа? Инженерные аллели CRISPR против миллионов лет эволюции человека». Артериосклероз, тромбоз и сосудистая биология . 44 (2): 328–333. doi : 10.1161/atvbaha.123.318069 . PMC 10948015. PMID 38059350 .
Дальнейшее чтение
[ редактировать ]- Doudna J , Mali P (23 марта 2016 г.). CRISPR-CAS: лабораторное руководство . Нью -Йорк: Cold Spring Harbor Laboratory Press. ISBN 978-1-62182-131-1 .
- Моханраджу П., Макарова К.С., Зцше Б., Чжан Ф. , Кунин Э.В. , Ван дер Оост Дж (август 2016 г.). «Разнообразные эволюционные корни и механистические изменения систем CRISPR-CAS» (PDF) . Наука . 353 (6299): AAD5147. doi : 10.1126/science.aad5147 . HDL : 1721.1/113195 . PMID 27493190 . S2CID 11086282 . Архивировано из оригинала 2021-10-23 . Получено 2019-08-29 .
- Сандер JD, Joung JK (апрель 2014 г.). «Системы CRISPR-CAS для редактирования, регулирования и нацеливания геномов» . Nature Biotechnology . 32 (4): 347–355. doi : 10.1038/nbt.2842 . PMC 4022601 . PMID 24584096 .
- Slaymaker IM, Gao L, Zetsche B, Scott DA, Yan WX, Zhang F (январь 2016 г.). «Рационально спроектированные нуклеазы CAS9 с улучшенной специфичностью» . Наука . 351 (6268): 84–88. Bibcode : 2016sci ... 351 ... 84 с . doi : 10.1126/science.aad5227 . PMC 4714946 . PMID 26628643 .
- Terns RM, Terns MP (март 2014 г.). «Технологии на основе CRISPR: прокариотическое оборонное оружие перепрофилировано» . Тенденции в генетике . 30 (3): 111–118. doi : 10.1016/j.tig.2014.01.003 . PMC 3981743 . PMID 24555991 .
- Westra Er, Buckling A, Fineran PC (май 2014 г.). «Системы CRISPR-CAS: за пределами адаптивного иммунитета». Nature Reviews Microbiology . 12 (5): 317–326. doi : 10.1038/nrmicro3241 . PMID 24704746 . S2CID 36575361 .
- Andersson AF, Banfield JF (май 2008 г.). «Динамика популяции вирусов и приобретенная устойчивость к вирусам в природных микробных сообществах». Наука . 320 (5879): 1047–1050. Bibcode : 2008Sci ... 320.1047a . doi : 10.1126/science.1157358 . PMID 18497291 . S2CID 26209623 .
- Hale C, Kleppe K, Terns RM, Terns MP (декабрь 2008 г.). «Прокариотическое молчание (PSI) РНК в пирококковом фуриосе» . РНК . 14 (12): 2572–2579. doi : 10.1261/rna.1246808 . PMC 2590957 . PMID 18971321 .
- Ван дер Плоэг -младший (июнь 2009 г.). «Анализ CRISPR у Streptococcus mutans предполагает частое появление приобретенного иммунитета против инфекции с помощью M102-подобных бактериофагов» (PDF) . Микробиология . 155 (Pt 6): 1966–1976. doi : 10.1099/mic.0.027508-0 . PMID 19383692 .
- Van der Oost J, Brouns SJ (ноябрь 2009 г.). «РНКИ: Прокариоты попадают в действие» . Клетка . 139 (5): 863–865. doi : 10.1016/j.cell.2009.11.018 . PMID 19945373 . S2CID 11863610 .
- Karginov FV, Hannon GJ (январь 2010 г.). «Система CRISPR: небольшая защита РНК в бактериях и археи» . Молекулярная клетка . 37 (1): 7–19. doi : 10.1016/j.molcel.2009.12.033 . PMC 2819186 . PMID 20129051 .
- Pul U, Wurm R, Arslan Z, Geissen R, Hofmann N, Wagner R (март 2010 г.). «Идентификация и характеристика промоторов E. coli crispr-cas и их молчание H-NS» . Молекулярная микробиология . 75 (6): 1495–1512. doi : 10.1111/j.1365-2958.2010.07073.x . PMID 20132443 . S2CID 37529215 .
- Díez-Villaseñor C, Almendros C, García-Martínez J, Mojica FJ (май 2010). «Разнообразие локусов CRISPR в Escherichia coli » . Микробиология . 156 (Pt 5): 1351–1361. Doi : 10.1099/mic.0.036046-0 . PMID 20133361 .
- Deveau H, Garneau JE, Moineau S (2010). «Система CRISPR/CAS и ее роль во взаимодействиях фага-бактерий». Ежегодный обзор микробиологии . 64 : 475–493. doi : 10.1146/annurev.micro.112408.134123 . PMID 20528693 .
- Koonin EV , Makarova KS (декабрь 2009 г.). «CRISPR-CAS: адаптивная система иммунитета у прокариот» . Отчеты о биологии F1000 . 1 : 95. doi : 10.3410/b1-95 . PMC 2884157 . PMID 20556198 .
- «Возраст красной ручки» . Экономист . 22 августа 2015 года. ISSN 0013-0613 . Архивировано с оригинала 2015-08-24 . Получено 2015-08-25 .
- Ran AF, HSU PD, Wright J, Agarwala V, Scott Da, Zhang F (2013). «Инженер генома с использованием системы CRISPR-CAS9» . Природные протоколы . 8 (11): 2281–2308. doi : 10.1038/nprot.2013.143 . PMC 3969860 . PMID 24157548 .
Внешние ссылки
[ редактировать ]- «Расширенное редактирование генов: CRISPR-CAS9» (PDF) . Исследовательская служба Конгресса .
- «Дженнифер Дудна Говорит: Инженер генома с CRISPR-CAS9: Рождение технологии прорыва» . 10 сентября 2022 года.
- «Человеческая природа» . Новая Сезон 47. Эпизод 9. 9 сентября 2020 года. PBS . WGBH . Получено 7 апреля 2023 года .
Банк белка
[ редактировать ]- Обзор всей структурной информации, доступной в PDB для Uniprot : Q46901 (каскадная субъединица CRISPR System Casa) в PDBE-KB .
- Обзор всей структурной информации, доступной в PDB для Uniprot : P76632 (каскадная субъединица CRISPR System CASB) в PDBE-KB .
- Обзор всей структурной информации, доступной в PDB для Uniprot : Q46899 (CRISPR System Cascade Subunit CASC) в PDBE-KB .
- Обзор всей структурной информации, доступной в PDB для Uniprot : Q46898 (каскадная субъединица CRISPR System CASD) в PDBE-KB .
- Обзор всей структурной информации, доступной в PDB для Uniprot : Q46897 (корпус субъединицы системы CRISPR) в PDBE-KB .