КРИСПР
| Каскад (CRISPR-ассоциированный комплекс противовирусной защиты) | |||
|---|---|---|---|
 Белок CRISPR Cascade (голубой), связанный с РНК CRISPR (зеленый) и ДНК фага (красный) [ 1 ] | |||
| Идентификаторы | |||
| Организм | |||
| Символ | КРИСПР | ||
| Входить | 947229 | ||
| ПДБ | 4QYZ | ||
| RefSeq (защита) | НП_417241.1 | ||
| ЮниПрот | P38036 | ||
| |||
| Часть серии о |
| КРИСПР |
|---|
| Редактирование генома : CRISPR-Cas |
|
варианты: Анти-CRISPR - CIRTS - CRISPeYCRISPR-Cas10 - CRISPR-Cas13 - CRISPR-BEST CRISPR-Disp - CRISPR-Gold - CRISPRa - CRISPRi Easi-CRISPR – СДЕЛАЙТЕ ЭТО |
| Фермент |
|
Cas9 - FokI - EcoRI - PstI - SmaI HaeIII – Cas12a (Cpf1) – xCas9 |
| Приложения |
| КАМЕРА - ICE - Направленная генетика |
| другой метод редактирования генома: |
| Прайм-редактирование - Pro-AG - RESCUE - TALEN - ZFN - LEAPER |
| Часть серии о |
| Генная инженерия |
|---|
| Генетически модифицированные организмы |
| История и регулирование |
| Процесс |
| Приложения |
| Споры |
CRISPR ( / ˈ k r ɪ s p ər / ) ( аббревиатура от кластеризованных , коротких регулярно расположенных между собой палиндромных повторов ) таких , — семейство последовательностей ДНК обнаруженных в организмов , геномах прокариотических как бактерии и археи . [ 2 ] Эти последовательности получены из фрагментов ДНК бактериофагов , ранее заразивших прокариот. Их используют для обнаружения и уничтожения ДНК подобных бактериофагов при последующих инфекциях. Следовательно, эти последовательности играют ключевую роль в противовирусной (т.е. антифаговой ) системе защиты прокариот и обеспечивают форму приобретенного иммунитета . [ 2 ] [ 3 ] [ 4 ] [ 5 ] CRISPR обнаружен примерно в 50% секвенированных геномов бактерий и почти в 90% секвенированных архей. [ 6 ]

Cas9 (или «CRISPR-ассоциированный белок 9») — это фермент , который использует последовательности CRISPR в качестве руководства для распознавания и открытия определенных цепей ДНК, которые комплементарны последовательности CRISPR. Ферменты Cas9 вместе с последовательностями CRISPR составляют основу технологии, известной как CRISPR-Cas9 , которую можно использовать для редактирования генов в живых организмах. [ 8 ] [ 9 ] Этот процесс редактирования имеет широкий спектр применений, включая фундаментальные биологические исследования, разработку биотехнологических продуктов и лечение заболеваний. [ 10 ] [ 11 ] Разработка метода редактирования генома CRISPR-Cas9 была отмечена Нобелевской премией по химии 2020 года, присужденной Эммануэль Шарпантье и Дженнифер Дудна . [ 12 ] [ 13 ]
История
[ редактировать ]Повторяющиеся последовательности
[ редактировать ]Открытие кластерных повторов ДНК произошло независимо в трех частях света. Первое описание того, что позже будет названо CRISPR, принадлежит из Университета Осаки исследователю Ёсидзуми Исино и его коллегам в 1987 году. Они случайно клонировали часть последовательности CRISPR вместе с геном «iap» (изоферментная конверсия щелочной фосфатазы) из генома эшерихий. палочка [ 14 ] [ 15 ] что было их целью. Организация повторов была необычной. Повторяющиеся последовательности обычно располагаются последовательно, без перемежения разных последовательностей. [ 11 ] [ 15 ] Они не знали функции прерванных кластерных повторов.
В 1993 году исследователи микобактерии туберкулеза в Нидерландах опубликовали две статьи о кластере прерванных прямых повторов (DR) у этой бактерии. Они признали разнообразие последовательностей, которые участвуют в прямых повторах среди разных штаммов M. Tuberculosis. [ 16 ] и использовал это свойство для разработки метода набора текста под названием сполиготипирование , который используется до сих пор. [ 17 ] [ 18 ]
Франсиско Мохика из Университета Аликанте в Испании изучал функцию повторов у архейных видов Haloferax и Haloarcula . Руководитель Мохики предположил, что кластерные повторы играют роль в правильном разделении реплицируемой ДНК на дочерние клетки во время клеточного деления, поскольку плазмиды и хромосомы с идентичными массивами повторов не могли сосуществовать в Haloferax volcanii . Транскрипция прерванных повторов также отмечена впервые; это была первая полная характеристика CRISPR. [ 18 ] [ 19 ] К 2000 году Мохика и его ученики после автоматического поиска опубликованных геномов идентифицировали прерывистые повторы у 20 видов микробов как принадлежащие к одному и тому же семейству. [ 20 ] Поскольку эти последовательности располагались через промежутки, Мохика первоначально назвал эти последовательности «короткими повторами с регулярными интервалами» (SRSR). [ 21 ] В 2001 году Мохика и Рууд Янсен , которые искали дополнительные прерванные повторы, предложили аббревиатуру CRISPR (Clustered Regularly Interspaced Short Palindromic Повторы), чтобы унифицировать многочисленные акронимы, используемые для описания этих последовательностей. [ 19 ] [ 22 ] В 2002 году Тан и др. продемонстрировали доказательства того, что области повторов CRISPR из генома Archaeoglobus fulgidus транскрибируются в длинные молекулы РНК, которые впоследствии обрабатываются в небольшие РНК единичной длины, а также в некоторые более длинные формы из 2, 3 или более единиц спейсерных повторов. [ 23 ] [ 24 ]
В 2005 году йогурта исследователь Родольф Баррангу обнаружил, что Streptococcus thermophilus после повторяющихся фаговых заражений развивает повышенную устойчивость к фагам из-за включения дополнительных спейсерных последовательностей CRISPR. [ 25 ] Работодатель Баррангу, датская пищевая компания Danisco, затем разработала устойчивые к фагам штаммы S. thermophilus для производства йогурта. Позднее Danisco была куплена компанией DuPont , которой принадлежит около 50 процентов мирового рынка молочных культур, и технология широко распространилась. [ 26 ]
Системы, связанные с CRISPR
[ редактировать ]Большим прорывом в понимании CRISPR стало наблюдение Янсена о том, что кластер повторов прокариот сопровождается четырьмя гомологичными генами, которые составляют CRISPR-ассоциированные системы, cas 1–4. Белки Cas обнаруживают геликазные и нуклеазные мотивы , что указывает на их роль в динамической структуре локусов CRISPR. [ 27 ] В этой публикации в качестве универсального названия этого паттерна использовалась аббревиатура CRISPR, но его функция осталась загадочной.

В 2005 году три независимые исследовательские группы показали, что некоторые спейсеры CRISPR происходят из фаговой ДНК и внехромосомной ДНК, такой как плазмиды . [ 31 ] [ 32 ] [ 33 ] По сути, спейсеры представляют собой фрагменты ДНК, полученные от вирусов, ранее атаковавших клетку. Источник спейсеров был признаком того, что система CRISPRcas может играть роль в адаптивном иммунитете бактерий . [ 28 ] [ 34 ] Все три исследования, предлагающие эту идею, первоначально были отвергнуты известными журналами, но со временем появились в других журналах. [ 35 ]
Первая публикация [ 32 ] предложив роль CRISPR-Cas в микробном иммунитете, Мохика и его коллеги из Университета Аликанте предсказали роль РНК-транскрипта спейсеров в распознавании мишени в механизме, который может быть аналогичен системе РНК-интерференции, используемой эукариотическими клетками. Кунин и его коллеги расширили эту гипотезу РНК-интерференции, предложив механизмы действия различных подтипов CRISPR-Cas в соответствии с предсказанной функцией их белков. [ 36 ]
Экспериментальная работа нескольких групп выявила основные механизмы иммунитета CRISPR-Cas. В 2007 году были опубликованы первые экспериментальные доказательства того, что CRISPR представляет собой адаптивную иммунную систему. [ 4 ] [ 11 ] Область CRISPR у Streptococcus thermophilus приобрела спейсеры из ДНК инфицирующего бактериофага . Исследователи манипулировали устойчивостью S. thermophilus к различным типам фагов, добавляя и удаляя спейсеры, последовательность которых соответствовала обнаруженной в тестируемых фагах. [ 37 ] [ 38 ] В 2008 году Браунс и Ван дер Ост идентифицировали комплекс белков Cas, названный Cascade, который в E. coli разрезал предшественник РНК CRISPR внутри повторов на зрелые молекулы РНК, содержащие спейсер, называемые CRISPR РНК (crRNA), которые оставались связанными с белком. сложный. [ 39 ] Более того, было обнаружено, что каскад, crRNA и хеликаза/нуклеаза ( Cas3 ) необходимы для обеспечения бактериального хозяина иммунитета против заражения ДНК-вирусом . Разработав антивирусный CRISPR, они продемонстрировали, что две ориентации crRNA (смысловая/антисмысловая) обеспечивают иммунитет, что указывает на то, что направляющие crRNA нацелены на дцДНК . В том же году Марраффини и Сонтхаймер подтвердили, что последовательность CRISPR S. epidermidis нацелена на ДНК, а не на РНК, чтобы предотвратить конъюгацию . Это открытие противоречило предложенному механизму иммунитета CRISPR-Cas, подобному РНК-интерференции, хотя система CRISPR-Cas, нацеленная на чужеродную РНК, была позже обнаружена у Pyrococcus Furiosus . [ 11 ] [ 38 ] Исследование 2010 года показало, что CRISPR-Cas разрезает нити как фаговой, так и плазмидной ДНК у S. thermophilus . [ 40 ]
Красный9
[ редактировать ]Более простая система CRISPR Streptococcus pyogenes основана на белке Cas9 . Cas9 Эндонуклеаза представляет собой четырехкомпонентную систему, включающую две небольшие молекулы: crRNA и трансактивирующую РНК CRISPR (tracrRNA). [ 41 ] [ 42 ] В 2012 году Дженнифер Дудна и Эммануэль Шарпантье модернизировали эндонуклеазу Cas9 в более управляемую двухкомпонентную систему, объединив две молекулы РНК в « РНК с одним проводником », которая в сочетании с Cas9 могла найти и разрезать ДНК-мишень. указанной направляющей РНК. [ 43 ] Этот вклад был настолько значительным, что он был отмечен Нобелевской премией по химии в 2020 году. Манипулируя нуклеотидной последовательностью направляющей РНК, искусственную систему Cas9 можно было запрограммировать на нацеливание на любую последовательность ДНК для разделения. [ 43 ] Другое сотрудничество, включающее Вирджиниюса Шикшниса , Гасюнаса, Баррангу и Хорвата, показало, что Cas9 из системы CRISPR S. thermophilus также можно перепрограммировать для нацеливания на выбранный ими сайт путем изменения последовательности его crRNA. Эти достижения стимулировали усилия по редактированию геномов с помощью модифицированной системы CRISPR-Cas9. [ 18 ]
Группы под руководством Фэн Чжана и Джорджа Черча одновременно впервые опубликовали описания редактирования генома в культурах клеток человека с использованием CRISPR-Cas9. [ 11 ] [ 44 ] [ 45 ] С тех пор он использовался для лечения широкого спектра микроорганизмов, включая пекарские дрожжи ( Saccharomyces cerevisiae ), [ 46 ] [ 47 ] [ 48 ] Candida условно-патогенный микроорганизм albicans , [ 49 ] [ 50 ] рыбка данио ( Danio rerio ), [ 51 ] плодовые мушки ( Drosophila melanogaster ), [ 52 ] [ 53 ] муравьи ( Harpegnathos saltator [ 54 ] и Ooceraea biroi [ 55 ] ), комары ( Aedes aegypti [ 56 ] ), нематоды ( Caenorhabditis elegans ), [ 57 ] растения, [ 58 ] мыши ( Mus musculus Domesticus ) , [ 59 ] [ 60 ] обезьяны [ 61 ] и человеческие эмбрионы. [ 62 ]
CRISPR был модифицирован для создания программируемых факторов транскрипции , которые позволяют активировать или подавлять целевые гены. [ 63 ]

Было показано, что система CRISPR-Cas9 эффективно редактирует гены в трехпронуклеарных зиготах человека , как впервые описано в статье 2015 года китайских ученых П. Ляна и Ю. Сюй. Система успешно расщепила мутантный бета-гемоглобин ( HBB ) в 28 из 54 эмбрионов. Четыре из 28 эмбрионов были успешно рекомбинированы с использованием донорской матрицы. Ученые показали, что во время рекомбинации ДНК расщепленной цепи гомологичная эндогенная последовательность HBD конкурирует с экзогенной донорской матрицей. Репарация ДНК в эмбрионах человека гораздо сложнее и специфичнее, чем в производных стволовых клетках. [ 64 ]
Кас12а
[ редактировать ]В 2015 году нуклеаза Cas12a (ранее называвшаяся Cpf1 [ 65 ] ) был охарактеризован в системе CRISPR-Cpf1 бактерии Francesella novicida . [ 66 ] [ 67 ] Его первоначальное название, взятое из TIGRFAMs, определения семейства белков созданного в 2012 году, отражает распространенность его подтипа CRISPR-Cas в линиях Prevotella и Francesella . Cas12a продемонстрировал несколько ключевых отличий от Cas9, в том числе: создание «шахматного» разреза двухцепочечной ДНК в отличие от «тупого» разреза, производимого Cas9, с использованием «T-богатого» PAM (обеспечивающего альтернативные сайты нацеливания для Cas9) и необходимости только РНК CRISPR (crRNA) для успешного нацеливания. Напротив, Cas9 требует как crRNA, так и трансактивирующей crRNA (tracrRNA).
Эти различия могут дать Cas12a некоторые преимущества перед Cas9. Например, небольшие crRNA Cas12a идеально подходят для мультиплексного редактирования генома, поскольку в один вектор можно упаковать больше из них, чем sgRNA Cas9. Липкие 5'-концы, оставленные Cas12a, также можно использовать для сборки ДНК, которая гораздо более специфична для мишени, чем традиционное клонирование ферментов рестрикции. [ 68 ] Наконец, Cas12a расщепляет ДНК на 18–23 пары оснований ниже сайта PAM. Это означает, что после репарации не происходит нарушения последовательности узнавания, и поэтому Cas12a обеспечивает несколько циклов расщепления ДНК. Напротив, поскольку Cas9 разрезает только 3 пары оснований выше сайта PAM, путь NHEJ приводит к индел -мутациям, которые разрушают последовательность узнавания, тем самым предотвращая дальнейшие циклы разрезания. Теоретически, повторные циклы расщепления ДНК должны увеличить вероятность желаемого редактирования генома. [ 69 ] Отличительной особенностью Cas12a по сравнению с Cas9 является то, что после разрезания своей мишени Cas12a остается связанным с мишенью и затем неизбирательно расщепляет другие молекулы оцДНК. [ 70 ] Это свойство называется активностью «побочного расщепления» или «транс-расщепления» и использовалось для разработки различных диагностических технологий. [ 71 ] [ 72 ]
Кас13
[ редактировать ]В 2016 году нуклеаза Cas13a (ранее известный как C2c2 ) из бактерии Leptotrichia shahii . Cas13 представляет собой РНК-ориентированную РНК-эндонуклеазу, что означает, что она не расщепляет ДНК, а только одноцепочечную РНК. Cas13 направляется с помощью crRNA к мишени оцРНК, связывает и расщепляет мишень. Подобно Cas12a, Cas13 остается связанным с мишенью, а затем без разбора расщепляет другие молекулы оцРНК. [ 73 ] Это побочное свойство расщепления было использовано для разработки различных диагностических технологий. [ 74 ] [ 75 ] [ 76 ]
В 2021 году доктор Хуэй Ян охарактеризовал новые миниатюрные варианты белка Cas13 (mCas13): Cas13X и Cas13Y. Использование небольшой части последовательности гена N SARS-CoV-2 в качестве мишени для характеристики mCas13 выявило чувствительность и специфичность mCas13 в сочетании с RT-LAMP для обнаружения SARS-CoV-2 как в синтетических, так и в клинических образцах по сравнению с другими доступные стандартные тесты, такие как RT-qPCR (1 копия/мкл). [ 77 ]
Структура локуса
[ редактировать ]Повторы и проставки
[ редактировать ]Массив CRISPR состоит из богатой АТ лидерной последовательности, за которой следуют короткие повторы, разделенные уникальными спейсерами. [ 78 ] Повторы CRISPR обычно имеют размер от 28 до 37 пар оснований (п.н.), хотя их длина может составлять от 23 до 55 п.н. [ 79 ] Некоторые демонстрируют диадную симметрию , подразумевающую образование вторичной структуры, такой как стебель-петля («шпилька») в РНК, в то время как другие спроектированы так, чтобы быть неструктурированными. Размер спейсеров в разных массивах CRISPR обычно составляет от 32 до 38 п.н. (диапазон от 21 до 72 п.н.). [ 79 ] Новые спейсеры могут быстро появляться как часть иммунного ответа на фаговую инфекцию. [ 80 ] Обычно в массиве CRISPR содержится менее 50 единиц последовательности повтор-спейсер. [ 79 ]
Структуры CRISPR РНК
[ редактировать ]-
 CRISPR-DR2: Вторичная структура взята из базы данных Rfam . Семья RF01315 .
CRISPR-DR2: Вторичная структура взята из базы данных Rfam . Семья RF01315 . -
 CRISPR-DR5: Вторичная структура взята из базы данных Rfam . Семья RF011318 .
CRISPR-DR5: Вторичная структура взята из базы данных Rfam . Семья RF011318 . -
 CRISPR-DR6: Вторичная структура взята из базы данных Rfam . Семья RF01319 .
CRISPR-DR6: Вторичная структура взята из базы данных Rfam . Семья RF01319 . -
 CRISPR-DR8: Вторичная структура взята из базы данных Rfam . Семья RF01321 .
CRISPR-DR8: Вторичная структура взята из базы данных Rfam . Семья RF01321 . -
 CRISPR-DR9: Вторичная структура взята из базы данных Rfam . Семья RF01322 .
CRISPR-DR9: Вторичная структура взята из базы данных Rfam . Семья RF01322 . -
 CRISPR-DR19: Вторичная структура взята из базы данных Rfam . Семья RF01332 .
CRISPR-DR19: Вторичная структура взята из базы данных Rfam . Семья RF01332 . -
 CRISPR-DR41: Вторичная структура взята из базы данных Rfam . Семья RF01350 .
CRISPR-DR41: Вторичная структура взята из базы данных Rfam . Семья RF01350 . -
 CRISPR-DR52: Вторичная структура взята из базы данных Rfam . Семья RF01365 .
CRISPR-DR52: Вторичная структура взята из базы данных Rfam . Семья RF01365 . -
 CRISPR-DR57: Вторичная структура взята из базы данных Rfam . Семья RF01370 .
CRISPR-DR57: Вторичная структура взята из базы данных Rfam . Семья RF01370 . -
 CRISPR-DR65: Вторичная структура взята из базы данных Rfam . Семья RF01378 .
CRISPR-DR65: Вторичная структура взята из базы данных Rfam . Семья RF01378 .










Гены Cas и подтипы CRISPR
[ редактировать ]Небольшие кластеры генов cas часто располагаются рядом с массивами повторов и спейсеров CRISPR. В совокупности 93 гена cas сгруппированы в 35 семейств на основе сходства последовательностей кодируемых белков. 11 из 35 семейств образуют ядро cas , в которое входят семейства белков от Cas1 до Cas9. Полный локус CRISPR-Cas имеет по крайней мере один ген, принадлежащий ядру cas . [ 81 ]
Системы CRISPR-Cas делятся на два класса. Системы класса 1 используют комплекс из нескольких белков Cas для расщепления чужеродных нуклеиновых кислот. Системы класса 2 используют для той же цели один большой белок Cas. Класс 1 делится на типы I, III и IV; 2-й класс делится на типы II, V и VI. [ 82 ] 6 типов систем разделены на 33 подтипа. [ 83 ] Каждый тип и большинство подтипов характеризуются «сигнатурным геном», встречающимся почти исключительно в этой категории. Классификация также основана на составе cas присутствующих генов . Большинство систем CRISPR-Cas содержат белок Cas1. Филогения белков Cas1 в целом соответствует принятой классификационной системе. [ 84 ] но существуют исключения из-за перетасовки модулей. [ 81 ] Многие организмы содержат несколько систем CRISPR-Cas, что позволяет предположить, что они совместимы и могут иметь общие компоненты. [ 85 ] [ 86 ] Спорадическое распространение подтипов CRISPR-Cas позволяет предположить, что система CRISPR-Cas подвержена горизонтальному переносу генов в ходе микробной эволюции .
В этой таблице отсутствует информация о перекрестных ссылках UniProt и InterPro. ( октябрь 2020 г. ) |
| Сорт | Тип Каса | Подтип Cas | Сигнатурный белок | Функция | Ссылка |
|---|---|---|---|---|---|
| 1 | я | — | Кас3 | Одноцепочечная ДНК-нуклеаза (домен HD) и АТФ-зависимая геликаза | [ 87 ] [ 88 ] |
| Я | Кас8а, Кас5 | Cas8 представляет собой субъединицу модуля интерференции, которая важна для нацеливания на вторгающуюся ДНК путем распознавания последовательности PAM . Cas5 необходим для процессинга и стабильности crRNA. | [ 84 ] [ 89 ] | ||
| ИБ | Cas8b | ||||
| IC | Cas8c | ||||
| ИДЕНТИФИКАТОР | Кас10д | содержит домен, гомологичный пальмовому домену полимераз нуклеиновых кислот и нуклеотидциклаз | [ 90 ] [ 91 ] | ||
| ИЕ | Ксе1, Ксе2 | ||||
| ЕСЛИ | Csy1, Csy2, Csy3 | Тип IF-3 участвует в транспозонах, связанных с CRISPR. | [ 84 ] | ||
| ИГ [ Примечание 1 ] | GSU0054 | [ 92 ] | |||
| III | — | Кас10 | Гомолог Cas10d и Cse1. Связывает целевую РНК CRISPR и способствует стабильности интерференционного комплекса. | [ 91 ] [ 93 ] | |
| III-А | КСМ2 | Не определено | [ 84 ] | ||
| III-Б | CMR5 | Не определено | [ 84 ] | ||
| III-С | Кас10 или Csx11 | [ 84 ] [ 93 ] | |||
| III-D | Csx10 | [ 84 ] | |||
| III-E | [ 92 ] | ||||
| III-F | [ 92 ] | ||||
| IV | — | Csf1 | [ 92 ] | ||
| IV-А | [ 92 ] | ||||
| IV-Б | [ 92 ] | ||||
| IV-С | [ 92 ] | ||||
| 2 | II | — | Красный9 | Нуклеазы RuvC и HNH вместе продуцируют DSB , а по отдельности могут производить одноцепочечные разрывы. Обеспечивает приобретение функциональных спейсеров при адаптации. | [ 94 ] [ 95 ] |
| II-А | CSN2 | Кольцевой ДНК-связывающий белок. Участвует в первичной адаптации в системе CRISPR типа II. | [ 96 ] | ||
| II-Б | Кас4 | Эндонуклеаза, которая работает с cas1 и cas2 для создания спейсерных последовательностей. | [ 97 ] | ||
| II-С | Характеризуется отсутствием Csn2 или Cas4. | [ 98 ] | |||
| V | — | Кас12 | Нуклеаза RuvC. Не хватает ХНХ. | [ 82 ] [ 99 ] | |
| V-A | Кас12а (Cpf1) | Автоматическая обработка активности пре-crRNA для мультиплексной регуляции генов | [ 92 ] [ 100 ] | ||
| V-B | Кас12б (С2с1) | [ 92 ] | |||
| V-C | Кас12с (С2с3) | [ 92 ] | |||
| V-D | Кас12д (CasY) | [ 92 ] | |||
| ВЕ | Кас12е (КасХ) | [ 92 ] | |||
| ВФ | Кас12f (Кас14, C2c10) | [ 92 ] | |||
| V-G | Кас12г | [ 92 ] | |||
| ВХ | Cas12h | [ 92 ] | |||
| V-I | Cas12i | [ 92 ] | |||
| V-K [ Примечание 2 ] | Кас12к (C2c5) | Тип VK вовлечен в CRISPR-ассоциированные транспозоны . | [ 92 ] | ||
| V-U | С2с4, С2с8, С2с9 | [ 92 ] | |||
| МЫ | — | Кас13 | РНК-ориентированная РНКаза | [ 82 ] [ 101 ] | |
| С ПОМОЩЬЮ | Кас13а(C2c2) | [ 92 ] | |||
| VI-Б | Кас13б | [ 92 ] | |||
| ШУТИТЬ | Кас13с | [ 92 ] | |||
| VI-Д | Кас13д | [ 92 ] | |||
| VI-X | Кас13x.1 | РНК-зависимая РНК-полимераза, профилактическое ингибирование РНК-вируса | [ 102 ] | ||
| VI-Y | [ 102 ] |
Механизм
[ редактировать ]
(1) Приобретение начинается с распознавания вторгшейся ДНК с помощью Cas1 и Cas2 и расщепления протоспейсера.
(2) Протоспейсер лигируют с прямым повтором, соседним с лидерной последовательностью, и
(3) удлинение одной цепи восстанавливает CRISPR и дублирует прямой повтор. Стадии обработки и интерференции crRNA происходят по-разному в каждой из трех основных систем CRISPR.
(4) Первичный транскрипт CRISPR расщепляется генами cas с образованием crRNA.
(5) В системах типа I Cas6e/Cas6f расщепляются в месте соединения оцРНК и дцРНК, образованных шпильками в прямом повторе. В системах типа II используется трансактивирующая (tracr) РНК для образования дцРНК, которая расщепляется Cas9 и РНКазой III. В системах типа III используется гомолог Cas6, который не требует наличия шпилек в прямом повторе для расщепления.
(6) В системах типа II и типа III вторичная обрезка выполняется либо на 5'-, либо на 3'-конце для получения зрелых crRNA.
(7) Зрелые crRNA связываются с белками Cas, образуя интерференционные комплексы.
(8) В системах типа I и типа II взаимодействие между белком и последовательностью PAM необходимо для деградации вторгающейся ДНК. Системы типа III не требуют PAM для успешной деградации, а в системах типа III-A спаривание оснований происходит между crRNA и мРНК, а не между ДНК, на которую нацелены системы типа III-B.



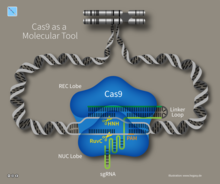

Иммунитет CRISPR-Cas — это естественный процесс бактерий и архей. [ 103 ] CRISPR-Cas предотвращает заражение бактериофагом, его конъюгацию и естественную трансформацию путем разрушения чужеродных нуклеиновых кислот, попадающих в клетку. [ 38 ]
Приобретение проставки
[ редактировать ]Когда микроб подвергается вторжению бактериофага , первым этапом иммунного ответа является захват ДНК фага и вставка ее в локус CRISPR в виде спейсера. Cas1 и Cas2 обнаружены в обоих типах иммунных систем CRISPR-Cas, что указывает на их участие в приобретении спейсера. Исследования мутаций подтвердили эту гипотезу, показав, что удаление Cas1 или Cas2 останавливает приобретение спейсера, не влияя на иммунный ответ CRISPR. [ 104 ] [ 105 ] [ 106 ] [ 107 ] [ 108 ]
Охарактеризовано множество белков Cas1 и решены их структуры. [ 109 ] [ 110 ] [ 111 ] Белки Cas1 имеют разнообразные аминокислотные последовательности. Однако их кристаллические структуры схожи, и все очищенные белки Cas1 представляют собой металлозависимые нуклеазы/ интегразы , которые связываются с ДНК независимым от последовательности образом. [ 85 ] Репрезентативные белки Cas2 были охарактеризованы и обладают либо (одноцепочечными) оцРНК- [ 112 ] или (двухцепочечная) дцДНК- [ 113 ] [ 114 ] специфическая эндорибонуклеазная активность.
В системе IE E. coli Cas1 и Cas2 образуют комплекс, в котором димер Cas2 соединяет два димера Cas1. [ 115 ] В этом комплексе Cas2 выполняет роль неферментативного каркаса. [ 115 ] связывание двухцепочечных фрагментов вторгшейся ДНК, тогда как Cas1 связывает одноцепочечные фланги ДНК и катализирует их интеграцию в массивы CRISPR. [ 116 ] [ 117 ] [ 118 ] Новые спейсеры обычно добавляются в начале CRISPR рядом с лидерной последовательностью, создавая хронологическую запись вирусных инфекций. [ 119 ] В E. coli называемый гистонподобный белок, фактором хозяина интеграции ( IHF ), который связывается с лидерной последовательностью. за точность этой интеграции отвечает [ 120 ] IHF также повышает эффективность интеграции в системе типа IF Pectobacterium atrosepticum . [ 121 ] но в других системах могут потребоваться другие факторы хоста [ 122 ]
Смежные мотивы протоспейсера (PAM)
[ редактировать ]Биоинформатический анализ областей фаговых геномов, которые были вырезаны в качестве спейсеров (называемых протоспейсерами), показал, что они не были выбраны случайным образом, а вместо этого были обнаружены рядом с короткими (3–5 п.о.) последовательностями ДНК, называемыми мотивами, прилегающими к протоспейсерам (PAM). Анализ систем CRISPR-Cas показал, что PAM важны для систем типа I и типа II, но не для систем типа III во время сбора данных. [ 33 ] [ 123 ] [ 124 ] [ 125 ] [ 126 ] [ 127 ] В системах типа I и типа II протоспейсеры вырезаются в позициях, прилегающих к последовательности PAM, при этом другой конец спейсера обрезается с помощью механизма линейки, что позволяет поддерживать регулярность размера спейсера в массиве CRISPR. [ 128 ] [ 129 ] Консервативность последовательности PAM различается в разных системах CRISPR-Cas и, по-видимому, эволюционно связана с Cas1 и лидерной последовательностью . [ 127 ] [ 130 ]
Новые спейсеры добавляются в массив CRISPR направленным образом. [ 31 ] происходит преимущественно, [ 80 ] [ 123 ] [ 124 ] [ 131 ] [ 132 ] но не исключительно, прилегающие [ 126 ] [ 129 ] к лидерской последовательности. Анализ системы типа IE из E. coli показал, что первый прямой повтор, соседний с лидерной последовательностью, копируется, при этом вновь приобретенный спейсер вставляется между первым и вторым прямыми повторами. [ 107 ] [ 128 ]
Последовательность PAM, по-видимому, важна при вставке спейсера в системах типа IE. Эта последовательность содержит сильно консервативный конечный нуклеотид (нт), соседний с первым нуклеотидом протоспейсера. Эта нт становится последней основой в первом прямом повторе. [ 108 ] [ 133 ] [ 134 ] Это предполагает, что механизм получения спейсера генерирует одноцепочечные выступы в предпоследнем положении прямого повтора и в PAM во время вставки спейсера. Однако, по-видимому, не все системы CRISPR-Cas разделяют этот механизм, поскольку PAM в других организмах не демонстрируют такой же уровень консервации в конечном положении. [ 130 ] Вполне вероятно, что в этих системах тупой конец образуется на самом конце прямого повтора и протоспейсера во время приобретения.
Варианты вставки
[ редактировать ]Анализ CRISPR Sulfolobus solfataricus выявил дополнительные сложности канонической модели вставки спейсеров, поскольку один из шести локусов CRISPR встраивал новые спейсеры случайным образом по всему массиву CRISPR, а не вставлял их ближе всего к лидерной последовательности. [ 129 ]
Множественные CRISPR содержат множество спейсеров к одному и тому же фагу. Механизм, вызывающий это явление, был открыт в системе типа IE E. coli . Значительное улучшение приобретения спейсеров было обнаружено там, где спейсеры уже нацелены на фаг, даже если они не соответствуют протоспейсеру. Этот «прайминг» требует, чтобы белки Cas, участвующие как в захвате, так и в интерференции, взаимодействовали друг с другом. Вновь приобретенные спейсеры, возникающие в результате механизма прайминга, всегда обнаруживаются на той же цепи, что и прайминг-спейсер. [ 108 ] [ 133 ] [ 134 ] Это наблюдение привело к гипотезе, что механизм сбора скользит по чужеродной ДНК после прайминга, чтобы найти новый протоспейсер. [ 134 ]
Биогенез
[ редактировать ]CRISPR-РНК (crRNA), которая позже направляет нуклеазу Cas к мишени на этапе интерференции, должна быть сгенерирована из последовательности CRISPR. Первоначально crРНК транскрибируется как часть одного длинного транскрипта, охватывающего большую часть массива CRISPR. [ 29 ] Затем этот транскрипт расщепляется белками Cas с образованием crRNA. Механизм производства crRNA различается в разных системах CRISPR-Cas. В системах типа IE и типа IF белки Cas6e и Cas6f соответственно распознают стеблевые петли. [ 135 ] [ 136 ] [ 137 ] создается путем спаривания идентичных повторов, фланкирующих crRNA. [ 138 ] Эти белки Cas расщепляют более длинный транскрипт на краю парной области, оставляя одну crRNA вместе с небольшим остатком парной области повтора.
Системы типа III также используют Cas6, однако их повторы не создают стебель-петли. Вместо этого расщепление происходит за счет обертывания более длинного транскрипта вокруг Cas6, что позволяет расщеплять непосредственно перед повторяющейся последовательностью. [ 139 ] [ 140 ] [ 141 ]
В системах типа II отсутствует ген Cas6, и вместо этого для расщепления используется РНКаза III. Системы функционального типа II кодируют очень малую РНК, которая комплементарна повторяющейся последовательности, известную как транс-активирующая crРНК (tracrRNA). [ 41 ] Транскрипция tracrRNA и первичного транскрипта CRISPR приводит к спариванию оснований и образованию дцРНК в повторяющейся последовательности, которая впоследствии подвергается воздействию РНКазы III для производства crРНК. В отличие от двух других систем, crRNA не содержит полного спейсера, который вместо этого усечен на одном конце. [ 94 ]
CrRNAs связываются с белками Cas, образуя рибонуклеотидные комплексы, которые узнают чужеродные нуклеиновые кислоты. CrRNA не демонстрируют предпочтения между кодирующими и некодирующими цепями, что указывает на систему нацеливания на ДНК, управляемую РНК. [ 5 ] [ 40 ] [ 104 ] [ 108 ] [ 142 ] [ 143 ] [ 144 ] Комплекс типа IE (обычно называемый каскадом) требует пяти белков Cas, связанных с одной crRNA. [ 145 ] [ 146 ]
Помехи
[ редактировать ]На стадии интерференции в системах типа I последовательность PAM распознается на цепи, комплементарной crRNA, и требуется наряду с отжигом crRNA. В системах типа I правильное спаривание оснований между crРНК и протоспейсером сигнализирует о конформационном изменении в каскаде, который рекрутирует Cas3 для деградации ДНК.
Системы типа II полагаются на один многофункциональный белок Cas9 на этапе интерференции. [ 94 ] Cas9 требует, чтобы как crRNA, так и tracrRNA функционировали и расщепляли ДНК, используя свои двойные HNH и RuvC/RNaseH-подобные эндонуклеазные домены. Спаривание оснований между PAM и геномом фага необходимо в системах типа II. Однако PAM распознается на той же цепи, что и crRNA (противоположная цепь системам типа I).
Системы типа III, как и типа I, требуют связывания шести или семи белков Cas с crRNA. [ 147 ] [ 148 ] Системы типа III, проанализированные на S. Solfataricus и P. Furiosus, нацелены на мРНК фагов, а не на геном фаговой ДНК. [ 86 ] [ 148 ] что может сделать эти системы уникальными способными воздействовать на геномы фагов на основе РНК. [ 85 ] Также было обнаружено, что системы типа III нацелены на ДНК в дополнение к РНК, используя в комплексе другой белок Cas, Cas10. [ 149 ] Было показано, что расщепление ДНК зависит от транскрипции. [ 150 ]
Механизм различения своей ДНК от чужеродной во время интерференции встроен в crRNA и поэтому, вероятно, является общим для всех трех систем. На протяжении отличительного процесса созревания каждого основного типа все crRNA содержат спейсерную последовательность и некоторую часть повтора на одном или обоих концах. Именно частичная повторяющаяся последовательность не позволяет системе CRISPR-Cas нацеливаться на хромосому, поскольку спаривание оснований помимо спейсерной последовательности сигнализирует о себе, и предотвращает расщепление ДНК. [ 151 ] Ферменты CRISPR, управляемые РНК, классифицируются как ферменты рестрикции типа V.
Эволюция
[ редактировать ]| CRISPR-ассоциированный белок Cas2 (адаптационная РНКаза) | |||
|---|---|---|---|
 Кристаллическая структура гипотетического белка tt1823 Thermus thermophilus. | |||
| Идентификаторы | |||
| Символ | CRISPR_Cas2 | ||
| Пфам | PF09827 | ||
| ИнтерПро | ИПР019199 | ||
| CDD | cd09638 | ||
| |||
| CRISPR-ассоциированный белок CasA/Cse1 (эффекторная ДНКаза типа I) | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | CRISPR_Cse1 | ||
| Пфам | PF09481 | ||
| ИнтерПро | ИПР013381 | ||
| CDD | cd09729 | ||
| |||
| CRISPR-ассоциированный белок CasC/Cse3/Cas6 (эффекторная РНКаза типа I) | |||
|---|---|---|---|
 Кристаллическая структура хруст-ассоциированного белка Thermus thermophilus | |||
| Идентификаторы | |||
| Символ | CRISPR_assoc | ||
| Пфам | PF08798 | ||
| Пфам Клан | CL0362 | ||
| ИнтерПро | ИПР010179 | ||
| CDD | cd09727 | ||
| |||
Считается, что гены cas в адаптерном и эффекторном модулях системы CRISPR-Cas произошли от двух разных предковых модулей. Транспозон -подобный элемент , называемый капозоном, кодирующий Cas1-подобную интегразу и, возможно, другие компоненты адаптационного модуля, был вставлен рядом с наследственным эффекторным модулем, который, вероятно, функционировал как независимая врожденная иммунная система. [ 152 ] Высококонсервативные гены cas1 и cas2 адаптерного модуля произошли от предкового модуля, в то время как различные эффекторные cas гены класса 1 произошли от предкового эффекторного модуля. [ 153 ] Эволюция этих различных генов cas эффекторных модулей класса 1 направлялась различными механизмами, такими как события дупликации. [ 154 ] С другой стороны, каждый тип эффекторного модуля класса 2 возник в результате последующих независимых вставок мобильных генетических элементов. [ 155 ] Эти мобильные генетические элементы заменили множество эффекторных модулей генов для создания эффекторных модулей с одним геном, которые производят большие белки, выполняющие все необходимые задачи эффекторного модуля. [ 155 ] Спейсерные области систем CRISPR-Cas взяты непосредственно из чужеродных мобильных генетических элементов, поэтому их долгосрочную эволюцию трудно проследить. [ 156 ] Было обнаружено, что неслучайная эволюция этих спейсерных областей сильно зависит от окружающей среды и конкретных чужеродных мобильных генетических элементов, которые она содержит. [ 157 ]
CRISPR-Cas может иммунизировать бактерии против определенных фагов и тем самым остановить передачу инфекции. По этой причине Кунин описал CRISPR-Cas как ламарковский механизм наследования. [ 158 ] Однако это было оспорено критиком, который отметил: «Мы должны помнить [Ламарка] за то добро, которое он внес в науку, а не за вещи, которые лишь внешне напоминают его теорию. и элегантный способ, которым действительно работает эволюция». [ 159 ] Но по мере проведения более поздних исследований стало очевидно, что приобретенные спейсерные области систем CRISPR-Cas действительно являются формой ламарковской эволюции, поскольку они представляют собой генетические мутации, которые приобретаются, а затем передаются дальше. [ 160 ] С другой стороны, эволюция генного механизма Cas, обеспечивающего работу системы, происходит посредством классической дарвиновской эволюции. [ 160 ]
Коэволюция
[ редактировать ]Анализ последовательностей CRISPR выявил коэволюцию генома хозяина и вируса. [ 161 ]
Базовая модель эволюции CRISPR — это новые спейсеры, которые заставляют фаги мутировать свои геномы, чтобы избежать бактериального иммунного ответа, создавая разнообразие как в популяциях фагов, так и в популяциях хозяев. Чтобы противостоять фаговой инфекции, последовательность спейсера CRISPR должна идеально соответствовать последовательности гена целевого фага. Фаги могут продолжать заражать своих хозяев заданными точечными мутациями в спейсере. [ 151 ] Аналогичная строгость требуется и при ПАМ, иначе бактериальный штамм останется чувствительным к фагам. [ 124 ] [ 151 ]
Цены
[ редактировать ]Исследование 124 штаммов S. thermophilus показало, что 26% всех спейсеров были уникальными и что разные локусы CRISPR демонстрировали разную скорость приобретения спейсеров. [ 123 ] Некоторые локусы CRISPR развиваются быстрее, чем другие, что позволило определить филогенетические взаимоотношения штаммов. Сравнительный геномный анализ показал, что E. coli и S. enterica эволюционируют гораздо медленнее, чем S. thermophilus . Штаммы последнего, разошедшиеся 250 000 лет назад, все еще содержали тот же спейсерный комплемент. [ 162 ]
Метагеномный с дренажем кислотных шахт анализ двух биопленок показал, что один из проанализированных CRISPR содержал обширные делеции и добавления спейсеров по сравнению с другой биопленкой, что указывает на более высокую активность/распространенность фагов в одном сообществе, чем в другом. [ 80 ] В полости рта временное исследование показало, что 7–22% спейсеров использовались совместно в течение 17 месяцев внутри одного человека, тогда как менее 2% были общими для разных людей. [ 132 ]
В той же среде один штамм был отслежен с использованием праймеров ПЦР, специфичных для его системы CRISPR. Результаты общего уровня присутствия/отсутствия спейсера показали значительное разнообразие. Однако в этот CRISPR за 17 месяцев были добавлены три спейсера. [ 132 ] предполагая, что даже в среде со значительным разнообразием CRISPR некоторые локусы эволюционируют медленно.
CRISPR были проанализированы на основе метагеномов, полученных для проекта «Микробиом человека» . [ 163 ] Хотя большинство из них были специфичны для конкретного участка тела, некоторые из них широко распространены среди людей. Один из этих локусов происходил от видов стрептококков и содержал около 15 000 спейсеров, 50% из которых были уникальными. Подобно целевым исследованиям полости рта, некоторые из них показали небольшую эволюцию с течением времени. [ 163 ]
Эволюцию CRISPR изучали в хемостатах с использованием S. thermophilus для непосредственного изучения скорости приобретения спейсеров. За одну неделю штаммы S. thermophilus приобретали до трех спейсеров при заражении одним фагом. [ 164 ] В течение того же периода времени у фага развились однонуклеотидные полиморфизмы , которые зафиксировались в популяции, что позволяет предположить, что нацеливание предотвратило репликацию фага в отсутствие этих мутаций. [ 164 ]
Другой эксперимент с S. thermophilus показал, что фаги могут инфицировать и реплицироваться в хозяевах, имеющих только один целевой спейсер. Еще один показал, что чувствительные хозяева могут существовать в средах с высокими титрами фагов. [ 165 ] Хемостатические и наблюдательные исследования указывают на множество нюансов CRISPR и (ко)эволюции фагов.
Идентификация
[ редактировать ]CRISPR широко распространены среди бактерий и архей. [ 90 ] и показать некоторые сходства последовательностей. [ 138 ] Их наиболее примечательной характеристикой являются повторяющиеся прокладки и прямые повторы. Эта характеристика позволяет легко идентифицировать CRISPR в длинных последовательностях ДНК, поскольку количество повторов снижает вероятность ложноположительного совпадения. [ 166 ]
Анализ CRISPR в метагеномных данных является более сложным, поскольку локусы CRISPR обычно не собираются из-за их повторяющегося характера или из-за вариаций штаммов, что сбивает с толку алгоритмы сборки. Если доступно много эталонных геномов, полимеразную цепную реакцию (ПЦР). для амплификации массивов CRISPR и анализа содержания спейсеров можно использовать [ 123 ] [ 132 ] [ 167 ] [ 168 ] [ 169 ] [ 170 ] Однако этот подход дает информацию только о специально нацеленных CRISPR и об организмах, достаточно представленных в общедоступных базах данных для разработки надежных полимеразных праймеров для ПЦР. Вырожденные праймеры, специфичные для повторов, можно использовать для амплификации спейсеров CRISPR непосредственно из образцов окружающей среды; ампликоны, содержащие два или три спейсера, можно затем собрать с помощью вычислений для реконструкции длинных массивов CRISPR. [ 170 ]
Альтернативой является извлечение и реконструкция массивов CRISPR на основе метагеномных данных. Это сложнее с вычислительной точки зрения, особенно при использовании технологий секвенирования второго поколения (например, 454, Illumina), поскольку короткая длина считывания предотвращает появление более двух или трех повторяющихся единиц в одном считывании. Идентификация CRISPR в необработанных считываниях была достигнута с использованием чисто de novo. идентификации [ 171 ] или с использованием прямых повторяющихся последовательностей в частично собранных массивах CRISPR из контигов (перекрывающихся сегментов ДНК, которые вместе представляют собой консенсусную область ДНК) [ 163 ] и прямые повторяющиеся последовательности из опубликованных геномов [ 172 ] в качестве крючка для выявления прямых повторов в отдельных прочтениях.
Использование фагами
[ редактировать ]Другой способ защиты бактерий от фаговой инфекции — наличие хромосомных островков . Подтип хромосомных островков, называемый индуцируемым фагом хромосомным островом (PICI), вырезается из бактериальной хромосомы при фаговой инфекции и может ингибировать репликацию фага. [ 173 ] PICI индуцируются, вырезаются, реплицируются и, наконец, упаковываются в небольшие капсиды определенными стафилококковыми умеренными фагами. PICI используют несколько механизмов для блокирования размножения фагов. В первом механизме Ppi, кодируемый PICI, дифференциально блокирует созревание фага путем связывания или специфического взаимодействия с фагом TerS, тем самым блокируя образование комплекса фага TerS/TerL, ответственного за упаковку фаговой ДНК. Во втором механизме PICI CpmAB перенаправляет морфогенетический белок фагового капсида, создавая 95% капсида размером с SaPI, и ДНК фага может упаковывать только 1/3 их генома в эти маленькие капсиды и, следовательно, становиться нежизнеспособным фагом. [ 174 ] Третий механизм включает два белка, PtiA и PtiB, которые нацелены на LtrC, который отвечает за выработку вириона и белков лизиса. Этот механизм интерференции модулируется модулирующим белком PtiM, который связывается с одним из белков, опосредующих интерференцию, PtiA и, следовательно, достигает необходимого уровня интерференции. [ 175 ]
Одно исследование показало, что литический фаг ICP1, специально нацеленный на Vibrio cholerae серогруппы O1, приобрел систему CRISPR-Cas, которая нацелена на PICI-подобный элемент V. cholerae . Система имеет 2 локуса CRISPR и 9 генов Cas. Кажется, она гомологична системе IF, обнаруженной у Yersinia pestis . Более того, как и бактериальная система CRISPR-Cas, ICP1 CRISPR-Cas может приобретать новые последовательности, что позволяет фагу и хозяину развиваться совместно. [ 176 ] [ 177 ]
Было показано, что некоторые архейные вирусы несут массивы мини-CRISPR, содержащие один или два спейсера. Было показано, что спейсеры в вирусных массивах CRISPR нацелены на другие вирусы и плазмиды, что позволяет предположить, что мини-чипы CRISPR представляют собой механизм исключения гетеротипической суперинфекции и участвуют в межвирусных конфликтах. [ 170 ]
Приложения
[ редактировать ]Редактирование генов CRISPR — это революционная технология, которая позволяет точно и целенаправленно модифицировать ДНК живых организмов. CRISPR-Cas9, разработанный на основе естественного защитного механизма, обнаруженного у бактерий, является наиболее часто используемой системой, которая позволяет «разрезать» ДНК в определенных местах и удалять, модифицировать или вставлять генетический материал. Эта технология изменила такие области, как генетика, медицина, [ 178 ] [ 179 ] и сельское хозяйство, [ 180 ] [ 181 ] предлагая потенциальные методы лечения генетических нарушений, достижения в области растениеводства и исследования фундаментальных механизмов жизни. Однако его этические последствия и потенциальные непредвиденные последствия вызвали серьезные споры. [ 182 ] [ 183 ]
См. также
[ редактировать ]- Активация CRISPR
- Анти-CRISPR
- Инструменты CRISPR/Cas
- Редактирование генов CRISPR
- Журнал CRISPR
- « Дизайнерский малыш »
- ДРАКО
- Джин нокаут
- Полногеномный нокаут CRISPR-Cas9
- Глоссарий генетики
- Инженерия зародышевой линии человека
- Человеческая природа (документальный фильм, 2019)
- МАГЕСТИК
- Новая евгеника
- Премьер-редактирование
- РНКи
- миРНК
- Нуклеазный анализ Surveyor
- Синтетическая биология
- Цинковый палец
Примечания
[ редактировать ]Ссылки
[ редактировать ]- ^ ВВП : 4QYZ : Мулепати С., Эру А., Бэйли С. (2014). «Кристаллическая структура комплекса наблюдения, управляемого РНК CRISPR, связанного с мишенью оцДНК» . Наука . 345 (6203): 1479–1484. Бибкод : 2014Sci...345.1479M . дои : 10.1126/science.1256996 . ПМЦ 4427192 . ПМИД 25123481 .
- ^ Перейти обратно: а б Баррангу Р. (2015). «Роль систем CRISPR-Cas в адаптивном иммунитете и не только». Современное мнение в иммунологии . 32 : 36–41. дои : 10.1016/j.coi.2014.12.008 . ПМИД 25574773 .
- ^ Редман М., Кинг А., Уотсон С., Кинг Д. (август 2016 г.). «Что такое CRISPR/Cas9?» . Архив детских болезней: издание для образования и практики . 101 (4): 213–215. дои : 10.1136/archdischild-2016-310459 . ПМЦ 4975809 . ПМИД 27059283 .
- ^ Перейти обратно: а б Баррангу Р. , Фремо С., Дево Х., Ричардс М., Бояваль П., Муано С. и др. (март 2007 г.). «CRISPR обеспечивает приобретенную устойчивость прокариот к вирусам». Наука . 315 (5819): 1709–1712. Бибкод : 2007Sci...315.1709B . дои : 10.1126/science.1138140 . hdl : 20.500.11794/38902 . ПМИД 17379808 . S2CID 3888761 . ( требуется регистрация )
- ^ Перейти обратно: а б Марраффини Л.А., Сонтхаймер Э.Дж. (декабрь 2008 г.). «Интерференция CRISPR ограничивает горизонтальный перенос генов у стафилококков путем воздействия на ДНК» . Наука . 322 (5909): 1843–1845. Бибкод : 2008Sci...322.1843M . дои : 10.1126/science.1165771 . ПМК 2695655 . ПМИД 19095942 .
- ^ Хилле Ф., Рихтер Х., Вонг С.П., Братович М., Рессель С., Шарпантье Э. (март 2018 г.). «Биология CRISPR-Cas: вперед и назад». Клетка . 172 (6): 1239–1259. дои : 10.1016/j.cell.2017.11.032 . hdl : 21.11116/0000-0003-FC0D-4 . ПМИД 29522745 . S2CID 3777503 .
- ^ Хорват П., Баррангу Р. (январь 2010 г.). «CRISPR/Cas, иммунная система бактерий и архей». Наука . 327 (5962): 167–170. Бибкод : 2010Sci...327..167H . дои : 10.1126/science.1179555 . ПМИД 20056882 . S2CID 17960960 .
- ^ Бак Р.О., Гомес-Оспина Н., Портеус М.Х. (август 2018 г.). «Редактирование генов в центре внимания». Тенденции в генетике . 34 (8): 600–611. дои : 10.1016/j.tig.2018.05.004 . ПМИД 29908711 . S2CID 49269023 .
- ^ Чжан Ф, Вэнь Ю, Го Икс (2014). «CRISPR/Cas9 для редактирования генома: прогресс, последствия и проблемы» . Молекулярная генетика человека . 23 (Р1): Р40–6. дои : 10.1093/hmg/ddu125 . ПМИД 24651067 . S2CID 19008095 .
- ^ CRISPR-CAS9, TALENS и ZFNS – битва в редактировании генов https://www.ptglab.com/news/blog/crispr-cas9-talens-and-zfns-the-battle-in-gene-editing/ Архивировано в 2021 г. -05-25 в Wayback Machine
- ^ Перейти обратно: а б с д и Сюй П.Д., Ландер Э.С., Чжан Ф. (июнь 2014 г.). «Разработка и применение CRISPR-Cas9 для генной инженерии» . Клетка . 157 (6): 1262–1278. дои : 10.1016/j.cell.2014.05.010 . ПМЦ 4343198 . ПМИД 24906146 .
- ^ «Пресс-релиз: Нобелевская премия по химии 2020» . Нобелевский фонд. Архивировано из оригинала 15 января 2021 года . Проверено 7 октября 2020 г.
- ^ Ву К.Дж., Пельтье Э. (7 октября 2020 г.). «Нобелевская премия по химии присуждена двум ученым за работу по редактированию генома — Эммануэль Шарпантье и Дженнифер А. Дудна разработали инструмент Crispr, который может с высокой точностью изменять ДНК животных, растений и микроорганизмов» . Нью-Йорк Таймс . Архивировано из оригинала 8 октября 2020 года . Проверено 7 октября 2020 г.
- ^ Рават А., Рой М., Джьоти А., Кошик С., Верма К., Шривастава В.К. (август 2021 г.). «Цистеиновые протеазы: борьба с патогенными паразитическими простейшими с помощью вездесущих ферментов» . Микробиологические исследования . 249 : 126784. doi : 10.1016/j.micres.2021.126784 . ПМИД 33989978 . S2CID 234597200 .
- ^ Перейти обратно: а б Исино Ю, Синагава Х, Макино К, Амемура М, Наката А (декабрь 1987 г.). «Нуклеотидная последовательность гена iap, ответственного за превращение изофермента щелочной фосфатазы в Escherichia coli, и идентификация продукта гена» . Журнал бактериологии . 169 (12): 5429–5433. дои : 10.1128/jb.169.12.5429-5433.1987 . ПМК 213968 . ПМИД 3316184 .
- ^ ван Соолинген Д., де Хаас П.Е., Херманс П.В., Гроенен П.М., ван Эмбден Дж.Д. (август 1993 г.). «Сравнение различных повторяющихся элементов ДНК как генетических маркеров штаммовой дифференциации и эпидемиологии микобактерий туберкулеза» . Журнал клинической микробиологии . 31 (8): 1987–1995. doi : 10.1128/JCM.31.8.1987-1995.1993 . ПМК 265684 . ПМИД 7690367 .
- ^ Гроенен П.М., Буншотен А.Е., ван Соолинген Д., ван Эмбден Дж.Д. (декабрь 1993 г.). «Природа полиморфизма ДНК в кластере прямых повторов микобактерии туберкулеза; применение для дифференциации штаммов новым методом типирования». Молекулярная микробиология . 10 (5): 1057–1065. дои : 10.1111/j.1365-2958.1993.tb00976.x . ПМИД 7934856 . S2CID 25304723 .
- ^ Перейти обратно: а б с Мохика Ф.Дж., Монтолиу Л. (2016). «О происхождении технологии CRISPR-Cas: от прокариот к млекопитающим». Тенденции в микробиологии . 24 (10): 811–820. дои : 10.1016/j.tim.2016.06.005 . ПМИД 27401123 .
- ^ Перейти обратно: а б Мохика Ф.Дж., Родригес-Валера Ф. (2016). «Открытие CRISPR у архей и бактерий» (PDF) . Журнал ФЭБС . 283 (17): 3162–3169. дои : 10.1111/февраль 13766 . hdl : 10045/57676 . ПМИД 27234458 . S2CID 42827598 . Архивировано (PDF) из оригинала 25 февраля 2021 г. Проверено 29 августа 2019 г.
- ^ Мохика Ф.Дж., Диес-Вильясеньор К., Сориа Э., Хуэс Дж. (апрель 2000 г.). «Биологическое значение семейства регулярно расположенных повторов в геномах архей, бактерий и митохондрий» . Молекулярная микробиология . 36 (1): 244–246. дои : 10.1046/j.1365-2958.2000.01838.x . ПМИД 10760181 .
- ^ Исааксон В. (2021). Взломщик кодов: Дженнифер Дудна, редактирование генов и будущее человечества . Нью-Йорк: Саймон и Шустер. п. 73. ИСБН 978-1-9821-1585-2 . OCLC 1239982737 . Архивировано из оригинала 14 января 2023 г. Проверено 20 октября 2021 г.
- ^ Баррангу Р. , ван дер Ост Дж. (2013). Системы CRISPR-Cas: РНК-опосредованный адаптивный иммунитет у бактерий и архей . Гейдельберг: Спрингер. п. 6. ISBN 978-3-642-34656-9 .
- ^ Тан Т.Х., Бачеллери Дж.П., Рождественский Т., Бортолин М.Л., Хубер Х., Друнговски М. и др. (май 2002 г.). «Идентификация 86 кандидатов на малые непосредственные РНК из археи Archaeoglobus fulgidus» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (11): 7536–7541. Бибкод : 2002PNAS...99.7536T . дои : 10.1073/pnas.112047299 . ПМК 124276 . ПМИД 12032318 .
- ^ Шарпантье Э., Рихтер Х., ван дер Ост Дж., Уайт М.Ф. (май 2015 г.). «Пути биогенеза РНК-направляющих в адаптивном иммунитете CRISPR-Cas архей и бактерий» . Обзоры микробиологии FEMS . 39 (3): 428–441. дои : 10.1093/femsre/fuv023 . ПМЦ 5965381 . ПМИД 25994611 .
- ^ Ромеро Д.А., Мэгилл Д., Миллен А., Хорват П., Фремо К. (ноябрь 2020 г.). «Взаимодействие молочного лактококкового и стрептококкового фага с хозяином: промышленная перспектива в развивающемся фаговом ландшафте». Обзоры микробиологии FEMS . 44 (6): 909–932. дои : 10.1093/femsre/fuaa048 . ПМИД 33016324 .
- ^ Молтени М., Хакинс Дж. (1 августа 2020 г.). «Руководство WIRED по Crispr» . Конде Наст. Проводной журнал. Архивировано из оригинала 23 октября 2021 года . Проверено 23 февраля 2021 г.
- ^ Янсен Р., Эмбден Дж. Д., Гаастра В., Шоулс Л. М. (март 2002 г.). «Идентификация генов, связанных с повторами ДНК у прокариот» . Молекулярная микробиология . 43 (6): 1565–1575. дои : 10.1046/j.1365-2958.2002.02839.x . ПМИД 11952905 . S2CID 23196085 .
- ^ Перейти обратно: а б Хорват П., Баррангу Р. (январь 2010 г.). «CRISPR/Cas, иммунная система бактерий и архей». Наука . 327 (5962): 167–170. Бибкод : 2010Sci...327..167H . дои : 10.1126/Science.1179555 . ПМИД 20056882 . S2CID 17960960 .
- ^ Перейти обратно: а б Марраффини Л.А., Сонтхаймер Э.Дж. (март 2010 г.). «CRISPR-интерференция: РНК-направленный адаптивный иммунитет у бактерий и архей» . Обзоры природы Генетика . 11 (3): 181–190. дои : 10.1038/nrg2749 . ПМЦ 2928866 . ПМИД 20125085 .
- ^ Грисса I, Верно Дж., Пурсель К. (май 2007 г.). «База данных CRISPRdb и инструменты для отображения CRISPR и создания словарей спейсеров и повторов» . БМК Биоинформатика . 8 : 172. дои : 10.1186/1471-2105-8-172 . ПМК 1892036 . ПМИД 17521438 .
- ^ Перейти обратно: а б Пурсель К., Сальвиньоль Дж., Верно Дж. (март 2005 г.). «Элементы CRISPR в Yersinia pestis приобретают новые повторы за счет преимущественного поглощения ДНК бактериофага и предоставляют дополнительные инструменты для эволюционных исследований» . Микробиология . 151 (Часть 3): 653–663. дои : 10.1099/mic.0.27437-0 . ПМИД 15758212 .
- ^ Перейти обратно: а б Мохика Ф.Х., Диес-Вильясеньор К., Гарсиа-Мартинес Х., Сория Э. (февраль 2005 г.). «Промежуточные последовательности прокариотических повторов с регулярными интервалами происходят от чужеродных генетических элементов». Журнал молекулярной эволюции . 60 (2): 174–182. Бибкод : 2005JMolE..60..174M . дои : 10.1007/s00239-004-0046-3 . ПМИД 15791728 . S2CID 27481111 .
- ^ Перейти обратно: а б Болотин А, Квинкис Б, Сорокин А, Эрлих С.Д. (август 2005 г.). «Кластерные короткие палиндромные повторы с регулярными интервалами (CRISPR) имеют спейсеры внехромосомного происхождения» . Микробиология . 151 (Часть 8): 2551–2561. дои : 10.1099/mic.0.28048-0 . ПМИД 16079334 .
- ^ Моранж М. (июнь 2015 г.). «Что говорит нам история XXXVII. CRISPR-Cas: открытие иммунной системы у прокариот» (PDF) . Журнал биологических наук . 40 (2): 221–223. дои : 10.1007/s12038-015-9532-6 . ПМИД 25963251 .
- ^ Лендер ES (январь 2016 г.). «Герои CRISPR» . Клетка . 164 (1–2): 18–28. дои : 10.1016/j.cell.2015.12.041 . ПМИД 26771483 .
- ^ Макарова К.С., Гришин Н.В., Шабалина С.А., Вольф Ю.И., Кунин Е.В. (март 2006 г.). «Предполагаемая иммунная система прокариот, основанная на РНК-интерференции: вычислительный анализ предсказанного ферментативного механизма, функциональные аналогии с эукариотическими РНКи и гипотетические механизмы действия» . Биология Директ . 1 :7. дои : 10.1186/1745-6150-1-7 . ПМК 1462988 . ПМИД 16545108 .
- ^ Пенниси Э. (август 2013 г.). «Увлечение CRISPR». Новости Фокус. Наука . 341 (6148): 833–836. Бибкод : 2013Sci...341..833P . дои : 10.1126/science.341.6148.833 . ПМИД 23970676 .
- ^ Перейти обратно: а б с Марраффини, Лос-Анджелес (октябрь 2015 г.). «CRISPR-Cas иммунитет у прокариот». Природа . 526 (7571): 55–61. Бибкод : 2015Природа.526...55М . дои : 10.1038/nature15386 . ПМИД 26432244 . S2CID 3718361 .
- ^ Браунс С.Дж., Джор М.М., Лундгрен М., Вестра Э.Р., Слиджхуис Р.Дж., Снейдерс А.П. и др. (август 2008 г.). «Маленькие РНК CRISPR обеспечивают противовирусную защиту прокариот» . Наука . 321 (5891): 960–964. Бибкод : 2008Sci...321..960B . дои : 10.1126/science.1159689 . ПМЦ 5898235 . ПМИД 18703739 .
- ^ Перейти обратно: а б Гарно Дж.Э., Дюпюи М., Виллион М., Ромеро Д.А., Баррангу Р. , Бояваль П. и др. (ноябрь 2010 г.). «Бактериальная иммунная система CRISPR/Cas расщепляет ДНК бактериофага и плазмиды». Природа 468 (7320): 67–71. Бибкод : 2010Природа.468...67Г . CiteSeerX 10.1.1.451.9645 . дои : 10.1038/nature09523 . ПМИД 21048762 . S2CID 205222849 .
- ^ Перейти обратно: а б Дельчева Е., Чилински К., Шарма С.М., Гонсалес К., Чао Ю., Пирзада З.А. и др. (март 2011 г.). «Созревание РНК CRISPR с помощью транскодируемой малой РНК и фактора хозяина РНКазы III» . Природа . 471 (7340): 602–607. Бибкод : 2011Natur.471..602D . дои : 10.1038/nature09886 . ПМК 3070239 . ПМИД 21455174 .
- ^ Баррангу Р. (ноябрь 2015 г.). «Разнообразие CRISPR-Cas иммунных систем и молекулярных машин» . Геномная биология . 16 : 247. дои : 10.1186/s13059-015-0816-9 . ПМЦ 4638107 . ПМИД 26549499 .
- ^ Перейти обратно: а б Джинек М., Чилински К., Фонфара И., Хауэр М., Дудна Дж.А. , Шарпантье Э. (август 2012 г.). «Программируемая ДНК-эндонуклеаза, управляемая двойной РНК, в адаптивном бактериальном иммунитете» . Наука . 337 (6096): 816–821. Бибкод : 2012Sci...337..816J . дои : 10.1126/science.1225829 . ПМК 6286148 . ПМИД 22745249 .
- ^ Конг Л., Ран Ф.А., Кокс Д., Лин С., Барретто Р., Хабиб Н. и др. (февраль 2013 г.). «Мультиплексная геномная инженерия с использованием систем CRISPR/Cas» . Наука . 339 (6121): 819–823. Бибкод : 2013Sci...339..819C . дои : 10.1126/science.1231143 . ПМЦ 3795411 . ПМИД 23287718 .
- ^ Мали П., Ян Л., Эсвелт К.М., Аах Дж., Гуэлл М., ДиКарло Дж.Э. и др. (февраль 2013 г.). «Инженерия генома человека под контролем РНК с помощью Cas9» . Наука . 339 (6121): 823–826. Бибкод : 2013Sci...339..823M . дои : 10.1126/science.1232033 . ПМЦ 3712628 . ПМИД 23287722 .
- ^ ДиКарло Дж.Э., Норвилл Дж.Э., Мали П., Риос Х, Аах Дж., Черч ГМ (апрель 2013 г.). «Геномная инженерия Saccharomyces cerevisiae с использованием систем CRISPR-Cas» . Исследования нуклеиновых кислот . 41 (7): 4336–4343. дои : 10.1093/нар/gkt135 . ПМЦ 3627607 . ПМИД 23460208 .
- ^ Чжан Г.К., Конг II, Ким Х, Лю Дж.Дж., Кейт Дж.Х., Джин Ю.С. (декабрь 2014 г.). «Создание четверного ауксотрофного мутанта промышленного полиплоидного штамма saccharomyces cerevisiae с использованием РНК-ориентированной нуклеазы Cas9» . Прикладная и экологическая микробиология . 80 (24): 7694–7701. Бибкод : 2014ApEnM..80.7694Z . дои : 10.1128/АЕМ.02310-14 . ПМЦ 4249234 . ПМИД 25281382 .
- ^ Лю Дж.Дж., Конг II, Чжан Г.К., Джаякоди Л.Н., Ким Х., Ся П.Ф. и др. (апрель 2016 г.). «Метаболическая инженерия пробиотиков Saccharomyces boulardii » . Прикладная и экологическая микробиология . 82 (8): 2280–2287. Бибкод : 2016ApEnM..82.2280L . дои : 10.1128/AEM.00057-16 . ПМЦ 4959471 . ПМИД 26850302 .
- ^ Вяс В.К., Барраса М.И., Финк Г.Р. (2015). « Система CRISPR Candida albicans позволяет осуществлять генную инженерию важнейших генов и семейств генов» . Достижения науки . 1 (3): e1500248. Бибкод : 2015SciA....1E0248V . дои : 10.1126/sciadv.1500248 . ПМЦ 4428347 . ПМИД 25977940 .
- ^ Нг Х, Дин Н (2017). « Candida albicans за счет увеличения экспрессии единой направляющей РНК» . мСфера . 2 (2): e00385–16. дои : 10.1128/mSphere.00385-16 . ПМЦ 5397569 . ПМИД 28435892 .
- ^ Хван Вай, Фу Ю, Рейон Д., Мэдер М.Л., Цай С.К., Сандер Дж.Д. и др. (март 2013 г.). «Эффективное редактирование генома рыбок данио с использованием системы CRISPR-Cas» . Природная биотехнология . 31 (3): 227–229. дои : 10.1038/nbt.2501 . ПМЦ 3686313 . ПМИД 23360964 .
- ^ Грац С.Дж., Каммингс А.М., Нгуен Дж.Н., Хэмм Д.К., Донохью Л.К., Харрисон М.М. и др. (август 2013 г.). «Геномная инженерия дрозофилы с помощью CRISPR-РНК-ориентированной нуклеазы Cas9» . Генетика . 194 (4): 1029–1035. дои : 10.1534/генетика.113.152710 . ПМЦ 3730909 . ПМИД 23709638 .
- ^ Бассетт А.Р., Тиббит С., Понтинг К.П., Лю Дж.Л. (июль 2013 г.). «Высокоэффективный таргетный мутагенез дрозофилы с помощью системы CRISPR/Cas9» . Отчеты по ячейкам . 4 (1): 220–228. дои : 10.1016/j.celrep.2013.06.020 . ПМЦ 3714591 . ПМИД 23827738 .
- ^ Ян Х., Опахалоэмфан С., Манчини Г., Ян Х., Галлитто М., Млейнек Дж. и др. (август 2017 г.). «Инженерная мутация орко приводит к аберрантному социальному поведению и дефектному развитию нервной системы у муравьев» . Клетка . 170 (4): 736–747.e9. дои : 10.1016/j.cell.2017.06.051 . ПМЦ 5587193 . ПМИД 28802043 .
- ^ Трайбл В., Оливос-Сиснерос Л., Маккензи С.К., Сарагости Дж., Чанг Н.К., Мэтьюз Б.Дж. и др. (август 2017 г.). «Мутагенез орко вызывает потерю клубочков антеннальных долей и нарушение социального поведения у муравьев» . Клетка . 170 (4): 727–735.e10. дои : 10.1016/j.cell.2017.07.001 . ПМК 5556950 . ПМИД 28802042 .
- ^ Кистлер К.Е., Восшалл Л.Б., Мэтьюз Б.Дж. (апрель 2015 г.). «Геномная инженерия с использованием CRISPR-Cas9 у комара Aedes aegypti» . Отчеты по ячейкам . 11 (1): 51–60. дои : 10.1016/j.celrep.2015.03.009 . ПМК 4394034 . ПМИД 25818303 .
- ^ Фридланд А.Е., Цур Ю.Б., Эсвелт К.М., депутат Колаяково, генеральный директор Черча, Каларко Х.А. (август 2013 г.). «Наследственное редактирование генома C. elegans с помощью системы CRISPR-Cas9» . Природные методы . 10 (8): 741–743. дои : 10.1038/nmeth.2532 . ПМЦ 3822328 . ПМИД 23817069 .
- ^ Цзян В., Чжоу Х., Би Х., Фромм М., Ян Б., Уикс Д.П. (ноябрь 2013 г.). «Демонстрация целевой модификации генов, опосредованной CRISPR/Cas9/sgRNA, у Arabidopsis, табака, сорго и риса» . Исследования нуклеиновых кислот . 41 (20): e188. дои : 10.1093/nar/gkt780 . ПМЦ 3814374 . ПМИД 23999092 .
- ^ Ван Х., Ян Х., Шивалила К.С., Давлати М.М., Ченг А.В., Чжан Ф. и др. (май 2013 г.). «Одноэтапное создание мышей, несущих мутации в нескольких генах, с помощью геномной инженерии, опосредованной CRISPR/Cas» . Клетка . 153 (4): 910–918. дои : 10.1016/j.cell.2013.04.025 . ПМЦ 3969854 . ПМИД 23643243 .
- ^ Сони Д., Ван Д.М., Регми С.К., Миттал М., Фогель С.М., Шлютер Д. и др. (май 2018 г.). «Дебиквитиназная функция А20 поддерживает и восстанавливает эндотелиальный барьер после повреждения сосудов легких» . Открытие клеточной смерти . 4 (60): 60. дои : 10.1038/s41420-018-0056-3 . ПМЦ 5955943 . ПМИД 29796309 .
- ^ Го X, Ли XJ (июль 2015 г.). «Направленное редактирование генома эмбрионов приматов» . Клеточные исследования . 25 (7): 767–768. дои : 10.1038/cr.2015.64 . ПМЦ 4493275 . ПМИД 26032266 .
- ^ Балтимор Д., Берг П., Ботчан М., Кэрролл Д., Чаро Р.А., Черч Дж. и др. (апрель 2015 г.). «Биотехнология. Разумный путь вперед в области геномной инженерии и модификации генов зародышевой линии» . Наука . 348 (6230): 36–38. Бибкод : 2015Sci...348...36B . дои : 10.1126/science.aab1028 . ПМЦ 4394183 . ПМИД 25791083 .
- ^ Ларсон М.Х., Гилберт Л.А., Ван X, Лим В.А., Вайсман Дж.С., Ци Л.С. (ноябрь 2013 г.). «CRISPR-интерференция (CRISPRi) для последовательно-специфического контроля экспрессии генов» . Протоколы природы . 8 (11): 2180–2196. дои : 10.1038/nprot.2013.132 . ПМЦ 3922765 . ПМИД 24136345 .
- ^ Лян П., Сюй Ю, Чжан Х, Дин С., Хуан Р., Чжан З. и др. (май 2015 г.). «CRISPR/Cas9-опосредованное редактирование генов в трехпронуклеарных зиготах человека» . Белок и клетка . 6 (5): 363–372. дои : 10.1007/s13238-015-0153-5 . ПМЦ 4417674 . ПМИД 25894090 .
- ^ Yan MY, Yan HQ, Ren GX, Zhao JP, Guo XP, Sun YC (сентябрь 2017 г.). «Рекомбинация с помощью CRISPR-Cas12a в бактериях» . Прикладная и экологическая микробиология . 83 (17). Бибкод : 2017ApEnM..83E.947Y . дои : 10.1128/AEM.00947-17 . ПМК 5561284 . ПМИД 28646112 .
- ^ Зетше Б., Гутенберг Дж.С., Абудайе О.О., Слеймейкер И.М., Макарова К.С., Эсслетцбихлер П. и др. (октябрь 2015 г.). «Cpf1 представляет собой единственную РНК-ориентированную эндонуклеазу системы CRISPR-Cas класса 2» . Клетка . 163 (3): 759–771. дои : 10.1016/j.cell.2015.09.038 . ПМК 4638220 . ПМИД 26422227 .
- ^ Фонфара И, Рихтер Х, Братович М, Ле Рун А, Шарпантье Э (апрель 2016 г.). «Связанный с CRISPR фермент, расщепляющий ДНК Cpf1, также обрабатывает РНК-предшественник CRISPR». Природа . 532 (7600): 517–521. Бибкод : 2016Natur.532..517F . дои : 10.1038/nature17945 . ПМИД 27096362 . S2CID 2271552 .
- ^ Ким Х, Ким С.Т., Рю Дж., Кан Б.С., Ким Дж.С. и Ким С.Г. (февраль 2017 г.). «Редактирование генома растений без ДНК, опосредованное CRISPR / Cpf1» . Природные коммуникации . 8 (14406): 14406. Бибкод : 2017NatCo...814406K . дои : 10.1038/ncomms14406 . ПМК 5316869 . ПМИД 28205546 .
- ^ «Нуклеаза Cpf1» . abmgood.com . Архивировано из оригинала 23 октября 2021 г. Проверено 14 декабря 2017 г.
- ^ Чен Дж.С., Ма Э., Харрингтон Л.Б., Да Коста М., Тянь Х., Палефски Дж.М. и др. (апрель 2018 г.). «Связывание CRISPR-Cas12a с мишенью высвобождает неизбирательную активность одноцепочечной ДНКазы» . Наука . 360 (6387): 436–439. Бибкод : 2018Sci...360..436C . дои : 10.1126/science.aar6245 . ПМК 6628903 . ПМИД 29449511 .
- ^ Бротон Дж.П., Денг Х., Ю.Г., Фашинг К.Л., Сервеллита В., Сингх Дж. и др. (июль 2020 г.). «Обнаружение SARS-CoV-2 на основе CRISPR-Cas12» . Природная биотехнология . 38 (7): 870–874. дои : 10.1038/s41587-020-0513-4 . ПМЦ 9107629 . ПМИД 32300245 .
- ^ Нгуен Л.Т., Смит Б.М., Джайн П.К. (сентябрь 2020 г.). «Усиление активности транс-расщепления Cas12a с помощью сконструированной crRNA позволяет обнаруживать амплифицированные нуклеиновые кислоты» . Природные коммуникации . 11 (1): 4906. Бибкод : 2020NatCo..11.4906N . дои : 10.1038/s41467-020-18615-1 . ПМЦ 7528031 . ПМИД 32999292 .
- ^ Абудайе О.О., Гутенберг Дж.С., Конерманн С., Йонг Дж., Слеймейкер И.М., Кокс Д.Б. и др. (август 2016 г.). «C2c2 представляет собой однокомпонентный программируемый РНК-направляемый эффектор CRISPR, нацеленный на РНК» . Наука . 353 (6299): aaf5573. doi : 10.1126/science.aaf5573 . ПМК 5127784 . ПМИД 27256883 .
- ^ Гутенберг Дж.С., Абудайе О.О., Ли Дж.В., Эсслетцбихлер П., Ди А.Дж., Йонг Дж. и др. (апрель 2017 г.). «Обнаружение нуклеиновой кислоты с помощью CRISPR-Cas13a/C2c2» . Наука . 356 (6336): 438–442. Бибкод : 2017Sci...356..438G . дои : 10.1126/science.aam9321 . ПМК 5526198 . ПМИД 28408723 .
- ^ Гутенберг Дж.С., Абудайе О.О., Келлнер М.Дж., Йонг Дж., Коллинз Дж.Дж., Чжан Ф. (апрель 2018 г.). «Мультиплексная и портативная платформа для обнаружения нуклеиновых кислот с Cas13, Cas12a и Csm6» . Наука . 360 (6387): 439–444. Бибкод : 2018Sci...360..439G . дои : 10.1126/science.aaq0179 . ПМЦ 5961727 . ПМИД 29449508 .
- ^ Ивасаки Р.С., Бати RT (сентябрь 2020 г.). «СПРИНТ: платформа на основе Cas13a для обнаружения малых молекул» . Исследования нуклеиновых кислот . 48 (17): е101. дои : 10.1093/nar/gkaa673 . ПМЦ 7515716 . ПМИД 32797156 .
- ^ Махас А., Ван Ц, Марсик Т., Махфуз М.М. (октябрь 2021 г.). «Новая миниатюрная система CRISPR-Cas13 для диагностики SARS-CoV-2» . ACS Синтетическая биология . 10 (10): 2541–2551. doi : 10.1021/acsynbio.1c00181 . ПМЦ 8482783 . ПМИД 34546709 .
- ^ Хилле Ф., Шарпантье Э (ноябрь 2016 г.). «CRISPR-Cas: биология, механизмы и актуальность» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 371 (1707): 20150496. doi : 10.1098/rstb.2015.0496 . ПМК 5052741 . ПМИД 27672148 .
- ^ Перейти обратно: а б с Баррангу Р. , Марраффини Л.А. (апрель 2014 г.). «Системы CRISPR-Cas: переход прокариот к адаптивному иммунитету» . Молекулярная клетка . 54 (2): 234–244. doi : 10.1016/j.molcel.2014.03.011 . ПМК 4025954 . ПМИД 24766887 .
- ^ Перейти обратно: а б с Тайсон Г.В., Банфилд Дж.Ф. (январь 2008 г.). «Быстро развивающиеся CRISPR, участвующие в приобретенной устойчивости микроорганизмов к вирусам». Экологическая микробиология . 10 (1): 200–207. Бибкод : 2008EnvMi..10..200T . дои : 10.1111/j.1462-2920.2007.01444.x . ПМИД 17894817 .
- ^ Перейти обратно: а б Кунин Е.В., Макарова К.С. (май 2019). «Происхождение и эволюция систем CRISPR-Cas» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 374 : 20180087.doi : (1772 ) 10.1098/rstb.2018.0087 . ПМК 6452270 . ПМИД 30905284 .
- ^ Перейти обратно: а б с Райт А.В., Нуньес Х.К., Дудна Х.А. (январь 2016 г.). «Биология и применение систем CRISPR: использование природного инструментария для генной инженерии» . Клетка . 164 (1–2): 29–44. дои : 10.1016/j.cell.2015.12.035 . ПМИД 26771484 .
- ^ Макарова К.С., Вольф Ю.И., Иранзо Дж., Шмаков С.А., Алхнбаши О.С., Браунс С.Дж. и др. (декабрь 2019 г.). «Эволюционная классификация систем CRISPR–Cas: взрыв класса 2 и производные варианты» . Обзоры природы Микробиология . 18 (1): 67–83. дои : 10.1038/s41579-019-0299-x . hdl : 10045/102627 . ПМЦ 8905525 . ПМИД 31857715 .
- ^ Перейти обратно: а б с д и ж г час Макарова К.С., Вольф Ю.И., Алхнбаши О.С., Коста Ф., Шах С.А., Сондерс С.Дж. и др. (ноябрь 2015 г.). «Обновленная эволюционная классификация систем CRISPR-Cas» . Обзоры природы. Микробиология . 13 (11): 722–736. дои : 10.1038/nrmicro3569 . ПМК 5426118 . ПМИД 26411297 .
- ^ Перейти обратно: а б с Виденхефт Б., Штернберг С.Х., Дудна Дж.А. (февраль 2012 г.). «Системы генетического молчания, управляемые РНК, у бактерий и архей». Природа . 482 (7385): 331–338. Бибкод : 2012Natur.482..331W . дои : 10.1038/nature10886 . ПМИД 22337052 . S2CID 205227944 .
- ^ Перейти обратно: а б Дэн Л., Гарретт Р.А., Шах С.А., Пэн X, Ше Q (март 2013 г.). «Новый механизм взаимодействия с помощью модуля CRISPR-Cmr типа IIIB в Sulfolobus». Молекулярная микробиология . 87 (5): 1088–1099. дои : 10.1111/mmi.12152 . ПМИД 23320564 .
- ^ Синкунас Т., Гасюнас Г., Фремо С., Баррангу Р. , Хорват П., Сикснис В. (апрель 2011 г.). «Cas3 представляет собой одноцепочечную ДНК-нуклеазу и АТФ-зависимую геликазу в иммунной системе CRISPR/Cas» . Журнал ЭМБО . 30 (7): 1335–1342. дои : 10.1038/emboj.2011.41 . ПМК 3094125 . ПМИД 21343909 .
- ^ Хуо Ю, Нам К.Х., Дин Ф., Ли Х., Ву Л., Сяо Ю. и др. (сентябрь 2014 г.). «Структуры CRISPR Cas3 предлагают механистическое понимание каскадно-активируемого раскручивания и деградации ДНК» . Структурная и молекулярная биология природы . 21 (9): 771–777. дои : 10.1038/nsmb.2875 . ПМК 4156918 . ПМИД 25132177 .
- ^ Брендель Дж., Столл Б., Ланге С.Дж., Шарма К., Ленц С., Стахлер А.Е. и др. (март 2014 г.). «Комплекс белков Cas 5, 6 и 7 необходим для биогенеза и стабильности кластеризованных, регулярно расположенных коротких палиндромных повторов (crispr), производных РНК (crrnas) в Haloferax volcanii» . Журнал биологической химии . 289 (10): 7164–77. дои : 10.1074/jbc.M113.508184 . ПМЦ 3945376 . ПМИД 24459147 .
- ^ Перейти обратно: а б Чилинский К., Макарова К.С., Шарпантье Э., Кунин Е.В. (июнь 2014 г.). «Классификация и эволюция систем CRISPR-Cas типа II» . Исследования нуклеиновых кислот . 42 (10): 6091–6105. дои : 10.1093/nar/gku241 . ПМК 4041416 . ПМИД 24728998 .
- ^ Перейти обратно: а б Макарова К.С., Аравинд Л., Вольф Ю.И., Кунин Е.В. (июль 2011 г.). «Объединение семейств белков Cas и простой сценарий возникновения и эволюции систем CRISPR-Cas» . Биология Директ . 6:38 . дои : 10.1186/1745-6150-6-38 . ПМК 3150331 . ПМИД 21756346 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В Макарова К.С., Вольф Ю.И., Иранзо Дж., Шмаков С.А., Алхнбаши О.С., Браунс С.Дж. и др. (февраль 2020 г.). «Эволюционная классификация систем CRISPR-Cas: взрыв класса 2 и производные варианты» . Обзоры природы. Микробиология . 18 (2): 67–83. дои : 10.1038/s41579-019-0299-x . hdl : 10045/102627 . ПМЦ 8905525 . ПМИД 31857715 . S2CID 209420490 .
- ^ Перейти обратно: а б Могила И, Казлаускене М, Валинските С, Тамулатиене Г, Тамулайтис Г, Сикснис В (март 2019 г.). «Генетическое исследование комплекса Csm системы CRISPR-Cas типа III-A выявляет роль отдельных субъединиц» . Отчеты по ячейкам . 26 (10): 2753–2765.e4. дои : 10.1016/j.celrep.2019.02.029 . ПМИД 30840895 .
- ^ Перейти обратно: а б с Гасюнас Г., Баррангу Р. , Хорват П., Сикснис В. (сентябрь 2012 г.). «Рибонуклеопротеиновый комплекс Cas9-crRNA опосредует специфическое расщепление ДНК для адаптивного иммунитета у бактерий» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (39): E2579–2586. Бибкод : 2012PNAS..109E2579G . дои : 10.1073/pnas.1208507109 . ПМЦ 3465414 . ПМИД 22949671 .
- ^ Хелер Р., Самай П., Моделл Дж.В., Вайнер С., Голдберг Г.В., Бикард Д. и др. (март 2015 г.). «Cas9 определяет функциональные вирусные мишени во время адаптации CRISPR-Cas» . Природа . 519 (7542): 199–202. Бибкод : 2015Natur.519..199H . дои : 10.1038/nature14245 . ПМЦ 4385744 . ПМИД 25707807 .
- ^ Нам К.Х., Куринов И., Ке А. (сентябрь 2011 г.). «Кристаллическая структура кластеризованных коротких палиндромных повторов с регулярными промежутками (CRISPR), ассоциированных с белком Csn2, выявила Ca2+-зависимую активность связывания двухцепочечной ДНК» . Журнал биологической химии . 286 (35): 30759–30768. дои : 10.1074/jbc.M111.256263 . ПМК 3162437 . ПМИД 21697083 .
- ^ Ли Х., Дхингра Ю., Сашитал Д.Г. (апрель 2019 г.). «Комплекс Cas4-Cas1-Cas2 обеспечивает точную обработку преспейсера во время адаптации CRISPR» . электронная жизнь . 8 . doi : 10.7554/eLife.44248 . ПМК 6519985 . ПМИД 31021314 .
- ^ Чилински К., Ле Рун А., Шарпантье Э. (май 2013 г.). «Семейства tracrRNA и Cas9 систем иммунитета CRISPR-Cas типа II» . Биология РНК . 10 (5): 726–737. дои : 10.4161/rna.24321 . ПМЦ 3737331 . ПМИД 23563642 .
- ^ Макарова К.С., Чжан Ф., Кунин Е.В. (январь 2017). «SnapShot: системы CRISPR-Cas класса 2» . Клетка . 168 (1–2): 328–328.e1. дои : 10.1016/j.cell.2016.12.038 . ПМИД 28086097 .
- ^ Пол Б., Монтойя Дж. (февраль 2020 г.). «CRISPR-Cas12a: Функциональный обзор и приложения» . Биомедицинский журнал . 43 (1): 8–17. дои : 10.1016/j.bj.2019.10.005 . ПМК 7090318 . ПМИД 32200959 .
- ^ Кокс Д.Б., Гутенберг Дж.С., Абудайе О.О., Франклин Б., Келлнер М.Дж., Йонг Дж. и др. (ноябрь 2017 г.). «Редактирование РНК с помощью CRISPR-Cas13» . Наука . 358 (6366): 1019–1027. Бибкод : 2017Sci...358.1019C . дои : 10.1126/science.aaq0180 . ПМЦ 5793859 . ПМИД 29070703 .
- ^ Перейти обратно: а б Сюй С, Чжоу Ю, Сяо Q, Хэ Б, Гэн Г, Ван Цз и др. (май 2021 г.). «Программируемое редактирование РНК с помощью компактных систем CRISPR-Cas13 из некультивируемых микробов» (PDF) . Природные методы . 18 (5): 499–506. дои : 10.1038/s41592-021-01124-4 . ПМИД 33941935 . S2CID 233719501 . Архивировано (PDF) из оригинала 12 мая 2024 г. Проверено 12 мая 2024 г.
- ^ Азангу-Хьяви М., Гасеми М., Ханали Дж., Бороманд-Сабур М., Джамалха М., Сулеймани М. и др. (2020). «CRISPR/Cas: от редактирования опухолевых генов к Т-клеточной иммунотерапии рака» . Границы в иммунологии . 11 : 2062. дои : 10.3389/fimmu.2020.02062 . ПМЦ 7553049 . ПМИД 33117331 .
- ^ Перейти обратно: а б Алияри Р., Дин С.В. (январь 2009 г.). «Вирусный иммунитет на основе РНК, инициируемый семейством иммунных рецепторов хозяина Dicer» . Иммунологические обзоры . 227 (1): 176–188. дои : 10.1111/j.1600-065X.2008.00722.x . ПМК 2676720 . ПМИД 19120484 .
- ^ Дугар Г., Хербиг А., Фёрстнер К.Ю., Хайдрих Н., Рейнхардт Р., Низельт К. и др. (май 2013 г.). «Карты транскриптома высокого разрешения выявляют штаммоспецифичные регуляторные особенности нескольких изолятов Campylobacter jejuni» . ПЛОС Генетика . 9 (5): e1003495. дои : 10.1371/journal.pgen.1003495 . ПМЦ 3656092 . ПМИД 23696746 .
- ^ Хатум-Аслан А., Манив И., Марраффини Л.А. (декабрь 2011 г.). «Длина зрелых кластеризованных, регулярно расположенных коротких палиндромных повторов РНК (crRNA) измеряется с помощью механизма линейки, закрепленного на сайте процессинга предшественника» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (52): 21218–21222. Бибкод : 2011PNAS..10821218H . дои : 10.1073/pnas.1112832108 . ПМК 3248500 . ПМИД 22160698 .
- ^ Перейти обратно: а б Йосеф I, Горен М.Г., Кимрон У (июль 2012 г.). «Белки и элементы ДНК, необходимые для процесса адаптации CRISPR у Escherichia coli » . Исследования нуклеиновых кислот . 40 (12): 5569–5576. дои : 10.1093/nar/gks216 . ПМЦ 3384332 . ПМИД 22402487 .
- ^ Перейти обратно: а б с д Свартс, округ Колумбия, Мостерд С., ван Пассел М.В., Браунс С.Дж. (2012). «Вмешательство CRISPR направляет приобретение спейсера, специфичного для цепи» . ПЛОС ОДИН . 7 (4): e35888. Бибкод : 2012PLoSO...735888S . дои : 10.1371/journal.pone.0035888 . ПМЦ 3338789 . ПМИД 22558257 .
- ^ Бабу М. , Белоглазова Н., Флик Р., Грэм С., Скарина Т., Ночек Б. и др. (январь 2011 г.). «Двойная функция системы CRISPR-Cas: обеспечение бактериального антивирусного иммунитета и восстановление ДНК» . Молекулярная микробиология . 79 (2): 484–502. дои : 10.1111/j.1365-2958.2010.07465.x . ПМК 3071548 . ПМИД 21219465 .
- ^ Хан Д., Леманн К., Краусс Г. (июнь 2009 г.). «SSO1450 - белок CAS1 из Sulfolobus solfataricus P2 с высоким сродством к РНК и ДНК». Письма ФЭБС . 583 (12): 1928–1932. Бибкод : 2009FEBSL.583.1928H . дои : 10.1016/j.febslet.2009.04.047 . ПМИД 19427858 . S2CID 22279972 .
- ^ Виденхефт Б., Чжоу К., Джинек М., Койл С.М., Ма В., Дудна Дж.А. (июнь 2009 г.). «Структурная основа ДНКазной активности консервативного белка, участвующего в CRISPR-опосредованной защите генома» . Структура . 17 (6): 904–912. дои : 10.1016/j.str.2009.03.019 . ПМИД 19523907 .
- ^ Белоглазова Н., Браун Г., Циммерман М.Д., Праудфут М., Макарова К.С., Кудрицка М. и др. (июль 2008 г.). «Новое семейство специфичных для последовательностей эндорибонуклеаз, связанных с кластеризованными короткими палиндромными повторами с регулярными промежутками» . Журнал биологической химии . 283 (29): 20361–20371. дои : 10.1074/jbc.M803225200 . ПМЦ 2459268 . ПМИД 18482976 .
- ^ Самай П., Смит П., Шуман С. (декабрь 2010 г.). «Структура CRISPR-ассоциированного белка Cas2 из Desulfovibrio vulgaris» . Acta Crystallographica Раздел F. 66 (Часть 12): 1552–1556. дои : 10.1107/S1744309110039801 . ПМК 2998353 . ПМИД 21139194 .
- ^ Нам К.Х., Дин Ф., Хайтджема С., Хуан К., ДеЛиса М.П., Ке А. (октябрь 2012 г.). «Двухцепочечная эндонуклеазная активность в белке Cas2, связанном с регулярно расположенными короткими палиндромными повторами (CRISPR) Bacillus halodurans» . Журнал биологической химии . 287 (43): 35943–35952. дои : 10.1074/jbc.M112.382598 . ПМЦ 3476262 . ПМИД 22942283 .
- ^ Перейти обратно: а б Нуньес Дж.К., Кранцуш П.Дж., Ноеске Дж., Райт А.В., Дэвис К.В., Дудна Дж.А. (июнь 2014 г.). «Образование комплекса Cas1-Cas2 опосредует приобретение спейсера во время адаптивного иммунитета CRISPR-Cas» . Структурная и молекулярная биология природы . 21 (6): 528–534. дои : 10.1038/nsmb.2820 . ПМК 4075942 . ПМИД 24793649 .
- ^ Нуньес Дж.К., Ли А.С., Энгельман А., Дудна Х.А. (март 2015 г.). «Приобретение спейсера, опосредованное интегразой, во время адаптивного иммунитета CRISPR-Cas» . Природа . 519 (7542): 193–198. Бибкод : 2015Natur.519..193N . дои : 10.1038/nature14237 . ПМК 4359072 . ПМИД 25707795 .
- ^ Ван Дж., Ли Дж., Чжао Х., Шэн Г., Ван М., Инь М. и др. (ноябрь 2015 г.). «Структурные и механистические основы PAM-зависимого приобретения спейсера в системах CRISPR-Cas» . Клетка . 163 (4): 840–853. дои : 10.1016/j.cell.2015.10.008 . ПМИД 26478180 .
- ^ Нуньес Дж.К., Харрингтон Л.Б., Кранцуш П.Дж., Энгельман А.Н., Дудна Х.А. (ноябрь 2015 г.). «Захват чужеродной ДНК во время адаптивного иммунитета CRISPR-Cas» . Природа . 527 (7579): 535–538. Бибкод : 2015Natur.527..535N . дои : 10.1038/nature15760 . ПМЦ 4662619 . ПМИД 26503043 .
- ^ Сорек Р., Лоуренс К.М., Виденхефт Б. (2013). «CRISPR-опосредованная адаптивная иммунная система у бактерий и архей» . Ежегодный обзор биохимии . 82 (1): 237–266. doi : 10.1146/annurev-biochem-072911-172315 . ПМИД 23495939 .
- ^ Нуньес Дж.К., Бай Л., Харрингтон Л.Б., Хиндер Т.Л., Дудна Х.А. (июнь 2016 г.). «Иммунологическая память CRISPR требует фактора хозяина для специфичности» . Молекулярная клетка . 62 (6): 824–833. doi : 10.1016/j.molcel.2016.04.027 . ПМИД 27211867 .
- ^ Фагерлунд Р.Д., Уилкинсон М.Е., Клыков О., Барендрегт А., Пирс Ф.Г., Кипер С.Н. и др. (июнь 2017 г.). «Захват и интеграция спейсера адаптационным комплексом CRISPR Cas1-Cas2–3 типа IF» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (26): Е5122–Е5128. Бибкод : 2017PNAS..114E5122F . дои : 10.1073/pnas.1618421114 . ПМЦ 5495228 . ПМИД 28611213 .
- ^ Ролли С., Грэм С., Руйон С., Уайт М.Ф. (февраль 2018 г.). «Обработка преспейсеров и специальная интеграция в систему CRISPR типа IA» . Исследования нуклеиновых кислот . 46 (3): 1007–1020. дои : 10.1093/нар/gkx1232 . ПМК 5815122 . ПМИД 29228332 .
- ^ Перейти обратно: а б с д Хорват П., Ромеро Д.А., Куте-Монвуазен А.С., Ричардс М., Дево Х., Муано С. и др. (февраль 2008 г.). «Разнообразие, активность и эволюция локусов CRISPR у Streptococcus thermophilus » . Журнал бактериологии . 190 (4): 1401–1412. дои : 10.1128/JB.01415-07 . ПМК 2238196 . ПМИД 18065539 .
- ^ Перейти обратно: а б с Дево Х., Баррангу Р. , Гарно Дж.Э., Лабонте Дж., Фремо С., Бояваль П. и др. (февраль 2008 г.). «Фаговый ответ на устойчивость, кодируемую CRISPR, у Streptococcus thermophilus » . Журнал бактериологии . 190 (4): 1390–1400. дои : 10.1128/JB.01412-07 . ПМК 2238228 . ПМИД 18065545 .
- ^ Мохика Ф.Д., Диес-Вильясеньор К., Гарсиа-Мартинес Х., Альмендрос К. (март 2009 г.). «Короткие последовательности мотивов определяют цели прокариотической защитной системы CRISPR» . Микробиология . 155 (Часть 3): 733–740. дои : 10.1099/mic.0.023960-0 . ПМИД 19246744 .
- ^ Перейти обратно: а б Лиллестёл Р.К., Шах С.А., Брюггер К., Реддер П., Фан Х., Кристиансен Дж. и др. (апрель 2009 г.). «Семейства CRISPR рода Sulfolobus кренархей: двунаправленная транскрипция и динамические свойства». Молекулярная микробиология . 72 (1): 259–272. дои : 10.1111/j.1365-2958.2009.06641.x . ПМИД 19239620 . S2CID 36258923 .
- ^ Перейти обратно: а б Шах С.А., Хансен Н.Р., Гарретт Р.А. (февраль 2009 г.). «Распределение спейсерных совпадений CRISPR в вирусах и плазмидах кренархейных ацидотермофилов и значение их ингибирующего механизма». Труды Биохимического общества . 37 (Часть 1): 23–28. дои : 10.1042/BST0370023 . ПМИД 19143596 . S2CID 19093261 .
- ^ Перейти обратно: а б Диес-Вильясеньор К., Гусман Н.М., Альмендрос К., Гарсиа-Мартинес Х., Мохика Ф.Дж. (май 2013 г.). «Репортерные плазмиды интеграции CRISPR-спейсера обнаруживают различные подлинные особенности приобретения среди вариантов CRISPR-Cas IE Escherichia coli » . Биология РНК . 10 (5): 792–802. дои : 10.4161/rna.24023 . ПМЦ 3737337 . ПМИД 23445770 .
- ^ Перейти обратно: а б с Эрдманн С., Гаррет Р.А. (сентябрь 2012 г.). «Избирательное и гиперактивное поглощение чужеродной ДНК адаптивной иммунной системой археи посредством двух различных механизмов» . Молекулярная микробиология . 85 (6): 1044–1056. дои : 10.1111/j.1365-2958.2012.08171.x . ПМЦ 3468723 . ПМИД 22834906 .
- ^ Перейти обратно: а б Шах С.А., Эрдманн С., Мохика Ф.Дж., Гаррет Р.А. (май 2013 г.). «Мотивы узнавания протоспейсеров: смешанные идентичности и функциональное разнообразие» . Биология РНК . 10 (5): 891–899. дои : 10.4161/rna.23764 . ПМЦ 3737346 . ПМИД 23403393 .
- ^ Андерссон А.Ф., Банфилд Дж.Ф. (май 2008 г.). «Динамика популяций вирусов и приобретенная устойчивость к вирусам в природных микробных сообществах». Наука . 320 (5879): 1047–1050. Бибкод : 2008Sci...320.1047A . дои : 10.1126/science.1157358 . ПМИД 18497291 . S2CID 26209623 .
- ^ Перейти обратно: а б с д Pride DT, Sun CL, Salzman J, Rao N, Loomer P, Armitage GC и др. (январь 2011 г.). «Анализ стрептококковых CRISPR из слюны человека выявляет существенное разнообразие последовательностей внутри и между субъектами с течением времени» . Геномные исследования . 21 (1): 126–136. дои : 10.1101/гр.111732.110 . ПМК 3012920 . ПМИД 21149389 .
- ^ Перейти обратно: а б Горен М.Г., Йосеф I, Остер О, Кимрон У (октябрь 2012 г.). «Экспериментальное определение сгруппированных коротких палиндромных дупликонов с регулярными промежутками в Escherichia coli ». Журнал молекулярной биологии . 423 (1): 14–16. дои : 10.1016/j.jmb.2012.06.037 . ПМИД 22771574 .
- ^ Перейти обратно: а б с Даценко К.А., Пугач К., Тихонов А., Ваннер Б.Л., Северинов К., Семенова Е. (июль 2012 г.). «Молекулярная память о предшествующих инфекциях активирует систему адаптивного бактериального иммунитета CRISPR/Cas» . Природные коммуникации . 3 : 945. Бибкод : 2012NatCo...3..945D . дои : 10.1038/ncomms1937 . ПМИД 22781758 .
- ^ Геснер Э.М., Шелленберг М.Дж., Гарсайд Э.Л., Джордж М.М., Макмиллан А.М. (июнь 2011 г.). «Распознавание и созревание эффекторных РНК в пути вмешательства CRISPR». Структурная и молекулярная биология природы . 18 (6): 688–692. дои : 10.1038/nsmb.2042 . ПМИД 21572444 . S2CID 677704 .
- ^ Сашитал Д.Г., Джинек М., Дудна Дж.А. (июнь 2011 г.). «Вызванное РНК конформационное изменение, необходимое для расщепления РНК CRISPR эндорибонуклеазой Cse3». Структурная и молекулярная биология природы . 18 (6): 680–687. дои : 10.1038/nsmb.2043 . ПМИД 21572442 . S2CID 5538195 .
- ^ Хаурвиц Р.Э., Джинек М., Виденхефт Б., Чжоу К., Дудна Дж.А. (сентябрь 2010 г.). «Обработка РНК, специфичная для последовательности и структуры, эндонуклеазой CRISPR» . Наука . 329 (5997): 1355–1358. Бибкод : 2010Sci...329.1355H . дои : 10.1126/science.1192272 . ПМК 3133607 . ПМИД 20829488 .
- ^ Перейти обратно: а б Кунин В., Сорек Р., Гугенхольц П. (2007). «Эволюционная консервативность последовательности и вторичных структур в повторах CRISPR» . Геномная биология . 8 (4): С61. дои : 10.1186/gb-2007-8-4-r61 . ПМК 1896005 . ПМИД 17442114 .
- ^ Карт Дж., Ван Р., Ли Х., Тернс Р.М., Тернс MP (декабрь 2008 г.). «Cas6 — это эндорибонуклеаза, которая генерирует направляющие РНК для защиты от захватчиков у прокариот» . Гены и развитие . 22 (24): 3489–3496. дои : 10.1101/gad.1742908 . ПМК 2607076 . ПМИД 19141480 .
- ^ Ван Р., Преамплюм Г., Тернс М.П., Тернс Р.М., Ли Х (февраль 2011 г.). «Взаимодействие рибоэндонуклеазы Cas6 с РНК CRISPR: распознавание и расщепление» . Структура . 19 (2): 257–264. дои : 10.1016/j.str.2010.11.014 . ПМК 3154685 . ПМИД 21300293 .
- ^ Нивонер О., Джинек М., Дудна Дж.А. (январь 2014 г.). «Эволюция распознавания и обработки РНК CRISPR эндонуклеазами Cas6» . Исследования нуклеиновых кислот . 42 (2): 1341–1353. дои : 10.1093/нар/gkt922 . ПМК 3902920 . ПМИД 24150936 .
- ^ Семенова Е., Жоре М.М., Даценко К.А., Семенова А., Вестра Э.Р., Ваннер Б. и др. (июнь 2011 г.). «Вмешательство кластеризованных коротких палиндромных повторов (CRISPR) РНК с регулярными промежутками регулируется затравочной последовательностью» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (25): 10098–10103. Бибкод : 2011PNAS..10810098S . дои : 10.1073/pnas.1104144108 . ПМК 3121866 . ПМИД 21646539 .
- ^ Гудбергсдоттир С., Дэн Л., Чен З., Дженсен Дж.В., Дженсен Л.Р., Ше К. и др. (январь 2011 г.). «Динамические свойства систем Sulfolobus CRISPR/Cas и CRISPR/Cmr при воздействии трансмиссивных вирусных и плазмидных генов и протоспейсеров» . Молекулярная микробиология . 79 (1): 35–49. дои : 10.1111/j.1365-2958.2010.07452.x . ПМК 3025118 . ПМИД 21166892 .
- ^ Маника А., Зебек З., Тайхманн Д., Шлепер С. (апрель 2011 г.). «In vivo активность CRISPR-опосредованной вирусной защиты у гипертермофильных архей» . Молекулярная микробиология . 80 (2): 481–491. дои : 10.1111/j.1365-2958.2011.07586.x . ПМИД 21385233 . S2CID 41442419 .
- ^ Джоре М.М., Лундгрен М., ван Дуйн Э., Бултема Дж.Б., Вестра Э.Р., Вагмаре С.П. и др. (май 2011 г.). «Структурная основа распознавания ДНК с помощью CRISPR-РНК с помощью Cascade» (PDF) . Структурная и молекулярная биология природы . 18 (5): 529–536. дои : 10.1038/nsmb.2019 . ПМИД 21460843 . S2CID 10987554 . Архивировано (PDF) из оригинала 23 октября 2021 г. Проверено 30 июня 2019 г.
- ^ Виденхефт Б., Ландер Г.К., Чжоу К., Джор М.М., Браунс С.Дж., ван дер Ост Дж. и др. (сентябрь 2011 г.). «Структуры РНК-ориентированного надзорного комплекса бактериальной иммунной системы» . Природа . 477 (7365): 486–489. Бибкод : 2011Natur.477..486W . дои : 10.1038/nature10402 . ПМК 4165517 . ПМИД 21938068 .
- ^ Чжан Дж., Руйон С., Керу М., Рикс Дж., Брюггер К., Грэм С. и др. (февраль 2012 г.). «Структура и механизм комплекса CMR для CRISPR-опосредованного противовирусного иммунитета» . Молекулярная клетка . 45 (3): 303–313. doi : 10.1016/j.molcel.2011.12.013 . ПМЦ 3381847 . ПМИД 22227115 .
- ^ Перейти обратно: а б Хейл С.Р., Чжао П., Олсон С., Дафф М.О., Грейвли Б.Р., Уэллс Л. и др. (ноябрь 2009 г.). «Расщепление РНК под контролем РНК белковым комплексом CRISPR РНК-Cas» . Клетка . 139 (5): 945–956. дои : 10.1016/j.cell.2009.07.040 . ПМЦ 2951265 . ПМИД 19945378 .
- ^ Эстрелла М.А., Куо Ф.Т., Бэйли С. (2016). «РНК-активируемое расщепление ДНК эффекторным комплексом CRISPR-Cas типа III-B» . Гены и развитие . 30 (4): 460–470. дои : 10.1101/gad.273722.115 . ПМЦ 4762430 . ПМИД 26848046 .
- ^ Самай П., Пьенсон Н. , Цзян В., Голдберг Г.В., Хатум-Аслан А., Марраффини Л.А. (2015). «Котранскрипционное расщепление ДНК и РНК во время иммунитета CRISPR-Cas типа III» . Клетка . 161 (5): 1164–1174. дои : 10.1016/j.cell.2015.04.027 . ПМЦ 4594840 . ПМИД 25959775 .
- ^ Перейти обратно: а б с Марраффини Л.А., Сонтхаймер Э.Дж. (январь 2010 г.). «Дискриминация себя и чужого во время иммунитета, направленного на РНК CRISPR» . Природа . 463 (7280): 568–571. Бибкод : 2010Natur.463..568M . дои : 10.1038/nature08703 . ПМЦ 2813891 . ПМИД 20072129 .
- ^ Крупович М., Беген П., Кунин Е.В. (август 2017 г.). «Каспозоны: мобильные генетические элементы, породившие механизм адаптации CRISPR-Cas» . Современное мнение в микробиологии . 38 : 36–43. дои : 10.1016/j.mib.2017.04.004 . ПМЦ 5665730 . ПМИД 28472712 .
- ^ Кунин Е.В. , Макарова К.С. (май 2013 г.). «CRISPR-Cas: эволюция системы адаптивного иммунитета на основе РНК у прокариот» . Биология РНК . 10 (5): 679–686. дои : 10.4161/rna.24022 . ПМЦ 3737325 . ПМИД 23439366 .
- ^ Кунин Е.В. , Макарова К.С., Чжан Ф (июнь 2017). «Разнообразие, классификация и эволюция систем CRISPR-Cas» . Современное мнение в микробиологии . 37 : 67–78. дои : 10.1016/j.mib.2017.05.008 . ПМЦ 5776717 . ПМИД 28605718 .
- ^ Перейти обратно: а б Шмаков С., Смаргон А., Скотт Д., Кокс Д., Пызоча Н., Ян В. и др. (март 2017 г.). «Разнообразие и эволюция систем CRISPR-Cas класса 2» . Обзоры природы. Микробиология . 15 (3): 169–182. дои : 10.1038/nrmicro.2016.184 . ПМЦ 5851899 . ПМИД 28111461 .
- ^ Купчок А., Болбак Дж.П. (февраль 2013 г.). «Вероятностные модели эволюции содержания спейсера CRISPR» . Эволюционная биология BMC . 13 (1): 54. Бибкод : 2013BMCEE..13...54K . дои : 10.1186/1471-2148-13-54 . ПМК 3704272 . ПМИД 23442002 .
- ^ Штернберг С.Х., Рихтер Х., Шарпантье Э., Кимрон У. (март 2016 г.). «Адаптация в системах CRISPR-Cas». Молекулярная клетка . 61 (6): 797–808. doi : 10.1016/j.molcel.2016.01.030 . hdl : 21.11116/0000-0003-E74E-2 . ПМИД 26949040 .
- ^ Кунин Е.В. , Вольф Ю.И. (ноябрь 2009 г.). «Является ли эволюция дарвиновской или/и ламаркистской?» . Биология Директ . 4:42 . дои : 10.1186/1745-6150-4-42 . ПМЦ 2781790 . ПМИД 19906303 .
- ^ Вайс А. (октябрь 2015 г.). «Ламарковские иллюзии» . Тенденции в экологии и эволюции . 30 (10): 566–568. Бибкод : 2015TEcoE..30..566W . дои : 10.1016/j.tree.2015.08.003 . ПМИД 26411613 .
- ^ Перейти обратно: а б Кунин Е.В. , Вольф Ю.И. (февраль 2016). «Насколько ламаркианский иммунитет CRISPR-Cas: континуум механизмов развития» . Биология Директ . 11 (1): 9. дои : 10.1186/s13062-016-0111-z . ПМК 4765028 . ПМИД 26912144 .
- ^ Гейдельберг Дж. Ф., Нельсон В. К., Шенфельд Т., Бхайя Д. (2009). Ахмед Н. (ред.). «Микробная война в сообществе микробных матов: CRISPR дают представление о совместной эволюции геномов хозяина и вируса» . ПЛОС ОДИН . 4 (1): е4169. Бибкод : 2009PLoSO...4.4169H . дои : 10.1371/journal.pone.0004169 . ПМЦ 2612747 . ПМИД 19132092 .
- ^ Touchon M, Rocha EP (июнь 2010 г.). Рандау Л. (ред.). «Маленькие, медленные и специализированные CRISPR и анти-CRISPR эшерихий и сальмонелл» . ПЛОС ОДИН . 5 (6): е11126. Бибкод : 2010PLoSO...511126T . дои : 10.1371/journal.pone.0011126 . ПМК 2886076 . ПМИД 20559554 .
- ^ Перейти обратно: а б с Ро М., Ву Ю.В., Тан Х., Доак Т.Г., Йе Ю. (2012). «Разнообразные CRISPR развиваются в микробиомах человека» . ПЛОС Генетика . 8 (6): e1002441. дои : 10.1371/journal.pgen.1002441 . ПМЦ 3374615 . ПМИД 22719260 .
- ^ Перейти обратно: а б Сан КЛ, Баррангу Р. , Томас БК, Хорват П., Фремо К., Банфилд Дж.Ф. (февраль 2013 г.). «Фаговые мутации в ответ на диверсификацию CRISPR в бактериальной популяции». Экологическая микробиология . 15 (2): 463–470. Бибкод : 2013EnvMi..15..463S . дои : 10.1111/j.1462-2920.2012.02879.x . ПМИД 23057534 .
- ^ Куно С., Сако Ю., Ёсида Т. (май 2014 г.). «Диверсификация CRISPR в пределах сосуществующих генотипов в естественной популяции цветущей цианобактерии Microcystis aeruginosa» . Микробиология . 160 (Часть 5): 903–916. дои : 10.1099/mic.0.073494-0 . ПМИД 24586036 .
- ^ Сорек Р., Кунин В., Гугенхольц П. (март 2008 г.). «CRISPR — широко распространенная система, обеспечивающая приобретенную устойчивость бактерий и архей к фагам» . Обзоры природы. Микробиология . 6 (3): 181–186. дои : 10.1038/nrmicro1793 . ПМИД 18157154 . S2CID 3538077 . Архивировано из оригинала 8 марта 2021 г. Проверено 25 ноября 2019 г.
Таблица 1. Веб-ресурсы для анализа CRISPR
- ^ Прайд Д.Т., Зальцман Дж., Релман Д.А. (сентябрь 2012 г.). «Сравнение кластеризованных коротких палиндромных повторов с регулярными промежутками и виромов в слюне человека выявляет бактериальную адаптацию к слюнным вирусам» . Экологическая микробиология . 14 (9): 2564–2576. Бибкод : 2012EnvMi..14.2564P . дои : 10.1111/j.1462-2920.2012.02775.x . ПМЦ 3424356 . ПМИД 22583485 .
- ^ Состоялось в Нидерландах, Эррера А., Уитакер Р.Дж. (ноябрь 2013 г.). «Реассортация локусов повторов-спейсеров CRISPR у Sulfolobus Islandicus». Экологическая микробиология . 15 (11): 3065–3076. Бибкод : 2013EnvMi..15.3065H . дои : 10.1111/1462-2920.12146 . ПМИД 23701169 .
- ^ Состоялось в Нидерландах, Эррера А., Кадилло-Кирос Х., Уитакер Р.Дж. (сентябрь 2010 г.). «Разнообразие, связанное с CRISPR, в популяции Sulfolobus Islandicus» . ПЛОС ОДИН . 5 (9): e12988. Бибкод : 2010PLoSO...512988H . дои : 10.1371/journal.pone.0012988 . ПМЦ 2946923 . ПМИД 20927396 .
- ^ Перейти обратно: а б с Медведева С., Лю Ю., Кунин Е.В., Северинов К., Прангишвили Д., Крупович М. (ноябрь 2019 г.). «Массивы мини-CRISPR, переносимые вирусами, участвуют в межвирусных конфликтах» . Природные коммуникации . 10 (1): 5204. Бибкод : 2019NatCo..10.5204M . дои : 10.1038/s41467-019-13205-2 . ПМК 6858448 . ПМИД 31729390 .
- ^ Скеннертон, Коннектикут, Имелфорт М, Тайсон Г.В. (май 2013 г.). «Crass: идентификация и реконструкция CRISPR на основе несобранных метагеномных данных» . Исследования нуклеиновых кислот . 41 (10): е105. дои : 10.1093/нар/gkt183 . ПМЦ 3664793 . ПМИД 23511966 .
- ^ Стерн А., Мик Э., Тирош И., Сэги О., Сорек Р. (октябрь 2012 г.). «Нацеливание на CRISPR выявляет резервуар распространенных фагов, связанных с микробиомом кишечника человека» . Геномные исследования . 22 (10): 1985–1994. дои : 10.1101/гр.138297.112 . ПМК 3460193 . ПМИД 22732228 .
- ^ Новик Р.П., Кристи Дж.Э., Пенадес-младший (август 2010 г.). «Фагосвязанные хромосомные острова грамположительных бактерий» . Обзоры природы Микробиология . 8 (8): 541–551. дои : 10.1038/nrmicro2393 . ПМЦ 3522866 . ПМИД 20634809 .
- ^ Рам Дж., Чен Дж., Кумар К., Росс Х.Ф., Убеда С., Дамле П.К. и др. (октябрь 2012 г.). «Вмешательство острова патогенности стафилококка в репродукцию фагов-помощников является парадигмой молекулярного паразитизма» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (40): 16300–16305. Бибкод : 2012PNAS..10916300R . дои : 10.1073/pnas.1204615109 . ПМЦ 3479557 . ПМИД 22991467 .
- ^ Рам Дж., Чен Дж., Росс Х.Ф., Новик Р.П. (октябрь 2014 г.). «Точно модулированное вмешательство острова патогенности в транскрипцию генов позднего фага» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (40): 14536–14541. Бибкод : 2014PNAS..11114536R . дои : 10.1073/pnas.1406749111 . ПМК 4209980 . ПМИД 25246539 .
- ^ Сид К.Д., Лазински Д.В., Колдервуд С.Б., Камилли А. (февраль 2013 г.). «Бактериофаг кодирует собственный адаптивный ответ CRISPR/Cas, чтобы избежать врожденного иммунитета хозяина» . Природа . 494 (7438): 489–491. Бибкод : 2013Natur.494..489S . дои : 10.1038/nature11927 . ПМЦ 3587790 . ПМИД 23446421 .
- ^ Бойд С.М., Ангермейер А., Хейс С.Г., Барт З.К., Патель К.М., Сид К.Д. (сентябрь 2021 г.). «Бактериофаг ICP1: стойкий хищник холерного вибриона » . Ежегодный обзор вирусологии . 8 (1): 285–304. doi : 10.1146/annurev-virology-091919-072020 . ISSN 2327-056X . ПМК 9040626 . ПМИД 34314595 .
- ^ Фэн Ц, Ли Ц, Чжоу Х, Ван З, Линь С, Цзян Цз и др. (август 2024 г.). «Технология CRISPR в болезнях человека» . Медкомм . 5 (8): е672. дои : 10.1002/mco2.672 . ПМЦ 11286548 . ПМИД 39081515 .
- ^ Ли Т, Ли С, Кан Ю, Чжоу Дж, И М (август 2024 г.). «Использование развивающегося CRISPR/Cas9 для точной онкологии» . Журнал трансляционной медицины . 22 (1): 749. doi : 10.1186/s12967-024-05570-4 . ПМЦ 11312220 . ПМИД 39118151 .
- ^ Мишра С., Наяк С., Тутеджа Н., Пусапати С., Суэйн Д.М., Саху Р.К. (июль 2024 г.). «CRISPR/Cas-опосредованная геномная инженерия растений: применение и перспективы» . Растения . 13 (14). Базель, Швейцария: 1884 г. doi : 10.3390/plants13141884 . ПМЦ 11279650 . ПМИД 39065411 .
- ^ Каур Р., Гупта С., Чаухан А., Мишра В., Шарма М.К., Сингх Дж. (август 2024 г.). «Использование возможностей микрофлюидики на основе кластеризованных коротких палиндромных повторов с регулярными интервалами (CRISPR) для молекулярной диагностики следующего поколения». Отчеты по молекулярной биологии . 51 (1): 896. doi : 10.1007/s11033-024-09840-8 . ПМИД 39115550 .
- ^ Шинвари З.К., Танвир Ф., Халил А.Т. (2018). «Этические проблемы редактирования генома с помощью CRISPR» . Актуальные проблемы молекулярной биологии . 26 : 103–110. дои : 10.21775/cimb.026.103 . ПМИД 28879860 .
- ^ Козан Д.В., Фарбер С.А. (февраль 2024 г.). «Разумно ли когда-нибудь редактировать аллели дикого типа? Сконструированные аллели CRISPR по сравнению с миллионами лет эволюции человека». Атеросклероз, тромбоз и сосудистая биология . 44 (2): 328–333. дои : 10.1161/ATVBAHA.123.318069 . ПМК 10948015. ПМИД 38059350 .
Дальнейшее чтение
[ редактировать ]- Дудна Дж. , Мали П. (23 марта 2016 г.). CRISPR-Cas: Лабораторное руководство . Нью-Йорк: Лабораторное издательство Колд-Спринг-Харбор. ISBN 978-1-62182-131-1 .
- Моханраджу П., Макарова К.С., Зетше Б., Чжан Ф. , Кунин Е.В. , ван дер Ост Дж. (август 2016 г.). «Различные эволюционные корни и механистические вариации систем CRISPR-Cas» (PDF) . Наука . 353 (6299): аад5147. дои : 10.1126/science.aad5147 . hdl : 1721.1/113195 . ПМИД 27493190 . S2CID 11086282 . Архивировано из оригинала 23 октября 2021 г. Проверено 29 августа 2019 г.
- Сандер Дж.Д., Йонг Дж.К. (апрель 2014 г.). «Системы CRISPR-Cas для редактирования, регулирования и нацеливания геномов» . Природная биотехнология . 32 (4): 347–355. дои : 10.1038/nbt.2842 . ПМК 4022601 . ПМИД 24584096 .
- Slaymaker IM, Гао Л., Зетше Б., Скотт Д.А., Ян В.С., Чжан Ф. (январь 2016 г.). «Рационально сконструированные нуклеазы Cas9 с улучшенной специфичностью» . Наука . 351 (6268): 84–88. Бибкод : 2016Sci...351...84S . дои : 10.1126/science.aad5227 . ПМЦ 4714946 . ПМИД 26628643 .
- Тернс Р.М., Тернс МП (март 2014 г.). «Технологии на основе CRISPR: перепрофилирование прокариотического защитного оружия» . Тенденции в генетике . 30 (3): 111–118. дои : 10.1016/j.tig.2014.01.003 . ПМЦ 3981743 . ПМИД 24555991 .
- Вестра Э.Р., Баклинг А., Финеран ПК (май 2014 г.). «Системы CRISPR-Cas: за пределами адаптивного иммунитета». Обзоры природы Микробиология . 12 (5): 317–326. дои : 10.1038/nrmicro3241 . ПМИД 24704746 . S2CID 36575361 .
- Андерссон А.Ф., Банфилд Дж.Ф. (май 2008 г.). «Динамика популяций вирусов и приобретенная устойчивость к вирусам в природных микробных сообществах». Наука . 320 (5879): 1047–1050. Бибкод : 2008Sci...320.1047A . дои : 10.1126/science.1157358 . ПМИД 18497291 . S2CID 26209623 .
- Хейл С., Клеппе К., Тернс Р.М., Тернс член парламента (декабрь 2008 г.). «Прокариотические молчащие (пси)РНК у Pyrococcus Furiosus» . РНК . 14 (12): 2572–2579. дои : 10.1261/rna.1246808 . ПМК 2590957 . ПМИД 18971321 .
- ван дер Плог-младший (июнь 2009 г.). «Анализ CRISPR у Streptococcus mutans предполагает частое возникновение приобретенного иммунитета против заражения M102-подобными бактериофагами» (PDF) . Микробиология . 155 (Часть 6): 1966–1976. дои : 10.1099/mic.0.027508-0 . ПМИД 19383692 .
- ван дер Ост Дж., Браунс С.Дж. (ноябрь 2009 г.). «RNAi: в дело вступают прокариоты» . Клетка . 139 (5): 863–865. дои : 10.1016/j.cell.2009.11.018 . ПМИД 19945373 . S2CID 11863610 .
- Каргинов Ф.В., Хэннон Г.Дж. (январь 2010 г.). «Система CRISPR: защита, управляемая малой РНК, у бактерий и архей» . Молекулярная клетка . 37 (1): 7–19. doi : 10.1016/j.molcel.2009.12.033 . ПМК 2819186 . ПМИД 20129051 .
- Пул У, Вурм Р., Арслан З., Гейссен Р., Хофманн Н., Вагнер Р. (март 2010 г.). «Идентификация и характеристика промоторов CRISPR-cas E. coli и их подавление с помощью H-NS» . Молекулярная микробиология . 75 (6): 1495–1512. дои : 10.1111/j.1365-2958.2010.07073.x . ПМИД 20132443 . S2CID 37529215 .
- Диес-Вильясеньор К., Альмендрос К., Гарсиа-Мартинес Х., Мохика Ф.Д. (май 2010 г.). «Разнообразие локусов CRISPR в Escherichia coli » . Микробиология . 156 (Часть 5): 1351–1361. дои : 10.1099/mic.0.036046-0 . ПМИД 20133361 .
- Дево Х., Гарно Ж.Э., Муано С. (2010). «Система CRISPR/Cas и ее роль во взаимодействии фагов и бактерий». Ежегодный обзор микробиологии . 64 : 475–493. дои : 10.1146/annurev.micro.112408.134123 . ПМИД 20528693 .
- Кунин Е.В. , Макарова К.С. (декабрь 2009 г.). «CRISPR-Cas: система адаптивного иммунитета у прокариот» . Отчеты о биологии F1000 . 1 : 95. дои : 10.3410/B1-95 . ПМЦ 2884157 . ПМИД 20556198 .
- «Эпоха красного пера» . Экономист . 22 августа 2015. ISSN 0013-0613 . Архивировано из оригинала 24 августа 2015 г. Проверено 25 августа 2015 г.
- Ран А.Ф., Сюй П.Д., Райт Дж., Агарвала В., Скотт Д.А., Чжан Ф. (2013). «Геномная инженерия с использованием системы CRISPR-Cas9» . Протоколы природы . 8 (11): 2281–2308. дои : 10.1038/nprot.2013.143 . ПМЦ 3969860 . ПМИД 24157548 .
Внешние ссылки
[ редактировать ]- «Расширенное редактирование генов: CRISPR-Cas9» (PDF) . Исследовательская служба Конгресса .
- «Доклад Дженнифер Дудна: Геномная инженерия с CRISPR-Cas9: рождение революционной технологии» . 10 сентября 2022 г.
- «Человеческая природа» . НОВА . 47 сезон. 9 серия. 9 сентября 2020. PBS . ВГБХ . Проверено 7 апреля 2023 г.
Банк данных по белкам
[ редактировать ]- Обзор всей структурной информации, доступной в PDB для UniProt : Q46901 (каскадный субблок CasA системы CRISPR) в PDBe-KB .
- Обзор всей структурной информации, доступной в PDB для UniProt : P76632 (каскадный субблок CasB системы CRISPR) в PDBe-KB .
- Обзор всей структурной информации, доступной в PDB для UniProt : Q46899 (каскадный субблок CasC системы CRISPR) в PDBe-KB .
- Обзор всей структурной информации, доступной в PDB для UniProt : Q46898 (каскадный субблок CasD системы CRISPR) в PDBe-KB .
- Обзор всей структурной информации, доступной в PDB для UniProt : Q46897 (каскадный субблок CasE системы CRISPR) в PDBe-KB .