Сетка
| Сетка Временный диапазон: ранний меховой
~ С 125 до 120 млн. - поздний | |
|---|---|

| |
| Гипотетическая реконструкция жизни трех Kinnareemimus | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Клада : | Динозаурия |
| Клада : | Сауриския |
| Клада : | Теропода |
| Клада : | Maniraptoriformes |
| Клада : | † Орнитомимозаврия |
| Род: | † Kinnareemimus Buffetaut et al. , 2009 |
| Тип видов | |
| † Kinnareemimus honkaenensis Buffetaut et al. , 2009
| |
| Синонимы | |
| |
Kinnareemimus ( / ˌ k ɪ n ə r i ˈ m ː i m ə s / , означающего « мимик Kinnaree », после фигуры из тайского фольклора) является родом орнитомимозаврного , динозавра который вымершим был обнаружен в раннем цвете в том, что сейчас есть в Таиланде . Род содержит только типовые виды , K. Khonkaenensis . Конкретный эпитет - провинция Хон Каен , которая находится в северо -восточном Таиланде, где были обнаружены останки животного. [ 1 ]
Открытие и именование
[ редактировать ]
орнитомимозавра Оскокающие окаменелости и ихнофоссили были зарегистрированы на Таиланде в плато Хорат еще в 1997 году в публикации Эрика Баффета и его коллег. В то время только два рода динозавров, Siamotyrannus и Phuwiangosaurus были названы из страны. Их доклад включал в себя рисунок плюсневых средств орнитомимозавра, который был обнаружен в местности под названием «Phu Wiang 5». Эти плюсневые кости явно показали производное арктометатасаловое состояние, аналогичное орнитомимозаврам, известным из Ларамидии . Однако этот образец не был идентифицирован ни одним именем, кроме «тайского орнитомимозавра». [ 2 ]
Следующее упоминание о «тайском орнитомимозавре» [ 3 ] был в первом выпуске журнала Dinosaur Press, который был опубликован в Японии в 2000 году. [ 3 ] В этой публикации таксон упоминается Nomen Nudum "Ginnareemimus" Рюичи Канеко, который написал статью в журнале, в которой была недавняя информация о палеонтологии в Таиланде . [ 4 ] [ 5 ]
Kinnareemimus получил формальное описание видов в 2009 году , когда Эрик Баффета, Варавуд Сютетерн и Хайян Тонг опубликовал анатомическую монографию в Бюллетене Геологического общества Лондона . Все кости, относящиеся к орнитомимозаврам, обнаруженным в Phu Wiang 5, были рассеяны, и поэтому невозможно точно знать, сколько отдельных животных были окаменели на месте. По этой причине, когда Kinnareemimus был описан в 2009 году, только один частичный мегапотисто III-обозначенный PW5A-100-был обозначен как голотип . Оставшиеся образцы получали уникальные обозначения (от 101 до 131), и каждый из них был направлен на новый род. В их описании замечание Buffetaut и Suteethorn, что они уверены, что весь материал принадлежит одному виду, но они консервативно не включали анатомию этих костей в диагностике видов. [ 1 ]
Когда было описано, авторы назвали род после Киннари , существо от фольклора тайского народа , у которого есть голова и торс женщины и ноги птицы и живет в легендарном лесу Химмапан. Buffetaut и Suteethorn отмечают, что выбор Kinnaree был в отношении птичьих ног Kinnareemimus . [ 1 ]
Описание
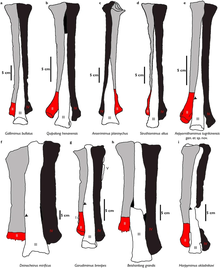
[ редактировать ]Поскольку голотип Kinnareemimus ) , является настолько фрагментарным (состоящий только из одного плюсневого оценки размера животного всегда были очень спекулятивными и неопределенными. Авторы, которые описали род, Эрик Баффета и его коллеги, не дали оценки массы животного. Они отметили, что соотношение длины между голени и третьей плюсневой кости Киннаримимуса наиболее похоже на Гарудимимус . Они интерпретируют это как означают, что его пропорции тела могли быть более похожи на этот род, чем на полученные роды, такие как Gallimimus или Struthiomimus . [ 1 ] Впоследствии Молина-Пресес и Ларраменди оценили, что они имели длину около 1,8 метра (5,9 фута) и, возможно, весили около 13 килограммов (29 фунтов). [ 6 ]
Голотип
[ редактировать ]| Образец | Включает |
|---|---|
| PW5A-100 | дистальная часть левой плюсневой кости III |
| PW5A-101 | левая плюсневка II |
| PW5A-102 | проксимальная часть левой плюсневой кости IV |
| PW5A-103 | Средняя часть левой плюсневой кости III |
| PW5A-104 | проксимальная часть левой плюсневой кости III |
| PW5A-105 | Большая часть левой плюсневой кости II |
| PW5A-106 | Правая плюсневка IV |
| PW5A-107 | дистальная часть правой плюсневой кости III |
| PW5A-108 | проксимальная часть мегапотистого IV |
| PW5A-109 | проксимальная часть мегапотистого IV |
| PW5A-110 | Правая голень |
| PW5A-111 | левая голень |
| PW5A-112 | проксимальная часть левой малоберцовой кости |
| PW5A-113 | проксимальная часть правого лобка |
| PW5A-114 | проксимальная часть левого лобка |
| PW5A-115 до 121 | педали фаланги |
| PW5A-122 | частичная педаль фаланга с разоблачением |
| PW5A-123 | Дорсальный позвоночный центр |
| PW5A-124 | Частичный каудальный позвонок |
| PW5A-125 | Средний каудальный позвоночный центр |
| PW5A-126 до 129 | Дистальные каудальные позвонки |
| PW5A-130 | проксимальный каудальный позвонок |
| PW5A-131 | проксимальная часть правой плюсневой кости III |
Holoytpe of Kinnareemimus (PW5A-100) является частично полной третьей плюсневой кости , которая сохраняет суставную поверхность с левой стороны. Неполнота образца затрудняла для Бафтаута и коллег оценить полный размер кости. Тем не менее, другой образец (PW5A-101) является второй плюсневой кости с одинаковым размером и надежностью и гораздо более завершен. Авторы, описывающие его, считают, что он мог принадлежать тому же человеку, и поэтому они использовали его, чтобы оценить полную длину третьей плюсневой кости, которая составила примерно 145 миллиметров (5,7 дюйма) в жизни. [ 1 ]
Этот образец использовался для диагностики kinnareemimus в отличие от всех других орнитомимозавров , основанных на следующих апоморфиях : полуарктомтатарсальском состоянии, где третья плюсневая часть все еще видна в проксимальном взгляде и кодризирует в дистальном направлении, прежде чем распахнуть в более широкий треугольный сегмент и кожа на далеком конце. [ 1 ] Этот диагноз был внесен в поправку в 2024 году Саматхи, чтобы включить вентрально-педальные педали с сгибательными ямками, что поддерживало включение Kinnareemimus в Ornithomisosauria. [ 7 ]
Справляемые образцы
[ редактировать ]- Осевой скелет
Единственными частями осевого скелета Kinnareemimus , которые сохраняются, являются восемь позвонков из различных частей позвоночника (один предполагаемый дорсальный позвонок и семь хвостовых позвонков ). Одиночный некаудальный позвонок (PW5A-124) был выведен как дорсальный позвонок, потому что в нем отсутствуют суставные грани для шевронов , хотя это неизвестно наверняка. Нейронная арка для позвонков не сохранилась, а часть отсутствующей кости соответствует местоположению незаметных швов у незрелого человека, что заставило авторов предположить, что образец поступил от юношеского человека. [ 1 ]
Каудальные позвонки, которые присваиваются Kinnareemimus , от нескольких отдельных людей. Centra в имеют песочные часы и пустоте без поперечных процессов, напоминающие состояние, наблюдаемое Gallimimus . Некоторые из сохранившихся позвонков имеют различные формы поперечного сечения и, вероятно, поступают из разных участков хвоста. По крайней мере, один (PW5A-130) может быть первым каудальным позвонком. Семь позвонков, сохранившихся на других частях хвоста, различаются в градусах слияния, что указывает на эти образцы, связанные с множественной сборкой. [ 1 ]
- Аппендикулярный скелет
Большинство останков, обнаруженных в Phu Wiang 5, являются костями из задних лимбов или бедер разных людей. две частичные лобковые кости Сохраняются , которые очень стройны по сравнению с лобковыми костями такого же размера орнитомимозавров аналогичного размера . людей Buffetaut и коллеги отмечают, что это может быть не настоящей апоморфией, а вместо этого артефактом онтогенерии . [ 1 ]
Два голени были сохранены в типовой местности. Они похожи по длине, а один из правой ноги, а другой - с левой ноги. Однако, как полагают, они от разных людей из -за различий в надежности между двумя, которые, возможно, являются результатом онтогена. Общая форма голени напоминает примитивные орнитомимозавры с несколькими отличительными чертами. Единственная малоберцовая кость , которая сохраняется, является лишь частично завершена и сохраняет широкую и глубокую канавку вдоль вала, который напоминает состояние в Gallimimus . Сохраненные фаланги похожи по форме с гарудимимусом и арпимимусом , но они более стройные. В одном частичном когти, который сохраняется, не хватает туберкулеза сгибателя, аналогичного третьему некушному в Struthiomimus . [ 1 ]
Большинство остатков Kinnareemimus - это плюсневые кости, которые поступают из нижней части ноги чуть выше ноги. Известны плюсновики нескольких людей, что позволяет реконструировать плюсневую кость с разумной точностью. Примечательной особенностью является наличие арктометатарасальского состояния , которое аналогично получению полученных орнитомимидов и тиранозавра . Тем не менее, третья плюсневка не была полностью закрыта проксимальными концами второго и четвертого плюсневых, так что это не было настоящим арктометатарсом. [ 1 ]
Классификация
[ редактировать ]Хотя Kinnareemimus является относительно фрагментарным, имеющиеся данные реконструкций плюсневых средств допускают его относительную эволюционную позицию в Ornithomimosauria . Развитие Arctometatarsus является примерно линейным в филогенетической топологии орнитомимозавров. Исходя из этого, было выведено, что Kinnareemimus находится в более производной положении, чем гарудимимус , но менее полученным, чем Ornithomimidae . Тем не менее, Kinnareemimus геологически намного старше, чем Гарудимимус , а также пелеканимимус и арпимимус (также базальные орнитомимозавры), что позволяет предположить, что эволюция арктометатарса у орнитомимозавров не была хронологической линейной. Buffetaut и коллеги используют это, чтобы предположить, что большое количество эволюции орнитомимозавра еще не было отобрано. [ 1 ] Они не проводят филогенетический анализ, включая Kinnareemimus в их описании, [ 1 ] Но последующие авторы попытались сделать это. [ 8 ] [ 9 ]
В своей филогенетической работе над Theropods в 2014 году Стив Брусатт и его коллеги составили филогенетическую матрицу с использованием 152 таксонов, кодированных для 853 дискретных символов. Эта матрица включала данные для Kinnareemimus , но таксон должен был быть обрезан из последних деревьев, произведенных для бумаги. Дерево консенсуса по консенсусу с максимальной страстью, произведенное в дополнительных материалах с этой статьей, включало Kinnareemimus , и это дерево не смогло восстановить большинство манирапторовских клад как монофилетические . [ 8 ] Последующие анализы с использованием этой матрицы и других матриц обрезали Kinnareemimus по аналогичным причинам. [ 10 ] [ 11 ]
Первая научная статья, которая включала Kinnareemimus в опубликованных деревьях с существенным разрешением, была в 2024 году Андреа Кау . Дополнительные материалы из этой статьи включали десятки консенсусных деревьев, некоторые из которых включали Kinnareemimus . Сокращенная версия деревьев, опубликованных CAU, показана ниже. [ 9 ]




Вскоре после публикации анализа Theropod CAU в 2024 году Adun Samathi опубликовал статью, которая проанализировала филогенетическое положение Kinnareemimus . В своем анализе он отмечает, что было очень мало поддержки Kinnareemimus в качестве орнитомимозавра в основных филогенетических анализах. Brusatte и коллеги (2014) отметили, что, вероятно, это был базальный колурозавр . Хартман и его коллеги (2019) нашли его тиранозавроидом , но также поддержка его, возможно, представляющего альварессавроид . Чтобы решить неопределенность в литературе до этого момента, Самати набрал анатомические данные для Kinnareemimus в трех независимых матрицах теропода. Это были матрицы Choiniere и коллег (2012), McFeeters and Colleagues (2016) и MALICE Hattori и коллеги (2023). [ 7 ]
Три матрица восстановили разные результаты. Матрица от Choiniere и коллег, которая также была модифицирована Samathi, чтобы включить Deinocheirus , восстановил Kinnareemimus как базальный член Deinocheiridae . Матрица McFeeters и коллег восстановила его как не макрошейриформный орнитомимозавр, и этот результат также был восстановлен матрицей от Хаттори и его коллег. Самати в конечном итоге не смог разрешить уверенную позицию для таксона, кроме того, что это определенно не является членом Ornithomimidae . Последнее дерево, опубликованное в работе Самати, основанную на матрице Хаттори, показано ниже. [ 7 ]
Палеобиология
[ редактировать ]
В своем описании Kinnareemimus , Баффетаут и коллеги пытались оценить возможную адаптацию животного в жизни. Сохраненные кости позволили авторам реконструировать большую часть задней конечности на основе оценки размера взрослых костей, которые, как считается, были от несовершеннолетних. Соотношение между длиной большеберцовой кости и плюсневой кости было ранее установлено в качестве метода вывода Cursoriality у других орнитомимозавров . Расчетная длина лапки в Kinnareemimus была сопоставима с известными останками гарудимимуса и была значительно меньше, чем коэффициенты, известные из Struthiomimus и Gallimimus , которые, как полагают, были хорошо адаптированы для бега. [ 1 ]
Тем не менее, авторы также отметили, что плюсневые виды Kinnareemimus были гораздо более стройными, чем у косому археорнитомимусу , связанного с близко связанными , что может указывать на то, что либо животное не было полностью выращено, либо оно, возможно, приобрело педорфные признаки. Они не делают никаких конкретных гипотез о бегущих способностях Kinnareemimus . [ 1 ]
Палеоэкология
[ редактировать ]Палеоэкола
[ редактировать ]До сих пор формация Sao Khua только дала континентальные окаменелости, без каких -либо признаков морских окаменелостей или морских отложений . [ 12 ] Исторически были сообщения о зубах Ichthyosaur и Plesiosaur из формирования Sao Khua, но теперь они были идентифицированы как принадлежащие Crocodiliformes , которые в основном не являются немяскими во время меловых . [ 13 ] [ 14 ] Отложения формирования Sao Khua состоит из красных глин , мудров , песчаников , алеолетов и конгломератных пород, которые указывают на речную среду , которая также обладала озерами, поймами и плетеной каналами . [ 12 ] [ 15 ]
Современная фауна
[ редактировать ]
Из всех мезозойских образований в северо -восточном Таиланде, Sao Khua имеет наиболее распространенные и разнообразные ископаемые . [ 16 ] В специально известно, что местность Phu Wiang 5 включала небольшие, возможно, ювенильные образцы Sauropod Phuwiangosaurus . Спинозаврид . Siamosaurus также известен из этого места от зубов [ 1 ]
Регион был организован множеством других теропод, хотя не известно, уверенно, если какой -либо из них сосуществовал с Kinnareemimus конкретно. К ним относятся симпатрические Megaraptorans Phuwiangvenator и Vayuraptor , [ 17 ] Большой теропод Siamotyrannus , неназванный спинозаурид (иногда называемый «phu wiang spinosaurid b») и неназванного карчадонтозаврида . [ 13 ] [ 18 ] Район также был домом для других небольших теропод, включая неназванного орнитомимозавра , неназванного компонен и возможных орниторинов . [ 19 ] [ 20 ] Останки Сауропода также известны из области, которая, как считается, отличается от Phuwiangosaurus , но ни один из них не был официально описан или назван. Тем не менее, еще предстоит обнаружены какие -либо орнитишки, обнаруженные из формирования. [ 13 ]
Пресноводная жизнь изобилует в формировании Сан -Хуа. К ним относятся Neosuchians Siamosuchus , Sunosuchus и Theriosuchus , многочисленные виды черепах , амиформные рыбы, несколько гибодонтообразных акул и пресноводных двустворчатых моллюсков . [ 15 ] Существует также неопределенный птерозавр , известный из формирования. [ 21 ]
Смотрите также
[ редактировать ]- 2009 по палеонтологии архозавра
- Аптианский вымирание
- Хорат Группа
- Список азиатских динозавров
- Палеонтология в Таиланде
- Временная шкала исследования орнитомимозавра
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Buffetaut, Eric; Suteethorn, Varavudh; Тонг, Хайян (2009). «Ранний« Острих -динозавр »(Theropoda: Ornithomimosauria) из раннего мелового образования Sao Khua в Таиланде». В Баффетауте, Эрик; Cuny, G.; Le Loeuff, Джин; Suteethorn, Varavudh (ред.). Поздние палеозойские и мезозойские экосистемы в Se Asia . Геологическое общество Лондона. С. 229–243. doi : 10.1144/sp315.16 . ISBN 978-1-86239-275-5 Полем S2CID 128633687 .
- ^ Buffetaut, Eric; Suteethorn, Varavudh; Тонг, Хайян; Chaimanee, Yaowalak; Khansubha, Sasidhorn (1997). «Новые открытия динозавров в юрском периоде и меловом северо -восточном Таиланде» (PDF) . Международная конференция по стратиграфии, тектоническая эволюция Юго -Восточной Азии и южной части Тихого океана : 157–161.
- ^ Jump up to: а беременный Канеко, Рюичи (2000). «Следующие треки динозавров в Таиланде». Dino Press Vol. 1 Япония: Aurora Oval Inc.
- ^ «Журнал Dino Press» . Архив списка рассылки динозавров . 2000.
«Следующие треки динозавров в Таиланде»: статья об ихнологии динозавра Рюичи Канеко, с некоторым обсуждением Phuwiangosaurus и Siamotyrannus
- ^ «Исправление родов динозавров № 150» . Архив списка рассылки динозавров . 2000.
Статья Рюичи Канеко в первом выпуске Dino Press содержит название, пока еще подписываемого орнитомимидом из Таиланда, Джиннаримимус (не указано название вида)
- ^ Молинина-Пресес, Рубен; Lerramendi, Asher (2019). Факты и цифры динозавров: Тероподы и другие динозавриформы Перевод Коннолли, Дэвида; Рамирес Круз, Гонсало Ангель. Иллюстрируется Андрей Атутахин и Санте Маззеей. ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. ISBN 978-0691180311 .
- ^ Jump up to: а беременный в Самати, Адун (2024). «Филогенетическое положение kinnareemimus honkaenensis (Dinosauria: Theropoda: Ornithomimosauria) из нижнего мела Таиланда» Зотакса . 5448 : 67–8 Doi : 10.11646/ Zotaxa.5448.1.4
- ^ Jump up to: а беременный Брусатт, Стивен Л.; Ллойд, Грэм Т.; Ван, Стив С.; Норелл, Марк А. (2014). «Постепенная сборка плана птичьего тела завершилась быстрыми темпами эволюции на переходе динозавров-птиц» (PDF) . Текущая биология . 24 (20): 2386–2392. doi : 10.1016/j.cub.2014.08.034 . PMID 25264248 . S2CID 8879023 .
- ^ Jump up to: а беременный Кау А. (2024). В унифицированной рамках для макроэволюции предрасположенных динозавров . Бюллетень Итальянского палеонтологического общества, 63 (1): 1-19.
- ^ Ли Сюй; Йошицугу Кобаяши; Джунчанг Лю; Юн-Нам Ли; Юнцин Лю; Кохей Танака; Xingliao Zhang; Сонхай Цзя; Джиминг Чжан (2011). «Новый орнитомимидный динозавр с североамериканскими сродством из поздней меловой формирования Цюпа в провинции Хэнань в Китае». Мерашные исследования . 32 (2): 213–222. doi : 10.1016/j.cretres.2010.12.004 .
- ^ Серрано-боньяс, Клаудия Инес; Эспиноса-чавес, Белинда; МакКракен, С. Августа; Гутирес Бландо, Кирен; Из Леон-Давила, Клаудио; Вентура, Хосе Флорес (2020). « Paraxenisaurus numberensis , большой орнитомимозавр Deinocheirid из формирования Cerro del Pueblo (верхний меловой), Коауила, Мексика». Журнал южноамериканских наук о Земле . 101 . Bibcode : 2020jsaes . Doi : 10.1016/j.jsames.2020.102610 . S2CID 218968100 .
- ^ Jump up to: а беременный Такер, Райан Т.; Hyland, Ethan G.; Гейтс, Терри А.; Кинг М. Райан; Робертс, Эрик М.; Фоли, Эллиот К.; Берндт, Дэвид; Ханта, Раттанафорн; Хансубха, Саса-он; Aswasereelert, Wasiee; Занно, Линдсей Э. (2022). «Возраст, история осаждения и палеоклиматическая обстановка ранних меловых динозавров из формирования Sao Khua (Horat Group), Таиланд» . Палеогеография, палеоклиматология, палеоэкология . 601 : 111107. DOI : 10.1016/j.palaeo.2022.111107 . S2CID 249702713 .
- ^ Jump up to: а беременный в Buffetaut, Eric; Suteethorn, Varavudh (1999). «Фауна динозавров в формировании Таиланда Сан -Хуа и начало мелового излучения динозавров в Азии» . Палеогеография, палеоклиматология, палеоэкология . 150 (1–2): 13–23. Bibcode : 1999ppp ... 150 ... 13b . doi : 10.1016/s0031-0182 (99) 00004-8 . ISSN 0031-0182 .
- ^ Buffetaut, Eric; Жиль, Куни; Le Loeuff, Джин; Suteethorn, Varavudh (2009). Поздние палеозойские и мезозойские континентальные экосистемы в SE Asia . Лондон: Геологическое общество Лондона . С. 2–3. doi : 10.1144/sp315 . ISBN 978-1862395633 .
- ^ Jump up to: а беременный Эрик, Баффетаут; Varavudh, Suteethorn (1998). Холл, Роберт; Холлоуэй, Джереми Д. (ред.). «Биогеографическое значение мезозойских позвоночных из Таиланда» . Биогеография и геологическая эволюция SE Asia . Лейден, Нидерланды: издатели Backuys: 83–90.
- ^ Самати, Адун; Сандер, П. Мартин; Чантхазит, Форнфен (2021). «Спинозаурид из Таиланда (формация Сан -Хуа, ранний меловой) и переоценка Camarillasaurus cirugedae из раннего мела Испании» . Историческая биология . 33 (12): 3480–3494. doi : 10.1080/08912963.2021.1874372 . S2CID 233884025 .
- ^ Самати, А.; Chanthasit, P.; Мартин Сандер, П. (май 2019). «Два новых базальных динозавров с коелурозаврией из нижнего мелового образования Sao Khua в Таиланде» . Acta Palaeontologica Polonica . 64 (2): 239–260. doi : 10.4202/app.00540.2018 .
- ^ Buffetaut, Eric; Suteethorn, Varavudh (январь 2012 г.). «Carcharodontosaurid Theropod (Dinosauria, Saurischia) в формировании Sao Khua (ранний меловой, Барреманский) Таиланд» . 2012 Аннотация EAVP : 27–30.
- ^ Buffetaut, Eric; Suteethorn, Varavudh; Le Loeuff, Джин; Хансубха, Саса-он; Тонг, Хайян; Вонгко, К. (январь 2005 г.). «Фауна динозавра из формирования Хук Круат (ранний меловой) Таиланда» . Международная конференция по геологии, геотехнологии и минеральным ресурсам Индокитая (Geoindo 2005) : 575–581.
- ^ Buffetaut, Eric; Дайк, Гарет; Suteethorn, Varavudh; Тонг, Хайян (2005-12-01). «Первая запись ископаемой птицы из раннего мела Таиланда» . Comptes rendus palevol . 4 (8): 681–686. doi : 10.1016/j.crpv.2005.06.002 . ISSN 1631-0683 .
- ^ Куни, Жиль; Suteethorn, V. (2003-01-01). «Птеродактилоидный зуб из формации Сан -Хуа (ранний меловой) Таиланда» . Махасаракхэмский университетский журнал . 22 : 92–98.
Внешние ссылки
[ редактировать ]- Краткое описание архивировано 2011-07-17 на The Wayback Machine в списке рассылки динозавров














