Lotka–Volterra equations
Уравнения Лотки-Вольтерра , также известные как модель хищник-жертва Лотки-Вольтерра , представляют собой пару нелинейных дифференциальных уравнений первого порядка , часто используемых для описания динамики биологических систем , в которых взаимодействуют два вида: один в качестве хищника , а другой в качестве хищника. другой как добыча. Популяции меняются со временем в соответствии с парой уравнений:

где
- переменная добычи ( x — плотность популяции например, количество кроликов на квадратный километр);
- переменная y — плотность популяции какого-либо хищника (например, количество лисиц на квадратный километр);
- и представляют мгновенные темпы роста двух популяций;
- т представляет время;
- добычи Параметры , α и β , описывают, соответственно, максимальную скорость роста добычи на душу населения и влияние присутствия хищников на уровень смертности жертв.
- Параметры хищника, γ , δ , соответственно описывают уровень смертности хищника на душу населения и влияние присутствия добычи на скорость роста хищника.
- Все параметры положительные и реальные.


Решение дифференциальных уравнений является детерминированным и непрерывным . Это, в свою очередь, означает, что поколения хищника и жертвы постоянно перекрываются. [1]
Система уравнений Лотки–Вольтерра является примером модели Колмогорова , [2] [3] [4] Это более общая структура, которая может моделировать динамику экологических систем с взаимодействием хищник-жертва, конкуренцией , болезнями и мутуализмом .
Биологическая интерпретация и модельные предположения
[ редактировать ]Предполагается, что жертва имеет неограниченный запас пищи и размножается в геометрической прогрессии, если только она не подвергается нападению хищников; этот экспоненциальный рост представлен в приведенном выше уравнении термином αx . Предполагается, что скорость нападения хищников на жертву пропорциональна скорости встречи хищников и жертвы; выше это представлено как βxy . Если хотя бы x или y равны нулю, хищничества быть не может. С помощью этих двух членов приведенное выше уравнение добычи можно интерпретировать следующим образом: скорость изменения популяции добычи определяется ее собственным темпом роста минус скорость, с которой на нее охотятся.
Термин δxy представляет собой рост популяции хищников. (Обратите внимание на сходство со скоростью хищничества; однако используется другая константа, поскольку скорость роста популяции хищников не обязательно равна скорости, с которой они поедают добычу). Термин γy представляет собой уровень потерь хищников из-за естественной смерти или эмиграции; это приводит к экспоненциальному затуханию в отсутствие добычи. Следовательно, уравнение выражает, что скорость изменения популяции хищника зависит от скорости, с которой он поедает добычу, за вычетом его собственного уровня смертности.
Модель «хищник-жертва» Лотки-Вольтерры делает ряд предположений об окружающей среде и биологии популяций хищников и жертв: [5]
- Популяция добычи всегда находит достаточно еды.
- Кормовая база популяции хищников полностью зависит от численности популяции жертв.
- Скорость изменения численности населения пропорциональна ее численности.
- В ходе этого процесса окружающая среда не меняется в пользу одного вида, и генетическая адаптация несущественна.
- Хищники имеют безграничный аппетит.
- Обе популяции могут быть описаны одной переменной. Это равнозначно предположению, что популяции не имеют пространственного или возрастного распределения, которое способствует динамике.
Биологическая значимость модели
[ редактировать ]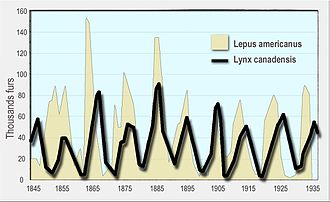
Ни одно из приведенных выше предположений вряд ли справедливо для естественных популяций. Тем не менее, модель Лотки-Вольтерра демонстрирует два важных свойства популяций хищников и жертв, и эти свойства часто распространяются на варианты модели, в которых эти предположения смягчены:
Во-первых, динамика популяций хищников и жертв имеет тенденцию к колебаниям. В естественных популяциях наблюдались колебания численности хищников и жертв, например, о рыси и зайце-снегоступе. данные компании Гудзонова залива [6] а также популяции лосей и волков в национальном парке Айл-Рояль . [7]
Во-вторых, популяционное равновесие этой модели обладает тем свойством, что равновесная плотность добычи (задаваемая выражением ) зависит от параметров хищника и равновесной плотности хищника (задаваемой выражением ) от параметров жертвы. Следствием этого является то, что увеличение, например, скорости роста добычи, , приводит к увеличению равновесной плотности хищников, но не равновесной плотности жертв. Улучшение окружающей среды для добычи приносит пользу хищнику, а не добыче (это связано с парадоксом пестицидов и парадоксом обогащения ). Проявлением этого явления служит возросший процент вылова хищных рыб, увеличившийся в годы 1-й мировой войны (1914–18), когда скорость роста добычи увеличилась за счет сокращения промыслового усилия.



Еще одним примером является экспериментальное удобрение океана железом. В нескольких экспериментах в океане было растворено большое количество солей железа. Ожидалось, что железо, которое является лимитирующим питательным веществом для фитопланктона, будет стимулировать рост фитопланктона и изолировать углекислый газ из атмосферы. Добавление железа обычно приводит к кратковременному цветению фиопланктона, который быстро потребляется другими организмами (например, мелкой рыбой или зоопланктоном ) и ограничивает эффект обогащения главным образом увеличением плотности хищников, что, в свою очередь, ограничивает секвестрацию углерода. Это предсказано равновесной плотностью населения в модели «хищник-жертва» Лотки-Вольтерры и является особенностью, которая переносится на более сложные модели, в которых ограничительные предположения простой модели смягчены. [8]
Приложения к экономике и маркетингу
[ редактировать ]Модель Лотки – Вольтерры имеет дополнительные приложения в таких областях, как экономика. [9] и маркетинг. [10] [11] Его можно использовать для описания динамики рынка с несколькими конкурентами, взаимодополняющими платформами и продуктами, экономикой совместного использования и многим другим. Существуют ситуации, в которых один из конкурентов вытесняет других конкурентов с рынка, и другие ситуации, в которых рынок достигает равновесия, при котором каждая фирма стабилизирует свою долю рынка. Также можно описать ситуации, в которых происходят циклические изменения в отрасли или хаотичные ситуации, в которых нет равновесия, а изменения часты и непредсказуемы.
История
[ редактировать ]«хищник-жертва» Лотки-Вольтерра Модель была первоначально предложена Альфредом Дж. Лоткой в теории автокаталитических химических реакций в 1910 году. [12] [13] По сути, это было логистическое уравнение : [14] первоначально выведен Пьером Франсуа Верхюльстом . [15] расширил модель В 1920 году Лотка через Андрея Колмогорова до «органических систем», используя в качестве примера виды растений и виды травоядных животных. [16] а в 1925 году он использовал эти уравнения для анализа взаимодействия хищник-жертва в своей книге по биоматематике . [17] Тот же набор уравнений был опубликован в 1926 году Вито Вольтеррой , математиком и физиком, который заинтересовался математической биологией . [13] [18] [19] Расследование Вольтерры было вдохновлено его общением с морским биологом Умберто Д'Анкона , который в то время ухаживал за его дочерью, а позже должен был стать его зятем. Д'Анкона изучал уловы рыбы в Адриатическом море и заметил, что в годы 1-й мировой войны (1914–18) процент выловленной хищной рыбы увеличился. Это озадачило его, поскольку за годы войны усилия рыболовства значительно сократились, а поскольку предпочтительным уловом является добыча рыбы, интуитивно можно было бы ожидать, что это приведет к увеличению процента добычи рыбы. Вольтерра разработал свою модель для объяснения наблюдения Д'Анконы и сделал это независимо от Альфреда Лотки. В своей публикации он упомянул более раннюю работу Лотки, после чего модель стала известна как «модель Лотки-Вольтерры». [20]
Позже модель была расширена, включив в нее зависящий от плотности рост добычи и функциональную реакцию формы, разработанной К.С. Холлингом ; модель, которая стала известна как модель Розенцвейга – Макартура. [21] Модели Лотки-Вольтерры и Розенцвейга-Макартура использовались для объяснения динамики естественных популяций хищников и жертв.
В конце 1980-х годов появилась альтернатива модели Лотки-Вольтерры «хищник-жертва» (и ее обобщения, зависящие от общей жертвы) - модель, зависящая от соотношения, или модель Ардити-Гинзбурга . [22] Обоснованность моделей, зависящих от добычи или соотношения, широко обсуждается. [23]
Уравнения Лотки-Вольтерра имеют долгую историю использования в экономической теории ; их первоначальную заявку обычно приписывают Ричарду Гудвину в 1965 году. [24] или 1967 год. [25] [26]
Решения уравнений
[ редактировать ]Уравнения имеют периодические решения. Эти решения не имеют простого выражения в терминах обычных тригонометрических функций , хотя они вполне сносны. [27] [28] [29]
Если ни один из неотрицательных параметров α , β , γ , δ не обращается в нуль, три можно включить в нормализацию переменных, чтобы оставить только один параметр: поскольку первое уравнение однородно по x , а второе по y , параметры β / α и δ / γ поглощаются при нормализации y и x соответственно, а γ — при нормализации t , так что только α / γ остается произвольным. Это единственный параметр, влияющий на характер решений.
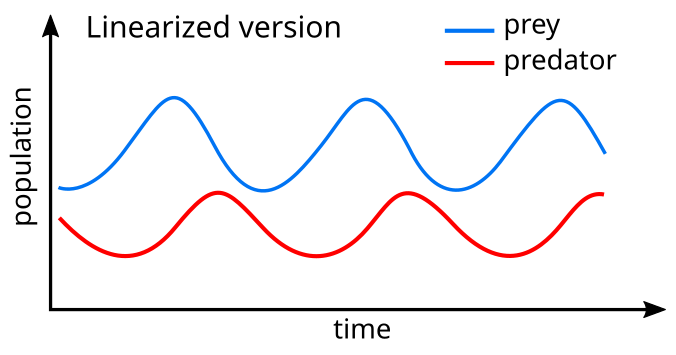
Линеаризация простому уравнений дает решение, подобное гармоническому движению. [30] при этом популяция хищников отстает от популяции добычи на 90° в цикле.
Простой пример
[ редактировать ]

Предположим, есть два вида животных: кролик (добыча) и лиса (хищник). Если первоначальная плотность составляет 10 кроликов и 10 лисиц на квадратный километр, можно построить график развития этих двух видов с течением времени; с учетом параметров темпы роста и смертности кроликов составляют 1,1 и 0,4, а у лисиц - 0,1 и 0,4 соответственно. Выбор временного интервала произволен.
Можно также построить решения параметрически в виде орбит в фазовом пространстве без представления времени, но с одной осью, представляющей количество добычи, а другой осью, представляющей плотность хищников за все время.
Это соответствует исключению времени из двух приведенных выше дифференциальных уравнений для получения одного дифференциального уравнения.

связывая переменные x (хищник) и y (добыча). Решениями этого уравнения являются замкнутые кривые. Он поддается разделению переменных : интегрированию

дает неявное отношение

где V — постоянная величина, зависящая от начальных условий и сохраняющаяся на каждой кривой.
Отступление: эти графики иллюстрируют серьезное потенциальное ограничение в применении в качестве биологической модели: при таком конкретном выборе параметров в каждом цикле популяция кроликов сокращается до чрезвычайно низкой численности, но восстанавливается (в то время как популяция лисиц остается значительной на уровне самая низкая плотность кроликов). Однако в реальных ситуациях случайные колебания дискретного числа особей могут привести к фактическому вымиранию кроликов, а, как следствие, и лисиц. Эту задачу моделирования назвали «проблемой атто-лисы», причем атто -лиса является условным 10 −18 лисы. [31] [32] Плотность 10 −18 лисиц на квадратный километр соответствует в среднем примерно 5×10 −10 лисиц на поверхности земли, что в практическом плане означает, что лисы вымерли.
Гамильтонова структура системы
[ редактировать ]Поскольку количество сохраняется во времени, она играет роль гамильтоновой функции системы. [33] Чтобы убедиться в этом, мы можем определить скобку Пуассона следующим образом: . Тогда уравнения Гамильтона будут иметь вид Переменные и не являются каноническими, поскольку . Однако, используя преобразования [34] и мы пришли к канонической форме уравнений Гамильтона с гамильтонианом :










График в фазовом пространстве еще одного примера
[ редактировать ]
Менее крайний пример охватывает:
α знак равно 2/3 , β знак равно 4/3 , γ знак равно 1 знак равно δ . Предположим, что x и y измеряют тысячи каждый. Круги обозначают начальные условия добычи и хищника от x = y = 0,9 до 1,8 с шагом 0,1. Фиксированная точка находится в (1, 1/2).
Динамика системы
[ редактировать ]В модельной системе хищники процветают, когда добыча многочисленна, но в конечном итоге превосходят запасы пищи и сокращаются. Поскольку популяция хищников невелика, популяция жертв снова увеличится. Эта динамика продолжается в численности населения цикле роста и сокращения .
Равновесие населения
[ редактировать ]Равновесие численности населения возникает в модели, когда ни один из уровней численности населения не меняется, т.е. когда обе производные равны 0:


Приведенная выше система уравнений дает два решения: и


Следовательно, существует два равновесия.
Первое решение фактически представляет собой вымирание обоих видов. Если обе популяции равны 0, то они будут оставаться такими до бесконечности. Второе решение представляет собой фиксированную точку, в которой обе популяции поддерживают свою текущую, ненулевую численность, и в упрощенной модели делают это бесконечно. Уровни численности населения, при которых достигается это равновесие, зависят от выбранных значений параметров α , β , γ и δ .
Стабильность фиксированных точек
[ редактировать ]Стабильность фиксированной точки в начале координат можно определить, выполнив линеаризацию с использованием частных производных .
Матрица Якоби модели хищник-жертва: и известна как матрица сообщества .

Первая фиксированная точка (затухание)
[ редактировать ]При оценке в установившемся состоянии (0, 0) матрица Якоби J становится

Собственные значения этой матрицы равны

В модели α и γ всегда больше нуля, и поэтому знаки приведенных выше собственных значений всегда будут различаться. Следовательно, фиксированная точка в начале координат является седловой точкой .
Нестабильность этой неподвижной точки имеет значение. Если бы она была стабильной, к ней могли бы быть привлечены ненулевые популяции, и поэтому динамика системы могла бы привести к исчезновению обоих видов во многих случаях начального уровня популяции. Однако, поскольку фиксированная точка происхождения является седловой и, следовательно, нестабильной, из этого следует, что вымирание обоих видов в модели затруднено. (На самом деле, это могло произойти только в том случае, если бы жертва была искусственно полностью уничтожена, в результате чего хищники умерли от голода. Если бы хищники были уничтожены, популяция жертв в этой простой модели росла бы без ограничений.) Популяции жертв и хищников могут приблизиться к нулю и все равно восстановиться.
Вторая фиксированная точка (колебания)
[ редактировать ]Оценка J во второй фиксированной точке приводит к

Собственные значения этой матрицы равны

Поскольку собственные значения являются чисто мнимыми и сопряжены друг с другом, эта неподвижная точка должна быть либо центром замкнутых орбит в локальной окрестности, либо спиралью притяжения или отталкивания. В консервативных системах должны существовать замкнутые орбиты в локальной окрестности неподвижных точек, существующих в минимумах и максимумах сохраняющейся величины. Сохраняющаяся величина получена выше как на орбитах. Таким образом, орбиты вокруг фиксированной точки замкнуты и эллиптические , поэтому решения являются периодическими, колеблющимися по небольшому эллипсу вокруг фиксированной точки с частотой и период .



Как показано на циркуляционных колебаниях на рисунке выше, кривые уровня представляют собой замкнутые орбиты, окружающие фиксированную точку: уровни популяций хищника и жертв циклически колеблются и колеблются без затухания вокруг фиксированной точки с частотой. .

Значение константы движения V или, что то же самое, K = exp(− V ) , , можно найти для замкнутых орбит вблизи неподвижной точки.

Увеличение K приближает замкнутую орбиту к фиксированной точке. Наибольшее значение константы K получается при решении оптимизационной задачи Таким образом, максимальное значение K достигается в стационарной (неподвижной) точке. и составляет где е — число Эйлера .



См. также
[ редактировать ]- Конкурентные уравнения Лотки – Вольтерра
- Обобщенное уравнение Лотки – Вольтерра.
- Мутуализм и уравнение Лотки – Вольтерра
- Матрица сообщества
- Динамика населения
- Динамика численности рыбного хозяйства
- Модель Николсона – Бейли
- Реакционно-диффузионная система
- Парадокс обогащения
- Законы Ланчестера , аналогичная система дифференциальных уравнений для военных сил.
- Random generalized Lotka–Volterra model
- Модель потребительских ресурсов
Примечания
[ редактировать ]- ^ Кук, Д.; Хиорнс, RW; и др. (1981). Математическая теория динамики биологических популяций . Том. II. Академическая пресса.
- ^ Фридман, Гавайи (1980). Детерминированные математические модели в популяционной экологии . Марсель Деккер .
- ^ Брауэр, Ф.; Кастильо-Чавес, К. (2000). Математические модели в популяционной биологии и эпидемиологии . Спрингер-Верлаг .
- ^ Хоппенстедт, Ф. (2006). «Модель Хищник-жертва» . Схоларпедия . 1 (10): 1563. Бибкод : 2006SchpJ...1.1563H . doi : 10.4249/scholarpedia.1563 .
- ^ «ДИНАМИКА ХИЩНИК-ЖЕРТВА» . www.tiem.utk.edu . Архивировано из оригинала 15 декабря 2012 г. Проверено 9 января 2018 г.
- ^ Гилпин, Мэн (1973). «Едят ли зайцы рысь?». Американский натуралист . 107 (957): 727–730. дои : 10.1086/282870 . S2CID 84794121 .
- ^ Йост, К.; Девулдер, Г.; Вучетич, Дж. А.; Петерсон, Р.; Ардити, Р. (2005). «Волки острова Рояль демонстрируют независимую от масштаба насыщенность и зависящую от плотности хищничество по отношению к лосям». Дж. Аним. Экол . 74 (5): 809–816. дои : 10.1111/j.1365-2656.2005.00977.x .
- ^ Пан, А.; Пурзиаи, Б.; Хуанг, Х. (3 июня 2015 г.). «Влияние удобрения океана железом на биологический углеродный насос фитопланктона». Успехи прикладной математики и механики . 3 (1): 52–64. дои : 10.4208/aamm.10-m1023 . S2CID 124606355 .
- ^ Прасолов, Александр В. (2016). Некоторые количественные методы и модели в экономической теории . Экономические вопросы, проблемы и перспективы. Нью-Йорк: Издательство Nova. ISBN 978-1-63484-937-1 .
- ^ Хун, Хуэй-Чи; Чиу, Ю-Чи; Ву, Мух-Чернг (2017). «Модифицированная модель Лотки – Вольтерра для распространения и замены многопоколенческих технологий обработки DRAM» . Математические проблемы в технике . 2017 : 1–12. дои : 10.1155/2017/3038203 . ISSN 1024-123X .
- ^ Орбах, Яир (2022). Прогнозирование динамики рынка и технологий . Израиль: Издательство Ариэльского университета. стр. 123–143. ISBN 978-965-7632-40-6 .
- ^ Лотка, AJ (1910). «Вклад в теорию периодической реакции» . Дж. Физ. хим. 14 (3): 271–274. дои : 10.1021/j150111a004 .
- ^ Jump up to: а б Гоэл, Н.С.; и др. (1971). О Вольтерре и других нелинейных моделях взаимодействующих популяций . Академическая пресса. ISBN 0-12-287450-1 .
- ^ Берриман, А.А. (1992). «Происхождение и эволюция теории хищник-жертва» (PDF) . Экология . 73 (5): 1530–1535. дои : 10.2307/1940005 . JSTOR 1940005 . Архивировано из оригинала (PDF) 31 мая 2010 г.
- ^ Верхюльст, PH (1838 г.). «Заметьте по закону, что численность населения продолжает увеличиваться» . Корр. Математика и физика . 10 : 113–121.
- ^ Лотка, AJ (1920). «Аналитическая записка о некоторых ритмических отношениях в органических системах» . Учеб. Натл. акад. наук. США 6 (7): 410–415. Бибкод : 1920PNAS....6..410L . дои : 10.1073/pnas.6.7.410 . ПМЦ 1084562 . ПМИД 16576509 .
- ^ Лотка, AJ (1925). Элементы физической биологии . Уильямс и Уилкинс .
- ^ Вольтерра, В. (1926). «Вариации и колебания численности особей сожительствующих видов животных». Мем. Линкейцы Рим . 2 :31–113.
- ^ Вольтерра, В. (1931). «Вариации и колебания численности особей у видов животных, живущих вместе». В Чепмене, Р.Н. (ред.). Экология животных . МакГроу-Хилл .
- ^ Кингсленд, С. (1995). Моделирование природы: эпизоды истории популяционной экологии . Издательство Чикагского университета. ISBN 978-0-226-43728-6 .
- ^ Розенцвейг, ML; Макартур, Р.Х. (1963). «Графическое представление и условия устойчивости взаимодействия хищник-жертва». Американский натуралист . 97 (895): 209–223. дои : 10.1086/282272 . S2CID 84883526 .
- ^ Ардити, Р.; Гинзбург, Л.Р. (1989). «Связь в динамике хищник-жертва: зависимость от соотношения» (PDF) . Журнал теоретической биологии . 139 (3): 311–326. Бибкод : 1989JThBi.139..311A . дои : 10.1016/s0022-5193(89)80211-5 . Архивировано из оригинала (PDF) 4 марта 2016 г. Проверено 26 июня 2013 г.
- ^ Абрамс, Пенсильвания; Гинзбург, Л.Р. (2000). «Природа хищничества: зависит от добычи, зависит от соотношения или нет?». Тенденции в экологии и эволюции . 15 (8): 337–341. дои : 10.1016/s0169-5347(00)01908-x . ПМИД 10884706 .
- ^ Гандольфо, Г. (2008). «Джузеппе Паломба и уравнения Лотки – Вольтерра». Рендиконти Линчеи . 19 (4): 347–357. дои : 10.1007/s12210-008-0023-7 . S2CID 140537163 .
- ^ Гудвин, Р.М. (1967). «Цикл роста» . В Файнштейне, CH (ред.). Социализм, капитализм и экономический рост . Издательство Кембриджского университета .
- ^ Десаи, М.; Ормерод, П. (1998). «Ричард Гудвин: краткая признательность» (PDF) . Экономический журнал . 108 (450): 1431–1435. CiteSeerX 10.1.1.423.1705 . дои : 10.1111/1468-0297.00350 . Архивировано из оригинала (PDF) 27 сентября 2011 г. Проверено 22 марта 2010 г.
- ^ Штайнер, Антонио; Гандер, Мартин Якоб (1999). «Параметрические решения уравнений хищник-жертва в сравнении» . Иль Вольтерриано . 7 :32–44.
- ^ Эванс, CM; Финдли, Г.Л. (1999). «Новая трансформация проблемы Лотки-Вольтерры». Журнал математической химии . 25 : 105–110. дои : 10.1023/A:1019172114300 . S2CID 36980176 .
- ^ Леконт, М.; Массон, П.; Ци, Л. (2022). «Предельные колебания цикла, время отклика и зависящее от времени решение модели хищник-жертва Лотки-Вольтерры». Физика плазмы . 29 (2): 022302. arXiv : 2110.11557 . дои : 10.1063/5.0076085 . S2CID 239616189 .
- ^ Тонг, Х. (1983). Пороговые модели в нелинейном анализе временных рядов . Спрингер-Верлаг.
- ^ Лобри, Клод; Сари, Тевфик (2015). «Миграции в модели Розенцвейга-Макартура и проблема «атто-лисы»» (PDF) . Арима . 20 : 95–125.
- ^ Моллисон, Д. (1991). «Зависимость темпов эпидемии и численности населения от основных параметров» (PDF) . Математика. Биосци . 107 (2): 255–287. дои : 10.1016/0025-5564(91)90009-8 . ПМИД 1806118 .
- ^ Нутку, И. (1990). «Гамильтонова структура уравнений Лотки-Вольтерра» . Буквы по физике А. 145 (1): 27–28. Бибкод : 1990PhLA..145...27N . дои : 10.1016/0375-9601(90)90270-X . hdl : 11693/26204 . S2CID 121710034 .
- ^ Байджент, Стив (02 марта 2010 г.). «Динамика Лотки-Вольтерры. Введение» (PDF) .
Дальнейшее чтение
[ редактировать ]- Хофбауэр, Йозеф; Зигмунд, Карл (1998). «Динамические системы и уравнения Лотки – Вольтерра». Эволюционные игры и демографическая динамика . Нью-Йорк: Издательство Кембриджского университета. стр. 1–54. ISBN 0-521-62570-Х .
- Каплан, Дэниел; Гласс, Леон (1995). Понимание нелинейной динамики . Нью-Йорк: Спрингер. ISBN 978-0-387-94440-1 .
- Ли, скорая помощь (1968). «Экологическая роль уравнений Вольтерра». Некоторые математические проблемы биологии . - современная дискуссия с использованием данных компании Гудзонова залива о рыси и зайцах в Канаде с 1847 по 1903 год.
- Мюррей, доктор юридических наук (2003). Математическая биология I: Введение . Нью-Йорк: Спрингер. ISBN 978-0-387-95223-9 . '
- Конспект лекций курса Стефано Аллесины по экологии сообщества: https://stefanoallesina.github.io/Theoretical_Community_Ecology/
Внешние ссылки
[ редактировать ]- Из демонстрационного проекта Wolfram — требуется проигрыватель CDF (бесплатно) :
- Алгоритмическое моделирование Лотки-Вольтерра (Веб-моделирование).