Убиквитин-активирующий фермент
| Убиквитин-активирующие ферменты | |||
|---|---|---|---|
 Кристаллическая структура дрожжевого убиквитин-активирующего фермента E1 / убиквитин-комплекса. [ 1 ] | |||
| Идентификаторы | |||
| ЕС №. | 6.2.1.45 | ||
| CAS №. | 74812-49-0 | ||
| Альт. имена | E1 ферменты | ||
| Базы данных | |||
| Intenz | Intenz View | ||
| Бренда | Бренда вход | ||
| Расширение | Вид Nicezyme | ||
| Кегг | Кегг вход | ||
| Метатический | Метаболический путь | ||
| Напрямую | профиль | ||
| PDB Структуры | RCSB PDB PDBE PDBSUM | ||
| Джин Онтология | Друг / Quickgo | ||
| |||
Убиквитин-активирующие ферменты , также известные как ферменты E1 , катализируют первый этап в реакции убиквитинирования , который (среди прочего) может нацелиться на белок для деградации через протеасому . Эта ковалентная связь убиквитина убиквитинподобных или белков с целевыми белками является основным механизмом регуляции функции белка у эукариотических организмов . [ 2 ] Многие процессы, такие как деление клеток , иммунные ответы и эмбриональное развитие, также регулируются посттрансляционной модификацией убиквитин и убиквитин-подобными белками. [ 2 ]
Убиквитинирование (UbiquityLation)
[ редактировать ]Убиквитин-активирующий фермент (E1) запускает процесс убиквитинирования (рис. 1). Фермент E1, наряду с АТФ , связывается с белком убиквитина. Затем фермент E1 передает белок убиквитин во второй белок, называемый убиквитин -носителем или конъюгационным белком (E2). Комплексы белков E2 с белковой лигазой убиквитин (E3). Эта убиквитин белка лигаза распознает, какой белок должен пометить, и катализирует перенос убиквитина в этот белок. Этот путь повторяется до тех пор, пока целевой белок не получит полную цепь убиквитина, прикрепленной к себе. [ 3 ]
Структура и механизм
[ редактировать ]В начале каскада убиквитинирования фермент E1 (рис. 2) связывает АТФ-мг 2+ и убиквитин и катализирует убиквитин С-концевой ацил аденилирование. [ 4 ] На следующем этапе каталитический цистеин (рис. 3) на ферменте E1 атакует комплекс убиквитин-AMP посредством замены ацила, одновременно создавая связь тиоэфира и группу, выходящую из усилителя. [ 2 ] Наконец, комплекс E1-убиквитина передает убиквитин в фермент E2 с помощью реакции транстеотерификации, в которой каталитический цистеин E2 атакует заднюю сторону комплекса E1-убиквитина. [ 5 ] Тем не менее, процесс транстеотеризации очень сложный, так как ферменты E1 и E2 образуют промежуточный комплекс, в котором оба фермента подвергаются ряду конформационных изменений , чтобы связываться друг с другом. [ 5 ]
На протяжении всего этого механизма фермент E1 связан с двумя молекулами убиквитина. Хотя этот вторичный убиквитин аналогично аденилируется, он не образует тот же комплекс тиоэфира, который описан ранее. Функция вторичного убиквитина остается в значительной степени неизвестной, однако считается, что он может способствовать конформационным изменениям, наблюдаемым в ферменте E1 во время процесса транстеотерификации. [ 2 ]
-
![Рисунок 1. показывает последовательность того, как активирующий фермент убиквитин присоединяется к АТФ и субстрату убиквитина. Это также показывает, как два субстрата убиквитина могут быть связаны за один раз. [6]](//upload.wikimedia.org/wikipedia/commons/thumb/0/01/Ubiquitin-activating_enzyme_bound_to_ATP_and_ubiquitin_substrate.png/300px-Ubiquitin-activating_enzyme_bound_to_ATP_and_ubiquitin_substrate.png) Рисунок 1. показывает последовательность того, как активирующий фермент убиквитин присоединяется к АТФ и субстрату убиквитина. Это также показывает, как два субстрата убиквитина могут быть связаны за один раз. [ 6 ]
Рисунок 1. показывает последовательность того, как активирующий фермент убиквитин присоединяется к АТФ и субстрату убиквитина. Это также показывает, как два субстрата убиквитина могут быть связаны за один раз. [ 6 ] -
![Рисунок 2. Белок E1 связывает молекулу убиквитина в каждом из двух идентичных активных сайтов (выделено). Важные остатки, цистеин и аргинин, помечены красным. [2]](//upload.wikimedia.org/wikipedia/commons/thumb/9/91/Ubiquitin_activating_enzyme_cysteine_highlighted.png/300px-Ubiquitin_activating_enzyme_cysteine_highlighted.png) Рисунок 2. Белок E1 связывает молекулу убиквитина в каждом из двух идентичных активных сайтов (выделено). Важные остатки, цистеин и аргинин, помечены красным. [ 2 ]
Рисунок 2. Белок E1 связывает молекулу убиквитина в каждом из двух идентичных активных сайтов (выделено). Важные остатки, цистеин и аргинин, помечены красным. [ 2 ] -
![Рисунок 3. Обзор крупным планом на несвязанном активном участке. Считается, что arg (603) перезаряжается каталитическую Cys (600) после того, как убиквитин был перенесен в фермент E2. [2]](//upload.wikimedia.org/wikipedia/commons/thumb/3/36/Cys_and_Arg_active_site_of_ubiquitin_activating_enzyme.png/300px-Cys_and_Arg_active_site_of_ubiquitin_activating_enzyme.png) Рисунок 3. Обзор крупным планом на несвязанном активном участке. Считается, что arg (603) перезаряжается каталитическую Cys (600) после того, как убиквитин был перенесен в фермент E2. [ 2 ]
Рисунок 3. Обзор крупным планом на несвязанном активном участке. Считается, что arg (603) перезаряжается каталитическую Cys (600) после того, как убиквитин был перенесен в фермент E2. [ 2 ] -
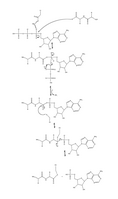 Рисунок 4. Полный механизм аденилирования убиквитина и последующего убиквитин -связывания с E1.
Рисунок 4. Полный механизм аденилирования убиквитина и последующего убиквитин -связывания с E1. -
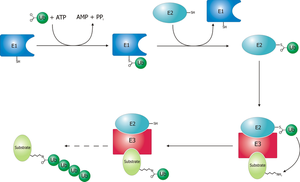 Схематическая диаграмма системы убиквитиляции.
Схематическая диаграмма системы убиквитиляции.
![Рисунок 1. показывает последовательность того, как активирующий фермент убиквитин присоединяется к АТФ и субстрату убиквитина. Это также показывает, как два субстрата убиквитина могут быть связаны за один раз. [6]](http://upload.wikimedia.org/wikipedia/commons/thumb/0/01/Ubiquitin-activating_enzyme_bound_to_ATP_and_ubiquitin_substrate.png/300px-Ubiquitin-activating_enzyme_bound_to_ATP_and_ubiquitin_substrate.png)
![Рисунок 2. Белок E1 связывает молекулу убиквитина в каждом из двух идентичных активных сайтов (выделено). Важные остатки, цистеин и аргинин, помечены красным. [2]](http://upload.wikimedia.org/wikipedia/commons/thumb/9/91/Ubiquitin_activating_enzyme_cysteine_highlighted.png/300px-Ubiquitin_activating_enzyme_cysteine_highlighted.png)
![Рисунок 3. Обзор крупным планом на несвязанном активном участке. Считается, что arg (603) перезаряжается каталитическую Cys (600) после того, как убиквитин был перенесен в фермент E2. [2]](http://upload.wikimedia.org/wikipedia/commons/thumb/3/36/Cys_and_Arg_active_site_of_ubiquitin_activating_enzyme.png/300px-Cys_and_Arg_active_site_of_ubiquitin_activating_enzyme.png)


Изозимы
[ редактировать ]Следующие гены кодируют убиквитин-активирующие ферменты:
Ассоциация болезней
[ редактировать ]Система убиквитина-протеасом имеет решающее значение для соответствующей деградации белка в клетках. Дисфункции этой системы может нарушать клеточный гомеостаз и привести к множеству расстройств. В нормально функционирующих клетках ковалентная связь убиквитина или убиквитин-подобного белка с целевым белком изменяет поверхность целевого белка. Эти убиквитинированные белки подвержены деградации протеолитическими и непротеолитическими путями. [ 7 ] Если эта система сработает, могут возникнуть в результате многочисленные наследственные и приобретенные заболевания, такие как рак, диабет , инсульт , болезнь Альцгеймера , боковой склероз , астма, рассеянный склероз , астма , воспалительное заболевание кишечника , аутоиммунный тиреоид , воспалительный артрит , лупс и синдром вексаса . [ 7 ]
Миссен в UBE1 и X-связанной инфантильной мышечной атрофии позвоночника (XL-SMA)
[ редактировать ]Среди различных расстройств, связанных с путем убиквитин-протеасом, находится X-связанная с инфантильной мышечной атрофией позвоночника (XL-SMA). [ 8 ] Фатальное детское расстройство связано с потерей клеток переднего рога и детской смерти. Клинические особенности включают гипотонию, ареплексию и множественные врожденные контрактуры. В крупномасштабном анализе мутаций скрининг шести семейств XL-SMA дал результаты, указывающие на две новые мисссенс-мутации в двух семействах и новую синонимичную замену C → T в еще три семейства. Все эти обнаруженные мутации были расположены в экзоне 15 гена UBE1 (ген, кодирующий убиквитин-активирующий фермент), и наблюдали, как в семействах разделяют заболевание. В краткости, Missense UBE1 может привести к нарушению сложного здания с гигаксонином , белком, участвующим в структуре аксонов и поддержанию нейронов. Это может привести к нарушению деградации микротрубочки, ассоциированного с белком 1B (MAP1B), что приводит к наращиванию белка MAP1B, что может усилить гибель нейрональных клеток. [ 8 ] Таким образом, предполагается, что мутации в UBE1 являются причиной генетических дефектов у индивидуумов XL-SMA.
Ссылки
[ редактировать ]- ^ PDB : 3CMM ; Lee I, Schindelin H (июль 2008 г.). «Структурное понимание активации убиквитина, катализируемой E1, и переносится в конъюгирующие ферменты» . Клетка . 134 (2): 268–78. doi : 10.1016/j.cell.2008.05.046 . PMID 18662542 .
- ^ Jump up to: а беременный в дюймовый и фон Schulman BA, Harper JW (май 2009 г.). «Убиквитиноподобная активация белка ферментами E1: вершина для нижестоящих сигнальных путей» . Природа обзор молекулярной клеточной биологии . 10 (5): 319–31. doi : 10.1038/nrm2673 . PMC 2712597 . PMID 19352404 .
- ^ Lecker SH, Goldberg AL, Mitch WE (июль 2006 г.). «Деградация белка путем убиквитин-протеасом в нормальных и болезненных состояниях» (PDF) . Журнал Американского общества нефрологии . 17 (7): 1807–19. doi : 10.1681/asn.2006010083 . PMID 16738015 .
- ^ Tokgöz Z, Bohnsack RN, Haas Al (май 2006 г.). «Плейотропные эффекты связывания ATP.MG2+ в каталитическом цикле убиквитин-активирующих фермента» . Журнал биологической химии . 281 (21): 14729–37. doi : 10.1074/jbc.m513562200 . PMID 16595681 .
- ^ Jump up to: а беременный Lee I, Schindelin H (июль 2008 г.). «Структурное понимание активации убиквитина, катализируемой E1, и переносится в конъюгирующие ферменты» . Клетка . 134 (2): 268–78. doi : 10.1016/j.cell.2008.05.046 . PMID 18662542 .
- ^ Эта цифра адаптирована из схемы 1 «Механизм убиквитина, активирующего фермента: кинетический и равновесный анализ» Артура Хааса. Хаас Ал, Роуз И.А. (сентябрь 1982 г.). «Механизм убиквитина, активирующий фермент. Кинетический и равновесный анализ» . Журнал биологической химии . 257 (17): 10329–37. PMID 6286650 .
- ^ Jump up to: а беременный Ван Дж., Мальдонадо, штат Массачусетс (август 2006 г.). «Система убиквитин-протеасом и ее роль в воспалительных и аутоиммунных заболеваниях» (PDF) . Клеточная и молекулярная иммунология . 3 (4): 255–61. PMID 16978533 .
- ^ Jump up to: а беременный Ramser J, Ahearn Me, Lenski C, et al. (Январь 2008 г.). «Редкие миссенс и синонимичные варианты в UBE1 связаны с X-связанной с инфантильной мышечной атрофией позвоночника» . Американский журнал человеческой генетики . 82 (1): 188–93. doi : 10.1016/j.ajhg.2007.09.009 . PMC 2253959 . PMID 18179898 .
Внешние ссылки
[ редактировать ]- Убиквитин-активирующие+ферменты в Национальной медицинской библиотеке Медицинской библиотеки США (Mesh)